(pallas. 1776) (campanulariidae)
И. А. Косевнч, Н. Н. Марфенин
Obelia longissima (Pallas, 1766) — широко распространенный, возможно, космополитичный нид (Степаньянц, 1980), обычный компонент биоценозов обрастаний (Зевина, 1963). Образующий большие по сравнению с другими гидроидами побеги — до 30 см высотой (Наумов, 1960), этот вид не остается незамеченным при фаунистических исследованиях. Однако морфология О. longissima исследована недостаточно. Описание вида дано на основании изучения строения отдельных побегов (Веггill, 1949, 1950; Cornelius, 1975), а о строении колонии в целом ничего не известно.
Эта статья — продолжение начатой ранее серии описаний морфологии и биологии наиболее доступных для лабораторных исследований видов гидроидов: Obelia loveni (Allm.) (Марфенин, Косевич, 1984а, б), Dynamena pumila (L.) (Марфенин, 1974; Бурыкин, 1980; Карлсен, 1980), Moerisia maeoiica (Ostr.) (Марфенин, 1984).
Исследования проводили на Беломорской биологической станции МГУ летом 1981 и 1982 гг. Материал брали с Еремеевского порога пролива Великая Салма. .Морфологию О. longissima изучали на живых колониях. Для итого их выращивали из отдельных побегов (Crowell, 1957) или из фрустул иа стеклянных пластинах или полосках полиэтилена. Полоски с колониями содержали в море на глубине 0,5, 1,0 и 1,5 м, а пластины — в лаборатории в кристаллизаторах с постоянной аэрацией (воду меняли ежедневно). Лабораторные колонии кормили однодневными науплиусами Artetnia salina в избыточном количестве в течение 2 ч ежедневно. Температура в море была от 5 до 12, в лаборатории от 13 до 21°.
Для регистрации основных компонентов колонии применяли модифицированный метод картирования (Марфенин, 1980).
Поселяется О. longissima на различном субстрате — камнях, створках раковин, ламинариях, гидросооружениях, днищах судов, буях и других плавающих предметах, предпочитая открытые места. Образует густые многоярусные заросли, в которых выделяются высокие (10— :20 см) и разветвленные побеги (рис. 1,А).
В условиях Белого моря гидроид активен круглый год и рано приступает к размножению (Марфенин, Карлсен, 1983). В июне в планктоне обнаруживается масса медуз, а в конце июня — начале июля происходит оседание планул. В августе наблюдается депрессия, во время которой происходит значительное рассасывание и нарушение целостности ценосарка больших побегов. А в начале сентября уже появляются гонангии в молодых колониях, развившихся за лето.
Реклама

Колонии О. longissima, как и других видов рода Obelia, построены по радиальному плану. От первичного, или материнского, побега отходят несколько столонов 1-го порядка. Ветвясь, они дают столоны 2-го порядка, которые в свою очередь могут образовывать боковые столоны 3-го порядка и т. д. На столонах расположены вертикальные симподиально ветвящиеся побеги. От ствола побега отходят ветви 1-го порядка, от них — ветви 2-го порядка и т. д.
Морфология основных элементов колонии.
Гидрориза нитевидная, с хорошо различимыми отдельными столонами. Столоны разных порядков устроены одинаково. Перисарк столона представляет собой почти правильную трубку с диаметром около 0,2 мм (рис. 2, А). Ценосарк лишь в верхушке роста столона вплотную прилегает к перисарку на участке длиной около 0,2—0,4 мм. Обычно имеется 3—4 столона 1-го порядка (максимально 5). Боковые столоны появляются в любом месте материнского столона, кроме участка, расположенного дистальнее последнего побега. Место ветвления тяготеет к основаниям побегов, но в крупных колониях боковые столоны отходят и между побегами. В таких колониях почка бокового столона быстро расширяется, образуя в месте ответвления перетяжку (рис. 2,5). Растут столоны преимущественно прямолинейно, анастомозов практически не образуют.
Побеги образуются по мере роста столона в его дистальной части, Расстояние между побегами варьирует от 0,4 до 15,5 мм (среднее 3,7— 3,9 мм), к периферии колонии оно несколько увеличивается (рис. 1,5). В основании больших побегов столоны слегка расширены. Там же образуется “узел гидроризы” за счет отхождения по обе стороны основания побега нескольких боковых столонов, которые в свою очередь сразу же дают боковые ветви (рис. 1,А) (Косевич, 1984).
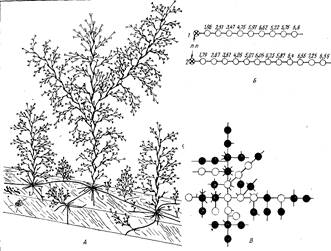
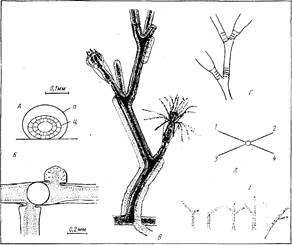
Рис. 2. Строение отдельных частей колонии: А — поперечное сечение столона (п — иерисарк, ц — ценосарк); Б — характер отхождения боковых ветвей столона (круг-место отхождения побега); В — проксимальная часть “гигантского” побега. Образование полисифонности за счет стелющихся столонов; Г — строение междоузлия “нормального” побега {В и Г — в одинаковом масштабе); Д — схема отхождения ветвей 1-го порядка от ствола побега (цифры — порядок ветвей по вертикали); Е — примеры аномалий щупалец гидрантов
Реклама

Побеги в колонии встречаются двух типов — “нормальные” и “гигантские” ] (Косевич, 1984). Они одинаковой формы, только у “гигантских” побегов длина и диаметр междоузлий в 1,5—2 раза больше, чем у “нормальных”, размеры которых более типичны для семейства Саmpanulariidae, и рода Obelia в частности (рис. 2, 5, Г). Материнский и несколько ближайших к нему побегов — “нормальные”. “Гигантские” побеги появляются в колонии, имеющей 6—10 побегов. В дальнейшем часто наблюдается чередование “гигантских” и “нормальных” побегов. С ростом колонии частота образования “гигантских” побегов возрастает (рис. 1,5). Расстояние между соседними “гигантскими” побегами составляет в среднем 4—5 мм, но никогда не бывает меньше. 2 мм (“нормальные” побеги могут появляться на расстоянии друг от друга менее 1 мм, в среднем — 2—3 мм).
Примечание. Термин “нормальные” побеги предложен нами для обозначения побегов той размерной группы, которая соответствует первому побегу, развивающемуся из пла-нулы или фрустулы. При недостатке питания в колонии имеются лишь “нормальные” побеги; при благоприятных условиях — и “нормальные”, и “гигантские”.
Все побеги состоят из междоузлий, однотипных по плану строения. Верхушка роста очередного междоузлия закладывается на границе гладкой части междоузлия и ножки гидранта. Угол между соседними междоузлиями составляет 30°. Каждое следующее междоузлие лежит нe в вертикальной плоскости, проходящей через предыдущее междоузлие, а несколько отклоняется от нее. Таким образом, ствол у побега О. longissima оказывается спирально-зигзагообразным.
Боковые ветви 1-го порядка отходят от каждого междоузлия стебля. Верхушка роста ветви закладывается на основании ножки гидранта с прилежащей стороны, несколько сбоку от плоскости, проходящей через ножку и междоузлие ствола. Существует довольно строгая последовательность отклонения ветвей 1-го порядка от этой плоскости: поочередно по две то влево, то вправо (рис, 2,Д). Это обеспечивает приблизительно крестообразное расположение их относительно оси стебля. Ветви 1-го порядка перпендикулярны стволу и параллельны субстрату. Эти правила ветвления полностью распространяются и на ветви 2-го и 3-го порядков.
Важная особенность “гигантских” побегов О. longissima — это прогрессивное уменьшение размеров междоузлии в ряду: ствол — ветви 1-го порядка — ветви 2-го порядка и т. д. (табл. 1). Чем выше порядок ветви, тем быстрее она перестает расти. Вследствие этого, а также из-за различий в размерах междоузлии с возрастанием порядка ветви ее длина уменьшается. Самые нижние ветви “гигантских” побегов обычно рассасываются, замещаясь более молодыми. Из оснований ножки вторичных гидрантов в проксимальной части побегов вырастают столоны, которые стелятся вниз по стволу побега. (рис. 2, Г). особенно важно при увеличении размеров побегов.
Таблица 1
Диаметр (d) и длина (I) междоузлии в стволе и ветвях разного порядка в трех типичных “гигантских” побегах Obelia longissitna, мм
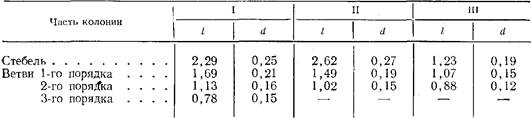
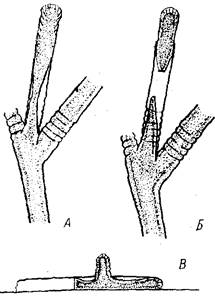
Достигнув его основания, столон переходит на субстрат и некоторое время растет по нему, формируя иногда один-два побега (рис 2,Г). Это увеличивает прочность и жесткость основания побега, усиливает прикрепление его к субстрату, что особенно важно при увеличении размеров побегов. фрустулы. Как в естественных условиях, так и при содержании в лаборатории у О. longissima образуются фрустулы (Иванова-Казас, 1977). При этом дистальные междоузлия ветвей любого порядка вмести образования гидранта продолжают расти, формируя гладкую трубку, превышающую в 1,5—2 раза длину междоузлия. Ценосарк внутри этой трубки сильно истончается в месте перехода кольчатой части перисарка в гладкую и в конце концов рвется (рис. 3,А,5). Отделившаяся верхушка роста продолжает перемещаться н дистальном направлении, при этом становится заметным (в электронном сканирующем микроскопе) отсутствие перисарка на ее переднем конце. Вскоре после этого верхушка роста, ставшая фрустулой, “вываливается” из перисарка и при первом же прикосновении к любому субстрату приклеивается к нему и продолжает рост наподобие отрезка столона. Примерно через 12 ч на ней появляется дорсальная верхушка роста (рис. 3,В), а через сутки из нее формируются междоузлие и гидрант, имеющие размеры меньшие, чем первое междоузлие и гидрант, сформировавшиеся на столоне,
Чаще всего фрустуляция наблюдается при обильном кормлении колонии или резком повышении температуры на 4—6°.
Гидранты. Все гидранты в колонии вне зависимости от того, на каких ветвях они расположены, имеют одинаковые размеры. Форма гидрантов О. longissima типична для рода Obeliaa. Своей базальной частью тело гидранта опирается на диафрагму гидротеки. Щупальца расположены одним венчиком диаметром 2 мм; поочередно изгибаясь вверх и вниз, они образуют два яруса. Обычно в венчике от 28 до 32 щупалец. Булавовидный гипостом способен широко раскрываться. Средняя высота гидранта от диафрагмы до кончика типостома — 0,8 мм. В расправленном состоянии щупальца нижнего яруса опираются на край гидротеки, фиксируя положение гидранта. Когда гидрант сжимается, щупальца складываются над ним конусом. Встречаются аномальные щупальца — двух- и трехвершинные (рис. 2,Е).
Рис. 3. Образование фрустул: Л — фруетулообразующая ветвь, Б — отделение фрустулы от ценосарка колонии, В — прорастание фрустулы на субстрате, вид сбоку
Гидротека — симметричной бокаловидной формы. Высота ее от устья до диафрагмы в среднем 0,5 мм (от 0,4 до 0,7 мм); высота от устья до ножки — в среднем 0,6 мм; диаметр устья — от 0,25 до 0,45 мм (средний — 0,31 мм). Отношения высоты гидротеки к диаметру устья соответственно равны: 1,54 (от 1,25 до 1,91) и 1,81 (от 1,5 до 2,18) (129 измерений).
Формирование гидранта О. longissima при температуре 18—20° происходит примерно за 24 ч, из которых около 4 ч уходит па выдавливание крышки гидротекн. Вышедший гидрант уже полностью сформирован и больше не растет. Продолжительность жизненного цикла гидранта в среднем составляет 6—7 сут и варьирует от 1 до 14 и более суток (130 измерений). Вслед за дедифференциацией и рассасыванием гидранта после диапаузы (не менее суток) формируется следующий гидрант. При этом происходит некоторое удлинение его ножки. По форме и размерам первичные и вторичные гидранты не различаются. Нам удалось проследить пять поколений гидрантов, однако остается неясным, ограниченно ли число таких поколений.
Количественные показатели структуры колонии. О структуре колонии в целом можно судить по количественным показателям, введенным ранее (Марфенин, 1977; Марфенин, Косевич, 1984а). Это попарные соотношения таких величин, как L — общая длина гидроризы, М — суммарная длина ценосарка побегов, С — общая длина ценосарка всей колонии (C=L+A1), Н — число функционирующих гидрантов, В — число всех верхушек роста, / — суммарное число междоузлии побегов. Каждый из показателей структуры колонии (табл. 2) имеет свой функциональный смысл.
Таблица 2
Показатели структуры колоний Obelia longissima разного размера в естественных и лабораторных условиях (июнь—июль 1981 г.)
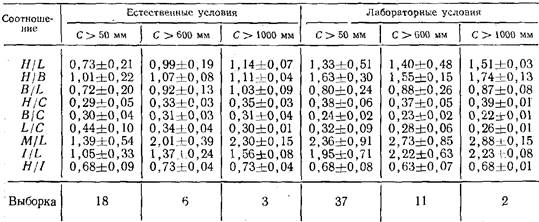
Так, M/L характеризует распределение массы колонии между побегами и гидроризой, 7/L — дает возможность сравнивать разные виды по соотношению числа структурных элементов побегов на единицу длины гидроризы (занимаемой площади), Н// — характеризует отношение числа имеющихся в колонии функционирующих гидрантов к их максимально возможному числу. Характеристики остальных показателей даны ранее (Марфенин, Косевич, 1984а).
Здесь структурные показатели рассчитаны на основании картирования 9 колоний, росших в естественных условиях, и 11 — из лаборатории. Их сравнение выявило некоторые морфологические различия. В естественных условиях гидрориза составляла большую часть колонии, чем у лабораторных колоний, в результате чего плотность расположения междоузлии и гидрантов на занимаемой площади оказалась меньше (H/L, I/L). В естественных условиях большей оказалась раз-ветвленность колонии (В/С) из-за большего числа верхушек роста гидроризы (сравнение показателей В/С, Н/В, H/L).
Обсуждение результатов. О. longissima приспособлена к обитанию на открытом пространстве при несильном течении, о чем свидетельствуют все особенности строения колонии.
В колонии наряду с “нормальными”, обеспечивающими облов прилежащего к субстрату слоя воды побегами, имеются “гигантские”, которые способствуют расширению пространства облова. Однако такие побеги в силу своей громоздкости и большой парусности становятся уязвимыми для всевозможных механических повреждений, которые неизбежны при обитании в густых зарослях водорослей или на сильном течении. Кроме этого, в сильном потоке все ветви больших побегов складываются вдоль ствола, а сам побег пригибается к субстрату, что сводит на нет основное преимущество такого типа строения.
Некоторые особенности морфологии гидроида позволяют понять, как на основе типичного для рода Obelia строения становится возможным формирование и само существование побегов, столь характерных для О. longissima.
Чередование на столоне “нормальных” и “гигантских” побегов уменьшает перекрывание крон последних и обеспечивает равномерный облов всего объема, занятого колонией. Уменьшение размеров междоузлии в “гигантском” побеге при переходе от ствола к ветвям 1-го порядка, а затем соответственно к ветвям 2-го и 3-го порядков и облегчает всю конструкцию, и делает ее более гармоничной, так как уменьшает перекрывание ветвей внутри побега. К этому же результату приводит упорядочение ветвления побегов,
Уменьшение изгиба междоузлии и утолщение псрисарка (особенно проявляющееся в стволе) должны увеличивать жесткость конструкции побега. Полисифонпость базальной части побега и “узел гидроризы” в его основании увеличивают прочность крепления побега к субстрату. Малая высота колец в проксимальной зоне кольчатости междоузлии по отношению к их диаметру при относительно большом числе колец (до 10) повышает гибкость побегов.
Значительная часть указанных особенностей О. longissima обусловлена усложнением морфогенеза побегов, проявляющимся в способности к изменению диаметра верхушек роста и в почковании боковых ветвей в определенном порядке. Это означает, что по морфологии побегов О. longissima устроена сложнее, чем О. loveni, хотя жизненный цикл у нее проще — с типичным незамаскированным метагенезом.
Размножение О. longissima с, помощью медуз позволяет этому виду осваивать новые места обитания. Присущая же ей способность к фрустуляции в благоприятных условиях делает возможным быстро увеличивать популяцию вида в уже освоенном месте.
При анализе строения колонии бросается в глаза то, что практически все структурные показатели изменяются с ее ростом. Становление структуры колонии происходит постепенно. Обнаруживается тенденция возрастания доли побегов в общей массе колонии, что приводит к увеличению числа верхушек роста и гидрантов.
Почти неизменными с момента начала развития колонии остаются три показателя: H/В, В/С, Я/I, что объясняется постоянством зависимости числа верхушек роста от числа гидрантов, обеспечивающих их пищей, необходимой для роста. Кроме этого, постоянство показателя В/С может быть следствием упорядоченности ветвления, что является характерной особенностью вида. С ростом колонии увеличиваются размеры и число “гигантских” побегов.
Таблица 3
Распределение верхушек роста (ВР) и междоузлии (МУ) в стволе и ветвях разного порядка в трех “гигантских” побегах Obelia longissitna
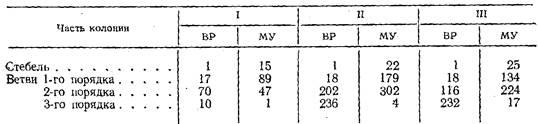
Это приводит к большей концентрации гидрантов на единице занимаемой колонией площади. У трех обследованных нами “гигантских” побегов 90% междоузлий, а соответственно гидрантов, и верхушек роста приходилось на боковые ветви побегов (табл. 3). Такая ситуация вообще характерна для видов с разветвленными побегами.
Итак, подробное изучение морфологии О. longissima позволяет не только полнее описать этот вид, известный до сих пор лишь по строению зооидов и отдельных междоузлии, но и сделать еще один шаг на пути уточнения границ между видами рода Obelia, введя новые признаки и показатели для их сравнения, а также продвинуться в понимании морфогенетических проявлений эволюции гидроидов семейства Campanulariidae.
Список литературы
Бурыкин Ю. Б. 1980. Регулирующая роль некоторых экологических факторов в процессах роста и интеграции колониальных гидроидов//Теоретическое и практическое значение кншечнополостных. Л, С. 16—19.
3евина Г. Б. 1963. Обрастание на Белом море/'/Тр. Ин-та океанол. АН СССР. 70. 52—71.
Иванова-Казас О. М- 1977. Бесполое размножение животных. Л.
Карлсен А. Г. 1980. Изучение строения, роста и процессов интеграции колоний у гидроидов при лабораторной культивации на примере Dynamena puinila (L.) //Теоретическое и практическое значение кишсчнополостных. Л. С. 34—40. Косевнч И. А. 1984. Диморфизм побегов в колонии гидроида Obelia longissima (Pallas, 1766) (Hydrozoa, Thecaphora, CampanuJariidac). Деп. в ВИНИТИ 16.03.84 1984, № 1507. Марфенин Н. Н. 1974. Структура колонии гидроидного полипа Dynamena pumila (Hydrozoa, Leptolida) // //Тр. Беломорск. биол- станции МГУ. 4. 140—147. Марфенин Н. Н. 1977. Опыт изучения интеграции колонии гидроида Dynamena pumila (L.) с помощью количественных морфологических показателей//Журн. общ. биол. 38, № 3. 409—422. Марфенин Н. Н. 1980. Метод картирования пространственной организации колониальных Hydrozoa и его значение при изучении частей колонии//Теоретическое и практическое значение кншечнополостпых. Л. С. 66—69. М а р ф е н и и Н. И. 1984. Морфофункииональный анализ временной колонии на примере гидроида Moerlsia rnaeotica (Ostr., 1896) (Leptolida, Limnomedusae) // Журн. общ. биол. 45,№ 5. 660—669.
Марфенин Н. Н., Карлсен Л. Г. 1983. Состояние фауны гидроидов Еремеевского порога Белого моря ранней весной// Науч. докл. высш. школы. Биол. науки. № 2. 26—30.
Марфенин Н. Н., Косевич И. А. 1984а. Морфология колонии у гидроида Obelia loucni (Ailm.) (Campanulariidae) // Всстн. Моск. ун-та. Сер. Биол. № 2, 37—46. Марфенин Н.Н., Косевнч И. А. 19846, Биология гидроида Obelia /оveni (AIIm.): образование колонии, поведение и жизненный цикл гидрантов, размножение//Вести. Моск. ун-та. Сер. Биол. № 3. 16—24. Наумов Д. В- I960. Гидроиды н гидромедузы морских, солоноватоводных и пресноводных бассейнов СССР. М.—Л. Степаньянц С, Д. 1980. О космополитизме у гидроидов//Теоретическое и практическое значение кишечнополостных. Л. С. 114—122 Berrill N. J. 1949. The polymorphic transformation of Obelia // Quart. J. Microscop. Sci. 90, part 3. 235—264. Berrill N. J. 1950. Growth and form in Calyptoblastic Hydroids. 1. Comparison of Campanulid, Campanularian, Serlularian and Plumularian//J. Morphol, 85. 297—335. Cornelius P. F. S. 1975. The hydroid species of Obelia (Coelenterata, Hydrozoa:Campanulariidae), with notes on the medusa stagey/Bull. Br, Mus, nat. Hist. (Zool.). 28. 249—293. Crowell S. 1957. Differential responses of growth zones to nutritive level, age, and temperature in the colonial hydroid Campanularia//J. Exp. Zool. 134, N I. 63—90.
|
