Стереотип кормового поведения птиц: эколого-эволюционные аспекты
А.Г. Резанов
Концепция кормового метода
При описании кормового поведения птиц используют различные подходы. Один из самых распространенных вариантов — описание поведения птиц
Принятая терминология. Кормовой манёвр («feeding manoeuvre (maneuver)») [41, 42, 60] — последовательность стереотипных ло- комоций и моторных актов фуражира при разыскивании и особенно при атаке добычи. Связи фуражира с субстратом (его элементами) при описании поведения в рамках «манёвра» не придается основополагающего значения. Кормовой метод. Русский вариант термина «feeding method» впервые использован автором [8]. Термин «кормовой метод» шире понятия «кормового манёвра», поскольку подразумевает под собой связь кормового манёвра с условиями окружающей среды [11, 12, 15] (рис. 1).
*
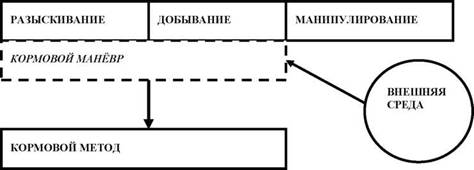
Рис. 1. Общая схема кормовой поведенческой последовательности; представление о кормовом манёвре и кормовом методе
(в основном древесно-кустарниковых Passeriformes) при помощи кормовых манёвров [41, 42, 60].
Описывая кормовое поведение птиц на уровне «кормового манёвра», исследователь вынужденно игнорирует его связь с элементами среды (ассоциации с другими животными, техническими средствами и пр.), не описывает типы клевков и пр. Такой подход к изучению кормового поведения представляется «непростительной роскошью», поскольку при этом теряется важная информация.
Например, кормовой манёвр «aerialfly-catching» («схватывающий полет» [9]) известен для большого числа, в том числе далеко не родственных видов, родов и даже семейств — Passeriformes: Muscicapidae, Laniidae, Bombycillidae, Campephagidae, Dicruridae, Tyrannidae и др.; Coraciiformes: Meropidae, Todidae, Momotidae, Coraciidae; Trogoniformes; Piciformes: Galbulidae, Bucconidae и др. [12]. Автором предложен подход, при котором кормовое поведение птиц рассматривается как кормовая поведенческая последовательность, центральным звеном которой является кормовой метод [11; 12; 15]. На основе концепции кормового метода разработана специальная система классификаторов [12], отражающая поведение фуражира, его связи с элементами среды, пространственное положение и тип пищевого объекта. При помощи цифровых кодов описывается любой вариант поведения от нормы до модификации. Закодированный кормовой метод представляет собой «цепочку» цифр, каждый из компонентов которой сравним с аналогичными компонентами других «цепочек». Система открыта для дополнений.
Реклама

О стереотипе кормового поведения птиц: норма и модификации
Птицы обладают определенным набором стереотипных двигательных актов, с помощью которых они добывают себе корм [30]. На практике под стереотипом кормового поведения понимают наиболее оригинальные («шаблонные») кормовые методы, на основе которых возможна полевая идентификация вида [6]. Некоторые стереотипные кормовые методы не являются оригинальными и присутствуют в арсенале других видов, т. е. являются конвергентными, что затрудняет использование признаков кормового поведения в систематике птиц [12, 21]. Исследованию стереотипа кормового поведения птиц посвящена целая серия работ [11, 12, 14, 22, 25, 31]. Стереотип кормового поведения определяется набором стабильных «фиксированных» элементов (локомоций и моторных актов), проявляющихся в зависимости от конкретной кормовой ситуации [11]. Существует понятие принципа «изначального разнообразия» — это своего рода программа, которая реализуется при определенных условиях в рамках морфо-физиологического оснащения вида, т. е. как видоспецифическое поведение. В контексте кормового поведения — это представление о кормовом наборе и кормовом репертуаре [14, 22]. Кормовой набор (арсенал) — некое число кормовых методов (стереотипных кормовых манёвров, связанных с конкретными средами/субстратами), соответствующее морфо-физиологическому оснащению вида (популяции). Это генетически детерминированная и в то же время вероятностная поведенческая программа, которая в формате одной особи может быть реализована только частично. Кормовой репертуар — реализованная часть видового (популяционного) набора кормовых методов, постоянно или периодически используемых конкретными особями (популяциями).
Изменения, происходящие в кормовой ситуации, как правило, приводят к «переключению» фуражира с одного стереотипного кормового метода на другой (достаточно смены одной из составляющих), более адекватный на данный момент времени. Происходит чередование кормовых методов, входящих в репертуар особи. Так, белая трясогузка за минуту кормежки на асфальте нередко использует до 4-5 кормовых методов, периодически чередуя их [14, 22]. В местностях с сезонной выраженностью климата у ряда видов птиц существует разделение стереотипа на «летний» и «зимний» [26].
Реклама

По встречаемости все кормовые методы можно разделить на фоновые, обычные, редкие и необычные. Суровые условия (зима, ранняя весна), ведущие к бескормице в привычных местообитаниях, вынуждают птиц либо «переключаться» на необычные корма, для добывания которых используются необычные кормовые методы, либо перемещаться в непривычные местообитания, иная структура которых уже сама по себе провоцирует «уклоняющиеся» формы кормового поведения. В послегнездовой период необычные методы могут приобретаться молодыми птицами (особенно у мигрирующих форм), попавших в непривычную для них экологическую обстановку [7]; со временем они могут входить в кормовой репертуар особи. Среди необычных кормовых методов встречаются и так называемые «случайные» методы (единичное упоминание для вида), однако их «случайность» может быть результатом элементарного отсутствия информации.
Под модификациями кормового поведения автор понимает формы поведения, уклоняющиеся от традиционной нормы (по типу локомоций, по связям с элементами среды и пр.), но не выходящие за пределы видовой адаптивной зоны. «Уклоняющиеся» формы кормового поведения, или необычные кормовые методы (unusual feeding methods, or modes), как правило, не выходят за пределы видоспецифических действий и являются поведенческими модификациями. Но далеко не все модификации поведения по встречаемости в кормовом репертуаре видов (популяций) входят в категорию необычных кормовых методов — например, такая антропогенная модификация поведения, как следование грачей (Corvus frugilegus) и чаек (Larus spp.) за плугом, давно уже стало фоновой для этих птиц [19]. Предлагается разделение модификаций кормового поведения на три категории:
Модификация кормового метода по субстратной составляющей (использование нетрадиционного для кормового поведения субстрата, элемента среды и пр.), без затрагивания кормового манёвра [2, 17, 23, 24, 45, 46, 48, 51-53, 55, 57, 62-65, 67-69].
Модификация кормового метода на уровне кормового манёвра (использование нетрадиционного для кормового поведения манёвра) [1, 3, 5, 7, 13, 16, 18, 20, 27, 35, 37, 49, 58, 61].
Модификация кормового метода на уровне манёвра и субстрата [9, 10, 33, 36, 38, 39, 59].
Некоторые модификации (особенно третьей категории), по-видимому, могут быть квалифицированы как поведенческие аберрации [5, 10, 16, 17, 20, 36-38, 62].
Возникающая между конспецифическими популяциями географическая дифференциация, в частности в отношении кормового поведения, как лабильного, или пластичного признака фенотипа, рассматривается как «микроэволюция кормового поведения» [34]. Ввиду дефицита информации по этому вопросу оценка того или иного кормового метода птиц как «географического» затруднена. Так, кормовые методы серой цапли (Ardea cinerea), используемые при добывании корма на глубоководных участках, необычны и документально зарегистрированы только в нескольких графствах Англии, в Германии и в России на Псковском озере. В районе Аббертонского водохранилища (Эссекс, Англия), такое поведение характерно для целой популяции цапель [10]. На Псковском озере добывание цаплями рыбы на глубоких местах отмечено только в течение 2 дней — 15 регистраций за два месяца наблюдений в течение двух летних сезонов [10]; вероятнее всего, такое поведение носит случайный характер.
Еще сложнее оценить географический аспект кормежки травника (Tringa Manus) в манере большого улита (T.nebularia), поскольку информация по этому вопросу крайне фрагментарна — Англия [61] и Дагестан, Россия [7].
Ярко выражена географическая изменчивость питания у палеаркти- ческих видов хищных птиц. Переход на преимущественное потребление иных видов добычи в большинстве случаев требует новых способов охоты. Е.В. Козлова [4] отмечала трофическую географическую дифференциацию степного орла даже в пределах подвида: A.rapax orientalis в Центральной Азии питается грызунами, а на зимовках в Сомали — падалью.
Отдельные кормовые методы, приуроченные к конкретному географическому пространству, функционируют в определенные периоды времени, затем «угасают» и через какое-то время проявляются вновь [12]. Предположительно, что наряду со стабильными (в рамках пространственно-временного континуума) кормовыми методами у локальных групп птиц могут иметь место, по-видимому, стереотипные, но нестабильные во времени и пространстве, кормовые методы. Конкретная окружающая обстановка, обуславливающая состояние кормовой базы и оцениваемая птицей-фуражиром, является своеобразным пусковым механизмом (или блокатором) того или иного кормового метода.
Уклоняющиеся формы кормового поведения: эколого-эволюционный аспект
Проявление «отклоняющихся» от традиционных кормовых методов, но не выходящих за пределы стереотипа, форм кормового поведения, как правило, наблюдается в послегнездовой период, особенно у мигрирующих видов. Вероятнее всего, новые кормовые методы начинают использовать молодые птицы, и в дальнейшем они могут входить в кормовой репертуар этих птиц. Таким образом, в пределах популяции формируется фенотипическая изменчивость по кормовому поведению, что определенно ведет к расширению видовой (популяционной) ниши.
Некоторые необычные специализированные кормовые методы практически выходят в промежуточные участки между зонами или даже входят в чужие адаптивные зоны. Например, травник, кормящийся в манере большого улита, по сути, использует видовой кормовой метод последнего [7, 61]. Исходя из принципа конкурентного исключения Г аузе, перспектив у травника в этом направлении нет. Действительно, большой улит морфологически (морфология челюстного аппарата) больше, чем травник, приспособлен к схватыванию и сдавливанию (удержанию) клювом мелких рыбок. Однако в природе возможно совместное существование видов со сходными потребностями практически в одной и той же нише (принцип балансирующего, или динамического, отбора). С другой стороны, встречаемость таких методов в пространственно-временном формате крайне невелика (большой разброс в пределах историко-географического пространства по травнику — Англия и Дагестан — с расхождением в 30 лет), чтобы говорить о серьезных эволюционных последствиях необычных форм кормового поведения. В лучшем случае возможно распространение и функционирование таких методов в пределах локальных популяций с возможной перспективой частичного выхода в новые адаптивные зоны.
Некоторые кормовые методы, проявляющиеся в локальных популяциях (группировках), могут существовать какое-то время и вновь исчезать [12]. Например, «пирсинг» («piercing») нектарников фритиллярии лазоревками (Parus caeruleus) в Оксфордском ботаническом саду [66] или хищничество большого пестрого дятла (Dendrocopus major) в одном лесничестве Германии, которое не проявлялось в течение 20 лет, а потом вновь (1992 г.) стало регистрироваться наблюдателями [56]. Возможно, существуют своего рода локальные «нестабильные» кормовые методы, периодически проявляющиеся в рамках определенного пространственно-временного континуума.
Представляет собой интерес ситуация с выкладыванием орехов на проезжую часть американскими воронами (Corvus brachyrhynchos). Такое поведение было признано целенаправленным и целесообразным [47, 54]. Позднее было показано, что вороны бросают на дорогу орехи как в присутствии, так и при отсутствии автомобилей. Действия птиц оценены как эпизодические, ненаправленные [40].
Возникновение (проявление) новых кормовых модификаций может происходить на уровне отдельных особей. Так, в локальной популяции попугаев кеа (Nestor notabilis) в Маунт Кук Вилладж (Новая Зеландия) некоторые особи с целью добывания пищевых отбросов научились при помощи клюва открывать крышки мусорных вёдер [44]. У больших синиц (Parus major) в Англии распространение необычного кормового метода (продалбливание клювом крышек молочных бутылок) начиналось с отдельных локальных популяций [43, 50].
Можно ли «уклоняющиеся» формы кормового поведения считать эволюционным резервом вида (популяции), хранящимся в кормовом арсенале? Вероятно, да. Также очевидно, что при этом происходит расширение реальной популяционной экологической ниши. Формирование стереотипа, по-видимому, приурочено к периоду становления вида (популяции) при его (её) переходе в новую адаптивную зону, в которой начинается его эволюция. Адаптивную зону [29] можно рассматривать как экологическую нишу, в которой эволюционирует таксон [28].
Возможный эволюционный сценарий. В период становления вида (популяции) в конкретной адаптивной зоне в рамках его морфо-физилогического оснащения происходит формирование видоспецифического набора (арсенала) кормовых методов на базе существующего набора, имеющегося в распоряжении предковой формы. При формировании у нового вида отличной морфологии какие-то исходные кормовые методы, не адекватные новому морфо-физиологическому оснащению вида, выпадают из видового набора. На базе сформированного кормового набора, в соответствующих условиях, путем составления новых комбинаций между кормовыми манёврами и средами (элементами среды, субстратами), возникают новые (модифицированные) формы кормового поведения. Новые кормовые методы появляются в репертуаре особи (популяции) и с этого момента уже входят в состав кормового арсенала вида (популяции). Также существуют так называемые нестабильные в формате пространственно-временного континуума кормовые методы [12], известные для некоторых локальных популяций птиц [56, 66].
Каждая, отдельно взятая особь использует часть кормового набора в виде кормового репертуара. Естественно, что суточный кормовой репертуар меньше сезонного, сезонный меньше годового, а годовой меньше репертуара, использованного особью в течение всей жизни. Это очевидно, поскольку в течение дня птица пребывает в более однообразной экологической ситуации, чем в течение всей своей жизни, а вид, в целом, за исторический отрезок мониторинга его кормового поведения сталкивается со значительно более разнообразными условиями существования. Так, дневной репертуар белой трясогузки, кормящейся на дорожках, может включать в себя до 7-8 кормовых методов, в то время как кормовой репертуар вида на пространстве ареала состоит более чем из 70 кормовых методов. В целом, трясогузка использует порядка 10 различных кормовых манёвров, которые в сочетании с элементами среды формируют всё многообразие кормовых методов [14, 22].
Исторические репертуары особей вида (популяции) слагают кормовой набор вида, т. е. его стереотипное (видоспецифическое) поведение. Такое поведение адекватно видовой (популяционной) адаптивной зоне. Появление в кормовом репертуаре уклоняющихся форм кормового поведения служит эволюционным резервом, благодаря которому в определенных ситуациях, вероятно, возможен выход в новую адаптивную зону. Помимо стабильных форм поведения, в составе кормового набора встречаются «нестабильные» («случайные») кормовые методы. В различных географических точках видового ареала нестабильные формы кормового поведения могут, без видимых на то причин, периодически «появляться» или «исчезать» из кормового репертуара вида (популяции). Тем не менее они остаются в составе кормового арсенала вида (популяции), являясь своеобразным его (ее) эволюционным резервом.
Список литературы
Бёме Л.Б. Некоторые особенности биологии птиц Центрального Казахстана // Бюллетень МОИП. Отд. биол. Т. 55. Вып. 5. М., 1950. С. 44-48.
Бородин О.В. Необычная охота тетеревятника // Современная орнитология 1991. М.: Наука, 1992. С. 266.
Жмуд М.Е. Необычный способ охоты малой белой цапли на рыб // Прикладная этология. М.: Наука, 1983. С. 250-251.
Козлова Е.В. Птицы зональных степей и пустынь Центральной Азии. Л.: Наука, 1975. 251 с.
Плешак Т.В. Необычное кормовое поведение клестов-еловиков Loxia curvirostra // Русский орнитологический журнал. 1998. Т. 7 (44). С. 22.
Промптов А.Н. Видовой стереотип поведения и его формирование у диких птиц // Доклады АН СССР. Нов. сер. Т. 27. Вып. 2. М., 1940. С. 171-175.
Резанов А.Г. Необычный способ охоты травника (Tringa totanus L.) // Фауна и экология позвоночных животных. М.: МГПИ, 1976. С. 98-101.
Резанов А.Г. Кормовое поведение и способы добывания пищи у белой трясогузки Motacilla alba (Passeriformes, Motacillidae // Зоологический журнал. Т. 60. Вып. 4. М., 1981. С. 548-556.
Резанов А.Г. Случаи воздушного кормления большого пестрого дятла // Орнитология. Вып. 17. М.: МГУ, 1982. С. 188.
Резанов А.Г. Способы охоты серой цапли Ardea cinerea при добывании рыбы на глубоководных участках // Русский орнитологический журнал. 1995. Т. 4 (1-2). С. 25-27.
Резанов А.Г. Кормовое поведение птиц как многовариантная поведенческая последовательность: изменчивость и стереотипность // Русский орнитологический журнал. 1996. Т. 5 (1-2). С. 53-63.
Резанов А.Г. Кормовое поведение птиц: метод цифрового кодирования и анализ базы данных. М.: Высшая школа, 2000. 224 с.
Резанов А.Г. Воздушная охота галок Corvus monedula и озерных чаек Larus ridibundus // Русский орнитологический журнал. 2002. Т.11 (179). С. 245-246.
Резанов А.Г. Кормовое поведение Motacilla alba L. 1758 (Aves, Passeriformes, Motacillidae): экологический, географический и эволюционный аспекты. М.: МГПУ, 2003. 390 с.
Резанов А.Г. Использование понятий «кормовой манёвр» и «кормовой метод» в описании кормового поведения птиц // Орнитологические исследования в Северной Евразии. Ставрополь: СГУ, 2006. С. 433-434.
Резанов А.Г. Серые вороны Corvus cornix кормятся плодами рябины, подвешиваясь вниз головой // Русский орнитологический журнал. 2006. Т. 15 (309). С.130-131.
Резанов А.Г. Добывание серой вороной Corvus cornix корма из воды // Русский орнитологический журнал. 2007. Т. 16 (352). С. 460-461.
Резанов А.Г. Серая ворона Corvus cornix использует «топанье» для добывания дождевых червей // Русский орнитологический журнал. 2007. Т. 16 (366). С. 896-898.
Резанов А.Г. Историко-географический анализ «следования за плугом» у птиц // Русский орнитологический журнал. 2008. Т. 17(410). С. 499-513.
Резанов А.Г. Серые вороны Corvus cornix кормятся вишнями, подвесившись вниз головой // Русский орнитологический журнал. 2008. Т. 17 (449). С. 1554-1555.
Резанов А.Г. О возможности использования признаков кормового поведения в систематике птиц // Вестник МГПУ. Серия «Естественные науки». 2008. № 2(24). С. 53-59.
Резанов А.Г. Кормовое поведение Motacilla alba L., 1758. LAP LAMBERT Academic Publishing, 2012. 436 с.
Резанов А.Г. Необычный кормовой метод пищухи Certhia familiaris: обследование глубокой автомобильной колеи // Русский орнитологический журнал. 2012. Т. 21 (759). С. 1181-1182.
Резанов А.Г. О кормовом поведении кваквы Nycticorax nycticorax // Русский орнитологический журнал. 2012. Т. 21 (791). С. 2131-2137.
Резанов А.Г., Резанов А.А. Историко-географический аспект стереотипного кормового поведения птиц // Орнитологические исследования в Северной Евразии. Ставрополь: СГУ, 2006. С. 434-436.
Резанов А.Г., Резанов А.А. Оценка разнообразия кормового поведения большого пёстрого дятла Dendrocopos // Русский орнитологический журнал. 2010. Т. 19 (570). С.831-860.
Резанов А.Г., Резанов А.А. О кормовом поведении малой белой Egretta garzetta и жёлтой Ardeola ralloides цапель на мелководьях и сплавинах Нила // Русский орнитологический журнал. 2010. Т. 19 (543). С. 61-66.
Северцов А.С. Теория эволюции. М.: ВЛАДОС, 2005. 380 с.
Сипсон Д. Темпы и формы эволюции. М.: ИЛ, 1948. 359 с.
Хайнд Р. Поведение животных. Синтез этологии и сравнительной психологии. М.: Мир, 1975. 855 с.
Хлебосолов Е.И. Стереотип кормового поведения птиц // Успехи современной биологии. 1993. Т. 113 (6). С. 717-730.
Хохлов А.Н. О необычном трофическом поведении обыкновенной иволги в Ставропольском крае // Кавказский орнитологический вестник. Ставрополь: СГУ, 1991. № 2. С. 110.
Allsopp E., Allsopp К. Night Herons swimming // British Birds. 1965. Vol. 58. № 7. P. 297.
ArnoldS.J. The microevolution of feeding behavior // Foraging behavior. Ecological, Ethological, and Psychological approaches, Garland STPM Press, New York & London, 1981. P. 409. 453.
BergstromB.J. Unusual prey-stalking behavior by a goshawk // J. Field Ornithology. 1985. Vol. 56. № 4. P. 415.
Boyle G. Heron fishing in deep water // British Birds. 1967. Vol. 60. № 5. P. 215.
Brewster W. The birds of Lake Umbagog region of Maine // Bull. Mus. Comp. Zool. 1925. Vol. 66. P. 211-402.
Collar N. Golden orioles (Oriolus oriolus) diving into water // British Birds. 1975. Vol. 68. P. 245-246.
CornishA.VJackdaws hawking flying ants // British Birds. 1947. Vol. 40. № 4. P. 115.
Cristol D.A., Switzer P.V., Johnson K.L., Walke L.S. Crows do not use automobiles as nutcrackers: putting an anecdote to the test // Auk. 1997. Vol. 114. № 2. P. 296-298.
Davies N.B., Green R.F. The development and ecological significance of feeding techniques in the Reed Warbler (Acrocephalus scirpaceus) // Anim. Behav. 1976. Vol. 24. № 1. P. 213-229.
Fitzpatrick J.W. Foraging behavior of Neotropical Tyrant Flycatchers // Condor. 1980. Vol. 82. № 1. C. 43-57.
Fisher J., Hinde R.A. The opening of milk bottles by birds // British Birds. 1949. Vol. 42. № 11. P. 347-357
Gajdon G.K., Fijn N., Huber L. Limited spread of innovation in a wild parrot, the kea (Nestor notabilis) // Anim. Cogn. 2006. Vol. 9. № 3. P. 173-181.
Goethe F Black Terns feeding after ploughs // British Birds. 1970. Vol. 63. № 1. P. 34.
Greenhalgh M. Ruffs following a plough with Black-headed Gulls // British Birds. 1965. Vol. 58. № 7. P. 299.
Grobecker D.B., Pietsch T.W. Crows use automobiles as nutcrackers // Auk. 1978. Vol. 95. № 4. P. 760-761.
Jackson G. Ruffs following a plough // British Birds. 1973. Vol. 66. № 7. P. 311-312.
HaymanR.W. Persistent ground-feeding by Buzzards // British Birds. 1970. Vol. 63. № 3. P. 133.
Hinde R., Ficher J. Further observations on the opening of milk bottles by birds // British Birds. 1951. Vol. 44. № 12. P. 393-396.
HoulstonR. Kestrel and Grey Heron associating with plough // British Birds. 1978. Vol. 71. № 6. P. 270.
Hugh J. Carrion Crows taking fish from water // British Birds. 1955. Vol. 48. № 2. P. 91.
Hughes S. Carrion Crows submerging to catch fish // British Birds. 1976. Vol. 69. № 7. P. 273.
Maple T. Do crows use automobiles as nutcrackers? // Western Birds. 1974. Vol. 5. № 3. P. 97-98.
Mayer E. Purple Heron following the plough // British Birds. 2002. Vol. 95. № 2. P. 85-86.
Meier W. Buntspechte plundern Mehlschwalbennester // Vogelk. Hefte Edertal. 1993. № 19. P. 71-72.
Mitchell K.D.G. Green Woodpecker following plough // British Birds. 1948. Vol. 41. № 4. P. 120.
Naunton C.R. Unusual hunting behaviour of Sparrowhawks // British Birds. 1973. Vol. 66. № 9. P. 39-397.
Podmore R.E. Jackdaws hawking insects, with a note on flight behaviour // British Birds. 1948. Vol. 41. № 9. P. 272.
Remsen J.V.Jr., Robinson S.K. A classification scheme for foraging behavior of birds in terrestrial habitats // Studies in Avian Biol. 1990. № 13. P. 144-160.
Reynolds J.F Feeding habits of Redshank // British Birds. 1949. Vol. 42. № 7. P. 220.
Rhodes R.W Hen Harrier apparently attempting to catch fish // British Birds. 1979. Vol. 72. № 6. P. 289.
Roberts E.L. Carrion Crows taking fish from water // British Birds. 1955. Vol. 48. № 2. P. 91.
Stevenson P.P.L. Kestrel following plough // British Birds. 1948. Vol. 41. № 9. P. 279.
Tichon M. Peche specialisee d’une corneille noire (Corvus corone corone) // Aves. 1989. Vol. 26. № 1. P. 57.
Thompson C.F., Ray G.F., Preston R.L. Nectar robbing in Blue Tits Parus caeruleus failure of a novel feeding trait to spread // Ibis. 1996. Vol. 138. № 3. P. 552-553.
Trelfa G. Carrion Crow alighting on water and taking fish // British Birds. 1954. Vol. 47. № 11. P. 405-406.
TullyH. Carrion Crow taking fish // British Birds. 1951. Vol. 44. № 9. P. 323.
Waters E. Common Tern diving from bridge // British Birds. 1966. Vol. 59. № 8. P. 341.
|
