Особенности фенологического развития Batrachium kaufmannii (Ranunculaceae) в малых реках Ярославской области
О. А. Лебедева
Cреди многочисленных видов водных растений, активно распространяющихся в водотоках Ярославской области, шелковники занимают особое место в силу их высокой адаптивной пластичности. Обладая способностью к существованию в разных средах обитания (водной и наземной) в связи с динамикой уровня воды, а также развитым вегетативным размножением, растения быстро занимают новые местообитания, внедряются в искусственные водоемы, приспосабливаясь к другим условиям. Разрастаясь, заросли шелковника способны изолировать отдельные участки реки, снижая интенсивность водообмена, что постепенно приводит к их заболачиванию.
Шелковник Кауфмана (Batrachium kaufmannii) - полностью погруженный вегетативный малолетник, предпочитает глубины от 40 до 70 см, составляющие средний предел распространения макрофитной растительности в области (гидрофит), является холодостойким растением (микротермит), светолюбив (гелиофит). Встречается в реках, ручьях, в сравнительно быстро текущих водах (реофит), на перекатах с каменистыми грунтами, в виде куртин и различных по площади зарослей [6].
В литературных источниках [1, 5] еведения о сезонном развитии шелковника Кауфмана очень кратки и не освещают даже половины фенологических фаз.
В связи с этим целью исследования явилось изучение фенологии B. kaufmannii с учетом влияния различных абиотических факторов на сезонную ритмику растения в водотоках, находящихся под влиянием гидрологического режима Рыбинского водохранилища.
Материал и методы исследования
Материалом для исследования послужили фенологические наблюдения за ходом наступления основных фаз развития B. kaufmannii в течение сезонов вегетации 2009-2010 гг. в бассейне р. Ильдь, являющейся притоком второго порядка Рыбинского водохранилища (Ярославская область). Как и другие реки области, р. Ильдь по водному режиму относится к восточноевропейскому типу, который характеризуется ярко выраженным весенним половодьем, летнеосенней низкой меженью, прерываемой иногда паводками, и зимней меженью [11].
Стационарные исследования проводили на 2 станциях, расположенных по продольному профилю реки в зоне подпора, представляющие собой проточные участки с глубинами от 35 см (зона прибрежья) до 70 см (стрежневая зона). В
Реклама

процессе наблюдения за растениями отмечали даты наступления и окончания основных фенологических фаз. Растения для анализа отбирали подекадно в течение сезонов вегетации, одновременно отмечали глубину произрастания и придонную температуру. В лабораторных условиях у растений определяли основные морфометрические параметры вегетативной и генеративной сфер.
Методика обработки и проведение морфологического анализа описаны нами ранее [6]. Обработка данных проводилась с помощью пакета анализа данных MS Excel 2009. Характер суточного цветения изучали по методике А. Н. Пономарева [8], фенологические наблюдения проводили по методике И. Н. Бейдеман [2].
Результаты исследований
Фенологические фазы, согласно Р. Е. Левиной [7], представляют собой этапы индивидуального развития растений, фиксируемые по морфологическим признакам. От сроков наступления фаз, продолжительности вегетации и других важных биологических особенностей растений (рост, ветвление и др.) зависит их зимостойкость и наличие генеративной фазы. Причем, как считает ряд отечественных и зарубежных исследователей [4, 5], важным критерием развития растений в природных условиях является соответствие ритма биологического развития ритму сезонных явлений района произрастания.
Установлено, что для шелковника Кауфмана в водотоках Ярославской области характерно раннее начало вегетации. Весеннее отрастание для растений второго года жизни в бассейне р. Ильдь приходится на начало мая, что совпадает с переходом суточных температур через 5°С и окончанием половодья. Самое раннее начало вегетации отмечено 27.04.10 г., самое позднее - 14.05.09 г.
Если сравнить метеорологические условия этих лет, то среднемесячная температура в первой декаде мая 2010 г. была на 7-10°С выше, чем в 2009 г. (табл. 1).
Активация пазушных почек у перезимовавших побегов B. kaufmannii приводит к формированию одноосных, вегетативных, ассимилирующих побегов второго года жизни, которые, итеративно нарастая, успевают пройти за вегетационный сезон все фазы развития.
В отличие от изученных ранее видов [6] шел- ковник Кауфмана в водотоках Ярославской области формирует не прямостоячую (ортотропную), а стелющуюся (плагиотропную) биоморфу. Полегание сопровождается придаточным укоренением по всей длине побега и ярко выраженной анизоризией. Большинство корней детерминированные и способны скручиваться в спираль, что позволяет им (выполняя роль амортизатора) лучше фиксировать растение на перекате.
Реклама

Переход в генеративный период жизни у растений, по мнению Т. А. Работнова [9], зависит от многих экологических факторов (температурный и световой режимы, минеральное питание и т. д.). Образование генеративных структур (заложение и развитие цветочных почек) у шелков- ника Кауфмана связано с повышением температуры воды в среднем до 17,5°С.
После закладки репродуктивных почек моно- подиальное нарастание сменяется симподиаль- ным, и уже в конце мая каждая особь формирует сложную побеговую систему. В этой фазе у шел- ковника начинается процесс морфологической дезинтеграции (термин: Ценопопуляции, 1976 [10]), присущей большинству водных растений. МД у B. kaufmannii приводит не только к клональному возобновлению неспециализированными диаспорами, но и, снижая интенсивность физиологических процессов, не дает вытягиваться междоузлиям, тем самым позволяя растению существовать в особых условиях высокой проточности.
При снижении уровня воды в водотоках шел- ковник (как и другие реофиты) стремится использовать создавшиеся благоприятные условия для цветения. Следуя методике А. Н. Пономарева [8], фенологическое начало цветения определяется по зацветанию первого цветка, а окончание - по отцветанию последнего, однако у растений с растянутым периодом цветения установление окончания данного процесса представляет значительные трудности.
Примером такого вида служит B. kaufmannii, у которого из-за неограниченного роста соцветия на одной особи можно обнаружить бутоны, цветки и плоды разной степени зрелости. Массовое цветение у шелковника Кауфмана обычно начинается в первой декаде июня и продолжается до конца месяца. В этом периоде интенсивность увеличения морфометрических показателей приостанавливается, что обусловлено сменой фазы роста растения фазой цветения. Большинство морфологических показателей достигают своих предельных значений (рис. 1).
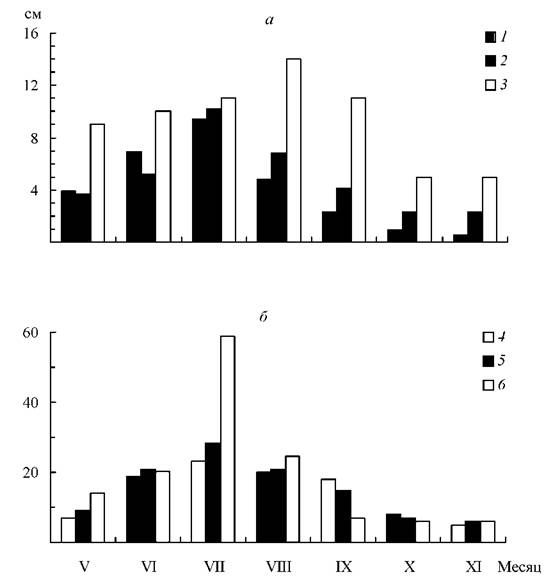
Рис. 1. Изменения средних за вегетационный период основных морфометрических показателей B. kaufmannii в водотоках Ярославской области. (а) - длина: 1 - междоузлия, 2 - листовой пластинки, 3 - придаточного корня, (б) - количество. Порядок остальных обозначений, как на рис. (а)
Для растений характерен дневной ритм раскрытия цветков, а интенсивность цветения зависит от количества солнечных дней. Фаза цветения в районе исследований совпадает с дальнейшим увеличением придонной температуры, устойчивым понижением уровня воды в реке (летняя межень) и, как следствие, значительным снижением проточности.
Период цветения у шелковника Кауфмана в 2009 г. пришелся на пасмурную и дождливую погоду, в результате чего его продолжительность составила от 20 до 25 суток, тогда как в 2010 г. с жарким и засушливым летом - всего 10 суток.
Во второй декаде июня растение вступает в фазу плодоношения и диссеминации. Одной из особенностей шелковника Кауфмана в водотоках области является неустойчивая семенная продуктивность, формирование большого количества орешков с одними покровами (без эндосперма и зародыша), а также редко встречающиеся всходы. Вероятно, особые условия произрастания шелковника Кауфмана (перекаты), создают определенные сложности с опылением, а также вынуждают растение тратить большие энергетические ресурсы на поддержание вегетативной сферы, вследствие чего падает семенная продуктивность.
Календарные сроки наступления фазы плодоношения у B. kaufmannii в большинстве случаев приходятся на 20 июня, а окончание может растянуться до середины августа.
С конца августа до начала сентября в реке наблюдается постепенное понижение придонной температуры до 7°С. Похолодание и ночные заморозки почти не отражаются на габитусе растений, а морфологические перестройки побега осенней формы шелковника касаются лишь формирования укороченных междоузлий верхушек вегетативных побегов.
Необходимо отметить, что у B. kaufmannii к этому времени (в отличие от B. circinatum и B. trichophyllum) не наблюдается полная МД и только незначительная часть побеговой системы распадается на фрагменты. Единственным свидетельством о переходе растения от активного состояния к покою является уменьшение емкости пазушных почек.
Признаки подготовки шелковника Кауфмана к зиме становятся более заметными лишь в конце октября, когда постепенное преобладание деструктивных процессов приводит к потере большей части надземной сферы растения. Для зимующего побега B. kaufmannii характерно формирование сближенных до 0,5-0,9 см междоузлий, 7-9 листьев, имеющих длину от 1,5 до 2,3 см и появление контрактильных корней, втягивающих базальную часть побега в каменистый грунт. В таком состоянии растение вступает в фазу относительного (ростового) покоя и минимальных морфометрических показателей (рис. 1).
Обсуждение результатов
Продолжительность периода от начала вегетации до массового созревания семян у B. kauf- mannii в водотоках Ярославской области составляет около 65-70 суток, что вполне вписывается в вегетационный период Центральных районов Нечерноземной зоны [12]. Сроки наступления основных фенологических фаз могут варьироваться, их амплитуды по итогам наблюдений могут достигать от 7 до 25 суток (табл. 1).
Таблица1
Фазы развития B. kaufmannii в водотоках Ярославской области (2009-2010 гг.)
| Фенологическая фаза
|
Дата
|
Амплитуда, количество дней
|
| min
|
max
|
| Начало вегетации
|
27.04.2010
|
14.05.2009
|
18
|
| Начало ветвления
|
20.05.2010
|
27.05.2009
|
7
|
| Начало бутонизации
|
29.05.2010
|
10.06.2009
|
11
|
| Начало цветения
|
|
|
|
| Созревание семян
|
10.06.2010
|
19.06.2009
|
10
|
| Окончание вегетации
|
20.06.2010
|
14.07.2009
|
25
|
| |
10.10.2009
|
20.10.2010
|
|
Весеннее развитие шелковника, выраженное в увеличении длины междоузлий, определяется наступлением положительных температур и окончанием весеннего половодья. Максимум морфологических показателей (изменения которых в течение вегетационного сезона идут по одновершинной кривой) наблюдается в середине июля, совпадая с продолжительной летней меженью и температурой воды > 20°С. Жаркие, солнечные дни сокращают период цветения (2010 г.), а длительные осадки при пониженной температуре (в том числе туманы) увеличивают (2009 г.). Реальная семенная продуктивность также определяется метеорологическими условиями в этом периоде. Поскольку вид является энтомофильным, то ненастье, препятствуя опылению, может снижать ее в несколько раз.
Прогрев воды > 25°С при низком ее уровне, приводит к выпадению фаз цветения, отмиранию надземной сферы шелковника и нарушению структурысообщества.Охлаждение дна р. Ильдь, как показали промеры придонных температур, начинается в августе. Первоначально идет равномерное понижение температуры воды по всем станциям, затем, в стрежневой части реки придонная температура воды оказывается на 1-2°С выше, чем на мелководье. Это обстоятельство, вместе с высокой проточностью и большим содержанием кислорода, позволяет шелковнику продолжать вегетацию на глубоководных станциях до наступления заморозков.
По характеру фенологического развития в годичном цикле B. kaufmanii следует отнести к ве- сеннее-летне-осеннезеленому типу [3] с зимним периодом относительного (ростового) покоя. Фенологические фазы растянуты и могут сдвигаться в разные годы в зависимости от климатических условий и уровенного режима водоема. В водотоках Ярославской области у B. kaufmanii существует вероятность выпадения фазы созревания семян, поэтому данный вид размножается преимущественно вегетативным путем.
Список литературы
1. Барыкина, Р. П. Особенности структуры развития водных лютиков [Текст] / Р. П. Барыкина. - Бюл. МОИП. Отд. биол. - 1988. - Т. 93, Вып. 2. - С. 134144.
2. Бейдеман, И. Н. Методика изучения фенологии растений и растительных сообществ [Текст] / И. Н. Бейдеман. - Новосибирск : Наука. 1974. - 156 с.
3. Борисова, И. В. Сезонная динамика растительного сообщества. [Текст] / И. В. Борисова. Полевая геоботаника. - Л. : Наука, 1972. - Т. 4. - С. 5-94.
4. Ворошилов, В. Н. Ритм развития у растений. [Текст] / В. Н. Ворошилов. - М. : Изд-во АН СССР, 1960. - 135 с.
5. Дубына, Д. В. Макрофиты - индикаторы изменения природной среды [Текст] / Д. В. Дубына. - Киев : Наук. думка, 1993. - 428 с.
6. Лебедева, О. А., Лапиров, А. Г. Ритм сезонного развития и морфологическая поливариантность Batrachium circinatum (Sibth.) Spach на Рыбинском водохранилище [Текст] / О. А. Лебедева, А. Г. Лапиров // Биология внутр. вод. - 2009. - №3. - С. 36-40.
7. Левина, Р. Е. Репродуктивная биология семенных растений. Обзор проблемы [Текст] / Р. Е. Левина. - М. : Наука, 1981. - 96 с.
8. Пономарев, А. Н. Цветение и опыление злаков [Текст] // А. Н. Пономарев // Учен. записки Перм. унта. - 1964. - Т. 39, № 5. - С. 706-720.
9. Работнов, Е. А. Вопросы изучения состава популяций для целей фитоценологии [Текст] // Е. А. Работнов. Проблемы ботаники. - М. ; Л. - 1950. Вып.1. - С. 465-483.
10. Смирнова, О. В., Заугольнова, Л. Б., Торопо- ва, Н. А. Критерии выделения возрастных состояний и особенности хода онтогенеза у растений различных биоморф [Текст] / О. В. Смирнова, Л. Б. Заугольнова,
Н.А. Торопова // Ценопопуляции растений (основные понятия и структура). - М. : 1976. - С. 14-43.
11. Экосистема малой реки в измененных условиях среды [Текст]. - М. : Товарищество научных изданий КМК, 2007. - 372 с.
12. Физико-географическое районирование Нечерноземного центра [Текст]. - М. : Изд-во МГУ, 1963. -184 с.
|
