Молекулярная эволюция в подсемействе Prunoideae Focke
В. И. Авдеев
Крупное подсемейство сливовых (Prunoideae Focke), как и остальные 4 подсемейства Rosaceae Juss., слабо изучены в эволюционном плане, хотя и являются очень ценными растениями. До конца не ясен даже родовой состав подсемейства, генетические связи родов и видов, произрастающих на разных континентах. Биосистематика сливовых, как и большинства других растений, построена на анализе внешних признаков. В этом случае вне поля зрения остаются молекулярные механизмы эволюции, в основе которых лежат явления рецессивности, доминантности, репрессии, дерепрессии генов и др. Поэтому у живых организмов при однородности внешних признаков весьма обычно значительное разнообразие особей, видов, родов по молекулярному составу, биохимическим признакам [1].
Попытки создания системы подсемейства сливовых предпринимались, но они имели чисто практический характер, для обслуживания селекционных нужд [2]. Это были формальные схемы, где не учитывались палеоданные, история флор и ареалов родов, даже полиморфизм популяций по внешним признакам. С учётом всего этого и использованием новейших экспериментальных данных по белковым (полипептидным, молекулярным) маркёрам впервые эволюция подсемейства сливовых была охарактеризована в докторской диссертации и статье [3, 4]. Однако на то время ещё не были уточнены молекулярные критерии эволюционного возраста таксонов, это было сделано уже позднее [5, 6] на основе разработок школы А. В. Благовещенского [7]. В итоге связи по белковым маркёрам внутри родов сливовых и межродовые отношения в процессе их эволюции представлены на дендрограммах (рис. 1 и 2) и разъяснены в описаниях этих рисунков. Краткие же комментарии к этим данным будут нижеследующими.
Древнейшими (верхнемелового возраста) были роды Cerasus Mill. и Prunus L. Различия по белковым маркёрам этих и всех остальных таксонов столь велики, что попытки объединить их в единый род Prunus (слива) не являются целесообразными. В роде Prunus в течение третичного периода вымерли в Евразии сначала древнейшие узколистные виды (некоторые из них, мигрировав в Северную Америку через древнюю Североатлантическую сушу, сохранились), затем широколистные виды. К числу древних видов сливы, возникших в тургайских лесах Евразии, относится тёрн (P spinosa L.). Попытки считать его очень молодым гибридным видом [2] — ошибочны. Древним потомком Prunus является род Louiseania Carr. Виды сливы из Восточной Азии и многие североамериканские — молодые виды (рис. 1А).
Реклама

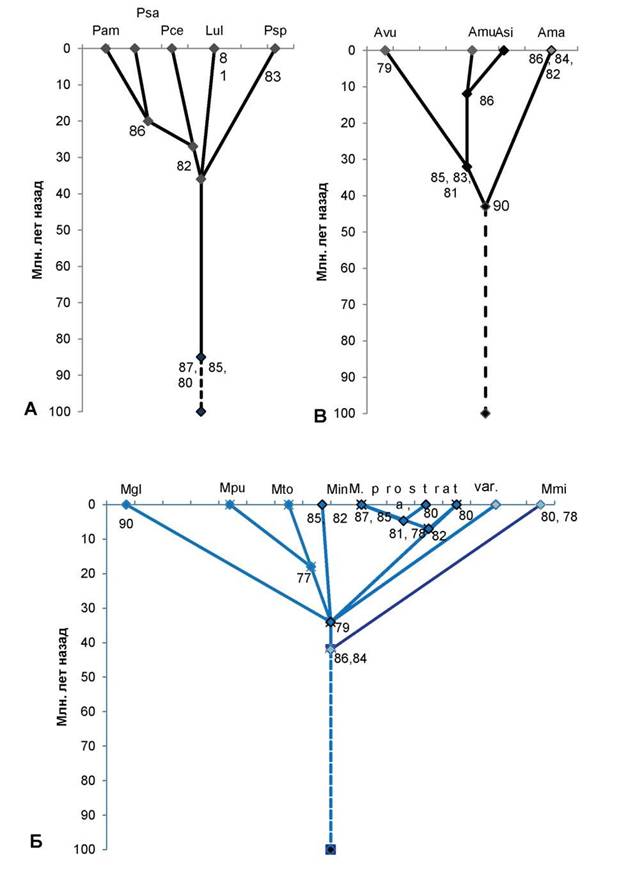
Рис. 1. Схема таксоногенеза внутри родов подсемейства Prunoideae Focke по основным 12S-полипептидам запасных белков-глобулинов семян
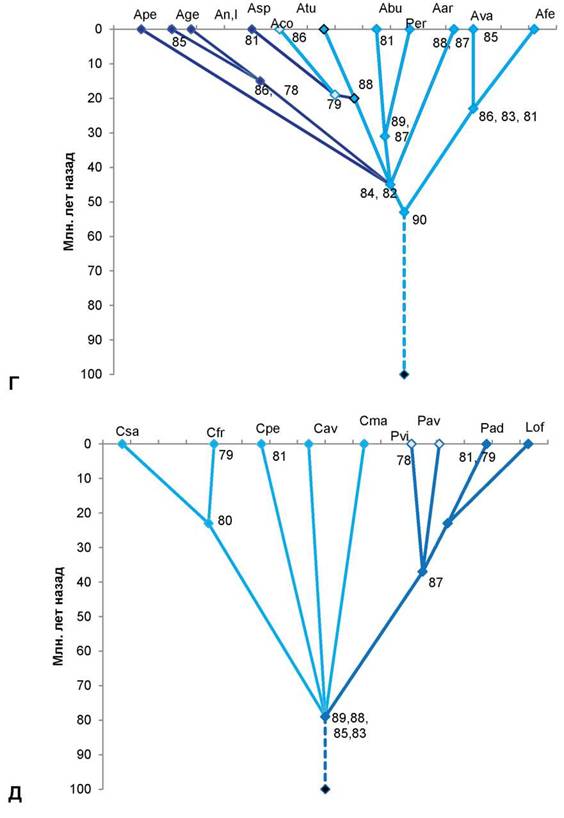
Условные обозначения:
А — род слива (Prunus L.): Pam — слива американская (Prunus americana Marsh.); Psa — слива ивовидная, или китайская (Prunus salicina Lindl.); Pce — слива вишневидная, или алыча (Prunus cerasifera Ehrh.); Psp — слива колючая, или тёрн (Prunus spinosa L.); Lul — луизеания (афлатуния) вязолистная [Lou- iseania ulmifolia (Franch.) Pachom.], или Aflatunia ulmifolia (Franch.) Vass. Оба рода выделяются общими древними полипептидными компонентами 87, 85, 80, алыча, сливы ивовидная и американская — молодым
компонентом 82, последние 2 вида сливы — также компонентом 86. Древний тёрн выделяется компонентом 83, а древняя луизеания вязолистная выделяется компонентом 81.
Б — род микровишня (Microcerasus Webb): Mgl — микровишня железистая [Microcerasus glandu- losa (Thunb.) M. Roem.]; Mpu — микровишня низкая [Microcerasus pumila (L.) Erem. et Yushev]; Mto — микровишня войлочная [Microcerasus tomentosa (Thunb.) Erem. et Yushev]; микровишня седая разновидность Блиновского [Microcerasus incana var. blinovskii (Totsch.) Erem. et Yushev]; далее слева направо: микровишня простёртая разновидность двусторонняя [Microcerasus prostrata var. bifrons (Fritsch) Erem. et Yushev], микровишня простёртая разновидность тяньшанская [Microcerasus prostrata var. tianschanica (Po- jark.) Erem. et Yushev], микровишня простёртая разновидность бородавчатая [Microcerasus prostrata var. verrucosa (Franch.) Erem. et Yushev], представленная двумя популяциями — слева типа микровишни красноплодной, справа — типичная микровишня бородавчатая; микровишня мелкоплодная [Microcerasus microcarpa (C. A. Mey.) Erem. et Yushev]. Древние общие компоненты 86, 84, моложе — общий компонент 79. Микровишня железистая выделяется древним компонентом 90, микровишни низкая и войлочная — более молодым компонентом 77, микровишня седая — компонентами 85, 82. Микровишня двусторонняя имеет компоненты 87, 85, тяньшанская — компонент 80 (обе они объединяются компонентами 81, 78), микровишня бородавчатая (типа красноплодной) имеет компонент 80 (объединяясь с предыдущими двумя разновидностями компонентом 82), микровишня бородавчатая (типичная) — самая древняя разновидность, микровишня мелкоплодная — древняя, имеет компоненты 80, 78.
Реклама

В — род абрикос (Armeniaca Scop.): Avu — абрикос обыкновенный (Armeniaca vulgaris Lam.); Amu — абрикос муме (Armeniaca mume Sieb.); Asi — абрикос сибирский [Armeniaca sibirica (L.) Lam.]; Ama — абрикос маньчжурский [Armeniaca mandshurica (Maxim.) Skvortz.]. Общий древний компонент — 90, абрикосы обыкновенный, муме, сибирский выделяются компонентами 85, 83, 81, из них последние 2 вида — компонентом 86, древний абрикос маньчжурский имеет компоненты 86, 84 и 82.
Г — род миндаль (Amygdalus L.): древний компонент 90 дал начало двум ветвям рода — малая ветвь (справа) и большая ветвь (слева). В правой ветви — 2 вида с общими древними компонентами 86, 83, 81, один из нихAfe — миндаль Фенцля [Amygdalusfenzliana (Fritsh) Lipsky]; другой Ava — миндаль Вавилова (Amygdalus vavilovii M. Pop) с компонентом 85. В левой ветви общими древними компонентами являются компоненты 84, 82. Они дали начало Ape — миндалю Петунникова (Amygdalus petunnicovii Litv.); Age — миндалю грузинскому (Amygdalus georgica Desf); An — миндалю низкому (Amygdalus nana L.) и очень близкому к нему Al — миндалю Ледебура (Amygdalus ledebouriana Schlecht.). Последние 3 вида миндаля имеют общий компонент 86, а миндаль грузинский выделяется компонентом 85. Asp—миндаль колючейший (Amygdalus spinosisssima Bunge) с компонентом 81; Atu — миндаль туркменский (Amygdalus turcomanica Lincz.) с компонентом 86, оба миндаля имеют общий компонент 79; Aco — миндаль обыкновенный (Amygdalus communis L.) имеет с миндалями туркменским и колючейшим общий древний компонент 88. Древние компоненты 89, 87 дали начало Abu — миндалю бухарскому (Amygdalus bucharica Korsh.) с компонентом 81 и Aar — миндалю арабскому (Amygdalus arabica Olivier); компонентами 88, 87 выделяется древний род Per — персик (Persica Mill.).
Д — роды вишня (Cerasus Mill.), черёмуха (Padus Mill.), паделлус (Padellus Vass.), лавровишня (Lau- rocerasus Duham.) с общими древними компонентами 89, 88, 85, 83. Csa — вишня сахалинская [Cerasus sachalinensis (Fr. Schmidt) Kom.]; компонентом 79 выделяется Cfr — вишня кустарниковая, или степная (Cerasus fruticosa Pall.), оба этих вида объединяет компонент 80; Cpe — вишня пенсильванская [Cerasus pennsylvanica (L.f.) Loisel.] с компонентом 81; Cav — вишня птичья, или черешня [Cerasus avium (L.) Mo- ench]; Cma — вишня Максимовича [Cerasus maximowiczii (Rupr.) Kom.]; роды с общим компонентом 87: Pav — черёмуха птичья (Padus avium Mill.) с компонентом 78; Pvi — черёмуха виргинская [Padus virginiana (L.) M. Roem.]; Pad — паделлус, магалебка, или антипка [Padellus mahaleb (L.) Vass.] с компонентами 81, 79; Lof— лавровишня лекарственная (Laurocerasus officinalisM. Roem.). Номера в узлах схемы — основные полипептидные электрофоретические компоненты 12S-глобулинов. По вертикальной оси нанесена (в миллионах лет назад) общепринятая в геологии магнитохронологическая шкала [8], где отрезок времени до 65 млн. лет назад — конец мелового периода; 65—54 млн. лет назад — эпоха палеоцена, 54—38 млн. лет назад — эпоха эоцена, 38—26 млн. лет назад — эпоха олигоцена, 26—8 млн. лет назад — эпоха миоцена (третичный период), 8—2 млн. лет назад — эпоха плиоцена; 1,5—2 млн. лет назад и до наших дней — эпоха плейстоцена (четвертичный период). Сплошные линии на схеме — установленные эволюционные связи, прерывистые линии — связи с неизвестными предками.
В роде Cerasus на территории Евразии, Северной Америки растут древнейшие виды (типа черешни, вишен Максимовича, пенсильванской). Но вишня сахалинская, как и степная, лесостепная вишня кустарниковая, являются молодыми видами. Род Padus возник от какого-то вымершего рода вишни во вторичном (урало-западносибирском)
центре происхождения вишен, а роды Padellus (магалебка) и особенно род Lauroceresus, продолжающие линию древних вишен, — наиболее молодые роды (рис. 1Д).
Остальные роды сливовых более молодые, возникли в эпохи палеогена. В роде Amyg- dalus (миндаль) издревле развивались две линии, из которых древнейшим был предок миндалей с бороздчатым эндокарпием (типа миндаля Вавилова, Фенцля и др.). Мнение о культигенном и гибридном происхождении миндаля Вавилова [2] — также ошибочное [3]. К древним миндалям второй линии относятся миндали, растущие в степной зоне (в том числе A. nana L.), их предок возник одновременно с родом Persica (персик). Персик расселялся в лесах Евразии, вымер, сохранившись в основном на востоке Азии. Из миндалей этой второй линии очень древним является миндаль Петунникова, который сохранился в горах на севере Средней Азии. Виды миндаля из районов Передней Азии, юга Средней Азии — сравнительно молодые, но старше северных степных миндалей (рис. 1Г).
Из рода микровишня (ошибочно относимого к вишне) древнейшими являются виды из западной части Евразии, в особенности микровишня мелкоплодная, имеющая резко реликтовый ареал в Передней Азии. Виды востока Евразии, Северной Америки — более молодые, но сохранили в себе признаки древних лесных видов Евразии (рис. 1Б). В Средней, особенно в Передней Азии между видами микровишни идут интенсивные процессы гибридизации, приводящие к взаимной интрогрессии части их геномов, но на автополиплоидной основе [3].
Много спорного было в возрасте и истории рода Armeniaca (абрикос). Древнейшим видом является абрикос маньчжурский с реликтовым типом ареала. Более молодой вид — абрикос обыкновенный, самые молодые — абрикос сибирский (с крупнейшим ареалом) и абрикос муме (или китайский) с ареалом на юге Восточной Азии (рис. 1А).
Общего предка подсемейства Prunoideae установить пока сложно. Не изучены по белковым маркёрам род Emplectocladus из Северной Америки, ряд родов, близких к вишне, черёмухе и растущих в Восточной Азии. Однако метод белковых маркёров показывает, что центром происхождения родов сливовых являются древние Тургайская и Древнесредиземноморская области Евразии в пределах современной западной (а не восточной) части Евразии [3, 4]. Учитывая, что древнейшими таксонообразующими белковыми маркёрами у двудольных растений являются низкомолекулярные полипептиды основных 128-глобулинов [5, 6], можно полагать, что предок был ближе к роду Cerasus. При формировании подсемейства сливовых самостоятельной была линия, давшая начало родам слива (с луизеанией), вишня (с черёмухой, паделлусом, лавровишней). В другой линии сливовых выделились миндали древнейшего происхождения (типа миндаля Вавилова), затем только предковая линия дивергировала на «абрикосовую» и предка родов микровишня, персик и остальных видов миндаля (рис. 2).
Таким образом, род миндаль имеет сборное происхождение, и это мнение часто высказывалось в литературе. Интересно, что данные по белковой эволюции подтверждают данные по анатомии листа сливовых [9] в части того, что род абрикос, хотя и внешне близок к сливе (и с нею частично гибридизирует), но родственен древней «миндальной» ветви.
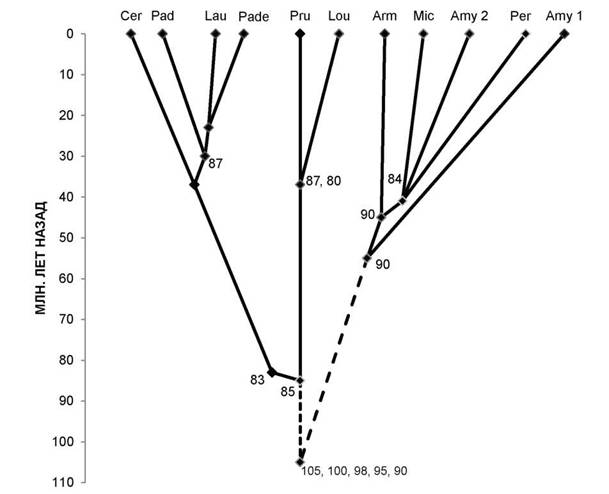
Рис. 2. Схема таксоногенеза родов подсемейства Prunoideae Focke по 12S-полипептидам. Условные обозначения: Cer — род Cerasus Mill. (вишня); Pad — PadusMill. (черёмуха); Lau — Laurocerasus Duham. (лавровишня); Pade — Padellus (L.) M. Roem. (паделлус, магалебка); Pru — Prunus L. (слива); Lou — Lou- iseania Carr, (луизеания), или Aflatunia Vass. (афлатуния); Arm — Armeniaca Scop. (абрикос); Mic — Micro- cerasus Webb (микровишня); Amy 2 — виды Amygdalus L. (миндаль, большая ветвь, см. рис. 1); Amy 1 — то же, но малая ветвь; Per — Persica Mill. (персик). Клада «вишнеслив» маркируется древним компонентом 85, при этом «вишнёвые» роды — компонентом 83, а роды слива и луизеания — молодыми компонентами 87 и 80 (компонент 87 маркирует также паделлус, черёмуху, лавровишню). Клада «абрикос, микровишня, персик, миндаль» возникла много позже на основе древнего компонента 90, а позднее — более молодого компонента 84.
На рисунках 1 и 2 существенно уточнён возраст ряда таксонов, известный в литературе (см. [7, с. 92]). Цифрами указаны полипептидные компоненты разной степени древности; сплошные линии — установленные эволюционные связи, прерывистые — связи с неизвестными предками. Остальные обозначения аналогичны рисунку 1.
Список литературы
Кимура М. Молекулярная эволюция: теория нейтральности. М. : Мир, 1985. 400 с.
Ерёмин Г. В. Отдалённая гибридизация косточковых плодовых растений. М. : Колос, 1985. 280 с.
Авдеев В. И. Плодовые растения Средней Азии, их происхождение, классификация, исходный материал для селекции : дис. ... д-ра с.-х. наук. СПб. : ВНИИР им. Н. И. Вавилова, 1997. 328 с.
Авдеев В. И. К истории происхождения видов подсемейства сливовых (Rosaceae) в степной зоне // Тр. Ин-та биоресурсов и прикладной экологии. Оренбург : ОГПУ, 2000. Вып. 1. С. 45—58.
Авдеев В. И. Этапы формирования степных ландшафтов в Евразии. Аспекты эволюции видов Poa- ceae // Известия ОГАУ 2009. № 2. С. 59—65.
Авдеев В. И., Саудабаева А. Ж. Сравнительный анализ адаптаций по полипептидным маркёрам у абрикосов Оренбуржья // Приёмы повышения адаптивности косточковых культур, вопросы осеверения и расширения границ садоводства : сб. материалов междунар. симпозиума. Челябинск : НПО «Сад и огород», 2011. С. 45—51.
Благовещенский А. В., Александрова Е. Г. Биохимические основы филогении высших растений. М. : Наука, 1974. 104 с.
Сузюмов А. Е. Земной магнетизм и тектоника дна океанов // Новое в жизни, науке, технике. М. : Знание, 1981. № 1. 48 с.
Соколова Е. А. Значение анатомических признаков для систематики представителей подсемейства Prunoideae (Rosaceae) : автореф. дис. ... д-ра биол. наук. СПб. : ВНИИР им. Н. И. Вавилова, 2000. 28 с.
|
