Белковые маркёры у ряда культивируемых злаков в Оренбуржье
В. И. Авдеев
А. Ж. Саудабаева
Метод маркирования фитогенофонда с помощью запасных белков семян (пролами- нов у злаков, глобулинов у двудольных растений) издавна и широко используется в растениеводстве, ботанике, генетике. Как арбитражный метод он рекомендован Международной ассоциацией по семенному контролю (ISTA).
Этот метод наиболее хорошо разработан на культивируемых злаковых растениях, имеющих большое экономическое значение. По многим из них (пшеница, ячмень и др.) создан соответствующий банк данных, но по целому ряду их сортов эти данные отсутствуют. В мире слабо изучены молекулярные маркёры проса. Так, попытки различить на основе ДНК-маркёров (RAPD, ISSR) культивируемые и сорные образцы проса самого различного географического происхождения (Азия, Европа, Австралия и др.) не дали результатов [1]. Во многом такие неудачи связаны с целым рядом недостатков всех методов ДНК-маркирования
. Однако не было и разработанной методики экстрагирования и электрофореза запасных белков проса (каферинов), так что этот пробел необходимо было восполнять в процессе проведения исследований. Для этого были использованы общепринятые методики выделения, вертикального гель-электрофореза белков, регистрации их полипептидных спектров
. Методом электрофореза запасных белков семян изучены 6 сортов ячменя, 7 сортов пшеницы и 2 образца сорного и культивируемого проса (табл. 1 и 2; рис. 1 и 2).
Ячмень. Из данных таблицы 1 видно, что изученные сорта ячменя являются крайне невыравненными по составу гордеинов — запасных белков-проламинов ячменя. Другими словами, внешне они выглядят как отдельные морфологические сорта, но по спектрам проламинов представляют собой, в сущности, сортосмеси, состоящие из отдельных биотипов. При этом наиболее смешанным является сорт Анна, менее смешанными — сорта Донецкий 8, Натали, Оренбургский Совместный. Среди спектров сорта Анна обнаружен спектр, близкий к сорту Донецкий 8. Исключение составил монотипный по спектрам сорт Оренбургский 17. Явление смешанного состава сортов давно установлено на разных сортах ячменя, пшеницы, ржи, злаковых и бобовых кормовых травах, огурца и других растений, при этом внутри отдельных сортов выявлено 2—20 и более биотипов каждого сорта [4]. Однако из ряда современных представлений [2] можно заключить, что такие естественные сортосмеси являются важными адаптациями самих сортов.
Реклама

Из наших же данных следует, что в спектрах гордеинов можно насчитать от 11 (сорт Анна) и до 15 компонентов полипептидов (сорт Натали, Донецкий 8), при этом на долю более высокомолекулярных («тяжёлых», ю-полипептидов) компонентов приходится менее 50% от всего их состава. Сорта различаются по всем зонам полипептидов (ю, у, в, а), по зоне же «у» схожи только сорта Анна и Оренбургский Совместный, отчасти сорт Скарлетт. Больше всех отличается сорт Оренбургский 17. Сорта Натали и Скарлетт имеют довольно яркие (2 балла) компоненты ю-полипептидов в позиции 9t (табл. 1).
Таблица 1
Формулы гордеина сортов ячменя
| |
Состав и интенсивность полипептидов четырёх фракций гордеина, балл
|
| Сорт, тип спектра
|
а
|
|
|
Y
|
ю
|
| 32
|
42
|
52
|
62
|
72
|
3i
|
32
|
4i
|
42
|
5i
|
52
|
22
|
23
|
52
|
43
|
22
|
4i
|
42
|
43
|
52
|
61
|
62
|
63
|
72
|
8i
|
82
|
9i
|
| Анна, первый тип
|
|
|
1
|
3
|
1
|
|
|
|
2
|
2
|
2
|
|
|
1
|
|
|
|
|
3
|
|
3
|
|
|
3
|
|
3
|
|
| Анна, второй тип
|
1
|
3
|
|
1
|
1
|
1
|
|
1
|
|
1
|
3
|
|
|
1
|
|
|
|
|
3
|
|
3
|
|
|
3
|
|
3
|
|
| Донецкий 8, первый тип
|
1
|
2
|
|
1
|
|
1
|
|
1
|
|
1
|
3
|
|
|
1
|
2
|
|
|
3
|
1
|
|
|
3
|
|
1
|
3
|
|
|
| Донецкий 8, второй тип
|
2
|
3
|
|
2
|
|
2
|
|
2
|
|
1
|
3
|
|
|
1
|
2
|
|
|
1
|
3
|
|
|
3
|
|
|
3
|
|
|
| Донецкий 8, третий тип
|
2
|
2
|
|
1
|
|
2
|
|
2
|
|
1
|
3
|
|
|
1
|
2
|
|
|
1
|
3
|
|
|
3
|
|
|
3
|
|
|
| Натали, первый тип
|
|
|
2
|
3
|
3
|
|
|
1
|
2
|
1
|
|
|
|
2
|
|
|
|
3
|
|
|
1
|
3
|
|
1
|
3
|
|
2
|
| Натали, второй тип
|
1
|
|
3
|
3
|
1
|
1
|
|
1
|
2
|
|
1
|
|
|
2
|
|
|
|
3
|
|
|
1
|
|
3
|
3
|
3
|
|
2
|
| Скарлетт, стандарт (st)
|
|
|
1
|
2
|
2
|
|
|
1
|
2
|
|
1
|
|
|
2
|
|
|
|
3
|
|
|
2
|
2
|
|
3
|
3
|
|
2
|
| Оренбургский Совместный, первый тип
|
|
1
|
|
3
|
1
|
|
|
1
|
2
|
|
2
|
|
|
1
|
|
|
3
|
|
|
1
|
|
3
|
1
|
3
|
|
2
|
|
| Оренбургский Совместный, второй тип
|
|
1
|
|
3
|
2
|
|
|
1
|
2
|
|
2
|
|
|
1
|
|
|
3
|
|
|
3
|
|
|
3
|
3
|
|
2
|
|
| Оренбургский 17
|
1
|
2
|
|
1
|
|
|
1
|
|
1
|
1
|
2
|
2
|
2
|
|
|
3
|
|
|
3
|
|
|
3
|
|
|
2
|
|
|
Таблица 2
Формулы глиадина сортов пшеницы и каферина сорго
| Сорт
|
Состав и интенсивность полипептидов глиадина, балл
|
| а
|
в
|
Y
|
ю
|
| 12
|
22
|
42
|
52
|
53
|
6i
|
62
|
7i
|
12
|
22
|
32
|
42
|
52
|
22
|
23
|
32
|
42
|
52
|
22
|
32
|
42
|
52
|
6i
|
62
|
63
|
72
|
8i
|
9i
|
93
|
i02
|
| Мягкие гексаплоидные пшеницы
|
| Мироновская 808 (st)
|
|
1
|
1
|
2
|
|
2
|
2
|
3
|
1
|
3
|
2
|
3
|
3
|
2
|
2
|
3
|
3
|
1
|
1
|
|
2
|
|
|
3
|
1
|
1
|
1
|
2
|
2
|
2
|
| Саратовская 90
|
1
|
2
|
2
|
2
|
|
2
|
|
3
|
|
3
|
2
|
3
|
3
|
3
|
3
|
3
|
1
|
1
|
2
|
|
1
|
|
3
|
1
|
|
1
|
2
|
2
|
2
|
2
|
| Пионерская 32
|
|
|
|
3
|
|
2
|
|
3
|
3
|
3
|
3
|
3
|
3
|
|
3
|
3
|
3
|
2
|
2
|
2
|
3
|
|
|
|
3
|
3
|
3
|
|
3
|
1
|
| Оренбургская 105
|
|
1
|
1
|
|
|
3
|
|
3
|
|
|
3
|
|
3
|
|
3
|
3
|
3
|
1
|
1
|
1
|
1
|
|
|
3
|
1
|
1
|
3
|
1
|
1
|
1
|
| Колос Оренбуржья
|
|
|
|
|
2
|
2
|
|
3
|
|
3
|
3
|
3
|
3
|
1
|
3
|
3
|
3
|
1
|
1
|
|
1
|
|
|
3
|
2
|
2
|
2
|
2
|
2
|
1
|
| Твёрдые тет
|
раплоидные пшеницы
|
| Гордеиформе (st)
|
1
|
1
|
|
2
|
2
|
|
3
|
|
3
|
3
|
3
|
3
|
3
|
|
1
|
|
1
|
3
|
3
|
2
|
2
|
2
|
3
|
3
|
2
|
3
|
|
|
|
|
| Мелинопус 6641
|
|
|
3
|
|
|
3
|
2
|
2
|
|
3
|
3
|
2
|
3
|
3
|
3
|
|
|
3
|
3
|
|
3
|
|
|
3
|
|
|
|
|
|
|
| Просо сорное и культивируемое
|
1
|
1
|
|
2
|
|
3
|
3
|
3
|
|
3
|
2
|
3
|
3
|
3
|
3
|
3
|
3
|
2
|
|
|
|
|
|
3
|
1
|
2
|
1
|
2
|
2
|
2
|
Примечание. Жирным шрифтом выделены сорта Мироновская 808, Саратовская 90, Оренбургская 105, имеющие блок полипептидов, который может быть связан с повышенной морозостойкостью сортов пшеницы (см. текст ниже).
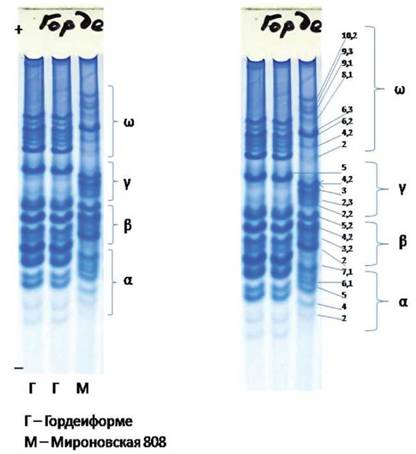
Рис. 1. Типовые проламиновые спектры запасных белков семян сортов ячменя и пшеницы (слева — 4 зоны полипептидов, справа — 4 зоны и позиции полипептидов по шкале)
Сорт ячменя Оренбургский 17
Сорт пшеницы Пионерская 32
Рис. 2. Некоторые проламиновые спектры изученных сортов ячменя и пшеницы (М — метчик молекулярных масс полипептидов, сорт пшеницы Мироновская 808)
Реклама

Известно, что зону высокомолекулярных компонентов (ю, у) кодируют сразу три группы генов (A, C, D). При этом рекомбинации затрагивают гены А (ю-полипептиды) и гены В (у- и в-полипептиды), тогда как гены, кодирующие зону низкомолекулярных («лёгких», а-полипептидов) компонентов, одинаковы, в рекомбинациях не участвуют [5]. Поскольку
же в зоне а-полипептидов имеются существенные сортовые различия (табл. 1), то можно сделать выводы, что, во-первых, гены этой зоны имеют важное значение для выживания (адаптации) генотипов (сортов, форм) ячменя и что, во-вторых, различия генотипов по зоне а-полипептидов определяются мутациями генов (возможно, не только генными, но и хромосомными). Сказанное выше подтверждает нашу гипотезу о том, что именно мутации в зоне а-полипептидов являются магистральным и молодым направлением в эволюции злаков [6].
Пшеница. Изученные проламиновые спектры сортов пшеницы (глиадины) однотипны и за счёт всех зон полипептидов более насыщены компонентами, чем спектры сортов ячменя. Из сортов пшеницы наиболее бедный спектр характерен для сорта Мелинопус 6641 (14 компонентов), у остальных сортов — по 20—25 компонентов. Сорта пшеницы также хорошо различаются, но более схожи по всем зонам полипептидов сорта Саратовская 90 и Колос Оренбуржья. Сорта твёрдой пшеницы Гордеиформе и Мелинопус 6641 выделяются отсутствием компонента 32 (у-полипептиды) и компонентов 9 9 1 02 и других компонентов у ю-полипептидов (табл. 2). Отметим, что в данных Ю. В. Перуанского, Е. В. Кожемякина (цит. по: [7, с. 216]) у пшеницы во всех зонах явно занижено число компонентов и не приведено разделение одних и тех же компонентов спектра (например, компонент 9 из зоны ю-полипептидов не был разделён на субкомпоненты 9 93). Таким образом, наши данные существенно уточняют такого рода исследования. В связи с этим хочется выразить мнение, что применяемая во ВНИИР им. Н. И. Вавилова методика написания формул проламинов злаков более прогрессивна, поскольку позволяет выявить более тонкие различия между изучаемыми сортами, формами, видами.
В молекулярном маркировании особенную трудность представляет выявление связи между полипептидными компонентами и фенотипическими (морфологическими, биохимическими и т.д.) признаками. Так, например, сорта морозостойких пшениц включают целые блоки компонентов, что обусловлено полигенным наследованием этого важного признака. У изученных 280 сортов озимой мягкой пшеницы повышенная морозостойкость связана с наличием в а-зоне блока компонентов 2467 или блока 24567, а в ю-зоне — блока компонентов 819193102 и ряд других блоков [8]. Из изученных нами сортов одновременное присутствие этих блоков характерно для сортов Мироновская 808, Саратовская 90, Оренбургская 105 (табл. 2). В условиях Оренбуржья сорт Мироновская 808 не отличается высокой зимостойкостью, но высокозимостойким является сорт Саратовская 90 [9]. По этим же данным, повышенную зимостойкость имеет сорт Колос Оренбуржья, но у него в полипептидном спектре имеются только компоненты 536^ (в а-зоне) и присутствуют все необходимые для проявления морозостойкости компоненты 819193102 (в ю-зоне). Отметим также, что у зимостойкого сорта Саратовская 90 есть дополнительные компоненты 12 (в а-зоне) и 6t (в ю-зоне) (см. табл. 2). Очевидно, что для этих и других сортов необходимо по единой методике установить их морозостойкость. Также нужно отметить, что все изученные сорта не имеют маркёров 8292 (в ю-зоне) низкой морозостойкости, но имеют маркёры 567 в а-зоне (зимостойкий сорт Колос Оренбуржья), которые якобы связаны с низкой морозостойкостью [8]. Вполне возможно, что низкая морозостойкость больше определяется ю-полипептидами.
Просо. Выше отмечалось, что представляло техническое затруднение получить спектры у проса, что связано с большим количеством танина в его семенах. Как видно из данных таблицы 2, по типовому спектру проламинов спектр проса близок к спектру пшеницы, но не ячменя. Всего у проса выделяются 22 компонента. При этом близость к пшенице маркируют общие с ней компоненты 102, 93 (среди ю-полипептидов), компонент 32 (среди у-полипептидов), компонент 2 частый компонент 32 (среди в-полипептидов), компоненты 7 6 2 12 (среди а-полипептидов). Из литературы известно, что роды трибы просовых слабо изучены по запасным белкам семян. Имеющиеся данные по близким
к просу родам Setaria (щетинник), Oplismenus (остянка), Brachiaria (ветвянка) показывают, что эта триба, как и триба пшеницевых, имеет в спектрах ю-полипептиды [10], что позволяет относить обе эти трибы к древним злакам [6]. Таким образом, перечисленные выше 9 полипептидных компонентов, общих с пшеницевыми, представляются одними из древнейших компонентов злаков.
Из этих же данных также видно, что изученные образцы сорного и культивируемого проса почти не различаются, но в высокомолекулярной части спектра обычно встречаются яркие (3 балла) полипептидные ю-компоненты. Как отмечалось, методом ДНК- маркёров не удалось распознать географические образцы культивируемого и сорного проса [1]. Опираясь на наши данные, можно предположить, что при использовании белковых маркёров можно-таки выявить существующие географические различия по испытываемым образцам проса. Поэтому есть перспективы более широкого использования метода белковых маркёров на сортах, формах, гибридах проса. На гибридах можно установить компоненты, которые передаются при гибридизации по наследству, ведь известно, что грязно-жёлтая окраска зерновки у сорного проса является в Ft доминантным признаком [11]. Кроме того, можно ожидать, что метод белковых маркёров обязательно выявит степень биологического «загрязнения» культивируемого проса со стороны сорного проса (за счёт их гибридизации) и установит неминуемую при этом процессе ин- трогрессию генов от сорного к культивируемому просу и наоборот. В этом плане просо является уникальным объектом для изучения генетических процессов в любой флоре.
В заключение авторы статьи выражают благодарность всем сотрудникам Оренбургского НИИСХ Россельхозакадемии и Оренбургского ГАУ МСХ РФ за предоставленные для молекулярного анализа семена сортов злаковых растений.
Список литературы
Введенская И. О., Ваухан Д. А., Курцева А. Ф., Дой К. Оценка генетического разнообразия проса обыкновенного (Panicum miliaceum) на основе использования ДНК-маркёров // Сельскохозяйственная биология. 2002. № 5. С. 56—64.
Конарев А. В. Использование молекулярных маркёров в решении проблем генетических ресурсов растений и селекции // Аграрная Россия. 2006. № 6. С. 4—22.
Идентификация сортов и регистрация генофонда культурных растений по белкам семян / под ред. академика РАСХН В. Г. Конарева. СПб. : ВИР, 2000. 186 с.
Гаврилюк И. П. Молекулярные маркёры в идентификации, регистрации и сохранении биоразнообразия растений // Биоразнообразие и биоресурсы Урала и сопредельных территорий : материалы между- нар. конф. Оренбург : ОГПУ, 2001. С. 68—69.
Поморцев А. А. Гордеин-кодирующие локусы как генетические маркёры у ячменя : презентация. М. : Ин-т генетики им. Н. И. Вавилова РАН, 2010. 25 с.
Авдеев В. И. Этапы формирования степных ландшафтов в Евразии. Аспекты эволюции видов Poaceae // Известия Оренбургского государственного аграрного университета. 2009. № 2. С. 59—65.
Краснова Л. И. Биология, селекция, семеноводство озимой пшеницы на Южном Урале. Оренбург : Издат. центр ОГАУ, 2003. 380 с.
Губарева Н. К., Алпатьева Н. В. К вопросу об использовании белковых маркёров в оценке морозостойкости озимой мягкой пшеницы // Аграрная Россия. 2002. № 3. С. 31—34.
Краснова Л. И. Сорта озимой пшеницы // Сортовые ресурсы Оренбуржья : коллективная монография / под ред. проф. А. Г. Крючкова. Оренбург : РАСХН, 2011. С. 43—101.
Конарев А. В., Введенская И. О. Белковые маркёры в анализе видов и популяций злаковых трав трибы мятликовых // Теоретические основы селекции. Т. 1. Молекулярно-биологические аспекты прикладной ботаники, генетики и селекции. М. : Колос, 1993. С. 193—232.
Красавин В. Д. Идентификация проса сорного (Panicum miliaceum subsp. ruderale) и проса посевного (Panicum miliaceum subsp. miliaceum). Оренбург : Оренб. НИИСХ РАСХН, 2002. 30 с.
|
