Эколого-географические закономерности сезонного развития декоративных видов природной флоры в лесостепной зоне Западной Сибири
Т.И. Фомина
Выявлен ритмологический спектр 149 видов, перспективных при интродукции в лесостепной зоне Западной Сибири. Состав биоморфологических групп по сезонным ритмам развития (срокам весеннего отрастания и начала цветения, продолжительности вегетации) проанализирован в связи с географическим и поясно-зональным распространением видов. Большинство их являются весенне-летнезелеными видами с ранним отрастанием, раннелетним или летним сроками начала цветения.
Создание и поддержание коллекций видов природной флоры в ботанических садах - один из приемов сохранения генетических ресурсов и рационального использования полезных растений [1-3]. Интродукция дикорастущих декоративных видов направлена, прежде всего, на обогащение ассортимента растений для оптимизации экологических и эстетических параметров урбанизированной среды. Изучение многих объектов природной флоры с последующей комплексной их оценкой позволяет проводить отбор видов и форм, наиболее перспективных для культивирования.
В Центральном сибирском ботаническом саду (г. Новосибирск) коллекция декоративных видов природной флоры была сформирована в середине 1990-х гг. Мобилизация видов флоры Сибири осуществлялась путем сборов растений и семян в природных популяциях, а видов инорайонных флор - делектусными семенами. Основной целью исследования было выявление особенностей сезонного развития видов при адаптации к условиям лесостепи Западной Сибири и в связи с их экологогеографическим распространением.
Климат района континентальный, умереннохолодный с недостаточным увлажнением [4]. Средняя температура зимних месяцев равна -15°, летних 16-19°. Сумма температур воздуха выше 10°C, обеспечивающих активную вегетацию растений, составляет в среднем 1920°. Безморозный период короткий или средней продолжительности, от 92 до 144 дней. Вегетационный период (со среднесуточными температурами воздуха выше 5°) продолжается 158 дней. Среднемноголетнее количество осадков равно 442 мм, в том числе за период с апреля по октябрь выпадает 338 мм [5].
В настоящей работе приводятся результаты изучения 149 перспективных в культуре видов, относящихся к 37 семействам и 88 родам покрытосеменных растений: 5 из них полукустарники и полукустарнички (Iberis sem- pervirens, Satureja montana, Hyssopus officinalis, Thymus serpyllum, Vinca minor), остальные - травянистые поли- карпики. Подавляющее большинство видов распространены в умеренном поясе Северного полушария, лишь 3 вида субтропического происхождения. При распределении видов по географическим и поясно-зональным группам использовали флористические и геоботаниче- ские источники [6-8]. Фенологические наблюдения проведены по методике И.Н. Бейдеман [9], феноритмотипы выделены в соответствии с работами И.В. Борисовой и Р. А. Карписоновой [11].
Реклама

По географическому распространению виды составляют 9 групп (рис. 1). Наиболее многочисленная азиатская группа объединяет общеазиатские, центрально-азиатские, восточно-азиатские, североазиатские и южносибирско-монгольскиевиды.Попоясно зональному распределению виды отнесены к 4 комплексам и 11 группам (рис. 2). В равной степени доминируют степной и лесной комплексы, а среди групп - бореальная и лесостепная.
В сезонной ритмике развития наиболее значимыми для оценки поведения видов и их декоративных качеств имеют сроки весеннего отрастания, сроки начала цветения и длительность вегетации. По этим признакам определены ритмологические группы видов.
В зависимости от среднемноголетних дат весеннего отрастания выделены ранние, средние и поздние виды. У ранних видов отрастание проходит вскоре после схода снега и оттаивания верхнего горизонта почвы, как правило, в третьей декаде апреля. Поздние виды начинают вегетацию во второй или третьей декадах мая при прогревании верхнего горизонта почвы. Даты начала вегетации сильно варьируют по годам в зависимости от среднесуточных температур воздуха, достигая у ранних и средних видов 33 дней. Отрастание поздних видов в меньшей степени зависит от погодных условий и проходит с максимальным сдвигом 22 дня.
По датам начала цветения виды распределены по 5 группам: ранневесенние (27.04-15.05), поздневесенние (16.05-5.06), раннелетние (6.06-25.06), летние (26.0615.07) и позднелетние (16.07-15.08). У весенних видов смещение дат начала цветения достигает 28-35 дней, у видов летних групп бывает более значительным. В то же время ритм цветения некоторых видов очень устойчивый, с варьированием даты начала цветения не более 3-5 дней (Eryngium planum, Veronica spicata, Euphorbia cyparissias, Geum coccineum, Viola cucullata и др.).
Реклама

По длительности периода вегетации виды представляют 2 ритмологические группы и 5 феноритмотипов: коротковегетирующие - эфемероиды, гемиэфемероиды и длительновегетирующие - весенне-летнезеленые, весенне-летне-осеннезеленые, весенне-летне-зимнезе- леные (табл. 1).
Коротковегетирующая группа в коллекции представлена 6 видами. Им свойственно раннее отрастание и весенние сроки цветения. Сезонный цикл эфемероидов (Erythronium sibiricum, Corydalis bracteata, Anemonoides altaica) длится 34-45 дней. Эти растения отрастают и зацветают в третьей декаде апреля, вегетацию заканчивают в первой декаде июня. У гемиэфемероидов вегетация завершается во второй половине ию- ля-начале августа, составляя от 83 дня у Allium aflatun- ense до 100-101 дня у Allium microdictyon и Adonis ver- nalis.
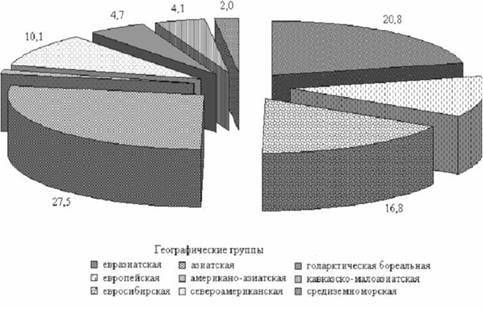
Рис. 1. Географические группы видов, %
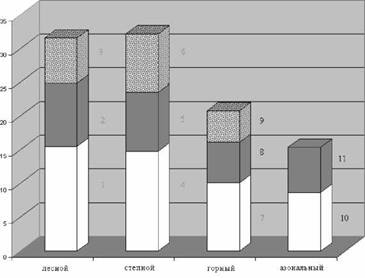
Рис. 2. Поясно-зональные комплексы и группы, %:
1 - неморальная; 2 - бореально-монтанная; 3 - бореальная; 4 - степная; 5 - горно-степная; 6 - лесо-степная; 7 - высокогорная; 8 - монтанная; 9 - скально-россыпная; 10 - луговая; 11 - лугово-болотная
Таблица 1
Группы видов по сезонной ритмике развития
| Фенофаза
|
Группа
|
Число видов по феноритмотипам
|
%
|
| Э
|
ГЭ
|
ВЛЗ
|
ВЛОЗ
|
ВЛЗЗ
|
всего
|
| Весеннее
отрастание
|
ранняя
|
3
|
3
|
33
|
16
|
28
|
83
|
55,7
|
| средняя
|
-
|
-
|
27
|
25
|
4
|
56
|
37,6
|
| поздняя
|
-
|
-
|
10
|
-
|
-
|
10
|
6,7
|
| Начало
цветения
|
ранневесенняя
|
3
|
1
|
2
|
1
|
4
|
11
|
7,4
|
| поздневесенняя
|
-
|
2
|
11
|
2
|
8
|
23
|
15,4
|
| раннелетняя
|
-
|
-
|
21
|
14
|
15
|
50
|
33,6
|
| летняя
|
-
|
-
|
27
|
18
|
5
|
50
|
33,6
|
| позднелетняя
|
-
|
-
|
9
|
6
|
-
|
15
|
10,0
|
| всего
|
3
|
3
|
70
|
41
|
32
|
149
|
|
| %
|
2,0
|
2,0
|
47,0
|
27,5
|
21,5
|
|
|
В самой многочисленной группе весенне-летне- зеленых видов отмечается наибольший разброс по срокам отрастания и продолжительности вегетации. Отрастание проходит преимущественно в ранние и средние сроки. В то же время все поздние виды принадлежат к весенне-летнезеленым. Цветение наступает в период с середины мая до середины июля в широком диапазоне сумм эффективных температур. Продолжительность вегетации составляет 100-160 дней.
Весенне-летне-осеннезеленые виды вегетируют с весны до появления снежного покрова и установления отрицательных температур воздуха (в условиях Новосибирска - третья декада октября). Длительная вегетация, в течение 161-178 дней, обусловлена у большинства видов развитием осенней генерации розеточных побегов. Однако их весеннее отрастание в среднем запаздывает по сравнению с летнезелеными видами. Осенние побеги в зимний период под снегом отмирают, следовательно, их развитие биологически нецелесообразно с точки зрения современных условий существования видов, что уже отмечалось ранее [12]. С излишним расходом пластических веществ, вероятно, и связано более позднее развертывание почек возобновления весной. Начало цветения сдвинуто на раннелетний и летний период - всего 7,3% весенних видов, тогда как в группе летнезеленых их 18,6%.
Весенне-летне-зимнезеленые виды сохраняют зеленые побеги круглогодично, поэтому весной их вегетация возобновляется вскоре после схода снега. К ним относятся хамефиты: полукустарники и полукустарнички, наземноползучие травы (Lysimachia nummularia, Antennaria dioica, виды Sedum L. и др.), Veronica incana, Dianthus arenarius, Linum perenne и др. Биологическое значение зимнезелености состоит в максимально раннем переходе к фотосинтезу, позволяющем растениям осуществлять наибольшую в данных условиях среды продуктивность. Как и осеннезеленость, этот признак сезонной ритмики связан с экологоисторическими обстоятельствами формирования видов в условиях с более мягкой зимой [12, 13]. Зимнезеленые виды цветут весной или рано летом. Среди них мало летних видов, доминирующих в других длитель- новегетирующих группах, - всего 15,6% и отсутствуют поздноцветущие виды.
Длительность вегетации растений в культуре обусловлена, в первую очередь, эволюционно сформированным феноритмом. В группе летнезеленых 63,1% видов заканчивают вегетацию до наступления осенних заморозков, во второй половине августа - первой половине сентября, т. е. независимо от погодных условий вегетационного периода. В основном это азиатские виды, включая все южносибирско-монгольские, подавляющее большинство общеазиатских и североазиатских. У остальных летнезеленых видов вегетация прекращается вынужденно, с наступлением осенних заморозков. Из 7 летнезеленых североамериканских видов 6 заканчивают вегетацию вынужденно, что обусловлено значительно большей продолжительностью вегетационного периода в приатлантической части Северной Америки.
Анализ ритмологического состава географических групп выявил следующие закономерности (табл. 2). В евразиатской группе доминируют весенне-летнезеленые виды с ранним отрастанием, раннелетними и летними сроками цветения. В европейской и евросибирской группах распределение видов по феноритмотипам с длительной вегетацией довольно равномерное. С усилением континентальности климата, при переходе от европейской группы к евросибирской и евразиатской, снижается процент видов, зимующих с зелеными листьями, и прослеживается тенденция к более раннему отрастанию. По срокам цветения среди европейских видов преобладают раннелетние, тогда как в евросибирской группе с ними успешно конкурируют виды с весенним началом цветения. Участие обеих групп в летнем цветении заметно ниже, чем евразиатских видов.
В азиатской группе, приуроченной по большей части к областям с резко континентальным климатом, резко снижается процент зимнезеленых и осеннезеленых видов. Распределение по срокам отрастания близко к евросибирской группе с доминированием ранних видов. Сроки начала цветения различны: южносибирско- монгольские виды зацветают преимущественно весной, североазиатские в июне, а восточно-азиатские цветут в основном в более поздние сроки. В североамериканской группе распределение феноритмотипов приближается к евразиатскому. Однако по другим ритмологическим признакам североамериканские виды резко выделяются на фоне групп евразиатского происхождения. Влияние более продолжительного вегетационного периода на родине этих видов при интродукции в Новосибирске проявляется в сильном смещении начала вегетации и начала цветения на поздние сроки.
Изученные голарктические виды характеризуются длительной вегетацией (57,1% из них летнезеленые) и ранним отрастанием (71,4%). Подавляющее их большинство цветет рано летом. Кавказско-малоазиатские и средиземноморские виды сохраняют зеленые листья круглогодично или до установления снежного покрова, отрастают довольно рано. Цвести начинают в период с середины мая до середины июля, при этом 55,6% видов летние. Коротковегетирующие виды относятся к азиатской и евросибирской группам. Ввиду малочисленности они не оказывают заметного влияния на ритмологический спектр своих географических групп.
Определенные закономерности проявляются в сезонной ритмике поясно-зональных комплексов и групп (табл. 3). Весенне-летнезеленые виды доминируют во всех комплексах, кроме горного, что соответствует общему распределению феноритмотипов в коллекции. В горных условиях наиболее распространены виды с розеточной биоморфой, обеспечивающей более длительную и более раннюю вегетацию, отсюда преобладание зимне- и осеннезеленых видов.
Более мягкие условия перезимовки травянистых растений в широколиственных лесах по сравнению с таежными повышают количество видов, длительно сохраняющих зеленые листья. Неморальные виды в среднем позже начинают вегетацию, а зацветают довольно равномерно в течение всего сезона. Бореально- монтанная группа отличается меньшим процентом зимнезелености, более равномерным отрастанием весной и более ранним цветением, чем равнинные виды. Для лесных видов позднее отрастание не характерно.
Ритмологический спектр основных географических групп, % видов
Т а б л и ц а 2
| Ритмологические группы
|
Евразиатская
|
Европейская
|
Евросибирская
|
Северо
американская
|
Азиатская
|
| По длительности вегетации
|
ВЛЗЗ
|
19,4
|
36,8
|
28,0
|
20,0
|
7,3
|
| ВЛОЗ
|
25,8
|
36,8
|
32,0
|
33,3
|
12,2
|
| ВЛЗ
|
54,8
|
26,4
|
32,0
|
46,7
|
70,7
|
| Э+ГЭ
|
-
|
-
|
8,0
|
-
|
9,8
|
| По срокам весеннего отрастания
|
Р
|
61,3
|
52,6
|
60,0
|
26,7
|
61,0
|
| Ср
|
32,2
|
42,1
|
36,0
|
53,3
|
34,1
|
| П
|
6,5
|
5,3
|
4,0
|
20,0
|
4,9
|
| По срокам начала цветения
|
РВ+ПВ
|
16,2
|
21,0
|
36,0
|
13,3
|
29,3
|
| РЛ
|
35,4
|
47,4
|
32,0
|
26,7
|
24,4
|
| Л
|
41,9
|
26,3
|
24,0
|
33,3
|
34,1
|
| ПЛ
|
6,5
|
5,3
|
8,0
|
26,7
|
12,2
|
Т а б л и ц а 3
Ритмологический спектр поясно-зональных комплексов и групп, % видов
| Ритмологические группы
|
Лесной
|
Степной
|
Горный
|
Азональный
|
| Б
|
БМ
|
Н
|
всего
|
ЛС
|
С
|
МС
|
всего
|
М
|
В
|
СР
|
всего
|
Л
|
ЛБ
|
Все
го
|
| 1
|
ВЛЗЗ
|
8,5
|
2,1
|
4,3
|
14,9
|
8,3
|
6,2
|
6,3
|
20,8
|
6,5
|
6,5
|
32,2
|
45,2
|
-
|
4,3
|
4,3
|
| ВЛОЗ
|
12,8
|
8,5
|
8,5
|
29,8
|
10,4
|
2,1
|
6,3
|
18,8
|
9,7
|
12,9
|
16,1
|
38,7
|
8,7
|
17,4
|
26,1
|
| ВЛЗ
|
27,7
|
14,8
|
8,5
|
51,0
|
25,0
|
18,7
|
14,6
|
58,3
|
3,2
|
3,2
|
-
|
6,4
|
34,8
|
34,8
|
69,6
|
| Э+ГЭ
|
-
|
4,3
|
-
|
4,3
|
2,1
|
-
|
-
|
2,1
|
9,7
|
-
|
-
|
9,7
|
-
|
-
|
-
|
| |
|
| 2
|
Р
|
34,1
|
17,0
|
6,4
|
57,5
|
27,1
|
8,3
|
14,6
|
50,0
|
22,6
|
19,4
|
35,4
|
77,4
|
26,1
|
8,7
|
34,8
|
| С
|
14,8
|
12,8
|
12,8
|
40,4
|
14,6
|
16,7
|
8,3
|
39,6
|
6,5
|
3,2
|
12,9
|
22,6
|
13,0
|
34,8
|
47,8
|
| П
|
-
|
-
|
2,1
|
2,1
|
4,2
|
2,0
|
4,2
|
10,4
|
-
|
-
|
-
|
-
|
4,3
|
13,1
|
17,4
|
| |
|
| 3
|
РВ+ПВ
|
10,7
|
12,8
|
6,3
|
29,8
|
8,3
|
4,2
|
4,2
|
16,7
|
12,8
|
9,7
|
6,5
|
29,0
|
4,3
|
8,7
|
13,0
|
| РЛ
|
17,0
|
10,6
|
4,3
|
31,9
|
16,7
|
6,2
|
8,3
|
31,2
|
12,8
|
6,5
|
19,4
|
38,7
|
8,7
|
26,1
|
34,8
|
| Л
|
17,0
|
6,4
|
6,4
|
29,8
|
12,5
|
14,6
|
10,4
|
37,5
|
3,2
|
-
|
22,6
|
25,8
|
26,1
|
17,4
|
43,5
|
| ПЛ
|
4,2
|
-
|
4,3
|
8,5
|
8,3
|
2,1
|
4,2
|
14,6
|
-
|
6,5
|
-
|
6,5
|
4,4
|
4,3
|
8,7
|
Примечание. Группы по: 1 - длительности вегетации, 2 - срокам весеннего отрастания, 3 - срокам начала цветения.
В лиственных лесах развитие побегов у травянистых растений приурочено к благоприятному по условиям освещения периоду - до развертывания листьев на деревьях и подлеске [11]. В зоне тайги раннее отрастание, кроме того, обусловлено меньшей продолжительностью вегетационного периода.
Степной комплекс по распределению феноритмо- типов не отличается существенно от лесных групп, но по срокам начала вегетации выделяется появлением поздних видов. Цветение приурочено в основном к раннелетнему и летнему периодам, как и у лесных видов. При этом ниже процент весенних и, наоборот, выше процент поздноцветущих видов. Лесостепные и монтанно-степные виды по ритмологическим признакам довольно близки. Степные виды отличаются сравнительно большей долей летнезеленых, причем 2/3 их отрастают в средние сроки, что позднее большинства видов предыдущих групп. В сезонном спектре цветения степных видов явно доминируют летние.
Для горного комплекса летнезеленые виды не характерны, а максимальный процент зимнезелености отмечается в скально-россыпной группе. Вегетация наиболее ранняя относительно остальных поясно-зональных комплексов. Основная часть видов цветет в раннелетний период, но сравнительно велика доля весенних видов - 29,0%. Летний спектр цветения образуют скальнороссыпные виды. Азональный комплекс характеризуется преобладанием летнезеленого феноритмотипа. Среди луговых и лугово-болотных видов зимнезеленые почти не встречаются. Приуроченность к пониженным местам, часто в сочетании с избыточным увлажнением, более длительное прогревание почвы весной обусловливают довольно позднее отрастание видов этих групп. Сроки их цветения также сдвинуты в направлении летних.
На основании вышесказанного можно сделать следующие выводы.
Исследована сезонная ритмика 149 видов природной флоры, перспективных для культуры в лесостепной зоне Западной Сибири в качестве декоративных многолетних растений. При интродукции выделены ритмологические группы: 1) по срокам весеннего отрастания - ранняя, средняя, поздняя; 2) по срокам начала цветения - ранневесенняя, поздневесенняя, раннелетняя, летняя и позднелетняя; 3) по длительности вегетации - короткове- гетирующая с феноритмотипами эфемероиды и гемиэфемероиды, длительновегетирующая с феноритмотипами весенне-летнезеленые, весенне-летне-осеннезеленые и весенне-летне-зимнезеленые виды.
Ритмологические признаки проанализированы в связи с географическим и поясно-зональным распространением видов. По географическому распространению виды отнесены к 9 группам, по поясно-зональному распределению - к 4 комплексам и 11 группам.
В лесостепной зоне Западной Сибири среди декоративных многолетних растений природной флоры доминируют весенне-летнезеленые виды с ранним отрастанием. Основную роль в сезонном спектре цветения играют раннелетние и летние виды.
Выявлен ритмологический состав географических групп. С усилением континентальности климата, при переходе от европейской группы к евросибирской и евразиатской, далее - к азиатской, снижается процент осенне- и зимнезеленых видов, а также выражена тенденция к более раннему отрастанию. Сроки цветения сдвигаются с раннелетних на более ранние или более поздние. Североамериканская группа выделяется сильным смещением начала вегетации и начала цветения на поздние сроки.
Выявлен ритмологический спектр пояснозональных комплексов и групп. Весенне-летнезеленые виды преобладают во всех комплексах, за исключением горного. Последний отличается доминированием зимне- и осеннезеленых видов. Азональный комплекс характеризуется низким процентом зимнезелености и смещением фенофаз отрастания и начала цветения на сравнительно более поздние сроки.
Список литературы
Соболевская К.А. Интродукция растений в Сибири / Отв. ред. И.Ю. Коропачинский. Новосибирск: Наука, 1991. 184 с.
Интродукция растений в Главном ботаническом саду им. Н.В. Цицина: К 50-летию основания / С.Е. Коровин, З.Е. Кузьмин, В.Н. Былов и др. М.: Наука, 1995. 188 с.
Андреев Л.Н., Горбунов Ю.Н. Роль ботанических садов России в сохранении биологического разнообразия растений // Биологическое разнообразие. Интродукция растений: Материалы Третьей Междунар. науч. конф. СПб., 2003. С. 5-7.
ШашкоД.И. Агроклиматические ресурсы СССР. Л.: Гидрометеоиздат, 1985. 247 с.
Научно-прикладной справочник по климату СССР. СПб.: Гидрометеоиздат, 1993. Сер. 3. Многолетние данные. Ч. 1-6. Вып. 20. 717 с.
Алехин В.В. География растений (основы фитогеографии, экологии и геоботаники) / Под ред. проф. Л.В. Кудряшова. 3-е изд. М.: Учпедгиз, 1950. 420 с.
Малышев Л.И., Пешкова Г.А. Особенности и генезис флоры Сибири (Предбайкалье и Забайкалье). Новосибирск: Наука, 1984. 265 с.
Седельников В.П. Высокогорная растительность Алтае-Саянской горной области. Новосибирск: Наука, 1988. 223 с.
Бейдеман И.Н. Методика изучения фенологии растений и растительных сообществ / Отв. ред. Г.И. Галазий. Новосибирск: Наука, 1974. 156 с.
Борисова И.В. Сезонная динамика растительного сообщества // Полевая геоботаника. 1972. Т. 4. С. 5-94.
КарписоноваР.А. Травянистые растения широколиственных лесов СССР: эколого-флористическая и интродукционная характеристика. М.: Наука, 1985. 205 с.
Горышина Т.К. Экология травянистых растений лесостепной дубравы. Л.: Изд-во Ленинград. ун-та, 1975. 127 с.
Голубев В.Н. Эколого-биологические особенности травянистых растений и растительных сообществ лесостепи. М.: Наука, 1965. 287 с. Статья представлена научной редакцией «Биология» 9 октября 2009 г.
|
