| МИНИСТЕРСТВО ПРИРОДНЫХ РЕСУРСОВ РОССИЙСКОЙ ФЕДЕРАЦИИ
ГОСУДАРСТВЕННЫЙ ПРИРОДНЫЙ ЗАПОВЕДНИК «БЕЛОГОРЬЕ»
УТВЕРЖДАЮ
Директор заповедника
«Белогорье»
_________________ А.С.Шаповалов
"_____"______________2006 г.
ЛЕТОПИСЬ ПРИРОДЫ
2004 год
Книга 5
Зам. директора по науке, к.г.н. В.А. Немченко
пос. Борисовка
| СОДЕРЖАНИЕ
|
|
| |
стр.
|
| Список исполнителей ……………………………………………………………………….
|
3
|
Реферат ………………………………………………………………………………………
|
4
|
Введение ……………………………………………………………………………………..
|
5
|
| 1. Территория заповедника …………………………………………………………………
|
6
|
| 1.1. Общие сведения ………………………………………………………………………
|
6
|
| 5. Погода …………………………………………………………………………………….
|
8
|
| 5.1. Климатические особенности 2004 года …………………………………………….
|
8
|
| 5.2. Времена года ………………………………………………………………………….
|
9
|
| 7. Флора и растительность …………………………………………………………………
|
30
|
| 7.1. Флора …………………………………………………………………………………
|
30
|
| 7.1.1. Аннотированный конспект флоры листостебельных мхов участка «Лес на
Ворскле»…………………………………………………………………………
|
30
|
| 7.1.2. Лишайники особо охраняемых природных территорий Белгородской области
|
59
|
| 7.1.3. Видовой состав семейства Lamiaceae заповедника «Белогорье»………………
|
71
|
| 7.2. Растительность ……………………………………………………………………….
|
77
|
| 7.2.1. Степная растительность …………………………………………………………..
|
77
|
| 7.2.1.1. Фенология видов травянистых растений участка «Ямская степь» …………
|
77
|
| 7.2.1.2. Продуктивность надземной массы травостоя участка «Ямская степь» ……
|
85
|
| 7.2.2. Лесная растительность …………………………………………………………..
|
101
|
| 7.2.2.1. Динамика опада древесно-кустарникового полога лесных насаждений ….
|
101
|
| 7.2.2.2. Структура и динамика лесных сообществ участка «Лес на Ворскле» …….
|
106
|
| 8. Фауна и животное население ……………………………………………………………
|
139
|
| 8.1. Фауна …………………………………………………………………………………
|
139
|
| 8.1.1. Список видов птиц ООПТ Белгородской области …………………………….
|
139
|
| 8.1.2. Териофауна ……………………………………………………………………….
|
142
|
| 8.2. Животное население …………………………………………………………………
|
143
|
| 8.2.1. Дикие пчелиные участка «Острасьевы яры» и прилегающих сельхозугодий ..
|
143
|
| 8.2.2. Учёты населения птиц участка «Лес на Ворскле» …………………………….
|
146
|
| 8.2.2. Население млекопитающих ………………………………………………………
|
149
|
| 8.2.2.1. Население рукокрылых ………………………………………………………..
|
149
|
| 8.2.2.2. Население мелких млекопитающих …………………………………………..
|
151
|
| 8.2.2.3. Население норных хищников …………………………………………………
|
153
|
| 8.2.2.4. Население диких копытных ……………………………………………………
|
154
|
| 8.3. Популяции отдельных видов ………………………………………………………..
Реклама

|
155
|
| 8.3.1. Бурозубка обыкновенная ………………………………………………………..
|
155
|
| 8.3.2. Вечерница рыжая …………………………………………………………………
|
155
|
| 8.3.3. Заяц-русак ………………………………………………………………………...
|
160
|
| 8.3.4. Пеструшка степная ……………………………………………………………….
|
161
|
| 8.3.5. Пеструшка восточноевропейская ……………………………………………….
|
161
|
| 8.3.6. Мышь полевая …………………………………………………………………….
|
162
|
| 8.3.7. Мышь лесная ……………………………………………………………………..
|
162
|
| 10. Состояние заповедного режима ……………………………………………………….
|
163
|
| 10.1. Прямые и косвенные воздействия (пожары) ……………………………………..
|
163
|
| Список использованных литературных и архивных материалов………………………..
|
164
|
СПИСОК ИСПОЛНИТЕЛЕЙ
Арбузова М.В. –
старший научный сотрудник, к.с.-х.н. (подпункт 7.2.2.1);
Жемчужников А.С. –
заместитель директора по охране заповедника ( разделы 1, 10);
Конорева Л.А. –
аспирант кафедры ботаники СПбГУ ( пункт 7.1.2);
Маслова Е.В. -
аспирант кафедры ботаники и методики преподавания биологии БелГУ
(пункт 7.1.3)
Мезенцев А.И. –
старший научный сотрудник, к.б.н. (пункт 8.2.1);
Немыкин А.А.
– магистрант кафедры ботаники СпбГУ (пункт 7.1.1);
Немченко В.А.
– ответственный исполнитель, заместитель директора по науке, к.г.н. (редактирование, разделы 1, 5, 10, подпункт 7.2.2.2);
Солнышкина Е.Н.
– младший научный сотрудник (пункт 7.2.1);
Шаповалов А.С. –
директор заповедника (редактирование, пункты 8.1.2, 8.2.2, подраздел
8.3);
Шерстаков Д.Н.
– аспирант кафедры зоологии позвоночных СпбГУ (пункты 8.1.1, 8.2.2);
Христофорова Н.В. –
техник – метеоролог (раздел 5).
РЕФЕРАТ
Наблюдение и изучение явлений и процессов в природном комплексе государственного природного заповедника «Белогорье» (Летопись природы).
Научный отчёт за 2004 г., 167 с., 6 рис., 62 табл., 66 источников.
МОНИТОРИНГ ПРИРОДНЫХ ПРОЦЕССОВ, ТЕРРИТОРИЯ, ПОЧВЫ,
ПОГОДА, ФЛОРА, РАСТИТЕЛЬНОСТЬ, ФАУНА, ЖИВОТНОЕ НАСЕЛНИЕ, ПОПУЛЯЦИИ ВИДОВ, СОСТОЯНИЕ ЗАПОВЕДНОГО РЕЖИМА.
Мониторинг природных процессов и биологического разнообразия – постоянная (бес-
срочная) научная работа заповедника.
Отчёт за 2004 г. составляет 5-ю книгу Летописи природы Государственного Природного заповедника «Белогорье».
Исследования проводятся как на всей территории заповедника (5 участков, 2131,0 га), так и на постоянных пробных площадях, маршрутах, а также на территории Белгородской области.
В отчёте даны характеристики процессов и объектов на основании исследований в 2004 г. В отдельных разделах для анализа используются многолетние данные.
Реклама

Исследования по программе Летописи природы заповедника должны быть продолжены для сохранения старых и формирования новых рядов многолетних наблюдений.
ВВЕДЕНИЕ
Научный отчёт по теме «Наблюдение и изучение явлений и процессов в природном комплексе Государственного Природного заповедника «Белогорье» за 2004 год (Летопись природы, книга 5 ) структурно представлен по схеме методического пособия «Летопись природы …» (Филонов, Нухимовская, 1985).
Летопись природы за 2004 год состоит из одной части, нумерация страниц и разделов – сквозная. Информация, приведённая в “Годовом информационном отчёте директора государственного природного заповедника “Белогорье” за 2004 год “, в Летописи природы не дублируется (сведения о всех научных исследованиях на территории заповедника за текущий год, практике студентов, публикациях, участии сотрудников в совещаниях и конференциях, охране заповедной территории).
Летопись природы подготовили научные сотрудники, техники и лаборанты научного отдела, заместитель директора по охране заповедника, сотрудники и специалисты различных научных учреждений и высших учебных заведений России и Украины. В сборе и обработке первичного материала принимали участие также инспектора по охране заповедника, аспиранты и студенты. Общее руководство работами в 2004 г. осуществляли директор заповедника А.С. Шаповалов и заместитель директора по науке В.А. Немченко. Подготоку и редактирование Летописи природы в 2005-06 гг. провели директор заповедника А.С. Шаповалов и заместитель директора по науке В.А. Немченко.
Кроме ежегодно представляемых материалов по динамике природных процессов на территории заповедника «Белогорье», в Летопись природы за 2004 год включены результаты обследования природных объектов на территории Белгородской области, в том числе проектируемые новые участки заповедника. Наряду с продолжением работ по многолетним рядам наблюдений за отдельными природными явлениями и процессами, начаты новые исследования по структуре и динамике лесостепных ландшафтов.
Из обработанных материалов, помещённых в Летопись природы, следует отметить:
Аннотированный конспект флоры листостебельных мхов участка «Лес на Ворскле» - первое по полноте исследование флоры данных мхов участка, позволившее обнаружить новые для Среднерусской возвышенности и Белгородской области виды (пункт 7.1.1); Аннотированный список видов лишайников ООПТ Белгородской области – пионерная работа по полноте охвата территории области (пункт 7.1.2); исследование Структуры и динамики лесных сообществ участка «Лес на Ворскле» - являющееся продолжением работ на лесотипологических постоянных пробных площадях, заложенных Ю.Н. Нешатаевым в 1963-76 гг. (подпункт 7.2.2.2); результаты исследований популяций отдельных видов, продолжающих многолетние ряды наблюдений данных групп животных (подраздел 8.3).
1.
ТЕРРИТОРИЯ ЗАПОВЕДНИКА
1.1. ОБЩИЕ СВЕДЕНИЯ
Распределение территории заповедника «Белогорье» по площадям и категориям земель по состоянию на 31.12.2004 г. приведено в таблице 1.1.
Таблица 1.1
Распределение территории государственного природного заповедника «Белогорье»
по участкам и категориям земель в 2002 году
| Категория земель
|
Площадь по участкам, га
|
Итого
|
| ЛнВ
|
ОЯ
|
ЯС
|
ЛГ
|
СИ
|
| I. Лесная площадь
|
997
|
62
|
111
|
180
|
1350
|
| 1. Покрытая лесом
|
994
|
33
|
111
|
157
|
1295
|
| в том числе:
|
| - условно естествен- ный лес
|
557
|
33
|
24
|
145
|
759
|
| - лесные культуры
|
434.5
|
87
|
12
|
533.5
|
| 2. Дендропарк
|
2.5
|
2.5
|
| 3. Непокрытая лесом
|
3
|
29
|
23
|
55
|
| в том числе:
|
| – естествен.редины
|
4
|
4
|
| – пустыри, прогалины
|
3
|
25
|
28
|
| –несомкнувкнувшие- ся лесные культуры
|
23
|
23
|
| II. Нелесная площадь
|
41
|
504
|
59
|
87
|
691
|
| 1. Степи и луга
|
| в том числе:
|
| – косимые(сенокосы)
|
388
|
1
|
32
|
421
|
| – некосимые(абс.запо-ведная степь, 2 участ.)
|
20.7
|
| – выпасаемые(пастб.)
|
4
|
15
|
19
|
| 2.Пашни
|
17
|
1
|
18
|
| 3. Залежи
|
| 4. Усадьбы
|
8
|
1
|
9
|
| 5. Сады, питомники
|
6
|
6
|
| 6. Воды
|
1
|
1
|
| 7. Болота
|
7
|
7
|
| 8. Кустарники
|
| 9. Дороги, просеки
|
9
|
1
|
4
|
1
|
15
|
| 10. Пески
|
1
|
1
|
| 11. Прочие земли
|
1
|
88
|
85.3
|
57
|
32
|
284
|
| Общая площадь
|
1038
|
90
|
566
|
170
|
267
|
2131
|
| Охранная зона
|
488
|
1400
|
860
|
2748
|
| Всего охраняемой территории
|
1526
|
90
|
1966
|
1030
|
267
|
4879
|
Примечание.
Участки заповедника: ЛнВ – Лес на Ворскле, ОЯ – Острасьевы яры, ЯС – Ямская степь, ЛГ – Лысые горы, СИ – Стенки Изгорья.
Данные по участку «Лес на Ворскле» соответствуют материалам лесоустройства 1986 г., участку «Ямская степь» – 1990г.; по участкам «Лысые горы» и «Стенки Изгорья» – материалам госучета лесного фонда 1998 г., полученным от Центрально-Черноземного государственного природного биосферного заповедника при передаче участков в 1999 г. Площадь участка «Острасьевы яры» определена согласно Распоряжения Правительства РФ от 26 сентября 1995 г. № 1342-р, земли которого на тот момент входили в состав сельхозугодий Борисовского района (структуирование земель участка проведено сотрудниками заповедника согласно «Экспликации земельных угодий совхоза «Прогресс» по состоянию на 1.12.94 г.» Комитета по земельным ресурсам и землеустройству Борисовского района Белгородской области).
Структура в целом и площади всех участков территории заповедника по сравнению с предыдущим годом остались без изменений и составляют 2131 га.
Заповедник «Белогорье» в настоящее время состоит из 5-ти обособленных участков, расположенных в пределах 3-х административных районах Белгородской области. На сопредельных с участком «Ямская степь» землях охранная зона шириной 1 км и Положение о ней установлены решением Исполнительного комитета Белгородского областного совета депутатов трудящихся от 30 августа 1977 года № 446, площадь охранной зоны ( согласно лесоустройству 1990 г.) составляет 1400 га. Вокруг участка «Лысые горы» охранная зона шириной 1 км установлена постановлением главы администрации Губкинского района от 12 апреля 1993 года № 135 и постановлением главы администрации Белгородской области от 10 марта 1994 года № 140, площадь которой составляет 860 га. Охранная зона вокруг участка «Лес на Ворскле» общей площадью 488 га и Положение о ней установлены постановлением главы администрации Борисовского района от 25 февраля 1994 года № 52 и постановлением главы администрации Белгородской области от 16 мая 1994 года № 261.
Вокруг участков «Острасьевы яры» (включён в состав заповедника в 1995 г.) и «Стенки Изгорья» (включён в состав заповедника в 1999 г.) охранные зоны пока не определены.
В результате, документально подтверждаемая общая площадь охранных зон участков заповедника «Белогорье» на 31.12.2002 года составляет 2748 га.
5.
ПОГОДА
5.1. КЛИМАТИЧЕСКИЕ ОСОБЕННОСТИ 2004 ГОДА
Метеонаблюдения проводятся на метеопосте заповедника «Белогорье» (192,8 м н.у.м.), расположенного на территории центральной усадьбы ( квартал 8, выдел 34 участка «Лес на Ворскле»), с постоянными четырёхсрочными наблюдениями – в 03, 09, 15 и 21 час. Штат метеопоста: инженер-метереолог и 4 техника-совместителя.
Наблюдения проводятся по следующим метеопараметрам и явлениям:
1. Облачность.
2. Состояние поверхности почвы.
3. Атмосферные явления.
4. Температура воздуха в градусах Цельсия (в метеобудке на высоте 2 м фиксируется срочная температура, минимальная и максимальная суточная температура).
5. Осадки, мм (осадкомер Третьякова).
6. Температура почвы в градусах Цельсия: на поверхности ( срочная, минимальная,
максимальная) и на глубине – 5, 10, 15, 20 см (коленчатые термометры Савинова).
7. Высота снежного покрова (см) по трём рейкам, расположенным на метеоплощадке.
2004 год был несколько более тёплым и особенно влажным по сравнению с 2003 годом. Cредняя температура воздуха за год составила 7,4° (в 2003 г. – 6,4°). Продолжительность периода с температурами выше 0° составила 252 дня (в 2003 г. – 241 день), сумма температур выше 0° равнялась 3023,1° (в 2003 г. – 2975,4°). Самым тёплым месяцем был август со среднемесячной температурой 18,9° (в 2003 г. июль – 19,6°), самым холодным – февраль с температурой –4,2° (в 2003 г. февраль –9,4°) (табл. 5.1, 5.2, 5.8). Абсолютный годовой минимум температуры воздуха отмечен 13 февраля (–21,5°), 28 и 29 июля отмечен годовой абсолютный максимум температуры воздуха – 30,6° (табл. 5.10).
Таблица 5.1
Продолжительность периодов (в днях) со средней температурой выше 0, 5, 10, и 15°
| Периоды
|
0°
|
5°
|
10°
|
15°
|
| 2004 г.
|
252
|
227
|
166
|
101
|
Таблица 5.2
Суммы температур выше 0, 5, 10, и 15°
| Периоды
|
0°
|
5°
|
10°
|
15°
|
| 2004 г.
|
3023,1
|
3018,0
|
2828,5
|
1801,7
|
Осадков за год выпало 819,7 мм (в 2003г. – 591,5 мм), при достаточно равномерном распределении по месяцам и сезонам года. Выделяются сентябрь (112,4 мм, абсолютный максимум месячных осадков за год) и ноябрь (28,9 мм, абсолютный минимум) (табл. 5.9).
Годовой и сезонный ход температуры почвы от поверхности до глубины 20 см представлен в табл. 5.12 – 5.17.
Среднегодовая температура поверхности почвы равна 8,7° (табл. 5.12), при среднегодовой температуре воздуха – 7,4°. В общем, годовой ход среднемесячной температуры поверхности почвы, с некоторым превышением, повторяет динамику среднемесячных температур воздуха. Но в январе и декабре она ниже температуры воздуха, а в ноябре обе температуры равны.
Повсеместно наблюдается превышение среднемесячной температуры почвы на глубинах 5, 10, 15 и 20 см (табл. 5.13 - 5.16) над среднемесячной температурой воздуха. Только в мае на глубинах 5 и 10 см температура почвы ниже чем температура воздуха. Наиболее заметно превышение температуры почвы над температурой воздуха в ноябре.
Экстремальные температуры поверхности почвы (табл. 5.17) следующие: абсолютный максимум отмечен 26 и 28 июля – 51,5°, абсолютный минимум 22 февраля (–23,0°).
Климатическая характеристика года по сезонам дана в таблице 5.3.
5.2. ВРЕМЕНА ГОДА.
Зима 2003-2004 годов
Зима 2003-2004 гг. была значительно короче, теплее и малоснежнее, чем зима 2002-2003 гг. Продолжительность зимнего периода составила 103 дня (в 2002-2003 гг. – 125 дней). Началась зима 1 декабря 2003 г. и закончилась 12 марта 2004 г. Средняя температура воздуха за сезон равнялась –3,0° (в 2002-2003 гг. –7,2°). Самым холодным зимним месяцем был февраль 2004 г. со среднемесячной температурой воздуха –4,2° и при наличии 4 дней с положительной среднесуточной температурой, которых в декабре 2003 г., феврале и марте 2004 г. было 5, 9 и 3 дня, соответственно.
Осадков за сезон выпало 194,4 мм (в 2003 г. – 137,0 мм). Выпадение твёрдых осадков зарегистрировано в течении 42 дней (снег, крупа), с дождём и моросью – 12 дней, со смешанными осадками – 10 дней. Постоянный снежный покров установился 8 декабря 2003 года и продержался до 17 марта 2004 г. Средняя высота снежного покрова в декабре равнялась 7 см, в январе – 19 см, феврале – 20 см и в марте – 5 см (табл.5.11).
Таблица 5.3
Климатическая характеристика года по сезонам
| Показатели
|
Зима
|
Весна
|
Лето
|
Осень
|
| 2003–2004г.
|
2004 г.
|
2004 г.
|
2004 г.
|
| Начало–конец
|
01.12 – 12.03
|
13.03 – 26.05
|
27.05 – 4.09
|
5.09 – 19.11
|
| Продолжитель–
ность (дни)
|
103
|
75
|
101
|
76
|
| Сумма темпера-
тур
|
- 314,9
|
646,0
|
1801,7
|
652,3
|
| Температура
воздуха средняя
|
- 3,0
|
8,6
|
17,8
|
8,6
|
| Сумма осадков (мм)
|
194,4
|
186,1
|
292,7
|
137,3
|
Весна 2004 года
Весна 2004 года началась 13 марта и длилась 75 дней (в 2003 г. – 34 дня и началась 4 апреля). Была она ощутимо теплее и влажнее весны 2003 г. Средняя температура воздуха за сезон равнялась 8,6° (в 2003 г. – 5,9°). Повышение температуры воздуха шло неравномерно. Очень быстро, всего за 8 дней, произошёл переход среднесуточной температуры воздуха с 0° до 5° (табл. 5.4). В дальнейшем повышение температурного фона шло значительно медленнее.
Таблица 5.4
Переход среднесуточных температур через 0°, 5°, 10° и 15° весной
Пределы перехода
|
0°
|
5°
|
10°
|
15°
|
| Даты перехода, 2004 г.
|
13.03
|
21.03
|
20.04
|
27.05
|
Осадков за весенний период выпало 186,1 мм (в 2003 г. – 23,4 мм), распределялись осадки в течении сезона достаточно равномерно. Выпадение снега отмечалось 1, 2 и 3 апреля.
Лето 2004 года
Продолжительность летнего периода 2004 года составила 101 день (в 2003 г. – 117 дней). Средняя температура воздуха за сезон равнялась 17,8° (в 2003 г. – 17,9°). Самым тёплым месяцем был август – 18,9°.
Осадков за лето выпало 292,7 мм (в2003 г. – 277,1 мм), при достаточно равномерном распределении месячных сумм осадков.
Осень 2004 года
Средняя температура за осенний период равнялась 8,6° (в 2003 г. – 6,8°). Продолжительность сезона составила 79 дней (в 2003 г. – 90 дней), начавшись 5 сентября и закончившись 19 ноября. Температура воздуха снижалась достаточно равномерно (табл. 5.5).
Таблица 5.5
Даты устойчивого перехода температур воздуха через 15°, 10°, 5°, 0° осенью
| Пределы перехода
|
15°
|
10°
|
5°
|
0°
|
| Даты перехода, 2004 г.
|
5.09
|
3.10
|
3.11
|
20.11
|
Осадков за осень выпало 137,3 мм (в 2003 г. – 119,3 мм). На фоне влажных сентября и октября (83,6 и 34,7 мм соответсьвенно) выделяется минимум осадков за ноябрь – 19,0 мм.
Январь
Средняя температура воздуха за месяц равнялась –3,5°. Характерен достаточно ровный отрицательный ход температуры воздуха, при постоянном повышении среднедекадных значений: –5,7° в 1-ю декаду, -2,6° во вторую и –2,3° в третью. Отмечено только четыре дня с положительными среднесуточными температурами – 15, 18, 29 и 30 января. Осадков выпало 96,3 мм. Выпадали осадки часто – 19 дней (16 дней с твёрдыми осадками). 15 января наблюдались жидкие осадки (морось). Вторая и третья декады были наиболее влажными.
Февраль
Февраль был холоднее и суше января. Средняя температура воздуха за февраль составила –4,2°. На протяжении месяца наблюдалось 9 дней с положительной среднесуточной температурой (все в I и III декадах). Вторая декада была наиболее холодной (–6.8°). Осадков за месяц выпало 47,4 мм. Выпадение осадков наблюдалось 23 дня (15 дней с твёрдыми осадками). Максимум осадков выпало во II декаду – 33,2 мм (70% месячной суммы).
Март
Средняя температура воздуха за месяц равнялась 2,8°. Наблюдался достаточно быстрый рост температуры воздуха. Переход через 0° произошёл 13 марта, через 5° - 21 марта. Нарастание температуры продолжалось достаточно интенсивно и средне декадная температура воздуха в третьей декаде равнялась 7,3°. Осадков за месяц выпало 63,2 мм (5 дней с твёрдыми осадками). Выпадение осадков росло к концу месяца – 2,8 мм в I декаду, 22,1 мм во вторую и 38,3 мм (61% месячной суммы) в третью. Постоянный снежный покров сошёл 17 марта.
Апрель
В апреле рост температуры воздуха несколько замедлился. Средняя температура воздуха за месяц равнялась 7,7°. На фоне общего подъёма температуры в первой декаде наблюдались дни с отрицательными среднесуточными температурами воздуха (2 и 3 апреля). Переход температуры воздуха через 10° произошёл 20 апреля. В дальнейшем повышение температуры стало ещё менее заметно и средне декадная температура в третьей декаде равнялась 10,3°. Осадков за месяц выпало 41,7 мм на протяжении 12 дней (три дня с твёрдыми осадками). Наиболее сухой была вторая декада – 8,6 мм (все они выпали 12 апреля).
Май
В мае продолжалось замедленное повышенние температуры воздуха на фоне существенного повышение увлажнения. Переход через 15° произошёл 27 мая. Средняя температура воздуха за месяц равнялась 12,6°. Даже в самой тёплой третьей декаде средне декадная температура воздуха составила 15,2°. Осадков за месяц выпало 90,5 мм, с максимумом в третьей декаде – 54,3 мм (60% месячной суммы).
Июнь
Средняя температура воздуха за месяц равнялась 16,3°. В первой декаде средне декадная температура воздуха даже снизилась по сравнению с третьей декадой мая – 14,1°. Лишь со второй декады температура воздуха начала повышаться – средне декадная 16,4° и в третьей декаде – 18,3°. В июне выпало 85.5 мм осадков, при постоянном повышении декадных сумм осадков: 4,0 мм – первая декада, 13,3 мм – вторая и 68,2 мм – третья.
Июль
Июль был теплее июня, температура воздуха постоянно росла на протяжение месяца. Средняя температура за месяц равнялась 18,7°. Самой тёплой была третья декада со средней температурой воздуха 21,4°. В июле выпало 88,3 мм осадков. Осадки выпадали неравномерно в течение 17 дней. Почти 60% месячной суммы осадков (52,9 мм) на протяжение 8 дней выпали во вторую декаду.
Август
В августе температурный фон имел нестабильный колебательный характер (в отличие от июльского постоянного роста) и повышенное увлажнение. Но при этом средняя температура воздуха за месяц незначительно превысила июльскую и равнялась 18,9°. Наиболее тёплой была первая декада со средней температурой воздуха 20.1°, вторая самая прохладная – 17,4°, третья – 21,0°. Осадков за месяц выпало 112,4 мм. Выпадение осадков имело нерегулярный характер. Отмечены осадки на протяжении 10 дней, около 60% (65,1 мм) осадков выпало в третьей декаде, 24 августа наблюдался годовой максимум суммы суточных осадков – 50,5 мм.
Сентябрь
В первой декаде сентября произошло резкое снижение температуры воздуха, среднедекадная температура упала до 14,4°. Переход температуры воздуха через 15° отмечен уже 5 сентября. В дальнейшем падение температуры замедлилось. Средняя температура воздуха за месяц равнялась 13,8°. Осадков за месяц выпало 96,0 мм. Выпадали осадки крайне нерегулярно, около 70% месячных осадков (64,7 мм) выпало в третью декаду, а во вторую декаду выпало всего 0,4 мм осадков.
Октябрь
В октябре средняя температура воздуха равнялась 7,5°. Переход температуры воздуха через 10° произошёл 3 октября. На общем фоне снижения температуры воздуха отмечено повышение средне декадной температуры в третьей декаде до 9,6° с 5,2° во второй декаде. За месяц осадков выпало 34,7 мм (1 день с твёрдыми осадками), с максимумом и по количеству и по интенсивности выпадения во второй декаде – 17,0 мм или около 50% месячной суммы.
Ноябрь
Для ноября характерено постоянное и достаточно стремительное снижение температурного фона. Переход температуры воздуха через 0° отмечен 20 ноября. Средняя температура воздуха за месяц равнялась 0,4°. Третья декада была стабильно морозная, средне декадная температура воздуха равнялась –6,5°. Осадков выпало 28,9 мм (5 дней с твёрдыми осадками). Более 90% месячных осадков выпало во второй декаде (26,4 мм) и 0% - в третьей.
Декабрь
В декабре среднесуточная температура воздуха была крайне изменчива и колебалась от –12,8° до 3,2°. Однако средняя температура воздуха за месяц составила –1,8°. Средне декадные температуры также были отрицательными, самой тёплой была первая декада (-0,4°), самой холодной – вторая (-3,0°). Осадков в декабре выпало 34,8 мм. Осадки выпадали в твёрдом виде 15 дней, в жидком и смешанном виде по 3 дня каждые.
Таблица 5.6
Число дней с атмосферными явлениями в 2004 г.
| Месяц
|
Атмосферные явления (число дней)
|
| с жидкими
осадками
|
с твердыми
осадками
|
со смешанными
осадками
|
с метелью
|
с инеем и
изморозью
|
с гололедом
|
со снежным
покровом
|
с грозой
|
с росой
|
с туманом
|
с дымкой
|
с пыльной бурей
|
с ветром 15 м/сек
|
без оттепели
|
с морозом в
воздухе
|
с морозом
на почве
|
с относительной влажностью > 80%
|
с относительной влажностью < 30%
|
с поземкой
|
| Январь
|
1
|
16
|
2
|
1
|
9
|
2
|
31
|
-
|
-
|
5
|
-
|
-
|
25
|
27
|
31
|
| Февраль
|
4
|
15
|
4
|
2
|
4
|
6
|
29
|
-
|
-
|
8
|
1
|
-
|
19
|
20
|
29
|
| Март
|
9
|
5
|
1
|
-
|
5
|
-
|
17
|
2
|
-
|
4
|
-
|
-
|
4
|
9
|
21
|
| Апрель
|
10
|
3
|
-
|
-
|
9
|
-
|
-
|
3
|
7
|
3
|
2
|
-
|
2
|
2
|
13
|
| Май
|
16
|
-
|
-
|
-
|
-
|
-
|
-
|
2
|
18
|
2
|
4
|
-
|
-
|
-
|
-
|
| Июнь
|
13
|
-
|
-
|
-
|
-
|
-
|
-
|
4
|
23
|
-
|
2
|
-
|
-
|
-
|
-
|
| Июль
|
17
|
-
|
-
|
-
|
-
|
-
|
-
|
8
|
22
|
-
|
-
|
-
|
-
|
-
|
-
|
| Август
|
10
|
-
|
-
|
-
|
-
|
-
|
-
|
6
|
26
|
1
|
1
|
-
|
-
|
-
|
-
|
| Сентябрь
|
13
|
-
|
-
|
-
|
-
|
-
|
-
|
8
|
21
|
5
|
2
|
-
|
-
|
-
|
-
|
| Октябрь
|
13
|
1
|
-
|
-
|
8
|
-
|
-
|
1
|
19
|
8
|
4
|
-
|
-
|
-
|
5
|
| Ноябрь
|
4
|
5
|
2
|
1
|
10
|
-
|
4
|
-
|
3
|
2
|
4
|
-
|
11
|
12
|
18
|
| Декабрь
|
3
|
15
|
3
|
-
|
4
|
-
|
15
|
-
|
-
|
7
|
5
|
-
|
13
|
17
|
25
|
| Сумма за год
|
113
|
60
|
12
|
4
|
49
|
-
|
96
|
34
|
139
|
45
|
25
|
-
|
74
|
87
|
142
|
Таблица 5.7
Число дней с осадками различных градаций в 2004 г.
| Месяц
|
Градации (мм)
|
| 0.0
|
0.1
|
0.5
|
1.0
|
5.0
|
10.0
|
20.0
|
30.0
|
60.0
|
70.0
|
| Январь
|
-
|
2
|
3
|
7
|
4
|
2
|
1
|
-
|
-
|
-
|
| Февраль
|
6
|
2
|
2
|
10
|
3
|
-
|
-
|
-
|
-
|
-
|
| Март
|
0
|
2
|
4
|
3
|
5
|
-
|
1
|
-
|
-
|
-
|
| Апрель
|
1
|
2
|
2
|
4
|
2
|
1
|
-
|
-
|
-
|
-
|
| Май
|
1
|
-
|
2
|
6
|
4
|
3
|
-
|
-
|
-
|
-
|
| Июнь
|
3
|
1
|
-
|
4
|
3
|
1
|
-
|
1
|
-
|
-
|
| Июль
|
1
|
4
|
1
|
6
|
1
|
3
|
1
|
-
|
-
|
-
|
| Август
|
1
|
-
|
-
|
4
|
1
|
3
|
-
|
1
|
-
|
-
|
| Сентябрь
|
1
|
1
|
1
|
3
|
3
|
3
|
1
|
-
|
-
|
-
|
| Октябрь
|
2
|
4
|
-
|
5
|
3
|
-
|
-
|
-
|
-
|
-
|
| Ноябрь
|
3
|
-
|
1
|
5
|
2
|
-
|
-
|
-
|
-
|
-
|
| Декабрь
|
1
|
3
|
7
|
8
|
2
|
-
|
-
|
-
|
-
|
-
|
| За год
|
20
|
21
|
23
|
65
|
33
|
16
|
4
|
2
|
-
|
-
|
Таблица 5.8
Средняя температура воздуха в 2004 г.
| Число
|
Месяц
|
| 01
|
02
|
03
|
04
|
05
|
06
|
07
|
08
|
09
|
10
|
11
|
12
|
| 1
|
-3,6
|
-6,1
|
3,5
|
3,9
|
7,1
|
13,3
|
14,2
|
24,0
|
17,5
|
12,7
|
8,6
|
-12,8
|
| 2
|
-2,8
|
-6,2
|
-0,6
|
-4,3
|
10,8
|
12,9
|
17,8
|
23,6
|
18,2
|
10,8
|
6,0
|
-2,5
|
| 3
|
-0,8
|
-5,8
|
-0,6
|
-5,4
|
10,0
|
12,4
|
16,0
|
22,1
|
16,2
|
8,7
|
1,7
|
1,4
|
| 4
|
-6,8
|
-5,4
|
-1,5
|
1,8
|
13,9
|
12,1
|
15,6
|
18,1
|
15,2
|
6,2
|
3,4
|
0,9
|
| 5
|
-8,5
|
-1,5
|
-5,3
|
5,8
|
15,8
|
15,6
|
16,9
|
19,9
|
14,2
|
5,7
|
4,8
|
1,0
|
| 6
|
-9,4
|
1,5
|
-6,8
|
7,6
|
16,2
|
15,4
|
17,8
|
19,8
|
14,2
|
6,8
|
4,7
|
2,0
|
| 7
|
-9,1
|
2,5
|
-2,6
|
8,7
|
13,0
|
16,9
|
16,5
|
17,6
|
13,4
|
7,6
|
4,8
|
0,8
|
| 8
|
-6,0
|
2,6
|
-1,3
|
9,2
|
13,1
|
18,6
|
16,8
|
18,2
|
13,5
|
9,5
|
0,0
|
1,5
|
| 9
|
-5,6
|
0,6
|
1,1
|
8,9
|
14,9
|
15,2
|
17,2
|
18,6
|
11,6
|
8,3
|
5,5
|
3,2
|
| 10
|
-4,5
|
-1,8
|
0,1
|
8,4
|
15,4
|
8,6
|
19,2
|
18,9
|
9,6
|
7,8
|
6,8
|
-0,2
|
| Сумма
|
-57,1
|
-19,6
|
-14,0
|
44,6
|
130,2
|
141,0
|
168,0
|
200,8
|
143,6
|
84,1
|
46,3
|
-4,7
|
| 11
|
-3,2
|
-5,3
|
-1,0
|
8,1
|
14,2
|
13,3
|
19,5
|
20,3
|
11,9
|
5,2
|
4,7
|
-2,4
|
| 12
|
-5,1
|
-11,6
|
-0,9
|
7,8
|
13,4
|
16,6
|
18,2
|
15,6
|
9,1
|
2,2
|
4,1
|
-1,0
|
| 13
|
-2,3
|
-16,5
|
1,9
|
6,9
|
8,5
|
18,0
|
18,9
|
17,9
|
14,0
|
3,0
|
6,6
|
-2,4
|
| 14
|
-1,4
|
-4,4
|
3,4
|
5,5
|
8,3
|
21,0
|
18,2
|
18,9
|
15,1
|
2,0
|
8,0
|
-6,9
|
| 15
|
0,8
|
-2,3
|
2,5
|
9,6
|
10,4
|
15,4
|
18,0
|
14,9
|
16,2
|
3,3
|
2,9
|
-9,4
|
| 16
|
-2,1
|
-4,2
|
1,8
|
10,6
|
9,7
|
16,4
|
14,3
|
15,3
|
15,8
|
4,2
|
2,6
|
-7,2
|
| 17
|
-3,6
|
-3,8
|
0,8
|
6,4
|
11,6
|
13,2
|
17,0
|
18,1
|
13,3
|
5,2
|
-0,2
|
-3,7
|
| 18
|
1,6
|
-6,4
|
3,2
|
8,1
|
9,3
|
14,2
|
16,8
|
17,9
|
16,3
|
10,4
|
0,2
|
1,5
|
| 19
|
-2,7
|
-2,5
|
5,6
|
9,9
|
11,2
|
17,7
|
17,6
|
17,8
|
11,9
|
8,6
|
2,4
|
2,0
|
| 20
|
-8,2
|
-12,9
|
3,8
|
10,6
|
11,6
|
18,2
|
18,8
|
17,9
|
14,6
|
7,8
|
-2,0
|
-0,8
|
| Сумма
|
-26,2
|
-68,1
|
21,1
|
83,5
|
108,2
|
164,0
|
177,3
|
174,6
|
138,2
|
51,9
|
29,3
|
-30,3
|
| 21
|
-2,8
|
-14,6
|
8,0
|
12,4
|
11,5
|
21,6
|
19,8
|
20,6
|
16,0
|
4,6
|
-3,6
|
-0,7
|
| 22
|
-7,5
|
-12,3
|
8,1
|
13,3
|
13,1
|
20,7
|
17,7
|
23,0
|
13,8
|
5,7
|
-6,2
|
-5,4
|
| 23
|
-6,8
|
-10,0
|
7,6
|
14,0
|
13,8
|
18,6
|
19,1
|
22,6
|
9,8
|
3,6
|
-6,6
|
-11,2
|
| 24
|
-1,1
|
-4,8
|
10,6
|
12,5
|
13,0
|
19,4
|
20,6
|
17,0
|
8,9
|
11,8
|
-5,9
|
-7,1
|
| 25
|
-0,7
|
0,8
|
10,6
|
10,6
|
10,4
|
19,9
|
22,2
|
15,9
|
8,7
|
12,3
|
-8,4
|
0,6
|
| 26
|
-1,3
|
0,8
|
7,6
|
10,8
|
10,2
|
20,7
|
22,6
|
16,1
|
17,1
|
11,5
|
-8,3
|
-1,0
|
| 27
|
-4,8
|
1,3
|
5,2
|
5,0
|
15,9
|
16,3
|
22,8
|
15,6
|
14,8
|
10,4
|
-6,4
|
1,6
|
| 28
|
-2,8
|
3,2
|
5,4
|
7,2
|
15,7
|
15,8
|
23,1
|
19,0
|
15,6
|
10,0
|
-5,9
|
2,1
|
| 29
|
2,3
|
2,0
|
5,2
|
8,9
|
15,7
|
15,2
|
22,9
|
19,9
|
14,9
|
7,7
|
-7,3
|
1,2
|
| 30
|
1,9
|
-
|
6,2
|
8,9
|
16,3
|
14,7
|
22,1
|
21,0
|
12,3
|
8,6
|
-6,4
|
0,4
|
| 31
|
-1,6
|
-
|
6,4
|
-
|
16,3
|
-
|
22,8
|
19,7
|
-
|
9,9
|
-
|
-1,8
|
| Сумма
|
-25,2
|
-33,6
|
80,9
|
103,6
|
151,9
|
182,8
|
235,7
|
210,4
|
131,9
|
96,1
|
-65,0
|
-21,3
|
| Сумма за месяц
|
-108,5
|
-121,3
|
88,0
|
231,7
|
390,3
|
487,9
|
581,0
|
585,8
|
413,7
|
232,1
|
10,6
|
-56,3
|
| Сред-няя за месяц
|
-3,5
|
-4,2
|
2,8
|
7,7
|
12,6
|
16,3
|
18,7
|
18,9
|
13,8
|
7,5
|
0,4
|
-1,8
|
Таблица 5.9
Ежемесячная сумма осадков в 2004 г.
| Число
|
Месяц
|
| 01
|
02
|
03
|
04
|
05
|
06
|
07
|
08
|
09
|
10
|
11
|
12
|
| 1
|
-
|
-
|
0,2
|
0,2
|
15,8
|
2,5
|
4,9
|
1,0
|
-
|
0,2
|
-
|
0,0
|
| 2
|
-
|
-
|
0,6
|
-
|
-
|
-
|
0,3
|
-
|
7,1
|
3,1
|
-
|
1,0
|
| 3
|
5,1
|
0,0
|
-
|
0,3
|
-
|
-
|
1,8
|
-
|
-
|
-
|
-
|
1,9
|
| 4
|
0,7
|
0,0
|
0,9
|
-
|
-
|
0,2
|
16,3
|
1,7
|
5,3
|
-
|
-
|
-
|
| 5
|
-
|
5,8
|
0,1
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
| 6
|
2,6
|
2,8
|
0,5
|
-
|
-
|
-
|
2,5
|
-
|
-
|
-
|
-
|
1,0
|
| 7
|
2,7
|
2,2
|
0,5
|
0,7
|
2,5
|
-
|
0,7
|
1,6
|
-
|
-
|
-
|
0,5
|
| 8
|
-
|
0,0
|
-
|
3,2
|
3,6
|
-
|
-
|
9,4
|
-
|
-
|
0,0
|
1,3
|
| 9
|
-
|
0,0
|
-
|
13,9
|
-
|
1,3
|
-
|
11,5
|
3,8
|
-
|
-
|
2,1
|
| 10
|
0,5
|
1,8
|
-
|
0,7
|
-
|
0,0
|
-
|
2,8
|
14,7
|
4,5
|
-
|
0,7
|
| Сумма
|
11,6
|
12,6
|
2,8
|
19,9
|
21,9
|
4,0
|
26,5
|
28,0
|
30,9
|
7,8
|
0,0
|
8,5
|
| 11
|
0,6
|
2,0
|
-
|
-
|
7,5
|
0,0
|
-
|
-
|
-
|
-
|
-
|
0,5
|
| 12
|
-
|
2,9
|
-
|
8,6
|
5,4
|
-
|
12,4
|
-
|
-
|
0,0
|
-
|
2,5
|
| 13
|
2,4
|
1,5
|
-
|
-
|
0,0
|
-
|
4,0
|
-
|
-
|
-
|
-
|
0,6
|
| 14
|
-
|
7,1
|
-
|
-
|
-
|
-
|
20,9
|
19,3
|
-
|
-
|
5,9
|
2,2
|
| 15
|
6,4
|
4,2
|
-
|
-
|
-
|
0,0
|
10,9
|
-
|
-
|
-
|
2,8
|
0,2
|
| 16
|
25,1
|
1,5
|
-
|
-
|
-
|
11,4
|
2,8
|
-
|
-
|
-
|
1,6
|
-
|
| 17
|
5,1
|
0,2
|
5,2
|
-
|
-
|
-
|
1,6
|
-
|
0,4
|
8,0
|
0,0
|
-
|
| 18
|
-
|
4,0
|
1,3
|
-
|
0,6
|
-
|
0,3
|
-
|
-
|
9,0
|
4,8
|
-
|
| 19
|
2,2
|
6,9
|
8,8
|
-
|
-
|
1,9
|
0,0
|
0,0
|
-
|
-
|
3,9
|
-
|
| 20
|
1,0
|
2,9
|
6,8
|
-
|
0,8
|
-
|
-
|
-
|
-
|
-
|
7,4
|
9,5
|
| Сумма
|
42,8
|
33,2
|
22,1
|
8,6
|
14,3
|
13,3
|
52,9
|
19,3
|
0,4
|
17,0
|
26,4
|
15,5
|
| 21
|
6,9
|
-
|
20,5
|
-
|
-
|
-
|
-
|
14,6
|
0,0
|
1,3
|
-
|
5,4
|
| 22
|
-
|
-
|
-
|
-
|
5,7
|
43,3
|
0,3
|
-
|
0,8
|
1,0
|
-
|
0,2
|
| 23
|
15,8
|
-
|
-
|
-
|
4,8
|
-
|
0,2
|
-
|
2,1
|
-
|
-
|
0,9
|
| 24
|
-
|
0,3
|
-
|
-
|
7,0
|
5,4
|
-
|
5,5
|
8,1
|
5,2
|
-
|
-
|
| 25
|
3,2
|
0,6
|
-
|
1,5
|
18,1
|
-
|
-
|
-
|
-
|
1,8
|
-
|
2,7
|
| 26
|
0,2
|
-
|
2,9
|
7,7
|
12,2
|
-
|
-
|
-
|
4,9
|
-
|
0,6
|
-
|
| 27
|
-
|
0,6
|
5,0
|
1,3
|
-
|
-
|
-
|
-
|
14,0
|
0,2
|
-
|
-
|
| 28
|
0,4
|
0,0
|
8,6
|
-
|
-
|
2,8
|
-
|
-
|
21,1
|
0,2
|
-
|
0,5
|
| 29
|
4,2
|
0,0
|
1,3
|
0,0
|
1,2
|
7,2
|
8,4
|
-
|
-
|
-
|
0,0
|
0,7
|
| 30
|
-
|
-
|
-
|
3,6
|
3,0
|
9,5
|
-
|
-
|
13,7
|
0,2
|
1,9
|
0,4
|
| 31
|
11,2
|
-
|
-
|
-
|
2,3
|
-
|
-
|
-
|
-
|
0,0
|
-
|
-
|
| Сумма
|
41,9
|
1,5
|
38,3
|
14,1
|
54,3
|
68,2
|
8,9
|
65,1
|
64,7
|
9,9
|
2,5
|
10,8
|
| Сумма за месяц
|
96,3
|
47,4
|
63,2
|
41,7
|
90,5
|
85,5
|
88,3
|
112,4
|
96,0
|
34,7
|
28,9
|
34,8
|
| Итого за год
|
819,7
|
Таблица 5.10
Экстремальные значения температуры воздуха в 2004 г.
| Число
|
Январь
|
Февраль
|
Март
|
Апрель
|
| макс
|
мин
|
макс
|
мин
|
макс
|
мин
|
макс
|
мин
|
| 1
|
-2,5
|
-5,5
|
-3,0
|
-8,0
|
7,6
|
1,0
|
8,6
|
-1,0
|
| 2
|
-1,4
|
-4,0
|
-2,5
|
-12,0
|
1,0
|
-2,4
|
-1,5
|
-0,6
|
| 3
|
-2,6
|
0,0
|
-3,5
|
-7,0
|
4,0
|
-4,6
|
-0,6
|
-8,4
|
| 4
|
-10,0
|
0,0
|
-2,5
|
-7,6
|
0,5
|
-4,5
|
6,0
|
-3,5
|
| 5
|
-6,7
|
-9,5
|
0,0
|
-4,0
|
-4,0
|
-7,0
|
12,7
|
-1,5
|
| 6
|
-7,5
|
-11,4
|
3,0
|
-0,3
|
-1,7
|
-12,2
|
15,0
|
1,0
|
| 7
|
-8,4
|
-10,5
|
4,6
|
1,0
|
-0,5
|
-5,5
|
13,2
|
4,2
|
| 8
|
-3,7
|
-9,5
|
4,9
|
1,0
|
5,5
|
-6,6
|
13,5
|
5,3
|
| 9
|
-3,5
|
-8,0
|
2,6
|
-1,0
|
7,0
|
-4,1
|
13,5
|
5,2
|
| 10
|
-3,0
|
-7,5
|
3,1
|
-5,5
|
4,0
|
-2,5
|
12,0
|
-1,5
|
| 11
|
-0,7
|
-5,5
|
-3,0
|
10,0
|
2,0
|
-2,2
|
12,5
|
3,0
|
| 12
|
-3,0
|
-7,0
|
-3,0
|
16,0
|
4,0
|
6,9
|
10,2
|
7,5
|
| 13
|
-1,0
|
-4,5
|
-10,0
|
-21,5
|
3,5
|
0,0
|
11,8
|
1,7
|
| 14
|
0,5
|
-3,0
|
-1,3
|
-11,0
|
7,2
|
1,0
|
11,0
|
0,5
|
| 15
|
2,5
|
1,5
|
0,2
|
-4,5
|
5,5
|
0,3
|
15,0
|
1,0
|
| 16
|
0,0
|
-3,7
|
-2,8
|
-8,1
|
5,5
|
0,2
|
17,0
|
6,2
|
| 17
|
-0,5
|
-7,0
|
-1,5
|
-5,0
|
1,2
|
0,3
|
23,5
|
1,0
|
| 18
|
2,6
|
-1,0
|
-2,0
|
-7,8
|
6,2
|
0,0
|
14,0
|
-0,5
|
| 19
|
2,6
|
-7,0
|
1,5
|
-9,5
|
11,5
|
3,0
|
16,6
|
3,0
|
| 20
|
-5,0
|
-12,0
|
-10,0
|
-15,0
|
6,2
|
1,0
|
16,7
|
5,5
|
| 21
|
0,0
|
-5,0
|
-7,5
|
-10,8
|
10,5
|
5,5
|
19,0
|
6,5
|
| 22
|
-4,0
|
-9,0
|
-6,0
|
18,0
|
12,1
|
5,6
|
21,5
|
6,5
|
| 23
|
-3,5
|
-9,3
|
-2,5
|
16,5
|
14,5
|
1,0
|
20,1
|
8,0
|
| 24
|
0,0
|
-4,0
|
-1,0
|
-10,1
|
15,9
|
5,5
|
18,5
|
8,5
|
| 25
|
0,0
|
-2,5
|
2,1
|
-1,5
|
14,5
|
8,1
|
17,0
|
5,5
|
| 26
|
0,9
|
-2,0
|
2,5
|
-0,7
|
12,0
|
4,7
|
16,0
|
7,5
|
| 27
|
-2,8
|
-6,2
|
2,8
|
0,0
|
6,0
|
3,9
|
8,5
|
3,5
|
| 28
|
-0,5
|
-5,0
|
5,2
|
0,4
|
7,2
|
4,6
|
12,0
|
-0,7
|
| 29
|
4,8
|
-3,0
|
4,0
|
0,3
|
7,3
|
4,0
|
15,0
|
6,1
|
| 30
|
4,8
|
1,4
|
11,0
|
3,5
|
17,0
|
-0,5
|
| 31
|
2,0
|
-6,8
|
12,0
|
0,6
|
| Сумма
за месяц
|
-49,6
|
-169,5
|
-25,6
|
-218,7
|
199,2
|
-4,7
|
405,3
|
69,1
|
| Средняя за месяц
|
-1,6
|
-5,5
|
-0,9
|
-7,5
|
6,4
|
-0,2
|
13,5
|
2,3
|
Продолжение таблицы 5.10
Экстремальные значения температуры воздуха в 2004 г.
| Число
|
Май
|
Июнь
|
Июль
|
Август
|
| макс
|
мин
|
макс
|
мин
|
макс
|
мин
|
макс
|
Мин
|
| 1
|
10,8
|
5,5
|
17,1
|
7,0
|
17,2
|
11,5
|
30,5
|
18,0
|
| 2
|
17,0
|
4,5
|
19,0
|
8,0
|
23,5
|
13,0
|
29,0
|
18,5
|
| 3
|
12,5
|
8,5
|
18,0
|
6,6
|
18,5
|
13,0
|
26,5
|
18,5
|
| 4
|
20,5
|
8,0
|
19,5
|
6,2
|
21,3
|
12,6
|
22,0
|
16,5
|
| 5
|
23,5
|
11,0
|
22,0
|
6,0
|
23,4
|
11,5
|
25,1
|
15,5
|
| 6
|
22,0
|
10,5
|
23,0
|
9,5
|
23,0
|
12,0
|
26,5
|
15,0
|
| 7
|
18,0
|
9,5
|
23,5
|
10,5
|
23,1
|
14,5
|
19,5
|
16,0
|
| 8
|
18,5
|
9,7
|
25,5
|
10,0
|
22,3
|
11,7
|
24,0
|
16,5
|
| 9
|
22,0
|
6,0
|
20,5
|
9,5
|
23,3
|
11,0
|
23,0
|
15,5
|
| 10
|
23,0
|
10,5
|
11,0
|
6,0
|
26,5
|
12,7
|
24,5
|
18,1
|
| 11
|
21,5
|
9,0
|
19,0
|
7,1
|
25,5
|
13,0
|
24,5
|
16,0
|
| 12
|
15,5
|
10,5
|
24,0
|
8,0
|
22,5
|
15,5
|
21,5
|
12,2
|
| 13
|
14,0
|
7,8
|
24,5
|
13,0
|
24,2
|
15,8
|
25,2
|
12,0
|
| 14
|
12,9
|
2,5
|
23,6
|
11,5
|
24,6
|
15,0
|
24,5
|
14,7
|
| 15
|
18,0
|
0,5
|
20,0
|
10,6
|
21,0
|
15,5
|
19,5
|
11,5
|
| 16
|
16,5
|
2,0
|
22,8
|
11,5
|
17,8
|
11,8
|
23,5
|
6,4
|
| 17
|
18,5
|
3,6
|
17,7
|
9,5
|
23,0
|
12,0
|
25,0
|
11,5
|
| 18
|
13,0
|
6,0
|
21,5
|
5,5
|
24,0
|
11,5
|
25,5
|
12,1
|
| 19
|
17,5
|
3,0
|
23,5
|
13,5
|
23,2
|
11,5
|
26,1
|
14,0
|
| 20
|
15,0
|
4,5
|
24,2
|
12,5
|
25,5
|
12,0
|
28,5
|
12,5
|
| 21
|
17,5
|
4,8
|
28,5
|
14,1
|
25,5
|
14,0
|
29,0
|
17,0
|
| 22
|
20,0
|
6,5
|
27,0
|
15,7
|
24,0
|
14,0
|
29,0
|
17,5
|
| 23
|
22,5
|
11,1
|
22,5
|
14,8
|
23,5
|
15,0
|
29,0
|
17,0
|
| 24
|
20,2
|
8,6
|
25,0
|
16,5
|
26,3
|
15,5
|
19,0
|
15,5
|
| 25
|
14,5
|
8,5
|
23,1
|
16,3
|
28,0
|
16,0
|
22,0
|
11,0
|
| 26
|
17,0
|
7,4
|
27,0
|
15,0
|
29,1
|
16,5
|
24,0
|
10,5
|
| 27
|
21,5
|
5,7
|
22,2
|
14,3
|
29,5
|
17,0
|
23,8
|
12,0
|
| 28
|
22,5
|
7,8
|
22,0
|
10,5
|
30,6
|
18,5
|
27,0
|
13,0
|
| 29
|
22,5
|
9,2
|
22,0
|
9,0
|
30,6
|
18,0
|
27,5
|
13,8
|
| 30
|
20,5
|
13,5
|
21,0
|
12,0
|
29,0
|
15,3
|
28,5
|
14,5
|
| 31
|
21,5
|
14,0
|
30,5
|
17,5
|
28,2
|
14,0
|
| Сумма
за месяц
|
570,4
|
230,2
|
660,2
|
320,2
|
760,0
|
434,4
|
781,4
|
446,8
|
| Средняя за месяц
|
18,4
|
7,4
|
22,0
|
10,7
|
24,5
|
14,0
|
25,2
|
14,4
|
Окончание таблицы 5.10
Экстремальные значения температуры воздуха в 2004 г.
| Число
|
Сентябрь
|
Октябрь
|
Ноябрь
|
Декабрь
|
| макс
|
мин
|
макс
|
мин
|
Макс
|
мин
|
макс
|
Мин
|
| 1
|
23,0
|
14,0
|
14,8
|
12,0
|
13,0
|
5,0
|
-5,5
|
-16,0
|
| 2
|
23,0
|
16,5
|
17,5
|
8,0
|
9,0
|
4,5
|
1,0
|
-9,0
|
| 3
|
22,0
|
11,5
|
12,4
|
4,4
|
8,0
|
-0,5
|
2,5
|
0,5
|
| 4
|
20,0
|
12,5
|
8,0
|
4,5
|
11,5
|
-2,2
|
2,5
|
0,5
|
| 5
|
23,0
|
8,5
|
9,5
|
2,5
|
13,0
|
-0,5
|
3,0
|
-0,5
|
| 6
|
18,5
|
5,2
|
14,1
|
1,0
|
12,0
|
1,0
|
4,5
|
0,0
|
| 7
|
22,0
|
5,5
|
14,0
|
1,7
|
12,2
|
1,0
|
1,0
|
-0,2
|
| 8
|
19,0
|
9,0
|
17,0
|
2,4
|
2,5
|
-2,5
|
3,0
|
0,0
|
| 9
|
17,5
|
5,6
|
16,5
|
1,5
|
9,5
|
1,5
|
2,8
|
1,0
|
| 10
|
14,0
|
6,5
|
11,5
|
8,3
|
9,5
|
6,0
|
1,7
|
-2,0
|
| 11
|
15,5
|
3,5
|
11,5
|
2,0
|
8,5
|
3,0
|
0,0
|
-4,0
|
| 12
|
17,5
|
7,0
|
6,5
|
-1,1
|
7,0
|
3,0
|
1,5
|
-2,0
|
| 13
|
20,8
|
7,3
|
9,0
|
-1,3
|
10,0
|
4,0
|
0,0
|
-3,0
|
| 14
|
22,0
|
11,8
|
8,5
|
-3,1
|
10,5
|
7,0
|
-4,8
|
-9,0
|
| 15
|
23,5
|
9,0
|
8,0
|
1,0
|
8,5
|
1,6
|
-7,8
|
-12,0
|
| 16
|
24,5
|
10,0
|
11,0
|
0,0
|
4,0
|
1,4
|
-2,0
|
-10,5
|
| 17
|
16,1
|
10,2
|
13,5
|
2,0
|
3,0
|
-2,1
|
-1,3
|
-6,5
|
| 18
|
18,0
|
7,5
|
15,0
|
6,5
|
6,5
|
-4,0
|
3,0
|
0,5
|
| 19
|
21,0
|
3,5
|
14,5
|
3,0
|
6,5
|
-4,5
|
5,0
|
0,5
|
| 20
|
24,2
|
7,5
|
15,5
|
4,5
|
9,5
|
-6,5
|
0,0
|
-17,
|
| 21
|
24,0
|
9,5
|
8,0
|
3,5
|
1,0
|
-5,5
|
1,0
|
-2,5
|
| 22
|
17,1
|
9,0
|
10,5
|
2,0
|
-2,5
|
-8,0
|
-3,0
|
-8,5
|
| 23
|
14,8
|
7,0
|
11,5
|
0,0
|
-1,4
|
-11,5
|
-5,5
|
-14,5
|
| 24
|
12,0
|
6,2
|
14,0
|
2,5
|
-1,5
|
-8,3
|
-3,0
|
-13,0
|
| 25
|
13,5
|
5,0
|
16,5
|
11,3
|
-3,0
|
-11,0
|
1,1
|
-3,5
|
| 26
|
22,0
|
12,0
|
19,5
|
5,5
|
-2,5
|
-14,0
|
3,0
|
-5,5
|
| 27
|
17,5
|
14,6
|
12,6
|
9,0
|
-0,3
|
-9,8
|
4,0
|
1,0
|
| 28
|
19,1
|
13,5
|
12,0
|
8,7
|
0,0
|
-8,5
|
4,5
|
1,0
|
| 29
|
18,9
|
9,1
|
13,0
|
1,8
|
-4,0
|
-9,5
|
4,5
|
0,5
|
| 30
|
18,5
|
11,7
|
19,5
|
7,5
|
-3,5
|
-7,5
|
1,0
|
2,0
|
| 31
|
-
|
-
|
12,0
|
8,0
|
0,0
|
-3,5
|
| Сумма
за месяц
|
582,5
|
269,7
|
397,4
|
119,6
|
156,5
|
-77,4
|
17,7
|
-123,9
|
| Средняя за месяц
|
19,4
|
9,0
|
12,8
|
3,8
|
5,2
|
-2,6
|
0,6
|
-4,0
|
Таблица 5.11
Высота снежного покрова (см.) по трем постоянным рейкам (метеоплощадка)
зимой 2003-2004 г.
| Даты
|
Декабрь
|
Январь
|
Февраль
|
Март
|
| I
|
II
|
III
|
ср.
|
I
|
II
|
III
|
ср.
|
I
|
II
|
III
|
ср.
|
I
|
II
|
III
|
ср.
|
| 1
|
5
|
6
|
5
|
5
|
21
|
20
|
20
|
20
|
8
|
8
|
8
|
8
|
| 2
|
5
|
6
|
5
|
5
|
21
|
20
|
20
|
20
|
8
|
8
|
8
|
8
|
| 3
|
5
|
6
|
5
|
5
|
21
|
20
|
20
|
20
|
5
|
6
|
5
|
5
|
| 4
|
12
|
13
|
12
|
12
|
21
|
20
|
20
|
20
|
6
|
7
|
6
|
6
|
| 5
|
11
|
12
|
11
|
11
|
21
|
20
|
20
|
20
|
6
|
7
|
6
|
6
|
| 6
|
12
|
13
|
13
|
13
|
19
|
18
|
18
|
18
|
7
|
7
|
7
|
7
|
| 7
|
12
|
13
|
13
|
13
|
16
|
15
|
15
|
15
|
6
|
6
|
6
|
6
|
| 8
|
14
|
15
|
16
|
15
|
17
|
16
|
16
|
16
|
12
|
11
|
12
|
12
|
5
|
6
|
5
|
5
|
| 9
|
14
|
15
|
16
|
15
|
16
|
15
|
15
|
15
|
10
|
10
|
10
|
10
|
5
|
6
|
5
|
5
|
| 10
|
14
|
14
|
15
|
14
|
16
|
15
|
15
|
15
|
10
|
10
|
10
|
10
|
5
|
6
|
5
|
5
|
| Сумма
|
42
|
44
|
47
|
44
|
111
|
115
|
110
|
110
|
172
|
164
|
165
|
165
|
61
|
67
|
61
|
61
|
| 11
|
14
|
14
|
15
|
14
|
16
|
16
|
16
|
16
|
10
|
10
|
10
|
10
|
5
|
6
|
5
|
5
|
| 12
|
14
|
14
|
15
|
14
|
16
|
16
|
16
|
16
|
11
|
12
|
12
|
12
|
5
|
6
|
5
|
5
|
| 13
|
13
|
13
|
14
|
13
|
16
|
16
|
16
|
16
|
13
|
14
|
13
|
13
|
1
|
2
|
2
|
2
|
| 14
|
10
|
11
|
12
|
11
|
17
|
16
|
17
|
17
|
23
|
22
|
24
|
23
|
1
|
2
|
2
|
2
|
| 15
|
-
|
1
|
1
|
1
|
16
|
15
|
16
|
16
|
24
|
23
|
24
|
24
|
1
|
1
|
1
|
1
|
| 16
|
-
|
-
|
-
|
-
|
14
|
13
|
13
|
13
|
24
|
23
|
24
|
24
|
1
|
1
|
1
|
1
|
| 17
|
1
|
2
|
1
|
1
|
24
|
26
|
25
|
25
|
23
|
23
|
23
|
23
|
1
|
1
|
1
|
1
|
| 18
|
1
|
2
|
1
|
1
|
24
|
26
|
25
|
25
|
23
|
23
|
23
|
23
|
| 19
|
1
|
2
|
1
|
1
|
13
|
17
|
14
|
15
|
28
|
30
|
29
|
29
|
| 20
|
2
|
2
|
1
|
2
|
13
|
14
|
14
|
14
|
28
|
30
|
29
|
29
|
| Сумма
|
56
|
61
|
61
|
58
|
169
|
175
|
172
|
173
|
207
|
210
|
211
|
210
|
15
|
19
|
17
|
17
|
| 21
|
2
|
2
|
1
|
2
|
21
|
22
|
21
|
21
|
30
|
31
|
30
|
30
|
| 22
|
-
|
-
|
-
|
-
|
21
|
22
|
21
|
21
|
30
|
31
|
30
|
30
|
| 23
|
-
|
-
|
-
|
-
|
27
|
28
|
26
|
27
|
30
|
31
|
30
|
30
|
| 24
|
10
|
12
|
12
|
12
|
32
|
32
|
31
|
32
|
28
|
29
|
28
|
28
|
| 25
|
10
|
12
|
12
|
12
|
32
|
32
|
31
|
32
|
28
|
29
|
28
|
28
|
| 26
|
9
|
10
|
10
|
10
|
35
|
35
|
34
|
35
|
22
|
23
|
21
|
22
|
| 27
|
9
|
10
|
10
|
10
|
35
|
35
|
34
|
35
|
20
|
21
|
20
|
20
|
| 28
|
9
|
10
|
10
|
10
|
34
|
34
|
34
|
34
|
15
|
14
|
16
|
15
|
| 29
|
8
|
8
|
8
|
8
|
29
|
28
|
27
|
28
|
11
|
10
|
10
|
10
|
| 30
|
6
|
7
|
6
|
6
|
19
|
18
|
17
|
18
|
| 31
|
5
|
6
|
5
|
5
|
23
|
22
|
22
|
22
|
| Сумма
|
68
|
77
|
74
|
75
|
308
|
308
|
298
|
305
|
214
|
219
|
213
|
213
|
| Сумма за м-ц
|
166
|
182
|
182
|
177
|
588
|
598
|
580
|
588
|
593
|
593
|
589
|
588
|
76
|
86
|
78
|
78
|
| Сред–
няя
|
7
|
19
|
20
|
5
|
Таблица 5.12
Средняя температура поверхности почвы в 2004 г.
| Число
|
Месяц
|
| 01
|
02
|
03
|
04
|
05
|
06
|
07
|
08
|
09
|
10
|
11
|
12
|
| 1
|
-2,5
|
-2,5
|
0,8
|
6,1
|
8,1
|
14,0
|
15,6
|
25,8
|
20,4
|
13,5
|
8,0
|
-5,6
|
| 2
|
-2,2
|
-4,6
|
-1,0
|
-0,7
|
13,8
|
17,8
|
21,6
|
25,9
|
19,1
|
12,0
|
6,0
|
-4,3
|
| 3
|
-0,9
|
-3,9
|
-0,7
|
1,3
|
9,8
|
19,5
|
17,0
|
24,9
|
18,6
|
9,2
|
2,2
|
0,8
|
| 4
|
-4,0
|
-3,3
|
-2,2
|
3,4
|
14,1
|
15,8
|
18,7
|
18,6
|
20,6
|
7,0
|
2,8
|
0,5
|
| 5
|
-5,3
|
-1,4
|
-2,9
|
6,1
|
15,6
|
23,4
|
21,1
|
20,5
|
11,0
|
7,5
|
4,3
|
-0,1
|
| 6
|
-5,1
|
0,2
|
-3,0
|
8,8
|
18,1
|
17,9
|
20,0
|
19,9
|
17,6
|
7,8
|
4,2
|
1,0
|
| 7
|
-6,6
|
0,0
|
-1,6
|
10,0
|
13,5
|
24,0
|
16,9
|
18,2
|
17,7
|
9,5
|
4,1
|
0,1
|
| 8
|
-4,9
|
0,4
|
-2,9
|
8,9
|
13,4
|
27,5
|
21,4
|
18,0
|
15,6
|
10,5
|
0,9
|
1,0
|
| 9
|
4,6
|
0,0
|
-2,1
|
9,3
|
15,3
|
20,1
|
22,4
|
18,6
|
13,5
|
10,2
|
5,9
|
1,2
|
| 10
|
-3,9
|
-1,9
|
-0,8
|
9,0
|
16,2
|
11,4
|
25,5
|
19,5
|
12,2
|
9,0
|
6,7
|
-0,1
|
| Сумма
|
-40,0
|
-17,0
|
-16,4
|
62,2
|
137,9
|
191,4
|
200,2
|
209,9
|
166,3
|
96,2
|
45,1
|
-5,5
|
| 11
|
-3,6
|
-4,8
|
-0,1
|
9,5
|
17,0
|
19,2
|
23,2
|
22,1
|
12.2
|
5,8
|
4,9
|
-1,9
|
| 12
|
-4,7
|
-9,4
|
1,6
|
8,4
|
14,5
|
24,1
|
20,2
|
19,7
|
11,9
|
0,7
|
4,4
|
-0,6
|
| 13
|
-2,2
|
-14,0
|
0,7
|
9,6
|
10,1
|
21,9
|
22,6
|
18,4
|
14,2
|
4,0
|
4,0
|
-0,7
|
| 14
|
-2,1
|
-3,0
|
3,6
|
7,8
|
12,6
|
27,2
|
18,8
|
18,2
|
17,2
|
3,4
|
8,0
|
-4,2
|
| 15
|
-0,3
|
-2,0
|
2,8
|
10,8
|
14,6
|
17,5
|
20,2
|
17,5
|
18,7
|
4,6
|
3,8
|
-4,8
|
| 16
|
-1,8
|
-3,6
|
3,3
|
12,0
|
15,4
|
20,4
|
16,4
|
15,8
|
18,4
|
4,8
|
2,9
|
-5,0
|
| 17
|
-4,5
|
-2,2
|
0,4
|
9,4
|
15,7
|
18,1
|
22,2
|
19,8
|
15,4
|
5,4
|
0,2
|
-3,5
|
| 18
|
0,0
|
-4,6
|
4,4
|
9,4
|
14,3
|
20,4
|
19,0
|
19,9
|
14,4
|
10,0
|
-0,6
|
0,2
|
| 19
|
-0,3
|
-0,4
|
8,2
|
11,1
|
17,6
|
22,6
|
21,8
|
18,4
|
15,7
|
7,2
|
0,9
|
0,9
|
| 20
|
-7,0
|
-9,4
|
4,6
|
9,9
|
17,9
|
22,7
|
22,9
|
17,0
|
17,7
|
8,0
|
0,6
|
-0,5
|
| Сумма
|
-29,2
|
-53,4
|
29,5
|
97,9
|
149,7
|
214,1
|
207,3
|
186,8
|
155,8
|
53,9
|
29,1
|
-20,1
|
| 21
|
-2,5
|
-12,0
|
8,1
|
14,9
|
14,0
|
27,1
|
24,9
|
23,4
|
18,6
|
4,9
|
-4,4
|
-1,2
|
| 22
|
-6,0
|
14,0
|
6,4
|
12,9
|
13,9
|
27,3
|
23,1
|
123,6
|
14,0
|
1,0
|
-5,8
|
-5,5
|
| 23
|
-5,6
|
-12,0
|
8,0
|
15,4
|
11,4
|
21,8
|
20,8
|
26,0
|
11,9
|
6,2
|
-5,8
|
-10,8
|
| 24
|
-1,2
|
-4,4
|
9,0
|
15,9
|
13,6
|
22,3
|
24,5
|
17,4
|
9,8
|
6,8
|
-5,9
|
-6,1
|
| 25
|
-1,1
|
0,1
|
8,2
|
10,2
|
12,1
|
22,5
|
29,7
|
17,7
|
9,5
|
11,4
|
-8,1
|
-0,1
|
| 26
|
-1,2
|
0,2
|
7,6
|
10,0
|
12,2
|
26,1
|
28,4
|
16,8
|
16,6
|
11,6
|
-8,8
|
-2,2
|
| 27
|
-4,5
|
0,2
|
5,5
|
6,3
|
17,2
|
18,6
|
27,3
|
15,0
|
15,1
|
10,5
|
-3,8
|
0,1
|
| 28
|
-2,7
|
1,2
|
6,4
|
8,0
|
17,6
|
21,6
|
27,8
|
19,9
|
15,6
|
10,0
|
-5,6
|
0,8
|
| 29
|
0,4
|
0,5
|
6,4
|
11,6
|
18,9
|
18,2
|
26,8
|
20,0
|
16,0
|
8,2
|
-4,5
|
0,6
|
| 30
|
0,1
|
6,0
|
12,4
|
17,4
|
16,2
|
25,1
|
22,4
|
13,0
|
9,1
|
-10,5
|
-0,1
|
| 31
|
-1,1
|
5,3
|
17,3
|
25,2
|
22,4
|
9,8
|
-1,2
|
| Сумма
|
-25,4
|
-40,2
|
76,9
|
177,6
|
165,6
|
221,7
|
283,6
|
224,6
|
140,1
|
89,5
|
-63,2
|
-25,7
|
| Сумма за месяц
|
-94,6
|
-110,6
|
90,0
|
277,7
|
453,2
|
627,2
|
691,1
|
621,3
|
462,2
|
239,6
|
11,0
|
-51,3
|
| Средняя за месяц
|
-3,0
|
-3,8
|
2,9
|
9,3
|
14,6
|
20,9
|
22,3
|
20,0
|
15,4
|
7,7
|
0,4
|
-1,6
|
Таблица 5.13
Средняя температура почвы на глубине 5 см в 2004 г.
| Число
|
Месяц
|
| 01
|
02
|
03
|
04
|
05
|
06
|
07
|
08
|
09
|
10
|
11
|
12
|
| 1
|
14,6
|
16,8
|
24,6
|
19,0
|
13,9
|
9,2
|
| 2
|
15,8
|
19,6
|
24,8
|
18,5
|
13,0
|
7,2
|
| 3
|
16,3
|
17,4
|
24,3
|
19,3
|
8,9
|
4,2
|
| 4
|
14,7
|
19,4
|
21,4
|
18,4
|
8,7
|
4,1
|
| 5
|
19,6
|
19,6
|
20,5
|
16,0
|
8,7
|
3,8
|
| 6
|
20,0
|
18,8
|
22,5
|
16,6
|
8,0
|
4,2
|
| 7
|
21,6
|
19,9
|
19,9
|
17,3
|
8,8
|
4,3
|
| 8
|
22,5
|
20,9
|
18,9
|
16,7
|
9,3
|
2,7
|
| 9
|
20,4
|
22,6
|
19,2
|
14,1
|
9,9
|
5,4
|
| 10
|
14,2
|
24,7
|
20,3
|
13,3
|
11,6
|
6,7
|
| Сумма
|
179,7
|
199,7
|
216,4
|
169,2
|
100,8
|
51,8
|
| 11
|
16,2
|
24,1
|
23,0
|
12,6
|
8,2
|
5,9
|
| 12
|
19,7
|
20,9
|
20,5
|
13,1
|
4,3
|
5,2
|
| 13
|
20,3
|
22,4
|
21,0
|
15,1
|
5,2
|
6,1
|
| 14
|
12,7
|
21,8
|
19,8
|
19,6
|
17,4
|
4,2
|
7,4
|
| 15
|
13,6
|
18,8
|
20,1
|
19,2
|
18,0
|
4,0
|
5,2
|
| 16
|
14,5
|
18,0
|
17,1
|
19,0
|
18,2
|
5,4
|
4,8
|
| 17
|
13,9
|
17,4
|
19,9
|
21,3
|
15,5
|
5,2
|
2,1
|
| 18
|
13,9
|
18,6
|
20,5
|
21,4
|
15,2
|
7,7
|
0,6
|
| 19
|
15,4
|
20,4
|
21,2
|
21,1
|
14,7
|
6,8
|
| 20
|
13,1
|
21,1
|
22,3
|
19,5
|
15,5
|
7,1
|
| Сумма
|
97,1
|
192,3
|
208,3
|
205,6
|
155,3
|
58,1
|
37,3
|
| 21
|
13,6
|
23,9
|
23,7
|
23,1
|
16,9
|
7,8
|
| 22
|
13,6
|
22,8
|
22,8
|
24,0
|
15,3
|
6,4
|
| 23
|
14,4
|
20,8
|
21,6
|
22,8
|
12,5
|
4,6
|
| 24
|
14,2
|
21,8
|
24,0
|
19,8
|
10,7
|
9,5
|
| 25
|
12,9
|
21,2
|
26,4
|
21,4
|
12,2
|
10,9
|
| 26
|
12,2
|
24,8
|
26,5
|
19,3
|
15,6
|
10,2
|
| 27
|
15,1
|
20,2
|
27,0
|
18,5
|
15,7
|
10,6
|
| 28
|
17,9
|
20,0
|
26,2
|
18,7
|
15,6
|
10,1
|
| 29
|
17,6
|
19,5
|
25,5
|
19,0
|
16,3
|
8,2
|
| 30
|
17,0
|
17,6
|
24,4
|
22,9
|
13,7
|
9,2
|
| 31
|
17,2
|
24,8
|
22,3
|
9,4
|
| Сумма
|
165,7
|
212,6
|
272,9
|
231,8
|
144,5
|
96,9
|
| Сумма за месяц
|
262,8
|
584,6
|
680,9
|
653,8
|
469,0
|
255,8
|
89,1
|
| Средняя за месяц
|
14,6
|
19,5
|
21,9
|
21,1
|
15,6
|
8,2
|
4,9
|
Таблица 5.14
Средняя температура почвы на глубине 10 см в 2004 г.
| Число
|
Месяц
|
| 01
|
02
|
03
|
04
|
05
|
06
|
07
|
08
|
09
|
10
|
11
|
12
|
| 1
|
14,8
|
17,2
|
24,3
|
18,8
|
14,1
|
9,5
|
| 2
|
15,2
|
18,6
|
24,6
|
18,4
|
13,7
|
8,0
|
| 3
|
15,6
|
17,7
|
24,0
|
19,4
|
9,9
|
5,3
|
| 4
|
15,7
|
18,8
|
22.0
|
18,0
|
9,6
|
4,6
|
| 5
|
17,9
|
18,8
|
20,4
|
15,8
|
9,3
|
4,3
|
| 6
|
19,1
|
18,5
|
22,7
|
16,4
|
8,8
|
5,0
|
| 7
|
20,1
|
19,4
|
20,4
|
18,0
|
9,2
|
5,4
|
| 8
|
20,9
|
19,7
|
19,2
|
16,6
|
9,2
|
3,8
|
| 9
|
19,8
|
21,8
|
18,8
|
14,0
|
9,9
|
5,2
|
| 10
|
15,2
|
23,0
|
19,8
|
13,5
|
11,0
|
6,6
|
| Сумма
|
174,3
|
193,5
|
216,2
|
168,9
|
104,7
|
57,7
|
| 11
|
15,8
|
23,2
|
21,8
|
12,7
|
8,7
|
6,4
|
| 12
|
18,4
|
20,8
|
19,9
|
13,0
|
6,0
|
5,8
|
| 13
|
19,4
|
21,6
|
20,9
|
14,6
|
6,2
|
6,1
|
| 14
|
12,1
|
20,5
|
19,9
|
19,8
|
17,3
|
5,5
|
7,2
|
| 15
|
12,5
|
18,6
|
19,7
|
19,4
|
17,3
|
5,0
|
6,1
|
| 16
|
13,6
|
17,9
|
17,4
|
18,8
|
17,7
|
5,6
|
5,3
|
| 17
|
13,5
|
17,1
|
19,2
|
20,6
|
15,6
|
5,6
|
3,3
|
| 18
|
16,9
|
17,9
|
19,8
|
20,9
|
15,1
|
7,3
|
1,8
|
| 19
|
14,4
|
19,3
|
20,4
|
20,9
|
14,8
|
7,4
|
| 20
|
12,9
|
19,9
|
21,2
|
19,9
|
16,3
|
8,0
|
| Сумма
|
95,9
|
184,8
|
203,2
|
202,9
|
154,4
|
65,3
|
42,0
|
| 21
|
13,0
|
22,7
|
22,8
|
22,6
|
16,6
|
7,2
|
| 22
|
13,1
|
22,9
|
22,5
|
23,3
|
16,1
|
6,8
|
| 23
|
13,5
|
20,4
|
21,1
|
19,8
|
13,4
|
5,6
|
| 24
|
13,9
|
20,1
|
23,1
|
19,7
|
11,7
|
8,5
|
| 25
|
12,9
|
20,8
|
24,9
|
21,2
|
11,8
|
10,7
|
| 26
|
11,9
|
23,1
|
25,3
|
19,1
|
14,9
|
10,2
|
| 27
|
13,9
|
20,5
|
25,8
|
18,1
|
15,3
|
10,3
|
| 28
|
16,3
|
19,3
|
25,5
|
18,4
|
15,7
|
10,0
|
| 29
|
16,7
|
18,9
|
24,5
|
19,0
|
15,9
|
8,8
|
| 30
|
16,6
|
18,4
|
23,8
|
22,6
|
13,8
|
9,4
|
| 31
|
16,7
|
24,1
|
22,1
|
9,4
|
| Сумма
|
142,2
|
207,0
|
263,4
|
225,9
|
145,2
|
96,9
|
| Сумма за месяц
|
238,1
|
566,1
|
660,1
|
645,0
|
468,
|
266,9
|
99,7
|
| Средняя за месяц
|
7,7
|
18,9
|
21,3
|
20,8
|
15,6
|
8,6
|
5,5
|
Таблица 5.15
Средняя температура почвы на глубине 15 см в 2004 г.
| Число
|
Месяц
|
| 01
|
02
|
03
|
04
|
05
|
06
|
07
|
08
|
09
|
10
|
11
|
12
|
| 1
|
14,7
|
17,2
|
23,0
|
18,6
|
14,0
|
9,8
|
| 2
|
14,6
|
17,8
|
23,9
|
18,3
|
13,5
|
9,1
|
| 3
|
15,1
|
17,7
|
23,6
|
19,2
|
10,6
|
7,3
|
| 4
|
15,4
|
18,0
|
22,4
|
17,7
|
10,7
|
6,3
|
| 5
|
16,9
|
18,3
|
20,8
|
15,3
|
10,5
|
5,8
|
| 6
|
17,9
|
18,2
|
22,3
|
16,1
|
10,0
|
6,1
|
| 7
|
18,6
|
19,1
|
20,5
|
18,3
|
9,8
|
6,2
|
| 8
|
19,5
|
19,0
|
19,5
|
16,4
|
9,9
|
5,3
|
| 9
|
19,3
|
20,5
|
18,6
|
14,0
|
10,4
|
5,7
|
| 10
|
15,6
|
22,0
|
19,1
|
13,4
|
11,2
|
6,6
|
| Сумма
|
167,6
|
187,8
|
213,7
|
167,3
|
110,6
|
68,2
|
| 11
|
15,4
|
22,0
|
21,6
|
12,4
|
10,1
|
6,5
|
| 12
|
17,1
|
20,6
|
19,7
|
12,7
|
8,0
|
6,5
|
| 13
|
18,6
|
20,9
|
20,2
|
14,1
|
7,6
|
6,5
|
| 14
|
11,7
|
19,5
|
19,9
|
19,7
|
17,0
|
6,9
|
7,2
|
| 15
|
12,2
|
18,2
|
19,2
|
19,2
|
17,1
|
6,4
|
6,9
|
| 16
|
12,9
|
17,5
|
17,4
|
18,3
|
16,9
|
6,6
|
6,0
|
| 17
|
13,1
|
16,7
|
18,4
|
19,8
|
16,4
|
7,0
|
5,0
|
| 18
|
13,0
|
17,4
|
19,1
|
20,6
|
15,1
|
7,5
|
3,7
|
| 19
|
13,5
|
18,4
|
19,3
|
20,4
|
14,9
|
7,8
|
| 20
|
12,8
|
19,0
|
20,8
|
20,4
|
14,5
|
8,2
|
| Сумма
|
89,2
|
177,8
|
197,6
|
199,9
|
151,1
|
76,1
|
48,3
|
| 21
|
12,6
|
20,6
|
21.7
|
21,5
|
16,2
|
8,2
|
| 22
|
12,7
|
21,2
|
21,8
|
22,5
|
16,0
|
7,3
|
| 23
|
13,0
|
19,9
|
21,3
|
19,6
|
14,5
|
6,8
|
| 24
|
12,9
|
20,1
|
21,8
|
19,3
|
13,0
|
8,2
|
| 25
|
12,9
|
20,1
|
23,8
|
21,0
|
12,2
|
9,8
|
| 26
|
11,8
|
21,7
|
24,1
|
19,0
|
14,5
|
9,8
|
| 27
|
13,1
|
20,3
|
24,2
|
17,7
|
14,9
|
10,4
|
| 28
|
15,0
|
19,1
|
24,9
|
18,0
|
15,0
|
9,8
|
| 29
|
15,7
|
18,4
|
24,2
|
19,4
|
15,6
|
9,5
|
| 30
|
15,8
|
17,7
|
23,3
|
21,9
|
14,0
|
9,5
|
| 31
|
16,1
|
23,1
|
21,7
|
9,6
|
| Сумма
|
151,6
|
199,1
|
254,2
|
221,6
|
145,9
|
98,9
|
| Сумма за месяц
|
240,8
|
544,5
|
639,6
|
635,2
|
464,3
|
285,6
|
116,5
|
| Средняя за месяц
|
13,4
|
18,2
|
20,6
|
20,5
|
15,5
|
9,2
|
6,5
|
Таблица 5.16
Средняя температура почвы на глубине 20 см в 2004 г.
| Число
|
Месяц
|
| 01
|
02
|
03
|
04
|
05
|
06
|
07
|
08
|
09
|
10
|
11
|
12
|
| 1
|
14,5
|
18,1
|
22,8
|
18,2
|
13,9
|
9,9
|
| 2
|
14,4
|
17,9
|
23,0
|
18,0
|
13,8
|
9,2
|
| 3
|
14,5
|
17,8
|
22,7
|
18,8
|
11,0
|
7,4
|
| 4
|
14,7
|
18,1
|
22,4
|
17,6
|
10,9
|
6,6
|
| 5
|
15,4
|
18,0
|
20,5
|
15,0
|
10,7
|
5,9
|
| 6
|
17,4
|
18,9
|
22,6
|
16,0
|
10,1
|
6,2
|
| 7
|
17,2
|
18,7
|
20,5
|
17,9
|
9,9
|
6,3
|
| 8
|
18,5
|
19,8
|
19,4
|
16,0
|
10,0
|
5,6
|
| 9
|
18,5
|
20,1
|
18,6
|
13,8
|
10,5
|
5,8
|
| 10
|
15,9
|
21,3
|
18,8
|
13,2
|
11,3
|
6,7
|
| Сумма
|
161,0
|
188,7
|
211,3
|
164,5
|
112,1
|
69,6
|
| 11
|
14,9
|
21,2
|
20,9
|
12,5
|
10,3
|
6,8
|
| 12
|
16,2
|
21,1
|
19,8
|
12,8
|
8,4
|
6,7
|
| 13
|
17,5
|
21,0
|
20,2
|
13,7
|
7,8
|
6,7
|
| 14
|
10,8
|
18,2
|
19,7
|
19,3
|
16,5
|
6,9
|
7,3
|
| 15
|
11,4
|
17,6
|
19,0
|
19,0
|
16,9
|
6,7
|
6,9
|
| 16
|
11,9
|
17,0
|
17,2
|
18,0
|
17,1
|
6,9
|
6,6
|
| 17
|
12,9
|
16,6
|
18,2
|
19,6
|
16,3
|
7,0
|
5,2
|
| 18
|
12,6
|
16,6
|
19,0
|
20,5
|
15,0
|
7,8
|
3,9
|
| 19
|
12,6
|
17,0
|
19,7
|
20,3
|
15,2
|
8,1
|
| 20
|
12,5
|
18,2
|
19,8
|
20,1
|
14,6
|
8,6
|
| Сумма
|
84,7
|
169,8
|
195,9
|
197,7
|
150,6
|
78,5
|
50,1
|
| 21
|
12,1
|
19,7
|
21,6
|
21,0
|
15,9
|
8,4
|
| 22
|
12,7
|
20,3
|
21,5
|
22,3
|
14,9
|
7,8
|
| 23
|
12,0
|
19,5
|
21,1
|
19,5
|
14,6
|
6,6
|
| 24
|
12,4
|
19,9
|
21,6
|
19,5
|
13,0
|
8,3
|
| 25
|
11,6
|
19,4
|
23,5
|
20,6
|
12,1
|
9,8
|
| 26
|
12,2
|
20,4
|
23,2
|
18,6
|
13,9
|
9,8
|
| 27
|
13,8
|
20,0
|
23,8
|
17,1
|
15,0
|
10,5
|
| 28
|
14,6
|
19,0
|
24,2
|
17,6
|
15,1
|
10,2
|
| 29
|
14,9
|
19,6
|
23,6
|
18,7
|
15,7
|
9,7
|
| 30
|
15,1
|
17,9
|
22,6
|
21,9
|
14,2
|
9,5
|
| 31
|
15,0
|
23,0
|
21,6
|
9,6
|
| Сумма
|
146,4
|
195,7
|
249,7
|
218,4
|
144,4
|
100,2
|
| Сумма за месяц
|
231,1
|
526,5
|
634,3
|
627,4
|
459,5
|
290,8
|
119,7
|
| Средняя за месяц
|
12,8
|
17,6
|
20,5
|
20,2
|
15,3
|
9,4
|
6,6
|
Таблица 5.17
Экстремальные температуры поверхности почвы в 2004 г.
| Число
|
Январь
|
Февраль
|
Март
|
Апрель
|
| макс
|
мин
|
макс
|
мин
|
макс
|
мин
|
макс
|
Мин
|
| 1
|
0,5
|
-5,0
|
-0,9
|
-7,5
|
3,0
|
-0,5
|
13,5
|
-3,2
|
| 2
|
-1,2
|
-4,0
|
-0,5
|
-10,0
|
1,0
|
-3,5
|
1,1
|
-7,1
|
| 3
|
0,0
|
-2,5
|
-1,5
|
-6,5
|
5,5
|
-9,5
|
11,6
|
-7,5
|
| 4
|
0,0
|
-6,0
|
-1,1
|
-6,0
|
1,2
|
-7,0
|
11,5
|
-4,0
|
| 5
|
-4,8
|
-7,0
|
0,0
|
-3,7
|
0,5
|
-8,5
|
20,5
|
-4,5
|
| 6
|
-5,0
|
-7,2
|
1,0
|
-1,0
|
2,0
|
-10,0
|
22,3
|
-4,0
|
| 7
|
-4,0
|
-8,5
|
2,6
|
-1,0
|
1,5
|
-8,0
|
20,8
|
3,1
|
| 8
|
-3,0
|
-6,0
|
2,5
|
-1,0
|
3,5
|
-7,0
|
15,5
|
1,5
|
| 9
|
-3,0
|
-9,0
|
1,5
|
-3,0
|
4,0
|
-8,5
|
20,2
|
1,5
|
| 10
|
-1,6
|
-7,0
|
2,5
|
-4,6
|
6,0
|
-6,0
|
19,5
|
-2,0
|
| 11
|
-0,5
|
-8,5
|
-1,6
|
-10,5
|
2,2
|
-2,5
|
19,5
|
-0,5
|
| 12
|
-2,5
|
-6,5
|
-1,5
|
-19,5
|
11,0
|
-7,3
|
13,0
|
4,0
|
| 13
|
0,5
|
-4,0
|
-5,0
|
-22,5
|
5,0
|
-2,0
|
21,4
|
-3,5
|
| 14
|
0,0
|
-4,0
|
-0,5
|
-8,0
|
10,5
|
-1,5
|
21,4
|
-4,0
|
| 15
|
1,0
|
-3,0
|
0,0
|
-4,5
|
9,8
|
-2,0
|
28,0
|
-3,5
|
| 16
|
0,0
|
-6,5
|
0,2
|
-6,8
|
10,0
|
-1,6
|
30,0
|
0,6
|
| 17
|
0,0
|
-9,0
|
2,0
|
-6,2
|
1,2
|
-1,5
|
33,5
|
2,0
|
| 18
|
1,7
|
-6,4
|
0,5
|
-9,5
|
10,8
|
-1,5
|
28,5
|
-11,0
|
| 19
|
2,0
|
-9,0
|
1,5
|
-3,0
|
23,0
|
2,0
|
29,5
|
3,0
|
| 20
|
-3,0
|
-12,0
|
-2,0
|
-22,0
|
7,2
|
-1,5
|
32,6
|
1,5
|
| 21
|
0,5
|
-6,5
|
-3,5
|
-14,2
|
15,0
|
4,0
|
34,5
|
2,0
|
| 22
|
-3,0
|
-11,5
|
-1,0
|
-23,0
|
14,7
|
2,0
|
35,7
|
4,0
|
| 23
|
-4,0
|
-8,6
|
0,5
|
-22,0
|
20,5
|
-2,5
|
33,0
|
6,5
|
| 24
|
0,0
|
-5,0
|
1,0
|
-12,5
|
22,5
|
0,5
|
33,0
|
7,5
|
| 25
|
1,5
|
-2,0
|
1,1
|
-2,5
|
20,2
|
3,4
|
31,0
|
3,5
|
| 26
|
0,9
|
-3,2
|
2,0
|
-1,0
|
16,2
|
1,0
|
14,5
|
6,5
|
| 27
|
-3,0
|
-6,0
|
1,0
|
-1,0
|
7,5
|
2,0
|
16,7
|
4,5
|
| 28
|
0,0
|
-5,2
|
2,5
|
-0,5
|
9,2
|
1,5
|
16,5
|
-0,2
|
| 29
|
0,9
|
-3,0
|
2,1
|
-3,0
|
12,0
|
2,8
|
28,0
|
5,0
|
| 30
|
2,0
|
-0,2
|
14,5
|
1,0
|
30,8
|
1,0
|
| 31
|
1,0
|
-8,5
|
15,5
|
-3,0
|
| Сумма за месяц
|
-26,1
|
-190,8
|
5,4
|
-236,4
|
286,7
|
-75,2
|
687,6
|
2,7
|
| Средняя за месяц
|
-0,8
|
-6,2
|
0,2
|
-8,2
|
9,2
|
-2,4
|
22,9
|
0,1
|
Таблица 5.17 (продолжение)
| Число
|
Май
|
Июнь
|
Июль
|
Август
|
| Макс
|
мин
|
макс
|
мин
|
макс
|
мин
|
макс
|
мин
|
| 1
|
18,0
|
5,0
|
20,0
|
11,5
|
21,2
|
11,5
|
47,6
|
17,0
|
| 2
|
29,5
|
3,0
|
37,6
|
7,3
|
35,4
|
12,0
|
49,5
|
17,0
|
| 3
|
15,6
|
8,0
|
38,5
|
6,4
|
23,5
|
13,5
|
47,2
|
17,0
|
| 4
|
30,5
|
7,1
|
40,4
|
6,8
|
33,0
|
14,5
|
22,5
|
16,5
|
| 5
|
32,6
|
7,5
|
33,5
|
7,2
|
35,6
|
11,0
|
37,5
|
12,5
|
| 6
|
33,0
|
7,2
|
42,3
|
9,0
|
31,0
|
11,0
|
48,0
|
12,5
|
| 7
|
24,5
|
11,5
|
45,0
|
10,1
|
36,0
|
14,0
|
24,5
|
13,5
|
| 8
|
25,5
|
12,5
|
49,5
|
10,0
|
41,6
|
10,0
|
35,0
|
13,5
|
| 9
|
31,5
|
9,3
|
40,8
|
11,5
|
42,9
|
10,5
|
31,5
|
16,0
|
| 10
|
33,0
|
10,0
|
15,2
|
8,5
|
48,0
|
11,0
|
36,0
|
14,1
|
| 11
|
34,5
|
9,6
|
35,7
|
6,4
|
37,5
|
12,0
|
37,0
|
16,5
|
| 12
|
18,5
|
11,5
|
44,5
|
8,5
|
31,7
|
16,0
|
38,0
|
10,5
|
| 13
|
14,0
|
7,0
|
42,0
|
12,0
|
36,0
|
16,6
|
43,0
|
10,5
|
| 14
|
32,0
|
4,5
|
42,0
|
11,5
|
32,5
|
15,0
|
31,0
|
12,5
|
| 15
|
35,0
|
1,5
|
35,0
|
12,0
|
30,6
|
16,0
|
36,0
|
12,0
|
| 16
|
34,5
|
2,0
|
33,8
|
12,5
|
27,5
|
13,3
|
41,0
|
7,5
|
| 17
|
36,7
|
3,5
|
31,2
|
10,5
|
36,5
|
12,0
|
47,5
|
9,5
|
| 18
|
27,5
|
8,5
|
35,5
|
6,5
|
37,5
|
12,0
|
42,0
|
10,5
|
| 19
|
34,5
|
3,5
|
42,5
|
12,2
|
39,6
|
11,0
|
44,0
|
11,6
|
| 20
|
22,5
|
4,0
|
42,7
|
13,0
|
42,0
|
11,5
|
45,5
|
11,0
|
| 21
|
35,3
|
3,5
|
49,2
|
12,5
|
44,1
|
13,0
|
47,0
|
16,0
|
| 22
|
30,0
|
5,0
|
43,5
|
13,0
|
41,0
|
14,8
|
39,5
|
18,0
|
| 23
|
23,0
|
11,5
|
29,4
|
15,6
|
38,5
|
13,8
|
40,2
|
15,7
|
| 24
|
24,5
|
5,6
|
32,7
|
17,5
|
46,5
|
14,3
|
24,1
|
15,5
|
| 25
|
15,5
|
10,6
|
34,5
|
15,3
|
45,0
|
15,3
|
33,5
|
10,0
|
| 26
|
24,5
|
7,5
|
43,5
|
15,0
|
51,5
|
15,8
|
32,5
|
9,0
|
| 27
|
36,0
|
4,0
|
25,5
|
15,6
|
49,5
|
12,5
|
15,5
|
10,0
|
| 28
|
28,5
|
6,2
|
35,8
|
10,5
|
51,5
|
18,0
|
27,0
|
13,0
|
| 29
|
36,0
|
8,5
|
31,5
|
9,2
|
49,6
|
17,5
|
27,5
|
13,8
|
| 30
|
28,5
|
9,0
|
28,5
|
13,5
|
41,5
|
15,5
|
28,5
|
14,5
|
| 31
|
29,6
|
14,0
|
48,5
|
16,5
|
28,2
|
14,0
|
| Сумма
за месяц
|
874,8
|
222,1
|
1101,8
|
331,1
|
1205,8
|
421,4
|
1127,8
|
411,2
|
| Средняя за месяц
|
28,2
|
7,2
|
36,7
|
11,0
|
38,9
|
13,6
|
36,4
|
13,3
|
Таблица 5.17 (окончание)
| Число
|
Сентябрь
|
Октябрь
|
Ноябрь
|
Декабрь
|
| Макс
|
Мин
|
макс
|
Мин
|
макс
|
мин
|
макс
|
Мин
|
| 1
|
36,3
|
11,0
|
17,5
|
12,0
|
14,0
|
8,0
|
-5,2
|
-15,5
|
| 2
|
32,0
|
15,0
|
17,5
|
8,2
|
13,0
|
4,5
|
0,5
|
11,5
|
| 3
|
34,5
|
8,4
|
12,5
|
4,0
|
16,1
|
-1,0
|
2,0
|
0,0
|
| 4
|
33,5
|
10,0
|
10,5
|
3,5
|
17,0
|
-1,0
|
3,0
|
-0,5
|
| 5
|
19,0
|
10,0
|
13,8
|
3,0
|
16,0
|
0,0
|
1,6
|
-1,0
|
| 6
|
32,5
|
10,0
|
22,1
|
-0,7
|
16,0
|
0,0
|
3,5
|
-1,0
|
| 7
|
34,6
|
4,5
|
22,0
|
0,5
|
16,5
|
2,0
|
0,6
|
-0,5
|
| 8
|
31,5
|
8,0
|
25,0
|
-0,5
|
4,5
|
0,0
|
2,1
|
0,0
|
| 9
|
29,1
|
4,5
|
25,0
|
1,5
|
9,7
|
4,0
|
4,0
|
0,8
|
| 10
|
24,6
|
6,9
|
20,0
|
4,5
|
10,5
|
5,0
|
3,0
|
-1,5
|
| 11
|
28,0
|
1,0
|
13,5
|
1,0
|
14,8
|
3,5
|
-0,5
|
-5,0
|
| 12
|
20,0
|
5,0
|
12,5
|
-3,0
|
7,0
|
3,3
|
0,0
|
-2,0
|
| 13
|
31,5
|
5,1
|
18,5
|
-2,5
|
9,0
|
4,0
|
0,0
|
-0,5
|
| 14
|
33,5
|
7,5
|
12,0
|
-2,5
|
9,5
|
6,5
|
-2,5
|
-6,5
|
| 15
|
37,0
|
8,5
|
13,4
|
0,0
|
8,0
|
2,8
|
-3,0
|
-8,5
|
| 16
|
36,0
|
7,5
|
19,5
|
0,0
|
5,5
|
0,0
|
-2,8
|
-9,0
|
| 17
|
22,6
|
8,0
|
7,5
|
1,5
|
8,0
|
-2,0
|
-1,5
|
-4,7
|
| 18
|
31,5
|
3,0
|
23,5
|
9,0
|
0,2
|
-5,5
|
1,5
|
-0,5
|
| 19
|
32,5
|
3,0
|
20,6
|
2,0
|
6,0
|
-4,5
|
2,3
|
-1,0
|
| 20
|
35,0
|
7,0
|
20,5
|
4,3
|
9,5
|
-7,0
|
1,5
|
-1,0
|
| 21
|
33,0
|
8,5
|
8,5
|
3,5
|
0,5
|
-6,0
|
-0,5
|
-2,0
|
| 22
|
25,0
|
12,5
|
14,1
|
2,5
|
5,4
|
-9,8
|
-3,0
|
-8,0
|
| 23
|
24,4
|
7,5
|
11,5
|
1,0
|
2,1
|
-10,0
|
-4,8
|
-17,5
|
| 24
|
13,3
|
6,5
|
13,0
|
5,0
|
1,2
|
-9,0
|
-3,5
|
-13,0
|
| 25
|
17,5
|
5,0
|
21,0
|
9,0
|
2,2
|
-13,0
|
0,0
|
-3,5
|
| 26
|
22,0
|
12,0
|
24,1
|
6,5
|
-2,5
|
-18,5
|
1,0
|
-10,5
|
| 27
|
19,1
|
14,5
|
12,6
|
8,5
|
0,0
|
-9,0
|
1,5
|
0,0
|
| 28
|
21,0
|
13,3
|
11,5
|
10,5
|
-2,9
|
-9,0
|
1,8
|
0,0
|
| 29
|
25,8
|
8,0
|
19,0
|
2,0
|
-1,4
|
-13,5
|
1,0
|
0,0
|
| 30
|
17,5
|
9,0
|
10,5
|
9,5
|
-3,5
|
-13,4
|
1,0
|
-1,5
|
| 31
|
11,5
|
9,5
|
0,5
|
-3,0
|
| Сумма за месяц
|
833,8
|
240,7
|
504,7
|
113,3
|
211,9
|
-88,6
|
5,1
|
-129,2
|
| Средняя за месяц
|
27,8
|
8,0
|
16,3
|
3,6
|
7,1
|
-2,9
|
0,2
|
-4,2
|
7. ФЛОРА И РАСТИТЕЛЬНОСТЬ
7.1. ФЛОРА
7.1.1. Аннотированный конспект флоры листостебельных мхов участка «Лес на
Ворскле»
Материалы и методы исследования
Сбор бриологического материала проводился на участке “Лес на Ворскле” заповедника “Белогорье” в полевой сезон 2003 года и в апреле 2004 года. Так же были учтены образцы, собранные нами в предыдущие годы, но долгое время остававшиеся неопределенными.
Сбор проводился маршрутным методом и методом пробных площадей. При обозначении пробных площадей проводился анализ карт лесотаксации, учитывался: состав древостоя, его возраст, рельеф территории. При разработке маршрута мы старались охватить биотопы, не вошедшие в точки для сбора - это нарушенный почвенный покров; антропогенные субстраты; пойма реки Ворсклы; старицы в лесу в пойме реки Локни и др.
Определение проводилось с использованием следующих определителей: Игнатов М.С. и др. Флора мхов средней части европейской России. Том 1. Sphagnaceae – Hedwigiaceae (2003), Абрамов И.И., и др. Определитель листостебельных мхов Карелии (1998), Абрамова А.Л. и др. Определитель листостебельных мхов Арктики СССР (1961), Бардунов Л.В. Определитель листостебельных мхов Центральной Сибири (1969), Мельничук В.М. Определитель лиственных мхов средней полосы и юга европейской части СССР (1970), Савич-Любицкая Л.И. Определитель листостебельных мхов СССР. Верхоплодные мхи (1970).
По образцам, определение которых вызывало затруднения, были проведены консультации в Главном ботаническом саду им. Н.В. Цицина РАН (г. Москва) и в Ботаническом институте им. В.Л. Комарова РАН (г. Санкт-Петербург).
Аннотированный конспект флоры листостебельных мхов участка “Лес на Ворскле” заповедника “Белогорье”
Данные по флоре мхов участка “Лес на Ворскле” заповедника “Белогорье” были получены на основе собственных сборов, а так же на основе анализа литературных источников. В результате исследования было просмотрено и определено более 900 гербарных экземпляров мохообразных.
Из наших сборов было определено 74 вида и 2 разновидности листостебельных мхов. Из литературных данных для “Леса на Ворскле” приводится еще 5 видов не найденных нами. Это Leptodictium
humile
, Tetraphis
pellucida
, Tortula
mucronifolia
, Plagiothecium
succulentum
(Попова, 2002, с.158), Thuidium
philihertii
(Максимов,1939, с.67). Таким образом, для участка “Лес на Ворскле” заповедника “Белогорье” на настоящий момент известно 79 видов и 2 разновидности листостебельных мхов.
Из найденных нами видов более 15 впервые указываются для “Леса на Ворскле”. Четыре вида нами впервые указываются для Белгородской области. Это виды: Cirriphyllum piliferum
, Orthotrichum affine
, Polytrichum formosum
, Pseudoscleropodium purum
.
Из наиболее интересных видов можно назвать Orthodicranum
strictum
, Tortula
subulata
, Eurhinchium
angustirete
, которые рекомендованы к занесению в Красную книгу Белгородской области, а так же Pseudoscleropodium
purum
, местонахождение которого в “Лесу на Ворскле” является первой достоверной находкой для территории Среднерусской возвышенности.
Из наиболее распространенных семейств первое место занимает Brachytheciaceae – 11 видов из 4 родов, затем следуют Hypnaceae – 8 видов из 6 родов, Amblystegiaceae – 6 видов и одна разновидность из 4 родов, Mniaceae – 7 видов из 3 родов, Pottiaceae – 6 видов из 3 родов. Семейства Polytrichaceae, Bryaceae и Dicranaceae – представлены 5 видами; Plagiotheciaceae - 4 видами; Orthotrichaceae, Anomodontaceae и Thuidiaceae – 3 видами; Leskeaceae, Fissidentaceae, Hylocomiaceae – 2 видами; Tetraphidaceae, Aulacomniaceae, Climaceaceae, Ditrichaceae, Funariaceae, Leucodontaceae, Neckeraceae, Grimmiaceae – представлены одним видом.
Систематический состав флоры мхов участка “Лес на Ворскле” заповедника “Белогорье” представлен в таблице 7.1.1.1.
Таблица 7.1.1.1
Систематический состав флоры листостебельных мхов “Леса на Ворскле”
| Семейство
|
Число
|
Род и число видов в нем
|
| видов
|
родов
|
| 1
|
2
|
3
|
4
|
| Tetraphidaceae
|
1
|
1
|
Tetraphis (1)
|
| Polytrichaceae
|
5
|
2
|
Atrichum (1)
Polytrichum (4)
|
| Окончание таблицы 7.1.1.1
|
| 1
|
2
|
3
|
4
|
| Funariaceae
|
1
|
1
|
Funaria (1)
|
| Pottiaceae
|
6
|
3
|
Barbula (1)
Phascum (1)
Tortula (4)
|
| Grimmiaceae
|
1
|
1
|
Grimmia (1)
|
| Fissidentaceae
|
2
|
1
|
Fissidens (2)
|
| Orthotrichaceae
|
3
|
1
|
Orthotrichum (3)
|
| Ditrichaceae
|
1
|
1
|
Ceratodon (1)
|
| Dicranaceae
|
5
|
3
|
Dicranum (2)
Dicranella (1)
Orthodicranum (2)
|
| Bryaceae
|
5
|
2
|
Bryum (4)
Pohlia (1)
|
| Mniaceae
|
7
|
3
|
Mnium (2)
Plagiomnium (4)
Rhizomnium (1)
|
| Aulacomniaceae
|
1
|
1
|
Aulacomnium (1)
|
| Climaceaceae
|
1
|
1
|
Climacium (1)
|
| Leucodontaceae
|
1
|
1
|
Leucodon (1)
|
| Anomodontaceae
|
3
|
1
|
Anomodon (3)
|
| Neckeraceae
|
1
|
1
|
Homalia (1)
|
| Leskeaceae
|
2
|
2
|
Leskea (1)
Leskeella (1)
|
| Thuidiaceae
|
3
|
2
|
Abietinella (1)
Thuidium (2)
|
| Amblystegiaceae
|
7
|
4
|
Amblystegium (3)
Calliergonella (1)
Leptodictyum (2)
Sanionia (1)
|
| Brachytheciaceae
|
11
|
4
|
Brachythecium (6)
Cirriphyllum (1)
Eurhynchium (3)
Pseudoscleropodium (1)
|
| Plagiotheciaceae
|
4
|
1
|
Plagiothecium (4)
|
Hypnaceae
|
8
|
6
|
Callicladium (1)
Hypnum (3)
Platydictya (1)
Ptilium (1)
Pylaisiella (1)
Platygyrium (1)
|
| Hylocomiaceae
|
2
|
2
|
Hylocomium (1)
Pleurozium
(1)
|
В приведенном ниже конспекте флоры мхов участка “Лес на Ворскле”
заповедника “Белогорье” объем, расположение и номенклатура таксонов даны в соответствии со списком мхов территории бывшего СССР (Игнатов и др.,1992).
В конспекте после названия вида указаны следующие данные о биологии и экологии данного вида, взятые из статьи Н.Н. Поповой “Бриофлора Среднерусской возвышенности” (2002):
Распределение полов:
1d – однодомный;
2d - двудомный;
1/2d – многодомный.
Репродуктивные органы:
V+ - регулярное образование специализированных выводковых органов;
V+/- - нерегулярное;
Наличие спорофитов указано в конце аннотации к виду.
LF - Жизненные формы:
T - высокие дерновинки;
t - низкие дерновинки;
Trh - высокие плотные дерновинки;
trh - низкие плотные дерновинки;
Tcu - высокие подушковидные дерновинки;
tcu - низкие подушковидные дерновинки;
D – древовидные;
W – сплетения;
Mr - грубые коврики;
Ms - плоские коврики;
Mt - нитевидные коврики.
С - Эколого-ценотические группы:
f – лесные;
st – степные;
f-st - лесо-степные;
f-m-b - лесо-лугово-болотные;
f-b - лесо-болотные;
m-b - лугово-болотные;
p – петрофиты;
p-f - петрофитно-лесные;
p-st - петрофитно-степные;
m-w-b - лугово-водно-болотные;
et – эвритопные.
G - Географические элементы.
ab - аркто-бореальный;
b – бореальный;
n – неморальный;
bn - бореально-неморальный;
x – аридный;
aa – арктоальпийский;
eh – эвриголарктический;
k – космополитный.
BRYOPSIDA
Bryidae
Tetraphidaceae
Tetraphis
pellucida
Hedw. – Тетрафис прозрачный.
2d; V+; LF: t; C: f; G: b.
Приводится Н.Н. Поповой. Произрастает почти исключительно на сильно разложившейся древесине во влажных и затененных условиях. Со спорофитами
(Попова, 2002, с.127).
Polytrichaceae
Atrichum
undulatum
(Hedw.) P. Beauv. –
Catharinea
undulata
Hedw. – Атрихум волнистый.
1/2d; LF: T; C: f; G: n.
Один из самых распространенных мхов, произрастающих на почвенных обнажениях – песчаных и глинистых субстратах, на вывалах, стенках рвов, канав и окопов, на склонах лесных оврагов, совместно с видами рода Brachythecium
,
Eurhynchium
hians
,
Plagiomnium
cuspidatum
,
Amblystegium
serpens
и другими видами. Часто со спорофитами. Найден в большом количестве в 1, 4, 6, 8, 9 кварталах.
Polytrichum
commune
Hedw. – Политрихум обыкновенный.
2d; LF: Trh; C: f-m-b; G: b.
Встречен на задернованной песчаной почве вместе с Dicranum
polysetum
на окраине сосняка, выходящего в пойму рек Ворсклы и Готни, в охранной зоне (далее ОЗ) заповедника на границе с 5 кварталом. Без спорофитов.
Polytrichum
juniperinum
Hedw. – Политрихум можжевельниковидный.
2d; LF: T; C: f-st; G: eh.
Встречен на песчаной почве на окраине сосняка, выходящего в пойму рек Ворсклы и Готни, в ОЗ на границе с 5 кварталом. Без спорофитов.
Polytrichum
formosum
Hedw. –
Polytrichastrum
formosum
(Hedw.) G.L. Smith – Политрихум красивый.
2d; LF: T; C: f; G: b.
Встречен единожды на обнаженной почве на вывале в 8 квартале в лесном овраге. Без спорофитов.
Polytrichum
piliferum
Hedw. – Политрихум волосоносный.
2d; LF: T; C: f-st; G: eh.
Довольно массово встречается на песчаной почве на границе 5 квартала с ОЗ, на опушке сосняка, выходящего в пойму реки Готни. Без спорофитов.
Funariaceae
Funaria
hygrometrica
Hedw. – Фунария гигрометрическая.
1d; LF: t; C: et; G: k.
Найдена на нарушенной глинистой почве вместе с Barbula
unguiculata
на обочине дороги в 9-м квартале. Со спорофитами.
Pottiaceae
Barbula
unguiculata
Hedw. – Барбула полудюймовая.
2d; LF: t; C: et; G: k.
На нарушенной глинистой почве, вместе с Funaria
hygrometrica
на обочине дороги в 9-м квартале. Со спорофитами.
Phascum cuspidatum
Hedw. – Tortula acaulon
(With.) Zander – Фаскум заостренный.
1d; LF: t; C: st; G: x.
На незадернованной почве - на кротовине в дендрарии заповедника (8квартал).Со спорофитами.
Tortula
mucronifolia
Schwaegr. – Тортула остроконечная.
1d; LF: T; C: p-f; G: aa.
Приводится Н.Н. Поповой. Наиболее типичные местообитания вида - склоны нагорных дубрав, где он произрастает вблизи корней деревьев на черноземовидной карбонатной почве. Со спорофитами (Попова,2002,с.134).
Tortula
muralis
Hedw. – Тортула стенная.
1d; LF: t; C: p; G: x.
На кирпичной кладке на усадьбе заповедника и на кордоне в лесу. Со спорофитами.
Tortula
ruralis
(Hedw.) Gaertn. et al. –
Syntrichia
ruralis
(Hedw.) Web. et Mohr – Тортула полевая.
2d; LF: T; C: st; G: eh.
Найдена в 8-м квартале в овраге на лежащей ветке дуба, на коре, растет отдельными дерновинками. Без спорофитов.
Tortula
subulata
Hedw. – Тортула шиловидная.
1d; LF: T; C: f; G: n.
Найдена на почве у корней дерева в неглубоком, но сыром лесном овраге в 21 выделе 1-го квартала. Редкий для Среднерусской возвышенности вид, планируемый к занесению в Красную книгу Белгородской области. Спорофиты обильно развиты.
Grimmiaceae
Grimmia
pulvinata
(Hedw.) Sm. – Гриммия подушковидная.
1d; LF: tcu; C: p; G: eh.
Найдена на кирпичной кладке в пойме реки Готни в охранной зоне заповедника (на границе с 1-м кварталом) на дороге к селу Красный куток. Со спорофитами.
Fissidentaceae
Fissidens
bryoides
Hedw. – Фиссиденс моховидный.
1d; LF: t; C: f; G: n.
Найден в дендрарии заповедника (8 квартал) на незадернованной почве под пихтой. Спорофиты присутствуют в весенних сборах.
Fissidens
taxifolius
Hedw. – Фиссиденс тиссолистный.
1d; LF: t; C: f; G: n.
Найден на почве на дне довольно глубокого лесного оврага - “Вервейкова яра” в 6-м квартале заповедника. Без спорофитов.
Orthotrichaceae
Orthotrichum
affine
Brid. – Ортотрихум близкий.
1d; LF: Cu; C: f; G: n.
Растет на коре деревьев (как живых, так и сухих), и на валежнике. Найден в 4, 8 и 9 кварталах. Со спорофитами.
Orthotrichum
pumilum
Sw. – Ортотрихум карликовый.
1d; LF: Cu; C: f; G: n.
Найден на коре как живых (яблоня, клен), так и сухих деревьев, и на валежнике. Кварталы 4, 8, 9. Распространен немного более широко, чем предыдущий вид. Со спорофитами.
Orthotrichum
speciosum
Nees. – Ортотрихум прекрасный.
1d; LF: Cu; C: f; G: b.
Довольно широко распространенный эпифит. Растет как на коре живых и сухих
деревьев, так и на валежинах. Найден в кварталах 2, 3, 4, 8, 9. Почти всегда со спорофитами.
Ditrichaceae
Ceratodon purpureus
(Hedw.) Brid. – Цератодон пурпурный.
2d; LF: t; C: et; G: k.
Найден на песчаной и глинистой почве, на валежине в кварталах 8, 10 и на границе 5 кв. с ОЗ. Иногда со спорофитами.
Dicranaceae
Dicranum
polysetum
Sw. – Дикранум многоножковый.
2d; LF: Trh; C: f-b; G: b.
Довольно обилен на почве в посадках сосны в ОЗ на границе с 5-м кварталом вместе с Polytrichum commune Hedw. Встречен так же на валежине в овраге в 1-м квартале. Без спорофитов.
Dicranum
scoparium
Hedw. – Дикранум метловидный.
2d; LF: Tcu; C: f; G: b.
Довольно распространен. Встречается отдельными дерновинками вместе с Hypnum
pallescens
,
Callicladium
haldanianum
,
Platygyrium
repens
,
видами рода Brachythecium
на почве, на коре у основания деревьев, валежнике (как на коре, так и на ксилеме), на сильно разложившейся древесине. Найден в 1, 4, 6, 8, 10 кварталах, в посадках сосны в ОЗ на границе с 5-м кварталом. Без спорофитов.
Dicranella heteromalla
(Hedw.) Schimp. – Дикранелла разнонаправленная.
2d; LF: t; C: f; G: bn.
Довольно часто встречается на почве на стенках рвов, вывалах, стенках лесных оврагов вместе с Atrichum
undulatum
и другими видами. 1, 6, 9 кварталы. Найдено несколько коробочек в сборах из 1-го квартала.
Orthodicranum montanum
(Hedw.) Loeske – Dicranum mintanum
Hedw. – Ортодикранум горный.
2d; LF: tcu; C: f; G: b.
Часто встречается на коре у основания берез, дуба, на валежнике, на сильно разложившейся древесине вместе с Hypnum
pallescens
,
Platygyrium
repens
,
Dicranum
scoparium
. Была найдена береза, основание которой покрывалось мхом до высоты 1,2 м. Найден в 3, 4, 6, 8, 10 кварталах. Без спорофитов.
Orthodicranum strictum
(Schleich. Ex Mohr) Gulm. – Dicranum tauricum
Sapehin. – Ортодикранум прямой.
2d; V+; LF: tcu; C: f; G: bn.
Единичная находка на коре у основания сосны в посадках сосны в ОЗ на границе
с 5-м кварталом. Вид планируется к занесению в Красную книгу Белгородской области. Без спорофитов.
Bryaceae
Bryum
argenteum
Hedw. –
Бриум серебристый.
2d; LF: t; C: et; G: k
Вид нарушенных местообитаний и антропогенных субстратов. Встречен на усадьбе заповедника и на кордонах на бетонных дорожках, кирпичных постройках, шифере. Без спорофитов.
Bryum
caespiticium
Hedw. – Бриум дернистый.
2d; LF: trh; C: et; G: k.
Найден в дендрарии заповедника (8 квартал) на незадернованной почве под пихтами. Без спорофитов.
Bryum
capillare
Hedw. – Бриум волосоносный.
2d; LF: t; C: f; G: eh.
Довольно распространенный вид. Растет на коре у основания деревьев, на нарушенной почве, на вывалах и валежинах вместе с Amblystegium
serpens
,
Atrichum
undulatum
и другими видами. Найден в 1, 4, 8 кварталах. Без спорофитов.
Bryum
capillare
var. flaccidum
(Brid.) B.S.G. –
Bryum
laevifilum
Syed. – Бриум гладконитьевый.
2d; V+; C: f; G: eh.
Более распространенный, чем предыдущий вид. Растет на коре у основания деревьев, на вывалах, валежнике, на сильно разложившейся древесине вместе с Amblystegium
serpens
,
Atrichum
undulatum
,
Platygyrium
repens
и другими видами. 1, 4, 6, 8, 10 кварталы, в посадках сосны в ОЗ заповедника на границе с 5-м кварталом. Без спорофитов.
Pohlia
nutans
(Hedw.) Lindb. – Полия поникшая.
1d; LF: trh; C: f; G: eh.
Часто можно встретить на почве между корнями и у основания деревьев, на стенках рвов, вывалах, реже – на валежинах и сильно разложившейся древесине, найдена так же на горизонтальном стволе липы на высоте около 1 м над землей. Встречается вместе с видами рода Brachythecium
,
Plagiomnium
cuspidatum
,
Eurhynchium
hians
,
Atrichum
undulatum
и другими видами. 1, 3, 6, 8, 9 кварталы, в посадках сосны в ОЗ на границе с 5-м кварталом. Часто со спорофитами.Mniaceae
Mnium marginatum
(Dicks.) P. Beauv. – Мниум окаймленный.
1d; LF: T; C: p-f; G: b
Найден на почве на склонах в глубоком и сыром лесном овраге – “Вервейковом яру” в 6-м квартале. Без спорофитов.
Mnium
stellare
Hedw. – Мниум звездчатый.
2d; LF: T; C: p-f; G: n.
Найден там же где и предыдущий вид, так же на почве на склонах оврага. Без спорофитов.
Plagiomnium
cuspidatum
(Hedw.) T. Kop. – Плагиомниум остроконечный.
1d; LF: T; C: f; G: bn.
Один из самых распространенных видов. Встречается на почве, вывалах, валежинах, сильно разложившейся древесине, на коре у основания и между корнями деревьев, на стенках рвов, окопов, на склонах лесных оврагов. Иногда покрывает сплошным ковром небольшие участки почвы во влажных местах. Растет совместно с видами рода Brachythecium
,
Eurhynchium
hians
,
Atrichum
undulatum
,
Amblystegium
serpens
и другими видами. Часто с обильным спороношением.1, 2, 3, 4, 6, 8, 9, 10 кварталы.
Plagiomnium medium
(B.S.G.) T. Kop. – Плагиомниум средний.
1d; LF: T; C: f; G: n.
Встречен на почве на тропинках в 1 и 8 кварталах, в дендрарии под кустами. Со спорофитами.
Plagiomnium rostratum
(Schrad.) T. Kop. – Плагиомниум клювовидный.
1d; LF: T; C: f; G: n.
Найден на почве на склонах в глубоком и сыром лесном овраге – “Вервейковом яру” в 6-м квартале. Найдены спорофиты в весенних сборах.
Plagiomnium
undulatum
(Hedw.) T. Kop. – Плагиомниум волнистый.
2d; LF: T; C: f; G: n.
Найден в 1-м квартале в неглубоком, но сыром лесном овраге. Там довольно обилен. Без спорофитов.
Rhizomnium
punctatum
(Hedw.) T. Kop. – Ризомниум точечный.
2d; LF: Trh; C: f; G: b.
Найден на разлагающейся древесине в “Вервейковом яру” в 6-м квартале, в лесном овраге в 8-м квартале и на почве на тропинке по просеке между 1 и 5 кварталами. В весенних сборах найдены спорофиты.
Aulacomniaceae
Aulacomnium palustre
(Hedw.) Schwaegr. –
Аулакомниум болотный.
2d; V+; LF: Trh; C: f-m-b; G: ab.
Найден на почве в посадках сосны в охранной зоне на границе с 5-м кварталом. Без спорофитов.
Climaceaceae
Climacium
dendroides
(Hedw.) Web. et Mohr – Климациум древовидный.
2d; LF: D; C: f-m-b; G: b.
Встречен на почве в дендрарии заповедника (8 квартал) и в посадках сосны на границе 5 квартала с ОЗ. Встречается как на открытых местах так и под деревьями и кустами. Без спорофитов.
Leucodontaceae
Leucodon sciuroides
(Hedw.) Schwaegr. – Левкодон беличий.
2d; LF: Mr; C: f; G: n.
Распространен в старовозрастных участках леса – в вековой дубраве в 10 квартале и вековом ясеннике в 4-м квартале. Встречен на коре клена, ясеня до высоты 1,5 м и выше, произрастает совместно с Anomodon attenuatus, A. longifolius. Без спорофитов.
Anomodontaceae
Anomodon attenuatus
(Hedw.) Hueb. – Аномодон утонченный.
2d; LF: Mr; C: p-f; G: n.
Довольно широко распространен на коре у основания дуба, ясеня, клена, так же на коре липы, встречается на валежинах и вывалах. 1, 2, 3, 6, 8, 9 кварталы. Без спорофитов.
Anomodon longifolius
(Brid.) Hartm.
– Аномодон длиннолистный.
2d; LF: Mr; C: p-f; G: n.
Встречается на коре у основания ясеня, клена, выше на стволе этих деревьев (на клене до 1,5 м) и липы (80 см над землей), на валежине. В основном в старовозрастных участках леса. 4, 6, 10 кварталы. Без спорофитов.
Anomodon viticulosus
(Hedw.) Hook. et Tayl. – Аномодон плетевидный.
2d; LF: Mr; C: p-f; G: n.
На коре живых старовозрастных деревьев до высоты 1,5 м и выше, так же найден на недавно упавшей валежине. В старовозрастных участках леса. 4, 8, 10 кварталы. Часто встречается совместно с другими видами рода Anomodon. Без спорофитов.
Neckeraceae
Homalia
trichomanoides
(Hedw.) B.S.G. – Гомалия трихомановидная.
1d; LF: Ms; C: f; G: n.
Встречается на коре у основания дуба, ясеня, клена, на почве на склонах лесных оврагов. 1, 3, 4, 6 кварталы. Иногда со спорофитами.Leskeaceae
Leskea
polycarpa
Hedw. – Лескея многоплодная.
1d; LF: Ms; C: f; G: n.
Распространенный вид. Встречается на стволах (иногда до высоты 1,7 м и выше) и у основания деревьев совместно с Leskeella
nervosa
,
Brachythecium
velutinum
,
B
.
velutinum
,
Amblystegium
serpens
и другими видами, на сильно разложившейся древесине, на стенках рвов. В 1, 2, 3, 6, 8, 9 кварталах. Часто со спорофитами.
Leskeella nervosa
(Brid.) Loeske. – Лескеелла жилковатая.
2d; V+; LF: Ms; C: f; G: bn.
Распространенный вид. На стволах (до 1,7 м и выше) и у основания клена, ясеня, липы, дуба, на валежинах и сильно разложившейся древесине. 1, 2, 4, 6, 9, 10 кварталы. Часто со спорофитами.
Thuidiaceae
Abietinella abietina
(Hedw.) Fleish. – Thuidium abietinum
(Hedw.) B.S.G. – Абиетинелла абиетина.
2d; LF: W; C: st; G: eh.
Найден на почве в дендрарии заповедника (8 квартал) на открытых местах, под кустами и пихтами совместно с Thuidium
recognitum
.
Без спорофитов.
Thuidium
philibertii
Limpr. – Туидиум Филибера.
2d; LF: W; C: f; G: bn.
Приводится в статье Л.Д. Максимова “Типы леса заповедника “Лес на Ворскле” (Максимов,1939,с. 67).
Thuidium recognitum
(Hedw.) Lindb. – Туидиум признанный.
2d; LF: W; C: f; G: bn.
Так же как и Abietinella
abietina
найден в дендрарии заповедника (8 квартал) на открытых местах и под кустами. Без спорофитов.
Amblystegiaceae
Amblystegium
serpens
(Hedw.) B.S.G. – Амблистегиум ползучий.
1d; LF: Mt; C: et; G: bn.
Один из самых массовых видов. Встречается на валежинах, на сильно разложившейся древесине, на коре у основания и на стволах деревьев, на почве, на вывалах, на стенках рвов и канав совместно с видами рода Brachythecium
,
Pylaisiella
polyantha
,
Hypnum
pallescens
,
Atrichum
undulatum
,
Platygyrium
repens
и другими видами. 1, 2, 3, 4, 6, 8, 9, 10 кварталы, в посадках сосны на границе 5 кв. с ОЗ. Часто со спорофитами.
Amblystegium serpens
var. juratzkanum
(Schimp.) Rau et Herv. – Amblystegium juratzkanum
Schimp.– Амблистегиум ползучий var. Юрацки.
1d; LF: Mt; C: et; G: n.
Найден на сильно разложившейся древесине в 1 и 10 кварталах. Со спорофитами.
Amblystegium
varium
(Hedw.) Lindb.- Амблистегиум разнообразный.
1d; LF: Ms; C: f-m-b; G: n.
Менее распространен, чем Amblystegium serpens, встречается на разложившейся древесине, на коре у основания деревьев, на почве, на вывалах. 1, 2, 3, 4, 8, 9 кварталы, в посадках сосны в ОЗ на границе с 5 кварталом. Иногда со спорофитами.
Calliergonella cuspidata
(Hedw.) Loeske – Каллиергонелла заостренная.
2d; LF: Mr; C: m-b; G: b.
Довольно обильна в дендрарии заповедника (8 квартал) на почве. Без спорофитов.
Leptodictyum humile
(P. Beauv.) Ochyra - Amblystegium curvipes
Guemb. – Лептодикциум приземистый.
1d; LF: Ms; C: f-m-b; G: n.
Приводится Н.Н. Поповой для переувлажненных местообитаний. Со спорофитами (9, 154).
Leptodictyum
riparium
(Hedw.) Warnst. – Лептодикциум береговой.
1d; LF: Ms; C: m-w-b; G: b.
Найден в болотце в 5-м квартале на корнях деревьев и валежнике, погруженных в воду. Со спорофитами.
Sanionia uncinata
(Hedw.) Loeske – Саниония крючковатая.
2d; LF: Ms; C: f; G: b.
Определена в сборах из 1-го квартала. Найдена на валежине. Без спорофитов.
Brachytheciaceae
Brachythecium
albicans
(Hedw.) B.S.G. – Брахитециум беловатый.
2d; LF: Mr; C: f-st; G: b.
На песчаной почве в траве в пойме р. Ворсклы и р. Готни совместно с Ceratodon
purpureus
в ОЗ на границе с 5 кварталом, и в дендрарии заповедника (8 квартал). Без спорофитов.
Brachythecium
oedipodium
(Mitt.) Jaeg. –
Brachythecium
curtum
(Lindb.) – Брахитециум вздутоножковый.
1d; LF: Ms; C: f; G: b.
Один из самых массовых видов. Встречается на валежинах, на сильно разложившейся древесине, на коре у основания деревьев, на почве, на вывалах, на стенках рвов и канав, на склонах оврагов совместно с другими видами Brachythecium
,
Amblystegium
serpens
,
Hypnum
pallescens
,
Platygyrium
repens
и другими видами. 1, 2, 4, 6, кварталы, в посадках сосны, на границе 5 квартала с ОЗ, в дендрарии (8 квартал). Часто со спорофитами.
Brachythecium
reflexum
(Starke) B.S.G. – Брахитециум отогнутый.
1d; LF: Ms; C: p-f; G: b.
Встречается на валежинах, на сильно разложившейся древесине, на коре у основания дубов и кленов совместно с другими видами Brachythecium
,
Amblystegium
serpens
,
Hypnum
pallescens
,
Platygyrium
repens
и другими видами.. 1, 2, 3, 4, 6 кварталы. Часто со спорофитами.
Brachythecium
rutabulum
(Hedw.) B.S.G. – Брахитециум кочерга.
1d; LF: Mr; C: f-m-b; G: b.
Один из самых массовых видов. Встречается на валежинах, на сильно разложившейся древесине, на коре у основания деревьев, на почве, вывалах, на стенках рвов и окопов совместно с другими видами Brachythecium
,
Amblystegium
serpens
,
Hypnum
pallescens
,
Platygyrium
repens
, на почве в посадках хвойных встречается совместно с Pleurozium
schreberi
. 1, 2, 3, 4, 6, 8, 9, 10 кварталы, в посадках сосны в ОЗ на границе с 5-м кварталом. Часто со спорофитами.
Brachythecium
salebrosum
(Web. et Mohr) B.S.G. – Брахитециум кочковатый.
1d; LF: Mr; C: et; G: bn.
Один из самых массовых видов. Встречается на валежинах, на сильно разложившейся древесине, на коре у основания деревьев, на почве, вывалах, на стенках рвов и окопов совместно с другими видами Brachythecium
,
Amblystegium
serpens
,
Hypnum
pallescens
,
Platygyrium
repens
и другими видами. 1, 2, 3, 4, 6, 8, 9, 10 кварталы, в посадках сосны в ОЗ на границе с 5-м кварталом. Часто со спорофитами.
Brachythecium
velutinum
(Hedw.) B.S.G. – Брахитециум бархатный.
1d; LF: Ms; C: f; G: b.
Один из самых массовых видов. Встречается на валежинах, на сильно разложившейся древесине, на коре у основания деревьев, на стволах деревьев (до высоты 1,7 м), на почве, вывалах, на стенках рвов и окопов совместно с другими видами Brachythecium
,
Amblystegium
serpens
,
Hypnum
pallescens
,
Platygyrium
repens
и другими видами. 1, 2, 3, 4, 6, 8, 9, 10 кварталы, в посадках сосны в ОЗ на границе с 5-м кварталом. Часто со спорофитами.
Cirriphyllum piliferum
(Hedw.) Grout. – Циррифиллум волосоносный.
2d; LF: W; C: f; G: b.
Встречен на разлагающейся древесине в посадках сосны в ОЗ на границе с 5 кварталом, и на почве под хвойными культурами в дендрарии заповедника (8 квартал) на почве. Без спорофитов.
Eurhynchium angustirete
(Broth.) T. Kop. – Эвринхиум узкоклеточный.
2d; LF: Mr; C: f; G: n.
Найден на валежине в 1-м квартале и на почве на склоне лесного оврага в 6-м квартале. Вид рекомендован к занесению в Красную книгу Белгородской области. Без спорофитов.
Eurhynchium
hians
(Hedw.) Sande Lac. – Эвринхиум зияющий.
2d; LF: Ms; C: et; G: n.
Один из самых массовых напочвенных видов. В основном встречается на почве, на вывалах, на склонах лесных оврагов, на стенках рвов и окопов, найден так же на коре у основания березы, на валежине и на сильно разложившейся древесине. Растет совместно с совместно видами рода Brachythecium
,
Amblystegium
serpens
,
Atrichum
undulatum
,
Plagiomnium
cuspidatum
и другими видами. 1, 4, 6, 8, 9, 10 кварталы. При весеннем сборе были найдены спорофиты.
Eurhynchium pul
с
hellum
(Hedw.) Jenn. – Эвринхиум красивенький.
2d; LF: Mr; C: f; G: b.
Найден на почве у водотока в лесном овраге, и на сильно разложившейся
древесине в “Вервейковом яру”, все в 6-м квартале. Без спорофитов.
Pseudoscleropodium
purum (Hedw.) Fleisch. ex Broth. – Псевдосклероподиум чистый.
Найден на почве в дендрарии (8 квартал) под кипарисами. Без спорофитов. Первая достоверная находка на территории Среднерусской возвышенности. Ранее отмечался Алексенко (1897г.) для двух районов Сумской области (Лебединский р-н: с. Кудановка – на опушке леса, Сумской р-н: с. Чернетчина)
(Алексеенко,1897,с.19). Но Н.Н. Попова считает это указание сомнительным, так как вид приводится для хвойного леса, что для него не характерно (Попова,2002, с.158).
Plagiotheciaceae
Plagiothecium cavifolium
(Brid.) Iwats. – Плагиотециум вогнутолистный.
2d; V+/-; LF: Ms; C: f; G: b.
На почве на склонах лесных оврагов, на валежине в овраге. Приурочен в основном к сырым местам. 1, 6, 8 кварталы. Иногда со спорофитами.
Plagiothecium
denticulatum
(Hedw.) B.S.G. – Плагиотециум мелкозубчатый.
1d; LF: Ms; C: f; G: b.
Встречается на валежинах, на сильно разложившейся древесине, на коре у основания дуба, сосны, на почве на склонах оврагов, стенках окопов, вывалах. 1, 4, 6, 9 кварталы, в посадках сосны в ОЗ на границе с 5-м кварталом. Спорофиты редко.
Plagiothecium
laetum
B.S.G. – Плагиотециум светло-зеленый.
1d; LF: Ms; C: f; G: n.
На валежинах, на коре у основания дубов, сосен. 1, 6, 10 кварталы. Спорофиты редко.
Plagiothecium succulentum
(Wils.) Lindb. – Плагиотециум суккулентный.
2d; LF: Ms; C: f; G: n.
Приводится Н.Н. Поповой для достаточно увлажненных и затененных местообитаний – на гнилой древесине, почвенных обнажениях и др. Без спорофитов (9, 159).
Hypnaceae
Callicladium haldanianum
(Grev.) Crum – Калликладиум Гальдони.
1d; LF: Ms; C: f; G: b.
Часто можно встретить на валежинах, на сильно разложившейся древесине, на коре у основания дуба. Растет совместно с видами рода Bachythecium
,
Leskea
polycarpa
,
Hypnum
pallescens
,
Platygyrium
repens
и другими видами. 1, 3, 6, 8, 9, 10 кварталы, в посадках сосны в ОЗ на границе с 5-м кварталом. Без спорофитов.
Hypnum
cupressiforme
Hedw. – Гипнум кипарисовидный.
2d; LF: Ms; C: p-f; G: eh.
Встречается в основном на валежинах, на сильно разложившейся древесине, иногда образует довольно мощные коврики, так же можно встретить на коре у основания деревьев, и на почве (в т.ч. и на вывалах) совместно с видами Brachythecium
,
Hypnum
pallescens
,
Platygyrium
repens
,
Callicladium
haldanianum
и другими видами. 1, 4, 6, 9, 10 квартал, в посадках сосны в охранной зоне заповедника на границе с 5-м кварталом. Иногда со спорофитами.
Hypnum
pallescens
(Hedw.) P. Beauv. – Гипнум бледноватый.
1d; LF: Ms; C: f; G: bn.
Один из самых массовых эпифитных и эпиксильных видов. Част и обилен на валежинах, на сильно разложившейся древесине, на коре деревьев, как у основания, так и до высоты 1,7 м. Растет совместно с видами Brachythecium
,
Platygyrium
repens
,
Callicladium
haldanianum
,
Amblystegium
serpens
и другими видами. Очень часто со спорофитами.
Hypnum
vaucheri
Lesq. – Гипнум Воше.
2d; LF: Ms; C: p-st; G: aa.
Найден на валежине в 10 квартале в вековой дубраве. Без спорофитов.
Platydictya
subtilis
(Hedw.) Crum –
Amblystegium
subtile
(Hedw.) B.S.G. – Платидиктия нежная.
1d; LF: Mt; C: p-f; G: n.
Встречается на сильно разложившейся древесине, на коре у основания ясеня, клена, на стволах дуба, клена. 1, 3, 6, 10 кварталы. Без спорофитов.
Ptilium crista-castrensis
(Hedw.) De Not. – Птилиум гребенчатый.
2d; LF: W; C: f; G: b.
Найдена одна веточка на валежине в овраге в 1-м квартале в вековой дубраве. Без спорофитов.
Pylaisiella
polyantha
(Hedw.) Grout – Pylaisia polyantha
(Hedw.) Schimp. – Пилезиелла многоцветковая.
1d; LF: Ms; C: f; G: n.
Часто можно встретить на валежинах, на сильно разложившейся древесине, у основания дуба, сосны, липы, на стволе яблони (в саду у усадьбы), липы, вяза. Растет совместно с видами Brachythecium
,
Hypnum
pallescens
,
Callicladium
haldanianum
,
Amblystegium
serpens
и другими видами. 1, 5, 8, 9, 10 кварталы, в посадках сосны в ОЗ на границе с 5-м кварталом. Часто со спорофитами.
Platygyrium
repens
(Brid.) B.S.G. – Платигириум ползучий.
1d; V+; LF: Ms; C: f; G: n.
Один из самых массовых видов. Очень часто можно встретить на валежинах, на сильно разложившейся древесине, у основания деревьев, на стволах до высоты 1,7 м и выше, на вывалах - совместно с видами Brachythecium
,
Hypnum
pallescens
,
Callicladium
haldanianum
,
Amblystegium
serpens
и другими видами, на песчаной почве (вместе с Polytrichum piliferum
) в пойме Ворсклы и Готни. 1, 2, 3, 4, 6, 8, 9 кварталы, в посадках сосны и в пойме Ворсклы и Готни в ОЗ на границе с 5-м кварталом. Часто со спорофитами.
Hylocomiaceae
Hylocomium
splendens
(Hedw.) B.S.G. – Гилокомиум блестящий.
2d; LF: W; C: f; G: b.
Встречен в посадках сосны в охранной зоне заповедника на границе с 5-м
кварталом. Без спорофитов.
Pleurozium
schreberi
(Brid.) Mitt. – Плевроциум Шребера.
2d; LF: W; C: f; G: b.
Довольно обилен на почве в посадках сосны в ОЗ на границе с 5-м кварталом и в дендрарии (8 квартал) заповедника. Встречается совместно с Brachythecium
rutabulum
.
Без спорофитов.
Анализ флоры мхов “Леса на Ворскле”
Анализ распределения мхов по субстратам
По типам субстрата мхи “Леса на Ворскле” можно разделить на мхи напочвенного покрова, эпифитные мхи, эпиксильные мхи, мхи каменистых антропогенных субстратов.
В дубраве с малонарушенным почвенным покровом напочвенные мхи практически отсутствуют в связи с большим количеством листового опада, и, вследствие чего, мертвопокровности этих участков. Здесь напочвенные мхи можно встретить в основном на нарушенной обнаженной почве на стенках оврагов, окопов, рвов, на кочках и тропинках. Среди наиболее распространенных видов напочвенного покрова, встречающихся практически повсеместно нужно назвать: Atrichum undulatum, Eurhynchium hians,
Brachythecium
salebrosum
,
B
.
velutinum
,
Plagiomnium
cuspidatum
. Немного реже можно встретить Pohlia
nutans
,
Brachythecium
rutabulum
,
B
.
oedipodium
,
Amblystegium
serpens
.
В лесных оврагах видовой состав видов мхов несколько обогащается в связи с повышенной, по сравнению с плакорными участками, относительной влажностью воздуха и часто наличию обнаженной почвы на дне и на отвесных стенках оврага, что создает большее разнообразие экотопических условий. Помимо вышеперечисленных видов здесь так же можно встретить Dicranella
heteromalla
,
Plagiothecium
cavifolium
,
Eurhynchium
pulchellum
,
Fissidens
taxifolius
,
Mnium
marginatum
,
Mnium
stellare
,
Plagiomnium
medium
,
Plagiomnium
rostratum
,
Rhizomnium
punctatum
,
Plagiothecium
denticulatum
,
Eurhynchium
angustirete
.
В посадках сосны в пойме рек Готни и Локни можно выделить моховой ярус, который составляют следующие виды бриофитов: наибольшую площадь моховых синузий имеют Brachythecium
rutabulum
и Pleurozium
schreberi
; так же в моховом покрове принимают участие Polytrichum
commune
,
Dicranum
polysetum
,
D
.
scoparium
,
Climacium
dendroides
,
Aulacomnium
palustre
,
Hylocomium
splendens
,
Pohlia
nutans
,
Brachythecium
oedipodium
.
Наличие в моховом покрове относительно молодых посадок сосны (35-40 летнего возраста) болотных видов мохообразных (таких как Aulacomnium
palustre
), а так же плауна обыкновенного, может быть связано с близким расположением сфагнового болота, находящегося в нескольких километрах от нашего сосняка.
В пойме реки Готни на песчаной почве на открытых местах можно встретить Ceratodon
purpureus
,
Polytrichum
piliferum
,
P
.
juniperinum
,
Tortula
ruralis
,
Brachythecium
albicans
.
Довольно богат и разнообразен видовой состав напочвенных мхов дендрария заповедника. На полянах довольно большую площадь моховых синузий имеют Plagiomnium
cuspidatum
,
P
.
medium
,
Abietinella
abietina
,
Thuidium
recognitum
,
Calliergonella
cuspidata
,
Pleurozium
schreberi
,
Atrichum
undulatum
,
Brachythecium
rutabulum
.
Так же на незадернованной почве здесь можно встретить Amblystegium
serpens
,
A
.
varium
,
Brachythecium
albicans
,
B
.
salebrosum
,
B
.
velutinum
,
Cirriphyllum
piliferum
,
Bryum
caespiticium
,
B
.
capillare
,
Ceratodon
purpureus
,
Climacium
dendroides
,
Fissidens
bryoides
,
Pohlia
nutans
,
Phascum
cuspidatum
,
Pseudoscleropodium
purum
.
Из мхов, участвующих в первичном заселении нарушенных почвенных субстратов можно назвать Funaria
hygrometrica
,
Barbula
unguiculata
,
Phascum
cuspidatum
,
Ceratodon
purpureus
,
Bryum
caespiticium
,
B
.
argenteum
Ряд видов мхов способен произрастать при достаточно развитом травяном покрове. Это такие виды, как Plagiomnium
cuspidatum
,
Brachythecium
albicans
,
Amblystegium
serpens
,
Eurhynchium
hians
и другие.
Среди видов каменистых антропогенных субстратов (бетонные и кирпичные постройки, бетонные дорожки) можно отметить: Ceratodon
purpureus
,
Bryum
caespiticium
,
B
.
argenteum
,
Tortula
ruralis
,
T
.
muralis
,
Grimmia
pulvinata
.
Характеризуя видовой состав эпифитных мхов необходимо отметить, что наибольшее количество видов обычно можно найти при основании стволов деревьев. Выше по стволу флора сильно обедняется. Характер обрастания стволов эпифитными мхами напоминает резко усеченный конус, так как мхи приурочены к наиболее влажной стороне дерева. Такая конфигурация определяется особенностями светового режима, преобладающим направлением ветров, распределением осадков. Эпифиты тяготеют к влажной стороне ствола (это верхняя сторона при даже незначительном наклоне ствола), даже если она обращена на юг. Это хорошо прослеживается в дубравах на склонах балок противоположных экспозиций. Благоприятные для поселения мхов условия создаются на коре, на наклоненных поверхностях стволов, где происходит аккумуляция влаги и гумуса. Именно на основаниях стволов и наклоненных поверхностях задерживается и большее, количество снега
(12, 124). Так же у основания стволов кора начинает немного подгнивать, поэтому основание по условиям больше похоже на валежник.
Из листостебельных мхов, приуроченных к основаниям деревьев необходимо отметить Amblystegium
serpens
,
Brachythecium
velutinum
,
B
.
oedipodium
,
B
.
rutabulum
,
B
.
reflexum
,
B
.
salebrosum
,
Hypnum
pallescens
,
Platygyrium
repens
,
Leskea
polycarpa
,
Leskeella
nervosa
,
Plagiothecium
denticulatum
,
P
.
laetum
,
Plagiomnium
cuspidatum
,
Amblystegium
varium
,
Anomodon
attenuatus
,
A
.
longifolius
,
Bryum
capillare
,
B
.
capillare
var. flaccidum
,
Callicladium
haldanianum
,
Dicranum
scoparium
,
Orthodicranum
montanum
,
Pylaisiella
polyantha
,
Homalia
trichomanoides
.
Такое большое разнообразие видов, растущих у основания, чаще наблюдается в порослевых дубравах, так как порослевые деревья часто расширены у основания, у их основания часто задерживается влага и может накапливаться мелкозем, что создает благоприятные условия для мхов.
Выше, до 1,5-2 м от земли по стволу поднимаются Amblystegium
serpens
,
Platydictya
subtilis
,
Anomodon
attenuatus
,
A
.
longifolius
,
A
.
viticulosus
,
Brachythecium
velutinum
,
Hypnum
pallescens
,
Leskea
polycarpa
,
Leskeella
nervosa
,
Leucodon
sciuroides
,
Pylaisiella
polyantha
,
Platygyrium
repens
,
Orthotrichum
speciosum
,
O
.
pumilum
, O
.
affine
.
Выше 2-х метров на стволах дерева можно встретить в основном Orthotrichum
speciosum
,
O
.
pumilum
, O
.
affine
,
а так же иногда по трещинам коры могут подниматься Pylaisiella
polyantha
,
Platygyrium
repens
.
Довольно интересен видовой состав листостебельных мхов, растущих на валежнике. Поваленные деревья являются начальным звеном при разложении древесины. Первое время валежник продолжает оставаться субстратом для эпифитных мхов, росших на коре еще живого дерева. Но по мере разложения древесины видовой состав мхов постепенно меняется. Здесь, помимо типичных эпифитных видов теперь можно встретить Hypnum
cupressiforme
,
Orthodicranum
montanum
,
Bryum
capillare
,
B
.
capillare
var. flaccidum
,
Sanionia
uncinata
,
Ptilium
crista
-
castrensis
,
Tortula
ruralis
.
Сильно разложившаяся древесина является конечным этапом при разложении древесных остатков. Во влажных и тенистых условиях разлагающаяся древесина активно заселяется мхами. Повышенная влажность и трофность гнилой древесины способствует массовому развитию экологически разнородных видов. Видовой состав мхов, растущих на сильно разложившейся древесине (эпиксилы) объединяет в себе как виды, которые часто встречаются у основания деревьев и на валежнике, так и виды почвенного покрова. Постепенно, по мере разложения древесины, эпифитные виды уступают место видам напочвенного покрова. Среди эпифитных видов, встречающихся на разложившейся древесине можно отметить Amblystegium
serpens
,
Amblystegium
varium
,
Brachythecium
velutinum
,
B
.
oedipodium
,
B
.
rutabulum
,
B
.
reflexum
,
B
.
salebrosum
,
Callicladium
haldanianum
,
Hypnum
pallescens
,
H
.
cupressiforme
,
Leskea
polycarpa
,
Leskeella
nervosa
,
Pylaisiella
polyantha
,
Platygyrium
repens
,
Dicranum
scoparium
.
Из видов напочвенного покрова здесь можно назвать Plagiomnium
cuspidatum
, Bryum
capillare
var. flaccidum
,
Cirriphyllum
piliferum
,
Orthodicranum
montanum
,
Eurhynchium
hians
,
Pohlia
nutans
,
Plagiomnium
rostratum
,
Rhizomnium
punctatum
,
Plagiothecium
denticulatum
.
Полученные нами данные вполне согласуются с данными К.Ф. Хмелева и Н.Н. Поповой по субстратной приуроченности мохообразных бассейна Среднего Дона (Хмелёв и др.,1988,с.123-135).
Фитоценотический анализ
Фитоценотический анализ флоры мхов участка “Лес на Ворскле” заповедника “Белогорье” показал, что больше всего в ней представлена лесная эколого-ценотическая группа. Лесная эколого-ценотическая группа листостебельных мохообразных “Леса на Ворскле” насчитывает 44 вида и представлена: Atrichum
undulatum
,
Brachythecium
oedipodium
,
Brachythecium
velutinum
,
Bryum
capillare
,
Bryum
capillare
var. flaccidum
,
Callicladium
haldanianum
,
Cirriphyllum
piliferum
,
Dicranella
heteromalla
,
Orthodicranum
montanum
,
Dicranum
scoparium
,
Orthodicranum
strictum
,
Eurhynchium
angustirete
,
Eurhynchium
pul
с
hellum
,
Fissidens
bryoides
,
Fissidens
taxifolius
,
Homalia
trichomanoides
,
Hylocomium
splendens
,
Hypnum
pallescens
,
Leskea
polycarpa
,
Leskeella
nervosa
,
Leucodon
sciuroides
,
Orthotrichum
speciosum
,
Orthotrichum
pumilum
,
Orthotrichum
affine
,
Plagiomnium
cuspidatum
,
Plagiomnium
medium
,
Plagiomnium
rostratum
,
Plagiomnium
undulatum
,
Plagiothecium
cavifolium
,
Plagiothecium
denticulatum
,
Plagiothecium
laetum
,
Plagiothecium
succulentum
,
Platygyrium
repens
,
Pleurozium
schreberi
,
Pohlia
nutans
,
Polytrichum
formosum
,
Ptilium
crista
-
castrensis
,
Pylaisiella
polyantha
,
Rhizomnium
punctatum
,
Sanionia
uncinata
,
Tetraphis
pellucida
,
Thuidium
recognitum
,
Thuidium
philibertii
,
Tortula
subulata
.
На втором месте находятся эвритопные виды, которые можно практически повсеместно на разных типах субстрата и в разных растительных группировках. Эта эколого-ценотическая группа представлена 9 видами: Bryum argenteum
, Amblystegium
serpens
var. Juratzkanum, Amblystegium serpens, Barbula unguiculata, Brachythecium salebrosum, Bryum caespiticium, Ceratodon purpureus, Eurhynchium hians, Funaria hygrometrica.
Петрофитно-лесная эколого-ценотическая группа так же представлена 9 видами: Platydictya subtilis, Anomodon attenuatus, Anomodon longifolius, Anomodon viticulosus, Brachythecium reflexum, Hypnum cupressiforme, Mnium marginatum, Mnium stellare, Tortula mucronifolia.
Шесть видов составляют лесо-лугово-болотную эколого-ценотическую группу: Aulacomnium palustre, Amblystegium varium, Brachythecium rutabulum, Climacium dendroides, Leptodictyum humile, Polytrichum commune.
Многие из этих видов были найдены в посадках сосны в охранной зоне на границе с 5-м кварталом.
Три вида входит в степную эколого-ценотическую группу: Tortula ruralis, Thuidium abietinum, Phascum cuspidatum
. Их можно встретить в пойме реки Готни на песчаной почве на открытых местах и в дендрарии заповедника на нарушенной незадернованной почве и на полянах.
Три вида входит в лесо-степную эколого-ценотическую группу. Это: Brachythecium albicans, Polytrichum juniperinum, Polytrichum piliferum
. Их можно встретить в посадках сосны в охранной зоне на границе с 5-м кварталом.
Из лесо-болотных видов был найден: Dicranum
polysetum
.
Из лугово-болотных Calliergonella
cuspidata
, массово произрастающая в дендрарии заповедника на открытых местах.
Leptodictyum riparium представляет лугово-водно-болотную эколого-ценотическую группу.
Петрофитная эколого-ценотическая группа: Grimmia
pulvinata
,
Tortula
muralis
, виды, которые встречаются на кирпичных постройках
Из петрофитно-степных Hypnum vaucheri,
Pseudoscleropodium
purum
.
В результате проведенного анализа выяснено, что самой многочисленной является лесная эколого-ценотическая группа. Участок “Лес на Ворскле” заповедника “Белогорье” представлен нагорной дубравой и, соответственно, видовой состав мхов так же представлен видами, приуроченными к лесным фитоценозам. Остальные эколого-ценотические группы в основном отражают разнообразие экотопов в пойме и в дендрарии заповедника.
Географический анализ флоры мхов
При географическом анализе флоры мхов участка “Лес на Ворскле” заповедника “Белогорье” выделено шесть географических элементов.
Бореальный географический элемент составляют 25 видов. Его образуют виды, распространение которых связано с зоной хвойных лесов Голарктики или имеющие в ней центры массовости. Это виды: Brachythecium
albicans
,
Brachythecium
oedipodium
,
Brachythecium
reflexum
,
Brachythecium
rutabulum
,
Brachythecium
velutinum
,
Callicladium
haldanianum
,
Calliergonella
cuspidata
,
Cirriphyllum
piliferum
,
Climacium
dendroides
,
Orthodicranum
montanum
,
Dicranum
polysetum
,
Dicranum
scoparium
,
Eurhynchium
pulchellum
,
Hylocomium
splendens
,
Leptodictyum
riparium
,
Mnium
marginatum
,
Plagiothecium
cavifolium
,
Plagiothecium
denticulatum
,
Pleurozium
schreberi
,
Polytrichum
formosum
,
Polytrichum
commune
,
Ptilium
crista
-
castrensis
,
Rhizomnium
punctatum
,
Sanionia
uncinata
,
Tetraphis
pellucida
.
Аркто-бореальный географический элемент составляет Aulacomnium
palustre
.
Неморальный географический элемент составляют 28 видов. Сюда относятся виды, распространенные в зоне широколиственных лесов Голарктики, на севере заходящие в зону хвойных лесов, а на юге – в лесостепь. Это: Leptodictium humile, Plagiothecium succulentum, Eurhynchium angustirete, Platydictya subtilis, Amblystegium serpens
var. juratzkanum, Amblystegium varium, Anomodon attenuatus, Anomodon longifolius, Anomodon viticulosus, Atrichum undulatum, Eurhynchium hians, Fissidens bryoides, Fissidens taxifolius, Homalia trichomanoides, Leskea polycarpa, Leucodon sciuroides, Mnium stellare, Orthotrichum affine, Orthotrichum pumilum, Orthotrichum speciosum, Plagiomnium medium, Plagiomnium rostratum, Plagiomnium undulatum, Plagiothecium laetum, Platygyrium repens, Pylaisiella polyantha, Pseudoscleropodium purum, Tortula subulata.
Бореально-неморальный географический элемент составляют 9 видов. Эти виды имеют центры массовости как в зоне хвойных, так и широколиственных лесов. Это: Thuidium philibertii, Amblystegium serpens, Brachythecium salebrosum, Dicranella heteromalla, Orthodicranum strictum, Hypnum pallescens, Leskeella nervosa, Plagiomnium cuspidatum, Thuidium recognitum.
Аридный географический элемент составляют виды, распространенные в основном в степной и полупустынной зонах или имеющие в ней центры массовости. Аридный географический элемент представлен двумя видами: Tortula muralis и Phascum cuspidatum.
Аркто-альпийский географический компонент представлен двумя видами: Tortula mucronifolia
и Hypnum vaucheri.
Эвриголарктический географический элемент составляют 9 видов. Виды мхов этого элемента распространены по всей или почти всей Голарктике, не отличаясь заметным тяготением к определенной растительно-климатической зоне. Это виды: Bryum capillare, Bryum capillare
var. flaccidum , Grimmia pulvinata, Hypnum cupressiforme, Pohlia nutans, Polytrichum juniperinum, Polytrichum piliferum, Tortula ruralis, Thuidium abietinum.
Пять видов составляют космополитный географический элемент. Эти виды произрастают на открытой почве и грунте, сопутствуют человеку, создающему своей деятельностью экотопы для их произрастания. Это: Bryum argenteum, Barbula unguiculata, Bryum caespiticium, Ceratodon purpureus, Funaria hygrometrica
(10, 21-26).
Из проведенного анализа видно, что во флоре мхов участка “Лес на Ворскле” заповедника “Белогорье” преобладают виды бореального и неморального, а так же бореально-неморального географических элементов, которые составляют виды, приуроченные в основном к лесным растительным группировкам. Остальные географические элементы по численности значительно уступают преобладающим элементам.
Заключение
В результате проведенных исследований, с учетом литературного материала, для участка “Лес на Ворскле” заповедника “Белогорье” выявлено 79 видов и 2 разновидности листостебельных мохообразных. Их них более 15 видов впервые указываются для “Леса на Ворскле”. Четыре вида впервые указываются для Белгородской области (
Cirriphyllum
piliferum
, Orthotrichum
affine
, Polytrichum
formosum
, Pseudoscleropodium
purum
)
, из них один вид (Pseudoscleropodium
purum
)
– первая достоверная находка на территории Среднерусской возвышенности. Составлен аннотированный конспект видов листостебельных мохообразных “Леса на Ворскле”. Был проведен таксономический анализ полученных данных, выявлены преобладающие семейства. Проведен анализ распределения мхов по типам субстрата, фитоценотический и географический анализы.
Три вида (
Orthodicranum
strictum
, Tortula
subulata
, Eurhinchium
angustirete
)
, найденных в “Лесу на Ворскле”, были рекомендованы к занесению в Красную книгу Белгородской области.
7.1.2. Лишайники особо охраняемых природных территорий Белгородской области
В итоге предпринятого критического обобщения результатов собственных полевых исследований, а также опубликованных и гербарных материалов, нами составлен список лишайников ООПТ Белгородской области, который в настоящее время включает 235 видов. Для ряда видов выявлены подвиды, разновидности и формы. Сбор и определение материала осуществлялись по стандартной методике (Окснер, 1974) с использованием ряда российских и зарубежных определителей (Окснер, 1956, 1993; Определитель, 1971-2004; Foucard, 2001; Servit, 1954; Wirth, 1995). Систематическое положение выявленных видов приводится в соответствии с работой О. Эрикссона и Д. Хоксворта (Eriksson, Hawksworth, 1998). Номенклатура таксонов указана по Р. Сантессону (Santesson, 2004), Виды в списке расположены в алфавитном порядке. После названия вида дается информация о его систематическом положении и синонимах, под которыми вид публиковался ранее для территории Белгородской области. Далее указаны административные районы в пределах области, где вид был отмечен (рис. 7.1.2.1). Даются также сведения о принадлежности видов к эколого-субстратной группе и встречаемости их на территории ООПТ. Встречаемость видов указана согласно следующей шкале: единично – вид встречен только на одной ООПТ, редко – на 2-х и 3-х ООПТ, часто – на 4-х и более ООПТ, но не является повсеместно распространенным; повсеместно – на всех обследованных ООПТ (доминирующий вид).
Условные обозначения
* - виды и внутривидовые таксоны, впервые найденные на территории ООПТ; часть этих сведений опубликована ранее (Конорева, 1999; Конорева, 2000; 2001; 2002; Конорева, Мучник, 2002а,б,в; Мучник, Конорева, 2003; Мучник, Урбанавичюс, 2001)
# - виды и внутривидовые таксоны, приведенные только по литературным данным (Бобырева, 1999а, б; Кашменский, 1906; Лебедева, 1997, неопубликованные гербарные данные; Макарова, 1971; Максимов, 1939; Мучник, 1995)
¢ - виды и внутривидовые таксоны, новые для Центрально-Черноземного региона
! – виды, включенные в Красную книгу Белгородской области
1. Аmandinea punctata
(Hoffm.) Coppins & Scheid. in Scheidegger (Lecanorales
, Physciaceae
) - syn. Buellia punctata
(Hoffm.) Massal.; 1, 3, 4, 5, 8, 14, 17, 19; эпифито-эпиксил, повсеместно.
2. Anaptychia ciliaris
(L.) Körb. (Lecanorales
, Physciaceae
) - 3, 5, 8, 10, 14, 18, 19; эпифит, часто.
*Anaptychia ciliaris
(L.) Körb. f. crinalis
Rabh. (Lecanorales
, Physciaceae
) - 8, 17; эпифит, редко.
3. *Arthonia dispersa
(Schrad.) Nyl. (Arthoniales
, Arthoniaceae
) – 8;
эпифит
,
единично
.
4. #Arthonia mediella
Nyl. (Arthoniales
, Arthoniaceae
) – 19;
эпифит
,
единично
.
5. *Arthonia punctiformis
Ach. (Arthoniales
, Arthoniaceae
) - 3, 5, 17, 19;
эпифит
,
часто
.
6. Arthonia radiata
(Pers.) Ach. (Arthoniales
, Arthoniaceae
) - 17, 19;
эпифит
,
редко
.
7. *Arthrosporum populorum
A.Massal. (Lecanorales
, Catillariaceae
) – 3; эпифит, единично.
8. *Aspicilia contorta
(Hoffm.) Kremp. (Lecanorales
, Hymeneliaceae
) – 11; эпилит, единично.
9. ¢*Aspicilia moenium
(Vain.) G.Thor & Timdal (Lecanorales
, Hymeneliaceae
) - 5, 17; эпилит, редко.
10. Bacidia bagliettoana
(A.Massal. & De Not.) Jatta (Lecanorales
, Bacidiaceae
) - syn. Bacidia
muscorum
(Sw.) Mudd; 4, 10, 14, 19; эпиксил, часто.
11. #Bacidia incompta
(Borrer ex Hook.) Anzi (Lecanorales
, Bacidiaceae
) – 3; эпифит, единично.
12. #Bacidia rubella
(Hoffm.) A.Massal. (Lecanorales
, Bacidiaceae
) - 3; эпифит, редко.
13. *Bacidia subincompta
(Nyl.) Arnold (Lecanorales
, Bacidiaceae
) - 4, 10; эпибриофит, редко.
14. #Bacidia trachona
(Ach.) Lettau (Lecanorales
, Bacidiaceae
) – 4; эпилит, единично.
15. #Bacidina arnoldiana
(Körb.) V.Wirth & Vĕzda (Lecanorales
, Bacidiaceae
) - syn. Bacidia arnoldiana
Körb.; 4;
эпилит
,
единично
.
16. ¢*Bacidina delicata
(Larbal. ex Leight.) Coppins (Lecanorales
, Bacidiaceae
) - 4, 10, 17; эпибриофит, редко. Новый вид для России.
17. !#Bryoria implexa
(Hoffm.) Brodo & D.Hawksw. (Lecanorales
, Parmeliaceae
) – syn. Alectoria implexa
(Hoffm.) Nyl. 3; эпифит, единично.
18. Buellia
disciformis
(Fr.) Mudd (Lecanorales
, Physciaceae
) - 8, 14, 15, 17, 18, 19; эпифито-эпиксил, часто.
19. Buellia schaereri
De Not. (Lecanorales
, Physciaceae
) - 3, 14, 19; эпифит, часто.
20. #Caloplaca borealis
(Vain.) Poelt (Teloschistales
, Teloschistaceae
) – 14; эпифит, единично.
21. Caloplaca cerina
(Ehrh. ex Hedw.) Th.Fr. (Teloschistales
, Teloschistaceae
) – 3, 4, 5, 8, 14, 15, 17, 18, 19; эпифито-эпиксил, часто.
22. *Caloplaca cerina
(Ehrh. ex Hedw.) Th.Fr. var. chloroleuca
(Sm.) Th.Fr. (Teloschistales
, Teloschistaceae
) – 10;
эпифито
-
эпиксил
,
единично
.
23. *Caloplaca cerinella
(Nyl.) Flagey (Teloschistales
, Teloschistaceae
) – 17;
эпифито
-
эпиксил
,
единично
.
24. Caloplaca decipiens
(Arnold) Blomb. & Forssell (Teloschistales
, Teloschistaceae
) – 3, 4, 5, 11, 17;
эпилит
,
часто
.
25. #Caloplaca ferruginea
(Huds.) Th.Fr. (Teloschistales
, Teloschistaceae
) – 1, 8, 14;
эврисубстратный
,
редко
.
26. Caloplaca holocarpa
(Hoffm. ex Ach.) A.E.Wade (Teloschistales
, Teloschistaceae
) - 1, 2, 3, 4, 8, 10, 11, 14, 17, 19; эврисубстратный, повсеместно.
*Caloplaca holocarpa
(Hoffm. ex Ach.) A.E.Wade var. chloroleuca
(Teloschistales
, Teloschistaceae
) – 4;
эпифито
-
эпиксил
,
единично
.
27. #Caloplaca lactea
(A.Massal.) Zahlbr. (Teloschistales
, Teloschistaceae
) – 1;
эпилит
,
единично
.
28. *Caloplaca obscurella
(J. Lahm ex Körb.) Th.Fr. (Teloschistales
, Teloschistaceae
) – 3;
эпифит
,
редко
.
29. ¢*Caloplaca raesaeneni
Bredk. (Teloschistales
, Teloschistaceae
) – 8;
эпигеидный
,
единично
.
30. Caloplaca stillicidiorum
(Vahl.) Lynge (Teloschistales
, Teloschistaceae
) – 10;
эпиксил
,
редко
.
31. Candelaria concolor
(Dicks) Stein (Lecanorales
, Candelariaceae
) - 3, 4, 5, 8, 14, 17, 19; эпифито-эпиксил, часто.
32. Candelariella aurella
(Hoffm.) Zahlbr. (Lecanorales
, Candelariaceae
) – 3, 5, 10, 11, 17; эврисубстратный, часто.
33. *Candelariella efflorescens
R.C.Harris & W.R.Buck (Lecanorales
, Candelariaceae
) - 3, 4, 5, 8, 14, 15, 17, 19; эпифит, часто.
34. Candelariella vitellina
(Hoffm.) Mǔll.Arg. (Lecanorales
, Candelariaceae
) - 2, 3, 4, 10, 14, 17; эврисубстратный, часто.
35. Candelariella xantostigma
(Ach.) Lettau (Lecanorales
, Candelariaceae
) - 3, 14, 17, 19; эпифит, часто.
36. *Catillaria nigroclavata
(Nyl.) Schuler (Lecanorales
, Catillariaceae
) - 3, 4, 8;
эпифит
,
часто
.
37. *Catinaria atropurpurea
(Schaer.) Vézda & Poelt (Lecanorales
, Bacidiaceae
) – syn. Catillaria atropurpurea
(Schaer.) Th
.
Fr
.; 14; эпифит, единично.
38. #Cetraria islandica
(L.) Ach. (Lecanorales
, Parmeliaceae
) - 19; эпигеидный, единично.
39. #Chaenotheca brunneola
(Ach.) Műll.Arg. (гр. Incertae sedis
, Coniocybaceae
) – 3; эпифит, единично.
40. *Chaenotheca chrysocephala
(Turner ex Ach.) Th.Fr. (гр. Incertae sedis
, Coniocybaceae
) – 3; эпифит, единично.
41. Chaenotheca ferruginea
(Turner ex Sm.) Mig. (гр. Incertae sedis
, Coniocybaceae
) – 3, 19; эпифит, редко.
42. Chaenotheca furfuracea
(L.) Tibell (гр. Incertae sedis
, Coniocybaceae
) – 18, 19; эпифито-эпиксил, редко.
43. Chaenotheca stemonea
(Ach.) Műll.Arg. (гр. Incertae sedis
, Coniocybaceae
) – 3, 18; эпифит, редко.
44. #Chaenotheca trichialis
(Ach.) Th.Fr. (гр. Incertae sedis
, Coniocybaceae
) – 3, 19; эпифито-эпиксил, редко.
45. Chaenothecopsis savonica
(Räsänen) Tibell (гр. Incertae sedis
, Mycocaliciaceae
) – 8, 19; эпиксил, редко.
46. Chrysothrix candelaris
(L.) J.R.Laundon (Arthoniales
, Chrysotrichaceae
) - syn. Lepraria
candelaria
(L.) Fr
.; 2, 3, 4, 8, 10, 17; эпифито-эпиксил, часто.
47. #Chrysothrix chlorina
(Ach.) J.R.Laundon (Arthoniales
, Chrysotrichaceae
) - syn. Lepraria chlorina
(Ach.) Ach.; 14, 18;
эпифито
-
эпиксил
,
редко
.
48. !*Cladonia arbuscula
(Wallr.) Flot. (Lecanorales
, Cladoniaceae
) - syn. Cladina
arbuscula
(Wallr.) Hale & W.L.Culb., Cladonia sylvatica
auct.; 3; эпигеидный, единично.
Cladonia arbuscula
(Wallr.) Flot. ssp. mitis
(Sandst.) Ruoss (Lecanorales
, Cladoniaceae
) - syn. Cladina mitis
(Sandst.) Mong., Cladonia mitis
Sandst.; 3, 14; эпигеидный, редко.
49. !#Cladonia botrytes
(K.G.Hagen) Willd. (Lecanorales
, Cladoniaceae
) – 3; эпиксил, единично.
50. !*Cladonia cenotea
(Ach.) Schaer. (Lecanorales
, Cladoniaceae
) – 3; эпигеидо-эпиксил, редко.
51. Cladonia chlorophaea
(Flőrke ex Sommerf.) Spreng. (Lecanorales
, Cladoniaceae
) - 1, 3, 8, 14, 17, 18, 19; эпигеидо-эпиксил, часто.
52. Cladonia coniocraea
(Flörke) Spreng. (Lecanorales
, Cladoniaceae
) - 1, 3, 4, 8, 14, 17, 18, 19; эпигеидо-эпиксил, повсеместно.
53. Cladonia cornuta
(L.) Hoffm. (Lecanorales
, Cladoniaceae
) - 3, 8, 17; эпигеидный, часто.
54. !*Cladonia crispata
(Ach.) Flot. (Lecanorales
, Cladoniaceae
) - 17;
эпигеидный
,
единично
.
#Сladonia crispata
(Ach.) Flot. var. elegans
Elenk. (Lecanorales
, Cladoniaceae
) - 3; эпигеидный, единично.
55. !#Сladonia deformis
(L.) Hoffm. (Lecanorales
, Cladoniaceae
) - 3; эпиксил, редко.
56. *Сladonia digitata
(L.) Hoffm. (Lecanorales
, Cladoniaceae
) - 3; эпиксил, единично.
57. Сladonia fimbriata
(L.) Fr. (Lecanorales
, Cladoniaceae
) - 1, 2, 3, 4, 7, 8, 10, 14, 15, 17, 18, 19; эпигеидо-эпиксил, повсеместно.
58. !#Сladonia
foliacea
(Huds.) Willd. (Lecanorales
, Cladoniaceae
) - 3; эпигеидный, редко.
59. !Сladonia furcata
(Huds) Schrad. (Lecanorales
, Cladoniaceae
) - 3; эпигеидный, редко.
#Сladonia furcata
(Huds) Schrad. var. pinnata
[(Flk.) Vain.] (Lecanorales
, Cladoniaceae
) - 3; эпигеидный, единично.
#Сladonia furcata
(Huds) Schrad. var. racemosa
[(Hoffm.) Flk.] (Lecanorales
, Cladoniaceae
) - 3; эпигеидный, единично.
60. !Сladonia gracilis
(L.) Willd. (Lecanorales
, Cladoniaceae
) – 3, 14, 17; эпигеидный, редко.
61. *Сladonia grayi
G.Merr. ex Sandst. (Lecanorales
, Cladoniaceae
) - 3; эпигеидный, единично.
62. Cladonia macilenta
Hoffm. (Lecanorales
, Cladoniaceae
) - syn. Cladonia bacillaris
Nyl.; 1, 3, 8, 14, 17, 19; эпигеидо-эпиксил, часто.
63. !Cladonia magyarica
Vain. (Lecanorales
, Cladoniaceae
) - 10, 14; эпигеидный, редко.
64. Cladonia parasitica
(Hoffm.) Hoffm. (Lecanorales
, Cladoniaceae
) – 3, 4, 8, 14, 18, 19; эпифито-эпиксил, часто.
65. Cladonia phyllophora
Hoffm. (Lecanorales
, Cladoniaceae
) - 3, 14; эпигеидный, редко.
66. Cladonia pyxidata
(L.) Hoffm. (Lecanorales
, Cladoniaceae
) – 3, 10, 14, 19; эпигеидо-эпиксил, часто.
67. !Cladonia rangiferina
(L.) F.H.Wigg. (Lecanorales
, Cladoniaceae
) - syn. Cladina
rangiferina
(L.) Nyl.; 3, 19; эпигеидный, редко.
68. Cladonia rei
Schaer. (Lecanorales
, Cladoniaceae
) - 1, 3, 8, 10, 14, 15, 17; эпигеидный, часто.
69. *Cladonia squamosa
Hoffm. (Lecanorales
, Cladoniaceae
) - 3; эпифито-эпиксил, редко.
70. Cladonia stellaris
(Opiz) Pouzar & Vezda (Lecanorales
, Cladoniaceae
) – syn. Cladina
stellaris
(Opiz) Brodo; 14; эпигеидный, редко.
71. !*Cladonia subrangiformis
Sandst. (Lecanorales
, Cladoniaceae
) - 14; эпигеидный, единично.
72. !#Cladonia subulata
(L.) Weber ex F.H.Wigg. (Lecanorales
, Cladoniaceae
) - syn. Cladonia cornutoradiata
Zopf; 3, 19; эпигеидо-эпиксил, редко.
73. !*Cladonia symphycarpa
(Flörke) Fr. (Lecanorales
, Cladoniaceae
) - 10, 14; эпигеидный, редко.
74. Cladonia uncialis
(L.) Weber ex F.H.Wigg. (Lecanorales
, Cladoniaceae
) - 3, 19; эпигеидный, редко.
75. !Cladonia verticillata
(Hoffm.) Schaer. (Lecanorales
, Cladoniaceae
) - syn. Cladonia
cervicornis
ssp. verticillata
(Hoffm.) Flot.; 3; эпигеидный, редко.
76. !Collema crispum
(Huds) Weber ex F.H.Wigg. (Lecanorales
, Collemataceae
) – 3, 4, 8, 10, 11, 14, 17; эпилит, часто.
77. !#ollema cristatum
(L.) Weber ex F.H.Wigg. (Lecanorales
, Collemataceae
) – 3; эпилит, редко.
78. #Collema minor
(Pakh.) Tomin (Lecanorales
, Collemataceae
) – 1, 14;. эпигеидный, редко.
79. Collema tenax
(Sw.) Ach. em. Degel. (Lecanorales
, Collemataceae
) – 1, 4, 8, 10, 11, 14, 17, 19; эпигеидный, часто.
80. *Diploschistes muscorum
(Scop.) R.Sant. in Hawksw., James & Coppins (Ostropales
, Thelotremataceae
) - 17; эврисубстратный, единично.
81. #Diplotomma alboatrum
(Hoffm) Flot. (Lecanorales
, Physciaceae
) - syn. Buellia
alboatra
(Hoffm) Th.Fr.; 14; эврисубстратный, единично.
82. Endocarpon pusillum
Hedw. (Verrucariales
, Verrucariaceae
) – 4, 8, 10, 11, 17;
эпигеидный
,
часто
.
83. !*Evernia mesomorpha
Nyl. (Lecanorales
, Parmeliceae
) - 3; эпифит, единично.
84. Evernia prunastri
(L.) Ach. (Lecanorales
, Parmeliceae
) - 1, 3, 4, 5, 7, 14, 17, 18, 19; эпифит, повсеместно.
85. #Graphis scripta
(L.) Ach. (Ostropales
, Graphidaceae
) – 3; эпифит, редко.
86. Hypocenomyce scalaris
(Ach.) M.Choisy (Lecanorales
, Lecideaceae
) - syn. Psora ostreata
Hoffm.; 1, 3, 14, 17, 18, 19; эпифито-эпиксил, часто.
87. Hypogymnia physodes
(L.) Nyl. (Lecanorales
, Parmeliceae
) - syn. Parmelia physodes
(L.) Ach.; 1, 2, 3, 4, 5, 7, 8, 14, 15, 17, 18, 19; эпифито-эпиксил, повсеместно.
88. Hypogymnia tubulosa
(Schaer.) Hav. (Lecanorales
, Parmeliceae
) - syn. Parmelia tubulosa
(Schaer.) Bitter; 3, 5, 7, 17; эпифито-эпиксил, часто.
89. !#Icmadophila ericetorum
(L.) Zahlbr. (Leothiales
, Icmadophilaceae
) – 3; эпиксил, единично.
90. Lecania cyrtella
(Ach.) Th.Fr. (Lecanorales
, Bacidiaceae
) – 4, 8, 10, 17, 19;
эпифит
,
часто
.
91. *Lecania cyrtellina
(Nyl.) Sandst. (Lecanorales
, Bacidiaceae
) - 4, 8, 17, 19;
эпифит
,
часто
.
92. *Lecania ephedrae
Elenk. (Lecanorales
, Bacidiaceae
) – syn. Lecania alexandrae
Tomin; 4, 11, 19;
эпифит
,
редко
.
93. Lecania fuscella
(Schaer.) A.Massal. (Lecanorales
, Bacidiaceae
) – 1, 3, 4, 8, 10, 11, 14, 17, 19;
эпифит
,
часто
.
94. *Lecania hyalina
(Fr.) R.Sant. comb.nov. (Lecanorales
, Bacidiaceae
) - syn. Lecania
globulosa
(Flörke) van den Boom & Serus, Catillaria globulosa
(Flörke) Th.Fr., Bacidia
globulosa
(Flörke) Hafellner & V.Wirth; 3, 8; эпифито-эпиксил, редко.
95. Lecania naegelii
(Hepp) Diederih & van den Boom (Lecanorales
, Bacidiaceae
) – syn Bacidia naegelii
(Hepp) Zahlbr.; 1, 3, 8, 14;
эпифит
,
часто
.
96. Lecanora albella
(Pers.) Ach. (Lecanorales
, Lecanoraceae
) – syn. Lecanora pallida
(Schreb.) Rabenh.; 3, 14, 15; эпифит, часто.
97. Lecanora allophana
Nyl. (Lecanorales
, Lecanoraceae
) – 1, 2, 3, 4, 5, 14, 18, 19; эпифито-эпиксил, повсеместно.
98. Lecanora argentata
(Ach.) Malme (Lecanorales
, Lecanoraceae
) – syn Lecanora
subfuscata
H.Magn.; 1, 3, 4, 5, 14, 19; эпифито-эпиксил, часто.
99. Lecanora carpinea
(L.) Vain. (Lecanorales
, Lecanoraceae
) – 1, 2, 3, 4, 5, 8, 14, 15, 17, 19; эпифито-эпиксил, повсеместно.
100. *Lecanora cateilea
(Ach.) A.Massal. (Lecanorales
, Lecanoraceae
) – 8, 19;
эпифит
,
редко
.
101. Lecanora chlarotera
Nyl. (Lecanorales
, Lecanoraceae
) - syn. Lecanora crassula
H.Magn.; 2, 3, 4, 5, 7, 8, 10, 14, 17, 19; эпифито-эпиксил, повсеместно.
102. #Lecanora circumborealis
Brodo & Vitik. (Lecanorales
, Lecanoraceae
) - 14; эпифит, единично.
103. *Lecanora conizaeoides Nyl. ex Cromb. (Lecanorales, Lecanoraceae) –
3; эпифит, единично.
104. *Lecanora dispersa
(Pers.) Sommerf. (Lecanorales
, Lecanoraceae
) – 3, 17; эпилит, редко.
105. Lecanora expallens
Ach. (Lecanorales
, Lecanoraceae
) – 3, 8, 14; эпифит, часто.
106. Lecanora hagenii
(Ach.) Ach. (Lecanorales
, Lecanoraceae
) – 2, 3, 4, 5, 8, 10, 11, 14, 17, 19; эврисубстратный, часто.
107. *Lecanora intumescens
(Rebent.) Rabenh. (Lecanorales
, Lecanoraceae
) – 7;
эпифит
,
единично
.
108. #Lecanora leptyrodes
(Nyl.) Degel. (Lecanorales
, Lecanoraceae
) – 19; эпифит, единично.
109. Lecanora populicola
(DC.) Duby (Lecanorales
, Lecanoraceae
) - syn Lecanora distans
(Ach.) Nyl.; 3, 8, 14, 18, 19; эпифит, часто.
110. Lecanora pulicaris
(Pers.) Ach. (Lecanorales
, Lecanoraceae
) - 3, 4, 5, 7, 14; эпифито-эпиксил, часто.
111. #Lecanora rugosella
Zahlbr. (Lecanorales
, Lecanoraceae
) – 14; эпифит, единично.
112. Lecanora saligna
(Schrad.) Zahlbr. (Lecanorales
, Lecanoraceae
) - 2, 3, 14, 19; эпифито-эпиксил, часто.
113. *Lecanora sambuci
(Pers.) Nyl. (Lecanorales
, Lecanoraceae
) – 4, 8, 14, 15, 17; эпифит, часто.
114. Lecanora subcarpinea
Szatala (Lecanorales
, Lecanoraceae
) – syn Lecanora nemoralis
Makar.; 3, 8, 19; эпифит, редко.
115. #Lecanora subrugosa
Nyl. (Lecanorales
, Lecanoraceae
) – 3; эпифит, единично.
116. Lecanora symmicta
(Ach.) Ach. (Lecanorales
, Lecanoraceae
) – syn Biatora symmicta
(Ach.) Mass.; 3, 15; эпифито-эпиксил, редко.
117. Lecanora umbrina
(Ach.) A.Massal. (Lecanorales
, Lecanoraceae
) – 4, 11, 14, 17; эпилит, часто.
118. Lecanora varia
(Hoffm.) Ach. (Lecanorales
, Lecanoraceae
) – 2, 3, 8, 17, 19; эпифито-эпиксил, часто.
119. #Lecidea albofuscescens
Nyl. (Lecanorales
, Lecideaceae
) - 14; эпифит, единично.
120. Lecidella elaeochroma
(Ach.) M.Choisy (Lecanorales
, Lecanoraceae
) - syn. Lecidea
olivacea
(Hoffm.) A.Massal., Lecidea elaeochroma
(Ach.) Ach.; 1, 2, 3, 5, 8, 14, 17, 18, 19;
эпифит
,
часто
.
121. Lecidella euphorea
(Flőrke) Hertel in Hawksw., James & Coppins (Lecanorales
, Lecanoraceae
) - syn. Lecidea euphorea
(Flőrke) Nyl.; 1, 3, 4, 5, 8, 14, 17, 19; эпифито-эпиксил, часто.
122. Lepraria incana
(L.) Ach. (
гр
. Fungi imperfecti
) - syn. Lepraria aeruginosa
auct. non (Weiss) Sm.; 1, 3, 4, 7, 8, 14, 19;
эврисубстратный
,
часто
.
123. Lepraria lobificans
Nyl. (гр. Fungi imperfecti
) – 3, 19; эпифит, редко.
124. #Lepraria membranacea
(Dicks.) Vain. (гр. Fungi imperfecti
) – syn Leproloma membranaceum
(Dicks.) J.R.Laundon; 4; эврисубстратный, единично.
125. Lepraria vouauxii
(Hue) R.C.Harris (гр. Fungi imperfecti
) – syn. Leproloma vouauxii
(Hue) J.R.Laundon; 3, 4, 14; эпибриофит, редко.
126. !Leptogium shraderi
(Bernh.) Nyl. (Lecanorales
, Collemataceae
) – 1, 10, 17; эпибриофит, редко.
127. Megaspora verrucosa
(Ach.) Hafellner & V.Wirth in Wirth (Pertusariales
, Megasporaceae
) - syn. Aspicilia verrucosa
(Ach.) Körb.; 3, 10; эпигеидный, редко.
128. #Melanelia elegantula
(Zahlbr.) Essl. (Lecanorales
, Parmeliceae
) - 3; эпифит, единично.
129. Melanelia exasperata
(De Not.) Essl. (Lecanorales
, Parmeliceae
) - syn. Parmelia exasperata
De Not.; 1, 3, 5, 8, 10, 14, 17, 19; эпифит, неморальный, голарктический, часто.
130. Melanelia exasperatula
(Nyl.) Essl. (Lecanorales
, Parmeliceae
) - syn. Parmelia
exasperatula
Nyl.; 3, 5, 10, 17, 19; эпифит, часто.
131. Melanelia fuliginosa
(Fr. ex Duby) Essl. in Egan (Lecanorales
, Parmeliceae
) - 3, 14, 17, 19; эпифит, часто.
132. !Melanelia glabra
(Schaer.) Essl. (Lecanorales
, Parmeliceae
) - syn. Parmelia glabra
(Schaer.) Nyl.; 3; 17, 19; эпифит, редко.
133. Melanelia subargentifera
(Nyl.) Essl. (Lecanorales
, Parmeliceae
) - syn. Parmelia
subargentifera
Nyl.; 1, 2, 3, 4, 5, 8, 15, 17, 18, 19; эпифит, часто.
#Melanelia subargentifera
(Nyl.) Essl. var. conspurcata
[Hillm.] (Lecanorales
, Parmeliceae
) - 3; эпифит, еднично.
134. #Micarea melaena
(Nyl.) Hedl. (Lecanorales
, Micareaceae
) - 4, 19; эпибриофит, редко.
135. #Micarea misella
(Nyl.) Hedl. (Lecanorales
, Micareaceae
) - 3; эпиксил, единично.
136. Micarea nitschkeana
(J.Lahm.ex Rabenh.) Harm. (Lecanorales
, Micareaceae
) - syn. Bacidia nitschkeana
(J.Lahm ex Rabenh.) Zahlbr.; 1, 3; эпиксил, редко.
137. #Mycobilimbia berengeriana
(A.Massal.) Hafellner & V.Wirth in Wirth (Lecanorales
, Porpidiaceae
) - 14; эпибриофит, единично.
138. *Mycobilimbia carneoalbida
(Műll.Arg.) Printzen (Lecanorales
, Porpidiaceae
) - syn. Bacidia sphaeroides
auct. non (Dicks.) Zahlbr., Biatora carneoalbida
(Müll.Arg.) Coppins; 17; эпибриофит, единично.
139. ¢*Mycobilimbia epixanthoides
(Nyl.) Vitik. et al. in Hafellner & Tǔrk (Lecanorales
, Porpidiaceae
) – 5;
эпибриофит
,
единично
.
140. *Mycobilimbia pilularis
(Körb.) Hafellner & Türk (Lecanorales
, Porpidiaceae
) - syn. Bacidia sphaeroides
(Dicks.) Zahlbr.; 17; эпифит, единично.
141. *Mycobilimbia tetramera
(De Not.) Vitik. et al. in Hafellner & Türk (Lecanorales
, Porpidiaceae
) – 4; эпибриофит, единично.
142. Mycoblastus fucatus
(Stirt.) Zahlbr. (Lecanorales
, Mycoblastaceae
) - syn. Mycoblastus
sterilis
Coppins & P.James; 3, 19; эпифит, редко.
143. *Mycocalicium subtile
(Pers.) Szatala (гр. Incertae sedis
, Mycocaliciaceae
) - 3, 14; эпифито-эпиксил, редко.
144. *Myxobilimbia microcarpa
(Th.Fr.) Hafellner in Hafellner & Türk (Lecanorales
, Porpidiaceae
) – 4; эпибриофит, единично.
145. *Myxobilimbia sabuletorum
(Schreb.) Hafellner in Hafellner & Türk (Lecanorales
, Porpidiaceae
) - syn. Mycobilimbia sabuletorum
(Schreb.) Hafellner; 19; эпибриофит, единично.
146. #О
pegrapha atra
Pers. (Ostropales
, Graphidaceae
) – 3; эпифит, единично.
147. #О
pegrapha
rufescens
Pers. (Ostropales
, Graphidaceae
) – syn. Opegrapha herpetica
(Ach.) Ach.; 3; эпифит; единично.
148. ¢*Pachyphyale fagicola
(Hepp) Zwackh (Gyalectales
, Gyalectaceae
) - 17; эпифит, единично.
149. Parmelia sulcata
Taylor (Lecanorales
, Parmeliceae
) - 1, 2, 3, 4, 5, 7, 8, 14, 15, 17, 18, 19; эпифито-эпиксил, повсеместно.
150. Parmelina quercina
(Willd.) Hale (Lecanorales
, Parmeliceae
) - syn. Parmelia quercina
(Willd.) Vain.; 1, 3, 4, 8, 14, 17; эпифит, часто.
151. Parmelina tiliacea
(Hoffm.) Hale (Lecanorales
, Parmeliceae
) - syn. Parmelia tiliacea
(Hoffm.) Ach., Parmelia scortea
(Ach.) Ach.; 2, 3, 4, 8, 14, 19; эврисубстратный, часто.
152. Parmeliopsis ambiqua
(Wulfen) Nyl. (Lecanorales
, Parmeliaceae
) – 3; эпифит, редко.
153. *Parmeliopsis hyperopta
(Ach.) Arnold (Lecanorales
, Parmeliaceae
) – 4, 14; эпифито-эпиксил, редко.
154. #Peltigera canina
(L.) Willd. (Peltigerales
, Peltigeraceae
) – 3, 14, 19; эпигеидный, редко.
#Peltigera canina
(L.) Willd var. subcanina
[(Gieln) Frey.] (Peltigerales
, Peltigeraceae
) –
14; эпигеидный, единично.
155. !#Peltigera
neckeri
Hepp ex Müll.Arg. (Peltigerales
, Peltigeraceae
) – 14, эпигеидный, единично.
156. !Peltigera ponojensis
Gyeln. (Peltigerales
, Peltigeraceae
) – 19; эпигеидный, редко.
157. !#Peltigera praetextata
(Flörke ex Sommerf.) Zopf (Peltigerales
, Peltigeraceae
) – 4, 14; эпигеидный, редко.
158. Peltigera rufescens
(Weiss) Humb. (Peltigerales
, Peltigeraceae
) – 4, 10, 14, 17; эпигеидный, часто.
159. Pertusaria albescens
(Huds.) M.Choisy & Werner in Werner (Pertusariales
, Pertusariaceae
) - syn. Pertusaria globulifera
(Turner) A.Massal.; 3, 14; эпифит, редко.
160. *Pertusaria amara
(Ach.) Nyl. (Pertusariales, Pertusariaceaea) - 3; эпифит, единично.
161. Pertusaria coccodes
(Ach.) Nyl. (Pertusariales
, Pertusariaceaea
) - 3; эпифит, редко.
162. #Phaeophyscia ciliata
(Hoffm.) Moberg (Lecanorales
, Physciaceae
) - 3; эврисубстратный, единично.
163. Phaeophyscia nigricans
(Flörke) Moberg (Lecanorales
, Physciaceae
) - 2, 3, 5, 8, 17; эврисубстратный, часто.
164. Phaeophyscia orbicularis
(Neck.) Moberg; (Lecanorales
, Physciaceae
) - syn. Physcia
orbicularis
(Neck.) Poetsch; 2, 3, 4, 5, 8, 10, 14, 15, 17, 18, 19; эврисубстратный, повсеместно.
165. #Phlyctis agelae
(Ach.) Flot. (Lecanorales
, Phlyctidaceae
) – 3; эпифит, единично.
166. Phlyctis argena
(Spreng.) Flot. (Lecanorales
, Phlyctidaceae
) - 1, 3, 8, 14, 19; эпифит, часто.
167. Physcia adscendens
H.Olivier nom. cons. (Lecanorales
, Physciaceae
) - 2, 3, 4, 5, 7, 8, 10, 14, 15, 19; эпифит, повсеместно.
168. Physcia aipolia
(Ehrh. ex Humb.) Fürnr. (Lecanorales
, Physciaceae
) - 2, 3, 4, 8, 14, 17, 19; эпифит, часто.
169. #Physcia caesia
(Hoffm.) Fürnr. (Lecanorales
, Physciaceae
) – 3;
эпилит
,
единично
.
170. *Physcia dubia
(Hoffm.) Lettau (Lecanorales
, Physciaceae
) – 3, 4, 5, 15, 17, 19;
эврисубстратный
,
часто
.
171. Physcia stellaris
(L.) Nyl. (Lecanorales
, Physciaceae
) - 1, 2, 3, 4, 5, 8, 10, 14, 15, 17, 19; эпифито-эпиксил, повсеместно.
172. Physcia tenella
(Scop.) DC. in Lamarck & Candolle nom. cons (Lecanorales
, Physciaceae
) - syn Physcia hispida
(Screb.) Frege; 2, 3, 4, 5, 7, 8, 14, 17, 18, 19; эврисубстратный, повсеместно.
173. Physcia tribacia
(Ach.) Nyl. (Lecanorales
, Physciaceae
) – 3, 4, 14;
эврисубстратный
,
редко
.
174. !*Physconia detersa
(Nyl.) Poelt (Lecanorales
, Physciaceae
) – 3, 4, 5, 8;
эпифит
,
часто
.
175. Physconia distorta
(With.) J.R.Laundon (Lecanorales
, Physciaceae
) - 1, 2, 3, 4, 5, 7, 8, 14, 15, 17, 18, 19; эпифит, повсеместно.
176. Physconia enteroxanta
(Nyl.) Poelt (Lecanorales
, Physciaceae
) - 1, 2, 3, 4, 5, 7, 8, 10, 14, 17, 18, 19; эпифито-эпиксил, повсеместно.
177. #Physconia grisea
(Lam.) Poelt; (Lecanorales
, Physciaceae
) –3;
эпифито
-
эпиксил
,
единично
.
178. Physconia perisidiosa
(Erichsen) Moberg (Lecanorales
, Physciaceae
) – 1, 3, 19;
эпифит
,
редко
.
179. Placidium lachneum
(Ach.) Breuss (Verrucariales
, Verrucariaceae
) - syn. Catapyrenium lachneum
(Ach.) Breuss
; 8, 11, 14, 17; эпилит, часто.
180. Placidium squamulosum
(Ach.) Breuss (Verrucariales
, Verrucariaceae
) - syn. Catapyrenium squamulosum
(Ach.) Breuss
; 4, 8, 10, 11, 14, 17; эпигеидный, часто.
181. Placynthiella dasaea
(Stirt.) T
Ø
nsberg (Lecanorales
, Trapeliaceae
) - 3, 5, 8, 14, 17, 19;
эпифито
-
эпиксил
,
часто
.
182. Placynthiella icmalea
(Ach.) Coppins & P.James (Lecanorales
, Trapeliaceae
) - 3, 4, 5, 14, 19; эпиксил, часто.
183. Placynthiella uliginosa
(Schrad.) Coppins & P.James (Lecanorales
, Trapeliaceae
) - 17, 19;
эпиксил
,
редко
.
184. !*Platismatia glauca
(L.) W.L.Culb. & C.F.Culb. (Lecanorales
, Parmeliaceae
) - syn. Cetraria glauca
(L.) Ach.; 3; эпифит, редко.
185. Pleurosticta acetabulum
(Neck.) Elix & Lumbsh in Lumbsch, Kothe & Elix (Lecanorales
, Parmeliceae
) - syn. Parmelia acetabulum
(Neck.) Duby, Melanelia acetabulum
(Neck.) Essl.; 1, 2, 3, 4, 5, 8, 10, 14, 15, 17, 18, 19; эпифито-эпиксил, повсеместно.
186. #Polychidium muscicola
(Sw.) Gray (Peltigerales
, Placynthiaceae
) – 19; эпибриофит, единично.
187. Protoblastenia rupestris
(Scop.) J.Steiner (Lecanorales
, Psoraceae
) - 4, 10, 14; эпилит, редко.
188. *Protoparmeliopsis muralis
(Schreb.) M.Choisy (Lecanorales
, Lecanoraceae
) – syn. Lecanora muralis
(Schreb.) Rabenh
.; 5, ур. Гнилое, степь, на бетоне; эпилит, единично.
189. !#Pseudevernia furfuracea
(L.) Zopf (Lecanorales
, Parmeliceae
) - syn. Evernia furfuracea
(L.) W.Mann; 3; эпифит, редко.
190. #Pyrenula laevigata
(Pers.) Arnold (Pyrenulales
, Pyrenulaceae
) – 3; эпифит, единично.
191. #Pyrenula nitida
(Weigel) Ach. (Pyrenulales
, Pyrenulaceae
) – 3; эпифит, единично.
192. #Pyrenula nitidella
(Flörke ex Schaer.) Műll.Arg. (Pyrenulales, Pyrenulaceae) – syn Pyrenula dermatodes
(Borrer) Schaer.; 3; эпифит, единично.
193. !Ramalina farinacea
(L.) Ach. (Lecanorales
, Ramalinaceae
) – 1, 3, 4; эпифит, часто.
194. Ramalina fraxinea
(L.) Ach. (Lecanorales
, Ramalinaceae
) – 3, 8, 14, 17; эпифит, часто.
195. Ramalina pollinaria
(Westr.) Ach. (Lecanorales
, Ramalinaceae
) – 1, 2, 3, 8, 14, 17, 19; эпифит, часто.
196. *Rinodina archaea
(Ach.) Arnold (Lecanorales
, Physciaceae
) – 19;
эпифит
,
единично
.
197. #Rinodina colobina
(Ach.) Th.Fr. (Lecanorales
, Physciaceae
) – 14;
эпифит
,
единично
.
198. *Rinodina exiqua
(Ach.) Gray (Lecanorales
, Physciaceae
) – 3, 8, 14, 17, 19;
эпифито
-
эпиксил
,
часто
.
199. Rinodina pyrina
(Ach.) Arnold (Lecanorales
, Physciaceae
) - 1, 3, 4, 5, 7, 8, 10, 15, 17, 19; эпифит, часто.
200. Rinodina septentrionalis
Malme (Lecanorales
, Physciaceae
) - 14, 17, 19; эпифит, редко.
201. Rinodina sophodes
(Ach.) A.Massal. (Lecanorales
, Physciaceae
) – 4, 14, 17; эпифит, редко.
202. ¢*Rinodina terrestris
Tomin. (Lecanorales
, Physciaceae
) – syn. Rinodina mucronatula
H.Magn.; 14; эпиксил, единично.
203. Sarcogyne regularis
Körb. (Lecanorales
, Acarosporaceae
) - 4, 8, 10, 14, 17; эпилит, часто.
204. Scoliciosporum chlorococcum
(Graewe ex Stenh.) Vézda (Lecanorales
, Lecanoraceae
) - 3, 4, 14, 18, 19; эврисубстратный, часто.
205. *Scoliciosporum umbrinum
(Ach.) Arnold (Lecanorales
, Lecanoraceae
) - 3; эпифит, редко.
206. #Squamarina lentigera
(Weber) Poelt (Lecanorales
, Bacidiaceae
) - 19; эпилит, единично.
207. Staurothele hymenogonia
(Nyl.) Th.Fr. (Verrucariales
, Verrucariaceae
) - 14;
эпилит
,
редко
.
208. #Sticta sylvatica
(Huds) Ach. (Peltigerales
, Lobariaceae
) – 19; эпигеидный, единично.
209. #Strangospora deplanata
(Almq.) Clauzade & Cl.Roux. (Lecanorales
, Lecanoraceae
) - 19; эпифит, единично.
210. *Strangospora moriformis
(Ach.) Stein (Lecanorales
, Lecanoraceae
) - 3; эпифито-эпиксил, единично.
211. Toninia sedifolia
(Scop.) Timdal (Lecanorales
, Catillariaceae
) – 1, 3, 4, 8, 10, 17; эпигеидный, часто.
212. #Toninia
cf. taurica
(Szatala) Oxner (Lecanorales
, Catillariaceae
) - 1; эпигеидный, единично.
213. Trapeliopsis flexuosa
(Fr.) Coppins & P.James (Lecanorales
, Trapeliaceae
) - 4, 5, 8, 14, 17, 19;
эпиксил
,
часто
.
214. Trapeliopsis granulosa
(Hoffm.) Lumbsch (Lecanorales
, Trapeliaceae
) - 8, 14, 19;
эпигеидный
,
часто
.
215. *Trapeliopsis viridescens
(Schrad.) Coppins & P.James (Lecanorales
, Trapeliaceae
) - 14, 17; эпибриофит, редко.
216. !#Tuckermannopsis chlorophylla
(Willd.) Hale in Egan (Lecanorales
, Parmeliceae
) - syn. Cetraria chlorophylla
(Willd.) Vain.; 3; эпифит, единично.
217. !Usnea hirta
(L.) Weber ex F.H.Wigg. (Lecanorales
, Parmeliceae
) - 3, 17; эпифит, редко.
218. *Usnea subfloridana
Stirt. (Lecanorales
, Parmeliaceae
) – 3; эпифит, единично.
219. #Verrucaria acrotella
Ach. (Verrucariales
, Verrucariaceae
) - 1, 14;
эпилит
,
редко
.
220. #Verrucaria cоerulea
DC. in Lamarck & Candolle (Verrucariales
, Verrucariaceae
) - 14;
эпилит
,
единично
.
221. Verrucaria calciseda
auct. (non DC.) (Verrucariales
, Verrucariaceae
) - 1, 4, 14, 17;
эпилит
,
часто
.
222. ¢*Verrucaria cretophila
Oxner (Verrucariales
, Verrucariaceae
) - 10, 14;
эпилит
,
редко
.
В настоящее время вид известен из двух точек – на территории России и Украины - в пределах выходов мела в долине р. Северский Донец. Ранее указывался как эндем Украины (Определитель…, 1977). Новой вид для России.
223. #Verrucaria floerkeana
Dalla Torre & Sarnth. (Verrucariales
, Verrucariaceae
) - 8;
эпилит
,
единично
.
224. ¢*Verrucaria foveolata
(Flörke) A.Massal. (Verrucariales
, Verrucariaceae
) - 8;
эпилит
,
единично
.
225. #Verrucaria fusca
Pers. (Verrucariales
, Verrucariaceae
) - 4;
эпилит
,
единично
.
226. Verrucaria fuscella
(Turner) Winch (Verrucariales
, Verrucariaceae
) - 8, 14, 17;
эпилит
,
часто
.
227. ¢*Verrucaria macrostoma
Dufour ex DC. in Lamarck & Candolle (Verrucariales
, Verrucariaceae
) - 14;
эпилит
,
редко
.
228. Verrucaria muralis
Ach. (Verrucariales
, Verrucariaceae
) - syn. Verrucaria rupestris
Schrad. non (Scop.) Weber ex F.H.Wigg.; 1, 4, 5, 8, 10, 11, 14, 17, 19;
эпилит
,
часто
.
229. ¢*Verrucaria murina
Leight. (Verrucariales
, Verrucariaceae
) - 11;
эпилит
,
единично
.
230. Verrucaria nigrescens
Pers. (Verrucariales
, Verrucariaceae
) - 11, 14, 17; эпилит, часто.
231. *Verrucaria viridula
(Schrad.) Ach. (Verrucariales
, Verrucariaceae
) - 14;
эпилит
,
единично
.
232. Vulpicida pinastri
(Scop.) J.-E.Mattsson et M.J.Lai (Lecanorales
, Parmeliceae
) - syn. Cetraria pinastri
(Scop.) Gray, Cetraria caperata
(L.) Vain.; 3, 14, эпифито-эпиксил, редко.
233. Xanthoria parietina
(L.) Th.Fr. (Teloschistales
, Teloschistaceae
) – 2, 3, 4, 5, 7, 8, 10, 14, 15, 17, 18; эврисубстратный, повсеместно.
234. Xanthoria polycarpa
(Hoffm.) Th.Fr. ex Rieber (Teloschistales
, Teloschistaceae
) – 1, 2, 3, 4, 5, 8, 10, 14, 17, 19; эпифито-эпиксил, повсеместно.
235. ¢*Xanthoria ukrainica
S. Kondr. (Teloschistales
, Teloschistaceae
) – 3; эпифит, единично.
Рис. 7.1.2.1. Степень изученности особо охраняемых природных территорий
Белгородской области
территории, сведения о лишайниках которых известны только из литературы
оригинальные исследования
Районы Белгородской области
1. Алексеевский 12. Краснояружский
2. Белгородский 13. Корочанский
3. Борисовский 14. Новооскольский
4. Валуйский 15. Прохоровский
5. Вейделевский 16. Ракитянский
6. Волоконовский 17. Ровеньской
7. Грайворонский 18. Старооскольский
8. Губкинский 19. Шебекинский
9. Ивнянский 20. Чернянский
10. Красненский 21. Яковлевский
11. Красногвардейский
7.1.3. Видовой состав семейства
Lamiaceae
заповедника «Белогорье»
Работа по выявлению видового состава семейства губоцветные (Lamiaceae) и сбора материала проводилась на территории заповедника “Белогорье” в летний период 2004 г. Обследованы три участка заповедника “Белогорье”: Ямская степь, Лысые горы, Астрасьевы яры (официально-документальное название участка – Острасьевы яры, прим. Ред.). В ходе экспедиционных исследований маршрутно-флористическим методом (Алехин, 1926) с составлением списков и указанием конкретных местообитаний каждого вида было обнаружено 29 видов семейства Lamiaceae. Из них: 18 - в Ямской степи, 20 - в Лысых горах, 17 - в Астрасьевых ярах.
В ходе работы собрано более сотни гербарных образцов. В приводимом ниже списке систематическое положение выявленных видов растений дано в соответствии с работой «Флора СССР» (1954-1955), в соответствии с системой А. Энглера. Номенклатура таксонов указана по С.К. Черепанову (Черепанов, 1995). После латинского названия в списке дано русское название, информация о местообитаниях вида в пределах обследованной территории для каждого участка заповедника “Белогорье”.
Условные обозначения: ЛГ – Лысые горы, ЯС – Ямская степь, АЯ – Астрасьевы яры.
Аннотированный список видов сем.
Lamiaceae
1. Ajuga
genevensis
L
.
– Живучка женевская. ЛГ: по опушкам, в березняке, по лесным культурам.
2. Scutellaria hastifolia L.
– Шлемник копьелистный. ЯС: лог Суры, вершина остепненного склона с кустарником.
3. Nepeta
pannonica
L
.
– Котовник венгерский. ЛГ: по лесным культурам, на опушках; ЯС: степной склон восточной экспозиции лога Суры.
4. Nepeta
cataria
L
.
– Котовник кошачий. ЛГ: в березняке.
5. Glechoma
hederaceae
L
.
– Будра плющевидная. ЛГ: по опушкам, остепненным склонам, в естественном широколиственном лесу.
6. Prunella
vulgaris
L
.
– Черноголовка обыкновенная. ЛГ: по опушкам леса; ЯС: дно лога Суры, дно балки Еремкин лог; АЯ: по опушкам леса.
7. Prunella grandiflora (L.) Scholl
.
– Черноголовка крупноцветковая. ЛГ: остепненные склоны; ЯС: по всей степи.
8. Phlomis
tuberosa
L
.
– Зопник клубненосный. ЛГ: по степным склонам, на опушках; ЯС: степной склон восточной экспозиции лога Суры; АЯ: изголовье балки, по опушкам.
9. Plomis
pungens
Willd
.
– Зопник колючий. ЛГ: остепненные склоны.
10. Galeopsis
ladanum
L
.
– Пикульник ладанниковый. ЛГ: по противопожарной вспаханной полосе, по обочинам дорог; АЯ: охранная зона, на границе дороги и пшеничного поля.
11. Galeopsis
tetrahit
L
.
– Пикульник обыкновенный. ЯС: остепненный склон западной экспозиции лога Суры.
12. Galeopsis
bifida
Boenn
.
– Пикльник двурасщепленный. ЯС: Лубушева степь, возле небольшой западины с кустарником; АЯ: вершина остепненного склона западной экспозиции.
13. Leonurus
quinquelobatus
Gilib
.
– Пустырник пятилопастный. ЛГ: по опушкам, среди лесных культур; ЯС: дно лога Суры; АЯ: по опушкам, зарослям кустарников.
14. Ballota
nigra
L
.
– Белокудренник черный. ЯС: дерезняки в степи; АЯ: по опушкам, зарослям кустарников, вдоль дорог.
15. Stachys
annua
(
L
.)
L
.
– Чистец однолетний. ЛГ: в березняке (88 кв., 15выд), по обочинам дорог.
16. Stachys
palustris
L
.
– Чистец болотный. ЛГ: в березняке дно оврага (88 кв., 13 выд.); ЯС: дно балки Еремкин лог; АЯ: дно лесной части балки, у ручья.
17. Stachys
recta
L
.
– Чистец прямой. – ЛГ: по степным склонам, по опушкам; ЯС: по всей степи; АЯ: степные склоны.
18. Stachys officinalis (L.) Trevis
.
– Чистец лекарственный (Буквица лекарственная). ЛГ: по опушкам; ЯС: по всей степи; АЯ: по опушкам леса.
19. Salvia verticillata L.
– Шалфей мутовчатый. ЛГ: в разреженных культурах, степных склонах.
20. Salvia
nutans
L
.
– Шалфей поникающий. ЛГ: остепненным склонам; АЯ: остепненные склоны балки.
21. Salvia tesquicola Klok.
&
Pobed
.
– Шалфей остепненный. ЯС: дно лога Вишняки; АЯ: южная часть балки, по бровке олесенного склона.
22. Salvia pratensis L.
– Шалфей луговой. ЛГ: на опушках, по степным склонам; ЯС: дно балки Еремкин лог; АЯ: по опушкам, зарослям кустарников.
23. Clinopodium
vulgare
L
.
– Пахучка обыкновенная. ЯС: дно балки Еремкин лог в кустарнике; АЯ: в зарослях кустарников остепненного склона западной экспозиции.
24. Acinos arvensis (Lam.) Dandy
– Щебрушка полевая. ЛГ: по степным склонам.
25. Origanum
vulgare
L
.
– Душица обыкновенная. ЛГ: по разреженным лесным культурам; ЯС: лог Суры; АЯ: изголовье балки, на опушке леса.
26. Thymus cretaceus Klok.
&
Shost
.
– Тимьян меловой. ЛГ: на меловых выходах остепненных склонов; ЯС: по меловым обнажением лога Вишняки.
27. Thymus marshallianus Willd. -
Тимьян Маршалла. ЯС: по всей степи; АЯ: по степным склонам.
28. Lycopus
europaeus
L
.
– Зюзник европейский. АЯ: дно лесной части балки, у ручья.
29. Mentha arvensis L.
– Мята полевая. АЯ: дно лесной части балки, у ручья.
В ходе исследований нами было выявлено 29 видов представителей семейства губоцветных, относящихся к 17 родам. Ранее не отмеченными для Лысых гор в списке Н.И. Золотухина (1995) является Stachys
palustris
L
.,
для Ямской степи в списке В.М. Покровской (1940) - Scutellaria
hastifolia
L
.,
Galeopsis
bifida
Boenn
.,
Salvia
tesquicola
Klok
. &
Pobed
.,
для Астрасьевых яров в флористическом списке Ю.А. Дорониной и др. (1993) - Galeopsis ladanum L., Galeopsis bifida Boenn., Stachys palustris L.
Таблица 7.1.3.1
Сводная таблица видов семейства Lamiaceae по трем участкам заповедника “Белогорье”
(в 2004 г.)
| Вид
|
Ямская степь
|
Лысые горы
|
Астрасьевы яры
|
| 1. Ajuga genevensis L.
|
+
|
| 2. Scutellaria hastifolia L.
|
+
|
| 3. Nepeta pannonica L.
|
+
|
+
|
| 4. Nepeta cataria L.
|
+
|
| 5. Glechoma hederaceae L.
|
+
|
| 6. Prunella vulgaris L.
|
+
|
+
|
+
|
| 7. Prunella grandiflora (L.) Scholl.
|
+
|
+
|
| 8. Phlomis tuberosa L.
|
+
|
+
|
+
|
| 9. Plomis pungens Willd.
|
+
|
| 10. Galeopsis ladanum L.
|
+
|
+
|
| 11. Galeopsis tetrahit L.
|
+
|
| 12. Galeopsis bifida Boenn.
|
+
|
+
|
| 13. Leonurus quinquelobatus Gilib.
|
+
|
+
|
+
|
| 14. Ballota nigra L.
|
+
|
+
|
| 15. Stachys annua (L.) L.
|
+
|
| 16. Stachys palustris L.
|
+
|
+
|
+
|
| 17. Stachys recta L.
|
+
|
+
|
+
|
| 18. Stachys officinalis (L.) Trevis.
|
+
|
+
|
+
|
| 19. Salvia verticillata L.
|
+
|
| 20. Salvia nutans L.
|
+
|
+
|
| 21. Salvia tesquicola Klok. & Pobed.
|
+
|
+
|
| 22. Salvia pratensis L.
|
+
|
+
|
+
|
| 23. Clinopodium vulgare L.
|
+
|
+
|
| 24. Acinos arvensis (Lam.) Dandy
|
+
|
| 25. Origanum vulgare L.
|
+
|
+
|
+
|
| 26. Thymus cretaceus Klok. & Shost.
|
+
|
+
|
| 27. Thymus marshallianus Willd.
|
+
|
+
|
| 28. Lycopus europaeus L.
|
+
|
| 29. Mentha arvensis L.
|
+
|
  

Рис. 7.1.3.2. Карта-схема Ямской степи
- Scutellaria hastifolia L.
- Nepeta pannonica L.
- Galeopsis tetrahit L.
- Galeopsis bifida Boenn.
- Stachys palustris L.
–

7.2. РАСТИТЕЛЬНОСТЬ
7.2.1. Степная растительность
7.2.1.1.а. Фенологическое состояние видов травянистых растений Ямской степи на период с 13 апреля по 29 мая 2004 года
Объектами фитофенологии были отдельные виды растений Ямской степи, встречающиеся на маршруте от ворот заповедного участка (у старого кордона) по степной дороге через большую ложбину к дальнему некосимому участку. Наблюдения осуществлялись фиксированием через 5 – 10 дней определенных фенофаз, по которым можно строить кривые вегетации, цветения и плодоношения. К сожалению во многие сроки наблюдений была дождливая погода, поэтому для некоторых видов не удалось зафиксировать все фенофазы. Принятые в таблице 7.2.1.1 сокращения: вег – вегетация; бут – бутонизация; нцв – начало цветения; цв – цветение; мцв – массовое цветение; пл – плодоношение; выс – высыпание; труб – выход в трубку (у злаков); кол – колошение (у злаков); нкол – начало колошения; аа – образует ярко выраженный аспект; а – принимает заметное участие в аспекте.
Таблица 7.2.1.1
Фенологическое состояние видов травянистых растений Ямской степи на период с 13 апреля по 29 мая 2004 года
| №
|
Название видов растений
|
Сроки наблюдений
|
| апрель
|
май
|
аспект
|
| 13
|
25
|
30
|
4
|
9
|
29
|
| 1
|
2
|
3
|
4
|
5
|
6
|
7
|
8
|
9
|
| 1
|
Adonis vernalis
|
бут
|
цв
|
цв
|
цв
|
нпл
|
пл
|
аа
|
| 2
|
Achillea millefolium
|
вег
|
вег
|
вег
|
вег
|
вег
|
вег
|
| 3
|
Alyssum calycinum
|
вег
|
вег
|
бут
|
нцв
|
цв
|
цв
|
| 4
|
Anthriscus sylvestris
|
вег
|
вег
|
вег
|
вег
|
вег
|
бут
|
| 5
|
Amoria montana
|
вег
|
вег
|
бут
|
нцв
|
а
|
| 6
|
Androsace elongata
|
бут
|
нцв
|
цв
|
цв
|
пл
|
пл
|
| 7
|
Androsace septentrionalis
|
вег
|
вег
|
бут
|
нцв
|
мцв
|
цв
|
| 8
|
Arenaria ucrainica
|
вег
|
вег
|
бут
|
бут
|
нцв
|
цв
|
| 9
|
Astragalus danicus
|
вег
|
вег
|
бут
|
бут
|
цв
|
| 10
|
Asparagus officinalis
|
вег
|
вег
|
бут
|
бут
|
нцв
|
| 11
|
Bromopsis inermis
|
вег
|
вег
|
вег
|
вег
|
вег
|
вег
|
аа
|
| 12
|
Bromopsis riparia
|
вег
|
вег
|
вег
|
вег
|
вег
|
нкол
|
аа
|
| 13
|
Bupleurum falcatum
|
вег
|
вег
|
вег
|
вег
|
вег
|
вег
|
| 14
|
Calamagrostis epigeios
|
вег
|
вег
|
вег
|
вег
|
вег
|
вег
|
аа
|
| 15
|
Campanula altaica
|
вег
|
вег
|
бут
|
нцв
|
| 16
|
Carex humilis
|
кол
|
пл
|
пл
|
пл
|
| 17
|
Carex caryophyllea
|
вег
|
бут
|
бут
|
нкол
|
кол
|
вег
|
| Таблица 7.2.1.1 (продолжение)
|
| 1
|
2
|
3
|
4
|
5
|
6
|
7
|
8
|
9
|
| 18
|
Centaurea marschalliana
|
бут
|
бут
|
бут
|
бут
|
нцв
|
мцв
|
| 19
|
Chamaecytisus ruthenicus
|
вег
|
вег
|
бут
|
бут
|
нцв
|
цв
|
а
|
| 20
|
Clematis integrifolia
|
вег
|
вег
|
вег
|
бут
|
а
|
| 21
|
Dianthus andrzejowskianus
|
вег
|
вег
|
бут
|
нцв
|
| 22
|
Draba sibirica
|
бут
|
цв
|
цв
|
цв
|
мцв
|
цв
|
аа
|
| 23
|
Dracocephalum ruyschiana
|
вег
|
вег
|
вег
|
бут
|
| 24
|
Echium russicum
|
вег
|
вег
|
вег
|
бут
|
| 25
|
Elytrigia intermedia
|
вег
|
вег
|
вег
|
вег
|
вег
|
вег
|
аа
|
| 26
|
Erysimum canescens
|
вег
|
вег
|
бут
|
нцв
|
цв
|
| 27
|
Falcaria vulgaris
|
вег
|
вег
|
вег
|
вег
|
вег
|
вег
|
а
|
| 13
|
25
|
30
|
4
|
9
|
29
|
| 28
|
Festuca valesiaca
|
вег
|
вег
|
вег
|
труб
|
труб
|
бут
|
а
|
| 29
|
F
ritillaria ruthenica
|
бут
|
бут
|
бут
|
нцв
|
цв
|
| 30
|
Galium tinctorium
|
вег
|
вег
|
вег
|
вег
|
бут
|
а
|
| 31
|
Genista tinctoria
|
вег
|
вег
|
вег
|
вег
|
бут
|
| 32
|
Geranium sanguineum
|
вег
|
вег
|
вег
|
вег
|
нцв
|
а
|
| 33
|
Hyacinthella leucophaea
|
бут
|
нцв
|
цв
|
цв
|
кцв
|
пл
|
| 34
|
Helictotrichon pubescens
|
вег
|
вег
|
труб
|
нкол
|
| 35
|
Inula hirta
|
вег
|
вег
|
вег
|
вег
|
вег
|
бут
|
| 36
|
Iris aphylla
|
вег
|
вег
|
вег
|
вег
|
бут
|
цв
|
а
|
| 37
|
Jurinea arachnoidea
|
вег
|
бут
|
бут
|
бут
|
бут
|
цв
|
| 38
|
Knautia arvensis
|
вег
|
вег
|
вег
|
бут
|
| 39
|
Lathyrus lacteus
|
вег
|
бут
|
бут
|
нцв
|
цв
|
мцв
|
а
|
| 40
|
Leucanthemum vulgare
|
вег
|
вег
|
вег
|
вег
|
бут
|
а
|
| 41
|
Linum nervosum
|
вег
|
вег
|
вег
|
бут
|
| 42
|
Melampyrum cristatum
|
вег
|
вег
|
вег
|
бут
|
| 43
|
Myosotis popovii.
|
вег
|
вег
|
нцв
|
цв
|
мцв
|
| 44
|
Peucedanum oreoselinum
|
вег
|
вег
|
вег
|
вег
|
вег
|
вег
|
| 45
|
Plantago urvillei
|
вег
|
вег
|
бут
|
бут
|
бут
|
цв
|
| 46
|
Potentilla alba
|
бут
|
нцв
|
цв
|
мцв
|
цв
|
цв
|
| 47
|
Potentilla humifusa
|
нцв
|
цв
|
цв
|
цв
|
мцв
|
цв
|
| Таблица 7.2.1.1 (окончание)
|
| 1
|
2
|
3
|
4
|
5
|
6
|
7
|
8
|
9
|
| 48
|
Primula veris
|
бут
|
нцв
|
цв
|
цв
|
цв
|
пл
|
| 49
|
Pyrethrum corymbosum
|
вег
|
вег
|
вег
|
вег
|
вег
|
бут
|
а
|
| 50
|
Pulsatilla patens
|
цв
|
кцв
|
пл
|
пл
|
пл
|
кпл
|
аа
|
| 51
|
Ranunculus illyricus
|
вег
|
вег
|
бут
|
нцв
|
| 52
|
Ranunculus polyanthemos
|
вег
|
бут
|
бут
|
бут
|
нцв
|
цв
|
а
|
| 53
|
Rumex acetosa
|
вег
|
вег
|
вег
|
бут
|
цв
|
| 54
|
Rumex acetosella
|
вег
|
бут
|
бут
|
бут
|
| 55
|
Salvia pratensis
|
вег
|
вег
|
вег
|
нцв
|
аа
|
| 56
|
Scorzonera purpurea
|
вег
|
вег
|
вег
|
бут
|
цв
|
| 57
|
Senecio integrifolius
|
бут
|
бут
|
бут
|
мцв
|
| 58
|
Serratula lycopifolia
|
вег
|
вег
|
бут
|
бут
|
бут
|
| 13
|
25
|
30
|
4
|
9
|
29
|
| 59
|
Seseli annuum
|
вег
|
вег
|
вег
|
вег
|
вег
|
вег
|
| 60
|
Stachys recta
|
вег
|
вег
|
вег
|
бут
|
а
|
| 61
|
Stellaria graminea
|
вег
|
бут
|
нцв
|
цв
|
| 62
|
Stipa pennata
|
вег
|
вег
|
вег
|
вег
|
трубка
|
нкол
|
аа
|
| 63
|
Stipa tirsa
|
вег
|
вег
|
вег
|
вег
|
вег
|
вег
|
а
|
| 64
|
Tragopogon pratensis
|
вег
|
вег
|
вег
|
вег
|
бут
|
| 65
|
Trifolium alpestre
|
вег
|
вег
|
вег
|
бут
|
а
|
| 67
|
Trinia multicaulis
|
вег
|
вег
|
вег
|
бут
|
бут
|
цв
|
| 68
|
Trommsdorfia maculata
|
вег
|
вег
|
вег
|
вег
|
бут
|
| 69
|
Turritis glabra
|
вег
|
вег
|
бут
|
нцв
|
| 70
|
Valeriana rossica
|
вег
|
бут
|
бут
|
бут
|
бут
|
нцв
|
а
|
| 71
|
Veratrum nigrum
|
вег
|
вег
|
вег
|
вег
|
вег
|
вег
|
а
|
| 72
|
Veratrum album L. var.lobelianum
|
вег
|
вег
|
вег
|
вег
|
вег
|
вег
|
| 73
|
Verbascum phoenicium
|
вег
|
вег
|
бут
|
цв
|
| 74
|
Veronica jacquinii
|
вег
|
вег
|
бут
|
бут
|
| 75
|
Veronica prostrata
|
вег
|
бут
|
бут
|
мцв
|
| 76
|
Veronica verna
|
вег
|
бут
|
бут
|
нцв
|
цв
|
цв
|
| 77
|
Vicia tenuifolia
|
вег
|
вег
|
вег
|
цв
|
аа
|
| 78
|
Viola ambigua
|
цв
|
цв
|
цв
|
7.2.1.1.б.
Фенология видов травянистых растений Ямской степи на период с 6 июня по 13 августа 2004 года
Объектами фитофенологии были отдельные виды растений Ямской степи, встречающиеся на маршруте от ворот заповедного участка по степной дороге через большую ложбину к дальнему некосимому участку. Наблюдения осуществлялись фиксированием через 5 – 10 дней определенных фенофаз, по которым можно строить кривые вегетации, цветения и плодоношения. К сожалению во многие сроки наблюдений была дождливая погода или плоды были съедены вредителями, поэтому для некоторых видов не удалось зафиксировать все фенофазы. Принятые в таблице сокращения: вег – вегетация; бут – бутонизация; нцв – начало цветения; цв – цветение; мцв – массовое цветение; пл – плодоношение; выс – высыпание; труб – выход в трубку (у злаков); кол – колошение (у злаков); нкол – начало колошения; аа – образует ярко выраженный аспект; а – принимает заметное участие в аспекте.
Таблица 7.2.1.2
Фенология видов травянистых растений Ямской степи на период с 6 июня по 13 августа 2004 года
| №
|
Название видов растений
|
Сроки наблюдений
|
| июнь
|
июль
|
август
|
| 13
|
25
|
30
|
4
|
9
|
29
|
13
|
25
|
30
|
4
|
9
|
| 1
|
2
|
3
|
4
|
5
|
6
|
7
|
8
|
9
|
10
|
11
|
12
|
13
|
| 1
|
A. nobilis
|
бут
|
бут
|
бут
|
н.цв
|
цв
|
мцв
|
мцв
|
цв
|
цв
|
цв
|
цв
|
| 2
|
Achillea millefolium а
|
вег
|
вег
|
вег
|
вег
|
вег
|
бут
|
бут
|
нцв
|
мцв
|
мцв
|
мцв
|
| 3
|
Aconitum nemorosum
|
вег
|
вег
|
вег
|
вег
|
вег
|
вег
|
вег
|
бут
|
бут
|
нцв
|
цв
|
| 4
|
Adonisvernalis аа
|
пл
|
пл
|
кпл
|
вег
|
вег
|
вег
|
вег
|
вег
|
вег
|
вег
|
вег
|
| 5
|
Agrostis tenuis
|
вег
|
вег
|
труб
|
нкол
|
кол
|
мкол
|
кол
|
пл
|
пл
|
выс
|
выс
|
| 6
|
Allium oleraceum а
|
вег
|
вег
|
бут
|
бут
|
бут
|
бут
|
нцв
|
цв
|
мцв
|
цв
|
кцв
|
| 7
|
Allium rotundum
|
вег
|
вег
|
вег
|
бут
|
бут
|
нцв
|
нцв
|
цв
|
цв
|
| 8
|
Amoria montana аа
|
нцв
|
цв
|
цв
|
мцв
|
цв
|
кцв
|
пл
|
пл
|
кпл
|
кпл
|
кпл
|
| 9
|
Androsace septentrionalis
|
пл
|
пл
|
пл
|
пл
|
К.пл
|
вег
|
вег
|
вег
|
отм
|
| 10
|
Anthericum ramosum а
|
вег
|
вег
|
бут
|
нцв
|
цв
|
мцв
|
цв
|
пл
|
пл
|
пл
|
пл
|
| 11
|
Anthriscus sylvestris
|
нцв
|
цв
|
цв
|
кцв
|
пл
|
пл
|
пл
|
пл
|
пл
|
пл
|
пл
|
| 12
|
Arenaria micradenia
|
мцв
|
кцв
|
пл
|
пл
|
пл
|
пл
|
кпл
|
вег
|
вег
|
вег
|
2цв
|
| 13
|
Artemisia vulgaris
|
вег
|
вег
|
вег
|
вег
|
бут
|
бут
|
бут
|
бут
|
нцв
|
м.цв
|
цв
|
| 14
|
Asparagus officinalis
|
цв
|
цв
|
цв
|
нпл
|
пл
|
пл
|
пл
|
пл
|
пл
|
пл
|
пл
|
| 15
|
Asperula cynanchica а
|
бут
|
бут
|
нцв
|
цв
|
мцв
|
цв
|
цв
|
пл
|
пл
|
пл
|
| 16
|
Aster amellus
|
вег
|
вег
|
вег
|
вег
|
вег
|
вег
|
бут
|
бут
|
нц
|
мцв
|
мцв
|
| 17
|
Astragalus danicus
|
мцв
|
цв
|
кцв
|
пл
|
пл
|
пл
|
| 18
|
Bromopsis inermis аа
|
вег
|
вег
|
вег
|
труб
|
нкол
|
кол
|
кол
|
кол
|
пл
|
пл
|
пл
|
| 19
|
Bromopsis riparia береговой аа
|
мкол
|
кол
|
кол
|
пл
|
пл
|
пл
|
пл
|
пл
|
пл
|
выс
|
выс
|
| 20
|
Bupleurum falcatum
|
вег
|
вег
|
вег
|
нбут
|
бут
|
нцв
|
цв
|
цв
|
цв
|
цв
|
цв
|
| 21
|
Calamagrostis epigeios аа
|
вег
|
вег
|
вег
|
труб
|
кол
|
нкол
|
мкол
|
кол
|
кол
|
пл
|
пл
|
| Таблица 7.2.1.2 (продолжение)
|
| 1
|
2
|
3
|
4
|
5
|
6
|
7
|
8
|
9
|
10
|
11
|
12
|
13
|
| 22
|
Campanula altaica
|
мцв
|
цв
|
цв
|
кцв
|
пл
|
пл
|
пл
|
пл
|
кпл
|
вег
|
вег
|
| 23
|
Campanula bononiensis
|
вег
|
вег
|
вег
|
бут
|
бут
|
нцв
|
цв
|
мцв
|
цв
|
кцв
|
пл
|
| 24
|
Campanula glomerata
|
вег
|
вег
|
бут
|
бут
|
нцв
|
цв
|
цв
|
цв
|
цв
|
цв
|
цв
|
| 25
|
Campanula persicifolia
|
вег
|
бут
|
бут
|
нцв
|
цв
|
цв
|
кцв
|
пл
|
пл
|
пл
|
пл
|
| 26
|
Carduus hamulosus
|
вег
|
вег
|
бут
|
нцв
|
цв
|
цв
|
кцв
|
пл
|
пл
|
пл
|
пл
|
| 27
|
Centaurea jacea
|
вег
|
вег
|
вег
|
вег
|
бут
|
бут
|
бут
|
нцв
|
цв
|
цв
|
мцв
|
| 28
|
Centaurea marschalliana
|
цв
|
цв
|
цв
|
кцв
|
пл
|
пл
|
| 29
|
Centaurea pseudophrygia
|
вег
|
вег
|
вег
|
бут
|
бут
|
бут
|
нцв
|
нцв
|
цв
|
цв
|
мцв
|
| 30
|
Centaurea scabiosa
|
бут
|
бут
|
бут
|
бут
|
бут
|
нцв
|
мцв
|
цв
|
цв
|
кцв
|
пл
|
| 31
|
Chamaecytisus ruthenicus
|
кцв
|
пл
|
пл
|
пл
|
| 32
|
Cichorium intybus
|
вег
|
вег
|
вег
|
вег
|
бут
|
нцв
|
цв
|
цв
|
мцв
|
цв
|
кцв
|
| 33
|
Cirsium arvense
|
вег
|
вег
|
вег
|
вег
|
бут
|
бут
|
нцв
|
цв
|
цв
|
пл
|
пл
|
| 34
|
Clematis integrifolia
|
нцв
|
цв
|
цв
|
нпл
|
пл
|
пл
|
пл
|
пл
|
кпл
|
вег
|
вег
|
| 35
|
Clematis recta
|
цв
|
цв
|
кцв
|
пл
|
пл
|
пл
|
пл
|
пл
|
кпл
|
вег
|
вег
|
| 36
|
Dactylis glomerata
|
кол
|
кол
|
мкол
|
пл
|
пл
|
пл
|
пл
|
пл
|
| 37
|
Delphinium cuneatum
|
бут
|
бут
|
бут
|
нцв
|
мцв
|
мцв
|
мцв
|
кцв
|
пл
|
| 38
|
Dianthus andrzejowskianus
|
нцв
|
цв
|
цв
|
мцв
|
цв
|
нпл
|
пл
|
пл
|
кпл
|
кпл
|
вег
|
| 39
|
Draba sibirica
|
пл
|
кпл
|
кпл
|
вег
|
вег
|
вег
|
вег
|
вег
|
вег
|
вег
|
вег
|
| 40
|
Dracocephalum ruyschiana
|
нцв
|
цв
|
мцв
|
мцв
|
кцв
|
пл
|
пл
|
пл
|
кпл
|
вег
|
вег
|
| 41
|
Echium russicum
|
бут
|
нцв
|
цв
|
мцв
|
пл
|
пл
|
пл
|
пл
|
пл
|
| 42
|
Elytrigia intermedia
|
вег
|
вег
|
труб
|
нкол
|
кол
|
кол
|
мкол
|
мкол
|
кол
|
пл
|
пл
|
| 43
|
Erysimum canescens
|
цв
|
кцв
|
пл
|
пл
|
пл
|
пл
|
пл
|
пл
|
кпл
|
вег
|
вег
|
| 44
|
Erysimum hieracifolium
|
бут
|
бут
|
нцв
|
цв
|
цв
|
мцв
|
цв
|
кцв
|
пл
|
пл
|
пл
|
| 45
|
Falcaria vulgaris
|
вег
|
вег
|
вег
|
нбут
|
бут
|
нцв
|
цв
|
цв
|
мцв
|
мцв
|
Нпл
|
| 46
|
Festuca pratensis
|
вег
|
труб
|
кол
|
мкол
|
кол
|
кол
|
| 47
|
Filipendula vulgaris
|
бут
|
нцв
|
цв
|
мцв
|
кцв
|
пл
|
пл
|
пл
|
кпл
|
вег
|
вег
|
| 48
|
Fragaria viridis
|
мцв
|
пл
|
пл
|
пл
|
пл
|
кпл
|
вег
|
вег
|
| 49
|
Galatella linosyris
|
вег
|
вег
|
вег
|
вег
|
вег
|
вег
|
вег
|
бут
|
бут
|
нцв
|
цв
|
| 50
|
Galium boreale
|
бут
|
нцв
|
цв
|
цв
|
| 51
|
Galium tinctorium
|
бут
|
нцв
|
цв
|
мцв
|
кцв
|
пл
|
пл
|
пл
|
пл
|
пл
|
кпл
|
| 52
|
Galium verum
|
бут
|
бут
|
бут
|
нцв
|
цв
|
мцв
|
цв
|
цв
|
пл
|
пл
|
пл
|
| 53
|
Genista tinctoria
|
нцв
|
цв
|
цв
|
мцв
|
цв
|
цв
|
пл
|
пл
|
пл
|
| 54
|
Geranium sanguineum
|
бут
|
нцв
|
цв
|
цв
|
цв
|
цв
|
пл
|
пл
|
пл
|
пл
|
кпл
|
| 55
|
Gladiolus imbricatus а
|
бут
|
нцв
|
мцв
|
цв
|
нпл
|
пл
|
пл
|
пл
|
пл
|
выс
|
вег
|
| 56
|
Gymnadenia conopsea
|
цв
|
цв
|
кцв
|
пл
|
пл
|
пл
|
пл
|
| 57
|
Helichrysum arenarium
|
вег
|
вег
|
бут
|
бут
|
бут
|
нцв
|
цв
|
цв
|
мцв
|
цв
|
кцв
|
| Таблица 7.2.1.2 (продолжение)
|
| 1
|
2
|
3
|
4
|
5
|
6
|
7
|
8
|
9
|
10
|
11
|
12
|
13
|
| 58
|
Helictotrichon pubescens
|
кол
|
кол
|
кол
|
пл
|
пл
|
| 59
|
Hieracium umbellatum
|
вег
|
вег
|
вег
|
вег
|
вег
|
бут
|
бут
|
бут
|
нцв
|
цв
|
цв
|
| 60
|
Hieracium virosum
|
вег
|
вег
|
вег
|
вег
|
вег
|
бут
|
нцв
|
цв
|
цв
|
мцв
|
| 61
|
Hylotelephium ruprechtii
|
вег
|
вег
|
вег
|
вег
|
бут
|
бут
|
бут
|
бут
|
нцв
|
цв
|
мцв
|
| 62
|
Hypericum perforatum
|
бут
|
нцв
|
цв
|
цв
|
цв
|
цв
|
цв
|
пл
|
пл
|
| 63
|
Inula hirta
|
бут
|
бут
|
бут
|
нцв
|
цв
|
цв
|
цв
|
кцв
|
пл
|
пл
|
пл
|
| 64
|
Iris aphylla а
|
кцв
|
вег
|
вег
|
вег
|
вег
|
вег
|
вег
|
вег
|
вег
|
| 65
|
Jurinea arachnoidea
|
кцв
|
пл
|
пл
|
кпл
|
вег
|
вег
|
вег
|
вег
|
вег
|
вег
|
вег
|
| 66
|
Knautia arvensis
|
бут
|
нцв
|
цв
|
мцв
|
цв
|
пл
|
пл
|
пл
|
кпл
|
вег
|
вег
|
| 67
|
Lathyrus lacteus
|
цв
|
кцв
|
пл
|
пл
|
пл
|
пл
|
пл
|
пл
|
выс
|
вег
|
вег
|
| 68
|
Lathyrus pratensis
|
цв
|
цв
|
цв
|
цв
|
цв
|
цв
|
пл
|
| 69
|
Leontodon hispidus
|
цв
|
цв
|
мцв
|
мцв
|
цв
|
нпл
|
пл
|
пл
|
выс
|
вег
|
вег
|
| 70
|
Leucanthemum vulgare
|
бут
|
нцв
|
цв
|
мцв
|
мцв
|
цв
|
кцв
|
пл
|
кпл
|
выс
|
выс
|
| 71
|
Lilium pilosiusculum
|
пл
|
пл
|
пл
|
| 72
|
Linaria biebersteinii
|
бут
|
бут
|
нцв
|
цв
|
цв
|
цв
|
цв
|
цв
|
цв
|
| 73
|
Linum flavum
|
бут
|
бут
|
нцв
|
цв
|
м.цв
|
мцв
|
мцв
|
цв
|
цв
|
кцв
|
пл
|
| 74
|
Linum nervosum
|
нцв
|
цв
|
цв
|
мцв
|
цв
|
кцв
|
пл
|
кпл
|
кпл
|
вег
|
вег
|
| 75
|
Lotus corniculatus
|
цв
|
мцв
|
цв
|
цв
|
цв
|
цв
|
цв
|
цв
|
цв
|
| 76
|
Lythrum virgatum
|
вег
|
вег
|
вег
|
вег
|
вег
|
вег
|
бут
|
бут
|
бут
|
нцв
|
цв
|
| 77
|
Medicago falcata
|
нцв
|
цв
|
цв
|
цв
|
цв
|
мцв
|
цв
|
цв
|
| 78
|
Melampyrum argyrocomum
|
бут
|
нцв
|
цв
|
мцв
|
цв
|
цв
|
цв
|
цв
|
цв
|
цв
|
кцв
|
| 79
|
Melampyrum cristatum
|
нцв
|
цв
|
цв
|
мцв
|
цв
|
кцв
|
пл
|
пл
|
кпл
|
кпл
|
вег
|
| 80
|
Myosotis popovii
|
цв
|
цв
|
кцв
|
пл
|
пл
|
пл
|
| 81
|
Nepeta pannonica
|
вег
|
бут
|
бут
|
бут
|
нцв
|
цв
|
цв
|
кцв
|
пл
|
| 82
|
Odontites vulgaris
|
вег
|
вег
|
бут
|
бут
|
нцв
|
мцв
|
| 83
|
Onobrychis arenaria
|
нцв
|
цв
|
мцв
|
цв
|
цв
|
цв
|
цв
|
цв
|
цв
|
цв
|
| 84
|
Origanum vulgare
|
вег
|
вег
|
вег
|
бут
|
бут
|
нцв
|
цв
|
цв
|
мцв
|
цв
|
цв
|
| 85
|
Pedicularis kaufmannii
|
цв
|
кцв
|
пл
|
пл
|
пл
|
пл
|
кпл
|
кпл
|
кпл
|
вег
|
вег
|
| 86
|
Peucedanum oreoselinum
|
вег
|
вег
|
вег
|
нбут
|
бут
|
нцв
|
мцв
|
мцв
|
мцв
|
нпл
|
пл
|
| 87
|
Phleum phleoides
|
вег
|
вег
|
вег
|
вег
|
вег
|
вег
|
труб
|
нкол
|
| 88
|
Phlomoides tuberosa
|
бут
|
бут
|
нцв
|
мцв
|
цв
|
цв
|
кцв
|
нпл
|
пл
|
пл
|
| 89
|
Picris hieracioides
|
вег
|
вег
|
вег
|
вег
|
бут
|
бут
|
нцв
|
цв
|
цв
|
цв
|
мцв
|
| 90
|
Pimpinella saxifraga
|
вег
|
вег
|
вег
|
вег
|
вег
|
нбут
|
бут
|
бут
|
нцв
|
цв
|
цв
|
| 91
|
Plantago urvillei
|
мцв
|
мцв
|
мцв
|
цв
|
кцв
|
пл
|
пл
|
пл
|
пл
|
выс
|
2цв
|
| 92
|
Potentilla argentea
|
цв
|
мцв
|
цв
|
цв
|
цв
|
пл
|
пл
|
пл
|
пл
|
| 93
|
Primula veris
|
пл
|
пл
|
пл
|
пл
|
кпл
|
вег
|
вег
|
вег
|
вег
|
вег
|
вег
|
| Таблица 7.2.1.2 (продолжение)
|
| 1
|
2
|
3
|
4
|
5
|
6
|
7
|
8
|
9
|
10
|
11
|
12
|
13
|
| 94
|
Prunella grandiflora
|
вег
|
бут
|
бут
|
нцв
|
цв
|
цв
|
цв
|
цв
|
цв
|
пл
|
пл
|
| 95
|
Pyrethrum corymbosum
|
бут
|
бут
|
бут
|
нцв
|
цв
|
мцв
|
мцв
|
цв
|
пл
|
пл
|
кпл
|
| 96
|
Ranunculus illyricus
|
ьцв
|
кцв
|
нпл
|
пл
|
кпл
|
вег
|
вег
|
вег
|
отм
|
| 97
|
Ranunculus polyanthemos
|
мцв
|
цв
|
цв
|
цв
|
пл
|
пл
|
пл
|
кпл
|
кпл
|
кпл
|
2цв
|
| 98
|
Rhinanthus glacialis subsp.subalpinus
|
нцв
|
цв
|
цв
|
мцв
|
цв
|
цв
|
пл
|
пл
|
кпл
|
| 99
|
Rumex acetosa
|
цв
|
цв
|
кцв
|
нпл
|
пл
|
пл
|
пл
|
пл
|
кпл
|
вег
|
вег
|
| 100
|
Rumex acetosella
|
цв
|
цв
|
цв
|
цв
|
пл
|
пл
|
пл
|
пл
|
пл
|
| 101
|
Salvia pratensis
|
мцв
|
цв
|
цв
|
кцв
|
пл
|
пл
|
пл
|
пл
|
кпл
|
вег
|
2цв
|
| 102
|
Sanguisorba officinalis
|
вег
|
бут
|
бут
|
н.цв
|
цв
|
цв
|
цв
|
цв
|
мцв
|
мцв
|
| 103
|
Scabiosa ochroleuca
|
вег
|
бут
|
бут
|
бут
|
нцв
|
цв
|
цв
|
цв
|
цв
|
цв
|
цв
|
| 104
|
Scorzonera purpurea
|
мцв
|
цв
|
цв
|
кцв
|
нпл
|
пл
|
кпл
|
кпл
|
кпл
|
вег
|
вег
|
| 105
|
Securigera varia
|
нцв
|
мцв
|
цв
|
цв
|
цв
|
цв
|
пл
|
пл
|
| 106
|
Sedum acre
|
цв
|
мцв
|
к.цв
|
пл
|
пл
|
пл
|
пл
|
пл
|
пл
|
| 107
|
Senecio erucifolius
|
вег
|
вег
|
вег
|
вег
|
вег
|
вег
|
бут
|
бут
|
бут
|
бут
|
нцв
|
| 108
|
Senecio jacobaea
|
вег
|
вег
|
вег
|
вег
|
бут
|
нцв
|
цв
|
мцв
|
мцв
|
цв
|
цв
|
| 109
|
Serratula lycopifolia.
|
бут
|
бут
|
бут
|
нцв
|
цв
|
мцв
|
цв
|
пл
|
пл
|
пл
|
| 110
|
Serratula radiata
|
бут
|
бут
|
бут
|
нцв
|
цв
|
| 111
|
Serratula tinctoria
|
вег
|
вег
|
вег
|
бут
|
бут
|
бут
|
нцв
|
цв
|
цв
|
цв
|
мцв
|
| 112
|
Seseli annuum
|
вег
|
вег
|
вег
|
вег
|
вег
|
нбут
|
бут
|
бут
|
нцв
|
цв
|
мцв
|
| 13
|
25
|
30
|
4
|
9
|
29
|
13
|
25
|
30
|
4
|
9
|
| 113
|
Solidago virgaurea
|
вег
|
вег
|
вег
|
вег
|
вег
|
бут
|
нцв
|
цв
|
цв
|
цв
|
мцв
|
| 114
|
Stachys officinalis
|
вег
|
вег
|
бут
|
бут
|
бут
|
н.цв
|
мцв
|
цв
|
цв
|
кцв
|
пл
|
| 115
|
Stachys recta
|
бут
|
нцв
|
цв
|
мцв
|
цв
|
цв
|
кцв
|
пл
|
пл
|
кпл
|
| 116
|
Stellaria graminea
|
ьцв
|
цв
|
цв
|
цв
|
цв
|
цв
|
кцв
|
нпл
|
пл
|
пл
|
пл
|
| 117
|
Stipa pennata
|
кол
|
мкол
|
пл
|
пл
|
пл
|
кпл
|
кпл
|
вег
|
вег
|
вег
|
вег
|
| 118
|
Stipa tirsa
|
вег
|
вег
|
труб
|
нкол
|
кол
|
кол
|
нпл
|
пл
|
пл
|
| 119
|
Tanacetum vulgare
|
вег
|
вег
|
вег
|
вег
|
бут
|
бут
|
нцв
|
цв
|
мцв
|
цв
|
кцв
|
| 120
|
Thymus marschallianus
|
бут
|
нцв
|
мцв
|
цв
|
цв
|
цв
|
пл
|
пл
|
пл
|
пл
|
| 121
|
Tragopogon pratensis
|
нцв
|
цв
|
цв
|
нпл
|
пл
|
пл
|
кпл
|
вег
|
вег
|
вег
|
вег
|
| 122
|
Trifolium medium
|
вег
|
вег
|
бут
|
бут
|
нцв
|
цв
|
цв
|
цв
|
мцв
|
цв
|
цв
|
| 123
|
Trifolium alpestre
|
нцв
|
цв
|
цв
|
мцв
|
мцв
|
цв
|
цв
|
кцв
|
пл
|
пл
|
| 124
|
Trommsdorfia maculata
|
бут
|
бут
|
нцв
|
мцв
|
цв
|
кцв
|
пл
|
кпл
|
кпл
|
вег
|
вег
|
| 125
|
Turritis glabra
|
цв
|
кцв
|
нпл
|
пл
|
пл
|
пл
|
пл
|
пл
|
пл
|
кпл
|
кпл
|
| 126
|
Veratrum nigrum а
|
вег
|
вег
|
бут
|
бут
|
бут
|
бут
|
нцв
|
цв
|
мцв
|
нпл
|
пл
|
| 127
|
Verbascum lychnitis
|
бут
|
бут
|
бут
|
бут
|
нцв
|
цв
|
мцв
|
цв
|
цв
|
кцв
|
пл
|
| 128
|
Verbascum phoenicium
|
мцв
|
цв
|
цв
|
кцв
|
нпл
|
пл
|
пл
|
| Таблица 7.2.1.2 (окончание)
|
| 1
|
2
|
3
|
4
|
5
|
6
|
7
|
8
|
9
|
10
|
11
|
12
|
13
|
| 129
|
Veronica incana
|
вег
|
вег
|
вег
|
бут
|
бут
|
нцв
|
цв
|
цв
|
цв
|
цв
|
цв
|
| 130
|
Veronica jacquinii
|
нцв
|
цв
|
цв
|
кцв
|
пл
|
пл
|
пл
|
пл
|
пл
|
пл
|
пл
|
| 131
|
Veronica longifolia
|
вег
|
вег
|
бут
|
бут
|
нцв
|
цв
|
цв
|
цв
|
пл
|
пл
|
пл
|
| 132
|
Veronica spicata
|
вег
|
бут
|
бут
|
нцв
|
цв
|
мцв
|
цв
|
цв
|
пл
|
пл
|
пл
|
| 133
|
Vicia tenuifolia
|
мцв
|
мцв
|
мцв
|
кцв
|
пл
|
пл
|
пл
|
пл
|
пл
|
нвыс
|
нвыс
|
| 134
|
Vincetoxicum hirundinaria
|
цв
|
мцв
|
цв
|
цв
|
цв
|
цв
|
цв
|
пл
|
пл
|
| 135
|
Xanthoselinum alsaticum
|
вег
|
вег
|
вег
|
вег
|
вег
|
нбут
|
бут
|
бут
|
нцв
|
цв
|
мцв
|
7.2.1.2. Продуктивность надземной массы травостоя участка «Ямская степь»
Методика исследования
Продуктивность степной растительности определялась методом взятия укосов (Семёнова-Тянь-Шанская и др., 1960; Собакинских, 1996).
Работа выполнялась в первую неделю июля 2004 года. На плакоре было заложено 6 пробных площадей (ПП) (10м х 10м) – 4 в косимом режиме (ПП - №№ 1, 2, 3, 4) и 2 в некосимом (ПП-№ 5 и 6), по периметру которых на площадках 0,25 м2
осуществлялось взятие укосов в пятикратной повторности. Травостой срезали большими ножницами на уровне земли с выстриганием всех плотных дерновин. Отчужденный травостой и подстилка (вместе с живым мхом) собирались в отдельные пакеты, снабженные соответствующей этикеткой. К подстилке относили лежащие на земле сухие части растений, потерявшие связь с материнскими экземплярами. К ветоши – прошлогодние, не потерявшие связи с материнскими растениями.
В лабораторных условиях (в тот же день на кордоне) зеленую массу разбирали по видам и хозяйственно-ботаническим группам (злаки, осоки, бобовые, разнотравье, мхи). Разобранный материал упаковывали в бумажные пакетики, подписывали и высушивали в естественных условиях до воздушно-сухого состояния. После высушивания все пробы взвешивали на весах с ценой деления 0,1 г, полученные данные заносили в таблицы. В таблицах виды группировались по семействам и хозяйственно-ботаническим группам. Масса изначально устанавливалась в г/0,25м2
(см. табл. 7.2.1.5, 7.2.1.7–7.2.1.9). Для определения урожайности эти показатели затем переводились в ц/га (см. табл. 7.2.1.3, 7.2.1.4). Доля семейств и хозяйственно-ботанических групп в укосах определялась в процентах от всей надземной зеленой фитомассы (Х). Доля Х и мёртвой массы (подстилка и ветошь) (Т) определялась в процентах от общей надземной массы (М).
Обзор литературы
На территории Губкинского района Белгородской области расположен один из шести уцелевших в мире участков плакорной луговой степи – Ямская степь, входящая в состав заповедника «Белогорье». Для сохранения видового разнообразия растений здесь соблюдается несколько режимов охраны: сенокосооборотный (косимый не ежегодно), ежегоднокосимый и абсолютно-заповедный (некосимый).
Травянистые группировки плакорных пространств комплексные. Микрорельеф водоразделов в заповеднике отличается хорошо выраженным чередованием различной величины бугорков, западинок между бугорками, постепенно образующихся благодаря неравномерному распределению осадков и пониженных седловинок, соединяющих бугорки с западинками. К растениям, имеющим максимум обилия на повышениях относятся – типчак, тимьян Маршалла, частично тонконог и др.
К растениям, имеющим максимум обилия в микрозападинах, относятся: звездчатка злаковидная, буквица лекарственная, трясунка средняя, девясил британский, лютик многоцветковый, валериана русская и др.
Не менее отчетливо выражена комплексность травянистых группировок заповедника в связи с сезонами года. Разность их строения и сложения по сезонам проявляется исключительно резко. Описывая одну и ту же площадь весной, летом и осенью, исследователь придет к совершенно разным выводам.
По инициативе В.В.Алехина уже в первые годы заповедования (с 1935 г.) в Ямской степи были выделены некосимые участки для изучения процессов развития целинной растительности при полном ее изъятии из хозяйственного использования.
Прекращение сенокоса в первый же год ведет к накоплению значительной массы отмерших растений, способствующих зимой снегонакоплению, более медленному таянию весеннего снега и ослабленному испарению влаги в летний период. Мезофитизация некосимых участков связана с изменением режима влажности почв.
Изменение растительного покрова некосимых участков выражается в следующем:
1. Резко возрастает количество корневищных злаков. В первые годы большое развитие получает костер прямой (кострец береговой), но затем он замещается более мощным и влаголюбивым костром безостым (кострецом безостым). Самые характерные виды – костер безостый и вейник наземный.
2. Резко уменьшается как количество видов, так и роль степных видов ковыльниковой свиты. Исключение составляют ковыли, которые могут долго сохраняться на некосимых участках и создавать аспект. Но при нарастании мертвого покрова постепенно разрыхляются и дернины ковылей. Типчак на некосимых участках очень быстро исчезает.
3. Исходные, косимые группировки полностью теряют свою красочность и аспективность. Некосимые участки из красочных имеют только хорошо выраженный аспект ракитника и на отдельных площадях перистых ковылей. Основной фон некосимых площадей – злаковый. Разнотравье в числе резко уменьшается
4. На некосимой степи чрезвычайно характерно сильное развитие травянистых лесных свит, особенно боровой свиты. Эти виды мощно разрастаются и жизненность их здесь намного превышает ее на косимых площадях.
5. На больших площадях некосимые участки покрываются зарослями ракитника русского и превращаются в «кустарниковые группировки».
6. Исходная степь в результате этих изменений буквально перерождается.
Остепнение «луговых степей» - результат вмешательства человека в естественные процессы взаимоотношений растительных типов. Быстрое перерождение «степных» целин при прекращении хозяйственного воздействия человека – одно из ярких доказательств их нестойкости, «вторичности» как растительной группировки (Зозулин, 1955).
На некосимых участках в течение ряда лет накапливаются нескашиваемые растительные остатки – ветошь, значительно изменившая их экологический режим и растительность.
Значение ветоши как экологического фактора определяется следующим:
- Ветошь задерживает влагу.
- Создается рыхлый увлажненный «субстрат», благоприятствующий внедрению и дальнейшему развитию корневищных растений.
- Увеличивается мощность подстилки и соответственно лучше удобряется почва.
- Происходит изоляция минерального субстрата, в результате чего корешки проростков повисают в толще ветоши и погибают.
- Изменяется режим увлажнения: а) позже начинается потребление почвенной влаги, приводящее соответственно к более позднему по сравнению с косимой степью развитию растительности некосимой степи; б) ветошь, насыщающаяся атмосферной влагой, задерживает ее просачивание в почву; в) потребление влаги на косимой степи после отчуждения растительности и до отрастания отавы замедляется в результате уменьшения транспирации, а на некосимой степи продолжается интенсивная транспирация до конца периода вегетации, иссушая нижние горизонты почвы (Дохман, 1966).
Многолетние наблюдения за состоянием травяных экосистем при различных режимах охраны позволили установить отличительные особенности их влияния на различные компоненты экосистем.
Исследования термического режим черноземов свидетельствуют о больших его различиях, обусловленных различными режимами охраны. Наличие мощной подстилки на абсолютно-заповедных участках тормозит весенний прогрев почвы и понижает ее общий температурный фон в течение всего вегетационного периода. Переход температуры почвы на глубине 20 см через +5, +10, +15о
С здесь происходит на 1 – 3 недели позже сроков, характерных для косимой степи. Примерно на две недели задерживается и прохождение фаз вегетации растений на участках абсолютно-заповедной степи по сравнению с таковыми на участках косимой степи. В условиях абсолютно-заповедного режима хорошее развитие получает первоцвет весенний, ковыль перистый, подмаренник настоящий. На сенокосооборотных участках – костер береговой, ковыль перистый, эспарцет песчаный, шалфей луговой. На постоянно косимых участках по сравнению с сенокосооборотным заметно сокращается число цветущих побегов у первоцвета весеннего, овсеца опушенного, а ковыль перистый, горицвет весенний не образуют аспекта.
Основной причиной изменения лугово-степных фитоценозов при абсолютно-заповедном режиме является накопление мертвых растительных остатков (подстилки, ветоши), масса которой в период максимального развития растительности примерно равна зеленой части и составляет 3,42 т/га. Общая фитомасса растительности на абсолютно-заповедных участках степи не превышает 6,83 т/га. Растительные остатки на участках степи с косимым, сенокосооборотным режимом не превышают 45,2 и 29,1% общей фитомассы травянистой растительности. Ветошь и подстилка значительно изменяют водно-термические условия и обуславливают мезофитизацию растительности, способствуют развитию корневищных злаков: мятлика узколистного, костра безостого, вейника наземного, пырея и др. Кроме того, здесь происходит резкое снижение участия в составе растительного покрова мхов и степных видов двудольных растений (Краснитский , 1984).
Для поддержания общего разнообразия степей участка необходимы соблюдение и оптимизация практиковавшихся с 1936 г. дифференцированных режимов, прежде всего в плакорных условиях. Сочетание различных режимов в степях Ямского участка ("абсолютно заповедный", ежегодно косимый, сенокосооборотный, пастбищный) обеспечивало долговременную сохранность высокого разнообразия флоры, особенно степных видов. Фактическое прекращение выкашивания в последние годы на значительных площадях может привести к постепенному обеднению степного разнообразия. В качестве некоторой альтернативы отчуждения фитомассы за счёт выкашивания и предотвращения значительного накопления ветоши следует проработать вопрос об умеренном контролируемом выпасе (на части территории) лошадей или крупного рогатого скота (Золотухин, 2004).
Степные экосистемы, исторически формировавшиеся в условиях постоянно действующих экзогенных факторов, при снятии или замене этих факторов оказываются очень неустойчивыми и подверженными весьма существенным трансформациям. Их изменение по существу ведет к полной деструкции степного биома в целом и отдельных степных экосистем (Дидух, 1993).
Надземная фитомасса Ямской степи в местах взятия укосов (данные на июль
2004 г.)
Косимые и некосимые травостои значительно отличаются по количеству общей
надземной массы (М). На некосимых площадях она почти в 2 раза больше и составляет 164,45 ц/га (табл.7.2.1.3).
При косимом режиме в надземной массе преобладает зеленая фитомасса (Х), которая составляет 50,01 ц/га (67,13% от М), при некосимом, наоборот, мертвая масса (Т) – 92.63 ц/га (56,33% от М). Эта закономерность подтверждается литературными данными (Семёнова-Тяньшанская и др.,1960).
Наибольшее участие в структуре укосов травостоя принимают злаки. Их продуктивность составляет в среднем 24,28 ц/га (44,95% от Х) при косимом режиме и 52,16 ц/га (72,63%) при некосимом, т.е. урожайность злаков на некосимых участках в 2,15 раза больше.
Продуктивность бобовых выше на косимых площадях, где она составляет 8,60 ц/га (15,92% от Х), на некосимых участках эти показатели значительно ниже (2,51 ц/га или 3,49%). Основная масса в этой хозяйственно-ботанической группе при всех режимах охраны приходится на горошек тонколистный Vicia tenuifolia.
Продуктивность разнотравья составляет 14,65 ц/га (27,12% от Х) на косимых площадях и 16,95 (23,66% от Х) на некосимых. Здесь наблюдается некоторое противоречие с литературными данными, которое объясняется присутствием на некосимых участках Cirsium setosum. Этот вид явно предпочитает режим абсолютного заповедования при полном отсутствии на косимых площадях и вносит существенный вклад в продуктивность разнотравья – в среднем 31,70% от массы этой хозяйственно-ботанической группы растений.
Таблица 7.2.1.3.
Средняя надземная фитомасса Ямской степи в ц/га
(возд.сух.вес) на косимых и некосимых пробных площадях (2004г.)
| Режим
охраны
|
Хозяйственно-ботанические группы
|
| Надзем
ная ма- сса, М
|
Зеленая масса,
Х
|
В том числе
|
Мертвая
масса, Т
(подстилка)
|
| злаки
|
бобовые
|
разнотравье
|
| ц/га
|
ц/га
|
%
|
ц/га
|
%
|
ц/га
|
%
|
ц/га
|
%
|
ц/га
|
%
|
| Косимый
|
80.46
|
54.01
|
67.13
|
24.28
|
44.95
|
8.60
|
15.92
|
14.65
|
27.12
|
26.45
|
32.87
|
| Некосимый
|
164.45
|
71.82
|
43.67
|
52.16
|
72.63
|
2.51
|
3.49
|
16.95
|
23.60
|
92.63
|
56.33
|
Таблица 7.2.1.4
Надземная фитомасса Ямской степи на заложенных пробных площадях в ц/га (возд.сух.вес)
по семействам и хозяйственно-ботаническим группам (средние значения) (2004г.)
| Название семейств и хозяйственно-ботанических групп
|
Надземная фитомасса в ц/га
|
| ПП №1*
|
ПП №2
|
ПП №3
|
ПП №4
|
ПП №5
|
ПП №6
|
| 1
|
2
|
3
|
4
|
5
|
6
|
7
|
| Надземная масса
|
52.6
|
73.7
|
101.78
|
93.78
|
156.45
|
172.45
|
| Подстилка
|
11.22
|
44.12
|
37.2
|
13.28
|
93.57
|
91.70
|
| Мох
|
9.29
|
0.98
|
12.26
|
3.03
|
0
|
0.36
|
| Зеленая масса
|
32.09
|
28.60
|
52.32
|
77.47
|
62.88
|
80.39
|
| Злаки
|
24.23
|
14.69
|
20.78
|
37.44
|
47.31
|
57.01
|
| Осоки
|
0.06
|
0.24
|
0.02
|
0
|
0
|
0.02
|
| Бобовые
|
2.10
|
2.16
|
7.66
|
22.50
|
1.01
|
4.02
|
| Разнотравье
:
|
5.70
|
11.51
|
23.86
|
17.53
|
14.56
|
19.34
|
| Alliaceae
|
0.04
|
0.33
|
0.21
|
0.12
|
0
|
0.13
|
| Apiaceae
|
0
|
0.12
|
0.91
|
3.8
|
0.39
|
0.17
|
| Asclepiadaceae
|
0.08
|
0.01
|
0
|
0
|
0.06
|
0
|
| Asteraceae:
|
0.57
|
0.58
|
1.95
|
1.64
|
1.58
|
9.66
|
| В т.ч. Cirsium setosum
|
0
|
0
|
0
|
0
|
1.44
|
9.30
|
| Asparagaceae
|
0
|
0
|
0
|
0
|
1.57
|
0
|
| Boraginaceae
|
0
|
0
|
0
|
0.01
|
0
|
0
|
| Таблица 7.2.1.4 (окончание)
|
| 1
|
2
|
3
|
4
|
5
|
6
|
7
|
| Brassicaceae
|
0.05
|
0.69
|
0.50
|
2.08
|
0
|
0.17
|
| Campanulaceae
|
0.01
|
0
|
0.01
|
0.31
|
0
|
0
|
| Caryophyllaceae
|
0.03
|
0.10
|
0
|
0.26
|
0
|
0
|
| Convolvulaceae
|
0.2
|
0
|
0.1
|
0
|
0
|
0
|
| Dipsacaceae
|
0
|
0.05
|
0.02
|
0
|
0
|
0
|
| Euphorbiaceae
|
0
|
0.04
|
0
|
0.01
|
0.04
|
0
|
| Geraniaceae
|
0
|
2.68
|
6.62
|
0
|
0
|
0
|
| Hypericaceae
|
0
|
0.02
|
0
|
0
|
0
|
0
|
| Iridaceae
|
0
|
0.62
|
0
|
0
|
0.75
|
2.21
|
| Lamiaceae
|
1.39
|
1.39
|
2.74
|
2.34
|
0.82
|
0.30
|
| Liliaceae
|
0
|
0
|
0
|
0.01
|
0.02
|
0.89
|
| Linaceae
|
0
|
0
|
0
|
0
|
0.03
|
0
|
| Plantaginaceae
|
0.10
|
0.01
|
0
|
0
|
0
|
0
|
| Polygonaceae
|
0
|
0
|
0.04
|
0
|
0
|
0
|
| Primulaceae
|
0
|
0
|
0.30
|
0
|
0
|
0
|
| Ranunculaceae
|
0.50
|
0
|
0.76
|
0.72
|
0.22
|
0
|
| Rosaceae :
|
1.71
|
4.01
|
6.25
|
1.86
|
6.83
|
3.66
|
| В т.ч. Filipendula vulg.
|
1.51
|
3.97
|
4.14
|
1.33
|
6.83
|
2.69
|
| Rubiaceae
|
0.68
|
0.68
|
3.33
|
2.98
|
0.93
|
2.17
|
| Scrophulariaceae
|
0.30
|
0.19
|
0.10
|
0.82
|
1.30
|
0
|
| Valerianaceae
|
0
|
0
|
0.02
|
0.31
|
0
|
0
|
| Violaceae
|
0
|
0.01
|
0.01
|
0
|
0.01
|
0
|
*
Расположение ПП: №1
– плакор напротив аншлага у старого кордона, косимый режим – 2002 – 2003 гг. выкашивалась; №2
– плакор на феномаршруте, напротив западной границы дальнего некосимого участка, косимый режим – 2002 – 2003 гг. не выкашивалась; №3
– плакор на экологическом профиле (б.Суры) рядом с точкой Щ 19/20 (сетка Б.К. Ганнибала, 2002 г., см. Летопись природы-2003г.), косимый режим – в 2002 г. выкашивалась, в 2003 г. не выкашивалась; №4
– плакор на экологическом профиле (б.Суры) рядом с точкой Щ 23/24, косимый режим – в 2002 – 2003 гг. выкашивалась; №5
– дальний некосимый участок на феномаршруте напротив №2; №6
– ближний некосимый участок.
Среди разнотравья по массе в косимой и некосимой степи выделяются семейства Розоцветных (в среднем 3,46 ц/га на косимых площадях и 5,24 ц/га на некосимых), Сложноцветных (1,18 и 5.62 ц/га), Мареновых (1,91 и 1,55 ц/га), Губоцветных (1,96 и 0,56 ц/га), виды которых присутствуют во всех укосах. Хочется отметить значительное участие в структуре укосов таволги шестилепестной Filipendula vulgris (в среднем 2,74 ц/га на косимых площадях и 4,76 ц/га на некосимых), подмаренников настоящего Gallium verum и красильного Galium tinctorium.
Небольшую долю в укосах в 2004 г. занимали осоки. Осоки составляют в среднем на косимых участках 0,08 ц/га, на некосимых – 0,01ц/га (табл. 7.2.1.4).
Мхи в большем количестве отмечены именно на косимых участках, особенно на тех, которые выкашивались в последние годы, в среднем 6,39 ц/га (табл. 7.2.1.4). При нарушении режима сенокошения их участие в структуре укосов существенно снижается. На некосимых участках масса мхов составляет 0,18 ц/га (табл. 7.2.1.4).
По неясной для нас причине при разборе укосов не удалось выделить ветошь, хотя зеленая часть и подстилка собирались в отдельные пакеты. Можно предположить, что из-за погодных условий отмирание надземной массы на период взятия укосов было незначительным, а прошлогодние сухие части уже потеряли связь с материнскими растениями и были собраны в подстилке.
Продуктивность степной растительности
при разных режимах охраны (на примере трех пробных площадей)
Более наглядно изменения, происходящие в степи при прекращении сенокошения, характеризуют количественные данные 2004г., полученные при учете продуктивности методом взятия укосов на плакорных участках с разными режимами охраны.
При прекращении сенокошения происходит увеличение надземной массы . На площади
возле аншлага, выкашиваемой последние четыре года (далее площадь №1 или первая), она
составляет 131,54 г/0,25м2
или 52,6 ц/га (табл. 7.2.1.4). На площади возле дальнего некосимого
участка, не выкашиваемой последние четыре года (далее – площадь №2 или вторая) она
составляет уже 184,26 г/0,25м2
или 73.7 ц/га (табл. 7.2.1.4), а на дальнем некосимом участке
(далее – площадь №3 или третья, или площадь №5 в табл.7.2.1.4) 391,12 г/0,25м2
(156,45 ц/га),
т.е. почти в три раза больше, чем на первой площади (табл. 7.2.1.5). Но если на первой площади большую долю от всей надземной массы составляет зелёная масса – 78.69% или
103.48 г / 0,25 м2
, то при прекращении сенокошения увеличивается доля отмерших остатков
(подстилки) 59,86 % или 110,30 г / 0,25м2
на второй площади и 59,81 % или 233,92 г /0,25 м2
на третьей (рис. 7.2.1.1).
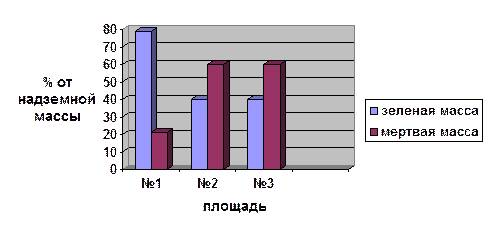
Рис. 7.2.1.1. Количественное соотношение зеленой (Х) и мертвой (Т) масс на площадях
с разными режимами охраны.
Наибольшее участие в структуре укосов на всех площадях принимают злаки. Их продуктивность составляет в среднем 60,58 г/0,25 м2
(58,54 % от Х) на первой площади при косимом режиме; 36,72 г/0,25 м2
(49,65 %) на второй площади при нарушении режима и 118,28 г/0,25 м2
(75,24 %) на некосимой площади. Таким образом, на некосимой площади на долю злаков приходится большая часть зеленой массы.
Продуктивность бобовых выше на косимых площадях (площадь №1 – 5,26 г/0,25м2
или 5,08 % от Х), даже при недлительном нарушении сенокошения (площадь №2) их доля существенно не снижается и составляет 5,40 г/0,25м2
или 7,30 % от Х. На некосимом участке этот показатель значительно ниже (2,52 г/0,25м2
или 1,60 %). Основная масса в этой хозяйственно-ботанической группе при всех режимах охраны приходится на горошек тонколистный Vicia tenuifolia (3.72 г/0,25м2
; 2.96 г/0,25м2
; 1.1 г/0,25м2
соответственно).
Таблица 7.2.1.5
Средняя надземная фитомасса Ямской степи в г/0,25 м2
на косимых и некосимых ПП
| Режим
охраны
|
Хозяйственно-ботанические группы
|
| Надзем-
ная мас- са, М
|
Зеленая масса,
Х
|
В том числе
|
Мертвая
масса,
Т
|
| злаки
|
бобовые
|
разнотравье
|
| г/0,25 м2
|
г/0,25 м2
|
% от надз.
|
г/0,25 м2
|
% от зел.
|
г/0,25 м2
|
%от зел.
|
г/0,25 м2
|
% от зел.
|
г/0,25 м2
|
% от надз.
|
| Коси-
мый №1
|
131.54
|
103.48
|
78.66
|
60.58
|
58.54
|
5.26
|
5.08
|
14.26
|
13.78
|
28.06
|
21.33
|
| Коси-
мый №2
|
184.26
|
73..96
|
40.14
|
36.72
|
49.65
|
5.40
|
7.30
|
28.78
|
38.91
|
110.30
|
59.86
|
| Некоси-
мый
№3
|
391.12
|
157.2
|
40.19
|
118.28
|
75.24
|
2.52
|
1.60
|
36.4
|
23.15
|
233.92
|
59.81
|
Продуктивность разнотравья составляет 14,26 г/0,25м2
(13,78 % от Х) на первой площади и 28,78 г/0,25м2
(38,91 % от Х) на второй, на третьей площади в некосимом режиме 36,4 г/0,25м2
или 23,15 %. На дальнем некосимом участке обилен бодяк полевой Cirsium setosum. Этот вид явно предпочитает режим абсолютного заповедования при полном отсутствии на косимых площадях, и даже единичные экземпляры вносят существенный вклад в продуктивность разнотравья – примерно 10 % от массы этой хозяйственно-ботанической группы растений.
Разнотравье на первой площади представлено видами 13 семейств, на второй площади – 17, на третьей – видами из 14 семейств. Среди этой хозяйственно-ботанической группы на первой площади в косимом режиме преобладают по массе семейства: Губоцветные Lamiaceae – 1,39 ц/га (3,48 г/0,25м2
), Розоцветные Rosaceae – 1,71 ц/га (4,28 г/0,25м2
), на второй площади Гераневые Geraniaceae – 2,68 ц/га (6,7 г/0,25м2
), Губоцветные Lamiaceae – 1,39 ц/га (3,48 г/0,25м2
) и Розоцветные Rosaceae – 4,01 ц/га (10,02 г/0,25м2
); на третьей площади в некосимом режиме Норичниковые Scrophulariaceae – 1,30 ц/га (3,24 г/0,25м2
), Розоцветные Rosaceae 6,83 ц/га (17,08 г/0,25м2
), Сложноцветные Asteraceae – 1,58 ц/га (3,46 г/0,25м2
), Спаржевые Asparagaceae – 1,57 ц/га (3,92 г/0,25м2
), виды которых присутствуют практически во всех укосах (табл.7.2.1.8 и 7.2.1.9).
Необходимо отметить значительное участие в структуре укосов лугового вида, представителя семейства Розоцветных - таволги шестилепестной Filipendula vulgris (см. табл. 7.2.1.4). Продуктивность этого вида нарастает при переходе от косимого режима к
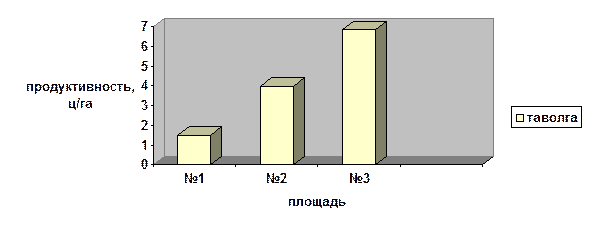
Рис.7.2.1.2. Продуктивность Filipendula vulgris на площадях с разными режимами охраны
некосимому и составляет в среднем 1,51 ц/га (3,78 г/0,25м2
) на первой площади, 3,97 ц/га (9,92 г/0,25м2
) на второй и 6,83 ц/га на третьей (17,08 г/0,25м2
).
На втором месте по обилию и продуктивности находятся подмаренники настоящий Gallium verum и красильный Galium tinctorium. Эти виды не делают различия между режимами, и нарушение режима сенокошения особого влияния на обилие этих видов не оказывает. Продуктивность составляет 1,64 г/0,25м2
на первой площади; 1,68 г/0,25м2
на второй и 2,32 г/0,25м2
на третьей.
Небольшую долю в укосах на всех площадях занимали осоки. Их продуктивность составляла всего 0,16 и 0,6 г/0,25м2
на первой и второй площадях (табл. 7.2.1.4, ПП №1 и 2). На третьей площади (дальний некосимый участок, ПП №5) осока в укосах не отмечена.
Мхи в большем количестве встречаются именно на первом участке (23,22 г/0,25м2
или 22,44% от Х), ежегодное сенокошение способствует их развитию и увеличению обилия. При нарушении режима сенокошения участие мхов в структуре укосов существенно снижается, например, на площади №2 составляет уже только 2,46 г/0,25м2
или 3,33 % от Х, а на некосимом участке мох в укосах отсутствует.
Сведения об обилии видов на ПП (10х10м) в местах взятия укосов, их жизненной форме и принадлежности к фитоценотической группе приведены в таблице 7.2.1.6.
Таблица 7.2.1.6
Характеристика видового состава сосудистых растений пробных площадей 100 м2
в местах взятия укосов на июль 2004 г
| Названия видов
|
Бм1
|
Эф2
|
Обилие видов по Друде на П П в %
|
| №1*
|
№2
|
№3
|
№4
|
№5
|
№6
|
| |
Achillea millefolium
|
п
|
лг
|
sol
|
-
|
sp
|
sp
|
sol
|
-
|
| |
Adonis vernalis
|
п
|
с
|
-
|
-
|
-
|
-
|
sol
|
-
|
| |
Allium oleraceum
|
п
|
л
|
sol
|
sol
|
sol
|
sol
|
-
|
sol
|
| |
Amoria montana
|
п
|
с
|
sp
|
sp
|
-
|
-
|
-
|
-
|
| |
Anthericum ramosum
|
п
|
с
|
-
|
-
|
-
|
sol
|
sol
|
-
|
| |
Asparagus officinalis
|
п
|
лг
|
-
|
-
|
-
|
-
|
sol
|
-
|
| |
Astragalus danicus
|
п
|
лг
|
sol
|
-
|
-
|
-
|
-
|
-
|
| |
Bromopsis inermis
|
п
|
лг
|
-
|
-
|
-
|
sol
|
sol
|
sol
|
| |
Bromopsis riparia
|
п
|
с
|
cop1
|
Cop1
|
Cop1
|
Cop1
|
sol
|
-
|
| |
Calamagrostis epigeios
|
п
|
лг
|
sp
|
-
|
-
|
-
|
sp
|
Cop1
|
| |
Campanula altaica
|
п
|
с
|
sol
|
-
|
sol
|
sol
|
-
|
-
|
| |
Carex michelii
|
п
|
лг
|
-
|
-
|
sol
|
-
|
-
|
-
|
| |
Carex sp
|
п
|
с
|
sol
|
sol
|
sol
|
-
|
-
|
sol
|
| |
Centaurea jacea
|
п
|
лг
|
-
|
-
|
sol
|
-
|
-
|
-
|
| |
Centaurea scabiosa
|
п
|
лг
|
-
|
-
|
sol
|
-
|
-
|
-
|
| |
Centaurea sumensis
|
п
|
с
|
-
|
sp
|
-
|
-
|
-
|
-
|
| |
Chamaecytisus ruthenicus
|
пк
|
с
|
sol
|
sol
|
sol
|
-
|
sol
|
-
|
| |
Cirsium setosum
|
п
|
лг
|
-
|
-
|
-
|
-
|
sol
|
sol
|
| |
Clematis integrifolia
|
п
|
с
|
-
|
-
|
sol
|
sol
|
-
|
-
|
| |
Convolvulus arvensis
|
п
|
с
|
sol
|
-
|
sol
|
-
|
-
|
-
|
| |
Dianthus andrzejowskianus
|
п
|
с
|
sol
|
sol
|
-
|
-
|
-
|
-
|
| |
Draba sibirica
|
п
|
с
|
sp
|
Cop1
|
Cop1
|
Cop1
|
-
|
sol
|
| |
Dracocephalum ruyschiana
|
п
|
с
|
-
|
sp
|
sol
|
-
|
-
|
-
|
| |
Elytrigia intermedia
|
п
|
с
|
sp
|
Cop1
|
sp
|
Cop1
|
Cop1
|
sp
|
| |
Eremogone micradenia
|
п
|
с
|
-
|
-
|
-
|
sol
|
-
|
-
|
| |
Euphorbia
|
п
|
с
|
-
|
sol
|
-
|
sol
|
sol
|
-
|
| |
Falcaria vulgaris
|
м
|
с
|
-
|
-
|
sp
|
sp
|
sp
|
sol
|
| |
Festuca pratensis
|
п
|
лг
|
sol
|
-
|
-
|
-
|
-
|
-
|
| |
Festuca valesiaca
|
п
|
с
|
sp
|
sp
|
sp
|
Cop1
|
Cop1
|
-
|
| |
Filipendula vulgaris
|
п
|
с
|
Cop1
|
Cop1
|
Cop1
|
sp
|
Cop1
|
Cop1
|
| |
Fragaria viridis
|
п
|
с
|
sp
|
sol
|
Cop1
|
sp
|
-
|
sp
|
| |
Galium boreale
|
п
|
лг
|
-
|
-
|
sol
|
-
|
-
|
-
|
| |
Galium tinctorium
|
п
|
с
|
sp
|
Cop1
|
sp
|
-
|
sol
|
sol
|
| |
Gallium verum
|
п
|
с
|
sp
|
sp
|
sp
|
sp
|
sp
|
sp
|
| |
Genista tinctoria
|
пк
|
с
|
sol
|
-
|
sol
|
sol
|
-
|
-
|
| |
Geranium sanquineum
|
п
|
с
|
-
|
sp
|
Cop1
|
-
|
-
|
-
|
| |
Hieracium
|
п
|
с
|
sp
|
-
|
-
|
sol
|
-
|
-
|
| |
Hypericum perforatum
|
п
|
с
|
-
|
sol
|
-
|
-
|
-
|
-
|
| |
Inula hirta
|
п
|
с
|
-
|
sol
|
sol
|
-
|
sol
|
-
|
| |
Iris aphylla
|
п
|
с
|
-
|
sp
|
-
|
-
|
sol
|
sp
|
| |
Knautia arvensis
|
п
|
с
|
-
|
-
|
sol
|
-
|
-
|
-
|
| |
Koeleria cristata
|
п
|
с
|
sol
|
-
|
-
|
-
|
-
|
-
|
| |
Lactuca serriola
|
м
|
ср
|
-
|
-
|
-
|
-
|
-
|
sol
|
| |
Lathyrus lacteus
|
п
|
с
|
-
|
sp
|
sol
|
sol
|
sol
|
-
|
| |
Leucanthemum vulgare
|
п
|
лг
|
sol
|
sol
|
sol
|
-
|
-
|
-
|
| |
Linaria ruthenica
|
п
|
с
|
-
|
-
|
-
|
sol
|
sol
|
-
|
| |
Linum nervosum
|
п
|
с
|
-
|
-
|
-
|
-
|
sol
|
-
|
| |
Medicago falcata
|
п
|
с
|
sol
|
sol
|
-
|
sp
|
-
|
-
|
| |
Myosotis arvensis
|
о
|
лг
|
-
|
-
|
-
|
sol
|
-
|
-
|
| |
Onobrychis arenaria
|
п
|
с
|
-
|
sol
|
-
|
sol
|
-
|
-
|
| |
Pedicularis kaufmannii
|
п
|
с
|
sol
|
-
|
-
|
-
|
-
|
-
|
| |
Phlomoides tuberosa
|
п
|
с
|
sol
|
sp
|
sol
|
sol
|
-
|
sol
|
| |
Plantago urvillii
|
п
|
с
|
sol
|
sol
|
-
|
-
|
-
|
-
|
| |
Poa angustifolia
|
п
|
с
|
sol
|
sp
|
sol
|
sp
|
sp
|
Cop1
|
| |
Potentilla argentea
|
п
|
с
|
sp
|
-
|
sol
|
-
|
-
|
-
|
| |
Potentilla humifusa
|
п
|
с
|
sol
|
-
|
-
|
-
|
-
|
-
|
| |
Primula veris
|
п
|
лг
|
-
|
-
|
sol
|
-
|
-
|
-
|
| |
Pulsatilla patens
|
п
|
с
|
-
|
-
|
-
|
-
|
sol
|
-
|
| |
Pyrethrum corymbosum
|
п
|
л
|
-
|
-
|
sp
|
-
|
-
|
-
|
| |
Ranunculus polyanthemos
|
п
|
с
|
sp
|
-
|
sol
|
sol
|
-
|
-
|
| |
Rumex acetosa
|
п
|
лг
|
-
|
-
|
sol
|
-
|
-
|
-
|
| |
Salvia nutans
|
п
|
с
|
sp
|
-
|
-
|
-
|
-
|
-
|
| |
Salvia pratensis
|
п
|
с
|
sol
|
sp
|
sp
|
sp
|
-
|
-
|
| |
Scabiosa ochroleuca
|
п
|
с
|
-
|
sol
|
-
|
-
|
-
|
-
|
| |
Scorzonera purpurea
|
п
|
с
|
-
|
-
|
sol
|
-
|
-
|
-
|
| |
Securigera varia
|
п
|
с
|
-
|
-
|
-
|
sol
|
-
|
-
|
| |
Serratula lycopifolia
|
п
|
с
|
-
|
-
|
sol
|
sol
|
-
|
-
|
| |
Serratula radiata
|
п
|
с
|
-
|
-
|
-
|
sol
|
-
|
-
|
| |
Seseli annuum
|
м
|
с
|
-
|
sp
|
sol
|
sol
|
-
|
-
|
| |
Stachys officinalis
|
п
|
л
|
-
|
-
|
-
|
-
|
sol
|
-
|
| |
Stachys recta
|
п
|
с
|
sp
|
sp
|
sol
|
sol
|
sol
|
-
|
| |
Stellaria graminea
|
п
|
лг
|
-
|
-
|
-
|
sp
|
-
|
-
|
| |
Stipa pennata
|
п
|
с
|
-
|
Cop1
|
sol
|
sol
|
Cop1
|
sol
|
| |
Thalictrum flexuosum
|
п
|
с
|
sol
|
-
|
sol
|
sp
|
-
|
-
|
| |
Thymus marschallianus
|
пк
|
с
|
sp
|
-
|
-
|
-
|
sol
|
-
|
| |
Trifolium alpestre
|
п
|
с
|
-
|
-
|
sp
|
sol
|
sol
|
-
|
| |
Valeriana officinalis
|
п
|
с
|
-
|
-
|
sol
|
sol
|
-
|
-
|
| |
Veratrum nigrum
|
п
|
л
|
-
|
-
|
-
|
-
|
-
|
sol
|
| |
Veronica chamaedrys
|
п
|
лг
|
-
|
-
|
sol
|
sp
|
-
|
-
|
| |
Veronica jacquinii
|
п
|
с
|
sol
|
sp
|
sol
|
sol
|
sol
|
-
|
| |
Veronica prostrata
|
п
|
с
|
-
|
sol
|
-
|
-
|
-
|
-
|
| |
Veronica spicata
|
п
|
с
|
sol
|
-
|
-
|
-
|
-
|
-
|
| |
Vicia tenuifolia
|
п
|
с
|
Cop1
|
sp
|
sp
|
sp
|
sp
|
sp
|
| |
Vincetoxicum hirundinaria
|
п
|
лг
|
sol
|
sol
|
-
|
-
|
sol
|
-
|
| |
Viola rupestris
|
п
|
с
|
-
|
sol
|
sol
|
-
|
-
|
-
|
| |
Viola
|
п
|
-
|
-
|
-
|
-
|
sol
|
-
|
Примечание: 1
– основные биоморфы: пк – полукустарники, полукустарнички, п – травянистые поликарпики, м – мало- и многолетние монокарпики, о – однолетники.
2
– эф – эколого-фитоценотическая приуроченность: с – степные виды, лг – луговые, л – лесные, ср – сорные, культурные.
* Расположение ПП: №1 – плакор напротив аншлага у старого кордона, косимый режим – в 2002 – 2003 гг. выкашивалась; №2 – плакор на феномаршруте напротив западной границы дальнего некосимого участка, косимый режим - 2002 – 2003 гг. не выкашивалась; №3 – плакор на экологическом профиле (б.Суры) рядом с точкой Щ 19/20, косимый режим – в 2002 г. выкашивалась, в 2003 г. не выкашивалась; №4 – плакор на экологическом профиле (б.Суры) рядом с точкой Щ 23/24, косимый режим – в 2002 – 2003 гг. выкашивалась; №5 – дальний некосимый участок на феномаршруте напротив №2; №6 – ближний некосимый участок.
Все данные по продуктивности видов растений на названных площадях, а так же расположенных на экологическом профиле и ближнем некосимом участке приведены в таблицах 7.2.1.7. – 7.2.1.9.
Таблица 7.2.1.7
Надземная фитомасса (возд.сух.вес) луговой степи в г/0,25 м2
на пробных площадях,
заложенных на экологическом профиле в районе балки Суры в июле 2004 г.
| №
|
Видовой состав
|
Надземная фитомасса, г/0,25 м2
|
| ПП № 3, Экологический профиль Щ 19/201
|
ПП № 4, Экологический профиль Щ 23/242
|
| №1
|
№2
|
№3
|
№4
|
№5
|
ср
|
№1
|
№2
|
№3
|
№4
|
№5
|
ср
|
| 1
|
2
|
3
|
4
|
5
|
6
|
7
|
8
|
9
|
10
|
11
|
12
|
13
|
14
|
| |
Злаки широколистные
|
21.9
|
12.4
|
45.6
|
34.0
|
18.6
|
26.5
|
18.8
|
41.0
|
68.7
|
64.8
|
26.8
|
44.02
|
| |
Злаки узколистные
|
16.4
|
35.0
|
35.9
|
29.0
|
10.9
|
25.44
|
66.4
|
28.7
|
69.6
|
52.7
|
30.5
|
49.58
|
| |
Carex
|
0
|
0.1
|
0
|
0.1
|
0
|
0.04
|
0
|
0
|
0
|
0
|
0
|
0
|
| |
Achillea
|
0.6
|
1.2
|
0
|
0.7
|
0.6
|
0.62
|
2.5
|
0.5
|
0.2
|
6
|
0.1
|
1.86
|
| |
Allium oleraceum
|
0.3
|
1.0
|
0
|
0.2
|
1.1
|
0.52
|
0.7
|
0.1
|
0.4
|
0.3
|
0
|
0.3
|
| |
Adonis vernalis
|
0
|
8.2
|
0
|
0
|
0
|
1.64
|
0
|
0
|
0
|
0
|
0
|
0
|
| |
Anthericum ramosum
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0.1
|
0
|
0
|
0
|
| |
Campanula altaica
|
0.1
|
0
|
0
|
0
|
0
|
0.02
|
0
|
0.4
|
2.8
|
0.2
|
0.5
|
0.78
|
| |
Centaurea
|
3.1
|
0
|
0
|
0
|
0
|
0.62
|
0
|
0
|
0
|
0
|
0
|
0
|
| |
Chamaecytisus ruthenicus
|
0
|
0.1
|
3.8
|
0
|
0
|
0.78
|
0
|
0
|
0
|
0
|
0
|
0
|
| |
Clematis integrifolia
|
0.4
|
0
|
0
|
0
|
0
|
0.08
|
2.2
|
0
|
0
|
0
|
0
|
0.44
|
| |
Convolvulus arvensis
|
0
|
0
|
0
|
0
|
1.2
|
0.24
|
0
|
0
|
0
|
0
|
0
|
0
|
| |
Dracocephalum ruyschiana
|
0
|
0.8
|
0
|
0
|
0
|
0.16
|
0
|
0
|
0
|
0
|
0
|
0
|
| |
Draba sibirica
|
1.4
|
2.0
|
0.3
|
1.9
|
0.6
|
1.24
|
6.1
|
8.3
|
0.4
|
7.2
|
4.0
|
5.2
|
| |
Euphorbia
|
0
|
0
|
0
|
0
|
0
|
0
|
0.1
|
0
|
0
|
0
|
0
|
0.02
|
| |
Eremogone micradenia
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0.5
|
0
|
0.1
|
| |
Falcaria vulgaris
|
3.2
|
1.3
|
3.9
|
0
|
0.7
|
1.82
|
5.7
|
9.3
|
24.7
|
0.8
|
5.8
|
9.26
|
| |
Fragaria viridis
|
6.4
|
2.2
|
2.1
|
3.7
|
11.9
|
5.26
|
0
|
0
|
0.5
|
0
|
6.1
|
1.32
|
| |
Filipendula vulgaris
|
12.5
|
2.5
|
7.6
|
15.6
|
13.5
|
10.34
|
1.6
|
2.3
|
2.7
|
3.4
|
6.6
|
3.32
|
| |
Galium
|
8.6
|
4.2
|
1.6
|
15
|
12.2
|
8.32
|
14.8
|
8.6
|
9.4
|
4.5
|
0
|
7.46
|
| |
Genista tinctoria
|
0
|
0
|
0
|
0.7
|
0
|
0.14
|
3.9
|
0
|
0
|
0.1
|
0.1
|
0.82
|
| |
Geranium sanquineum
|
2.2
|
30
|
2.4
|
15.1
|
33
|
16.54
|
0
|
0
|
0
|
0
|
0
|
0
|
| |
Hieracium
|
0
|
0
|
0
|
0
|
0
|
0
|
0.4
|
0
|
0
|
0
|
0
|
0.08
|
| |
Inula hirta
|
0
|
0
|
3.5
|
0.6
|
0.1
|
0.84
|
0
|
0
|
0
|
0
|
0
|
0
|
| Таблица 7.2.1.7 (окончание)
|
| 1
|
2
|
3
|
4
|
5
|
6
|
7
|
8
|
9
|
10
|
11
|
12
|
13
|
14
|
| 25.
|
Knautia arvensis
|
0
|
0
|
0
|
0.2
|
0
|
0.04
|
0
|
0
|
0
|
0
|
0
|
0
|
| |
Lathyrus lacteus
|
0.7
|
0.6
|
1.1
|
0.1
|
0
|
0.5
|
0
|
0.1
|
0
|
0
|
0
|
0.02
|
| |
Leucanthemum vulgare
|
0.1
|
0.1
|
0
|
0.1
|
0.1
|
0.08
|
0
|
0
|
0
|
0
|
0
|
0
|
| |
Linaria ruthenica
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0.1
|
0
|
0.1
|
0
|
0.04
|
| |
Medicago falcate
|
0
|
0
|
0
|
0
|
0
|
0
|
3.3
|
29.6
|
12.3
|
2.6
|
14.2
|
12.4
|
| |
Myosotis arvensis
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0.1
|
0
|
0
|
0.02
|
| |
Onobrychis arenaria
|
0
|
0
|
0
|
0
|
0
|
0
|
3.2
|
0
|
0
|
0
|
0
|
0.64
|
| |
Pyrethrum corymbosum
|
4.9
|
0
|
0
|
0
|
5.7
|
2.12
|
0
|
0
|
0
|
0
|
0
|
0
|
| |
Phlomoides tuberosa
|
1.3
|
2.4
|
0
|
0
|
2.0
|
1.14
|
0.1
|
0
|
4.5
|
0
|
12.5
|
3.42
|
| |
Potentilla argentea
|
0
|
0
|
0
|
0.1
|
0
|
0.02
|
0
|
0
|
0
|
0
|
0
|
0
|
| |
Primula veris
|
0.8
|
0.5
|
0.4
|
1.7
|
0.4
|
0.76
|
0
|
0
|
0
|
0
|
0
|
0
|
| |
Ranunculus polyanthemos
|
0.1
|
0
|
0
|
0
|
0
|
0.02
|
0
|
0
|
0.1
|
1.9
|
0
|
0.4
|
| |
Rumex acetosa
|
0
|
0
|
0
|
0.1
|
0.4
|
0.1
|
0
|
0
|
0
|
0
|
0
|
0
|
| |
Salvia pratensis
|
5.9
|
2.2
|
0.3
|
0
|
2.3
|
2.74
|
0
|
0
|
0
|
0.7
|
0
|
0.14
|
| |
Scorzonera purpurea
|
0.6
|
0
|
0
|
0.1
|
0
|
0.14
|
0
|
0
|
0
|
0
|
0
|
0
|
| |
Securigera varia
|
0
|
0
|
0
|
0
|
0
|
0
|
2.2
|
0
|
0
|
0
|
0
|
0.44
|
| |
Serratula lycopifolia
|
0
|
1.0
|
0
|
0.8
|
0.5
|
0.46
|
2
|
0
|
1.1
|
0
|
7.7
|
2.16
|
| |
Seseli annuum
|
0.5
|
0
|
0
|
0.4
|
1.4
|
0.46
|
0
|
0
|
0
|
0.2
|
0.9
|
0.22
|
| |
Stachys recta
|
0
|
0
|
17
|
0
|
0
|
3.4
|
0
|
6.8
|
0
|
0
|
4.7
|
2.3
|
| |
Stellaria graminea
|
0
|
0
|
0
|
0
|
0
|
0
|
0.2
|
0
|
1.5
|
0.1
|
1.0
|
0.56
|
| |
Thalictrum flexuosum
|
0
|
0.1
|
0.5
|
0
|
0.2
|
0.16
|
2.4
|
0
|
0
|
1.9
|
0.5
|
0.96
|
| |
Trifolium
|
4.7
|
0.8
|
1.7
|
2.0
|
0
|
1.84
|
5.6
|
0
|
0
|
0
|
0
|
1.12
|
| |
Vicia tenuifolia
|
12.1
|
2.2
|
5.3
|
23.7
|
36.1
|
15.88
|
23.0
|
52.4
|
20.4
|
80.1
|
28.2
|
40.82
|
| |
Valeriana rossica
|
0
|
0.2
|
0.1
|
0
|
0
|
0.06
|
2
|
0
|
0
|
0
|
1.9
|
0.78
|
| |
Veronica
|
0.5
|
0.4
|
0.1
|
0
|
0.3
|
0.26
|
2.8
|
3.1
|
0.3
|
1.2
|
2.6
|
2
|
| |
Viola sp
|
0
|
0
|
0
|
0
|
0.1
|
0.02
|
0
|
0
|
0
|
0
|
0
|
0
|
| |
подстилка
|
87.1
|
83.8
|
102.1
|
101.1
|
90.9
|
93.0
|
22.4
|
64.8
|
21.9
|
35.1
|
31.4
|
35.12
|
| |
мох
|
21.2
|
67.9
|
0
|
52.1
|
12.1
|
30.66
|
16.2
|
0
|
8.4
|
0
|
13.3
|
7.58
|
Примечание: 1
– в 2003 и 2004 г не выкашивался; 2
– в 2003 г. выкашивался, в 2004 г. не выкашивался
Таблица 7.2.1.8
Надземная фитомасса (возд.сух.вес) луговой степи в г / 0,25 м2
на пробных площадях,
заложенных на плакоре в июле 2004 г.
| №
|
Видовой состав
|
Надземная фитомасса, г/0,25 м2
|
| ПП № 1, плакор возле аншлага1
|
ПП № 2, плакор на косимом феномаршруте2
|
| №1
|
№2
|
№3
|
№4
|
№5
|
ср
|
№1
|
№2
|
№3
|
№4
|
№5
|
ср
|
| 1
|
2
|
3
|
4
|
5
|
6
|
7
|
8
|
9
|
10
|
11
|
12
|
13
|
14
|
| 1
|
Злаки широколистные
|
48.0
|
67.4
|
86.9
|
41.7
|
23.9
|
53.58
|
20.5
|
45
|
25.3
|
17.5
|
16.6
|
24.98
|
| 2
|
Злаки узколистные
|
11.1
|
13.5
|
1.2
|
6.7
|
2.5
|
7
|
10.2
|
5.0
|
14.0
|
16.9
|
12.60
|
11.74
|
| 3
|
Carex
|
0
|
0.7
|
0
|
0
|
0.1
|
0.16
|
0.3
|
0
|
2.7
|
0
|
0
|
0.6
|
| 4
|
Achillea
|
0.2
|
0.2
|
0.1
|
1.4
|
0
|
0.38
|
0
|
0
|
0
|
0
|
0
|
0
|
| 5
|
Allium oleraceum
|
0
|
0
|
0.3
|
0.2
|
0
|
0.1
|
0.9
|
0.5
|
0.3
|
0.6
|
0
|
0.46
|
| 6
|
Astragalus danicus
|
0
|
0
|
0.1
|
1.0
|
0
|
0.22
|
0
|
0
|
0
|
0
|
0
|
0
|
| 7
|
Campanula altaica
|
0
|
0
|
0
|
0.6
|
0.5
|
0.22
|
0
|
0
|
0
|
0
|
0
|
0
|
| 8
|
Centaurea
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
1.4
|
0.28
|
| 9
|
Chamaecytisus ruthenicus
|
0
|
0
|
0
|
0
|
0
|
0
|
2.0
|
0
|
0
|
1.3
|
0
|
0.66
|
| 10
|
Convolvulus arvensis
|
0.2
|
2.1
|
0
|
0
|
0.2
|
0.5
|
0
|
0
|
0
|
0
|
0
|
0
|
| 11
|
Dianthus andrzejowskianus
|
0.4
|
0
|
0
|
0
|
0
|
0.08
|
1.3
|
0
|
0
|
0
|
0
|
0.26
|
| 12
|
Dracocephalum ruyschiana
|
0
|
0
|
0
|
0
|
0
|
0
|
2.3
|
0
|
0
|
0
|
0
|
0.46
|
| 13
|
Draba sibirica
|
0
|
0
|
0.4
|
0
|
0.2
|
0.12
|
1.1
|
1.8
|
0.5
|
2.2
|
3
|
1.72
|
| 14
|
Euphorbia
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0.5
|
0
|
0.1
|
| 15
|
Fragaria viridis
|
0
|
0.2
|
0.1
|
1
|
0.5
|
0.36
|
0
|
0
|
0
|
0.5
|
0
|
0.1
|
| 16
|
Filipendula vulgaris
|
6.0
|
0.2
|
2.0
|
6.0
|
4.7
|
3.78
|
12.2
|
8.3
|
11.5
|
10.1
|
7.5
|
9.92
|
| 17
|
Galium
|
2.0
|
0.7
|
0.7
|
3
|
1.8
|
1.64
|
2.4
|
1.5
|
1.9
|
2.6
|
0
|
1.68
|
| 18
|
Genista tinctoria
|
0
|
0
|
0
|
0.6
|
0
|
0.12
|
0
|
0
|
0
|
0
|
0
|
0
|
| 19
|
Geranium sanquineum
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0.6
|
0
|
32.9
|
0
|
6.7
|
| 20
|
Hieracium
|
0
|
0.1
|
0.1
|
2.2
|
2.7
|
1.02
|
0
|
0
|
0
|
0
|
0
|
0
|
| 21
|
Hypericum perforatum
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0.2
|
0
|
0
|
0.04
|
| 22
|
Inula hirta
|
0
|
0
|
0
|
0
|
0
|
0
|
0.2
|
0
|
0
|
0
|
0
|
0.04
|
| 23
|
Iris aphylla
|
0
|
0
|
0
|
0
|
0
|
0
|
3.3
|
0
|
0
|
0
|
4.4
|
1.54
|
| Таблица 7.2.1.8 (окончание)
|
| 1
|
2
|
3
|
4
|
5
|
6
|
7
|
8
|
9
|
10
|
11
|
12
|
13
|
14
|
| 24
|
Lathyrus lacteus
|
0
|
0
|
0
|
0
|
0
|
0
|
0.4
|
5.2
|
0
|
0
|
0
|
1.12
|
| 25
|
Leucanthemum vulgare
|
0
|
0
|
0.1
|
0
|
0
|
0.02
|
0
|
5.6
|
0
|
0
|
0
|
1.12
|
| 26
|
Medicago falcate
|
0
|
0
|
0
|
0
|
0.1
|
0.02
|
0
|
0
|
0
|
1.3
|
0
|
0.26
|
| 27
|
Onobrychis arenaria
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
1.3
|
0
|
0.26
|
| 28
|
Phlomoides tuberosa
|
0
|
2.0
|
0
|
0
|
0.3
|
0.46
|
0
|
0
|
0
|
4.2
|
0
|
0.84
|
| 29
|
Pedicularis kaufmannii
|
0
|
0
|
0
|
0
|
2.7
|
0.54
|
0
|
0
|
0
|
0
|
0
|
0
|
| 30
|
Potentilla argentea
|
0.6
|
0
|
0.1
|
0
|
0
|
0.14
|
0
|
0
|
0
|
0
|
0
|
0
|
| 31
|
Plantago urvillei
|
0.2
|
0
|
0.5
|
0
|
0.5
|
0.24
|
0
|
0
|
0.1
|
0
|
0
|
0.02
|
| 32
|
Ranunculus polyanthemos
|
0
|
0
|
0
|
3.8
|
1.9
|
1.14
|
0
|
0
|
0
|
0
|
0
|
0
|
| 33
|
Salvia pratensis
|
5.8
|
0.3
|
0
|
1.2
|
0.2
|
1.5
|
0.8
|
0
|
2.8
|
0
|
2.8
|
1.28
|
| 34
|
Scabiosa ochroleuca
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0.6
|
0
|
0
|
0.12
|
| 35
|
Seseli annuum
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0.3
|
0.7
|
0.5
|
0
|
0.3
|
| 36
|
Stachys recta
|
1.0
|
0
|
0
|
2.2
|
0
|
0.64
|
0
|
2.0
|
2.0
|
0.5
|
0
|
0.9
|
| 37
|
Thalictrum flexuosum
|
0.5
|
0
|
0
|
0
|
0
|
0.1
|
0
|
0
|
0
|
0
|
0
|
0
|
| 38
|
Thymus marschallianus
|
0
|
0
|
1.5
|
0
|
2.9
|
0.88
|
0
|
0
|
0
|
0
|
0
|
0
|
| 39
|
Trifolium
|
3.7
|
0
|
0
|
0
|
2.2
|
1.18
|
0.5
|
0
|
0.2
|
0
|
0
|
0.14
|
| 40
|
Vicia tenuifolia
|
0
|
11.1
|
3.2
|
0
|
4.3
|
3.72
|
1.4
|
0.9
|
5.6
|
2.9
|
4.0
|
2.96
|
| 41
|
Veronica
|
0
|
0
|
0
|
1.0
|
0
|
0.2
|
0.3
|
0
|
0.9
|
1.2
|
0
|
0.48
|
| 42
|
Viola sp
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0.1
|
0
|
0
|
0.02
|
| 43
|
Vincetoxicum hirundinaria
|
0
|
0
|
0
|
1.0
|
0
|
0.2
|
0.1
|
0
|
0
|
0
|
0
|
0.02
|
| 44
|
подстилка
|
7.1
|
39.2
|
32.2
|
35.9
|
25.9
|
28.06
|
111.2
|
87.7
|
107.7
|
173.3
|
110.3
|
110.3
|
| 45
|
мох
|
56.8
|
28.2
|
1.9
|
6.9
|
22.3
|
23.22
|
2.8
|
0
|
9.5
|
0
|
0
|
2.46
|
1
– выкашивался в 2003 и 2004 гг.
2
– не выкашивался в 2003 и 2004 гг.
Таблица 7.2.1.9
Надземная фитомасса (возд.сух.вес) луговой степи в г / 0,25 м2
на пробных площадях,
заложенных на некосимых участках в июле 2004 г.
| №
|
Видовой состав
|
Надземная фитомасса, г/0,25 м2
|
| ПП № 5, дальний некосимый
|
ПП № 6, ближний некосимый
|
| №1
|
№2
|
№3
|
№4
|
№5
|
ср
|
№1
|
№2
|
№3
|
№4
|
№5
|
ср
|
| 1
|
2
|
3
|
4
|
5
|
6
|
7
|
8
|
9
|
10
|
11
|
12
|
13
|
14
|
| 1
|
Злаки широколистные
|
43.8
|
53.7
|
38.8
|
8.1
|
55.6
|
40.0
|
28.4
|
51.0
|
50.4
|
107.8
|
120.2
|
71.56
|
| 2
|
Злаки узколистные
|
84.2
|
52.1
|
95.3
|
103.8
|
56.0
|
78.28
|
138.2
|
42.7
|
120.7
|
7.4
|
45.8
|
70.96
|
| 3
|
Carex
|
0
|
0
|
0
|
0
|
0
|
0
|
0.2
|
0
|
0
|
0
|
0
|
0.04
|
| 4
|
Achillea
|
0
|
0.6
|
0
|
0.4
|
0
|
0.2
|
0
|
0
|
0
|
0
|
0
|
0
|
| 5
|
Allium oleraceum
|
0
|
0
|
0
|
0
|
0
|
0
|
1.6
|
0
|
0
|
0
|
0
|
0.32
|
| 6
|
Anthericum ramosum
|
0.3
|
0
|
0
|
0
|
0
|
0.06
|
0
|
0
|
0
|
0
|
0
|
0
|
| 7
|
Asparagus officinalis
|
0
|
19.6
|
0
|
0
|
0
|
3.92
|
0
|
0
|
0
|
0
|
0
|
0
|
| 8
|
Chamaecytisus ruthenicus
|
0
|
0
|
2.7
|
0
|
0
|
0.54
|
0
|
0
|
0
|
0
|
0
|
0
|
| 9
|
Cirsium setosum
|
18.0
|
0
|
0
|
0
|
0
|
3.6
|
16.3
|
1.0
|
96.6
|
2.0
|
0.4
|
23.26
|
| 10
|
Draba sibirica
|
0
|
0
|
0
|
0
|
0
|
0
|
2.1
|
0
|
0
|
0
|
0
|
0.42
|
| 11
|
Euphorbia
|
0.1
|
0.2
|
0
|
0.1
|
0.1
|
0.1
|
0
|
0
|
0
|
0
|
0
|
0
|
| 12
|
Falcaria vulgaris
|
2.6
|
0.9
|
0
|
0.4
|
1.0
|
0.98
|
2.1
|
0
|
0
|
0
|
0
|
0.42
|
| 13
|
Fragaria viridis
|
0
|
0
|
0
|
0
|
0
|
0
|
5.3
|
5.1
|
0
|
1.2
|
0.4
|
2.4
|
| 14
|
Filipendula vulgaris
|
26.8
|
8.8
|
22.7
|
17.2
|
9.9
|
17.08
|
9.4
|
1.3
|
7.6
|
9.7
|
5.7
|
6.74
|
| 15
|
Galium
|
1.6
|
4.0
|
2.7
|
2.0
|
1.3
|
2.32
|
12.9
|
5.2
|
4.3
|
2.6
|
2.1
|
5.42
|
| 16
|
Inula hirta
|
0.8
|
0
|
0
|
0
|
0
|
0.16
|
0
|
0
|
0
|
0
|
0
|
0
|
| 17
|
Iris aphylla
|
0
|
0
|
0
|
1.7
|
7.7
|
1.88
|
0
|
0
|
0
|
27.6
|
0
|
5.52
|
| 18
|
Lactuca serriola
|
0
|
0
|
0
|
0
|
0
|
0
|
4.4
|
0
|
0
|
0
|
0
|
0.88
|
| 19
|
Linum nervosum
|
0
|
0.3
|
0.1
|
0
|
0
|
0.08
|
0
|
0
|
0
|
0
|
0
|
0
|
| 20
|
Lathyrus lacteus
|
0.4
|
0
|
0.1
|
1.7
|
0
|
0.44
|
0
|
0
|
0
|
0
|
0
|
0
|
| 21
|
Linaria ruthenica
|
1.2
|
1.9
|
7.6
|
0
|
4.9
|
3.12
|
0
|
0
|
0
|
0
|
0
|
0
|
| 22
|
Phlomoides tuberosa
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
2.5
|
0
|
0
|
1.2
|
0.74
|
| 23
|
Pulsatilla patens
|
2.2
|
0
|
0.6
|
0
|
0
|
0.56
|
0
|
0
|
0
|
0
|
0
|
0
|
| Таблица 7.2.1.9 (окончание)
|
| 1
|
2
|
3
|
4
|
5
|
6
|
7
|
8
|
9
|
10
|
11
|
12
|
13
|
14
|
| 24
|
Stachys recta
|
0
|
0
|
0
|
2.4
|
3.7
|
1.22
|
0
|
0
|
0
|
0
|
0
|
0
|
| 25
|
Stachys officinalis
|
0
|
0
|
0
|
4.1
|
0
|
0.82
|
0
|
0
|
0
|
0
|
0
|
0
|
| 26
|
Thymus marschallianus
|
0
|
0
|
0.1
|
0
|
0
|
0.02
|
0
|
0
|
0
|
0
|
0
|
0
|
| 27
|
Trifolium
|
0
|
0
|
0
|
2.2
|
0
|
0.44
|
0
|
0
|
0
|
0
|
0
|
0
|
| 28
|
Vicia tenuifolia
|
0.2
|
0.2
|
0.8
|
4.2
|
0.1
|
1.1
|
2.8
|
2.4
|
45.0
|
0.1
|
0
|
10.06
|
| 29
|
Veronica
|
0
|
0
|
0
|
0.6
|
0
|
0.12
|
0
|
0
|
0
|
0
|
0
|
0
|
| 30
|
Viola sp
|
0
|
0
|
0
|
0.1
|
0
|
0.02
|
0
|
0
|
0
|
0
|
0
|
0
|
| 31
|
Veratrum nigrum
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
11.1
|
0
|
2.22
|
| 32
|
Vincetoxicum hirundinaria
|
0
|
0.4
|
0
|
0
|
0.3
|
0.14
|
0
|
0
|
0
|
0
|
0
|
0
|
| 33
|
подстилка
|
278.0
|
138.6
|
276.1
|
276.9
|
209.0
|
233.92
|
30.9
|
63.8
|
420.6
|
330.2
|
300.7
|
229.24
|
| 34
|
мох
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
0
|
4.5
|
0
|
0
|
0.9
|
7.2.2. ЛЕСНАЯ РАСТИТЕЛЬНОСТЬ
7.2.2.1. Динамика опада древесно-кустарникового полога лесных насаждений
7.2.2.1а. Динамика опада на постоянной пробной площади № 7 (Ямской участок)
На ПП №7 размещены 10 опадоуловителей ящичного типа размером 1 кв.м. каждый. Ящики расположены по склону и пронумерованы от 1 до 10 снизу вверх. Из каждого ящика опад собирали ежемесячно, исключая снежный период. Опад за декабрь вошел в общий зимний сбор 2005 г. Разобранный по фракциям опад сушили при температуре 105 градусов в сушильном шкафу, а затем взвешивали с точностью до 0,01 г. Итоги (сумма по всем опадоуловителям) приведены в таблице 7.2.2.1.
Таблица 7.2.2.1
Количество опада (кг/га) на ПП-7 (Ямской участок) за 2004 год по месяцам.
| Фракции
|
Месяцы
|
Итого
|
| по видам
|
I-III
|
IV
|
V
|
VI
|
VII
|
VIII
|
IX
|
X
|
XI
|
за год
|
| 1
|
2
|
3
|
4
|
5
|
6
|
7
|
8
|
9
|
10
|
11
|
| Ветки Д:
|
| облом
|
19,92
|
16,1
|
21,6
|
3,6
|
12,6
|
9,6
|
57,4
|
11,9
|
0,8
|
153,52
|
| опад
|
13,05
|
6,2
|
14,2
|
7,44
|
10,9
|
12,12
|
34,3
|
1,3
|
99,51
|
| Итого
|
32,97
|
22,3
|
35,8
|
11,04
|
23,5
|
21,72
|
91,7
|
13,2
|
0,8
|
253,03
|
| Листья Д:
|
| летние
|
36,9
|
33,7
|
70,6
|
| осенние
|
150,73
|
18,57
|
0,81
|
1,22
|
125
|
231,4
|
108,1
|
635,83
|
| весен.
|
42,6
|
20,1
|
62,7
|
| Итого
|
150,73
|
18,57
|
43,41
|
21,32
|
36,9
|
33,7
|
125
|
231,4
|
108,1
|
698,53
|
| Прочее по Д:
|
| цветы
|
0
|
| желуди
|
0,7
|
0,02
|
13,6
|
27,3
|
12,8
|
1,22
|
55,64
|
| кора
|
15,12
|
1,64
|
3,62
|
3,3
|
10,6
|
7,7
|
37
|
8,4
|
1,4
|
88,78
|
| почки
|
0,82
|
10,54
|
12,6
|
6,8
|
5,9
|
9
|
0,2
|
0,42
|
46,28
|
| Итого
|
16,64
|
1,64
|
14,16
|
15,92
|
31
|
40,9
|
58,8
|
9,82
|
1,82
|
190,7
|
| Всего по Дубу
|
200,34
|
42,51
|
93,37
|
48,28
|
91,4
|
96,32
|
275,5
|
254,42
|
110,72
|
1142,26
|
| Неопр. опад
|
3,08
|
4,9
|
16,44
|
10,77
|
7,1
|
8,88
|
9,52
|
1,71
|
2,4
|
64,8
|
| Лщ: листья
|
5,18
|
0,32
|
12,12
|
5,8
|
11,44
|
29,2
|
20,4
|
16,5
|
100,96
|
| облом
|
0
|
| плоды
|
0
|
| сережки
|
0,01
|
5
|
4,52
|
1
|
10,53
|
| кора
|
0
|
| Гр: листья
|
0,09
|
0,12
|
1,32
|
0,5
|
0,2
|
7,57
|
29,4
|
4,42
|
6,12
|
49,74
|
| плоды
|
0
|
| цветы
|
0
|
| облом
|
0
|
| Лп: листья
|
0,05
|
0,02
|
0,02
|
0,4
|
0,49
|
| цветы
|
0
|
| плоды
|
0
|
| Ивк: листья
|
0
|
| Чрм: листья
|
0
|
| облом
|
0
|
| плоды
|
0
|
| Окончание табл. 7.2.2.1
|
| 1
|
2
|
3
|
4
|
5
|
6
|
7
|
8
|
9
|
10
|
11
|
| Ос: листья
|
0,4
|
2,4
|
1,52
|
4,32
|
| почки
|
0,8
|
0,8
|
| сережки
|
0
|
| Клт: листья
|
0,71
|
5,71
|
6,5
|
0,8
|
0,78
|
14,5
|
| облом
|
0
|
| цветы
|
| семена
|
0,06
|
4,6
|
4,66
|
| Кл.яс.Листья
|
0
|
| плоды
|
0
|
| Кл.о.плоды
|
| Ирга. листья
|
| Калина.плоды
|
| Ябл: листья
|
0,12
|
7,14
|
3,62
|
2,6
|
1,4
|
5,6
|
0,02
|
20,5
|
| облом
|
0,12
|
0,12
|
| опад
|
0
|
| плоды
|
0
|
| Трн: листья
|
12
|
24,1
|
36,1
|
| плоды
|
0
|
| опад
|
| облом
|
0
|
| Бузина.облом
|
| Листья
|
| Жст: листья
|
0
|
| плоды
|
0
|
| Крш.листья
|
0
|
| плоды
|
| облом
|
0
|
| Свид.листья
|
0,02
|
0,87
|
1,84
|
8,2
|
2,14
|
13,07
|
| опад
|
| плоды
|
| Боярышник,листья
|
0
|
| Берескл.лист
|
| Береза.лист
|
0
|
| Ил: семена
|
| листья
|
0,02
|
0,02
|
| Ясп: семена
|
0
|
| Акация б.семена
|
4,3
|
| Акация ж.семена
|
1,5
|
1,5
|
| Травян. опад
|
6,7
|
0,24
|
0,02
|
8,2
|
5,2
|
0,6
|
20,96
|
| Лишайники
|
0,15
|
0,02
|
0,17
|
| Галлы
|
0,2
|
0,02
|
| Животн. опад
|
0,02
|
0,02
|
1,6
|
3,9
|
7,7
|
1,32
|
14,56
|
| Насекомые
|
4,15
|
4
|
0,04
|
0,62
|
0,24
|
9,05
|
| Грибы
|
| Экскременты
|
| насекомых
|
0
|
| животных
|
0
|
| Итого
|
21,7
|
10,56
|
41,96
|
35,83
|
20
|
52,39
|
116,14
|
44,81
|
27,98
|
366,85
|
| Всего
|
222,04
|
53,07
|
135,33
|
84,11
|
111,4
|
148,71
|
391,64
|
299,23
|
138,7
|
1509,11
|
Распределение опада по основным фракциям приведено в таблице 7.2.2.2.
Таблица 7.2.2.2.
Количество опада ( кг/га ) на ПП-7 (Ямской участок) за 2004 г. по основным фракциям.
| Фракции
|
Количество (кг./га.)
|
% от валового опада
|
| Средний валовой опад за 9 лет наблюдений (1990-1998) по данным ЦЧЗ – 4595,33
Средний валовой опад за последние 3 года наблюдений (2001-2003) – 3514,36
|
| Валовой опад
|
1509,11
|
100
|
| Листья дуба
|
698,53
|
46,29
|
| Ветки и кора дуба
|
341,81
|
22,65
|
| Прочее по дубу
|
101,92
|
6,75
|
| Листья прочих древесно-кустарниковых пород
|
239,7
|
15,89
|
| Прочий древесно-кустарниковый опад (без листьев)
|
17,61
|
1,16
|
| Прочий опад
|
109,54
|
7,26
|
За 2004 год. общее количество опада оказалось наименьшим за все время наблюдений. Преобладает дубовая фракция, составляющая более 75% от валового опада.
7.2.2.1б. Динамики опада на стационаре участка «Лес на Ворскле».
С 2004 г. начаты наблюдения на участке «Лес на Ворскле». В 10 квартале установлены 10 опадоуловителей стандартных размеров (1х1м). Методика работы и время сбора опада аналогичны применяемым на Ямском участке. В апреле 3 опадоуловителя были повреждены и в течение 3 месяцев, до их восстановления, опад собирали из 7 ящиков. За этот период итоговые данные получены методом интерполяции. Результаты приведены в табл. 7.2.2.3.
Таблица 7.2.2.3
Количество опада (кг/га) на стационаре участка «Лес на Ворскле» за 2004 год по месяцам.
| Фракции
|
Месяцы
|
Итого
|
| по видам
|
I-IV
|
V
|
VI
|
VII
|
VIII
|
IX
|
X
|
XI
|
XII
|
за год
|
| 1
|
2
|
3
|
4
|
5
|
6
|
7
|
8
|
9
|
10
|
11
|
| Ветки Д:
|
| облом
|
16,7
|
20,5
|
25,9
|
274,6
|
5,36
|
31,7
|
65,4
|
33,72
|
6,3
|
480,18
|
| опад
|
1,5
|
1,1
|
11,6
|
11,6
|
0,02
|
5,9
|
5,3
|
2,3
|
39,32
|
| Итого
|
18,2
|
21,6
|
37,5
|
286,2
|
5,38
|
37,6
|
70,7
|
36,02
|
6,3
|
519,5
|
| Листья Д:
|
| летние
|
7,7
|
58,1
|
11,02
|
76,82
|
| осенние
|
4,1
|
3,5
|
1,4
|
31,57
|
166,3
|
15,02
|
4,46
|
226,35
|
| весен.
|
2,3
|
1,1
|
3,4
|
| Итого
|
4,1
|
5,8
|
9,1
|
59,2
|
11,02
|
31,57
|
166,3
|
15,02
|
4,46
|
306,57
|
| Прочее по Д:
|
| цветы
|
0
|
| желуди
|
0,02
|
0,02
|
| кора
|
8,1
|
21
|
31,4
|
27,3
|
4,4
|
9,25
|
10,9
|
18,3
|
9,52
|
140,17
|
| почки
|
2,7
|
8,2
|
4,1
|
1,7
|
1,56
|
0,4
|
1,1
|
19,76
|
| Итого
|
10,8
|
29,22
|
35,5
|
29
|
5,96
|
9,65
|
12
|
18,3
|
9,52
|
159,95
|
| Продолжение табл. 7.2.2.3
|
| 1
|
2
|
3
|
4
|
5
|
6
|
7
|
8
|
9
|
10
|
11
|
| Всего по Д
|
33,1
|
56,62
|
82,1
|
374,4
|
22,36
|
78,82
|
249
|
69,34
|
20,28
|
986,02
|
| Неопр. опад
|
33,8
|
34,7
|
31,9
|
26,4
|
11,82
|
4,7
|
6,14
|
4,06
|
9,72
|
163,24
|
| Лщ: листья
|
1,6
|
3,8
|
| облом
|
0
|
| плоды
|
0
|
| сережки
|
0
|
| кора
|
0
|
| Гр: листья
|
1,3
|
5
|
6,3
|
| плоды
|
0
|
| цветы
|
0
|
| облом
|
0
|
| Лп: листья
|
0,02
|
10,6
|
11,7
|
11,42
|
138,4
|
7,72
|
0,32
|
180,18
|
| цветы
|
5,5
|
4
|
2,7
|
29,65
|
41,85
|
| плоды
|
4,1
|
3,74
|
17,32
|
6,66
|
2,42
|
34,24
|
| Ивк: листья
|
0
|
| Чрм: листья
|
0
|
| облом
|
0
|
| плоды
|
0
|
| Ос: листья
|
1,1
|
2,4
|
30,7
|
3,1
|
2,72
|
15,9
|
116,92
|
4,4
|
177,24
|
| почки
|
3,6
|
9,8
|
13,4
|
| облом
|
2,3
|
18,6
|
20,9
|
| сережки
|
7,1
|
1
|
8,1
|
| Клт: листья
|
5,6
|
14,9
|
20,5
|
| облом
|
2,6
|
2,6
|
| цветы
|
0
|
| семена
|
0
|
| Кл.яс.Листья
|
0
|
| плоды
|
0
|
| Кл.о.плоды
|
7,54
|
4,62
|
0,95
|
13,11
|
| облом
|
44,9
|
22,9
|
4
|
63,5
|
12,1
|
6,8
|
154,2
|
| листья
|
0,3
|
35,8
|
69,6
|
24,3
|
18,9
|
121,6
|
1040,6
|
16,9
|
1,56
|
1329,56
|
| почки
|
5,1
|
11,7
|
16,8
|
| кора
|
4,2
|
4,2
|
| цв.и прицв.
|
4,9
|
3
|
7,9
|
| Ирга. листья
|
0
|
| Калина.плоды
|
0
|
| Ябл: листья
|
0,51
|
0,51
|
| облом
|
2,5
|
2,5
|
| опад
|
0
|
| плоды
|
0
|
| Трн: листья
|
0
|
| плоды
|
0
|
| опад
|
0
|
| облом
|
0
|
| Бузина.облом
|
0
|
| Листья
|
0
|
| Береза,облом
|
2
|
2
|
| листья
|
4,9
|
4,9
|
| Окончание табл. 7.2.2.3
|
| 1
|
2
|
3
|
4
|
5
|
6
|
7
|
8
|
9
|
10
|
11
|
| Жст: листья
|
0
|
| плоды
|
0
|
| Крш.листья
|
0
|
| плоды
|
0
|
| облом
|
0
|
| Свид.листья
|
0
|
| опад
|
0
|
| плоды
|
0
|
| Боярышник,лист
|
0
|
| Бересклет.лист
|
0
|
| Береза.лист
|
0
|
| Ильм: семена
|
7,1
|
46,04
|
0,01
|
53,15
|
| опад
|
2,45
|
2,45
|
| кора
|
4,2
|
4,2
|
| листья
|
14,8
|
126,75
|
0,96
|
142,51
|
| Берест,листья
|
4,3
|
4,3
|
| Ясп: семена
|
0
|
| Акация б.семена
|
0
|
| Акация ж.семена
|
0
|
| Омела,плоды
|
5,4
|
1,6
|
0,22
|
1,8
|
0,2
|
9,22
|
| ветки
|
0,6
|
11,02
|
6
|
17,62
|
| листья
|
13,19
|
25,6
|
0,86
|
2,53
|
42,18
|
| Травян. опад
|
0,4
|
0,31
|
0,3
|
5,62
|
0,8
|
1,43
|
8,86
|
| Лишайники
|
8,3
|
0,02
|
8,32
|
| Галлы
|
0,02
|
0,2
|
0,22
|
| Животн. опад
|
0,9
|
17,3
|
17,5
|
21,3
|
57
|
| Насекомые
|
6,9
|
0,9
|
8
|
4,4
|
0,3
|
0,8
|
21,3
|
| Грибы
|
0
|
| Экскременты
|
0
|
| насекомых
|
0
|
| животных
|
12
|
0,07
|
12,07
|
| Итого
|
64
|
164,24
|
219,2
|
117,1
|
111,82
|
262,84
|
1558,09
|
68,78
|
26,96
|
2587,63
|
| Всего
|
97,1
|
220,86
|
301,3
|
491,5
|
134,18
|
341,66
|
1807,09
|
138,12
|
47,24
|
3573,65
|
Распределение опада по основным фракциям приведено в таблице 7.2.2.4.
Таблица 7.2.2.4.
Количество опада ( кг./га ) на стационаре участка «Лес на Ворскле» за 2004 г. по основным фракциям.
| Фракции
|
Количество (кг./га.)
|
% от валового опада
|
| Валовой опад
|
3573,65
|
100
|
| Листья дуба
|
306,57
|
8,57
|
| Ветки и кора дуба
|
659,67
|
18,46
|
| Прочее по дубу
|
19,78
|
0,55
|
| Листья прочих древесно-кустарниковых пород
|
1866
|
52,22
|
| Прочий древесно-кустарни- ковый опад (без листьев)
|
381,6
|
10,68
|
| Прочий опад
|
340,03
|
9,52
|
Общее количество опада на стационаре «Леса на Ворскле» существенно превышает таковой на Ямском участке, но в составе фракций преобладают листья прочих древесно-кустарниковых пород. В целом опад прочих древесно-кустарниковых пород составляет почти 63%. Из прочих пород львиная доля опада приходится на клен остролистный, остальная часть примерно поровну распределена между липой мелколистной, осиной и ильмом. На долю дуба приходится менее трети валового опада.
7.2.2.2. Структура и динамика лесных сообществ участка «Лес на Ворскле»
В 2004 г. начаты работы на лесотипологических постоянных пробных площадях (ППП) заложенных в 1963-76 гг., с сохранением методики предыдущих исследований (Проект организации …Т.I, 1976-1977). В сентябре-октябре 2004 г. в кв. 10 на ППП-7, -8, -12 (каждая ППП площадью 1 га – 100х100 м) проводилась сплошная перечислительная таксация (древостоя, подроста и подлеска). Учёты велись по секциям (0,25 га), на которые разбиты ППП.
Диаметры и высоты деревьев определялись мерной вилкой. При сплошном перечёте учитывались следующие показатели: 1). Номер дерева (если номер 1976 г. сохранился); 2). Диаметр ствола (см) на высоте груди (измерялся в двух взаимно-перпендикулярных направлениях). 3). Высота ствола (м) (выборочно, по 10-15 экземпляров из разных ступеней толщины; остальные деревья, подрост и кусты визуально или с помощью мерной вилки, если их высота не превышала 80 см).
На ППП определялось санитарное состояние каждого дерева, при этом отмечалось:
1). Наличие водяных побегов и комлевой поросли. 2). Раковые заболевания ствола. 3). Морозобойные трещины. 4). Дупла и др. нарушения ствола (сломанные вершины и скелетные ветки, суховершинность, фаутность и т.п.). 5). Отмечались сухие деревья, упавшие стволы (их длина на почве), пеньки – их диаметры и высоты.
Камеральная обработка производилась отдельно по каждой секции и в целом по всей ППП. К древостою относились стволы диаметром 4 см и более. Обработка проводилась отдельно по ярусам, а в пределах ярусов по отдельным древесным породам. Яруса выделялись на графике высот согласно распределения количества стволов по метровым ступеням высоты. Ступени толщины для I и II ярусов приняты 4 см, для III яруса – 2 см. В камеральных условиях определялся средний таксационный диаметр древостоя через сумму площадей сечений стволов, делённую на количество стволов. Далее на графике высот по каждой породе и вычесленному среднему диаметру определялась средняя высота древесной породы в ярусе. Затем для каждой ступени толщины и соответствующей высоты устанавливался разряд высот (бонитет) по каждой породе яруса (Логутов и др., 1959). После чего по объёмным таблицам при среднем коэффициенте формы определялся запас древостоя.
При начислении запасов использовались объёмные таблицы следующих авторов:
по дубу – проф. Б.А. Шустова (Тюрин и др.,1946);
по ясеню, клёну, липе, ильму – проф. Ф.П. Моисеенко (Логутов и др., 1959).
Далее по средней высоте и сумме площадей сечений определяли полноту отдельной породы яруса (Третьяков и др., 1965).
Результаты исследований и расчётов приведены в табл. 7.2.2.5 – 7.2.2.16. Для сравнительного анализа динамики структуры исследованных древостоев приведены данные сплошной перечислительной таксации тех же ППП в 1975-76 гг. (табл. 7.2.2.7, 7.2.2.11, 7.2.2.15) (Проект организации …Т.IV, Кн. VI., 1977).
В таблицах приняты следующие сокращения: Дуб – дуб черешчатый (Quercus robur); Клён о. – клён остролистный или платановидный (Acer platanoides); Клён п. – клён полевой или равнинный (Acer campestre); Липа – липа сердцелистная или мелколистная (Tilia cordata); Ильм – ильм голый или шершавый (Ulmus glabra Huds. (U. scabra Mill.)), Груша – груша обыкновенная (Pyrus communis), Ясень – ясень обыкновенный (Fraxinus excelsior), Берес. евр. – бересклет европейский (Euonymus europaea), Берес. бор. – бересклет бородавчатый (E. verrucosa), бузина чёр. – бузина чёрная (Sambucus nigra) (Определитель сосудистых …1995).
Таблица 7.2.2.5
Лесотаксационная характеристика постоянной пробной площади №7 (1.0 га) в 2004 году
| Порода
|
I ярус
|
| Ступени
толщины,
см
|
Высота,
м
|
Число сырорастущих деревьев по классам роста
|
Число
сухост.
дерев.,
шт
|
Сумма
площ. сечений, м2
|
Объём
одного
ствола,
м3
|
Запас стволов.
древесины,
м3
|
| I
|
II
|
III
|
IV
|
итого
|
растущ.
леса
|
сухост.
леса
|
растущ.
леса
|
сухост.
леса
|
| 1
|
2
|
3
|
4
|
5
|
6
|
7
|
8
|
9
|
10
|
11
|
12
|
13
|
14
|
| Дуб
|
24
|
25,3
|
5
|
5
|
0,226
|
0,54
|
2,7
|
| 28
|
27,2
|
10
|
10
|
0,161
|
0,78
|
7,8
|
| 32
|
26,8
|
8
|
8
|
0,,64
|
1,01
|
8,08
|
| 32
|
27
|
1
|
0,08
|
1,01
|
1,01
|
| 36
|
27,5
|
7
|
7
|
0,71
|
1,3
|
9,12
|
| 36
|
24
|
1
|
0,102
|
1,15
|
1,15
|
| 40
|
38,6
|
24
|
24
|
3,02
|
1,66
|
39,8
|
| 44
|
28,8
|
16
|
16
|
2,43
|
2,02
|
32,32
|
| 48
|
28,8
|
22
|
22
|
3,98
|
2,4
|
52,8
|
| 52
|
29,5
|
7
|
7
|
1,49
|
2,89
|
20,23
|
| 56
|
29,5
|
11
|
11
|
2,72
|
3,33
|
36,63
|
| 60
|
29,3
|
5
|
5
|
1,41
|
3,8
|
19,00
|
| 64
|
29
|
4
|
4
|
1,29
|
4,3
|
17,2
|
| Итого на пробе
|
119
|
119
|
2
|
18,52
|
0,18
|
245,68
|
2,16
|
| Окр.
|
119
|
119
|
2
|
18,5
|
0,2
|
246
|
2,2
|
| Клен.о
|
12
|
21,1
|
13
|
13
|
0,147
|
0,09
|
1,17
|
| 16
|
22,6
|
15
|
15
|
0,302
|
0,17
|
2,55
|
| 16
|
23
|
1
|
0,020
|
0,17
|
0,17
|
| 20
|
24
|
18
|
18
|
0,566
|
0,3
|
5,4
|
| 24
|
25,5
|
16
|
16
|
0,724
|
0,47
|
7,52
|
| 28
|
27,3
|
14
|
14
|
0,862
|
0,68
|
9,52
|
| 32
|
28,1
|
17
|
17
|
1,3
|
0,94
|
15,98
|
| 36
|
28,9
|
10
|
10
|
1,02
|
1,26
|
12,6
|
Таблица 7.2.2.5 (продолжение)
Лесотаксационная характеристика постоянной пробной площади №7 (1.0 га) в 2004 году
| Порода
|
I ярус
|
| Ступени
толщины,
см
|
Высота,
м
|
Число сырорастущих деревьев по классам роста
|
Число
сухост.
дерев.,
шт
|
Сумма
площ. сечений, м2
|
Объём
одного
ствола,
м3
|
Запас стволов.
Древесины,
м3
|
| I
|
II
|
III
|
IV
|
итого
|
растущ.
леса
|
сухост.
леса
|
растущ.
леса
|
сухост.
леса
|
| 1
|
2
|
3
|
4
|
5
|
6
|
7
|
8
|
9
|
10
|
11
|
12
|
13
|
14
|
| 40
|
28,8
|
13
|
13
|
1,63
|
1,61
|
20,93
|
| 44
|
29,3
|
6
|
6
|
0,914
|
2,02
|
12,12
|
| 48
|
29,7
|
3
|
3
|
0,541
|
2,47
|
7,41
|
| 52
|
29,7
|
5
|
5
|
1,06
|
2,97
|
14,85
|
| Итого на пробе
|
130
|
130
|
1
|
9,13
|
0,02
|
110,05
|
0,17
|
| Окр.
|
130
|
130
|
1
|
9
|
0,02
|
110
|
0,2
|
| Липа
|
12
|
21
|
8
|
8
|
0,09
|
0,1
|
0,8
|
| 16
|
21,8
|
6
|
6
|
0,12
|
0,2
|
1,2
|
| 16
|
20
|
1
|
0,02
|
0,2
|
0,2
|
| 20
|
25,2
|
13
|
13
|
0,409
|
0,34
|
4,42
|
| 24
|
26
|
13
|
13
|
0,588
|
0,53
|
6,89
|
| 28
|
26,5
|
6
|
6
|
0,37
|
0,78
|
4,68
|
| 32
|
26,8
|
14
|
14
|
1,12
|
1,06
|
14,84
|
| 36
|
28,3
|
6
|
6
|
0,612
|
1,36
|
8,16
|
| 40
|
28
|
10
|
10
|
1,252
|
1,66
|
16,6
|
| 44
|
28,5
|
2
|
2
|
0,304
|
2,03
|
4,06
|
| 48
|
29
|
1
|
1
|
0,181
|
2,47
|
2,47
|
| 52
|
28,5
|
1
|
1
|
0,21
|
2,93
|
2,93
|
| 56
|
29,5
|
1
|
1
|
0,25
|
3,42
|
3,42
|
| Итого на пробе
|
81
|
81
|
1
|
5,506
|
70,47
|
0,2
|
| Окр.
|
81
|
1
|
5,5
|
0,02
|
70,5
|
0,2
|
Таблица 7.2.2.5 (продолжение)
Лесотаксационная характеристика постоянной пробной площади №7 (1.0 га) в 2004 году
| Порода
|
I ярус
|
| Ступени
толщины,
см
|
Высота,
м
|
Число сырорастущих деревьев по классам роста
|
Число
сухост.
дерев.,
шт
|
Сумма
площ. сечений, м2
|
Объём
одного
ствола,
м3
|
Запас стволов.
Древесины,
м3
|
| I
|
II
|
III
|
IV
|
итого
|
растущ.
леса
|
сухост.
леса
|
растущ.
леса
|
сухост.
леса
|
| 1
|
2
|
3
|
4
|
5
|
6
|
7
|
8
|
9
|
10
|
11
|
12
|
13
|
14
|
| Ильм
|
8
|
21
|
1
|
1
|
0,02
|
0,03
|
0,03
|
| 12
|
22
|
1
|
1
|
0,011
|
0,09
|
0,09
|
| 16
|
21,5
|
3
|
3
|
0,06
|
0,17
|
0,51
|
| 16
|
20
|
1
|
0,02
|
0,17
|
0,17
|
| 20
|
24
|
1
|
1
|
0,031
|
0,3
|
0,3
|
| 20
|
26
|
1
|
0,031
|
0,3
|
0,3
|
| 24
|
23
|
1
|
1
|
0,045
|
0,47
|
0,47
|
| 24
|
21
|
1
|
0,045
|
0,47
|
0,47
|
| 36
|
27,5
|
1
|
1
|
0,102
|
1,25
|
1,25
|
| 44
|
38
|
1
|
1
|
0,152
|
2,02
|
2,02
|
| Итого на пробе
|
9
|
3
|
0,406
|
0,096
|
4,67
|
0,94
|
| Окр.
|
9
|
3
|
0,4
|
0,1
|
4,7
|
1,0
|
| Всего на пробе
|
339
|
7
|
33,562
|
0,316
|
430,87
|
3,47
|
| Окр.
|
339
|
7
|
33,6
|
0,3
|
430,9
|
3,5
|
Таблица 7.2.2.5 (продолжение)
Лесотаксационная характеристика постоянной пробной площади №7 (1.0 га) в 2004 году
| Порода
|
II ярус
|
III ярус
|
| Ступе-ни
толщ.,
см /
кл.роста
|
Выс-ота,
м
|
Число дер.,
шт
|
Сумма площ.
сечений,
м2
|
Объём
одного
ствола,
м3
|
Запас ствол.
древесины,
м3
|
Ступени
толщины,
см /
кл.роста
|
Высо-
та,
м
|
Число дер.,
шт
|
Сумма площ.
сечений,
м2
|
Объём
одного
ствола,
м3
|
Запас ствол.
древесины,
м3
|
| раст.
|
сух.
|
раст.
|
сух.
|
раст.
|
сух.
|
раст.
|
сух.
|
раст.
|
сух.
|
раст.
|
сух.
|
| 1
|
15
|
16
|
17
|
18
|
19
|
20
|
21
|
22
|
23
|
24
|
25
|
26
|
27
|
28
|
29
|
30
|
31
|
32
|
| Дуб
|
40
|
15
|
1
|
0,126
|
0,97
|
0,97
|
4
|
5,7
|
1
|
0,001
|
0,00053
|
0,0053
|
| 24
|
6,5
|
1
|
0,045
|
0,16
|
0,16
|
| 28
|
4,2
|
2
|
0,123
|
0,11
|
0,11
|
| 36
|
1,7
|
1
|
0,102
|
0,09
|
0,09
|
| Итого на пробе
|
1
|
0,126
|
0,97
|
5
|
0,271
|
0,37
|
| Окр.
|
1
|
0,1
|
1,0
|
5
|
0,3
|
0,4
|
| Клен.о
|
4
|
12
|
1
|
0,001
|
0,014
|
0,014
|
4
|
6,7
|
51
|
0,066
|
0,012
|
0,612
|
| 6
|
14
|
15
|
0,042
|
0,02
|
0,30
|
4
|
7,4
|
3
|
0,016
|
0,009
|
0,027
|
| 6
|
13
|
1
|
0,003
|
0,02
|
0,02
|
6
|
7,7
|
50
|
0,162
|
0,015
|
0,75
|
| 8
|
15
|
47
|
0,236
|
0,03
|
1,41
|
6
|
7,5
|
2
|
0,006
|
0,023
|
0,046
|
| 8
|
14
|
1
|
0,005
|
0,03
|
0,03
|
8
|
9,8
|
29
|
0,145
|
0,02
|
0,58
|
| 12
|
16,2
|
32
|
0,362
|
0,09
|
2,8
|
8
|
8
|
2
|
0,01
|
0,02
|
0,04
|
| 12
|
12
|
1
|
0,011
|
0,06
|
0,06
|
12
|
9
|
6
|
0,068
|
0,06
|
0,36
|
| 16
|
18
|
5
|
0,1
|
0,17
|
0,85
|
12
|
2,6
|
2
|
0,022
|
0,019
|
0,038
|
| 20
|
5,5
|
1
|
0,031
|
0,08
|
0,08
|
| Всего на пробе
|
100
|
3
|
0,741
|
0,019
|
5,374
|
0,11
|
136
|
10
|
0,441
|
0,085
|
2,3
|
0,231
|
| Окр.
|
100
|
3
|
0,7
|
0,02
|
5,4
|
0,1
|
136
|
10
|
0,4
|
0,09
|
2,3
|
0,2
|
Таблица 7.2.2.5 (окончание)
Лесотаксационная характеристика постоянной пробной площади №7 (1.0 га) в 2004 году
| Поро-да
|
II ярус
|
III ярус
|
| Ступе-ни
толщ.,
см /
кл.роста
|
Выс-ота,
м
|
Число дер.,
шт
|
Сумма площ.
сечений,
м2
|
Объём
одного
ствола,
м3
|
Запас ствол.
древесины,
м3
|
Ступени
толщины,
см /
кл.роста
|
Высо-
та,
м
|
Число дер.,
шт
|
Сумма площ.
сечений,
м2
|
Объём
одного
ствола,
м3
|
Запас ствол.
древесины,
м3
|
| раст.
|
сух.
|
раст.
|
сух.
|
раст.
|
сух.
|
раст.
|
сух.
|
раст.
|
сух.
|
раст.
|
сух.
|
| 1
|
15
|
16
|
17
|
18
|
19
|
20
|
21
|
22
|
23
|
24
|
25
|
26
|
27
|
28
|
29
|
30
|
31
|
32
|
| Липа
|
8
|
14,4
|
8
|
0,04
|
0,03
|
0,24
|
4
|
3,7
|
5
|
0,007
|
0,006
|
0,03
|
| 10
|
12
|
1
|
0,008
|
0,06
|
0,06
|
6
|
4,9
|
4
|
0,012
|
0,016
|
0,06
|
| 12
|
16
|
7
|
0,079
|
0,1
|
0,7
|
6
|
6,5
|
1
|
0,003
|
0,014
|
0,014
|
| 16
|
19
|
2
|
0,04
|
0,19
|
0,38
|
8
|
7,5
|
8
|
0,04
|
0,02
|
0,16
|
| 20
|
16
|
1
|
0,031
|
0,25
|
0,25
|
8
|
4
|
1
|
0,005
|
0,01
|
0,01
|
| 10
|
6,5
|
3
|
0,024
|
0,028
|
0,084
|
| 10
|
10
|
1
|
0,008
|
0,03
|
0,03
|
| 28
|
3,7
|
1
|
0,062
|
0,11
|
0,11
|
| Всего на пробе
|
18
|
1
|
0,17
|
0,031
|
1,38
|
0,25
|
20
|
4
|
0,083
|
0,078
|
0,334
|
0,164
|
| Окр.
|
18
|
1
|
0,2
|
0,03
|
1,4
|
0,3
|
20
|
4
|
0,08
|
0,08
|
0,3
|
0,2
|
| Ильм
|
8
|
13,3
|
5
|
0,025
|
0,03
|
0,15
|
4
|
5,7
|
6
|
0,008
|
0,015
|
0,09
|
| 12
|
15
|
10
|
0,113
|
0,084
|
0,84
|
4
|
6,5
|
4
|
0,005
|
0,015
|
0,06
|
| 12
|
13
|
1
|
0,011
|
0,07
|
0,07
|
6
|
7,3
|
18
|
0,051
|
0,015
|
0,27
|
| 16
|
15
|
2
|
0,04
|
0,15
|
0,3
|
6
|
8,2
|
4
|
0,012
|
0,0127
|
0,051
|
| 8
|
8,9
|
11
|
0,055
|
0,02
|
0,22
|
| 8
|
8,4
|
5
|
0,021
|
0,02
|
0,1
|
| 10
|
8
|
1
|
0,008
|
0,04
|
0,04
|
| 12
|
10
|
1
|
0,011
|
0,06
|
0,06
|
| 12
|
4,5
|
1
|
0,011
|
0,02
|
0,02
|
| 20
|
5
|
1
|
0,031
|
0,07
|
0,07
|
| Всего на пробе
|
17
|
1
|
0,178
|
0,011
|
1,29
|
0,07
|
37
|
15
|
0,133
|
0,08
|
0,68
|
0,301
|
| Окр.
|
17
|
1
|
0,0
|
0,01
|
1,3
|
0,1
|
37
|
15
|
0,1
|
0,1
|
0,7
|
0,3
|
| Всего на пробе
|
135
|
7
|
1,089
|
0,187
|
8,044
|
1,4
|
193
|
34
|
0,657
|
0,514
|
3,314
|
1,066
|
| Окр.
|
135
|
7
|
1,1
|
0,2
|
8,0
|
1,4
|
193
|
34
|
0,7
|
0,5
|
3,3
|
1,1
|
Таблица 7.2.2.6
Основные итоги обработки постоянной пробной площади № 7 (в переводе на 1 га) в 2004 г.
ГПЗ «Белогорье», уч. «Лес на Ворскле», кв. 10 , выд. 6
Величина пробной площади 1,0
га
Происхождение: естественное семенное и порослевое
| Ярус
|
Древесн.
входящ.
в ярус
|
Таксационные показатели
|
| Сос-
тав
|
Средние
|
Бони-
тет
|
Число
ствол.
шт
|
Сум-
ма
площ.
сече-
ния,
м2
|
Пол-
нота
|
Запас
ствол., м3
|
Прирост, м3
|
| воз-раст,
лет
|
Высо-
та,
м
|
диам.,
см
|
раст.
леса
|
сух.
леса
|
сред-
ний
|
теку-
щий
|
| 1
|
2
|
3
|
4
|
5
|
6
|
7
|
8
|
9
|
10
|
11
|
12
|
13
|
14
|
| I
|
Дуб
|
6
|
108
|
28,2
|
44
|
I
|
119
|
18,5
|
0,52
|
246
|
2,2
|
| Липа
|
1
|
26,6
|
34
|
81
|
5,5
|
0,11
|
70,5
|
0,2
|
| Клен.о
|
3
|
26,8
|
32
|
130
|
9,0
|
0,26
|
110
|
0,2
|
| Ильм
|
25,3
|
23
|
9
|
0,4
|
0,01
|
4,7
|
1,0
|
| Итого
|
10
|
108
|
I
|
339
|
33,4
|
0,9
|
431,2
|
3,6
|
| II
|
Дуб
|
15
|
40
|
1
|
0,1
|
0,004
|
1,0
|
| Липа
|
2
|
15,4
|
12,0
|
18
|
0,2
|
0,008
|
1,4
|
0,3
|
| Клен.о
|
7
|
15
|
9,0
|
100
|
0,7
|
0,03
|
5,4
|
0,1
|
| Ильм
|
1
|
14,4
|
12
|
17
|
0,2
|
0,009
|
1,3
|
0,1
|
| Итого
|
10
|
78
|
136
|
1,2
|
0,05
|
8,1
|
1,5
|
| III
|
Дуб
|
| Липа
|
1
|
5,7
|
7
|
20
|
0,08
|
0,006
|
0,3
|
0,2
|
| Клен.о
|
7
|
8,3
|
8
|
136
|
0,4
|
0,03
|
2,3
|
0,2
|
| Ильм
|
2
|
8
|
8
|
37
|
0,1
|
0,007
|
0,7
|
0,3
|
| Итого
|
10
|
193
|
0,58
|
0,04
|
3,3
|
0,7
|
| Всего
|
I
|
668
|
35,2
|
0,99
|
442,6
|
5,8
|
Фаутность и прочие особенности: Насаждение в удовлетвор. состоянии. Поперечный надлом. рак – единично (~1%); суховершинность, сухие ветви первой величины и в кроне – 6% слабое; дупла, морозобойные трещины, коровые повреждения ствола – 2% слабое; водяные побеги – 73% сильное; снеговые заломы – 3% слабое.
Пдр. Кл.о., Ил, Лп Пдл. Ил, Кл.о., бересклет евр. (см. табл 7.2.2.8)
Пкр. Сныть. Почва: тёмно-серая лесная средне оподз.
Подпочва: Лессовидная, суглинок. Рельеф и положение: слабый склон СЗ эксп.
Тип лесораст. условий: Д2 Тип леса: клёно-дубняк снытьевый
Дата перечёта 29 октября - 5 ноября 2004 г.
Обработал: Немченко В.А., Немченко М.Ю.
Таблица 7.2.2.7
Основные итоги обработки постоянной пробной площади № 7 (в переводе на 1 га) в 1976 г.
ГПЗ «Белогорье», уч. «Лес на Ворскле», кв. 10 , выд. 6.
Величина пробной площади 1,0
га
Происхождение: естественное семенное и порослевое
| Ярус
|
Древесн.
входящ.
в ярус
|
Таксационные показатели
|
| Сос-
тав
|
Средние
|
Бони-
тет
|
Число
ствол.
шт
|
Сум-
ма
площ.
сече-
ния,
м2
|
Пол-
нота
|
Запас
ствол., м3
|
Прирост, м3
|
| воз-раст,
лет
|
Высо-
та,
м
|
диам.,
см
|
раст.
леса
|
сух.
леса
|
сред-
ний
|
теку-
щий
|
| 1
|
2
|
3
|
4
|
5
|
6
|
7
|
8
|
9
|
10
|
11
|
12
|
13
|
14
|
| I
|
Дуб
|
8
|
80
|
24,9
|
35,7
|
I
|
182
|
18,4
|
0,54
|
215
|
21
|
| Липа
|
1
|
22,2
|
27,3
|
48
|
2,8
|
0,08
|
30
|
| Клен.о
|
1
|
23,5
|
26,5
|
39
|
2,2
|
0,06
|
23
|
| Ильм
|
2
|
0,1
|
2
|
| Итого
|
10
|
80
|
I
|
271
|
23,5
|
0,68
|
270
|
21
|
3,4
|
| II
|
Дуб
|
4
|
0,1
|
2
|
| Липа
|
3
|
17,0
|
17,9
|
53
|
1,3
|
0,04
|
11
|
| Клен.о
|
5
|
18,2
|
18,4
|
78
|
2,1
|
0,08
|
18
|
| Ильм
|
2
|
18,6
|
20,0
|
31
|
1,0
|
0,03
|
9
|
| Груша
|
3
|
0,1
|
| Итого
|
10
|
50
|
169
|
4,6
|
0,15
|
40
|
0,8
|
| Всего
|
I
|
440
|
28,1
|
0,83
|
310
|
21
|
4,2
|
5,0
|
Фаутность и прочие особенности: Насаждение в удовлетв. состоянии. Корневая гниль от опенка 10%;
поперечный надлом. рак – 5 % слабое
Пдр._Кл, Лп, Ил / 5 – 10 / Пдл. Груша, бересклет евр, клен полев. – единично.
Пкр. Осока, сныть. Почва: темно-серая лесная, средне оподзол.
Подпочва: Лессовидная, суглинок. Рельеф и положение: слабый склон СЗ эксп.
Тип лесораст. условий: Д2 Коренной тип леса: Л – 1б (липо-дубняк осоково-снытьевый)
Дата перечёта: 12 июля 1976 г.
Начальник л/у партии: Прозоров В.В.
Инженер: Миронова, Фоминых
Таблица 7.2.2.8
Учёт подроста и подлеска на постоянной пробной площади № 7 (1 га)
в 2004 г.*
| Высота, м
“
Д
|
Древесные и кустарниковые породы, шт / га
|
| Клён остр.
|
Ильм
|
Липа
|
Ясень
|
Груша
|
Берес. евр.
|
Берес. бор.
|
Бузина чёр.
|
Итого:
|
| до 0,5 1
|
84
|
195
|
4
|
2
|
-
|
524
|
-
|
-
|
809
|
| до 2 -
|
143
|
258
|
29
|
-
|
1
|
6
|
5
|
1
|
443
|
| свыше 2 -
|
354
|
93
|
12
|
-
|
-
|
-
|
1
|
-
|
460
|
| Итого: 1
|
581
|
546
|
45
|
2
|
1
|
530
|
6
|
1
|
| Всего:
|
1712+1
|
Примечание: * подрост и подлесок повреждается дикими животными (особенно высотой до 0,5 м).
“ опечатка - на ППП №7 обнаружен 1 дуб высотой менее 0,5 м.
в 1976 г.*
| Высота,
м
|
Древесные и кустарниковые породы, тыс. шт / га
|
Итого:
|
| Клён остр.
|
Липа
|
Ильмовые
|
Дуб
|
| до 5 лет
|
до 10 лет
|
бол.10 л.
|
до 5 лет
|
до 10 лет
|
бол.10 л.
|
до 5 лет
|
до 10 лет
|
бол.10 л.
|
до 5 лет
|
| до 0,5
|
2,1
|
-
|
-
|
-
|
-
|
-
|
0,1
|
-
|
-
|
57,4
|
59,6
|
| до 2
|
1,2
|
-
|
-
|
-
|
0,8
|
-
|
-
|
0,1
|
1,3
|
-
|
3,4
|
| свыше 2
|
-
|
-
|
1,8
|
-
|
-
|
0,1
|
-
|
-
|
-
|
-
|
1,9
|
| Итого:
|
3,3
|
-
|
1,8
|
-
|
0,8
|
0,1
|
0,1
|
0,1
|
1,3
|
57,4
|
| Всего:
|
64,9
|
Примечание: * подрост дуба (всходы) поражён мучнистой росой, неблагонадёжный;
подрост и подлесок повреждается дикими животными.
Таблица 7.2.2.9
Лесотаксационная характеристика постоянной пробной площади № 8 (1.0 га) в 2004 году
| Порода
|
I ярус
|
| Ступени
толщины,
см
|
Высота,
м
|
Число сырорастущих деревьев по классам роста
|
Число
сухост.
дерев.,
шт
|
Сумма
площ. сечений, м2
|
Объём
одного
ствола,
м3
|
Запас стволов.
Древесины,
м3
|
| I
|
II
|
III
|
IV
|
итого
|
растущ.
леса
|
сухост.
леса
|
растущ.
леса
|
сухост.
леса
|
| 1
|
2
|
3
|
4
|
5
|
6
|
7
|
8
|
9
|
10
|
11
|
12
|
13
|
14
|
| Дуб
|
20
|
28
|
1
|
1
|
0,031
|
0,41
|
0,41
|
| 36
|
28
|
1
|
1
|
0,0102
|
1,32
|
1,32
|
| 40
|
29
|
1
|
1
|
0,126
|
1,68
|
1,68
|
| 48
|
29
|
1
|
1
|
0,181
|
2,42
|
2,42
|
| 60
|
31
|
1
|
1
|
0,28
|
4,01
|
4,01
|
| 76
|
29
|
1
|
1
|
0,45
|
6,06
|
6,06
|
| 96
|
34
|
1
|
1
|
0,72
|
11,15
|
11,15
|
| 100
|
33
|
2
|
2
|
1,57
|
11,77
|
23,54
|
| 104
|
33,8
|
5
|
5
|
4,25
|
13,02
|
65,12
|
| 108
|
33,4
|
5
|
5
|
4,58
|
13,89
|
69,45
|
| 112
|
33,7
|
3
|
3
|
2,96
|
15,03
|
45,09
|
| 120
|
32
|
1
|
1
|
1,13
|
16,50
|
16,50
|
| 136
|
34
|
1
|
1
|
1,45
|
22,37
|
22,37
|
| Итого на пробе (1 га)
|
24
|
24
|
17,83
|
269,12
|
| Окр.
|
24
|
17,8
|
269
|
| Липа
|
24
|
24,7
|
3
|
3
|
0,136
|
0,53
|
1,59
|
| 36
|
31
|
2
|
2
|
0,20
|
1,36
|
2,72
|
| 40
|
29,3
|
3
|
3
|
0,38
|
1,66
|
4,98
|
| 44
|
25
|
1
|
1
|
0,152
|
1,85
|
1,85
|
| 48
|
26,3
|
5
|
5
|
0,903
|
2,266
|
11,33
|
| 52
|
28,8
|
6
|
6
|
1,27
|
2,88
|
17,3
|
| 56
|
28
|
4
|
4
|
0,98
|
3,265
|
13,06
|
| 60
|
28,8
|
6
|
6
|
1,70
|
3,95
|
23,7
|
Таблица 7.2.2.9 (продолжение)
Лесотаксационная характеристика постоянной пробной площади № 8 (1.0 га) в 2004 году
| Порода
|
I ярус
|
| Ступени
толщины,
см
|
Высота,
м
|
Число сырорастущих деревьев по классам роста
|
Число
сухост.
дерев.,
шт
|
Сумма
площ. сечений, м2
|
Объём
одного
ствола,
м3
|
Запас стволов.
Древесины,
м3
|
| I
|
II
|
III
|
IV
|
итого
|
растущ.
леса
|
сухост.
леса
|
растущ.
леса
|
сухост.
леса
|
| 1
|
2
|
3
|
4
|
5
|
6
|
7
|
8
|
9
|
10
|
11
|
12
|
13
|
14
|
| Липа
|
64
|
31
|
1
|
1
|
0,32
|
4,53
|
4,53
|
| 68
|
28,5
|
2
|
2
|
0,72
|
4,91
|
9,82
|
| 76
|
30,3
|
2
|
2
|
0,9
|
6,47
|
12,94
|
| 80
|
31
|
2
|
2
|
1,01
|
7,16
|
14,32
|
| 84
|
30
|
1
|
1
|
0,55
|
7,84
|
7,84
|
| Итого на пробе (1га)
|
38
|
38
|
9,3
|
126,0
|
| Окр .
|
38
|
9
|
126
|
| Кл. о.
|
8
|
22
|
2
|
2
|
0,01
|
0,03
|
0,06
|
| 12
|
23,5
|
12
|
12
|
1
|
0,136
|
0,011
|
0,09
|
1,08
|
0,09
|
| 16
|
24,2
|
20
|
20
|
0,403
|
0,17
|
3,4
|
| 20
|
25,5
|
34
|
34
|
1,068
|
0,30
|
10,2
|
| 20
|
25
|
1
|
0,031
|
0,30
|
0,30
|
| 24
|
27,3
|
46
|
46
|
2,081
|
0,47
|
21,62
|
| 28
|
27,4
|
21
|
21
|
1,293
|
0,68
|
14,28
|
| 32
|
28,3
|
23
|
23
|
1,85
|
0,94
|
21,62
|
| 36
|
29,9
|
8
|
8
|
0,812
|
1,25
|
10,0
|
| 36
|
31
|
1
|
0,102
|
1,25
|
1,25
|
| 40
|
29,8
|
4
|
4
|
0,502
|
1,61
|
6,44
|
| 44
|
29,3
|
4
|
4
|
0,604
|
2,02
|
8,08
|
| 56
|
28
|
1
|
1
|
0,25
|
3,13
|
3,13
|
| 60
|
29
|
1
|
1
|
0,28
|
4,05
|
4,05
|
| 64
|
30
|
1
|
1
|
0,32
|
4,64
|
4,64
|
| Итого на пробе (1га)
|
177
|
177
|
3
|
9,609
|
0,144
|
108,6
|
1,64
|
| Окр
|
177
|
3
|
9,61
|
0,14
|
109
|
1,6
|
Таблица 7.2.2.9 (продолжение)
Лесотаксационная характеристика постоянной пробной площади № 8 (1.0 га) в 2004 году
| Порода
|
I ярус
|
| Ступени
толщины,
см
|
Высота,
м
|
Число сырорастущих деревьев по классам роста
|
Число
сухост.
дерев.,
шт
|
Сумма
площ. сечений, м2
|
Объём
одного
ствола,
м3
|
Запас стволов.
Древесины,
м3
|
| I
|
II
|
III
|
IV
|
итого
|
растущ.
леса
|
сухост.
леса
|
растущ.
леса
|
сухост.
леса
|
| 1
|
2
|
3
|
4
|
5
|
6
|
7
|
8
|
9
|
10
|
11
|
12
|
13
|
14
|
| Ильм
|
12
|
25
|
1
|
1
|
0,011
|
0,09
|
0,09
|
| 16
|
22,5
|
2
|
2
|
0,04
|
0,17
|
0,34
|
| 16
|
23
|
1
|
0,02
|
0,17
|
0,17
|
| 20
|
24,5
|
4
|
4
|
0,125
|
0,30
|
1,2
|
| 24
|
27,2
|
10
|
10
|
0,453
|
0,47
|
4,7
|
| 28
|
27
|
7
|
7
|
0,431
|
0,68
|
4,76
|
| 32
|
28,4
|
7
|
7
|
0,56
|
0,94
|
6,58
|
| 40
|
28
|
1
|
1
|
0,126
|
1,61
|
1,61
|
| 44
|
27
|
1
|
1
|
0,152
|
1,81
|
1,81
|
| 48
|
29
|
2
|
2
|
0,36
|
2,47
|
4,94
|
| 64
|
30
|
1
|
1
|
0,32
|
4,64
|
4,64
|
| Итого на пробе (1 га)
|
36
|
1
|
2,578
|
0,02
|
30,67
|
0,17
|
| Окр.
|
36
|
1
|
2,6
|
0,02
|
31
|
0,2
|
| Всего на пробе (1 га)
|
275
|
4
|
39,317
|
0,164
|
534,39
|
1,81
|
| Окр.
|
275
|
4
|
39,0
|
0,16
|
535
|
1,8
|
Таблица 7.2.2.9 (продолжение)
Лесотаксационная характеристика постоянной пробной площади № 8 (1.0 га) в 2004 году
| Порода
|
II ярус
|
III ярус
|
| Ступени
толщ.,
см /
кл.роста
|
Высо-
та,
м
|
Число дер.,
шт
|
Сумма площ.
сечений,
м2
|
Объём
одного
ствола,
м3
|
Запас ствол.
древесины,
м3
|
Ступени
толщины,
см /
кл.роста
|
Высо-
та,
м
|
Число дер.,
шт
|
Сумма площ.
сечений,
м2
|
Объём
одного
ствола,
м3
|
Запас ствол.
древесины,
м3
|
| раст.
|
сух.
|
раст.
|
сух.
|
раст.
|
сух.
|
раст.
|
сух.
|
раст.
|
сух.
|
раст.
|
сух.
|
| 1
|
15
|
16
|
17
|
18
|
19
|
20
|
21
|
22
|
23
|
24
|
25
|
26
|
27
|
28
|
29
|
30
|
31
|
32
|
| Липа
|
20
|
20
|
1
|
0,031
|
0,32
|
0,32
|
| Итого на пробе (1 га)
|
1
|
0,031
|
0,32
|
| Окр.
|
1
|
0,03
|
0,3
|
| Клен о.
|
4
|
16,3
|
3
|
0,004
|
0,031
|
0,04
|
4
|
7,4
|
59
|
0,076
|
0,01
|
0,62
|
| 8
|
15,4
|
52
|
0,26
|
0,03
|
1,56
|
6
|
8,5
|
110
|
0,036
|
0,019
|
2,04
|
| 12
|
17,7
|
52
|
0,59
|
0,088
|
4,56
|
6
|
7,4
|
6
|
0,017
|
0,015
|
0,09
|
| 16
|
18,9
|
19
|
0,383
|
0,222
|
4,23
|
8
|
10,3
|
30
|
0,151
|
0,026
|
0,77
|
| 20
|
19,2
|
5
|
0,156
|
0,30
|
1,50
|
8
|
7
|
1
|
0,005
|
0,03
|
0,03
|
| 24
|
19
|
4
|
0,181
|
0,42
|
1,68
|
10
|
11,5
|
10
|
0,08
|
0,04
|
0,42
|
| 28
|
19
|
1
|
0,062
|
0,56
|
0,56
|
12
|
11,5
|
2
|
0,022
|
0,06
|
0,12
|
| 12
|
3
|
2
|
0,023
|
0,01
|
0,02
|
| 14
|
1,8
|
1
|
0,015
|
0,10
|
0,10
|
| 24
|
6
|
1
|
0,45
|
0,01
|
0,01
|
| 36
|
1
|
1
|
0,102
|
0,05
|
0,05
|
| 54
|
8
|
1
|
0,230
|
0,88
|
0,88
|
| Итого на пробе (1 га)
|
136
|
1,636
|
14,13
|
211
|
13
|
0,635
|
0,437
|
3,97
|
1,18
|
| Окр.
|
136
|
1,6
|
14,1
|
211
|
13
|
0,6
|
0,4
|
4
|
1,2
|
Таблица 7.2.2.9 (окончание)
Лесотаксационная характеристика постоянной пробной площади №7 (1.0 га) в 2004 году
| Порода
|
II ярус
|
III ярус
|
| Ступени
толщ.,
см /
кл.роста
|
Высо-
та,
м
|
Число дер.,
шт
|
Сумма площ.
сечений,
м2
|
Объём
одного
ствола,
м3
|
Запас ствол.
древесины,
м3
|
Ступени
толщины,
см /
кл.роста
|
Высо-
та,
м
|
Число дер.,
шт
|
Сумма площ.
сечений,
м2
|
Объём
одного
ствола,
м3
|
Запас ствол.
древесины,
м3
|
| раст.
|
сух.
|
раст.
|
сух.
|
раст.
|
сух.
|
раст.
|
сух.
|
раст.
|
сух.
|
раст.
|
сух.
|
| 1
|
15
|
16
|
17
|
18
|
19
|
20
|
21
|
22
|
23
|
24
|
25
|
26
|
27
|
28
|
29
|
30
|
31
|
32
|
| Кл. п.
|
4
|
5
|
1
|
0,001
|
0,003
|
0,003
|
| Итого на пробе (1 га)
|
1
|
0,001
|
0,003
|
| Ильм
|
12
|
15
|
2
|
0,022
|
0,08
|
0,16
|
4
|
5
|
2
|
0,003
|
0,01
|
0,02
|
| 16
|
16,5
|
2
|
0,04
|
0,16
|
0,31
|
6
|
5,8
|
2
|
0,006
|
0,01
|
0,02
|
| Итого на пробе ( 1 га)
|
4
|
0,62
|
0,47
|
4
|
0,009
|
0,04
|
| Окр.
|
4
|
0,06
|
0,5
|
4
|
0,01
|
0,04
|
| Всего на пробе ( 1 га)
|
141
|
1,729
|
14,92
|
216
|
13
|
0,645
|
0,437
|
4,013
|
1,18
|
| Окр.
|
141
|
1,7
|
15
|
216
|
13
|
0,6
|
0,4
|
4,0
|
1,2
|
Таблица 7.2.2.10
Основные итоги обработки постоянной пробной площади № 8 (в переводе на 1 га) в 2004 г.
ГПЗ «Белогорье», уч. «Лес на Ворскле», кв. 10, выд. 10
Величина пробной площади 1,0
га
Происхождение: естественное семенное
| Ярус
|
Древесн.
входящ.
в ярус
|
Таксационные показатели
|
| Сос-
тав
|
Средние
|
Бони-
тет
|
Число
ствол.
шт
|
Сум-
ма
площ.
сече-
ния,
м2
|
Пол-
нота
|
Запас
ствол., м3
|
Прирост, м3
|
| воз-раст,
лет
|
Высо-
та,
м
|
диам.,
см
|
раст.
леса
|
сух.
леса
|
сред-
ний
|
теку-
щий
|
| 1
|
2
|
3
|
4
|
5
|
6
|
7
|
8
|
9
|
10
|
11
|
12
|
13
|
14
|
| I
|
Дуб
|
5
|
309
|
32,4
|
97,2
|
I
|
24
|
17,8
|
0,5
|
269
|
| Липа
|
2
|
179
|
29,3
|
55,8
|
38
|
9,3
|
0,2
|
126
|
| Клен.о
|
2
|
28,0
|
26,2
|
177
|
9,6
|
0,3
|
109
|
1,6
|
| ильм
|
1
|
27,2
|
30,3
|
36
|
2,6
|
0,07
|
31
|
0,2
|
| Итого
|
10
|
275
|
39,0
|
1,07
|
535
|
1,8
|
| II
|
Липа
|
20
|
19,9
|
1
|
0,03
|
0,003
|
0,3
|
| Клен.о
|
10
|
18
|
12,2
|
136
|
1,6
|
0,06
|
14,1
|
| Ильм
|
15
|
13,7
|
4
|
0,06
|
0,003
|
0,5
|
| Итого
|
10
|
141
|
1,7
|
0,07
|
15
|
| III
|
Клен.о
|
10
|
9,2
|
6
|
211
|
0,6
|
0,04
|
4
|
1,2
|
| Клен.п
|
5
|
4
|
1
|
0,001
|
0,0006
|
0,003
|
| Ильм
|
6,8
|
5,3
|
4
|
0,01
|
0,0009
|
0,04
|
| Итого
|
10
|
216
|
0,6
|
0,042
|
4
|
1,2
|
| Всего
|
I
|
632
|
41,3
|
1,18
|
554
|
3
|
Фаутность и прочие собенности:
суховершинность, фаутность, усыхание ветвей первой велечины, поперечный надлом. рак и стволовая гниль от ложного трутовика - 3% слабая; водяные побеги – 27% сильная; дупла, морозобойные трещины, коровые повреждения ствола – более 8% средняя; снеговые заломы – менее 1%.
Пдр.: Кл.о., Ил.,
Пдл.:_Ил., Кл.о., Кл. п., бузина, бересклет бород (см. табл. 7.2.2.12).
Пкр.: Сныть.
Почва: темно-серая лесная ср. оподзол.
Подпочва: лессов.сугл.
Рельеф и положение: пологий юго-зап. склон
Тип лесораст. условий: Д2_
Тип леса: липо-дубняк снытьевый
_Дата перечёта: 20-28_октября___2004 г.
Обработал: Немченко В.А., Немченко М.Ю.
Таблица 7.2.2.11
Основные итоги обработки постоянной пробной площади № 8 (в переводе на 1 га) в1975 г.
ГПЗ «Белогорье», уч. «Лес на Ворскле», кв.10 , выд. 10
Величина пробной площади 1,0
га
Происхождение: естественное семенное
| Ярус
|
Древесн.
входящ.
в ярус
|
Таксационные показатели
|
| Сос-
тав
|
Средние
|
Бони-
тет
|
Число
ствол.
шт
|
Сум-
ма
площ.
сече-
ния,
м2
|
Пол-
нота
|
Запас
ствол., м3
|
Прирост, м3
|
| воз-раст,
лет
|
Высо-
та,
м
|
диам.,
см
|
раст.
леса
|
сух.
леса
|
сред-
ний
|
теку-
щий
|
| 1
|
2
|
3
|
4
|
5
|
6
|
7
|
8
|
9
|
10
|
11
|
12
|
13
|
14
|
| I
|
Дуб
|
10
|
280
|
29,7
|
93,2
|
II
|
32
|
21,8
|
0,56
|
298
|
8
|
1,10
|
| II
|
Липа
|
10
|
150
|
24,8
|
52,2
|
61
|
13,0
|
0,34
|
155
|
| Клен.о
|
24,6
|
44,2
|
3
|
0,5
|
0,01
|
5
|
| Ильм
|
23,6
|
44,2
|
3
|
0,5
|
0,01
|
5
|
2
|
| Итого
|
67
|
14,0
|
0,36
|
165
|
2
|
1,10
|
| III
|
Дуб
|
16,3
|
15,2
|
11
|
0,2
|
0,01
|
2
|
| Липа
|
12,2
|
8,0
|
26
|
0,1
|
1
|
| Клен.о
|
7
|
12,5
|
8,0
|
949
|
4,6
|
0,19
|
33
|
| Ильм
|
3
|
13,0
|
10,8
|
233
|
2,1
|
0,09
|
15
|
| Итого
|
1219
|
7,0
|
0,29
|
51
|
1,46
|
| Всего
|
II
|
1318
|
42,8
|
1,21
|
514
|
10
|
3,66
|
7,1
|
Фаутность и прочие особенности - суховершинность, усыхание ветвей первой велечины и стволовая
гниль от ложного трутовика - 20% средн.
Пдр._Кл,Лп, Ил ср. густ. Пдл._отсутствует
Пкр.__Сныть,осока___Почва:__темно-серая лесная ср. оподзол.
Подпочва:_лессов.сугл____Рельеф и положение:_пологий юго-зап. склон
Тип лесораст. условий:_Д2__Коренной тип леса:_Л – 1а (липо-дубняк снытьевый)
Дата перечёта:__май____1975 г.
Начальник л/у партии: Прозоров В.В.
Обработал: Фоминых, Миронова Н.А.
Таблица 7.2.2.12
Учёт подроста и подлеска на постоянной пробной площади № 8 (1,0 га) в 2004 г.*
| Высота, м
|
Древесные и кустарниковые породы, шт / га
|
Итого:
|
| Клён остр.
|
Ильм
|
Липа
|
Клён пол.
|
Бузина чёр.
|
Берес. бор.
|
| до 0,5
|
12
|
245
|
-
|
5
|
-
|
3
|
265
|
| до 2
|
16
|
153
|
6
|
1
|
5
|
-
|
181
|
| свыше 2
|
171
|
17
|
-
|
-
|
2
|
-
|
190
|
| Итого:
|
199
|
415
|
6
|
6
|
7
|
3
|
| Всего:
|
636
|
Примечание: * подрост и подлесок повреждается дикими животными (преимущественно
высотой до 0,5 м); материалы учётов 1976 г. в архиве заповедника отсутствуют.
Таблица 7.2.2.13
Лесотаксационная характеристика постоянной пробной площади № 12 (1.0 га) в 2004 году
| Порода
|
I ярус
|
| Ступени
толщины,
см
|
Высота,
м
|
Число сырорастущих деревьев по классам роста
|
Число
сухост.
дерев.,
шт
|
Сумма
площ. сечений, м2
|
Объём
одного
ствола,
м3
|
Запас стволов.
Древесины,
м3
|
| I
|
II
|
III
|
IV
|
итого
|
растущ.
леса
|
сухост.
леса
|
растущ.
леса
|
сухост.
леса
|
| 1
|
2
|
3
|
4
|
5
|
6
|
7
|
8
|
9
|
10
|
11
|
12
|
13
|
14
|
| Дуб
|
20
|
25
|
1
|
1
|
0,031
|
0,37
|
0,37
|
| 24
|
29
|
2
|
2
|
0,091
|
0,60
|
1,20
|
| 28
|
28,5
|
2
|
2
|
0,124
|
0,81
|
1,62
|
| 36
|
33
|
2
|
2
|
0,20
|
1,53
|
3,06
|
| 80
|
31
|
1
|
1
|
0,50
|
7,12
|
7,12
|
| 88
|
29,5
|
2
|
2
|
1,22
|
8.255
|
16,51
|
| 92
|
29,8
|
2
|
2
|
1,32
|
9,09
|
18,18
|
| 96
|
29,3
|
3
|
3
|
2,16
|
9,77
|
29,32
|
| 96
|
30
|
1
|
0,72
|
9,97
|
9,97
|
| 100
|
27,3
|
3
|
3
|
2,35
|
10,17
|
30,53
|
| 104
|
31
|
1
|
1
|
0,85
|
12,03
|
12,03
|
| 108
|
30
|
1
|
1
|
0,92
|
12,61
|
12,61
|
| 112
|
30,3
|
2
|
2
|
1,96
|
13,67
|
27,37
|
| 116
|
32,5
|
2
|
2
|
2,11
|
15,63
|
31,26
|
| 120
|
28,5
|
1
|
1
|
1,13
|
14,89
|
14,89
|
| 128
|
32
|
1
|
1
|
1,29
|
18,78
|
18,78
|
| Итого на
|
пробе
|
26
|
1
|
16,256
|
0,72
|
224,82
|
9,97
|
| Окр.
|
26
|
1
|
16,3
|
0,7
|
225
|
10
|
| Липа
|
12
|
22,9
|
8
|
8
|
0,091
|
0,10
|
0,8
|
| 16
|
24,2
|
25
|
25
|
0,504
|
0,20
|
5,0
|
| 20
|
26,1
|
27
|
27
|
0,849
|
0,35
|
9,54
|
| 20
|
23
|
1
|
0,031
|
0,34
|
0,34
|
| 24
|
26,5
|
16
|
16
|
0,723
|
0,53
|
8,48
|
Таблица 7.2.2.13 (продолжение)
Лесотаксационная характеристика постоянной пробной площади № 12 (1.0 га) в 2004 году
| Порода
|
I ярус
|
| Ступени
толщины,
см
|
Высота,
м
|
Число сырорастущих деревьев по классам роста
|
Число
сухост.
дерев.,
шт
|
Сумма
площ. сечений, м2
|
Объём
одного
ствола,
м3
|
Запас стволов.
Древесины,
м3
|
| I
|
II
|
III
|
IV
|
итого
|
растущ.
леса
|
сухост.
леса
|
растущ.
леса
|
сухост.
леса
|
| 1
|
2
|
3
|
4
|
5
|
6
|
7
|
8
|
9
|
10
|
11
|
12
|
13
|
14
|
| 28
|
27,8
|
13
|
13
|
0,8
|
0,78
|
10,14
|
| 32
|
28,5
|
2
|
2
|
0,16
|
1,06
|
2,12
|
| 36
|
27
|
1
|
1
|
0,102
|
1,36
|
1,36
|
| 40
|
25
|
1
|
1
|
0,126
|
1,66
|
1,66
|
| 44
|
27,8
|
3
|
3
|
0,452
|
2,03
|
6,09
|
| 48
|
26
|
1
|
1
|
0,181
|
2,47
|
2,47
|
| 52
|
29,3
|
4
|
4
|
0,84
|
2,93
|
11,72
|
| 56
|
29
|
4
|
4
|
0,98
|
3,42
|
13,68
|
| 60
|
30,5
|
2
|
2
|
0,56
|
3,95
|
7,90
|
| 68
|
30
|
1
|
1
|
0,36
|
5,15
|
5,15
|
| 72
|
26
|
1
|
1
|
0,41
|
5,80
|
5,80
|
| Итого на пробе
|
106
|
3
|
109
|
1
|
7,138
|
0,031
|
91,91
|
0,34
|
| Окр.
|
109
|
1
|
7,1
|
0,03
|
91,9
|
0,3
|
| Ильм
|
12
|
24
|
1
|
1
|
0,011
|
0,09
|
0,09
|
| 16
|
25,5
|
2
|
2
|
0,040
|
0,17
|
0,34
|
| 20
|
26
|
2
|
2
|
0,063
|
0,30
|
0,60
|
| 24
|
26
|
5
|
5
|
0,227
|
0,47
|
2,35
|
| 28
|
28,5
|
4
|
4
|
0,247
|
0,68
|
2,72
|
| 32
|
29
|
1
|
1
|
0,080
|
0,94
|
0,94
|
| Итого на пробе
|
15
|
15
|
0,668
|
7,04
|
| Окр.
|
15
|
0,7
|
7,0
|
Таблица 7.2.2.13 (продолжение)
Лесотаксационная характеристика постоянной пробной площади № 12 (1.0 га) в 2004 году
| Порода
|
I ярус
|
| Ступени
толщины,
см
|
Высота,
м
|
Число сырорастущих деревьев по классам роста
|
Число
сухост.
дерев.,
шт
|
Сумма
площ. сечений, м2
|
Объём
одного
ствола,
м3
|
Запас стволов.
Древесины,
м3
|
| I
|
II
|
III
|
IV
|
итого
|
растущ.
леса
|
сухост.
леса
|
растущ.
леса
|
сухост.
леса
|
| 1
|
2
|
3
|
4
|
5
|
6
|
7
|
8
|
9
|
10
|
11
|
12
|
13
|
14
|
| Кл. пол.
|
12
|
22
|
1
|
1
|
0011
|
0.09
|
0,09
|
| 16
|
20
|
1
|
1
|
0,020
|
0,17
|
0,17
|
| 20
|
25,4
|
4
|
4
|
0,125
|
0,30
|
1,2
|
| 32
|
28
|
1
|
1
|
0,080
|
0,94
|
0,94
|
| Итого на
|
пробе
|
7
|
0,236
|
2,4
|
| Окр.
|
7
|
0,2
|
2,4
|
| Груша
|
12
|
22
|
2
|
2
|
0,023
|
0,09
|
0,18
|
| 16
|
24
|
1
|
1
|
0,02
|
0,17
|
0,17
|
| 20
|
27
|
2
|
2
|
0,063
|
0,3
|
0,6
|
| 24
|
25
|
1
|
1
|
0,045
|
0,47
|
0,47
|
| 28
|
26
|
1
|
1
|
0,062
|
0,68
|
0,68
|
| Итого на
|
пробе
|
7
|
0,213
|
2,1
|
| Окр.
|
7
|
0.2
|
2,0
|
| Кл. о.
|
8
|
21
|
2
|
2
|
0,010
|
0,03
|
0,06
|
| 8
|
21
|
1
|
0,005
|
0,03
|
0,03
|
| 12
|
22
|
38
|
38
|
0,429
|
0,09
|
3,42
|
| 16
|
23,3
|
56
|
56
|
1,126
|
0,17
|
9,52
|
| 20
|
26,3
|
47
|
47
|
1,477
|
0,3
|
14,1
|
| 24
|
26,3
|
35
|
35
|
1,584
|
0,47
|
16,45
|
Таблица 7.2.2.13 (продолжение)
Лесотаксационная характеристика постоянной пробной площади № 12 (1.0 га) в 2004 году
| Порода
|
I ярус
|
| Ступени
толщины,
см
|
Высота,
м
|
Число сырорастущих деревьев по классам роста
|
Число
сухост.
дерев.,
шт
|
Сумма
площ. сечений, м2
|
Объём
одного
ствола,
м3
|
Запас стволов.
Древесины,
м3
|
| I
|
II
|
III
|
IV
|
итого
|
растущ.
леса
|
сухост.
леса
|
растущ.
леса
|
сухост.
леса
|
| 1
|
2
|
3
|
4
|
5
|
6
|
7
|
8
|
9
|
10
|
11
|
12
|
13
|
14
|
| 28
|
28
|
40
|
40
|
2,464
|
0,68
|
27,2
|
| 32
|
27,2
|
24
|
24
|
1,828
|
0,94
|
22,56
|
| 36
|
28.6
|
13
|
13
|
1,33
|
1,25
|
16,25
|
| 40
|
28,8
|
9
|
9
|
1,132
|
1,61
|
14,49
|
| 40
|
28
|
1
|
0,126
|
1,61
|
1,61
|
| 44
|
29
|
4
|
4
|
0.604
|
2,02
|
8,08
|
| 48
|
30
|
1
|
1
|
0,181
|
2,47
|
2,47
|
| Итого на
|
пробе
|
269
|
2
|
12,165
|
0,131
|
134,6
|
1,64
|
| Окр.
|
269
|
2
|
12,2
|
0,1
|
135
|
1,6
|
| Всего на
|
пробе
|
433
|
4
|
36,708
|
0,882
|
462,83
|
11,95
|
| Окр.
|
433
|
4
|
36,7
|
0,9
|
463
|
11,9
|
Таблица 7.2.2.13 (продолжение)
Лесотаксационная характеристика постоянной пробной площади № 12 (1.0 га) в 2004 году
| Порода
|
II ярус
|
III ярус
|
| Ступени
толщ.,
см /
кл.роста
|
Высо-
та,
м
|
Число дер.,
шт
|
Сумма площ.
сечений,
м2
|
Объём
одного
ствола,
м3
|
Запас ствол.
древесины,
м3
|
Ступени
толщины,
см /
кл.роста
|
Высо-
та,
м
|
Число дер.,
шт
|
Сумма площ.
сечений,
м2
|
Объём
одного
ствола,
м3
|
Запас ствол.
древесины,
м3
|
| раст.
|
сух.
|
раст.
|
сух.
|
раст.
|
сух.
|
раст.
|
сух.
|
раст.
|
сух.
|
раст.
|
сух.
|
| 1
|
15
|
16
|
17
|
18
|
19
|
20
|
21
|
22
|
23
|
24
|
25
|
26
|
27
|
28
|
29
|
30
|
31
|
32
|
| Дуб
|
16
|
18
|
2
|
0,04
|
0,18
|
0,36
|
28
|
6
|
1
|
0,062
|
0,21
|
0,21
|
| 104
|
18
|
1
|
0,85
|
7,25
|
7,25
|
84
|
4,5
|
1
|
0,55
|
1,22
|
1,22
|
| 88
|
2,7
|
1
|
0,61
|
0,80
|
0,80
|
| Итого на пробе
|
2
|
1
|
0,04
|
0,85
|
0,36
|
7,25
|
3
|
1,22
|
2,23
|
| Окр.
|
2
|
1
|
0,04
|
0,9
|
0,4
|
7,3
|
3
|
1,2
|
2,2
|
| Липа
|
8
|
17
|
1
|
0,005
|
0,03
|
0,03
|
8
|
9
|
1
|
0,005
|
0,02
|
0,02
|
| 12
|
16,8
|
9
|
0,102
|
0,10
|
0,86
|
8
|
2
|
1
|
0,005
|
0,004
|
0,004
|
| 12
|
16
|
1
|
0,011
|
0,10
|
0,10
|
12
|
7
|
1
|
0,011
|
0,04
|
0,04
|
| 16
|
17,7
|
3
|
0,06
|
0,18
|
0,54
|
16
|
8,5
|
1
|
0,020
|
0,09
|
0,09
|
| 16
|
5,5
|
1
|
0,02
|
0,06
|
0,06
|
| 28
|
3,3
|
1
|
0,062
|
0,03
|
0,03
|
| Итого на пробе
|
13
|
1
|
0,167
|
0,011
|
1,43
|
0,10
|
2
|
4
|
0,025
|
0,098
|
0,11
|
0,134
|
| Окр.
|
13
|
1
|
0,2
|
0,01
|
1,4
|
0,1
|
2
|
4
|
0,03
|
0,1
|
0,1
|
0,1
|
| Ильм
|
6
|
12
|
1
|
0,003
|
0,025
|
0,025
|
4
|
7
|
4
|
0,005
|
0,015
|
0,06
|
| 8
|
13
|
2
|
0,01
|
0,03
|
0,06
|
4
|
7
|
3
|
0,004
|
0,015
|
0,045
|
| 6
|
6,6
|
6
|
0,017
|
0,022
|
0,14
|
| 6
|
5,6
|
2
|
0,006
|
0,02
|
0,04
|
| 8
|
9,7
|
3
|
0,015
|
0,02
|
0,06
|
| 8
|
11
|
1
|
0,005
|
0,02
|
0,02
|
| Итого на пробе
|
3
|
0,013
|
0,085
|
13
|
6
|
0,037
|
0,015
|
0,26
|
0,105
|
| Окр.
|
3
|
0,01
|
0,09
|
13
|
6
|
0,04
|
0,02
|
0,3
|
0,1
|
Таблица 7.2.2.13 (продолжение)
Лесотаксационная характеристика постоянной пробной площади № 12 (1.0 га) в 2004 году
| Порода
|
II ярус
|
III ярус
|
| Ступени
толщ.,
см /
кл.роста
|
Высо-
та,
м
|
Число дер.,
шт
|
Сумма площ.
сечений,
м2
|
Объём
одного
ствола,
м3
|
Запас ствол.
древесины,
м3
|
Ступени
толщины,
см /
кл.роста
|
Высо-
та,
м
|
Число дер.,
шт
|
Сумма площ.
сечений,
м2
|
Объём
одного
ствола,
м3
|
Запас ствол.
древесины,
м3
|
| раст.
|
сух.
|
раст.
|
сух.
|
раст.
|
сух.
|
раст.
|
сух.
|
раст.
|
сух.
|
раст.
|
сух.
|
| 1
|
15
|
16
|
17
|
18
|
19
|
20
|
21
|
22
|
23
|
24
|
25
|
26
|
27
|
28
|
29
|
30
|
31
|
32
|
| Кл.п.
|
8
|
15
|
1
|
0,005
|
0,03
|
0,03
|
4
|
6,5
|
1
|
0,001
|
0,01
|
0,01
|
| 12
|
14
|
1
|
0,011
|
0,07
|
0,07
|
6
|
6,8
|
4
|
0,012
|
0,021
|
0,085
|
| 16
|
17
|
1
|
0,02
|
0,16
|
0,16
|
8
|
9,5
|
2
|
0,01
|
0,02
|
0.04
|
| Итого
|
На проб
|
3
|
0,036
|
0,26
|
6
|
1
|
0.022
|
0,001
|
0,125
|
0,01
|
| Окр
|
3
|
0,04
|
0,3
|
6
|
1
|
0,02
|
0.001
|
0,13
|
0,01
|
| Груша
|
16
|
15
|
1
|
0,02
|
0,16
|
0,16
|
| Итого
|
На проб
|
1
|
0.02
|
0,16
|
| Окр
|
1
|
0,02
|
0.2
|
| Кл. ост
|
4
|
12
|
1
|
0,001
|
0,025
|
0,025
|
4
|
6,9
|
50
|
0,064
|
0,01
|
0,425
|
| 6
|
13
|
25
|
0,071
|
0,021
|
0.535
|
4
|
56
|
5
|
0,006
|
0,011
|
0,055
|
| 6
|
14
|
0,003
|
0,02
|
0.02
|
6
|
8,4
|
68
|
0,19
|
0,019
|
1,325
|
| 8
|
15
|
78
|
0,391
|
0,03
|
2,34
|
6
|
5.1
|
4
|
0,002
|
0,018
|
0,07
|
| 8
|
13,4
|
5
|
0,025
|
0,03
|
0,15
|
8
|
9
|
26
|
0,13
|
0,02
|
0,52
|
| 12
|
16,9
|
64
|
0,725
|
0,09
|
5,76
|
8
|
4.5
|
1
|
0,005
|
0,02
|
0,02
|
| 12
|
17
|
2
|
0,022
|
0,09
|
0.18
|
12
|
9.3
|
3
|
0,033
|
0,06
|
0,18
|
| 16
|
13
|
12
|
0,241
|
0.17
|
2,01
|
12
|
3
|
3
|
0,034
|
0,06
|
0,18
|
| 20
|
19
|
1
|
0,031
|
0,30
|
0.30
|
16
|
5,6
|
3
|
0,06
|
0,13
|
0,39
|
| 24
|
17
|
1
|
0,045
|
0,35
|
0,35
|
20
|
8
|
1
|
0,031
|
0,23
|
0,23
|
| 24
|
6,5
|
1
|
0,045
|
0,35
|
0,35
|
Таблица 7.2.2.13 (окончание)
Лесотаксационная характеристика постоянной пробной площади № 12 (1.0 га) в 2004 году
| Порода
|
II ярус
|
III ярус
|
| Ступени
толщ.,
см /
кл.роста
|
Высо-
та,
м
|
Число дер.,
шт
|
Сумма площ.
сечений,
м2
|
Объём
одного
ствола,
м3
|
Запас ствол.
древесины,
м3
|
Ступени
толщины,
см /
кл.роста
|
Высо-
та,
м
|
Число дер.,
шт
|
Сумма площ.
сечений,
м2
|
Объём
одного
ствола,
м3
|
Запас ствол.
древесины,
м3
|
| раст.
|
сух.
|
раст.
|
сух.
|
раст.
|
сух.
|
раст.
|
сух.
|
раст.
|
сух.
|
раст.
|
сух.
|
| 1
|
15
|
16
|
17
|
18
|
19
|
20
|
21
|
22
|
23
|
24
|
25
|
26
|
27
|
28
|
29
|
30
|
31
|
32
|
| Итого на пробе
|
182
|
8
|
1,505
|
0,05
|
11,32
|
0,35
|
148
|
17
|
0,448
|
0,162
|
2,68
|
1,065
|
| Окр.
|
182
|
8
|
1,5
|
0,05
|
11,3
|
0,4
|
148
|
17
|
0,4
|
0,2
|
2,7
|
1,07
|
| Всего на пробе
|
204
|
10
|
1,781
|
0,911
|
13,615
|
7,7
|
169
|
31
|
0,532
|
1,496
|
3,175
|
3,541
|
| Окр.
|
204
|
10
|
1,8
|
0,9
|
13,6
|
7,7
|
169
|
31
|
0,5
|
1,5
|
3,2
|
3,5
|
Таблица 7.2.2.14
Основные итоги обработки постоянной пробной площади № 12 (в переводе на 1 га) в 2004 г.
ГПЗ «Белогорье», уч. «Лес на Ворскле», кв. 10, выд.13
Величина пробной площади 1,0
га
Происхождение: естественное семенное и частично порослевое
| Ярус
|
Древесн.
входящ.
в ярус
|
Таксационные показатели
|
| Сос-
тав
|
Средние
|
Бони-
тет
|
Число
ствол.
шт
|
Сум-
ма
площ.
сече-
ния,
м2
|
Пол-
нота
|
Запас
ствол., м3
|
Прирост, м3
|
| воз-раст,
лет
|
Высо-
та,
м
|
диам.,
см
|
раст.
леса
|
сух.
леса
|
сред-
ний
|
теку-
щий
|
| 1
|
2
|
3
|
4
|
5
|
6
|
7
|
8
|
9
|
10
|
11
|
12
|
13
|
14
|
| I
|
Дуб
|
5
|
308
|
29,8
|
89
|
II
|
26
|
16,3
|
0,44
|
225
|
10
|
| Липа
|
2
|
27,5
|
29
|
109
|
7,1
|
0,14
|
91,9
|
0,3
|
| Ильм
|
26,5
|
24
|
15
|
0,7
|
0,02
|
7,0
|
| Кл. п.
|
23,9
|
19
|
7
|
0,2
|
0,006
|
2,4
|
| Груша
|
24,8
|
19
|
7
|
0,2
|
0,006
|
2,0
|
| Кл. ост
|
3
|
26,4
|
24
|
269
|
12,2
|
0,36
|
135
|
1,6
|
| Итого
|
10
|
433
|
36,7
|
0,97
|
463,3
|
11,9
|
| II
|
Дуб
|
1
|
18
|
16
|
2
|
0,04
|
0,002
|
0,4
|
7,3
|
| Липа
|
1
|
17,2
|
14
|
13
|
0,2
|
0,006
|
1,4
|
0,1
|
| Ильм
|
12,5
|
6
|
3
|
0,01
|
0.0004
|
0,09
|
-
|
| Кл. п.
|
15,3
|
13
|
3
|
0,04
|
0,002
|
0,3
|
-
|
| Груша
|
15
|
16
|
1
|
0,02
|
0,0008
|
0,2
|
-
|
| Кл ост
|
8
|
15,3
|
10
|
182
|
1,5
|
0,06
|
11,3
|
0,4
|
| Итого
|
10
|
204
|
1,81
|
0,07
|
13,69
|
7,8
|
| III
|
Дуб
|
-
|
-
|
-
|
-
|
2,2
|
| Липа
|
1
|
8,8
|
14
|
2
|
0,03
|
-
|
0,1
|
0,1
|
| Ильм
|
1
|
7,8
|
6
|
13
|
0,04
|
0,003
|
0,3
|
0,1
|
| Кл. п.
|
8,2
|
6
|
6
|
0,02
|
0,001
|
0,13
|
0,01
|
| Кл. о.
|
8
|
8,3
|
6
|
148
|
0,4
|
0,03
|
2,7
|
1,07
|
| Итого
|
10
|
169
|
0,49
|
0,034
|
3,23
|
3,48
|
| Всего
|
II
|
806
|
39
|
1,07
|
480,22
|
23,18
|
Фаутность и прочие собенности: поперечный надломов. рак, стволовые трутовики – 3% слабое; суховершинность, усыхание ветвей – более 2% слабое; дупла, коровые повреждения ствола, морозобойные трещины – 5% слабое.
Пдр._Ил, Кл. о., Лп _________________Пдл._Ил, Кл. о., Кл. п., берес.евр., бузина_________Пкр._сныть__
Почва: темно-серая лесная ср. оподзолен _
Подпочва: лессов.сугл __Рельеф и положение: слабый южный склон
Тип лесораст. условий: Д1-2_________Тип леса: клёно-дубняк снытьевый__
_Дата перечёта_10-20 ноября___2004 г.
Обработал:Немченко В.А., Немченко М.Ю.____
Таблица 7.2.2.15
Основные итоги обработки постоянной пробной площади № 12 (в переводе на 1 га) в 1976 г.
ГПЗ «Белогорье», уч. «Лес на Ворскле», кв.10, выд. 13
Величина пробной площади 1,0
га
Происхождение: естественное семенное и частично порослевое.
| Ярус
|
Древесн.
входящ.
в ярус
|
Таксационные показатели
|
| Сос-
тав
|
Средние
|
Бони-
тет
|
Число
ствол.
шт
|
Сум-
ма
площ.
сече-
ния,
м2
|
Пол-
нота
|
Запас
ствол., м3
|
Прирост, м3
|
| воз-раст,
лет
|
Высо-
та,
м
|
диам.,
см
|
раст.
леса
|
сух.
леса
|
сред-
ний
|
теку-
щий
|
| 1
|
2
|
3
|
4
|
5
|
6
|
7
|
8
|
9
|
10
|
11
|
12
|
13
|
14
|
| I
|
Дуб
|
10
|
280
|
30,4
|
98,5
|
II
|
29
|
22,1
|
0,56
|
305
|
78
|
1,1
|
| II
|
Липа
|
8
|
24,9
|
52,3
|
29
|
6,2
|
0,16
|
73
|
| Дуб
|
1
|
24,2
|
71,5
|
3
|
0,12
|
0,04
|
14
|
| Ильм
|
1
|
21,8
|
39,5
|
5
|
0,6
|
0,02
|
6
|
| Клен
|
4
|
0,2
|
2
|
| Итого
|
10
|
150
|
41
|
8,2
|
0,22
|
95
|
0,6
|
| III
|
Клен
|
6
|
11,1
|
9,6
|
802
|
5,7
|
0,28
|
33
|
1
|
| Липа
|
2
|
12,5
|
10,7
|
256
|
2,3
|
0,09
|
14
|
| Ильм
|
1
|
12,8
|
11,7
|
89
|
1,0
|
0,04
|
7
|
1
|
| Груша
|
1
|
13,2
|
12,0
|
53
|
0,6
|
0,02
|
4
|
| Дуб
|
13
|
0,2
|
2
|
1
|
| Итого
|
10
|
35
|
1213
|
9,8
|
0,43
|
60
|
3
|
1,7
|
| Всего
|
II
|
1283
|
40,1
|
1,21
|
460
|
81
|
3,4
|
Фаутность и прочие особенности: I яр. суховерш. и усыхание крупных ветвей – центральн. и стволовая гниль от ложного трутовика – 20%, среднее. II яр. центральн. стволовая гниль липы от настоящего трутовика и
коллибии – 60%, среднее. III яр. состояние хорошее.
Пдр._Д, Кл, Ил Пдл._клен полевой, бересклет европейский
Пкр.__Сныть, осока___Почва:_темно-серая лесная ср. оподзолен
Подпочва:_лессовидн.сугл.____Рельеф и положение: слабый южный склон
Тип лесораст. условий: Д 1 – 2 Коренной тип леса: Д – 1а (дубняк снытьево-осоковый)
Дата перечёта: 20 /
VII 1976 г.
Начальник л/у партии: Прозоров В.В.
Обработал: инженер В.В., Миронова, Фоминых _
Таблица 7.2.2.16
Учёты подроста и подлеска на постоянной пробной площади № 12 (1,0 га)
в 2004 г.*
| Высота, м
|
Древесные и кустарниковые породы, шт / га
|
Итого:
|
| Клён остр.
|
Ильм
|
Липа
|
Клён пол.
|
Бузина чёр.
|
Берес. евр.
|
Берес. бор.
|
| до 0,5
|
19
|
332
|
-
|
35
|
11
|
142
|
1
|
540
|
| до 2
|
23
|
506
|
6
|
44
|
11
|
2
|
-
|
592
|
| Свыше 2
|
86
|
159
|
4
|
28
|
5
|
-
|
-
|
282
|
| Итого:
|
128
|
997
|
10
|
107
|
27
|
144
|
1
|
| Всего:
|
1414
|
Примечание: * подрост и подлесок повреждается дикими животными (преимущественно
высотой до 0,5 м).
в 1976 г.*
| Высота, м
|
Древесные и кустарниковые породы, тыс. шт / га
|
Итого:
|
| Клён остр.
|
Ильмовые
|
Дуб
|
| до 5 лет
|
до 10 лет
|
бол.10 лет
|
До 5 лет
|
До 5 лет
|
| до 0,5
|
0,8
|
-
|
-
|
0,2
|
32,5
|
33,5
|
| до 2
|
-
|
-
|
-
|
-
|
-
|
-
|
| свыше 2
|
-
|
1,5
|
-
|
-
|
-
|
1,5
|
| Итого:
|
0,8
|
1,5
|
-
|
0,2
|
32,5
|
| Всего:
|
35,0
|
Примечание: * однолетние всходы дуба поражены мучнистой росой;
подрост и подлесок повреждаются дикими животными.
8.
ФАУНА И ЖИВОТНОЕ НАСЕЛЕНИЕ
ФАУНА
Список видов птиц ООПТ Белгородской области
Исследования проводились в мае 2004 года на разных участках ООПТ Белгородской области: заказник «Бекарюковский бор» (пойма р.Нежеголь у с. Маломихайловка Шебекинского р-на), урочище «Борки» (р.Козинка у с. Борки Валуйского р-на).
Исследования авифауны участков «Бекарюковский бор» и «Борки» заключались в регистрации всех встреченных птиц при однократных посещениях этих мест.
Наблюдения птиц в « Бекарюковском бору »
Участок образован несколькими стациями: водоемом (старица р. Нежеголь), прилегающим к нему лугом, склоном на котором расположены сельскохозяйственные поля, лесом, в основном образованным сосной, растущей на меловых склонах и заброшенным фруктовым садом. Таким образом наблюдается высокая мозаичность участка, что в свою очередь определяет видовое разнообразие птиц, населяющих его.
05.05.04
-
06.05.04 г.
Водоем и прилегающие луг со склоном :
Скворец(Sturnus
vulgaris
) -
многочисленен на гнездовании в полосе старых ив и клена,отделяющих водоем от луга.
Вертишейка (Jynx
torquilla
) -
обычна,не раз регистрировалась за время экскурсии.
Ворон(Corvus
corax
) -
одна встреча.
Зарянка ( Erithacus
rubecula
)
- одна встреча.
Пеночка теньковка (Philloscopus
collybita
) -
одна встреча.
Щегол (Carduelis
carduelis
) -
одна встреча.
Луговой конек (Anthus
pratensis
) -
3 самца и самка.
Лесной конек (Anthus
trivialis
) -
3 самца.
Поползень (Sitta
europaea
) -
одна встреча.
Перепел (Coturnix
coturnix
) -
одна встреча.
Луговой чекан (Saxicola
rubetra
) -
обычен на гнездовании не раз регистрировался за время экскурсии.
Жаворонок полевой (Alauda
arvensis
) -
одна встреча.
Жулан (Lanius
collurio
) -
одна встреча.
Желтая трясогузка (Motacilla
flava
) -
одна встреча поющего самца.
Ремез (Remiz
pendulinus
) -
одна встреча.
Славка завирушка (Sylvia
curruca
) -
одна встреча поющего самца.
Зяблик ( Fringilla
coelebs
) -
обнаружено гнездо с насиживающей самкой.
Дубонос (Coccothraustes
coccothraustes
) -
обнаружено гнездо с насиживающей птицей.
Кукушка (
Cuculus
canorus
)
- одна встреча.
Канюк (Buteo
buteo
) -
одна встреча.
Болотный лунь (Circus
aeruginosus
) -
одна встреча охотящейся самки.
Лес:
Черный дрозд ( Turdus
merula
) -
одна встреча.
Зяблик ( Fringilla
coelebs
) - обычен,отмечался не однократно.
Певчий дрозд (Turdus
philomelos
)
- одна встреча.
Овсянка обыкновенная (Emberiza
citrinella
) -
2 самца.
Соловей (Luscinia
luscinia
) -
3 самца.
Лесной конек (Anthus
trivialis
) -
одна встреча.
Синица большая (Parus
major
) -
одна встреча.
Пеночка теньковка (Philloscopus
collybita
) -
одна встреча.
Дубонос (Coccothraustes
coccothraustes
) -
отмечена стайка птиц из 5 особей.
Пищуха (Certhia
familiaris
) -
одна встреча.
Пухляк (Parus
montanus
) -
поющий самец.
Горлица обыкновенная (Streptopelia
turtur
) -
одна встреча.
Удод (Upupa
epops
) -
1 поющий самец.
Неясыть серая (Strix
aluco
) -
одна встреча.
Фруктовый сад:
Лесной конек (Anthus
trivialis
) -
одна встреча.
Дубонос (Coccothraustes
coccothraustes
) - многочисленен на гнездовании.
Вертишейка (Jynx
torquilla
) -
одна встреча.
Зеленушка (Chloris
chloris
) -
обычна на гнездовании;
регистрировались как сами птицы,так и их гнезда.
Овсянка обыкновенная (Emberiza
citrinella
) -
обычна, не раз отмечалась во время экскурсии.
Лазоревка (Parus
caeruleus
) -
обнаружено гнездо с кладкой.
Ласточка деревенская(Hirundo
rustica
) -
стайки птиц,кормящиеся над садом.
Наблюдения в урочище « Борки »
Участок представляет собой меловые склоны , луг и небольшую речку (р.Козинка) возле них.
06.05.04 – 07.05.04 г.
Птицы,отмеченные возле реки:
Полевой воробей (Passer
montanus
)
Овсянка обыкновенная (Emberiza
citrinella
)
Пеночка теньковка (Philloscopus collybita)
Ласточка деревенская(Hirundo
rustica
)
Трясогузка белая (Motacilla
alba
)
Певчий дрозд (Turdus philomelos)
Соловей (Luscinia luscinia)
Иволга (Oriolus oriolus)
Зяблик ( Fringilla coelebs
)
Зимородок(Alctdo atthis)
Кукушка (Cuculus canorus)
Щегол (Carduelis carduelis)
Вертишейка (Jynx torquilla)
Скворец(Sturnus vulgaris)
Славка завирушка (Sylvia
curruca
)
Ворона серая (Corvus
cornix
)
Ремез (Remiz pendulinus)
Коноплянка (Cannabina cannabina)
Вяхирь (Columba palumbus)
Удод (Upupa epops)
Птицы,отмеченные на лугу:
Луговой чекан (Saxicola
rubetra
)
Каменка (Oenanthe oenanthe)
Овсянка обыкновенная (Emberiza citrinella)
Птицы,отмеченные на меловых склонах,поросших редким кустарником:
Славка серая (Sylvia communis)
Лесной конек (Anthus trivialis)
Сорока (Pica pica)
Каменка (Oenanthe oenanthe)
Кобчик (Falco vespertinus)
Коноплянка (Cannabina cannabina)
Териофауна
Материалы по фауне млекопитающих (табл. 8.1.2.1) подготовлены по результатам полевых исследований на территории участка «Острасьевы яры» 14.06-19.06.2004г. Аксеновой Т.Г. (Санкт-Петербургский государственный университет) в дополнение к списку видов млекопитающих Острасьевых яров (Летопись природы 2003). В ходе изучения мелких млекопитающих на участке «Острасьевы яры» в совокупности отработано 200 давилко-суток и 10 канавко-суток (2 ловчие канавки по 50 м с 5 конусами в каждой). Всего отловлено 153 экз. мелких млекопитающих 12 видов. Видовые названия млекопитающих приведены по Павлинову И.Я. и др. (2002).
Таблица 8.1.2.1
Дополнение к списку видов млекопитающих участка «Острасьевы яры»
| №
п/п
|
Наименование вида
|
Место
|
Учтено особей
|
| 1.
|
Хомячок серый - Cricetulus migratorius Pallas, 1773
|
степь
|
2
|
| 2.
|
Полевка рыжая – Сlethrionomys glareolus Schreber, 1780
|
опушка леса
|
1
|
| 3.
|
Пеструшка степная - Lagurus lagurus Pallas, 1773
|
степь, опушка леса
|
8
|
| 4.
|
Полевка восточноевропейская - Microtus rossiaemeridionalis Ognev, 1924
|
степь, опушка леса
|
57
|
| 5.
|
Мышь-малютка - Micromys minutus Pallas, 1771
|
опушка леса
|
1
|
8.2. ЖИВОТНОЕ НАСЕЛЕНИЕ
8.2.1. Дикие пчелиные участка «Острасьевы яры» и прилегающих сельхозугодий
В 2004 году было продолжено изучение заселения искусственных гнезд из тростника и других материалов одинерами (дикими одиночными пчелами и осами). Как оказалось, заселены были не все тростниковые трубочки, а в тех трубочках, которые были заселены, не всегда личиночные ячейки заполняли всю полость, то есть часть трубочек была заселена частично и эти трубочки не были запечатаны снаружи комочком почвы или обрывками сухих листьев (табл.8.2.1.1).
Таблица 8.2.1.1
Заселённость искусственных гнёзд из тростника одинёрами (дикими одиночными пчёлами
и осами), «Острасьевы яры», 2004 г.
| №
гнезда
|
Трубочек
в гнезде,
штук
|
Из них заселены одинёрами
|
| полностью
|
частично
|
Всего
|
| штук
|
%
|
штук
|
%
|
штук
|
%
|
| 1
|
47
|
9
|
19.1
|
2
|
4.3
|
11
|
23.4
|
| 2
|
33
|
4
|
12.1
|
7
|
21.2
|
11
|
33.3
|
| 3
|
45
|
0
|
0
|
5
|
11.1
|
5
|
11.1
|
| 4
|
28
|
0
|
0
|
1
|
3.6
|
1
|
3.6
|
| 5
|
26
|
0
|
0
|
3
|
11.5
|
1
|
11.5
|
| 6
|
26
|
4
|
15.4
|
6
|
23.0
|
10
|
38.4
|
| 7
|
34
|
1
|
2.9
|
0
|
0
|
1
|
2.9
|
| 8
|
26
|
1
|
3.8
|
14
|
53.8
|
15
|
57.6
|
| 9
|
47
|
3
|
6.4
|
10
|
21.3
|
13
|
27.7
|
| 10
|
37
|
0
|
0
|
2
|
5.4
|
2
|
5.4
|
| 11
|
47
|
1
|
2.1
|
3
|
6.4
|
4
|
8.5
|
| 12
|
29
|
0
|
0
|
3
|
10.3
|
3
|
10.3
|
| 13
|
48
|
0
|
0
|
4
|
8.3
|
4
|
8.3
|
| 14
|
55
|
0
|
0
|
4
|
7.3
|
4
|
7.3
|
| 15
|
52
|
4
|
7.7
|
5
|
9.6
|
9
|
17.3
|
| 16
|
37
|
2
|
5.4
|
4
|
10.8
|
6
|
16.2
|
| 17
|
45
|
0
|
0
|
2
|
4.4
|
2
|
4.4
|
| 18
|
23
|
1
|
4.3
|
3
|
13.0
|
4
|
17.4
|
| 19
|
43
|
0
|
0
|
4
|
9.3
|
4
|
9.3
|
| 20
|
33
|
0
|
0
|
2
|
6.1
|
2
|
6.1
|
| 21
|
50
|
0
|
0
|
4
|
8.0
|
4
|
8.0
|
| 22
|
35
|
0
|
0
|
3
|
8.6
|
3
|
8.6
|
| Всего
|
846
|
30
|
3.5
|
91
|
10.8
|
121
|
14.3
|
В 2004 г., как видно из таблицы 8.2.1.2, заселенность гнезд пчелами оказалась выше, чем в 2002 и 2003 гг. Причем по годам наблюдается тенденция к увеличению заселенности гнездовых трубочек, что косвенно свидетельствует об увеличении численности одинеров (пчёл, без ос) на исследуемом участке. Возможно, это связано с особенностями погоды в годы исследований, наличия достаточной кормовой базы. Но могло сказаться и присутствие сотен искусственных гнезд, что в значительной степени улучшило условия местообитания диких пчел.
Таблица 8.2.1.2
Заселенность тростниковых гнезд дикими одиночными пчелами.
Заповедный участок «Острасьевы яры», 2002 – 2004 гг.
| Год
|
Тростниковых трубочек в опыте,
шт
|
Из них заселенных пчелами
|
| штук
|
%
|
| 2002
|
1250
|
15
|
1.0
|
| 2003
|
1127
|
73
|
6.5
|
| 2004
|
846
|
108
|
12.8
|
Анализ заселенных в 2004 году дикими одиночными пчелами тростниковых трубочек
показал, что 81.4% их имели диаметр от 10 до 13 мм, а диаметр 50% всех заселенных
пчелами трубочек составлял 11-12 мм, табл. 8.2.1.3.
Таблица 8.2.1.3
Распределение тростниковых трубочек, заселенных одиночными пчелами, по диаметру.
Заповедный участок «Острасьевы яры», 2004 г.
| Диаметр трубочки, мм
|
Количество заселенных пчелами трубочек
|
| штук
|
%
|
| 7
|
2
|
1.9
|
| 8
|
5
|
4.6
|
| 9
|
5
|
4.6
|
| 10
|
21
|
19.4
|
| 11
|
29
|
26.9
|
| 12
|
26
|
24.1
|
| 13
|
13
|
12.0
|
| 14
|
7
|
6.5
|
| Всего:
|
108
|
100,0
|
Вскрытие заселенных трубочек позволило установить некоторые особенности устройства
ячеек внутри полости трубочек, а также их количество и состояние находившихся там пчел,
табл. 8.2.1.4.
Таблица 8.2.1.4
Результаты вскрытия тростниковых трубочек, заселенных дикими одиночными пчелами. Заповедный участок «Острасьевы яры», 2004 г.
| Конструкция и содержимое ячеек
|
Всего ячеек
|
Из них
|
| С живыми пчелами
|
С погибшими пчелами
|
Пустые ячейки
|
| штук
|
%
|
штук
|
%
|
Штук
|
%
|
| Ячейки из листьев, коконы темно-серые
|
224
|
167
|
74.6
|
5
|
2.2
|
52
|
23.2
|
| Ячейки с земляными перегородками, коконы до 11 мм в длину и 6 мм диаметром, светло-серые, плотные
|
339
|
248
|
73.2
|
22
|
6.5
|
69
|
20.3
|
| Ячейки с земляными перегородками, коконы до 8 мм длиной и 4 мм диаметром, полупрозрачные, серые
|
36
|
25
|
69.4
|
0
|
0
|
11
|
30.6
|
| Всего:
|
599
|
440
|
73.5
|
27
|
4.5
|
132
|
22.0
|
Тростниковые трубочки были заселены по крайней мере тремя видами пчел ( видовой состав
будет определен после выведения взрослых насекомых). Коконы с живыми пчелами составляли
73.5% от общего числа, а среднее количество ячеек на одну заселенную тростниковую трубочку составило: (599 : 108) = 5.54 ячейки, в том числе с живыми пчелами – 4.07 (440 : 108).
Выводы:
1. Заселённость искусственных гнёзд из тростника одинерами (дикими одиночными пчелами и осами) в 2004 г. на участке «Острасьевы яры» составило: из 846 выставленных трубочек (в 22-х гнёздах) - полностью заселены 30 шт (3.5%), частично заселены 91 шт (10.8%), всего заселено 121 шт (14.3%).
2. По годам наблюдается тенденция к увеличению заселенности гнездовых трубочек одиночными пчёлами (без ос): в 2002 г. из выставленных 1250 трубочек заселено пчёлами 15 шт (1%); в 2003 г. из 1127 шт заселено 73 шт (6.5%); в 2004 г. из 846 шт заселено 108 шт (12.8%) , что косвенно свидетельствует о росте численности пчёл одинеров на исследуемом участке. Возможно, это связано с особенностями погоды в годы исследований, наличием достаточной кормовой базы и постоянным присутствием сотен искусственных гнезд, что в значительной степени улучшило условия обитания диких пчел.
3. Тростниковые трубочки были заселены, по крайней мере, тремя видами пчел (видовой состав будет определен после выведения имаго).
4. Анализ заселенных в 2004 году дикими одиночными пчелами тростниковых трубочек показал, что 81.4 % их имели диаметр от 10 до 13 мм, а диаметр 50 % всех заселенных пчелами трубочек составлял 11-12 мм.
5. Не все тростниковые трубочки были заселены пчелами. В заселенных пчелами трубочках не всегда личиночные ячейки заполняли всю полость, то есть часть трубочек была заселена частично, и эти трубочки не были запечатаны снаружи комочком почвы или обрывками сухих листьев. Коконы с живыми пчелами составляли 73.5 % (440 ячеек из 599 обнаруженных), а среднее количество ячеек на одну заселенную тростниковую трубочку составило 5.54 шт, в том числе с живыми пчелами – 4.07.
6. Анализ заселенных в 2004 году дикими одиночными пчелами тростниковых трубочек показал, что 81.4 % их имели диаметр от 10 до 13 мм, а диаметр 50 % всех заселенных пчелами трубочек составлял 11-12 мм.
8.2.2. Учёты населения птиц на участке «Лес на Ворскле»
Исследования проводились в весеннее-летний период 2004 года.
Учет птиц в « Лесу на Ворскле » проводился на постоянном маршруте по стандартной методике (Новиков, 1953). Протяженность маршрута 1,0 км. Он проходил диагональю через 8-й выдел 10 квартала.
Также учеты птиц проводились на пробной орнитологической площадке площадью 10,5 га (кв. 10, выд. 8), которая включает лесотипологическую постоянную пробную площадь «Сукачевская делянка» площадью 1,0 га. Учеты проводились путем многократного посещения пробной площадки в ранние утренние часы и регистрации птиц сначала при обходе внешних границ участка, а затем при дополнительных заходах в глубь участка. Все встречи птиц наносились на небольшой контурный план площадки, для каждого вида птиц отдельный на каждую экскурсию. В результате повторных посещений участка накапливался ряд карточек с указанием местонахождения птиц. Сопоставляя их удалось выявить постоянных обитателей пробной орнитологической площадки.
Учеты птиц на постоянном маршруте « Лес на Ворскле »
17.05.04
Начало маршрута 5ч.30 мин.
Пасмурно, ветер слабый.
Черный дрозд ( Turdus
merula
) -
5 поющих самцов.
Зяблик ( Fringilla
coelebs
) - 9 поющих самцов.
Мухоловка белошейка (Ficedula
albicollis
) -
11 поющих самцов,1 самка.
Певчий дрозд (Turdus
philomelos
)
- 3 поющих самца.
Славка черноголовая (Sylvia
atricapilla
)
- 2 поющих самца.
Зарянка ( Erithacus
rubecula
) -
2 поющих самца.
Пеночка трещотка (Philloscopus
sibilatrix
) -
4 поющих самца.
Мухоловка пеструшка (Ficedula
hypoleuca
) -
1 поющий самец.
Лазоревка (Parus
caeruleus
)
- 2 птицы.
Пищуха (Certhia familiaris) -
2 птицы.
Большой пестрый дятел ( Dendrocopos
major
) - 3
птицы.
Поползень (Sitta
europaea
) -
2 поющих самца.
Синица большая (Parus
major
) -
5 поющих самцов.
Дубонос (Coccothraustes coccothraustes) -
1 птица.
Кукушка (Cuculus canorus) -
1 поющий самец.
Удод (Upupa epops) -
1 поющий самец.
Горлица обыкновенная (Streptopelia turtur) -
1 поющий самец.
19.05.04
Начало маршрута 5ч. 30 мин.
Переменная облачность.
Черный дрозд ( Turdus
merula
) -
5 поющих самцов.
Зяблик ( Fringilla
coelebs
) - 8 поющих самцов.
Мухоловка белошейка (Ficedula
albicollis
) -
10 поющих самцов.
Певчий дрозд (Turdus
philomelos
)
- 2 поющих самца.
Славка черноголовая (Sylvia
atricapilla
)
- 2 поющих самца.
Зарянка ( Erithacus
rubecula
) -
1 поющий самец.
Пеночка трещотка (Philloscopus
sibilatrix
) -
3 поющих самца.
Пищуха (Certhia familiaris) -
1 птица.
Большой пестрый дятел ( Dendrocopos
major
) - 4
птицы.
Поползень (Sitta
europaea
) -
3 поющих самца.
Синица большая (Parus
major
) -
3 поющих самцов.
22.05.04
Начало маршрута 5ч. 10 мин.
Переменная облачность.
Черный дрозд ( Turdus
merula
) -
5 поющих самцов.
Зяблик ( Fringilla
coelebs
) - 9 поющих самцов.
Мухоловка белошейка (Ficedula
albicollis
) -
10 поющих самцов.
Певчий дрозд (Turdus
philomelos
)
- 3 поющих самца.
Славка черноголовая (Sylvia
atricapilla
)
- 2 поющих самца.
Зарянка ( Erithacus
rubecula
) -
1 поющий самец.
Пеночка трещотка (Philloscopus
sibilatrix
) -
4 поющих самца.
Лазоревка (Parus
caeruleus
)
- 1 птица.
Пищуха (Certhia familiaris) -
3 птицы.
Большой пестрый дятел ( Dendrocopos
major
) - 1
птица.
Поползень (Sitta
europaea
) -
3 поющих самца.
Синица большая (Parus
major
) -
3 поющих самца.
Удод (Upupa
epops
) -
1 поющий самец.
Горлица обыкновенная (Streptopelia
turtur
) -
1 поющий самец.
24.05.04
Начало маршрута 5ч.30мин.
Пасмурно.
Черный дрозд ( Turdus
merula
) -
4поющих самца.
Зяблик ( Fringilla
coelebs
) - 8поющих самцов.
Мухоловка белошейка (Ficedula
albicollis
) -
10 поющих самцов.
Певчий дрозд (Turdus
philomelos
)
- 1 поющий самец.
Славка черноголовая (Sylvia
atricapilla
)
- 1 поющий самец.
Зарянка ( Erithacus
rubecula
) -
1 птица,1 поющий самец.
Пеночка трещотка (Philloscopus
sibilatrix
) -
3поющих самца.
Пищуха (Certhia familiaris) -
1тица.
Поползень (Sitta europaea) -
2 птицы.
Синица большая (Parus
major
) -
1поющий самец
Кукушка (
Cuculus
canorus
) -
1 поющий самец.
Пеночка теньковка (Philloscopus
collybita
) -
1 поющий самец.
07.06.04
Начало маршрута 5ч.15мин.
Ясно.
Черный дрозд ( Turdus
merula
) -
2оющих самца.
Зяблик ( Fringilla
coelebs
) - 5оющих самцов.
Мухоловка белошейка (Ficedula
albicollis
) -
9поющих самцов.
Певчий дрозд (Turdus
philomelos
)
- 1поющий самец.
Славка черноголовая (Sylvia
atricapilla
)
- 1поющих самца.
Зарянка ( Erithacus rubecula) -
1 птица.
Пеночка трещотка (Philloscopus sibilatrix) -
2 поющих самца.
Пеночка теньковка (Philloscopus collybita) -
3 поющих самца.
Пищуха (Certhia familiaris) -
2 птицы.
Большой пестрый дятел ( Dendrocopos
major
) -
2 птицы.
Поползень (Sitta
europaea
) -
1 поющий самец.
Синица большая (Parus
major
) -
3 поющих самцов.
Учеты птиц на пробной орнитологической площадке участка « Лес на Ворскле »
18.05.04
Начало обхода 5ч.20 мин.
Переменная облачность
Черный дрозд ( Turdus
merula
) -
1поющий самец.
Зяблик ( Fringilla
coelebs
) - 1поющий самец, 1 самец пел и перелетал с дерева на дерево.
Мухоловка белошейка (Ficedula
albicollis
) -
1 поющий самец.
Певчий дрозд (Turdus
philomelos
)
- 1 поющий самец.
Пеночка трещотка (Philloscopus
sibilatrix
) -
1поющий самец.
Пищуха (Certhia
familiaris
) -
1тица, кормящаяся на стволе дуба.
Синица большая (Parus
major
) -
2 кормящиеся на подросте клена птицы.
Сойка (Garrulus
glandarius
)
- 1 птица,издававшая крик тревоги при обнаружении наблюдателя.
20.05.04
Начало обхода 5ч.30мин.
Пасмурно.
Черный дрозд ( Turdus
merula
) -
1поющий самец.
Зяблик ( Fringilla
coelebs
) - 2поющих самца.
Мухоловка белошейка (Ficedula
albicollis
) -
1 поющий самец.
Певчий дрозд (Turdus
philomelos
)
- 1 поющий самец.
Пеночка трещотка (Philloscopus
sibilatrix
) -
1поющий самец.
Синица большая (Parus
major
) -
1 кормящиеся птица.
Поползень (Sitta
europaea
) -
1 поющий самец, перелетающий южную границу участка.
Зарянка ( Erithacus
rubecula
) -
1 птица,кормящаяся у ствола поваленного дерева в глубине участка.
23.05.04
Начало обхода 5ч.10мин.
Пасмурно.
Черный дрозд ( Turdus
merula
) -
1поющий самец.
Зяблик ( Fringilla
coelebs
) - 3поющих самца.
Мухоловка белошейка (Ficedula
albicollis
) -
2 поющих самца, 1 самец без песни сидел на дереве возле западной границы участка,1 самец в глубине участка издавал крик тревоги.
Пеночка трещотка (Philloscopus
sibilatrix
) -
1поющий самец.
Пеночка теньковка (Philloscopus
collybita
) -
2 поющих самца.
Пищуха (Certhia familiaris) -
1 птица.
Большой пестрый дятел ( Dendrocopos
major
) -
1 птица, перелетавшая с дерево на дерево с криком тревоги.
Синица большая (Parus
major
) -
самец,издающий тревожные позывы.
Поползень (Sitta
europaea
) -
1 кормящаяся в глубине участка.
Зарянка ( Erithacus rubecula) -
1 птица.
25.05.04
Начало обхода 5ч.30мин.
Дождь,пасмурно.
Черный дрозд ( Turdus
merula
) -
1поющий самец.
Зяблик ( Fringilla
coelebs
) - 2поющих самца.
Мухоловка белошейка (Ficedula
albicollis
) -
1 поющий самец.
Синица большая (Parus
major
) -
4 взрослые птицы,кормящиеся в подлеске.
05.06.04
Начало обхода 5ч.30мин.
Облачно.
Черный дрозд ( Turdus
merula
) -
1поющий самец.
Зяблик ( Fringilla
coelebs
) - 2поющих самца.
Мухоловка белошейка (Ficedula
albicollis
) -
2 поющих самца.
Пеночка трещотка (Philloscopus
sibilatrix
) -
1поющий самец.
Пеночка теньковка (Philloscopus
collybita
) -
2 поющих самца.
Пищуха (Certhia familiaris) -
1 птица.
Синица большая (Parus
major
) -
кормящийся выводок из 5 птиц.
Поползень (Sitta
europaea
) -
1 кормящаяся в глубине участка.
Зарянка ( Erithacus rubecula) -
1 птица.
8.2.3. Население млекопитающих
8.2.3.1. Население рукокрылых
Материалы по населению рукокрылых (табл. 8.2.3.1) подготовлены в результате полевых исследований на территории участка «Лес на Ворскле» 18.07-26.07.2004 г. Влащенко А.С. (Харьковский национальный университет им. В.Н. Каразина). В местах охоты рукокрылые учитывались при помощи ультразвукового детектора Peterson D200. Проведено 10 детекторных учетов, в ходе которых фиксировались: видовая принадлежность, количество особей, направление лета, характер и время кормодобывания.
Таблица 8.2.3.1
Результаты учета рукокрылых с помощью ультразвукового детектора
на участке «Лес на Ворскле» (сведения Влащенко А.С.)
| Наименование вида
|
Учтено особей
|
Обилие, %
|
| Ушан бурый - Plecotus auritus Linnaeus, 1758
|
4
|
2
|
| Вечерница малая - Nyctalus leisleri Kuhl, 1817
|
3
|
1
|
| Вечерница рыжая - Nyctalus noctula Schreber, 1774
|
45
|
20
|
| Нетопырь-карлик - Pipistrellus pipistrellus Schreber, 1774
|
108
|
49
|
| Нетопырь лесной - Pipistrellus nathusii Keyserling, Blasius, 1839
|
21
|
9
|
| Кожан поздний - Eptesicus serotinus Schreber, 1774
|
41
|
18
|
| Всего
|
222
|
100
|
Таблица 8.2.3.2
Результаты учета рукокрылых паутинными сетями на поле в квартале 9 участка «Лес на Ворскле» и на центральной усадьбе заповедника (сведения Влащенко А.С.)
| Наименование вида
|
Учтено особей
|
Обилие, %
|
| Вечерница малая - Nyctalus leisleri Kuhl, 1817
|
1
|
8
|
| Вечерница рыжая - Nyctalus noctula Schreber, 1774
|
4
|
31
|
| Нетопырь-карлик - Pipistrellus pipistrellus Schreber, 1774
|
3
|
23
|
| Кожан поздний - Eptesicus serotinus Schreber, 1774
|
5
|
38
|
| Всего
|
13
|
100
|
8.2.3.2. Население мелких млекопитающих
Таблица 8.2.3.3
Результаты учетов мелких млекопитающих ловушко-линиями на участке
«Острасьевы яры» (Сведения Аксеновой Т.Г.)
| Сроки проведения
учетов
|
Отработано
ловушко-суток
|
Наименование вида
|
Учтено особей
|
Обилие, %
|
| всего
|
на 100 ловушко-суток
|
| Опушка леса в районе ловчей канавки № 1
|
| 14.06-16.06
|
100
|
Бурозубка обыкновенная
Полевка восточноевроп.
Мышь полевая
Мышь лесная
Мышь желтогорлая
Мышь домовая
Всего
|
1
14
18
9
4
1
47
|
1
14
18
9
4
1
47
|
2
30
38
19
9
2
100
|
| Нижняя часть степного склона в районе ловчей канавки № 2
|
| 17.06-19.06
|
100
|
Бурозубка малая
Полевка восточноевроп.
Мышь полевая
Мышь лесная
Всего
|
1
16
18
5
40
|
1
16
18
5
40
|
3
40
45
13
100
|
| В результате всех учетов ловушко-линиями
|
| 14.06-19.06
|
200
|
Бурозубка малая
Бурозубка обыкновенная
Полевка восточноевроп.
Мышь полевая
Мышь лесная
Мышь желтогорлая
Мышь домовая
Всего
|
1
1
30
36
14
4
1
87
|
0.5
0.5
15.0
18.0
7.0
2.0
0.5
43.5
|
1
1
34
41
16
5
1
100
|
Таблица 8.2.3.4
Результаты учетов мелких млекопитающих ловчими канавками на участке
«Острасьевы яры» (Сведения Аксеновой Т.Г.)
| Сроки проведения
учета
|
Отработано
конусо-суток
|
Наименование вида
|
Учтено особей
|
Обилие, %
|
| всего
|
на 10 конусо-суток
|
| 1
|
2
|
3
|
4
|
5
|
6
|
| Ловчая канавка № 1 (опушка леса)
|
| 14.06-19.06
|
25
|
Бурозубка обыкновенная
Полевка рыжая
Пеструшка степная
Полевка восточноевроп.
Мышь-малютка
Мышь полевая
Мышь лесная
Мышь желтогорлая
Мышь домовая
Всего
|
7
1
4
12
1
5
2
1
1
34
|
2.8
0.4
1.6
4.8
0.4
2.0
0.8
0.4
0.4
13.6
|
21
3
12
35
3
15
6
3
3
100
|
| Ловчая канавка № 2 (степь)
|
| 14.06-19.06
|
25
|
Бурозубка обыкновенная
Слепыш обыкновенный
Хомячок серый
Пеструшка степная
Полевка восточноевроп.
Мышь полевая
Мышь лесная
Мышь домовая
Всего
|
1
1
2
4
15
1
5
3
32
|
0.4
0.4
0.8
1.6
6.0
0.4
2.0
1.2
12.8
|
3
3
6
13
47
3
16
9
100
|
| В результате всех учетов ловчими канавками
|
| 14.06-19.06
|
50
|
Бурозубка обыкновенная
Слепыш обыкновенный
Хомячок серый
Полевка рыжая
Пеструшка степная
Полевка восточноевроп.
Мышь-малютка
Мышь полевая
Мышь лесная
Мышь желтогорлая
Мышь домовая
Всего
|
8
1
2
1
8
27
1
6
7
1
4
66
|
1.6
0.2
0.4
0.2
1.6
5.4
0.2
1.2
1.4
0.2
0.8
13.2
|
12
2
3
2
12
41
2
9
11
2
6
100
|
8.2.3.3. Население норных хищников
Таблица 8.2.3.5
Результаты учетов выводков норных хищников на участке «Лес на Ворскле»
| Наименование вида
|
Учтено выводков
|
Номер норы*
|
| Лисица обыкновенная
|
3
|
1.1, 3.2, 10.1
|
| Барсук
|
1
|
6.4
|
Примечание. * Первая цифра номера норы указывает квартал, где она расположена, вторая - порядковый номер норы в квартале.
8.2.3.4. Население диких копытных
Абсолютный учёт численности диких копытных в 2004 г. проводился методом прогона (Жарков,1952). Обследование проводилось поквартально ( за исключением кварталов 1, 4, 5, 9, 10, где из-за величины или конфигурвции территории квартала прогон проводился в два этапа) с очерёдностью, исключающей, либо минимизирующей повторный учёт животных. Загонщики раставлялись с интервалом около 100 м, учётчики – в зависимости от рельефа и захламлённости просек (Шаповалов и др., 1997).
Таблица 8.2.3.6
Результаты учетов численности диких копытных на участке «Лес на Ворскле»*
| №
п/п
|
Место
|
Дата
|
Учтено особей
|
| кабан
|
косуля
|
лось
|
| абс.
|
отн.,%
|
абс.
|
отн.,%
|
абс.
|
отн.,%
|
| 1.
|
Северная часть квартала 1
|
31.01
|
19
|
-
|
7
|
-
|
0
|
-
|
| 2.
|
Южная часть квартала 1
|
31.01
|
6
|
-
|
8
|
-
|
0
|
-
|
| 3.
|
Квартал 2
|
28.01
|
0
|
-
|
0
|
-
|
0
|
-
|
| 4.
|
Квартал 3
|
28.01
|
3
|
-
|
2(3)
|
-
|
0
|
-
|
| 5.
|
Северная часть квартала 4
|
31.01
|
6(14)
|
-
|
3(5)
|
-
|
0
|
-
|
| 6.
|
Южная часть квартала 4
|
31.01
|
(1)
|
-
|
5(8)
|
-
|
0
|
-
|
| 7.
|
Восточная часть квартала 5
|
28.01
|
10
|
-
|
12(17)
|
-
|
0
|
-
|
| 8.
|
Западная часть квартала 5
|
28.01
|
0
|
-
|
1(2)
|
-
|
0
|
-
|
| 9.
|
Квартал 6
|
29.01
|
15
|
-
|
19
|
-
|
0
|
-
|
| 10.
|
Квартал 7
|
27.01
|
6
|
-
|
13(15)
|
-
|
0
|
-
|
| 11.
|
Квартал 8
|
27.01
|
0
|
-
|
6
|
-
|
0
|
-
|
| 12.
|
Квартал 9
|
29.01
|
13
|
-
|
22
|
-
|
0
|
-
|
| 13.
|
Квартал 10
|
-
|
0
|
-
|
(10)**
|
-
|
-
|
-
|
| 14.
|
Участок сада к северу от леса (охранная зона)
|
01.02
|
23(115)
|
-
|
18(90)***
|
-
|
4
|
-
|
| 16.
|
Сад к востоку от лесного массива (охранная зона)
|
01.02
|
0
|
-
|
16(19)
|
-
|
0
|
-
|
| Всего
|
-
|
101(202)
|
43(46)
|
132(231)
|
56(53)
|
4
|
2(1)
|
| Плотность на 1 тыс. га лесопокрытой площади (997 га) заповедника
|
-
|
101(202)
|
-
|
132(231)
|
-
|
4
|
-
|
| Плотность на 1 тыс. га территории заповедника и охранной зоны (1526 га)
|
-
|
66(132)
|
-
|
87(151)
|
-
|
3
|
-
|
Примечание.
* На других участках заповедника учеты диких копытных не проводились.
** В общую численность косули включена ориентировочная численность косули в 10 квартале (10 особей), где учет не проводился.
*** В садах к северу от лесного массива заповедника (охранная зона) учет проводится на участке, примыкающем к кварталам 1, 2 и ограниченном долиной р.Локня и лесополосами. При экстраполяции на всю территорию садов к северу от заповедника, входящих в охранную зону, численность кабана определена в 115 особей, косули - 90 особей.
8.3.ПОПУЛЯЦИИ ОТДЕЛЬНЫХ ВИДОВ
8.3.1. Бурозубка обыкновенная -
Sorex
araneus
Linnaeus
, 1758
Таблица 8.3.1
Сведения о половозрастной структуре популяции бурозубки обыкновенной
на участке «Острасьевы яры» (сведения Аксеновой Т.Г.)
| Пол
|
Учтено особей
|
| Всего
абс./ отн., %
|
Ad
|
subad
|
juv
|
| абс.
|
отн., %
|
абс.
|
отн., %
|
абс.
|
отн., %
|
| Самец
|
4
50
|
4
|
100
|
0
|
-
|
0
|
-
|
| Самка
|
4
50
|
3
|
75
|
1
|
25
|
0
|
-
|
| Итого
|
8
100
|
7
|
88
|
1
|
13
|
0
|
-
|
8.3.2.
Вечерница
рыжая
- Nyctalus noctula Schreber, 1774
Поиск убежищ проводился во время дневных экскурсий по лесному массиву на слух и на усадьбе заповедника при помощи ультразвукового детектора в предрассветные часы. Зверьки отлавливались из дупел при помощи пластиковой ловушки с применением сигаретного дыма. В местах охоты рукокрылых отлавливали при помощи паутинных сетей. У отловленных зверьков определялся вид, пол, возраст, измерялись длинна левого предплечья и вес.
Дупла в деревьях, заселенные рукокрылыми, описывались отдельно. Определялась порода дерева, диаметр ствола на уровне груди, высота до летка, ориентация летка по сторонам света, высота и ширина летка, происхождение дупла (естественное или устроенное дятлами).
За указанный период было обнаруженно пять дупел заселенных рукокрылыми. Из них отловленно 77 особей вечерницы рыжей. Характеристика убежищ и поло-возрастная структура популяции вечерницы рыжей представленны в табл. 8.3.2.
Таблица 8.3.2
Характеристика убежищ и сведения о половозрастной структуре популяции
вечерницы рыжей на участке «Лес на Ворскле» (Сведения Влащенко А.С.)
| №
квартала
|
Порода дерева,
диаметр ствола,
см
|
Высота до летка, м
|
Ориен-тация летка
|
Размеры летка (ширина х
высота), мм
|
Проис-хожде-ние дупла
|
Учтено особей
|
| 1
|
2
|
3
|
4
|
5
|
6
|
7
|
| 3
|
дуб, 67
|
-
|
-
|
-
|
-
|
*
|
| 6
|
дуб, 38
|
9.2
|
С
|
50 х 75
|
дятел
|
12 самок ad
6 самок juv
8 самцов juv
7 неопред.
|
| 8
|
дуб, 35
|
3.8
|
С
|
43 х 47
|
дятел
|
5 самок ad
6 самок juv
6 самцов juv
1 неопред.
|
| 8
|
дуб, 41
|
6.6
|
З
|
45 х 39
|
естеств.
|
11 самок ad
12 самок juv
8 самцов juv
|
| 10
|
дуб, 38
|
4.6
|
СВ
|
45 х 45
|
дятел
|
*
|
| 1
|
2
|
3
|
4
|
5
|
6
|
7
|
| Всего
|
28 самок ad
24 самок juv
22 самцов juv
8 неопред.
|
Примечание. * Рыжие вечерницы отмечены по характерному писку из дупла, но не были отловленны.
Таблица 8.3.3
Морфометрические данные особей вечерницы рыжей, отловленных в убежищах
и паутинной сетью (сведения Влащенко А.С.)
| №
п/п
|
Пол
|
Возраст
|
Длина левого предплечья, мм
|
Вес, гр.
|
| 1
|
2
|
3
|
4
|
5
|
| Дупло дуба в квартале 6; 20.07.2004 г.
|
| 1.
|
Самка
|
ad
|
55.1
|
28.8
|
| 2.
|
Самка
|
juv
|
56.2
|
23.3
|
| 3.
|
Самка
|
ad
|
55.2
|
29.5
|
| 4.
|
Самка
|
ad
|
54.9
|
28.0
|
| 5.
|
Самка
|
ad
|
55.8
|
27.8
|
| 6.
|
Самка
|
ad
|
55.4
|
31.3
|
| 7.
|
Самка
|
ad
|
57.4
|
29.5
|
| 8.
|
Самец
|
Juv
|
52.9
|
20.9
|
| 9.
|
Самка
|
ad
|
54.8
|
29.2
|
| 10.
|
Самка
|
ad
|
50.0
|
25.2
|
| 11.
|
Самец
|
Juv
|
52.9
|
22.6
|
| 12.
|
Самка
|
ad
|
55.7
|
29.9
|
| 13.
|
Самка
|
Juv
|
55.3
|
24.8
|
| 14.
|
Самка
|
Juv
|
53.3
|
24.3
|
| 15.
|
Самец
|
Juv
|
54.2
|
25.9
|
| 16.
|
Самка
|
Juv
|
56.5
|
23.9
|
| 17.
|
Самец
|
Juv
|
53.0
|
18.5
|
| 18.
|
Самка
|
Juv
|
54.0
|
25.3
|
| 19.
|
самка
|
juv
|
54.6
|
23.9
|
| 20.
|
самец
|
juv
|
54.8
|
23.5
|
| 21.
|
самец
|
juv
|
52.6
|
19.5
|
| 22.
|
самец
|
juv
|
53.3
|
22.9
|
| 23.
|
самка
|
Ad
|
55.5
|
27.9
|
| 24.
|
самка
|
Ad
|
54.0
|
30.7
|
| 25.
|
самка
|
Ad
|
55.0
|
29.1
|
| 26.
|
самец
|
juv
|
53.9
|
25.8
|
| Таблица 8.3.3 (продолжение)
|
| 1
|
2
|
3
|
4
|
5
|
| Дупло дуба в квартале 8; 21.07.2004 г.
|
| 27.
|
самка
|
juv
|
53.4
|
21.7
|
| 28.
|
самка
|
juv
|
49.3
|
16.8
|
| 29.
|
самец
|
juv
|
53.2
|
21.6
|
| 30.
|
самка
|
Ad
|
56.5
|
28.2
|
| 31.
|
самка
|
juv
|
53.2
|
22.1
|
| 32.
|
самец
|
juv
|
52.9
|
24.0
|
| 33.
|
самка
|
juv
|
52.6
|
22.3
|
| 34.
|
самец
|
juv
|
53.0
|
22.4
|
| 35.
|
самец
|
juv
|
52.9
|
22.0
|
| 36.
|
самец
|
juv
|
56.2
|
20.0
|
| 37.
|
самка
|
Ad
|
54.1
|
26.5
|
| 38.
|
самка
|
Ad
|
52.5
|
24.9
|
| 39.
|
самец
|
juv
|
50.7
|
19.5
|
| 40.
|
самка
|
Ad
|
53.0
|
29.1
|
| 41.
|
самка
|
Ad
|
54.8
|
28.0
|
| 42.
|
самка
|
juv
|
54.9
|
22.5
|
| 43.
|
самка
|
Ad
|
52.9
|
26.3
|
| 44.
|
самка
|
juv
|
53.8
|
22.6
|
| Дупло дуба в квартале 8; 23.07.2004 г.
|
| 45.
|
самка
|
juv
|
53.0
|
24.4
|
| 46.
|
самец
|
juv
|
52.4
|
27.4
|
| 47.
|
самка
|
juv
|
54.0
|
24.6
|
| 48.
|
самка
|
juv
|
51.4
|
20.3
|
| 49.
|
самка
|
Ad
|
55.0
|
31.3
|
| 50.
|
самка
|
Ad
|
54.5
|
31.0
|
| 51.
|
самец
|
juv
|
55.3
|
25.9
|
| 52.
|
самец
|
juv
|
53.4
|
26.5
|
| 53.
|
самка
|
juv
|
52.4
|
21.1
|
| 54.
|
самка
|
juv
|
56.4
|
25.7
|
| Таблица 8.3.3 (окончание)
|
| 1
|
2
|
3
|
4
|
5
|
| 55.
|
самец
|
juv
|
52.5
|
23.1
|
| 56.
|
самка
|
juv
|
50.3
|
15.5
|
| 57.
|
самка
|
juv
|
55.0
|
24.5
|
| 58.
|
самка
|
Ad
|
54.4
|
30.3
|
| 59.
|
самка
|
Ad
|
55.2
|
30.7
|
| 60.
|
самка
|
Ad
|
55.5
|
30.1
|
| 61.
|
самка
|
juv
|
54.9
|
24.8
|
| 62.
|
самка
|
Ad
|
51.0
|
26.5
|
| 63.
|
самка
|
Ad
|
57.7
|
32.0
|
| 64.
|
самка
|
Ad
|
54.6
|
28.1
|
| 65.
|
самка
|
juv
|
53.5
|
25.5
|
| 66.
|
самец
|
juv
|
53.5
|
25.0
|
| 67.
|
самец
|
juv
|
55.2
|
26.6
|
| 68.
|
самка
|
juv
|
53.7
|
21.9
|
| 69.
|
самка
|
juv
|
55.7
|
27.3
|
| 70.
|
самка
|
Ad
|
52.6
|
26.9
|
| 71.
|
самец
|
juv
|
53.7
|
26.9
|
| 72.
|
самец
|
juv
|
54.4
|
24.9
|
| 73.
|
самец
|
juv
|
53.0
|
24.8
|
| 74.
|
самка
|
juv
|
55.3
|
26.0
|
| 75.
|
самка
|
Ad
|
54.3
|
31.6
|
| 76.
|
самка
|
juv
|
54.2
|
24.5
|
| 77.
|
самка
|
Ad
|
55.3
|
29.8
|
| Паутинная сеть на поле в квартале 9; 24-25.07.2004 г.
|
| 78.
|
самка
|
juv
|
54.3
|
35.1
|
| 79.
|
самец
|
juv
|
54.0
|
25.7
|
| 80.
|
самец
|
juv
|
53.0
|
24.6
|
| 81.
|
самка
|
juv
|
54.9
|
25.2
|
8.3.3.
Заяц
-
русак
- Lepus europaeus Pallas, 1778
Учёты численности зайца-русака на участке «Лес на Ворскле» в 2004 г. проводились одновременно с учётами численности диких копытных (см. подпункт 8.2.2.4).
Таблица 8.3.4
Результаты учетов численности зайца-русака на участке «Лес на Ворскле»
| №
п/п
|
Место
|
Дата
|
Учтено особей
|
| 1.
|
Северная часть квартала 1
|
31.01
|
0
|
| 2.
|
Южная часть квартала 1
|
31.01
|
1
|
| 3.
|
Квартал 2
|
28.01
|
0
|
| 4.
|
Квартал 3
|
28.01
|
1
|
| 5.
|
Северная часть квартала 4
|
31.01
|
2(3)
|
| 6.
|
Южная часть квартала 4
|
31.01
|
3(4)
|
| 7.
|
Восточная часть квартала 5
|
28.01
|
1
|
| 8.
|
Западная часть квартала 5
|
28.01
|
1
|
| 9.
|
Квартал 6
|
29.01
|
3(4)
|
| 10.
|
Квартал 7
|
27.01
|
0
|
| 11.
|
Квартал 8
|
27.01
|
1(2)
|
| 12.
|
Квартал 9
|
29.01
|
0
|
| 14.
|
Квартал 10
|
-
|
(1)
|
| 15.
|
Участок сада к северу от леса *
|
01.02
|
1(5)
|
| 16.
|
Сад к востоку от лесного массива
|
01.02
|
5(6)
|
| Всего
|
-
|
19(29)
|
Примечание. *В садах к северу от лесного массива заповедника (охранная зона) учет проводится на участке, примыкающем к кварталам 1, 2 и ограниченном долиной р.Локня и лесополосами. При экстраполяции на всю территорию садов к северу от заповедника, входящих в охранную зону, численность зайца-русака определена в 5 особей.
8.3.4. Пеструшка степная -
Lagurus
lagurus
Pallas
, 1773
Таблица 8.3.5
Сведения о половозрастной структуре популяции пеструшки степной
на участке «Острасьевы яры» (сведения Аксеновой Т.Г.)
| Пол
|
Учтено особей
|
| всего
абс./ отн., %
|
ad
|
subad
|
juv
|
| абс.
|
отн., %
|
абс.
|
отн., %
|
абс.
|
отн., %
|
| Самец
|
2
25
|
1
|
50
|
1
|
50
|
0
|
-
|
| Самка
|
6
75
|
1
|
17
|
5
|
83
|
0
|
-
|
| Всего
|
8
100
|
2
|
25
|
6
|
75
|
0
|
-
|
8.3.5. Полевка восточноевропейская -
Microtus
rossiaemeridionalis
Ognev
, 1924
Таблица 8.3.6
Сведения о половозрастной структуре популяции полевки восточноевропейской
на участке «Острасьевы яры» (сведения Аксеновой Т.Г.)
| Пол
|
Учтено особей
|
| всего
абс./ отн., %
|
Ad
|
subad
|
juv
|
| абс.
|
Отн., %
|
абс.
|
отн., %
|
абс.
|
отн., %
|
| Самец
|
36
63
|
23
|
64
|
10
|
28
|
3
|
8
|
| Самка
|
21
37
|
14
|
67
|
4
|
19
|
3
|
14
|
| Всего
|
57
100
|
37
|
65
|
14
|
25
|
6
|
11
|
8.3.6.
Мышь
полевая
- Apodemus agrarius Pallas, 1771
Таблица 8.3.7
Сведения о половозрастной структуре популяции мыши полевой
на участке «Острасьевы яры» (сведения Аксеновой Т.Г.)
| Пол
|
Учтено особей
|
| всего
абс./ отн., %
|
ad
|
subad
|
juv
|
| абс.
|
отн., %
|
абс.
|
отн., %
|
абс.
|
отн., %
|
| Самец
|
21
51
|
13
|
62
|
2
|
10
|
6
|
29
|
| Самка
|
20
49
|
9
|
45
|
4
|
20
|
7
|
35
|
| Всего
|
41
100
|
22
|
54
|
6
|
15
|
13
|
32
|
8.3.7.
Мышь
лесная
- Apodemus uralensis Pallas, 1811
Таблица 8.3.8
Сведения о половозрастной структуре популяции мыши лесной
на участке «Острасьевы яры» (сведения Аксеновой Т.Г.)
| Пол
|
Учтено особей
|
| всего
абс./ отн., %
|
ad
|
subad
|
juv
|
| абс.
|
отн., %
|
абс.
|
отн., %
|
абс.
|
отн., %
|
| Самец
|
15
71
|
10
|
67
|
5
|
33
|
0
|
-
|
| Самка
|
6
29
|
4
|
67
|
2
|
33
|
0
|
-
|
| Всего
|
21
100
|
14
|
67
|
7
|
33
|
0
|
-
|
10. СОСТОЯНИЕ ЗАПОВЕДНОГО РЕЖИМА.
Влияние антропогенных
факторов на природу заповедника и охранной зоны
10.3. ПРЯМЫЕ И КОСВЕННЫЕ ВНЕШНИЕ ВОЗДЕЙСТВИЯ (пожары)
Таблица 10.3.1
Пожары в заповеднике в 2004 году
| №
п/п
|
Тип пожара, причина и
время возникновения
|
Участок, урочи- ще, квартал, вы- дел, характер растительности
|
Выгорев-шая пло- щадь,
га
|
Средства ту- шения, число учасвовавших
людей
|
Последствия
|
| 1
|
Низовой беглый, с/х пал, 16.04.2004 г.
|
Уч. «Острасье-
вы яры», приус-
тьевая С-З часть
уч-ка, остепнён-
ный луг
|
24.0
|
Самозатуха-ние пожара
|
Уничрожение
травяной подстилки и ветоши,
задымление атмосферы
|
| Итого: 24.0 га
|
Список использованных литературных и архивных материалов
- Абрамов И.И., Волкова Л.А. Определитель листостебельных мхов Карелии. – М.: КМК, 1998. – 390 с.
- Абрамова А.Л., Савич-Любицкая Л.И., Смирнова З.Н. Определитель листостебельных мхов Арктики СССР. – М.-Л., 1961. – 716 с.
- Алексенко М.А. Лиственные мхи северной части Харьковской губернии и смежных с ней уездов Курской губернии // Тр. о-ва испыт. природы при Харьковском ун-те. - №31, 1897. - С. 3-23.
- Алехин В.В., Сырейщиков Д.П. Методика полевых ботанических исследований. – Вологда: “Северный печатник”, 1926. - 141 с.
- Бардунов Л.В. Определитель листостебельных мхов Центральной Сибири. – Л., 1969. – 319 с.
- Бобырева С.В. Материалы к лихенофлоре заповедника «Лес на Ворскле» // Тезисы конференции, посвященной 85 - летию биостанции ХГУ. – Харьков, 1999а – с. 29-30.
- Бобырева С.В. Лихенофлора заповедника «Лес на Ворскле» - Дипломная работа. Харьков, 1999б. – 87 с.
- Дидух Я.П., Лысенко Г.Н. Экологические проблемы степей Украины. / Степи Евразии: Проблемы сохранения и восстановления. СПб-М.: Институт географии РАН, 1993. – 146 с.
- Доронина Ю.А., Нешатаев Ю.Н., Ухачева В.Н. Флористический список степной балки “Астрасьев яр” (Борисовский район Белгородской области) / Вестник С-Петерб. ун. Сер. 3. Вып. 4 (№24). – С. 50-55.
- Жарков И.В. Основные методы учёта диких копытных // Методы учёта численности и географического распределения наземных позврнрчных. М.: Изд-во АН СССР, 1952. - С. 214 - 238.
- Золотухин Н.И., Золотухина И.Б. Состав и многолетняя динамика флоры Ямского заповедного участка (Белгородская область) // «Летопись природы» ГПЗ «Белогорье», 2005. С.35-47.
- Золотухин Н.И.. Золотухина И.Б. Сосудистые растения / Природа Лысых гор – нового заповедного участка в Белгородской области // Тр. Центрально-Черноземного запов., вып. 14. – Москва, КМК Scientific Press Ltd., 1995. – С. 29-44.
- Игнатов М.С., Афонина О.М. Список мхов территории бывшего СССР // Arctoa. – №1, 1992. – С. 1-87.
- Игнатов М.С., Игнатова Е.А. Флора мхов средней части европейской России. – М.: КМК, 2003. Том 1. Sphagnaceae-Hedwigiaceae – 608 с.
- Кашменский Б.Ф. Лишайники Курской и Харьковской губернии // Ботанический журнал общества естествоиспытателей. - №3, 1906. – С. 6-25.
- Конорева Л.А. Дополнение к лихенофлоре заповедника “Лес на Ворскле” // материалы межвузовской конференции молодых ученых “Растение, микроорганизмы и среда”. – СПб, 1999. - С.36-37.
- Конорева Л.А. Лишайники заповедника “Лес на Ворскле” // Первая Российская Лихенологическая Школа и Международный Симпозиум Молодых Лихенологов “Аркто-альпийская флора. Охрана лишайников”. - Апатиты, 2000. – С.38-39.
- Конорева Л.А. «Лишайники заповедника «Белогорье» (заповедный участок «Лес на Ворскле») // Труды Первой Российской лихенологической школы. - Петрозаводск, 2001. – С. 67-85.
- Конорева Л.А. К изучению лишайников заповедника «Белогорье» (заповедный участок «Лес на Ворскле») // Сборник студенческих научных работ, выпуск 5, Белгород, 2002. С. 57-58.
- Конорева Л.А., Мучник Е.Э. Материалы к изучению лишайников Айдарского участка // Роль особо охраняемых природных территорий Центрального Черноземья в сохранении и изучении биоразнообразия лесостепи: Материалы научно-практической конференции, посвященной 75-летию Воронежского государственного природного биосферного заповедника. – Воронеж, издательство «Кривичи», 2002а. – С. 49-56.
- Конорева Л.А., Мучник Е.Э. Материалы к изучению лишайников заповедного участка «Лысые горы» (заповедник «Белогорье») // Тезисы Пущинской школы-конференции молодых ученых «Биология – наука XXI века». – т. 2, Пущино, 2002б. – С. 88-89.
- Конорева Л.А., Мучник Е.Э. Лихенобиота заповедника «Белогорье» и его окрестностей // Бореальная лихенофлора. Лихеноиндикация. Программа и тезисы докладов третьей международной лихенологической школы и симпозиума. – Екатеринбург: Изд-во Уральского ун-та, 2002в. – С. 42.
- Краснитский А.М. и др. Принципы охраны и современное состояние травяных экосистем Европейской лесостепи // Проблемы охраны генофонда и управления экосистемами в заповедниках степной и пустынной зон. Тезисы докладов Всесоюзного совещания (21 – 25 мая 1984 г. Аскания-Нова). М.:, 1984. С.131 –135.
- Логутов Д.П., Моисеенко Ф.П. Сортиментные таблицы для таксации леса на корню. Киев: Госстройиздат, 1959. 685 с.
- Макарова И.И. Лишайники учлесхоза «Лес на Ворскле» // Ученые записки ЛГУ, №351, вып. 52, том V. – ЛГУ, 1971. – С. 32 – 40.
- Максимов Л.Д. Типы леса заповедника “Лес на Ворскле” // Учен. зап. ЛГУ. Сер. биол. наук, №28(7), 1939 – С. 17-67.
- Мельничук В.М. Определитель лиственных мхов средней полосы и юга европейской части СССР. – Киев, 1970. – 442 с.
- Мучник Е.Э. Лихенологические исследования нового заповедного участка «Стенки-Изгорья» (Центрально-Черноземный заповедник) // Материалы Российско-Украинской научной конференции «Проблемы сохранения разнообразия природы степных и лесостепных регионов». - М., 1995.- С.87-88.
- Мучник Е.Э., Конорева Л.А. К изучению лишайников степных участков заповедника «Белогорье» (Белгородская обасть) // Материалы III Международного симпозиума «Степи Северной Евразии. Эталонные степные ландшафты: проблема охраны, экологической реставрации и использования. – Оренбург, ИПК «Газпромпечать» ООО «Оренбурггазпромсервис», 2003. – С. 356-358.
- Мучник Е.Э., Урбанавичюс Г.П. Находка Leptogium schraderi (Collemataceae, Lichenes) в России // Бот. журн. – 2001, Т.86, №3. – С. 120-121.
- Окснер А.Н. Определитель лишайников СССР. - Л.: Наука, 1974. - Вып. 2: Морфология, систематика и географическое распространение. - 281 с.
- Окснер А.М. Флора лишайникiв Украiнi. - Киев: Наукова думка, 1956. Т.1 - 496 с.
- Окснер А.М. Флора лишайникiв Украiнi. - Киев: Наукова думка, 1993. Т 2 - 500 с.
- Определитель лишайников СССР. – Вып. 1 Пертузариевые. Леканоровые. Пармелиевые. – Л.: Наука, 1971. – 410с.
- Определитель лишайников СССР. – Вып. 3. Калициевые. Гиалектовые. – Л.: Наука, 1975. – 275с.
- Определитель лишайников СССР. – Вып. 4. Веррукариевые. Пилокарповые. - Л.: Наука, 1977. – 343 с.
- Определитель лишайников СССР. – Вып. 5. Кладониевые. Акаросповые. – Л.: Наука, 1978. – 303 с.
- Определитель лишайников России. – Вып. 6. Санкт–Петербург: Наука, 1995. – 203 с.
- Определитель лишайников России. – Вып. 7. Санкт–Петербург: Наука, 1996. – 166 с.
- Определитель лишайников России – Вып. 8. Санкт-Петербург: Наука, 2003. – 277 с.
- Определитель лишайников России – Вып. 9. Санкт-Петербург: Наука, 2004. – 135 с.
- Определитель сосудистых растений центра Европейской России. Изд. 2-е, доп. и перераб. (авторы: И.А. Губанов, К.В. Киселёв, В.С. Новиков, В.Н. Тихомиров). – М.: «Аргус», 1995. 558 с.
- Павлинов И.Я., Крускоп С.В., Варшавский А.А., Борисенко А.В. Наземные звери России. Справочник-определитель. - М.:Изд-во КМК, 2002. - 298 с.
- Покровская В.М. Описание растительности Ямской степи / Тр. Центрально-Черноземного гос. заповедника. – М., 1940. – Вып. 1. С. 369-407.
- Попова Н.Н. Бриофлора Среднерусской возвышенности. I // Arctoa, №11. – 2002. – С. 101-168.
- Проект организации и ведения лесного хозяйства учебно-опытного лесхоза «Лес на Ворскле» ЛГУ. Том I. Объяснительная записка. I-я Воронежская лесоустроительная экспедиция. - Воронеж, 1976-1977. 493 с.
- Проект организации и ведения лесного хозяйства учебно-опытного лесхоза «Лес на Ворскле» ЛГУ. Постоянные пробные площади. Том IV. Книга VI. Воронеж, 1977. 798 с.
- Рыковский Г.Ф. Мохообразные Березинского биосферного заповедника. – Минск, 1980. – 135 с.
- Савич-Любицкая Л.И., Смирнова З.Н. Определитель листостебельных мхов СССР. Верхоплодные мхи. – Л., 1970. – 826 с.
- Семенова Тян-Шанская А.М., Никольская Н.И. Состав и динамика надземной массы травостоев в степях и на лесных полянах./ Труды ЦЧЗ им.проф.В.В.Алехина. Вып.6., 1960 г. С.82 – 114.
- Семенова Тян-Шанская А.М. Динамика степной растительности. М. – Л.: Наука, 1966. 174 с.
- Собакинских В.Д. Изучение динамики травянистых сообществ в заповедниках. Методическое пособие. Почвенный и биотический мониторинг заповедных экосистем. М.: КМК Scientific Press Ltd., 1996. С.39-47.
- Собакинских В.Д. Динамика надземной фитомассы луговой степи в Центрально-Черноземном заповеднике (1956 – 1995 годы).// Многолетняя динамика природных процессов и биологическое разнообразие заповедных экосистем Центрального Черноземья и Алтая: Труды Центрально-Черноземного государственного заповедника. Вып.15. М., 1997. С. 65 – 73.
- Третьяков Н.В., Горский П.В., Самойлович Г.Г. Справочник таксатора. Таблицы для таксации леса. Второе, переработанное издание. М.: Изд-во “Лесная промышленность”, 1965. 457 с.
- Тюрин А.В., Науменко И.М., Воропанов П.В. Лесная вспомогательная книжка. М.: Гослестехиздат, 1946. 407 с.
- Филатова Т.Д. и др. Растительность залежей Центрально_Черноземного заповедника./Растетельный покров Центрально-Черноземного заповедника: Труды Центрально-Черноземного государственного заповедника. Выпуск 18. Тула: Гриф и К0
, 2001 г. – С.23 – 81.
- Флора СССР // Под ред. Б.К. Шишкина, С.В. Юзепчук. – Т. ХХ. – М.-Л., 1954. – 555 с.
- Флора СССР // Под ред. Б.К. Шишкина, С.В. Юзепчук. – Т. ХХI. – М.-Л., 1954. – 703 с.
- Хмелев К.Ф., Попова Н.Н. Флора мохообразных бассейна Среднего Дона. – Воронеж, 1988. – 168 с.
- Черепанов С.К. Сосудистые растения России и сопредельных государств (в пределах бывшего СССР). – С-Пб.: “Мир и семья - 95”, 1995. – 992 с.
- Шаповалов А.С., Леонтьева О.А., Абсалон Д.А., Гонгальский К.Б., Кузнецов В.А., Панов И.Н. К оценке численности и опримизации методического обеспечения учётов численности диких копытных в заповеднике «Лес на Ворскле» // Проблемы сохранения и оценки состояния природных комплексов и объектов. Матеоиалы научно-практической конференции, посвящённой 70-летию Воронежского заповедника, 1997. С. 128-129.
- Eriksson O.E., Hawkswort D.L. Outline of the Ascomycetes // Systema Ascomycetum. Vol. 16, part 1-2. – 1998. – 161 p.
- Foucard T. Svenska scorplavar och svampar som vaxer pa dem. – Stockholm: Interpublishing, 2001. – 392 р.
- Santesson R., Moberg R., Nordin A. at al. Lichenforming and lichenicolous fungi of Fennoscandia. Museum of Evolution, Uppsala University, 2004. – 360 p.
- Servit M. Ceskoslovenske Lisejnikj celedi Verrucariaceae Lichenes Familiae Verrucariacearum. – Nakladatelstyf Ceskoslovenske Academie ved Praha, 1954. – 121 p.
- With V. Die Flechten Baden-Wŭrttembergs. Teil 1, 2. Stuttgart: Ulmer, 1995. – 1006 s.
|
