Зорина Зоя Александровна, Полетаева Инга Игоревна
Общая характеристика тех проявлений индивидуальной приспособительной деятельности, в основе которых лежит обучение. Краткое описание классических и инструментальных условных рефлексов. Примеры методов исследования процессов обучения, в частности анализируется формирование пространственных представлений. Ассоциативные процессы, отражающие механизм образования условных рефлексов, сопоставляются с основными видами когнитивной деятельности. Описание методов обучения (дифференцировки и их системы, выбор по образцу), по которым можно судить о способности животных к обобщению и умозаключению.
Индивидуально-приспособительная деятельность животного, т.е. адаптация особи к конкретным условиям среды, позволяет ему с большим или меньшим успехом выживать, преодолевая трудности и опасности повседневного существования. Эта деятельность многообразна по составу и включает приобретенные компоненты разной природы. Рассмотрим виды индивидуально-приспособительного поведения, в основе которых лежат процессы обучения.
1. Обучение и пластичность.
Из существующих в настоящее время определений феномена «обучение» предпочтение отдается определению У. Торпа (Thorpe, 1963).
Обучение — это появление адаптивных изменений индивидуального поведения в результате приобретения опыта.
На обучении основаны события естественной жизни животных, например формирование навыков отыскания определенной пищи, избегания опасных участков местности, выбора удобных троп, взаимодействия с сородичами и животными других видов и т.п. В лабораторных опытах можно наблюдать, как животное обучается действиям, заданным экспериментатором. Примерами могут быть «слюнные» условные рефлексы собаки, т.е. выделение слюны уже в момент попадания животного в комнату, где в процессе опытов оно получало подкормку; отыскание крысой выхода из лабиринта; избегание болевого раздражения; клевание птицей кнопки при действии определенных стимулов и т.п.
Способность к обучению базируется на присущем центральной нервной системе свойстве пластичности.
Пластичность — это свойство системы изменять свою реакцию на внешние воздействия как результат тех или иных внутренних преобразований на основе предшествующих воздействий.
Реклама

Она проявляется в способности системы изменять реакции на повторяющийся многократно раздражитель, а также в случаях его совместного действия с другими факторами. Пластичность может иметь разную направленность: чувствительность к раздражителю может повышаться — это явление называется сенсишизацией, или снижаться, тогда говорят о привыкании.
По определению нейрофизиологов, анализирующих пластические изменения в мозге (Конорски, 1970; Котляр, 1986),
пластичность — это относительно устойчивые функциональные изменения в системах нейронов, которые по длительности превышают время обычных синаптических процессов и определяют эффективность и направленность межнейронных связей.
Это достаточно формальное определение показывает, что изменения ответа системы при повторном действии стимула можно описать на языке математики.
2. Классификация форм индивидуально-приспособительной деятельности.
Формы обучения животных весьма разнообразны и обычно их подразделяют на три основные категории: неассоциативное обучение, ассоциативное обучение и когнитивные процессы. О. Меннинг (1982) и Д. Дьюсбери включали в свои классификации также и «инсайт-обуче-ние» (см. 3.4.5). Приводим систематизированный перечень этих феноменов (по данным разных авторов).
Классификация включает некоторые сведения об элементах рассудочной деятельности, которым посвящена основная часть пособия. Ряд авторов выделяют в своих книгах раздел «Complex cognition», который соответствует разделу «Когнитивные процессы» настоящей главы, а также содержанию ряда следующих глав, где рассматривается формирование у животных понятий, усвоение языков-посредников и т.д.
Для изучения механизмов обучения нередко используют различные модели, в том числе так называемые клеточные аналоги обучения или процесс посттетанической потенциации (см. гл. 9 и, например: Шеперд, 1987).
Классификация форм обучения и когнитивных процессов [составлена нами на основе классификаций О. Меннинга (Manning, Dawkins, 1992), — Д. Дьюсбери (1981), Р. Томаса (Thomas, 1996), Дж. Пирса (Реагсе, 1998) и др.].
> Неассоциативное обучение:
— привыкание.
> Ассоциативное обучение:
классические условные рефлексы;
инструментальные условные рефлексы.
> Когнитивные процессы:
латентное обучение;
выбор по образцу;
обучение, основанное на представлениях о:
* пространстве;
* порядке стимулов;
* времени (Обширный экспериментальный материал по восприятию и оценке животными параметров времени в книге не затронут, поскольку эти вопросы практически не имеют прямой связи с проблемой мышления животных);
Реклама

* числе.
— элементарное мышление (Классификация видов мышления животных рассматривается в главе 4)..
Далее рассматриваются основные формы обучения, приведенные в схеме классификации.
2.1. Неассоциативное обучение (привыкание).
Неассоциативное обучение (привыкание) заключается в ослаблении реакции при повторных предъявлениях раздражителя.
Изначально любой раздражитель (стимул), действующий на органы чувств животного, вызывает у него соответствующую ответную реакцию: поворот головы в сторону света или звука, отдергивание конечности и др., у моллюсков — втягивание жабр и т.д. При повторном систематическом предъявлении того же стимула реакция постепенно ослабевает и может исчезнуть совсем, т.е. происходит привыкание к раздражителю. Так, например, только что установленное в огороде чучело распугивает птиц, однако с течением времени их страх ослабевает и они перестают реагировать на этот предмет.
Привыкание — наиболее примитивная форма обучения, отчетливо выраженная даже у низших организмов — беспозвоночных животных. Его успешно исследуют на имеющих простую нервную систему ки-шечно-полостных, червях, моллюсках, насекомых. Для изучения привыкания используются также модельные системы — так называемые клеточные аналоги привыкания или срезы мозга.
Более строго привыкание определяют как снижение вероятности появления реакции или уменьшение интенсивности при неоднократном повторении вызывающего ее раздражителя. Ослабление ответной реакции можно считать истинным привыканием только в том случае, когда оно обусловлено изменениями в ЦНС, а не адаптацией рецепторов или утомлением.
Применение какого-либо нового стимула прекращает процесс привыкания к прежнему раздражителю, и угасшая было реакция на исходный раздражитель полностью восстанавливается. Для привыкания характерно и так называемое спонтанное восстановление, если действие стимула временно прекращается. Более подробные сведения о свойствах процесса привыкания можно найти в учебниках Кэнделла (1987), Шеперда (1987) и Эккерта и др. (1991).
2.2. Ассоциативное обучение.
При ассоциативном обучении в ЦНС формируется временная связь между двумя стимулами, один из которых изначально был для животного безразличен, а другой выполнял роль вознаграждения или наказания.
Формирование этой связи обнаруживается в виде изменений в поведении животного, которые в зависимости от своей «структуры» называются либо классическими, либо инструментальными условными рефлексами.
2.2.1. Основные характеристики условнорефлекторной деятельности
В 1902 году, анализируя нервную регуляцию процесса пищеварения у собак, И.П.Павлов обнаружил так называемое «психическое слюноотделение». Феномен заключался в выделении у животного слюны, которая стекала в пробирку через фистулу на щеке еше до попадания пищи в рот, т.е. заранее, «в опережающем режиме», как только собаку приводили в экспериментальную комнату и помещали в специальный станок (рис.1).
Таким же образом повторное сочетание любого нейтрального для животного раздражения (например, звонка) с кормлением вызывает выделение слюны еще до того, как собака получит пищу.
И. П. Павлов назвал звонок условным сигналом (УС), пищу — безусловным раздражителем (или стимулом), реакцию животного (слюноотделение) в ответ на предъявление пищи — безусловным рефлексом (БР), а в ответ на звонок — условным рефлексом (УР).
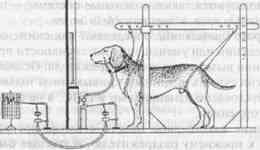
Рис. 1. Схема установки по выработке слюнных условных рефлексов в экспериментах И. П. Павлова.
Через фистулу в щеке слюна поступает в устройство, измеряющее ее объем. Перед животным имеется панель, где расположены источники нейтральных раздражителей (света и звука), которые могут стать УС.
Принцип образования условного рефлекса состоит в следующем: действие какого-либо нейтрального (или индифферентного) для животного раздражителя совместно со стимулом, вызывающим у него определенную реакцию (например, отдергивание конечности при болевой стимуляции), приводит к тому, что постепенно этот ранее нейтральный стимул начинает вызывать такую же реакцию.
Предъявление безусловного стимула вслед за условным в процессе выработки УР называется его подкреплением. Если при выработке УР применяется подкрепление, соответствующее имеющейся у животного мотивации (например, пищевое подкрепление в состоянии голода), то оно называется положительным и вырабатываемый УР также называется положительным. Можно выработать УР и с применением отрицательного подкрепления (наказания), т.е. такого воздействия, которого животное стремится избежать. В эксперименте в качестве отрицательного подкрепления чаще всего применяют удар электрического тока, вызывающий отдергивание конечности или перебегание в безопасное отделение камеры. Используют также действие воздушной струи, направленной на роговицу глаза и вызывающей мигание. Динамика реакций мигательной перепоныки кролика при выработке такого УР показана на рис. 2А.
Подкрепление, которое используется для выработки УР, оказывается эффективным, если оно применяется в период достаточно сильного мотивационного возбуждения. Так, удар воздушной струи, направленной на роговицу глаза, может стать отрицательным подкреплением, только если он действительно неприятен животному.
Слюнной и мигательный УР относятся к категории классических («павловских») условных рефлексов (см. также 2.2.2).
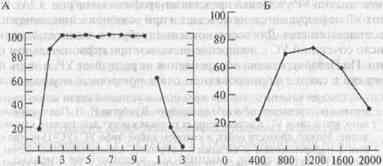
Рис. 2. Изменения условной реакции мигания у кролика.
Безусловный стимул — струя воздуха, направленная на роговицу; условный — звук; А — его угашение; Б — генерализация ответа при использовании в качестве УС тонов разных частот. По оси ординат — доля УР (в %), по оси абсцисс — число предъявлений (А) и частота тона (Б).
На основе изучения слюнных УР у собак И. П. Павлов сформулировал основные общие правила образования УР:
условный сигнал должен предшествовать безусловному раздражению, но не наоборот;
действие условного и безусловного раздражителей должно частично перекрываться во времени;
сочетание условного и безусловного раздражителей должно повторяться многократно.
Эти правила приложимы не только к классическим, но и к инструментальным (см. ниже) УР. И формирование, и проявления уже выработанных слюнных УР у собак в значительной степени чувствительны к влиянию посторонних раздражителей. В лаборатории И. П. Павлова было обнаружено, что любой посторонний и достаточно сильный стимул, подействовавший в течение эксперимента, уменьшает условнорефлекторное слюноотделение у собаки, как бы «отвлекает» ее. Павлов объяснял это явление на основе своего понимания механизма формирования УР: вмешательство постороннего стимула вызывает в коре головного мозга собаки сильный очаг возбуждения, который в силу природы условнорефлекторньгх связей подавляет уже сформированный УР, «индуцируя» торможение участка коры, ответственного за этот УР. Такое торможение И. П. Павлов назвал внешним.
Помимо внешнего торможения в лаборатории Павлова было описано и внутреннее торможение. Одно из проявлений этого процесса можно наблюдать в опытах, когда предъявление УС перестает сопровождаться подкреплением. Отмена подкрепления ведет к постепенному исчезновению внешних проявлений УР, к его угашению (ход угашения мигательного УР у кролика представлен графически на рис. 2А). Однако этот УР не разрушается, не исчезает и при возобновлении подкрепления восстанавливается. Для восстановления требуется значительно меньшее число сочетаний УС с подкреплением, чем при первоначальном обучении. По Павлову, отмена подкрепления не разрушает УР, а лишь подавляет его в связи с формированием очага внутреннего торможения.
Следует отметить, что для проявления условной связи между двумя стимулами подкрепление не всегда необходимо. В работах И. П. Павлова было показано, что и сами УС в определенных условиях могут действовать как подкрепление. Можно провести опыт, в котором какой-либо УС (УС-1) всегда сочетается с подкреплением и вызывает, например, слюноотделение. Затем некий второй УС (УС-2) многократно применяется вместе с УС-1 (без сочетания с безусловным раздражением). Если после этого применить только УС-2, можно убедиться, что он также вызывает условную реакцию. Это явление было названо условным рефлексом второго порядка.
Несколько отличается от описанного другой опыт, который также показывает, что для формирования УР подача подкрепления не всегда обязательна. Если два УС предъявлять животному совместно (УС-1 + УС-2) много раз еще до применения безусловного стимула, а затем предлагать только сочетание УС-1 и безусловного раздражения, то затем условная реакция проявится при даче одного лишь УС-2. Таким образом, хотя УС-2 никогда сам по себе не подкреплялся, между УС-1 и УС-2 сформировалась связь, которая позволила проявиться условной реакции при действии только УС-2. Процесс формирования связи между индифферентными раздражителями иногда называют сенсорным предобуславливанием {sensory preconditioning, Мак-Фарленд, 1987). Вряд ли это можно считать правильным, потому что обнаружить наличие подобной связи можно лишь с помощью соответствующего тестирования — путем выявления реакции на второй индифферентный стимул.
На основе огромного опыта изучения условных рефлексов у собак И. П. Павлов и его ученики создали учение о высшей нервной деятельности.
Оно базировалось на ряде постулатов, которые довольно хорошо соответствовали накопленным к тому времени экспериментальным данным. Концепция Павлова содержала следующие положения:
высшая нервная деятельность (т.е. образование условных рефлексов) есть результат взаимодействия двух основных нервных процессов — возбуждения и торможения;
при действии УС в коре головного мозга формируется очаг возбуждения;
из этого очага возбуждение иррадиирует (распространяется) по коре; внешнее проявление иррадиации возбуждения — процесс генерализации, т.е. появление УР не только на данный стимул, но и на близкие к нему по параметрам раздражители (например, не только на звуковой тон определенной высоты, который использовался при обучении, но и на другие звуки близких диапазонов) (см. рис. 3.2Б);
свойством генерализации обладает и тормозный процесс;
очаги возбуждения и торможения обладают свойством отрицательной индукции, благодаря которому на периферии очага возбуждения в коре появляется очаг торможения (таким же свойством обладают очаги торможения);
процессы возбуждения и торможения взаимодействуют на основе не только их иррадиации, но и концентрации; если иррадиации нервных процессов соответствует явление генерализации, то концентрация процесса возбуждения проявляется в формировании дифференцировочных условных рефлексов (см. 3.3);
формирование УР заключается в образовании связи между двумя очагами возбуждения, вызванными условным и безусловным раздражителями.
Фундаментальное значение открытия И. П. Павловым условных рефлексов заключается в том, что такой вид психической активности, как ассоциативное обучение, стал предметом экспериментальных физиологических исследований (ранее психологи изучали его только на основе интроспективных заключений).
Павловская концепция физиологии высшей нервной деятельности логично описывала полученные в тот период и теми методами экспериментальные данные. Она сыграла большую роль в науке, объясняя механизм формирования целого ряда сложных поведенческих реакций. Однако постепенно, с расширением методической базы, а также с переходом к экспериментам на других животных, стала очевидной ее ограниченная применимость для объяснения многих фактов, прежде всего потому, что упомянутые закономерности нервных процессов далеко не всегда подтверждались прямыми нейрофизиологическими исследованиями функций головного мозга. Так, например, представления об иррадиации и концентрации нервных процессов не подтвердились при использовании в качестве моделей других УР. В настоящее время отдельные ученые продолжают использовать положения павловской концепции при трактовке результатов изучения высшей нервной деятельности, получаемых традиционными методами павловской школы. В этом нет ничего парадоксального, так как в любой теории, прошедшей проверку временем, основная идея сохраняется.
Сущность павловского учения составляет идея об условных рефлексах как элементарной единице приспособительной деятельности. Методологический подход к изучению высшей нервной деятельности базируется на четырех принципах: детерминизм, анализ и синтез, приуроченность функции к структуре.
Ассоциативное обучение, включающее классические и инструментальные условные рефлексы, интенсивно исследовалось на протяжении всего XX века. Рассмотрим более подробно основные типы условных рефлексов.
2.2.2. Классические условные рефлексы
Условнорефлекторное слюноотделение, которому уделялось основное внимание в лаборатории Павлова, относится к классическим УР.
При выработке классического УР последовательность событий в опыте никак не зависит от поведения животного. Она устанавливается либо экспериментатором, либо специальной программой, в соответствии с которой включаются те или иные стимулы, в ответ на них можно наблюдать образование условных реакций.
В настоящее время в связи с использованием разнообразных экспериментальных животных (не только традиционных собак), а также благодаря разнообразным методам регистрации изучаемых реакций на смену павловской методике классических слюнных УР пришли другие, более удобные лабораторные модели, которые продолжают быть предметом многочисленных исследований (см. также Мак-Фарленд, 1987; Реагсе, 1998).
Разнообразные классические условнорефлекторные реакции можно наблюдать в экспериментах на животных, если проводить полиграфическую регистрацию ряда физиологических процессов организма (ЭКГ, ЭЭГ, плетизмог-рамму и др.). Как и в случае слюнного рефлекса, сочетание положительного (например, пищевого) безусловного раздражения с нейтральным приведет к тому, что последний начнет вызывать изменения в вегетативных функциях организма, которые до этого провоцировались только непосредственно самим безусловным воздействием. Полиграфическая регистрация обычно используется при изучении отрицательных (аверсивных) классических УР. Много работ такого рода проводится также для оценки реактивности нервной системы к действию стрессорных агентов и ее устойчивости к стрессу.
В настоящее время классические УР наиболее часто исследуют на моделях, использующих вкусовое отвращение и реакцию третьего века (мигательной перепонки).
Было, например, обнаружено, что классические УР образуются с разной скоростью в зависимости от того, какие физиологические системы вовлечены в их формирование. Так, УР избегания пищевого яда у крыс (как правило, используют хлорид лития) формируется легко, если введение его в организм сочетается с предложением животному пищи определенного вкуса. Однако УР избегания яда формируется с трудом или не образуется совсем, если его введение сочетается, например, со звуковым раздражением. На схеме (рис. 3.3) показано, как авторы (Garcia et al., 1970) представляют себе гипотетический механизм формирования такого УР вкусового отвращения. Вкусовой стимул, сочетающийся с пищевым отравлением, ведет к образованию УР вкусового отвращения. При сочетании звука с ударом тока образуется УР на боль. В то же время иное сочетание, например вкусового и болевого стимулов, не ведет к образованию УР.
Классический УР — сокращение мигательной перепонки при действии тактильного или звукового УС — обычно изучают на кроликах. При действии на роговицу глаза безусловного раздражителя — воздушной струи (или слабого удара тока) — мигательная перепонка сокращается. Это сокращение можно регистрировать специальным прибором и оценивать его интенсивность. Если безусловное раздражение сочетать с каким-либо нейтральным стимулом, например звуком, то после нескольких сочетаний мигательная перепонка будет сокращаться уже при изолированном действии этого звука, который становится условным сигналом (см. рис. 2).
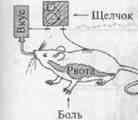
Рис. 3. Схема, показывающая избирательность ассоциаций между УС и подкреплением при формировании классических УР.
| Стимулы |
Последствия |
| рвота |
боль |
| Сладкий вкус |
формирование УР вкусового отвращения |
УР не формируется |
| Щелчок |
УР не формируется |
Формирование оборонительного УР |
2.2.3. Инструментальные условные рефлексы (или обучение методом проб и ошибок)
Начало исследований инструментальных УР связано с именем Э. Торндайка (см. 4.1), хотя их анализ проводился и в лаборатории И. П. Павлова.
В лаборатории И. П. Павлова «классические» УР носили название условных рефлексов 1-го рода, а инструментальные — условных рефлексов 2-го рода. В монографии Ю. Конорски (1969) на большом экспериментальном материале дается анализ сходства и различия между ними.
В опытах с «проблемными ящиками» Торндайк наблюдал, как посаженная в ящик кошка ищет выход, пытаясь открыть дверцу разными способами (рис. 3.4А) (для этого нужно было нажать на задвижку или потянуть за пружину). Кошка сначала совершает много разных действий (проб), которые в своем большинстве бывают неверными (ошибки), пока случайно не откроет яшик. При повторении опытов она выходит из ящика все быстрее и быстрее. Идея эксперимента была подсказана Торндайку К. Ллойдом-Морганом, наблюдавшим, как его собака манипулирует с задвижкой калитки, чтобы «улизнуть» из дома (рис. 3.5). Такое обучение Торндайк и назвал «методом проб и ошибок». В дальнейшем обучение животного подобным действиям получило название инструментальных (или оперантных) УР.
Торндайк первым предложил количественную оценку динамики обучения животного инструментальному навыку. Для этого он ввел так называемые «кривые научения», примеры которых приведены на рис. 4Б.
Между классическими и инструментальными УР существуют определенные различия.
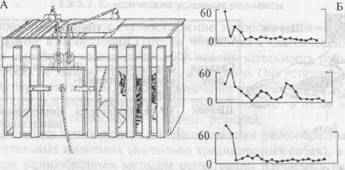
Рис. 4. Инструментальные УР.
А — один из вариантов «проблемного ящика», предложенный Э. Торндайком для исследования способности животного к решению.задачи методом «проб и ошибок»; Б — примеры кривых научения при использовании этого метода. По оси ординат — время, затраченное на решение задачи, по оси абсцисс — последовательные предъявления теста.
При классических УР временная связь между условным сигналом и безусловной реакцией возникает непроизвольно при действии безусловного раздражителя (подкрепления).
При инструментальных УР подкрепление, например пища, дается только после того, как животное совершает определенное действие, которое не имеет прямой связи с безусловным раздражителем.
В классическом УР выделение слюны происходит в ответ на контакт пищи с рецепторами полости рта, и эта реакция может стать услов-норефлекторной. В отличие от этого при выработке инструментального УР пищевое подкрепление изначально никак не связано ни с протягиванием лапы, ни с побежкой в лабиринте. Однако мы можем давать животному пищу сразу после того, как оно сделало такое движение, и вскоре животное будет его совершать, чтобы получить подкрепление.
К категории инструментальных УР относится формирование навыка нажатия на рычаг (или клевания кнопки, как на рис. 3.6) для получения порции пищи. После работ Б.Скиннера этот УР стал основным в исследованиях бихевиористов (анализ связей «стимул-реакция»). К инструментальным УР относится также обучение подопытного животного находить путь к пище или избегать неприятных стимулов в лабиринте. Инструментальным УР является и реакция избегания — навык переходить в тот отсек экспериментальной камеры, где отсутствует болевое подкрепление (рис. 7).

Рис. 5. Рисунок, сделанный по фотографии «исторического» эпизода. Собака К. Л. Моргана научилась открывать задвижку калитки, получая тем самым возможность «обрести свободу». Эта ситуация считается прототипом торндайковского «проблемного ящика».

Рис. 6. Камера Б. Скиннера для обучения голубей.
В верхней части левой стенки камеры размещены 3 диска-«ключа». Они различаются не только по положению, но и по цвету лампочки, которая загорается за каждым из них. В кормушку (ниже) подают подкрепление. Клевание ключа (или, например, каждое 5-е клевание) автоматически подает зерно в кормушку.

Рис. 7. Челночные камеры для исследования у лабораторных крыс и мышей реакции активного избегания.
А — современная установка для изучения условной реакции активного избегания, которая обычно состоит из 4 челночных камер с автоматизированным управлением и выводом результатов на экран монитора; Б — отдельная челночная камера.
Включению тока предшествует включение УС — звука или света. Чтобы избежать болевого воздействия, животное перебегает в другую половину камеры. По прошествии небольшого периода времени (как правило, его длину варьируют, чтобы не вырабатывать у животного УР на время) ток включают в той половине камеры, куда оно перед этим перебежало. УР считается выполненным, если животное перебегает в безопасную половину камеры во время действия УС и до включения тока.
Обширные исследования, выполненные сторонниками идей бихевиоризма, привели к появлению целого ряда новых терминов и понятий и созданию специфического языка для описания закономерностей процесса обучения, обнаруженных только благодаря примененным ими подходам. Многие термины сначала использовались как чисто технические — для объективного описания данных (знакомство с ними может помочь при чтении научных статей бихевиористов). К таким терминам, в частности, относятся:
оперантное поведение — спонтанные действия, не вызванные каким-либо очевидным стимулом;
реактивное поведение — всякое поведение, которое совершается в ответ на определенный стимул;
режим подкрепления (reinforcement schedule): соотношение числа реакций (например, нажатий на рычаг) и вознаграждений (например, кусочков пищи; подробнее см. ниже);
постоянный (или переменный) интервал (fixed or variable interval) —
вознаграждение дается через определенные интервалы времени, в период которых животное может осуществлять двигательные реакции (эти интервалы могут быть постоянными или варьировать по длительности), но по истечении этого интервата подкрепляется только первая реакция;
фиксированное (или переменное) соотношение (fixed ratio or variable ratio) — подкрепление дается животному только после того, как оно произведет какое-то количество реакций (нажатий или клеваний. Можно давать подкрепление только после определенного числа реакций (фиксированное соотношение) или это число может каким-то образом варьировать (переменное соотношение);
смежность (contiguity) — этим термином обозначают постулируемое возникновение связей между стимулами и возникающими на их основе реакциями, а также совпадение УС с реакцией животного или УС с подкреплением, которое необходимо для возникновения УР;
ключ (сие) — обычно это либо рычаг, на который нажимает крыса, либо освещенный кружок, по которому наносит удар клювом голубь (этот термин, обычный для работ бихевиористов, не следует путать с термином «ключевой стимул», принятым в этологии для обозначения видоспецифических стимулов, вызывающих инстинктивные реакции (см. 2.11).
На рис. 6 схематически изображена камера для выработки инструментальных УР у голубей. Скиннер считал, что любое поведение, относящееся к категории «оперантное», можно модифицировать, если при его выполнении давать животному подкрепление. Именно такой подход он предложил как эффективный способ анализа поведения. Если для выработки классических УР необходимы сочетания условных сигналов и подкрепления, то при методике свободного оперантно-го поведения, предложенной Скиннером, экспериментатор сопровождает подкреплением выполнение животным только определенного, намеченного им действия.
Например, крысу можно обучить нажимать на рычаг, если сначала сопровождать подкреплением любые ее действия в той части камеры, где он находится. Постепенно крыса обучается держаться вблизи рычага, и тогда подкрепление дают только, если она касается рычага мордой или лапой (для этого иногда на рычаг даже кладут пишу). Через некоторое время подкрепление дается только после выполнения четких движений — нажатий (одного или нескольких) лапой на рычаг.
Такое постепенное видоизменение поведения животного в результате вмешательства экспериментатора называется методом последовательного приближения или формированием (shaping) поведения.
Деление условных рефлексов на классические и инструментальные, удобное методически, не означает, что они имеют совершенно разную природу. В их основе лежат сходные нейрофизиологические механизмы, а любое «чисто» инструментальное действие животного всегда сопровождается реакцией, которая относится к классическим УР. И наоборот, в любом «чисто» классическом УР можно обнаружить двигательный компонент, который по своим свойствам относится к инструментальным (Борукаев, 1982).
О связи классических и инструментальных УР свидетельствует также «самоформирование» условной реакции {autoshaping). Например, если голодного голубя поместить в камеру Скиннера и в течение 5 с освещать ключ для клевания с интервалом в 1 мин, а после прекращения освещения давать порцию пиши, то голубь сначала не обращает внимания на ключ. Однако после нескольких таких сеансов формируется инструментальная реакция, и он быстро начинает клевать ключ независимо от освещения. Постепенно голубь научается клевать ключ только при его освещении, т.е. реакция становится более точной — она самоформируется. В данном случае она очень близка к классическому УР, поскольку голубь клюет ключ как бы вместо зерна, даже в отсутствие подкрепления.
С помощью метода «последовательного приближения» у животных удается сформировать не только клевание ключа при его освещении, но и самые разнообразные, сложные и иногда неожиданные навыки. В опытах Скиннера голуби «играли» в пинг-понг, а крысы могли подтянуть к себе с помощью бечевки бильярдный шар, взять его в передние лапы и засунуть в трубку, расположенную на 5 см выше пола клетки (описание экспериментов П. Эпштейна, посвященных формированию сложных навыков у голубей, будет дано в гл. 4.8.2).
Метод последовательного приближения составляет основу дрессировки цирковых и служебных животных. Примером продуктивного использования принципов бихевиоризма, и в частности роли метода последовательного приближения в формировании поведения, является работа знаменитой американской дрессировщицы дельфинов и психолога К. Прайор(1995).
Тезис Скиннера о том, что любые движения, на которые способно животное данного вида, равновероятно можно использовать для инструментального научения, был достаточно обоснованно подвергнут сомнению в работе его коллег - К. и М. Бреландов (Breland, Breland, 1961). Некоторые виды животных при обучении манипуляциям с предметами производят «заданные» экспериментатором действия только после выполнения некоторых врожденных (инстинктивных) действий либо совсем не могут им научиться. Так, например, обучая енота нажимать на рычаг, Бреланды заметили, что нажатию предшествует видоспецифи-ческое движение лап — «полоскание», характерное для этого вида при добыче пищи из ручья (за что енота и называют «полоскун»). На основании таких наблюдений возникло представление о предрасположенности к определенным видам обучения (подробнее см.: Зорина и др., 1999).
Образование инструментальных УР по механизму «последовательного приближения» играет важную роль в организации поведения животных не только в эксперименте, но и в естественных условиях.
Обучение по методу последовательного приближения может происходить самым неожиданным образом. Об этом, в частности, свидетельствует получившая широкую известность в начале XX века история «умного Ганса». Это был конь, который демонстрировал способность «считать», «складывать», «извлекать корни», отвечать на вопросы и т.п. (рис. 3.8). Столь «разумное» поведение объяснялось тем, что конь научился замечать малоуловимые движения дрессировщика, которые тот непроизвольно совершал, видя, что конь вот-вот даст правильный ответ. Ориентируясь на эти условные сигналы, конь в нужный момент прекращал стучать копытом.

Рис. 8. Инструментальные условные рефлексы у дрессированной лошади «умного Ганса». На фотографии видно, как ударами копыта по специальной доске Ганс сигнализировал «правильный» ответ (по Н. Н. Ладыгиной-Котс, 1914).
В начале XX века (1900—1904) барон В. фон Остен, убежденный в огромных умственных способностях лошадей, обучал нескольких из них различению цветов, азбуке и «счету». Узнавание каждой буквы или цифры лошадь обозначала соответствующим числом ударов копыта. Друг фон Остена художник Редлих обучил таким же образом свою собаку. Наиболее способным учеником оказался орловский рысак Ганс, который производил достаточно сложные арифметические подсчеты, отвечал на разнообразные вопросы, а иногда высказывался по собственной инициативе. Так, супруги Н. Н. и А. Ф. Котс, специально приехавшие для знакомства с ним в 1913 году, рассказывали, что после нескольких относительно коротких ответов на вопросы Ганс заявил: «В поле я встретил милую госпожу Краль, которая меня кормила».
Поведение его было столь впечатляющим, что вводило в заблуждение не только публику, но даже членов специальных комиссий, включая Н. Н. Лады-гину-Котс. Предполагали, что хозяин подает коню некие скрытые сигналы (как дрессировщик — цирковым животным), однако его обследовали 13 экспертов (комиссия психолога К. Штумпфа) и не обнаружили никакого обмана. Они засвидетельствовали, что Ганс действительно «считает» и никаких скрытых сигналов ему не подают. И лишь много позднее наблюдатели постепенно заметили, что Ганс отвечает только на те вопросы, ответ на которые знает сам экспериментатор. Специальный анализ, проведенный психологом О. Пфунгстом, показал, что животное реагирует на мельчайшие непроизвольные (идеомоторные) движения экспериментатора, например на отклонения корпуса на 2 мм, микродвижения бровей, мимику и т.п. Эта невольная подача сигналов происходила, по-видимому, из-за эмоционального напряжения человека, по мере того как число ударов копытом приближалось к искомому, Даже картонный щит, которым пробовал отгородиться от Ганса экспериментатор, не помогал: животное все равно улавливало какие-то только ему понятные знаки для определения правильного ответа.
Для проверки своего предположения Пфунгст специально научил Ганса реагировать на микродвижения, которые он совершал уже сознательно, и продемонстрировал комиссии механизм и природу «математических способностей» этой лошади.
История «умного Ганса» оставила заметный след в развитии науки о поведении животных:
она показала, сколь сложное поведение могло быть результатом обучения методом проб и ошибок;
продемонстрировала справедливость «канона Ллойда Моргана» (см. 2.3) и актуальность его применения, поскольку в основе поведения этой «мыслящей лошади» лежали чисто условнореф-лекторные, а не связанные с мышлением механизмы;
впервые привлекла внимание к проблеме чистоты эксперимента с точки зрения возможности неосознанного влияния экспериментатора на его результаты;
заставила по-новому взглянуть на возможности восприятия животных: способность уловить едва заметные движения человека свидетельствовала об их большой наблюдательности и способности концентрировать внимание.
3. Дифференцировочные условные рефлексы.
Перестав подкреплять выполнение условной реакции, можно не только «угасить» УР, но и «усовершенствовать» его, т.е. добиться, что условная реакция будет осуществляться точно на данный и только на данный стимул, а близкие по своим физическим свойствам раздражители ее не вызовут. Как известно, в начале формирования УР животное реагирует не только на строго конкретный условный стимул, например звук определенной частоты, но и на сходные с ним звуки, хотя и не полностью ему идентичные, например звуки близких частот (см. рис. 3.2Б). Эта стадия выработки УР называется генерализацией. Далее исследователь продолжает подкреплять (пищей или избавлением от опасности) условную реакцию только на определенный сигнал и не подкрепляет ответов на сходные, и в результате они постепенно угасают. Таким способом животное обучают различать — дифференцировать — весьма похожие сигналы.
В основе формирования дифференцировочных УР лежит днфферен-цировочное торможение — один из видов внутреннего торможения.
Процедура такого «совершенствования» сигнального значения условных раздражителей была названа И. П. Павловым дифференци-ровкой, или выработкой дифференцированных условных рефлексов (см. также рис. 3.2).
Выработка дифференцировочных УР (разными методами) успешно используется для оценки сенсорных способностей (т.е. возможностей органов чувств) животных разных таксономических групп (см., например: Дьюсбери, 1981; Реагсе, 1998). Так, выработав у животного УР дифференцирования (различения) двух цветовых стимулов, можно, выравнивая их другие параметры (например, яркость), изучать механизмы восприятия цвета. При этом если животное, в силу особенностей органов чувств, не воспринимает различий между двумя зрительными стимулами (цветами спектра), то выработать у него устойчивую дифференцировку такой пары стимулов невозможно.
Методики выработки дифференцировочных УР весьма многочисленны и разнообразны по процедуре эксперимента. Перечислим основные параметры, по которым различаются такие методики.
Порядок предъявления стимулов может быть последовательным или одновременным. При последовательном предъявлении голубь должен научиться клевать кнопку (диск) в ответ на стимул А и воздерживаться от реакции при включении стимула Б (методика go— no go). Выработка дифференцировки, таким образом, состоит в торможении реакции на второй стимул. При одновременном предъявлении конкретной пары стимулов животное учится различать (дифференцировать) их абсолютные признаки. Например, при дифференцировке стимулов по их конфигурации животному одновременно показывают две фигуры — круг и квадрат, и подкрепляют выбор одной из них, например, круга. Это наиболее распространенный вид дифференцировочных УР. Выработка и упрочение такой реакции требует, как правило, многих десятков сочетаний.
Животному предлагают сделать выбор стимулов: альтернативный или множественный.
Предъявление стимулов может осуществляться в соответствии с двумя режимами: повторение одной пары стимулов до достижения критерия и чередование нескольких пар стимулов при систематическом варьировании второстепенных параметров.
Рассмотрим некоторые методы более подробно.
При систематическом варьировании второстепенных параметров стимулов можно оценивать способность животных различать не только данную конкретную пару раздражителей, но и их «обобщенные» (отвлеченные) признаки, совпадающие у многих пар.
Например, животных можно обучить различать не конкретные круг и квадрат, а любые круги и квадраты независимо от их размера, цвета, ориентации и т.п. С этой целью в процессе обучения каждый следующий раз им предлагают новую пару стимулов (новые круг и квадрат). Новая пара отличается от остальных по всем второстепенным признакам стимулов — цвету, форме, размерам, ориентации и т.п., но сходна по их основному параметру — геометрической форме, различения которой и предполагается добиться. В результате такой тренировки у животного постепенно происходит обобщение основного признака и отвлечение от второстепенных. В приведенном примере обобщается признак «круг» (подробнее см. гл. 5).
Данный режим оказался весьма эффективным для выработки диф-ференцировочного УР на обобщенный признак «мерность», когда животное училось выбирать любую объемную (трехмерную) фигуру и не реагировать на плоские (двумерные) фигуры (см. 4.6.3). В таких опытах с собаками, обезьянами и воронами у одной группы животных каждого вида вырабатывали УР выбора любой объемной (трехмерной) фигуры, а у второй группы — любой плоской (двумерной). Каждый раз животному предъявляли новую пару стимулов из набора, изображенного на рис. 4.15 (всего 30 пар), причем плоская фигура всегда представляла собой фронтальную проекцию объемной.
Может возникнуть предположение, что задача по выработке таких дифференцировок для животных очень сложна, однако с ней справляются не только приматы, но и хищные млекопитающие (Дашев-ский, Детлаф, 1974), а также птицы — врановые (Крушинский и др., 1981) и даже голуби. При этом животным всех указанных видов требовалось примерно столько же сочетаний, как и при выработке диффе-ренцировки с одной парой стимулов. После такой серии обучения они способны выбирать любую новую объемную (или плоскую) фигуру без дополнительной тренировки.
При систематическом варьировании второстепенных признаков стимулов можно исследовать не только способность животных к обучению, но и более сложную форму их высшей нервной деятельности — способность к обобщению, которая составляет одно из важнейших свойств довербального мышления животных (гл. 5).
Как уже упоминалось, животные с разным уровнем структурно-функциональной организации мозга практически не различаются по способности к простым формам условнорефлекторного обучения. Образование отдельных дифференцировочных УР в этом плане не составляет исключения. Однако благодаря использованию дифференцировочных УР в качестве элементарных единиц обучения и созданию их разнообразных комбинаций было разработано несколько экспериментальных процедур, которые называют «сложными формами обучения», или серийным обучением (serial learning). Появление этих методов было связано со стремлением специалистов все же выявить какие-то различия в способностях у животных разных таксономических групп с разным уровнем организации мозга (см. 8.1). Наиболее важные из этих методов:
последовательные переделки сигнального значения дифферен-
цировочных стимулов;
формирование установки на обучение;
формирование «систем» дифференцировочных УР.
3.1. Последовательные переделки сигнального значения дифференцировочных стимулов.
Переделка сигнального значения условных раздражителей (ее часто называют переделкой дифференцировки) — это разновидность дифференцировочного обучения.
После достижения определенного, выбранного экспериментатором критерия выработки различения (например, 80% правильных решений) условия получения подкрепления меняются на противоположные: теперь реакция на стимул, ранее подкреплявшийся, перестает сопровождаться пищей, а реакцию на стимул, на который животное только что научилось не реагировать, начинают подкреплять.
Американский исследователь М. Биттерман (1973) предложил проводить такие переделки многократно. Эта процедура получила название метода последовательных (или обратимых) переделок (discrimination reversal learning). Автор исходил из того, что после многократных смен сигнального значения стимулов у животных может сформироваться единая стратегия (или правило), и они не будут каждый раз обучаться совершенно заново.
Опыты проводились на разных видах позвоночных. При каждой следующей переделке число ошибок снижалось, и в конце концов наступал момент, когда при очередной смене сигнального значения стимулов животное уже со второй пробы начинало выбирать «правильно» без дополнительного обучения.
Снижение числа проб, необходимого для очередной переделки реакции, происходит тем быстрее, чем выше уровень эволюционного развития данного животного.
3.2. Формирование «установки на обучение».
Метод формирования «установки на обучение» (object learning-set formation) разработал американский исследователь Г. Харлоу (Harlow, 1949; 1958). Согласно классическому варианту этой методики, животное (в опытах Харлоу макака-резуса) обучают простой дифференци-ровке — выбору одного из двух стимулов: игрушек или мелких предметов обихода. По достижении определенного критерия выработки дифференцировки начинают следующую серию: животному предлагают два новых стимула, ничем не похожих на первые. По завершении второй стадии обучения формируют третью дифференцировку и т.д. После выработки 100—150 таких реакций (их число зависит от уровня организации данного вида и может доходить до 1500 у крыс и голубей, занимая многие месяцы) животное уже при втором предъявлении новой пары стимулов действует не наугад, не методом проб и ошибок, а в соответствии с усвоенным им ранее правилом, которое принято называть установкой. Оно запоминает результат первой пробы и далее реагирует в соответствии с этой «установкой».
Правило состоит в том, чтобы «выбирать тот же предмет, что и в первой пробе, если его выбор сопровождался подкреплением, или другой, если подкрепление получено не было (в дальнейшем эта стратегия получила название "win-stay, lose-shift")».
Принято считать, что при обучении с использованием метода последовательных переделок и при формировании установки происходят сходные процессы, так что первый можно считать частным случаем формирования установки.
Впоследствии было создано несколько модификаций исходной методики. Например, Д. Уоррен (Warren, 1977) в своих опытах не вырабатывал каждую дифференцировку до достижения критерия обученности, а предъявлял каждую пару определенное число раз и затем предлагал следующую. Именно этот вариант был использован рядом авторов для сравнения обучаемости разных видов приматов (Passingham, 1982; Rumbaugh et al., 1987, 2000).
Методом формирования установки на обучение впервые была получена широкая сравнительная характеристика обучаемости животных разных систематических групп, которая в определенной степени коррелировала с показателями организации мозга. Вместе с тем, как и данные М. Биттермана, эти результаты свидетельствовали о существовании у животных каких-то процессов, выходящих за рамки простого образования дифференцировочных УР. Считают, что в ходе такой процедуры животное «учится учиться». Оно освобождается от связи «стимул-реакция» и «переходит от ассоциативного обучения к инсайт-подобному обучению с одной пробы» (Harlow, 1958). При этом, по мнению Р. Бирна (Byrne, 1998), у животного должна сформироваться характеристика подкрепляемого стимула в какой-то отвлеченной форме, т.е. должно сформироваться абстрактное правило. Человек выразил бы это правило, как «выбирай подкрепляемое» вместо того, чтобы запоминать каждый раз целый список стимулов (например, красный мяч «+», блюдце «—»; штопор «+», свисток «—» и т.д.).
Аналогичной точки зрения придерживается Л. А. Фирсов. Он считает, что этот вид обучения по своей сути и по лежащим в его основе механизмам близок к процессу обобщения (подробнее см. гл. 5), и с какого-то момента «первичное» обучение — выработка конкретной дифференцировки — сопровождается «вторичным» обучением — некими когнитивными процессами, при которых выявляется общее правило решения многих однотипных задач.
То же мнение разделяет английский специалист по теории обучения Н. Макинтош (Mackintosh, 2000): в основе формирования установки на обучение лежит переход от простых ассоциативных механизмов (образования УР и дифференцировок) к когнитивным, т.е. к процессам, основанным на формировании представлений об общем правиле решения серии однотипных задач. По его образному выражению, животные «переходят от заучивания наизусть к заучиванию по смыслу».
Данные, полученные с помощью метода последовательных переделок и метода формирования «установки», впервые создали основу для широкой сравнительной характеристики обучаемости животных и показали, что в процессе такого обучения наряду с ассоциативными участвуют механизмы другого уровня — когнитивные (см. 8.1 и 3.4).
3.3. формирование «систем» дифференцировочных условных рефлексов.
В работах Л. Г. Воронина (1984) также было показано, что при комбинации отдельных УР возникают новые свойства, не сводимые к свойствам суммы исходных рефлексов. Происходит образование так называемых систем дифференцировочных УР, или просто «систем». Такие системы Л. Г. Воронин считал целостными функциональными единицами, из которых складывается психическая деятельность.
У животного вырабатывают несколько дифференцировочных УР, объединенных в систему таким образом, что выполнение одного УР зависит от усвоения другого.
Одна из таких процедур составляет необходимый этап в изучении способности к транзитивному заключению — форме дедуктивного мышления, свойственной и животным, и человеку.
Программа эксперимента по выработке систем дифференцировок состоит в следующем. Животное учится дифференцировать несколько пар раздражителей (как правило, цветовых стимулов). При выработке дифференцировки каждой следующей пары стимулов производится смена сигнального значения части из них в соответствии с определенным правилом: стимул, бывший положительным в паре А— В+ (где «В+» — подкрепляемый стимул, а «А-» — неподкрепляемый), становится отрицательным в паре В— С+. В процессе обучения животное должно усвоить информацию о том, что значения стимулов В, С, D могут быть как положительными (В+, С+, D+), так и отрицательными (В—, С—, D—), в зависимости от пары, в которой они в данный момент предъявляются. Стимулы А— и F+ , т.е. первый и последний в последовательности, остаются во всех комбинациях только положительным (F) или только отрицательным (А). Высокая доля правильных выборов при чередовании всех пар стимулов, когда животное выбирает, например, стимул С в паре В— С+, но не реагирует на него в паре С— D+, свидетельствует, что оно усвоило принцип данной системы. Такую систему дифференцировок успешно усваивают дети, начиная с 4 лет (Bryant, Trabasso, 1971), взрослые шимпанзе (Gillan, 1981; Boysen, Bermtson, 1995), а также голуби и вороны (Зорина и др., 1989). Межвидовые различия в динамике и успешности формирования такой системы дифференцировок у этих видов птиц оказались незначительными и недостоверными. В то же время при обратимых последовательных переделках и формировании установки на обучение голуби существенно отставали как от врановых, так и от приматов.
Метод выработки дифференцировочных УР в разных его комбинациях является одним из ведущих в физиологии ВНД и используется как инструмент для исследования целого ряда проблем.
К этим проблемам, помимо оценки возможностей органов чувств животных разных видов, относится изучение следующих более сложных когнитивных способностей:
к обобщению;
к транзитивному заключению;
к формированию установки на обучение.
Такие экспериментальные исследования будут подробнее рассмотрены ниже.
4. Когнитивные (познавательные) процессы.
4.1. Общая характеристика.
Термин «когнитивные», или «познавательные», процессы употребляют для обозначения тех видов поведения животных и человека, в основе которых лежит не условнорефлекторный ответ на воздействие внешних стимулов, а формирование внутренних (мысленных) представлений о событиях и связях между ними.
И. С. Бериташвили называл их психонервными образами, или психонервными представлениями, Е. Н. Соколов — нервной моделью стимула, Л. А. Фирсов (1972; 1993) и Т. А. Натишвили (1987) — образной памятью. Д. Мак-Фарленд (1988) подчеркивает, что когнитивная деятельность животных относится к мыслительным процессам, которые зачастую недоступны прямому наблюдению, однако их существование возможно выявить в эксперименте.
Наличие представлений обнаруживается в тех случаях, когда субъект (человек или животное) совершает действие без влияния какого бы то ни было физически реального стимула. Такое возможно, например, когда он извлекает информацию из памяти или мысленно восполняет отсутствующие элементы действующего стимула. В то же время формирование мысленных представлений может никак не проявляться в исполнительной деятельности организма и обнаружится лишь позднее, в какой-то определенный момент.
Внутренние представления могут отражать самые разные типы сенсорной информации, не только абсолютные, но и относительные признаки стимулов, а также соотношения между разными стимулами и между событиями прошлого опыта. По образному выражению, животное создает некую внутреннюю картину мира, включающую комплекс представлений «что», «где», «когда». Они лежат в основе обработки информации о временных, числовых и пространственных характеристиках среды и тесно связаны с процессами памяти. Различают также образные и абстрактные (отвлеченные) представления (Premack, 1983). Последние рассматривают как основу формирования довербальных понятий (см. гл. 5).
Метод отсроченных реакций. Мысль о существовании у животных некоего «процесса представления», т.е. такой активности мозга, которая соответствует полученной ранее стимуляции, но которая может поддерживаться в ее отсутствие, была впервые высказана У.Хантером в 1913 г. Для оценки способности животного реагировать на воспоминание о стимуле в отсутствие этого реального стимула У. Хантер предложил метод отсроченных реакций (2.4.4).
Животное (в опытах Хантера — енота) помещали в клетку с тремя одинаковыми и симметрично расположенными дверцами для выхода. Над одной из них на короткое время зажигали лампочку, а потом еноту давали возможность подойти к любой из дверец. Если он выбирал дверцу, над которой зажигалась лампочка, то получал подкрепление. При соответствующей тренировке животные выбирали нужную дверцу даже после 25-секундной отсрочки — интервала между выключением лампочки и возможностью сделать выбор.
В опытах других исследователей задача ставится иначе. На глазах у голодного животного в один из двух (или трех) ящиков помещают корм. По истечении периода отсрочки животное выпускают из клетки или (как на рис. 3.9) убирают отделяющую его преграду. Его задача выбрать ящик с кормом.
Выполнение теста на отсроченные реакции на неслучайном уровне считается доказательством наличия у животного мысленного представления о спрятанном предмете (его образа), т.е. существования какой-то активности мозга, которая в этом случае подменяет информацию от органов чувств. С помощью этого метода были исследованы представители различных видов животных и было подтверждено, что их поведение может направляться не только действующими в данный момент стимулами, но также и хранящимися в памяти следами, образами или представлениями об отсутствующих стимулах (рис. 3.9).
Опыты на разных животных были дополнены исследованиями на детях. Эту методику стали довольно широко применять и в нашей стране, начиная с 30-х годов, но многие ученики И. П. Павлова (например, Э. Г. Вацуро) отождествляли ее с методом следового УР, когда выполнение условной реакции происходит как бы на основе «следов действия УС». Метод отсроченных реакций стали использовать как тест на форму особой (образной или декларативной) памяти, которая не идентична памяти, формирующейся при УР (см. словарь терминов).
Между условнорефлекторной (ассоциативной или процедурной) и образной (декларативной) памятью существуют тонкие функциональные различия. От особенностей их взаимодействия зависят, например, различия в организации сложного поведения низших и высших обезьян (Фирсов, 1972; 1993).
В классическом тесте на отсроченные реакции виды животных проявляют себя по-разному. Собаки, например, после того как корм положен в один из ящиков, ориентируют тело по направлению к нему и сохраняют эту неподвижную позу в течение всего периода отсрочки, а по ее окончании сразу бросаются вперед и выбирают нужный ящик. Другие животные в подобных случаях не сохраняют определенной позы и могут даже разгуливать по клетке, что не мешает им, тем не менее, правильно обнаруживать приманку. У шимпанзе формируется не просто представление об ожидаемом подкреплении, но ожидание определенного его вида. Так, если вместо показанного в начале опыта банана после отсрочки обезьяны обнаруживали салат (менее ими любимый), то отказывались его брать и искали банан. Мысленные представления контролируют и гораздо более сложные формы поведения. Многочисленные свидетельства этого были получены и в специальных экспериментах, и в наблюдениях за повседневным поведением обезьян в неволе и естественной среде обитания (см., например: Л адыги на-Котс, 1923; 1935; Гудолл, 1992; подробнее об этом см. гл. 7).
Тест на оценку представлений о константности свойств предметов. Один из способов изучения роли представлений в поведении и психике животного — тест на «константность свойств предмета» («object permanence»), т.е. на способность субъекта понимать, что предмет, скрывшийся из поля зрения, продолжает существовать, может быть отыскан и не меняет при этом своих свойств. Тест широко применяется в исследованиях психического развития животных и человека (Пиаже, 1969; Бауэр, 1979).
Для оценки способности животных оперировать представлениями о константности свойств предмета существуют тесты:
на «неисчезаемость»;
на «вмещаемость»;
на «перемещаемость» и др.
Термины были введены Л. В. Крушинским (1986) и соответствующие тесты названы им «эмпирическими законами». Они соответствует 4-й и 6-й стадиям развития по Пиаже (1969) (подробнее об оценке этих способностей у животных см. 4.6.1).
В основе понимания животным или ребенком принципа «неисчезаемости» лежит эмпирическое знание того, что предметы существуют, даже если в данный момент они недоступны непосредственному восприятию органами чувств.
По мнению Л. В. Крушинского, понимание животным этого принципа составляет необходимое условие для проявления способности к экстраполяции направления движения стимула и решению ряда других элементарных логических задач (см. гл. 4).
«Мысленный план» лабиринта. Одним из первых гипотезу о роли представлений в обучении животных выдвинул Э. Толмен в 30-х годах XX века (1930, 1997). Исследуя поведение крыс в лабиринтах разной конструкции, он пришел к выводу, что общепринятая в то время схема «стимул-реакция» не может удовлетворительно описать поведение животного, усвоившего ориентацию в такой сложной среде, как лабиринт. Толмен высказал предположение, что в период между действием стимула и ответной реакцией в мозге совершается определенная цепь процессов («внутренние или промежуточные переменные»), которые определяют последующее поведение. Сами эти процессы, по мнению Толмена, можно исследовать строго объективно по их функциональному проявлению в поведении.
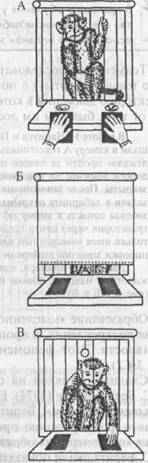
Рис. 9. Схема опытов по изучению отсроченных реакций.
А — демонстрация приманки и пустого контейнера; Б — приманку и пустой контейнер закрывают от животного непрозрачным экраном; В — экран убирают, и животное имеет возможность достать приманку, положение которой не изменилось. При следующем предъявлении этого теста контейнер с приманкой может располагаться слева.
В процессе обучения у животного формируется «когнитивная карта» всех признаков лабиринта, или его «мысленный план». Затем на основе этого «плана» животное выстраивает свое поведение.
Толмен и его последователи исследовали формирование «мысленного плана» («карты») с помощью лабиринтов разных конструкций, траекторию движения в которых животные могли менять в зависимости от того, были ли им доступны более короткие пути.
В работе Н. Чапиуса и П. Скардигли (цит. по: Реагсе, 1998) хомячка помещали в камеру А гексагонального лабиринта (рис. 10А), в котором ему «надлежало» пройти за пищей по пути, указанному штриховой линией. От этой дороги хомячок не отклонялся, поскольку все входы в другие отсеки были закрыты. После запоминания этого пути во время целого ряда предъявлений задачи в лабиринте открывали все дверцы, за исключением той, которая позволяла попасть к корму по прямой. Теперь кратчайшим путем к пище стала траектория через центр (сплошная линия). Этот путь можно уверенно найти, только имея «мысленный план» данного лабиринта. После определенной тренировки хомячки выбирали этот путь достоверно чаще, чем остальные.
Два других примера, взятых из работы самого Толмсна, показывают, как животное находит новый путь к пище, если знакомая дорога перекрыта (рис. 10Б и В).
Образование «мысленного плана» может происходить и в отсутствие подкрепления, в процессе ориентировочно-исследовательской активности. Этот феномен Толмен назвал латентным обучением (см. 4.2).
Сходных взглядов на организацию поведения придерживался И. С. Бериташвили (1974). Ему принадлежит термин — «поведение, направляемое образом». Бериташвили продемонстрировал способность собак к формированию представлений о структуре пространства, а также «психонервных образов» предметов. Ученики и последователи И. С. Бериташвили показали пути видоизменения и совершенствования образной памяти в процессе эволюции (Натишвили, 1987), а также в онтогенезе (Бадридзе, 1987), базируясь на данных по пространственной ориентации животных.
Гипотеза о когнитивных механизмах многих форм поведения закономерно возникала у исследователей, пытавшихся осмыслить данные самых разнообразных экспериментов. Американский ученый Д. Гриф-фин (Griffin, 1984) в книге «Мышление животных» («Animal thinking») в популярной форме рассматривает факты, хотя и не всегда действительно относящиеся к сфере мышления животных, но явно не получавшие объяснения в терминах бихевиоризма (например, орудийная деятельность и сложные виды коммуникации).
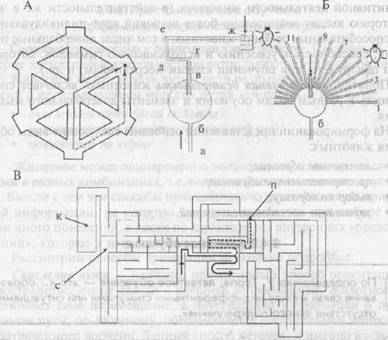
Рис. 10. Лабиринты, в которых исследовали формирование «мысленных карт» у животных.
А — гексагональный лабиринт; Б — усвоение общего плана строения лабиринта при изменении его конфигурации. Слева — голодная крыса обучается находить путь к пище, следуя по траектории а-б-в-г-д-е. После упрочения навыка животное помещают в точку а, но теперь из круглой камеры можно выйти по множеству рукавов за исключением одного (в). При наличии у животного «мысленного плана» оно чаще, чем случайно, выходит к пище по рукаву 5. В — крысу помещали в точку старта (с) и обучили находить корм (К). Постепенно у нее сформировался «мысленный план» лабиринта. После того как убрали перегородку (П), она стала бегать по более короткому пути, который обозначен пунктирной линией.
Когнитивная психология человека — направление, возникшее как реакция на господство идей бихевиоризма в области изучения психики человека, — оказала влияние на исследования когнитивных процессов у животных. Проведение аналогий между работой мозга и вычислительной машиной позволило описывать память животных как процесс переработки информации с привлечением соответствующего математического аппарата и понятий теории информации (Солсо, 1996).
Одно из наиболее популярных направлений в анализе когнитивных процессов у животных — это анализ обучения «пространственным» навыкам (главным образом, благодаря введению в лабораторную практику методов водного и радиального лабиринтов, см. 4.2). Существует даже тенденция считать эти феномены основным проявлением когнитивной деятельности животных (в действительности же в эту категорию входит значительно более широкий круг индивидуально-приспособительных реакций животных, в том числе довербальные понятия, способность к усвоению и использованию символов, которая была обнаружена при обучении языкам-посредникам, и др.).
Понятие, «когнитивная деятельность животных» включает способность к разным видам обучения и элементы довербального мышления.
На формировании представлений основаны следующие виды обучения животных:
латентное обучение;
пространственное обучение;
выбор по образцу;
заучивание последовательностей.
4.2. Латентное обучение.
По определению У. Торпа, патентное обучение — это «...образование связи между индифферентными стимулами или ситуациями в отсутствие явного подкрепления».
Элементы латентного обучения присутствуют практически в любом процессе обучения, но могут быть выявлены только в специальных опытах.
В естественных условиях латентное обучение возможно благодаря исследовательской активности животного в новой ситуации. Оно обнаружено не только у позвоночных. Эту или сходную способность для ориентации на местности используют, например, многие насекомые. Особенно хорошо латентное обучение изучено у перепончатокрылых. Так, пчела или оса, прежде чем улететь от гнезда, совершает «рекогносцировочный» полет над ним, что позволяет ей фиксировать в памяти «мысленный план» данного участка местности.
Наличие такого «латентного знания» выражается в том, что животное, которому предварительно дали ознакомиться с обстановкой опыта, обучается быстрее, чем контрольное, не имевшее такой возможности.
В настоящее время термин «латентное обучение» употребляется редко и лишь в определенном контексте. Однако на самом деле этот феномен достаточно широко распространен. Например, будет показано, что оно сопровождает выработку простых инструментальных УР (открывание кормушек разного цвета). Птицы (вороны и голуби) запоминают число единиц подкрепления, получаемого при действии стимулов разного цвета, а затем используют эту информацию в новой ситуации (Зорина и др., 1991). Феномен, который Л. А.Фирсов называет «вторичным обучением», по существу можно считать одним из вариантов латентного обучения.
4.3. Пространственное обучение. Современная теория «когнитивных карт».
Способность животных к ориентации в пространстве. Сведения об этой форме когнитивной деятельности вкратце заключаются в следующем.
Животное может искать путь к цели разными способами. По аналогии с прокладыванием морских путей эти способы называют:
счислением пути (dead reckoning);
использованием ориентиров (landmark use);
навигацией по карте.
Животное может одновременно пользоваться всеми тремя способами в разных комбинациях, т.е. они взаимно не исключают друг друга. Вместе с тем эти способы принципиально различаются по природе той информации, на которую животное опирается при выборе того или иного поведения, а также по характеру тех внутренних «представлений», которые у него при этом формируются.
Рассмотрим способы ориентации несколько подробнее:
Счисление пути — наиболее примитивный способ ориентации в пространстве; он не связан с внешней информацией. Животное отслеживает свое перемещение, а интегральная информация о пройденном пути, по-видимому, обеспечивается соотнесением этого пути и затраченного времени. Данный способ неточен, и именно из-за этого у высокоорганизованных животных его практически нельзя наблюдать в изолированном виде.
Использование ориентиров нередко сочетается со «счислением пути». Этот тип ориентации в большой степени близок формированию связей типа «стимул-реакция». Особенность «работы по ориентирам» состоит в том, что животное использует их строго поочередно, «по одному». Путь, который запоминает животное, представляет собой цепь ассоциативных связей.
При ориентации по местности («навигации по карте») животное использует встречающиеся ему предметы и знаки как точки отсчета для определения дальнейшего пути, включая их в интегральную картину представлений о местности.
В качестве примера расскажем о способностях к пространственной ориентации у столь разных видов млекопитающих, как шимпанзе и бурый медведь.
Наблюдения Э. Мензела (Menzel, 1979) за группами молодых шимпанзе, живших на огороженной, но достаточно обширной территории, показали, что при отыскании корма они гибко пользуются мысленным планом окружающей местности. Дж. Гудолл (1992) также приводит многочисленные подтверждения роли пространственной памяти в механизмах ориентировки шимпанзе в пространстве. По ее наблюдениям «карта», хранящаяся в памяти этих животных, весьма обширна. Она позволяет им легко находить пищевые ресурсы, разбросанные на площади до 24 кв. км в пределах Гомбе, и до сотен кв. км у популяций, обитающих в других частях Африки.
Пространственная память обезьян хранит не только расположение крупных источников пищи, например больших групп обильно плодоносящих деревьев, но и местонахождение отдельных таких деревьев и даже одиночных термитников. В течение по крайней мере нескольких недель они помнят о том, где происходили те или иные важные события, например конфликты между сообществами.
Знание своей территории обитания — важнейший фактор приспособленности хищных млекопитающих. Многолетние наблюдения В. С. Пажетнова (1991) за бурыми медведями в Тверской области позволили объективно охарактеризовать, какую роль играет мысленный план местности в организации их поведения. По следам животного натуралист может воспроизвести детали его охоты на крупную добычу, перемещения медведя весной после выхода из берлоги и в других ситуациях. Оказалось, что медведи часто используют такие приемы, как «срезание пути» при одиночной охоте, обход жертвы за многие сотни метров и др. Это возможно лишь в том случае, если у взрослого бурого медведя есть четкая мысленная карта района своего обитания.
Начало лабораторным исследованиям пространственной ориентации животных было положено в электрофизиологическом исследовании — при анализе реакций клеток гиппокампа (старой коры).
В 1976 г. появилась статья американского ученого Дж. О'Кифа, обнаружившего «клетки места» {place units) в гиппокампе свободноподвижной крысы. Эти нейроны разряжались избирательно только при попадании животного в определенное место замкнутого пространства. Исследования О'Кифа, изложенные позднее совместно с Л. Наделом (O'Keefe, Nadel, 1978), положили начало настоящему «взрыву» работ. Часть из них была посвящена роли гиппокампа в формировании «когнитивной карты» животного, а часть — разработке новых методических приемов тестирования ориентации животных в пространстве.
Для ориентации «по карте» у животного должны быть представления о расположении как близких, так и далеких объектов среды. Именно пространственные представления такого рода О'Киф и Надел назвали вслед за Толменом когнитивной картой. По их мнению, такие внутренние представления содержат аллоцентрическую пространственную информацию (т.е. информацию, содержание которой не зависит от собственного положения в данном пространстве). Она подобна той, которую можно извлечь при пользовании географической картой. Это, однако, не означает, что такие представления в физическом смысле напоминают карту. Подобные «карты» не являются ни совокупностью некоторого числа связей типа «стимул-реакция», ни цепями УР, а представляют собой, скорее, устойчивые инвариантные ассоциации, вовлекающие ряд стимулов. Такие ассоциации, по-видимому, сходны с теми, что формируются в физиологических экспериментах между двумя условными сигналами (см. 3.2).
Успешность использования животным внутренних пространственных карт основана на стабильности расположения объектов внешней среды. Экспериментально показано, что изменение их положения вносит ошибки в ориентацию.

Рис. 11. Восьмилучевой радиальный лабиринт.
А — автоматизированный радиальный лабиринт с закрытыми рукавами; Б — изображение лабиринта на экране монитора.
Существует целый ряд подходов к исследованию формирования у животного пространственных представлений. Некоторые, как мы только что показали на примере работ Мензела и Пажетнова, связаны с оценкой ориентации животных в естественных условиях. В лаборатории наиболее часто используются две методики — радиальный и водный лабиринты. Роль пространственных представлений и пространственной памяти в формировании поведения в основном исследуется на грызунах, а также некоторых видах птиц.
4.3.1. Обучение в радиальном лабиринте
Методика изучения способности животных к обучению в радиальном лабиринте была предложена американским исследователем Д.Олтоном (Olton, 1978).
Обычно радиальный лабиринт (рис. 3.11) состоит из центральной камеры и 8 (или 12) лучей, открытых или закрытых (называемых в этом случае отсеками, или коридорами). В опытах на крысах длина лучей лабиринта варьирует от 100 до 140 см. Для экспериментов на мышах лучи делают короче. Перед начатом опыта в конец каждого коридора помещают пищу. После процедуры приучения к обстановке опыта голодное животное сажают в центральный отсек, и оно начинает заходить в лучи в поисках пищи. При повторном заходе в тот же отсек животное пищи больше не получает, а такой выбор классифицируется экспериментатором как ошибочный.
По ходу опыта у крыс формируется мысленное представление о пространственной структуре лабиринта. Животные помнят о том, какие отсеки они уже посетили, а в ходе повторных тренировок «мысленная карта» данной среды постепенно совершенствуется. Уже после 7—10 сеансов обучения крыса безошибочно (или почти безошибочно) заходит только в те отсеки, где есть подкрепление, и воздерживается от посещения тех отсеков, где она только что была.
При помещении в радиальный лабиринт многие животные испытывают страх, что затрудняет оценку их способности к пространственному обучению. Возможной причиной этого являются узкие, стесняющие движение коридоры-лучи. В целях более полного анализа способности к усвоению пространственного расположения пищевого подкрепления недавно были начаты исследования обучения животных и птиц разных видов в «гигантском» радиальном лабиринте. Он расположен на открытой местности, а его лучи и центральный отсек в несколько раз больше, чем у лабиринтов, обычно используемых в лабораториях (Lipp et al., 2001).
Методом радиального лабиринта можно оценивать:
формирование пространственной памяти животных;
соотношение таких категорий пространственной памяти, как рабочая и референтная.
Рабочейпамятью называют обычно сохранение информации в пределах одного опыта. Референтная память хранит информацию, существенную для освоения лабиринта в целом.
Для раздельного анализа рабочей и референтной памяти часто используют следующую схему опыта. Животных обучают искать пищу в 4 открытых рукавах, 4 других все время остаются закрытыми, так что в них они зайти не могут (и, следовательно, пищи никогда в них не бывает). В ситуации теста все отсеки открывают, и в поиске пищи животное имеет возможность зайти в любой. Заход только в ранее открытые отсеки, где пищи уже нет, — это ошибка рабочей памяти. Заход в отсек, который при обучении был всегда закрыт, — ошибка референтной памяти.
Пространственная рабочая память достаточно долговечна. Ее длительность определяется временем, в течение которого крыса по окончании эксперимента помнит, в какие лучи она уже заходила. В некоторых случаях это время достигало 24 часов.
Исследуя пространственную память в радиальном лабиринте и проводя с животным длительные опыты, мы исследуем его долгосрочную память (которая может быть референтной или рабочей). Однако, если нам интересно выяснить, как реагирует животное на экстренное изменение ситуации в лабиринте и какова его немедленная реакция, — мы будем иметь дело с его краткосрочной памятью.
Подразделение памяти на референтную и рабочую основано на запоминании пространственных ориентиров и событий (посещение отсека).
Деление памяти на краткосрочную и долгосрочную основано на другом критерии — на продолжительности сохранения следов во времени.
Подразделение памяти на декларативную и процедурную мы рассматривали выше (см. 4.1).
Работы с радиальным лабиринтом позволили выявить у животных (главным образом, крыс) наличие определенных стратегий поиска пищи.
В самой общей форме такие стратегии подразделяются на алло- и эгоцентрические:
при аллоцентрической стратегии животное при поиске пищи полагается на свое мысленное представление о пространственной структуре данной среды;
эгоцентрическая стратегия основана на знании животным конкретных ориентиров и сопоставлении с ними положения своего тела.
Такое деление в большой степени условно, и животное, в особенности в процессе обучения, может параллельно использовать элементы обеих стратегий. Доказательства использования крысами аллоцентрической стратегии (мысленной карты) базируются на многочисленных контрольных экспериментах, в ходе которых либо вводятся новые, «сбивающие» с пути ориентиры (или, наоборот, подсказки), либо меняется ориентация всего лабиринта относительно ранее неподвижных координат и т.д.
4.3.2. Обучение в водном лабиринте Морриса (водный тест) В начале 80-х годов шотландский исследователь Р. Моррис (Morris, 1984) предложил для изучения способности животных к формированию пространственных представлений использовать «водный лабиринт». Метод приобрел большую популярность, и его стали называть «водным лабиринтом Морриса».
Принцип метода заключается в следующем. Животное (обычно мышь или крысу) выпускают в бассейн с водой (рис. 3.12). Из бассейна нет выхода, но имеется невидимая (вода замутнена) подводная платформа, которая может послужить убежищем: отыскав ее, животное может выбраться из воды. Мышь вынимают из бассейна, а через некоторое время снова выпускают плавать, однако уже из другой точки периметра. Постепенно время, которое проходит от пуска животного до отыскания платформы, укорачивается, а путь упрощается. Это свидетельствует о формировании у него представления о пространственном расположении платформы на основе внешних по отношению к бассейну ориентиров. Подобная мысленная карта может быть более или менее точной, а определить, в какой степени животное помнит положение платформы, можно, переместив ее в новое положение. В этом случае время, которое животное проведет, плавая над старым местоположением платформы, будет показателем прочности следа памяти.
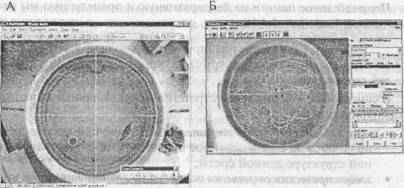
Рис. 12. Водный лабиринт Морриса.
А — проекция на экран монитора круглого бассейна с водой, плавая в котором животное отыскивает невидимую платформу; Б — траектория движений животного, зарегистрированная специальным устройством.
Создание специальных технических средств автоматизации эксперимента с водным лабиринтом (например, системы Noldus) и программного обеспечения для анализа результатов (например, Wolfer, Lipp, 1992) позволило использовать такие данные для точных количественных сравнений поведения животных в тесте. При этом можно оценивать:
динамику формирования пространственного навыка;
стратегии поведения животного в ходе опыта;
обнаруживать слабые отличия в поведении, например у мышей-нокаутов (см. гл. 9).
Такие возможности делают водный тест Морриса важным инструментом не только для изучения когнитивных функций, но и для решения ряда вопросов современной нейрогенетики (см. гл. 9).
Исследование роли отдельных структур мозга в формировании навыка поиска пиши в радиальном и водном лабиринтах показало, что ключевую роль в этом процессе играет гиппокамп. Фармакологические воздействия, относительно избирательно повреждающие эту структуру, нарушают поведение крыс и мышей и в радиальном, и в водном лабиринте.
4.4. Обучение и память животных в ситуациях, приближенных к естественным.
Все большую роль в изучении когнитивных функций животных начинают играть работы, выполненные в естественных для вида или приближенных к ним условиях. Такой подход позволяет отбросить предположение, что в неадекватных лабораторных условиях способность животного к тому или иному виду обучения может проявиться не полностью (например, слишком тесные рукава лабиринта, нечеткие ориентиры, стресс при плавании). Ввиду этого поиск биологически адекватных моделей для изучения способностей к обучению и памяти составляет одну из актуальных задач современной нейробио-логии. Важную роль в разработке таких моделей играет знание видос-пецифического (инстинктивного) поведения животных.
Один из аспектов этих исследований — оценка пространственной памяти в естественной среде обитания.
Пищедобывательное поведение ряда видов птиц из семейств врано-вых (Corvidae) и синицевых (Paridae) характеризуется тем, что осенью они откладывают многочисленные небольшие запасы пищи, а зимой и весной точно находят их. Эту способность и исследуют в качестве экологической модели пространственной памяти птиц. В обоих семействах выраженность такого поведения имеет существенные межвидовые различия. Большинство видов врановых птиц (в том числе грачи, вороны, сороки, галки) делают запасы эпизодически, тогда как сойки и кедровки разных видов (и Нового, и Старого Света) запасают корм систематически. Зимой и весной они отыскивают свои запасы, причем проявляют при этом удивительную точность, которая свидетельствует о запоминании координат каждой из таких «кладовок». Создание запасов пищи и запоминание их пространственных координат входит в обязательный видоспецифический репертуар поведения указанных видов.
Изучение процесса запасания и отыскания пищи кедровками (Nucifraga caryocatactes) имеет длительную историю. Удивительная точность, с которой они отыскивают кладовки, была отмечена натуралистами еще в конце XIX в. В 60-е годы XX в. его впервые начали исследовать экспериментально.
Опыты были проведены в 1962—1963 годах на Телецком стационаре Биологического института СО АН СССР. В сезон плодоношения кедров в построенную прямо В тайге большую вольеру выпускали по очереди кедровок после небольшого периода голодания. Получив доступ к шишкам и утолив первый голод, птицы начинали рассовывать орехи под мох, корни кустарника, под стволы деревьев (по нескольку штук в каждую «кладовку»). Наблюдатели точно картировали расположение кладовок, а птиц затем удаляли из вольера на разные промежутки времени (от нескольких часов до нескольких дней).
По возвращении в вольеру все подопытные птицы (перед этим остававшиеся без корма в течение нескольких часов) безошибочно обнаружили подавляющее число своих кладовок, причем практически не трогали чужих. Кедровки действовали при этом целенаправленно и, по-видимому, совершенно точно помнили, где их запасы находятся.
Характер поведения кедровок полностью соответствовал тому, что наблюдали в естественных условиях орнитологи (Воробьев, 1982). Точность обнаружения кладовок кедровками нарушается при смещении внешних пространственных ориентиров (Крушинская, 1966). Это экспериментатьно подтверждало предположение зоологов о том, что эти птицы точно запоминают место каждой кладовки, а не ищут их наугад. Н. Л. Крушинская, занимавшаяся в 60-е годы исследованием ней-роморфологического субстрата обучения и памяти птиц, предположила, что способность запоминать местоположение кладовки можно рассматривать как модель для изучения механизмов пространственной памяти, и поставила цель проанализировать роль гиппокампа в его осуществлении. Как известно, мозг птиц по общему плану строения радикально отличается от мозга млекопитающих (см.: Обухов, 1999). Тем не менее опыты Н. Л. Крушинской на голубях, а позднее опыты Зиновьевой и Зориной (1976) на врановых и курах свидетельствовали о том, что гиппокамп у птиц, как и у млекопитающих, играет решающую роль в механизме памяти. После разрушения гиппокампа кедровки отыскивали кладовки лишь наугад (Крушинская, 1966). Таким образом, экспериментально было показано, что в основе способности птиц к отысканию спрятанного корма действительно лежит точная фиксация и хранение в памяти местоположения своих кладовок.
Описанные выше опыты стали практически первым исследованием ней-рофизиологческих механизмов поведения птиц в естественной среде обитания. Начиная с 70-х годов феномен пространственной памяти птиц интенсивно изучается в ряде лабораторий США, Канады и Англии. Исследования проводятся на колумбийских кедровках (Nucifraga columbiana) и нескольких видах американских соек, у которых тенденция к устройству кладовок выражена в разной степени (Balda, Kamil, 1992).
Способность к систематическому запасанию корма накладывает отпечаток на общую структурно-функциональную организацию мозга и поведения птиц. Прослеживаются следующие корреляции (подробнее см. обзоры: Clayton, Krebs, 1994, 1995; Shettleworth, 1995).
У птиц тех видов, которые активно запасают корм, отношение объема гиппокампа к объему конечного мозга положительно коррелирует с выраженностью поведения запасания (Basil et al., 1996). Чем большую роль в выживании вида играет использование запасенной осенью пищи, тем большим бывает относительный объем гиппокампа.
У врановых тех видов, которые систематически запасают корм, взаимодействия между полушариями мозга в процессе обработки зрительной информации и фиксации следов памяти организованы иначе, чем у незапасающих. Запасающие виды отличаются от незапасающих по степени межполушарной асимметрии зрительных функций. У первых система структур, связанных с правым глазом, обрабатывает информацию главным образом о специфических признаках предметов, тогда как «система левого глаза» передает информацию преимущественно о пространственных признаках среды. У незапасающих видов такое разделение выражено значительно в меньшей степени (Clayton, Krebs, 1994, 1995).
Существует прямая зависимость между выраженностью запасания у 5 видов врановых (4 вида американских соек и колумбийской кедровки) и их способностью к некоторым видам обучения, включая радиальный лабиринт (Balda, Kamil, 1992; Shettleworth, 1995). Чем более важную роль в выживании вида играет способность создавать запасы корма, тем точнее птицы запоминают пространственные координаты кладовок и тем лучше решают аналогичные задачи в эксперименте.
Представляло интерес выяснить, какие характеристики событий, наряду с запоминанием пространственных координат, фиксируют птицы при устройстве запасов. Работы английской исследовательницы Н.Клэйтон показывают, что они запоминают более сложную информацию и могут ответить не только на вопрос, где спрятана пища, но также что спрятано и когда это произошло. В одном из экспериментов сойки запасали два вида корма: орехи и мучных червей. Оказалось, что при испытании через 4 часа они одинаково точно и активно находили оба вида запасов, тогда как через 104 часа искали только орехи и не подходили к кладовкам со «скоропортящимся продуктом». На основании этих и ряда подобных фактов такого рода когнитивную деятельность птиц можно рассматривать как проявление «памяти на эпизоды».
Птицы способны запоминать и хранить информацию не только о месте расположения запаса, но и о времени события, а также о пространственно-временных соотношениях.
Данные о том, что птицы помнят не только, где спрятано, но также что и когда спрятано (Clayton et al., 2000), представляются весьма актуальными, поскольку такой вид памяти (episodic memory) до недавнего времени считался прерогативой человека. Особый интерес вызывают исследования, в которых показано увеличение размеров гиппокампа у запасающих видов синиц в период устройства кладовок.
Пространственную память исследуют и у почтовых голубей. В этих экспериментах также учитывается биология изучаемого вида. Как известно, возвращение домой, в голубятню особенно четко выражено у голубей специальных пород, а опыт такой селекции восходит к достаточно далекому времени. Считается, что именно голуби приносили домой сообщения о победе спортсменов в периоды Олимпийских игр в Древней Греции.
Вопрос о том, в какой степени голубь, увезенный за сотни километров от дома, ориентируется по мысленной пространственной карте (Lipp, 1983), а в какой степени — по заученным ориентирам, насколько в этом участвуют обоняние (Papi et al., 1995) и восприятие магнитного поля, не имеет окончательного ответа (Bingman et al., 1995). Эти формы когнитивной деятельности голубей можно исследовать анатомо-физиологическими методами, например путем удаления некоторых структур мозга, в частности гиппокампа, а также с помощью фармакологических препаратов с известным типом действия. Формирование пространственных представлений у голубей при естественной навигации можно сопоставить с данными по их пространственному обучению в чисто лабораторных опытах.
4.5 Обучение «выбору по образцу».
«Выбор по образцу» — один из видов когнитивной деятельности, также основанный на формировании у животного внутренних представлений о среде. Однако в отличие от обучения в лабиринтах этот экспериментальный подход связан с обработкой информации не о пространственных признаках, а о соотношениях между стимулами — наличии сходства или отличия между ними.
Метод «выбора по образцу» был введен в начале XX в. Н. Н. Ладыгиной-Котс и с тех пор широко используется в психологии и физиологии. Он состоит в том, что животному демонстрируют стимул-образец и два или несколько стимулов для сопоставления с ним, подкрепляя выбор того, который соответствует образцу. Существует несколько вариантов «выбора по образцу»:
выбор из двух стимулов — альтернативный:
выбор из нескольких стимулов — множественный;
отставленный выбор — подбор «пары» предъявленному стимулу животное производит в отсутствие образца, ориентируясь не на реальный стимул, а на его мысленный образ, на представление о нем.
Когда животное выбирает нужный стимул, оно получает подкрепление. После упрочения реакции стимулы начинают варьировать, проверяя, насколько прочно животное усвоило правила выбора. Следует подчеркнуть, что речь идет не о простой выработке связи между определенным стимулом и реакцией, а о процессе формирования правила выбора, основанного на представлении о соотношении образца и одного из стимулов.
Успешное решение задачи при отставленном выборе также заставляет рассматривать данный тест как способ оценки когнитивных функций мозга и использовать его для изучения свойств и механизмов памяти.
Используются в основном две разновидности этого метода:
выбор по признаку сходства (соответствия) с образцом (matching to sample);
выбор по признаку отличия (несоответствия) от образца (oddity problem, non-matching to sample).
Отдельно надо отметить так называемый символьный, или знаковый, выбор по образцу (symbolic matching, arbitrary matching). В этом случае животное обучают выбирать стимул А при предъявлении стимула X и стимул В — при предъявлении Y в качестве образца. При этом стимулы А и X, В и Y не должны иметь ничего общего между собой. В обучении по этой методике на первых порах существенную роль играют чисто ассоциативные процессы — заучивание правила «если..., то...».
Первоначально опыт ставился так: экспериментатор показывал обезьяне какой-либо предмет (образец), а она должна была выбрать такой же из других предлагаемых ей двух или более предметов. Затем на смену прямому контакту с животным, когда экспериментатор держал в руках стимул-образец и забирал из рук обезьяны выбранный ею стимул, пришли современные экспериментальные установки, в том числе и автоматизированные, полностью разделившие животное и экспериментатора. В последние годы для этой цели используют компьютеры с монитором, чувствительным к прикосновению, а правильно выбранный стимул автоматически перемещается по экрану и останавливается рядом с образцом.
Голубей и крыс обучают выбору по образцу в камере Скиннера (см. рис. 3.6). На диск, расположенный в центре, проецируется образец, на боковые диски — стимулы для сравнения. В качестве стимулов используется, как правило, подсвет дисков разными цветами. Животное осуществляет выбор путем клевания одного из боковых дисков (голубь) или нажатия на него мордой или лапой (крыса).
Иногда ошибочно считают, что обучение выбору по образцу — это то же самое, что выработка дифференцировочных УР. Однако это не так: при дифференцировке происходит только образование реакции на присутствующие в момент обучения стимулы.
При «выборе по образцу» основную роль играет мысленное представление об отсутствующем в момент выбора образце и выявление на его основе соотношения между образцом и одним из стимулов.
Метод обучения выбору по образцу наряду с выработкой диффе-ренцировок используется для выявления способности животных к обобщению.
4.6. Заучивание последовательностей стимулов.
Интерес к этой форме когнитивной деятельности животных возник в связи с обучением обезьян языкам-посредникам, при котором выяснилось, что шимпанзе могут составлять «фразы» из нескольких «слов»-жестов и понимать смысл обращенных к ним «высказываний» (см. гл. 6). Г. Террес (Terrace et al., 1977), один из авторов этих ранних работ, проанализировал структуру таких фраз и высказал гипотезу, что в основе подобного поведения лежит не истинное понимание правильного порядка слов в английском предложении, а более простая и, вероятно, более универсальная для животных способность запоминать длинные ряды («списки») стимулов.
Процесс запоминания цепей стимулов путем их разделения на подгруппы называется «делением на куски» («chunking»). Террес подробно исследовал этот процесс на голубях и показал, что в основе такого запоминания лежит не фиксация некоей последовательности стимулов и ответов, как это предполагалось бы с позиций бихевиоризма и теории условных рефлексов (цепи УР). Эксперименты показали, что здесь происходит формирование иерархически организованных внутренних представлений о структуре такой последовательности, т.е. проявляется способность мысленно проанализировать ряд стимулов и запомнить их, разбивая такой ряд на отрезки, подобно тому, как человек запоминает номера телефонов группами по 3 цифры (Terrace, 1991; Terrace, Chien, 1991a, b).
Исследования Терреса позволяют понять, каким образом животные обрабатывают информацию о совокупностях, или цепях воспринимаемых стимулов, чтобы выполнить в ответ серию действий.
Предполагают, что внутренняя организация таких представлений осуществляется сходным образом у животных разного уровня организации. У голубей подобное обучение протекает так же, как у высших приматов, однако скорость обучения у них гораздо ниже, а последовательности, которые они в состоянии запомнить, менее сложные (D'Amato, Colombo, 1988).
4.7. Инсайт-обучение.
Термин «инсайт-обучение» (Thorpe, 1963) был введен в 60-е годы для описания ряда случаев сложных форм обучения, а также проявлений мышления, которые явно нельзя было отнести ни к одной из упомянутых выше простых категорий. Его использовали в случаях, когда решение задачи происходило слишком быстро для обычного обучения методом «проб и ошибок». К инсайт-обучению относили описанное В.Келером (1925; 1997) поведение шимпанзе, соединившего две палки, чтобы достать недоступное лакомство, а также опыты Н. Майера (Maier, 1929), в которых, как выражался автор, можно было тестировать способность крыс «к рассуждению» (см. 4.8). Однако согласно современным представлениям эти формы поведения относятся к проявлениям мышления, и они будут рассмотрены в следующих главах. В настоящее время термин «инсайт-обучение» употребляется все реже, уступая место конкретным определениям тех или иных форм обучения или рассудочной деятельности. Так, в 4-м и 5-м изданиях одного из самых известных учебников по поведению животных (Manning, Dawkins, An Introduction in Animal Behavior, 1992; 1998) появился раздел «Могут ли животные думать и осмысливать, планировать свои действия?» В нем авторы приводят ряд примеров разумных действий животных в природе (которые ранее они классифицировали как инсайт-обучение), а также описывают некоторые эксперименты, уделяя особое внимание необходимости строгого анализа и возможности использования «канона К. Л. Моргана» (см. 2.3) при трактовке подобных данных.
Резюме.
Индивидуально-приспособительная деятельность животного и в эксперименте, и в естественных условиях представляет собой сплав способностей к ассоциативному обучению и когнитивным процессам разного уровня сложности. Не только в естественных условиях жизни, но и в спланированных экспериментах бывает трудно разделить ассоциативное обучение и когнитивные процессы. Тем не менее существуют экспериментальные подходы, позволяющие анализировать эти явления раздельно. О них будет рассказано в последующих главах книги.
Какие категории поведения относятся к основным типам индивидуально-приспособительной деятельности?
Каковы основные принципы учения о высшей нервной деятельности?
В чем сходство и отличие классических и инструментальных условных рефлексов?
Какие методы позволяют оценивать способность животных к
обобщению?
Как исследуют формирование у животных пространственных представлений и в чем оно выражается?
Можно ли исследовать формирование пространственных представлений у животных в естественных условиях?
|
