| Банк рефератов содержит более 364 тысяч рефератов, курсовых и дипломных работ, шпаргалок и докладов по различным дисциплинам: истории, психологии, экономике, менеджменту, философии, праву, экологии. А также изложения, сочинения по литературе, отчеты по практике, топики по английскому. |

|
Ошибка опыта: Sx=0,811666 Xc=0,476875 Sd=0,1147869 НСР-05=0,236461 Анализируя данную таблицу можно сделать выводы, что форма сорта Ренет Черненко О-43 превосходит контроль на 3,56% (0,19г.), но это является несущественной разницей, поэтому ее преимущества не могут считаться значимыми, но следует обратить внимание на эту форму в дальнейшем, т. к. результат может измениться. Остальные формы в большей или меньшей степени уступают контролю. Худший результат показала форма Ренет Черненко о-35 в-2 (49,4%), что превышает контрольное значение приблизительно в 3 раза. Таблица 5.Сравнение потери влаги сорта Уэлси с его мутантными и гибридными формами.
Ошибка опыта: Sx=0,0263625 Xc=0,3545 Sd=0,1168653 НСР-05=0,2489231 Проанализировав данные таблицы делаем вывод, что формы Мутанты Уэлси и Уэлси 2 побег превосходят контрольный сорт на 2,42% (0,27г) и 2,21% (0,23 г.). Форма Мутанты Уэлси существенно выше контроля по засухоустойчивости. Преимущества другой формы несущественны и считаются ошибкой в пределах опыта. Таблица 6.Сравнение потери влаги сорта Мелба с его мутантными и гибридными формами.
Ошибка опыта: Sx=0,04551155 Xc=0,440556 Sd=0,454756 НСР-05=0,1319443 Из данных таблицы можно сделать вывод, что ни одна форма не превосходит контроль по засухоустойчивости. Средние значения формы 5-69 2 побег и 7-119 6 побег практически не отличаются от данных контроля, а формы о-58 и о-62 превысили контрольные данные приблизительно в 3 раза. Следует проводить дальнейшее наблюдения за ними в данном направлении, а так же проводить оценку на другие ценные признаки. Рис.3 Плоды сорта Мелба. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 ис.1
Сад ВНИИС им.
Мичурина.
ис.1
Сад ВНИИС им.
Мичурина.

|
Сорта, мутанты и гибриды |
Первичная масса |
Масса после сушки |
Разница |
% соотношение |
Среднее из 4-х повт. %/г. |
|
Мартовское (контроль) |
3,09 |
2,36 |
0,73 |
23,62% |
18,58%/0,60 |
|
3,42 |
2,86 |
0,56 |
16,37% |
||
|
2,58 |
2,19 |
0,39 |
15,12% |
||
|
3,70 |
2,99 |
0,71 |
19,19% |
||
|
Мутанты Мартовского 6-38 |
4,45 |
3,33 |
1,12 |
25,17% |
25,20%/1,16 |
|
4,65 |
3,42 |
1,23 |
26,45% |
||
|
4,69 |
3,34 |
1,35 |
28,78% |
||
|
4,51 |
3,59 |
0,92 |
20,40% |
||
|
Мартовское с-52 |
3,99 |
3,69 |
0,30 |
7,52% |
9,62%/0,37 |
|
4,25 |
3,81 |
0,44 |
10,35% |
||
|
3,91 |
3,57 |
0,34 |
8,70% |
||
|
3,36 |
2,96 |
0,40 |
11,90% |
||
|
Мартовское с-54 |
2,57 |
2,30 |
0,27 |
10,51% |
10,43%0,26 |
|
3,06 |
2,72 |
0,34 |
11,11% |
||
|
1,80 |
1,61 |
0,19 |
10,56% |
||
|
2,30 |
2,08 |
0,22 |
9,57% |
||
|
Мартовское с-55 |
2,41 |
2,20 |
0,21 |
8,71% |
9,05%/0,27 |
|
3,29 |
2,99 |
0,30 |
9,12% |
||
|
2,64 |
2,41 |
0,23 |
8,71% |
||
|
3,42 |
3,09 |
0,33 |
9,65% |
||
|
Мартовское с-56 |
3,70 |
3,40 |
0,30 |
8,11% |
7,56%/0,23 |
|
3,66 |
3,41 |
0,25 |
6,83% |
||
|
2,54 |
2,32 |
0,22 |
8,66% |
||
|
2,11 |
1,97 |
0,14 |
6,64% |
||
|
Мартовское с-57 |
2,23 |
2,01 |
0,22 |
9,87% |
10,17%/0,32 |
|
3,80 |
3,42 |
0,38 |
10,00% |
||
|
3,10 |
2,80 |
0,30 |
9,68% |
||
|
3,50 |
3,11 |
0,39 |
11,14% |
||
|
Мартовское с-58 |
1,59 |
1,40 |
0,19 |
11,95% |
11,56%/0,21 |
|
1,90 |
1,69 |
0,21 |
11,05% |
||
|
1,82 |
1,59 |
0,23 |
12,64% |
||
|
1,79 |
1,60 |
0,19 |
10,61% |
Ошибка опыта: Sx=0,502495
Xc=0,425
Sd=0,07106352
НСР-05=0,1463908
Проанализировав таблицу можно сделать вывод, что сразу несколько форм (с-52; с-54; с-55; с-56; с-57; с-58; ) превосходят контроль, ( рис. 4) причем достаточно значительно, лишь одна форма (6-38) уступает ему. Форма с-52 превосходит контроль на 8,96% (0,23 г); форма с-54 превосходит на 8,14%(0,34 г); форма с-55—на 9,53%(0,33 г); форма с-56—на 11,02%(0,36 г); форма с-57—на 8,4%(028 г); форма с-58—на 7,01%(0,39 г). В отличие от них форма 6-38 уступает контролю на 6,62%(0,56 г.) Разница у всех форм превосходящих контроль является существенной. С-52—51,78%; с-54—56,17%; с-55—48,71%; с-56—40,69%; с-57—54,76%; с-58—62,25%. Форма, наиболее сильно превосходящая контроль—с-58. Следует все эти формы вовлечь в селекционный процесс, для получения возможных источников и доноров засухоустойчивости.
Рис.4 Селекционная школка, годе высажены изучаемые формы Мартовского

Таблица 8.
Сравнение потери влаги сорта Жигулевское с его мутантными и гибридными формами.
|
Сорта, мутанты и гибриды |
Первичная масса |
Масса после сушки |
Разница |
% соотношение |
Среднее из 4-х повт. %/г. |
|
Жигулевское(контроль) |
11,90 |
10,80 |
1,10 |
9,24% |
9,56%/1,02 |
|
9,69 |
8,71 |
0,98 |
10,11% |
||
|
10,67 |
9,60 |
1,07 |
10,03% |
||
|
10,52 |
9,59 |
0,93 |
8,84% |
||
|
Жигулевское 100 грей о-24 №21 |
2,20 |
1,70 |
0,50 |
22,73% |
22,96%/0,54 |
|
2,21 |
1,72 |
0,49 |
22,17% |
||
|
2,52 |
2,00 |
0,52 |
20,63% |
||
|
2,51 |
1,85 |
0,66 |
26,29% |
||
|
Жигулевское 100 грей о-25 |
2,60 |
2,40 |
0,20 |
7,69% |
11,04%/0,20 |
|
0,99 |
0,86 |
0,13 |
13,13% |
||
|
1,80 |
1,65 |
0,15 |
8,33% |
||
|
2,00 |
1,70 |
0,30 |
15,00% |
||
|
Жигулевское 100 грей о-28 |
2,38 |
1,16 |
1,22 |
51,26% |
32,99%/0,86 |
|
3,03 |
2,01 |
1,02 |
33,66% |
||
|
2,80 |
2,30 |
0,50 |
17,86% |
||
|
2,40 |
1,70 |
0,70 |
29,17% |
||
|
Жигулевское 100 грей о-29 |
3,20 |
2,70 |
0,50 |
15,63% |
16,41%/0,51 |
|
3,30 |
2,71 |
0,59 |
17,88% |
||
|
3,00 |
2,60 |
0,40 |
13,33% |
||
|
2,82 |
2,29 |
0,53 |
18,79% |
||
|
Жигулевское 100 грей о-30 |
2,00 |
1,62 |
0,38 |
19,00% |
16,94%/0,24 |
|
0,70 |
0,59 |
0,11 |
15,71% |
||
|
1,50 |
1,20 |
0,30 |
20,00% |
||
|
1,38 |
1,20 |
0,18 |
13,04% |
||
|
Жигулевское 2-в о-34 |
1,40 |
1,10 |
0,30 |
21,43% |
21,67%/0,29 |
|
0,90 |
0,65 |
0,25 |
27,78% |
||
|
1,30 |
0,99 |
0,31 |
23,85% |
||
|
2,20 |
1,90 |
0,30 |
13,64% |
||
|
Жигулевское 2-в о-37 |
3,98 |
3,19 |
0,79 |
19,85% |
21,98%/0,82 |
|
3,78 |
2,82 |
0,96 |
25,40% |
||
|
3,52 |
2,78 |
0,74 |
21,02% |
||
|
3,60 |
2,82 |
0,78 |
21,67% |
||
|
Жигулевское 2-в о-38 |
2,40 |
1,90 |
0,50 |
20,83% |
21,03%/0,66 |
|
3,00 |
2,25 |
0,75 |
25,00% |
||
|
3,60 |
3,00 |
0,60 |
16,67% |
||
|
3,70 |
2,90 |
0,80 |
21,62% |
Ошибка опыта: Sx=6,907475
Xc=0,5705556
Sd=9,768645
НСР-05=0,2002572
Проанализировав таблицу можно придти к заключению, что лучшим из выборки оказался контрольный сорт (рис. 5). Все формы (кроме о-25) значительно уступают контролю, следовательно, из данной выборки ни одна форма не превышает контроля, следует проводить дальнейшие наблюдения за ними. В среднем все образцы отличаются от контрольного худшими в 2 раза показателями, и только форма Жигулевское 100 грей о-28 показала результаты превышающие контрольные значения более чем в три раза.
Рис. 5 Цветение Жигулевского.

Таблица 9.
Сравнение потери влаги сорта Лобо с его мутантными и гибридными формами.
|
Сорта, мутанты и гибриды |
Первичная масса |
Масса после сушки |
Разница |
% соотношение |
Среднее из 4-х повт. %/г. |
|
Лобо (контроль) |
14,30 |
13,05 |
1,25 |
8,74% |
9,81%//1,13 |
|
8,65 |
7,69 |
0,96 |
11,10% |
||
|
11,40 |
10,30 |
1,10 |
9,65% |
||
|
12,32 |
11,12 |
1,20 |
9,74% |
||
|
Лобо в-2 №10 о-10 |
0,60 |
0,50 |
0,10 |
16,67% |
19,25%/0,14 |
|
0,90 |
0,70 |
0,20 |
22,22% |
||
|
0,70 |
0,55 |
0,15 |
21,43% |
||
|
0,60 |
0,50 |
0,10 |
16,67% |
||
|
Лобо в-2 №9 о-6 |
3,80 |
3,52 |
0,28 |
7,37% |
13,24%/0,55 |
|
4,50 |
3,68 |
0,82 |
18,22% |
||
|
4,00 |
3,10 |
0,90 |
22,50% |
||
|
4,10 |
3,90 |
0,20 |
4,88% |
||
|
Лобо в-2 о-17 |
2,05 |
1,55 |
0,50 |
24,39% |
27,07%/0,62 |
|
1,61 |
1,10 |
0,51 |
31,68% |
||
|
2,53 |
1,87 |
0,66 |
26,09% |
||
|
3,10 |
2,29 |
0,81 |
26,13% |
||
|
Лобо в-2 о-8 |
2,65 |
1,98 |
0,67 |
25,28% |
24,64%/0,44 |
|
2,10 |
1,62 |
0,48 |
22,86% |
||
|
1,20 |
0,96 |
0,24 |
20,00% |
||
|
1,15 |
0,80 |
0,35 |
30,43% |
||
|
Лобо в-3 о-20 |
12,60 |
11,52 |
1,08 |
8,57% |
8,41%/1,21 |
|
13,09 |
11,99 |
1,10 |
8,40% |
||
|
17,30 |
15,81 |
1,49 |
8,61% |
||
|
14,38 |
13,22 |
1,16 |
8,07% |
||
|
Лобо о-65 |
1,78 |
1,21 |
0,57 |
32,02% |
32,94%/0,75 |
|
2,20 |
1,40 |
0,80 |
36,36% |
||
|
2,60 |
1,85 |
0,75 |
28,85% |
||
|
2,52 |
1,65 |
0,87 |
34,52% |
||
|
Лобо О-43 |
4,12 |
3,20 |
0,92 |
22,33% |
21,41%/0,74 |
|
2,90 |
2,28 |
0,62 |
21,38% |
||
|
3,39 |
2,65 |
0,74 |
21,83% |
||
|
3,38 |
2,70 |
0,68 |
20,12% |
Ошибка опыта: Sx=0,09259
Xc=0,695621
Sd=0,11309421
НСР-05=0,2697407
Анализ таблицы показывает, что одна форма (в-3 о-20) превосходит контроль. Различия в показателях являются несущественными (0,08 г. 16,564 %). Следовательно хотя данная форма хотя и имеет более высокий показатель не существенно отличается от контрольной. Остальные формы хуже контроля в 1,5-2 раза, а форма Лобо о-65 показала результат худший практически в 4 раза.
Рис.6 Сад ВНИИС им. Мичурина, где были отобраны контрольные пробы.

Таблица 10.
Сравнение потери влаги сорта Спартак с его мутантными и гибридными формами.
|
Сорта, мутанты и гибриды |
Первичная масса |
Масса после сушки |
Разница |
% соотношение |
Среднее из 4-х повт. %/г. |
|
Спартак (контр) |
2,90 |
2,45 |
0,45 |
15,52% |
16,89%/0,44 |
|
2,50 |
1,99 |
0,51 |
20,40% |
||
|
2,60 |
2,10 |
0,50 |
19,23% |
||
|
2,50 |
2,19 |
0,31 |
12,40% |
||
|
Спартак 100 грей о-1 |
3,70 |
3,05 |
0,65 |
17,57% |
22,75%/0,63 |
|
3,00 |
2,32 |
0,68 |
22,67% |
||
|
2,39 |
1,72 |
0,67 |
28,03% |
||
|
2,20 |
1,70 |
0,50 |
22,73% |
||
|
Спартак 100 грей о-12 №13 |
2,60 |
2,10 |
0,50 |
19,23% |
17,18%/0,39 |
|
2,50 |
2,15 |
0,35 |
14,00% |
||
|
2,30 |
1,79 |
0,51 |
22,17% |
||
|
1,50 |
1,30 |
0,20 |
13,33% |
||
|
Спартак 100 грей о-2 |
2,50 |
2,10 |
0,40 |
16,00% |
20,32%/0,51 |
|
1,60 |
1,30 |
0,30 |
18,75% |
||
|
2,80 |
2,27 |
0,53 |
18,93% |
||
|
2,90 |
2,10 |
0,80 |
27,59% |
||
|
Спартак 100 грей о-3 |
3,20 |
2,30 |
0,90 |
28,13% |
25,68%/0,67 |
|
2,40 |
1,82 |
0,58 |
24,17% |
||
|
2,00 |
1,51 |
0,49 |
24,50% |
||
|
2,70 |
2,00 |
0,70 |
25,93% |
||
|
Спартак 100 грей о-4 |
2,40 |
2,05 |
0,35 |
14,58% |
17,45%/0,29 |
|
1,70 |
1,42 |
0,28 |
16,47% |
||
|
1,60 |
1,38 |
0,22 |
13,75% |
||
|
1,20 |
0,90 |
0,30 |
25,00% |
||
|
Спартак 1-в о-5 |
2,30 |
2,05 |
0,25 |
10,87% |
11,72%0,30 |
|
2,50 |
2,22 |
0,28 |
11,20% |
||
|
2,70 |
2,42 |
0,28 |
10,37% |
||
|
2,70 |
2,31 |
0,39 |
14,44% |
||
|
Спартак 2-в о-11 |
2,60 |
1,38 |
1,22 |
46,92% |
26,73%/0,52 |
|
1,25 |
1,00 |
0,25 |
20,00% |
||
|
1,50 |
1,20 |
0,30 |
20,00% |
||
|
1,50 |
1,20 |
0,30 |
20,00% |
||
|
Спартак 2-в о-19 №16 |
1,08 |
0,92 |
0,16 |
14,81% |
15,25%/0,23 |
|
1,90 |
1,70 |
0,20 |
10,53% |
||
|
1,65 |
1,38 |
0,27 |
16,36% |
||
|
1,40 |
1,13 |
0,27 |
19,29% |
||
|
Спартак 2-в о-7 |
2,60 |
2,18 |
0,42 |
16,15% |
20,08%/0,36 |
|
1,80 |
1,41 |
0,39 |
21,67% |
||
|
1,60 |
1,20 |
0,40 |
25,00% |
||
|
1,20 |
0,99 |
0,21 |
17,50% |
Ошибка опыта: Sx=0,94122
Xc=0,43175
Sd=0,1331086
НСР-05=0,2715415
Проанализировав таблицу можно сказать, что две формы (1-в о-5 и 2-в о-19 №16) выявили более высокий показатель по сравнению с контрольным вариантом (рис. 7). Форма 1-в о-5 превзошла контроль на 5,17% (0,14 г), а форма 2-в о-19 № 16 на 1,64 % (0,21 г.), что не является существенной разницей. Следовательно, эти две формы с высокой степенью достоверности можно считать менее засухоустойчивыми. Из данной выборки нет резких отклонений, результаты не превышают контроль более чем в два раза. Следует обратить внимание на данную выборку и провести оценку на ее стабильность.
Рис. 7 Исследуемые формы из селекционной школки.

Таблица 11.
Сравнение потери влаги сорта Северный синап с его мутантными и гибридными формами.
|
Сорта, мутанты и гибриды |
Первичная масса |
Масса после сушки |
Разница |
% соотношение |
Среднее из 4-х повт. %/г. |
|
Сев синап (контр) |
3,80 |
3,28 |
0,52 |
13,68% |
16,45%/,57 |
|
3,10 |
2,49 |
0,61 |
19,68% |
||
|
3,30 |
2,75 |
0,55 |
16,67% |
||
|
3,80 |
3,20 |
0,60 |
15,79% |
||
|
Сев Синап 100 грей о-49 |
1,00 |
0,68 |
0,32 |
32,00% |
26,83%/0,31 |
|
1,30 |
0,99 |
0,31 |
23,85% |
||
|
0,70 |
0,49 |
0,21 |
30,00% |
||
|
1,91 |
1,50 |
0,41 |
21,47% |
||
|
Сев Синап 2-в о-54 |
2,60 |
1,82 |
0,78 |
30,00% |
25,34%/0,62 |
|
2,78 |
1,82 |
0,96 |
34,53% |
||
|
1,90 |
1,59 |
0,31 |
16,32% |
||
|
2,00 |
1,59 |
0,41 |
20,50% |
||
|
Сев синап 2-в о-55 |
1,59 |
1,38 |
0,21 |
13,21% |
22,21%/0,62 |
|
2,80 |
2,22 |
0,58 |
20,71% |
||
|
3,10 |
2,30 |
0,80 |
25,81% |
||
|
3,09 |
2,19 |
0,90 |
29,13% |
||
|
Сев Синап 2-в о-57 |
1,80 |
1,32 |
0,48 |
26,67% |
24,08%/0,40 |
|
1,55 |
1,29 |
0,26 |
16,77% |
||
|
1,72 |
1,26 |
0,46 |
26,74% |
||
|
1,53 |
1,13 |
0,40 |
26,14% |
||
|
Сев Синап 2-в о-59 |
1,78 |
1,50 |
0,28 |
15,73% |
13,74%/0,27 |
|
1,90 |
1,60 |
0,30 |
15,79% |
||
|
2,08 |
1,79 |
0,29 |
13,94% |
||
|
2,21 |
2,00 |
0,21 |
9,50% |
||
|
Сев Синап о-50 |
1,30 |
1,05 |
0,25 |
19,23% |
30,51%/0,42 |
|
1,10 |
0,62 |
0,48 |
43,64% |
||
|
1,20 |
0,82 |
0,38 |
31,67% |
||
|
2,00 |
1,45 |
0,55 |
27,50% |
||
|
Сев Синап о-51 |
1,15 |
0,39 |
0,76 |
66,09% |
51,92%/0,64 |
|
1,20 |
0,49 |
0,71 |
59,17% |
||
|
1,20 |
0,72 |
0,48 |
40,00% |
||
|
1,39 |
0,80 |
0,59 |
42,45% |
||
|
Сев Синап о-52 2-в |
1,61 |
1,09 |
0,52 |
32,30% |
25,64%/0,58 |
|
2,85 |
2,20 |
0,65 |
22,81% |
||
|
2,75 |
2,10 |
0,65 |
23,64% |
||
|
2,10 |
1,60 |
0,50 |
23,81% |
||
|
Сев Синап о-53 2-в |
3,50 |
2,58 |
0,92 |
26,29% |
26,58%/0,78 |
|
3,40 |
2,51 |
0,89 |
26,18% |
||
|
3,30 |
2,56 |
0,74 |
22,42% |
||
|
1,75 |
1,20 |
0,55 |
31,43% |
Ошибка опыта: Sx=0,1091853
Xc=0,6264286
Sd=0,1544113
НСР-05=0,3211755
Анализируя данную таблицу можно заявить, что форма 2-в о-59 показала более высокие результаты по сравнению с контрольным вариантом. Разница--19,72% 0,27 г. Это незначительный показатель, следовательно, с высокой степенью достоверности позволяет сказать, что данная форма находится на уровне контрольного варианта (рис. 8). Форма о-51 показала результат (51,92%), превышающий контрольное значение более чем в 3 раза.
Рис. 8 Северный Синап

Таблица 12.
Сравнение потери влаги сорта Декабренок с его мутантными и гибридными формами.
|
Сорта, мутанты и гибриды |
Первичная масса |
Масса после сушки |
Разница |
% соотношение |
Среднее из 4-х повт. %/г. |
|
Декабренок (контроль) |
4,75 |
4,08 |
0,67 |
14,11% |
15,24%/0,64 |
|
4,00 |
3,40 |
0,60 |
15,00% |
||
|
3,70 |
3,11 |
0,59 |
15,95% |
||
|
4,40 |
3,70 |
0,70 |
15,91% |
||
|
Декабренок о-48 |
2,08 |
1,41 |
0,67 |
32,21% |
30,44%/0,65 |
|
2,12 |
1,21 |
0,91 |
42,92% |
||
|
1,80 |
1,40 |
0,40 |
22,22% |
||
|
2,50 |
1,89 |
0,61 |
24,40% |
||
|
Декабренок 2-в о-45 |
1,30 |
1,05 |
0,25 |
19,23% |
22,15%/0,30 |
|
1,20 |
0,92 |
0,28 |
23,33% |
||
|
1,52 |
1,20 |
0,32 |
21,05% |
||
|
1,40 |
1,05 |
0,35 |
25,00% |
||
|
Декабренок 100 грей о-17 |
1,52 |
1,30 |
0,22 |
14,47% |
12,96%/0,33 |
|
3,54 |
2,80 |
0,74 |
20,90% |
||
|
2,50 |
2,25 |
0,25 |
10,00% |
||
|
1,70 |
1,59 |
0,11 |
6,47% |
||
|
Декабренок 100 грей о-19 |
3,50 |
2,50 |
1,00 |
28,57% |
26,65%/0,79 |
|
2,60 |
2,00 |
0,60 |
23,08% |
||
|
3,00 |
2,30 |
0,70 |
23,33% |
||
|
2,75 |
1,88 |
0,87 |
31,64% |
||
|
Декабренок 100 грей о-20 |
1,50 |
1,00 |
0,50 |
33,33% |
27,70%/0,43 |
|
1,20 |
0,80 |
0,40 |
33,33% |
||
|
2,61 |
2,30 |
0,31 |
11,88% |
||
|
1,55 |
1,05 |
0,50 |
32,26% |
||
|
Декабренок 100 грей о-21 |
1,00 |
0,75 |
0,25 |
25,00% |
28,42%/0,36 |
|
1,50 |
1,00 |
0,50 |
33,33% |
||
|
1,30 |
0,75 |
0,55 |
42,31% |
||
|
1,15 |
1,00 |
0,15 |
13,04% |
||
|
Декабренок 100 грей о-22 |
1,22 |
0,80 |
0,42 |
34,43% |
25,56%/0,29 |
|
1,12 |
0,90 |
0,22 |
19,64% |
||
|
1,00 |
0,80 |
0,20 |
20,00% |
||
|
1,10 |
0,79 |
0,31 |
28,18% |
||
|
Декабренок 100 грей о-23 |
2,59 |
1,50 |
1,09 |
42,08% |
43,40%/0,96 |
|
2,48 |
1,03 |
1,45 |
58,47% |
||
|
1,60 |
1,10 |
0,50 |
31,25% |
||
|
1,89 |
1,10 |
0,79 |
41,80% |
Ошибка опыта: Sx=01027785
Xc=0,5272233
Sd=0,1453508
НСР-05=0,2979691
Проанализировав данные таблицы можно заявить, что одна форма о-17, показала более высокие результаты, чем контроль. Разница составляет 55,039 %-0,28 г, что не превышает существенной разницы. Про данную форму можно с высокой степенью достоверности можно заявить, что она не существенно не превышает контрольного сорта. Худший показатель у формы Декабренок 100 грей о-23, превышающий контроль почти в 3 раза.
4.2 Сравнительная оценка потери H2O между перспективными мутантными и гибридными формами яблони.
Таблица 13.
Сравнение наиболее перспективных гибридных и мутантных форм яблони между собой.
|
Сорта, мутанты и гибриды |
Первичная масса |
Масса после сушки |
Разница |
% соотношение |
Средний % из 4-х повт. |
|
Мутанты Уэлси |
2,22 |
2,00 |
0,22 |
9,91% |
8,53% |
|
2,52 |
2,26 |
0,26 |
10,32% |
||
|
2,85 |
2,68 |
0,17 |
5,96% |
||
|
3,40 |
3,13 |
0,27 |
7,94% |
||
|
Мартовское с-52 |
3,99 |
3,69 |
0,30 |
7,52% |
9,62% |
|
4,25 |
3,81 |
0,44 |
10,35% |
||
|
3,91 |
3,57 |
0,34 |
8,70% |
||
|
3,36 |
2,96 |
0,40 |
11,90% |
||
|
Мартовское с-54 |
2,57 |
2,30 |
0,27 |
10,51% |
10,43% |
|
3,06 |
2,72 |
0,34 |
11,11% |
||
|
1,80 |
1,61 |
0,19 |
10,56% |
||
|
2,30 |
2,08 |
0,22 |
9,57% |
||
|
Мартовское с-55 |
2,41 |
2,20 |
0,21 |
8,71% |
9,05% |
|
3,29 |
2,99 |
0,30 |
9,12% |
||
|
2,64 |
2,41 |
0,23 |
8,71% |
||
|
3,42 |
3,09 |
0,33 |
9,65% |
||
|
Мартовское с-56 |
3,70 |
3,40 |
0,30 |
8,11% |
7,56% |
|
3,66 |
3,41 |
0,25 |
6,83% |
||
|
2,54 |
2,32 |
0,22 |
8,66% |
||
|
2,11 |
1,97 |
0,14 |
6,64% |
||
|
Мартовское с-57 |
2,23 |
2,01 |
0,22 |
9,87% |
10,17% |
|
3,80 |
3,42 |
0,38 |
10,00% |
||
|
3,10 |
2,80 |
0,30 |
9,68% |
||
|
3,50 |
3,11 |
0,39 |
11,14% |
||
|
Мартовское с-58 |
1,59 |
1,40 |
0,19 |
11,95% |
11,56% |
|
1,90 |
1,69 |
0,21 |
11,05% |
||
|
1,82 |
1,59 |
0,23 |
12,64% |
||
|
1,79 |
1,60 |
0,19 |
10,61% |
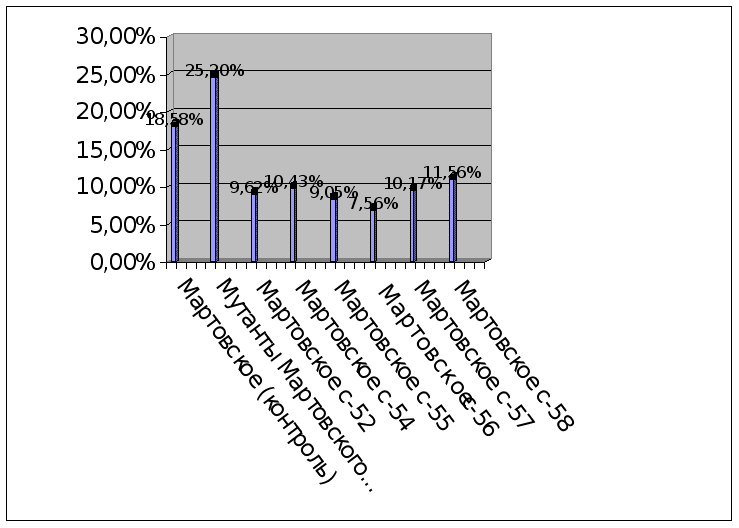
Как видно из данных, представленных в таблице, лучшими являются формы Мартовское с-56 и форма Мутанты Уэлси. Эти формы мы относим к наиболее адаптивным . Следует обратить особое внимание на эти формы и вовлечь их в дальнейший селекционный процесс для выделения доноров засухоустойчивости. Так же надо исследовать эти формы на другие хозяйственно-ценные признаки, и на стабильность по годам, сократить ювенильный период и ускорить получение первых плодов различными методами возможно в дальнейшем получить новый сорт, превосходящий районированные сорта по целому ряду хозяйственно ценных признаков.
Формы Мартовское с-52, Мартовское с-54, Мартовское с-55, Мартовское с-57 и Мартовское с-58 также показали хорошую засухоустойчивость. Их относим в группу засухоустойчивые. Следует вовлекать эти формы в селекционный процесс с целью получения исходного материала в селекции на засухоустойчивость.
Формы Декабренок о-17, Сев. Синап 2-в о-59, Спартак 1-в о-5, Спартак 2-в о-19 №16, Устойчивое в-3 о-20, Мутанты Уэлси 2 побег, Ренет Черненко о-43, хотя и показали результаты более высокие чем у контрольных форм, но не превысили существенной разницы, поэтому мы относим эти формы в группу слабозасухоустойчивые. За данными формами следует проводить регулярные наблюдения с целью выявления других хозяйственно-ценных признаков. Также можно провести многолетние наблюдения, в результате которых результат может варьировать и измениться в лучшую сторону.
Все остальные мутантные и гибридные формы показали результаты хуже, чем у контрольных форм, следовательно, они предположительно относятся в группу незасухоустойчивые. Следует проанализировать данные формы по результатам будущих лет, а так же на другие хозяйственно-ценные признаки, в течении нескольких лет, а формы не имеющие таковых исключить из селекционной школки.
4.3 Сравнительная оценка наиболее перспективных мутантных и гибридных форм с районированными контрольными сортами
Таблица 14.
Сравнение наиболее перспективных формы и гибридов с районированными контрольными сортами.
|
Сорта, мутанты и гибриды |
Средняя потеря влажности (%) |
|
Мартовское с-54 |
10,43% |
|
Мартовское с-55 |
9,05% |
|
Мартовское с-56 |
7,56% |
|
Мартовское с-57 |
10,17% |
|
Мартовское с-58 |
11,56% |
|
Мутанты Уэлси |
8,53% |
|
Антоновка обыкновенная (контроль) |
10,81% |
|
Ренет Черненко (контр) |
17,45% |
|
Уэлси 2 побег (контроль) |
10,95% |
|
Мелба (контроль) |
9,63% |
|
Мартовское (контроль) |
18,58% |
|
Красивое (контроль) |
7,91% |
|
Жигулевское(контроль) |
9,56% |
|
Устойчивое(контроль) |
9,81% |
|
Спартак (контр) |
16,89% |
|
Сев синап (контр) |
16,45% |
|
Декабренок (контроль) |
15,24% |
|
Голубь мира(контроль) |
10,00% |
Анализируя таблицу 14 можно отметить, что среди мутантных и гибридных форм яблони наиболее высокий результат по засухоустойчивости наблюдается у формы Мартовское с-56 (7,56%). Далее следуют формы: Мутанты Уелси (8,56%), Мартовское с-55 (9,05%) и другие, но из перечисленных форм только Мартовское с-56 превосходит районированные сорта, а свой контрольный сорт—Мартовское, приблизительно в 2 раза. Поэтому эта форма представляет интерес, наряду с другими, для дальнейшего изучения. Это особенно важно в связи с неравномерным распределением осадков в ЦЧР в течение вегетационного периода, что в сочетании с высокой температурой воздуха и почвы ведет к воздушной и почвенной засухе.
5. Выводы.
Наиболее засухоустойчивой из всех мутантных и гибридных форм оказалась форма Мартовское с-56.
Формы: Мартовское с-52, Мартовское с-54, Мартовское с-55, Мартовское с-57 и Мартовское с-58, Мутанты Уэлси характеризуются более высокой степенью засухоустойчивости, по сравнению с контрольными сортами.
Формы Декабренок о-17, Сев. Синап 2-в о-59, Спартак 1-в о-5, Спартак 2-в о-19 №16, Устойчивое в-3 о-20, Мутанты Уэлси 2 побег, Ренет Черненко о-43 показали засухоустойчивость на уровне с контрольными сортами.
Среди изучаемых мутантных и гибридных форм большинство растений яблони имели показатели потери свободной воды выше чем у контрольных сортов.
Результаты потери свободной воды у форм, превосходящих контроль, были в пределах 8,53-11,56%.
Наиболее слабой засухоустойчивостью обладает форма Синап о-51, у которой потеря свободной воды составила около 52%.
Результаты данных исследований возможно могут использоваться для приблизительной оценки зимостойкости форм и сортов, т. к. более высокий уровень засухоустойчивости характерен для зимостойких сортов.
6. Охрана окружающей среды.
Нужды и потребности человеческого общества растут с каждым десятилетием. Население планеты вырастает стремительно. Индустриализация сопровождается интенсивной эксплуатацией природных ресурсов и глубоким изменением природной среды. Распашка земельных массивов, использование земель под пастбища, вырубка лесов сооружение плотин и каналов, добыча полезных ископаемых, применение удобрений, эрозия почв и многие другие результаты деятельности человека приводят к значительным изменениям в природе.
Независимо от прошлых и нынешних ошибок, человек уже в прошлом пытался предотвращать ухудшение естественной среды, а новые идеи и совершенная техника вызывают у него желание и предоставляют возможности создавать или перестраивать окружающую среду таким образом, чтобы она приобретала качества, необходимые для его умственного здоровья, а также обеспечивала его нужды продуктами и удовлетворяла его разносторонние потребности (эстетика, спорт, сохранение естественных ландшафтов). Интенсификация сельского хозяйства, технический прогресс в промышленности, на транспорте привели к образованию диспропорций в окружающей среде, к деформации установившихся равновесий экосистем, к ухудшению экологической ситуации во всех сферах деятельности человека. Промышленные предприятия загрязняют атмосферу газообразными и твёрдыми выбросами, водоёмы – стоками, которые содержат большое количество вредных и сильно ядовитых веществ, от которых страдают флора и фауна. Эти вредные вещества через растения и животных поступают в пищу человека. Химизация сельскохозяйственного производства приводит к загрязнению почвы, водоёмов, воздуха, пищевых продуктов. Можно пожелать более строгое отношение к природоохранным мероприятиям со стороны хозяйства. Любой вид начинается с осмысления цели этой деятельности и путей ее достижения, что непосредственно зависит от имеющихся в наличии знаний. Человек с его все возрастающим влиянием на окружающую среду должен внимательно отнестись к изменениям в среде. Ведь он сам есть составная часть этой среды. Изменив что-либо обязательно подвергнется изменениям сам. Культура поведения и образование должны стать гарантами разумности поведения человека по отношению к природе. А образование и воспитание есть процесс длительный. И начинать его надо с самого раннего детства и неформально, а собственным примером. Вот почему экология как наука взаимоотношений человека с природой должна стать на подобающее ей место в общеобразовательном процессе детского воспитания. Только тогда можно ожидать каких-либо радикальных изменений в мышлении и действиях по отношении к системе «человек-природа».
В связи с ухудшением экологической обстановки изучается влияние некоторых техногенных факторов на почву, растения, разрабатываются приёмы реабилитации почв, загрязнённых тяжёлыми металлами и пестицидами.
На опытных участках ВНИИС им. Мичурина правильно налажено хранение и применение ядохимикатов, удобрений и нефтепродуктов. Для подготовки рабочих растворов пестицидов имеются специальные площадки. Иногда работа с пестицидами осложняется близостью жилых массивов.
Санитарно-гигиенические условия в учреждении поддерживаются на высоком уровне, что объясняется тем, что институт находится в черте г. Мичуринска.
7. Список использованной литературы.
Будаговский В.И. Зимостойкость корневой системы у карликовых и полукарликовых подвоев яблони // Изв. АН СССР. - Сер. Биология, 1954. - № 6.-С.11-25
Вигоров Л.И. Фонды высоковитаминных сортов яблонь для селекции / Труды по витаминам из природного сырья. - Уфа, 1971. - С. 134 - 140.
Генкель П.А. Методические указания по диагностике засухоустойчивости культурных растений. - М., Колос,!968. - 24с.
Генкель П.А. Физиология жаро- и засухоустойчивости растений. - М., 1982.-280с,
Генкель П.А., Окнина Е.З. Состояние покоя и морозоустойчивость плодовых растений. - М.: Наука, 1964. - 242с.
Грязев В.А. Выращивание саженцев для высокопродуктивных садов. -Ставрополь: Кавказский край, 1999. - 208с.
Гудковский В.А. Окислительный стресс плодовых культур // Научные основы устойчивого садоводства в России. - Мичуринск, 1999. - С.3-23.
Дорошенко Т.Н. Биологические основы размножения плодовых растений. - Краснодар: КТАУ, 1996. - 112 с.
Доспехов Б.А. Методика полевого опыта. - М.: Агропромиздат, 1985. -351с.
Жученко А.А. Экологическая генетика культурных растений. - Кишинев: Штиница, 1980.-587с.
Ищенко Л.А., Соловченко А.Е. Всхожесть семян яблони в условиях биотических и абиотических стрессов // Научные основы устойчивого садоводства в России. - Мичуринск, 1999. - С. 268-270.
Кичина В.В. Колоновидные яблони-М. ВСТИСП, 2002-160 с.
Кичина В.В. Методические указания по селекции яблони,-М.,1988.-63с.
Крысанов Ю.В. Культура слаборослых плодовых деревьев и ее значение в интенсификации плодоводства. - Воронеж, 1983. - 33 с.
Курсаков Г.А. Отдаленная гибридизация плодовых растений-М. Агропромиздат, 1986-112 с.
Кушниренко М.Д. Физиология водообмена и засухоустойчивости плодовых растений. - Кишинев: Штиница, 1975. - 215с.
Лебедев С.И. Физиология растений. - М.: Колос. - 1982. - 463 с.
Методы генной инженерии. Молекулярное клонирование: Пер. с англ. / Маниатис Т., Фриг Э., Сембук Дж.—М. Мир.1984—480 с.
Мичурин И.В. Сочинения. - Т. 1-4. - М.-Л.: Сельхозгиз, 1948. - 791с.
Программа и методика сортоизучения плодовых, ягодных и орехоплодных культур / Под общ. ред. Е.Н. Седова. - Орел, 1995. - 502 с.
Савельев Н.И. Генетические основы селекции яблони. - Мичуринск: Изд-во ВНИИГиСПР им.И.В. Мичурина, 1998. - 304 с.
Савельев Н.И. Яблоня/ Создание новых сортов и доноров ценных признаков на основе идентификации генов плодовых растений.—Мичуринск: Изд-во ВНИИГИСПР им. И.В. Мичурина 2002 -20 с.
Форте А.В., Д.Б. Дорохов, Н.И. Савельев-Техника RAPD анализа генотипов яблони. (Методические указания).-Мичуринск 2002 г.-48 с.
Шило А.А. Хозяйственно-биологическая характеристика индуцированных мутантов яблони. Дис. ... канд. с.-х. наук. - Орел, 1997. - 234 с.
ПРИЛОЖЕНИЯ
Приложение 1.
Погодные условия в годы исследований.
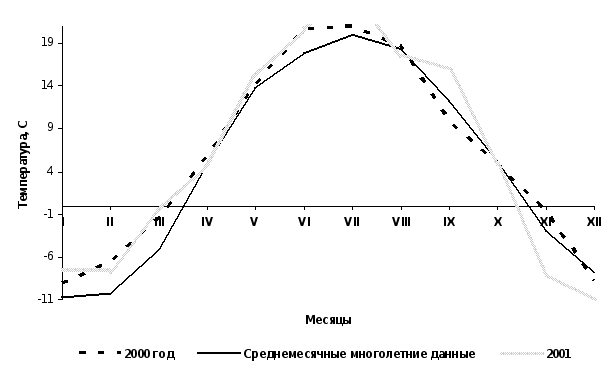
Рис.11 График среднемесячных температур
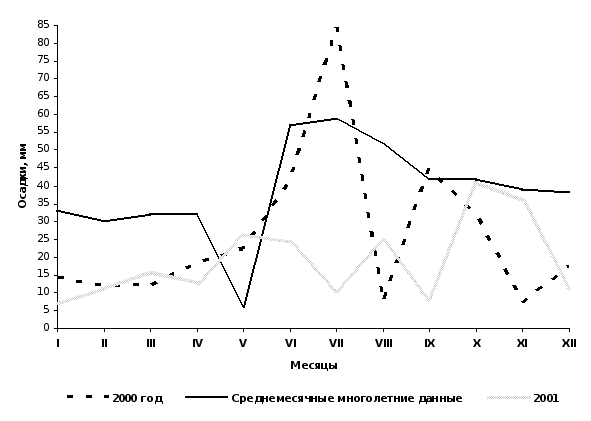
Рис.12 Среднемесячное количество осадков
Рис.13 Сравнение сорта Уэлси с его мутантными и гибридными формами.
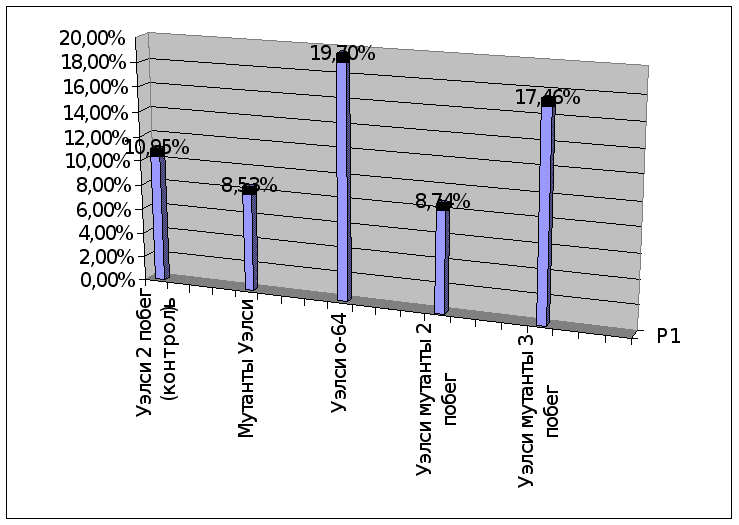
Рис. 14 Сравнение сорта Мартовское с его мутантными и гибридными формами.
Приложение 3.
Экономическая эффективность деятельности ВНИИС им. Мичурина.
ВНИИС им. И.В. Мичурина является многопрофильным учреждением по научному обеспечению садоводства, решающим комплексные отраслевые проблемы – организационно-экономические, технологические, селекционные, отраслевой и межотраслевой инфраструктуры.
РАЗРАБОТКА НОВЫХ И УСОВЕРШЕНСТВОВАНИЕ СУЩЕСТВУЮЩИХ ТЕХНОЛОГИЙ ПРОИЗВОДСТВА ЯБЛОК. На основе результатов эколого-физиологических и экономических исследований института, обобщения результатов исследований и передового зарубежного опыта применительно к конкретным эколого-экономическим условиям средней зоны РФ определены и научно обоснованы основные необходимые технологические параметры перспективных садов для средней зоны РФ, отвечающих современным требованиям хозяйствования в условиях рыночной экономики:
| назначение продукции | хранение и потребление в свежем виде |
| срок эксплуатации сада | 12-15 лет |
| срок вступления в плодоношение | 2-3-й год после посадки |
| схема размещения |
3-4 Ч 1-1,5 м |
| формировки | веретеновидные |
| подвои | слаборослые, клоновые |
| сорта | высокотоварные, устойчивые, пригодные к эксплуатации в высокоплотных садах |
| уровень рентабельности | 140-150% |
| прибыль на 1 га сада | 80000-100000 тыс. руб. |
Полученные данные показывают высокую эффективность изучаемой технологии производства подвоев с применением поверхностных органо-мульчирующих субстратов. Выход продукции с качественными параметрами, соответствующими новым стандартным требованиям, по основным подвоям составил свыше 70%. Общая продуктивность маточника за 3 года эксплуатации от 100 до 180 тыс. подвоев с гектара.
В результате 3-летних исследований институт разработал предварительный комплексный технологический регламент по производству высококачественных подвоев для высокодоходных слаборослых садов яблони, который можно применять в производственных условиях.
Институт осуществляет широкую внедренческую деятельность, как на коммерческой, так и на безвозмездной основе по освоению, как результатов своих исследований, так и передового опыта и информации отечественного и зарубежного садоводства по всем направлениям – технология производства яблок, земляники, черной смородины, груши, вишни и плодов других культур, хранение, механизация, прежде всего в своих базовых хозяйствах.
Переход на рыночную экономику, повышение уровня экстремальности окружающей среды, поставили перед учеными новые задачи, которые решаются путем разработки и осуществления комплексных программ. В этой связи в институте развернут широкий комплекс работ по фундаментальным и приоритетным прикладным направлениям исследований в садоводстве. Для надежного обоснования организационно-технологических систем по эффективному производству качественных плодов и ягод решаются следующие научно-теоретические проблемы:
• разработка эколого-физиологических основ оптимизации и стабилизации продукционных процессов при производстве ягод, получения продукции высокого качества. Конструирование рациональных моделей садов и ягодников;
• выявление и изучение механизмов устойчивости к биотическим и абиотическим стрессам плодовых и ягодных агроэкосистем и разработка методов повышения их устойчивости. Изучение антиоксидантного комплекса плодов и ягод;
• разработка системы формирования и управления качеством продукции садоводства на всех этапах - производство, хранение, транспортирование;
• эколого-экономическое обоснование оптимального размещения производства плодов и ягод, оптимизация породно-сортового состава садов и ягодников по регионам.
На основе выявленных общетеоретических закономерностей в институте разрабатываются комплексные технологические системы по производству плодов и ягод (семечковые - яблоки, груши; косточковые - вишня, черешня, слива; ягодные -смородина, земляника, крыжовник). При этом учитываются следующие задачи:
• эколого-физиологическое и технологическое изучение и подбор сортов и сорто-подвойных комбинаций;
• разработка технологий производства высококачественного посадочного материала плодовых и ягодных культур;
• разработка конструкций насаждений и. приемов их формирования, поддержания высокой продуктивности и качества плодов и ягод;
• разработка технологических регламентов по применению ростовых регуляторов, микроэлементов, фоторегуляторов и др.;
• разработка систем содержания почв в садах и ягодниках и почвенной агротехники;
• разработка систем оптимального минерального питания;
• разработка комплексных систем защиты садов и ягодников от вредителей, болезней, сорной растительности;
• разработка и создание новых машин и орудий труда для садоводства и питомниководства;
• изучение закономерностей физиолого-биохимических процессов плодов и ягод в послеуборочный период и разработка на их основе прогрессивных систем хранения и транспортирования в регулируемой атмосфере.
ОПХ ВНИИС им. И.В. Мичурина организовано в 1960 году. С 1992 года зарегистрировано как Опытно-производственное хозяйство ВНИИС им. И.В. Мичурина.
ОПХ ВНИИС является многоотраслевым хозяйством со сложившейся структурой и многоотраслевыми кадрами.
Имущество ОПХ ВНИИС состоит из 66% остаточной стоимости основных фондов, 18% незавершенного строительства, 10% запасов сырья и материалов. Общая земельная площадь составляет 2706 га, сельхозугодий—2230 га, пашня—1128 га. За 2001 год в ОПХ произведено продукции: Плодов—25313 ц. Ягод—496 ц. Зерна—8988 ц. Молока—2169 ц. Привес КРС—159 ц.
В хозяйстве используется цеховая структура, которая объединяет следующие цеха: Садоводство, Кормопроизводство, Животноводство, МТП, Мехмастерская, Автопарк, Строительный цех
В данной работе будут рассмотрены характеристики цеха садоводства.
Цех садоводства.
2001 год.
Цех садоводства является главной отраслью хозяйства. Основное направление—производство плодов и ягод, а также насаждений плодово-ягодного характера, маточников, уход и закладка многолетних насаждений.
2001 год сложился благоприятно для производства плодов и ягод. Несмотря на сильные весенние заморозки, благодаря обильному цветению получен высокий урожай плодов и ягод.
Производство яблок составило 25313 ц. При плане 10770 ц. или в 2,3 раза больше. Урожайность составила 124 ц/га. Фактическая себестоимость—0,45 руб/кг при плане 1,01 руб. это лучший показатель по Тамбовской области.
Высокорентабельной культурой является земляника. Собранно 496 ц при плане 450 ц. урожайность—71 ц/га. Себестоимость 1,66 при плане 4,2. Уровень рентабельности—672%.
В 2001 году заложено 4 га земляники и 1 га маточников земляники. В 2001 году планируется расширить плантацию как самой рентабельной культуры еще на 5 га.
Производство саженцев черной смородины оказалось также выше запланированного. Получено 60,5 тыс. шт. саженцев при плане 50 тыс. Весь посадочный материал полностью реализован осенью. Проведена затравка молодых плодоносящих садов против мышей, обрезано 25 га садов. Ремонтируется сельхозтехника, приобретена насосная станция для полива земляники, запчасти к плодовозам, трактор ДТ-75.
2000 год.
Произведено плодов –228т. Прибыль составила—321 тыс. руб. земляники произведено 51 т. урожайность—73 ц/га. Прибыль—570 тыс. руб. уровень рентабельности—713%. Заложенно 4,5 га земляники, семечкового сада—2,8 га. Увеличена площадь молодых насаждений земляники на 5 га. Малины—0,6 га.
1999 год.
Производство плодов—1308т. урожайность—68,8 ц/га. От реализации яблок полученна прибыль—20 тыс. руб. земляники собранно 42 тонны. Урожай—70 ц/га. Прибыль 142 т/руб. уровень рентабельности 322%. План: заложить 5 га земляники. Саженцев черной смородины выращено 31 тыс шт. почти все остались нереализованными. По реализации продукции на садоводство приходится 73%. Обрезано садов 25 га., произведена затравка против мышей, куплено ядохимикатов на сумму 200 тыс. рублей.
Площадь под плодовыми и ягодными культурами составляет 205 га (все данные приведены на 2000 год). Среди сортов яблони возделываются летние (15%), осенние (12%) и зимние (73%). Сады находятся в удовлетворительном состоянии средний возраст 11 лет. За последние годы заложено несколько кварталов на карликовых подвоях 62-396.
Земляника возделывается на площади в 7 га. Также имеются маточники по размножению данной культуры.
Большую площадь занимают насаждения смородины, большая часть из которых занимает черная, но также выращивается красная, белая и золотистая. В основном смородину выращивают и размножают для реализации населению и организациям посадочного материала. Имеется большая коллекция сортов.
Кроме того имеются небольшие участки груши, вишни, малины и др. плодово-ягодных культур на которых ведутся различные научные исследования.
| |||||
| |||||
| |||||
| |||||
|
Смотреть все комментарии (22)
Работы, похожие на Реферат: Сравнительная оценка засухоустойчивости мутантных и гибридных форм яблони
Меню |
||||
| Главная Рефераты Благодарности |
||||
Опрос |
||||
|
|
||||