| Министерство образования Российской Федерации
Уссурийский государственный педагогический институт
Черных Татьяна Анатольевна
Размножение папоротников в оранжерейных условиях
Дипломная работа
Научный руководитель:
к.б.н., доцент
В.Д. Фролов
Работа допущена к защите
________________________
Зав. кафедрой ботаники,
к.б.н., доцент
_____________ Прокопенко О.И.
Уссурийск, 2000
Оглавление
Введение ...........................................................................................................
Глава I. Общие представления о размножении растений............................
1.1. Вегетативное размножение......................................................
1.2. Собственно бесполое размножение......................................
1.3. Половое размножение...........................................................
Глава II. Материал и методика исследования..........................................
Глава III. Ботаническая характеристика объектов исследования..........
3.1. Общая характеристика отдела Папоротниковидные.....
3.2. Адиант венерин волос (Adiantum
capillus-veneris L.)........................................................................
3.3. Птерис критский (Pteris cretica L.)...................................
3.4. Асплениум луковиценосный (Asplenium bulbiferum
G. Forst.)......................................................................................
3.5. Листовик обыкновенный (Phyllitis
scolopendrum (L.) Newm.)...............................................................
3.6. Циртомиум серповидный (Cyrtomium falcatum (L.)
Presl.)..............................................................................................
3.7. Нефролепис возвышенный (Nephrolepis
exaltata Schott.)...............................................................................
3.8. Флебодиум золотистый (Phlebodium
aureum (L.) G. Smith).....................................................................
Глава IV. Размножение папоротников.......................................................
4.1. Вегетативное размножение папоротников.......................
4.2. Размножение папоротников спорами..............................
4. Цикл развития Циртомиума серповидного..........................
Реклама

4.4. Использование полученных материалов в
курсе ботаники средней и высшей школы.................................
Выводы.........................................................................................................
Литература...................................................................................................
Введение
Актуальность темы
. Роль растений в жизни человека и всех живых существ велика. Только зеленые растения способны преобразовывать энергию Солнца, поставляя тем самым для других организмов кислород и органические вещества. В результате деятельности растений воздух в городах очищается от газообразных примесей, сажи, пыли, выделяемых автотранспортом и промышленными предприятиями. Аналогичную роль выполняют и комнатные растения. Но, выращивая в своих квартирах цветы, человек преследует отнюдь не утилитарные цели. Зеленые растения украшают наше жилище, скрашивают тоскливую зимнюю пору, поднимают настроение, воспитывают доброту, любознательность, приучают к аккуратности.
Выращивание растений в закрытых помещениях уходит в глубокую древность. Вначале человек отдавал предпочтение лекарственным, пряноароматическим, пищевым и другим хозяйственно ценным растениям. Позднее в квартирах появились декоративнолистные и декоративноцветущие растения. Существенно пополнился ассортимент оранжерейных растений благодаря крупным географическим открытиям. Первопроходцы привозили из экспедиций новые, до сего времени неизвестные диковинные растения. В результате в наших оранжереях и комнатах появились растения из тропиков и субтропиков Америки, Африки, Азии, Австралии. Популярность комнатных растений с каждым годом продолжает расти. На это указывает и большое количество литературы, издаваемой по данному вопросу. В последние годы вышли из печати книги Н.Г. Никонова (1978), Г.К. Тавлиновой (1982, 1996), Г.А. Агеевой и К.Г. Лавровой (1992), Н.В. Вороновой (1992), Д.Б. Кудревца (1993), Д.Г. Хессайона (1995), С.О. Герасимова и др. (1997) и многие другие.
Не последнее место среди комнатных растений занимают папоротники. Это очень многоликая и разнообразная группа высших растений. В настоящее время насчитывают более 12 000 видов папоротников. Около двух третей из них растут в тропиках, а оставшаяся треть населяет умеренные области земного шара (Рейвн и др., 1990). В оранжерейной культуре папоротники появились во второй половине XVIII в. и сразу привлекли внимание садоводов. В Англии особенно они были популярны в викторианские времена (XIX в.). В настоящее время самая крупная коллекция папоротникообразных (около 1 200 таксонов) собрана в ботанических садах Кью, в Англии (Catalogue..., 1984). В России лучшая оранжерейная коллекция папоротников находится в Ботаническом саду БИНа и насчитывает свыше 400 видов (Арнаутова, 1987). По утверждению Д.Г. Хессайона (1995), более двух тысяч видов папоротников пригодны для выращивания в закрытом грунте. Некоторые виды папоротников (Птерис критский, Нефролепис возвышенный, Циртомиум серповидный, Флебодиум золотистый, Адиантум венерин волос, Платицерум и другие) можно с успехом выращивать в условиях гидрокультуры (Kühle, 1980).
Реклама

В жизни человека комнатные растения находят широкое применение. Некоторые из них используются в медицине при лечении заболеваний (Гаммерман и др.. 1963; Машковский, 1977; Муравьева, 1983; Пастушенков и др., 1990; Шретер, 1992), в косметике (Пашина, 1993), в ветеринарии (Рабинович, 1987). В качестве пищевых растений в оранжерейных условиях выращивают отдельные сорта томата, перца, огурца, баклажана (Хессайон, 1995). Но, все же, основная роль, отводимая растениям – это украшение наших квартир.
Комнатные растения, и, в частности, папоротники, могут дать интересный материал для учителя биологии. Целый ряд интересных опытов с комнатными растениями, которые можно провести в школе, изложены в книге Н.М. Верзилина (1954). На примере ряда папоротников (Асплениум, Нефролепис, Флебодиум и др.) ученики могут познакомиться с различными формами вегетативного размножения. Размножение и выращивание папоротников из спор предоставляет для учителя хорошую возможность познакомить учащихся с одним из основных явлений в растительном мире – чередованием спорофита и гаметофита в цикле развития растений. Здесь учитель может показать особенности анатомического и морфологического строения гаметофита папоротников, познакомить со строением гаметангиев – антеридиев и архегониев. Представители трех семейств папоротников (Марсилеевые, Сальвиниевые, Азолловые) освоили водную среду и являются прекрасными аквариумными растениями (Комнатный аквариум..., 1964; Жданов, 1973; Махлин, 1976, 1990; Золотницкий, 1993; Плонский, 1998). На их примере преподаватель имеет возможность иллюстрировать материал и проводить наблюдения в условиях, приближающихся к естественным.
Из вышеизложенного видно, что хорошее знание комнатных растений, и, в частности, папоротников, имеет большое значение в работе учителя, что и определило выбор настоящей темы.
Цель и задачи работы
. Целью настоящей работы является освоение методики выращивания папоротников и изучение начальных этапов их онтогенеза. Для достижения этой цели потребовалось выполнение следующих задач:
1. Познакомиться со способами размножения растений.
2. Дать ботаническую характеристику исследуемых видов папоротников.
3. Вырастить из спор исследуемые виды папоротников.
4. Изучить начальные этапы онтогенеза Циртомиума серповидного.
Структура и объем дипломной работы
. Дипломная работа состоит из введения, 4 глав, выводов и списка литературы. Основной текст изложен на 65 страницах машинописного текста и содержит 2 таблицы, 20 рисунков, в том числе 9 фотографию. Список литературы включает 67 названий.
Глава I.
Общие представления о размножении растений
Размножение – это свойство, присущее всем живым организмам и заключающееся в вопсроизведении себе подобных особей. Благодаря размножению каждый вид поддерживает свою численность и обеспечивает непрерывность и преемственность жизни (Васильев и др., 1978). В ходе эволюции возникали новые группы растений, усложнялось их внутреннее и внешнее строение. Одновременно с этим соверщенствовались и способы размножения растений. Различают три способа размножения – вегетативное, собственно бесполое и половое.
1.1. Вегетативное размножение.
Вегетативное размножение – это увеличение числа самостоятельных особей за счет отделения от материнского растения его неспециализированных или специализзированных частей, способных к самостоятельному существованию и развитию. Способность к вегетативному размножению присуща многим растениям. Ею обладают более 90% видов травянистых растений широколиственных лесов Европы (Федоров и др., 1962). Особенно широко вегетативное размножение выражено у растений, произрастающих в условиях, наблагоприятных для семенного размножения, таких как короткий вегетационный период, сильное затенение, избыточное увлажнение, отсутствие опылителей и т.п. Благодаря широкому распространению в природе, вегетативное размножение играет большую роль в сапоподдержании популяции того или иного вида. По словам Р.Е. Левиной (1964), способность к обильному вегетативному размножению имеет огромное приспособительное значение и часто обеспечивает доминирующее положение вида в фитоценозе. По сравнению с половым размножением, вегетативное имеет целый ряд преимуществ. Оно просто, надежно и к тому же часто обеспечивает быстрое расселение вида на территории (Даддингтон, 1972). Выживание вегетативного потомства у одного и того же вида выше, чем выживание семенного (Ценопопуляции растений, 1988).
В ходе эволюции появились самые разнообразные формы вегетативного размножения: размножение естественными отводками, размножение корневвыми отпрысками, размножение специализированными частями растений – корневищами, клубнями, луковицами, наземными и подземными столонами, выводковыми почками.
Естественными отводками
размножаются многие виды деревьев и кустарников: Пихта, Ель, Калина, Лещина и другие. На ветвях этих растений в месте контакта с почвой формируются придаточные корни, что способствует возникновению новых побегов и самостоятельных особей.
Корневыми отпрысками
размножаются Тополь серебристый, некоторые виды Сливы, Вишни, ряд сорных многолетних растений – Осот, Бодяк и другие. На корнях этих растений образуются придаточные почки, дающие начало побегам, которые часто называются корневыми отпрысками.
Корневище
– это подземный или ползучий по поверхности почвы побег, выполняющий функции отложения запасов, возобновления и вегетативного размножения (Серебряков, Серебрякова, 1965, цит. по: Таршис, 1975). Размножение с помощью корневищ широко распространено в природе. Оно характерно для многих видов папоротников и цветковых растений.
Луковица
представляет собой видоизмененный укороченный побег. Она состоит из крайне редуцированного стебля (донца), на котором сидят сочные видоизмененные листья. В пазухах этих листьев (чешуй) формируются “детки” – дочерние луковицы, которые отделяются от материнской в первый год или остаются связанными с ней до гибели последней. Размножение с помощью луковиц особенно широко распространено среди однодольных растений.
Распространенным видоизменением побега являются клубни
. Клубень – это укороченный побег, служащий для запаса питательных веществ и вегетативного размножения. Из дальневосточных растений размножение клубнями характерно для видов рода Хохлатка, Сердечника трехнадрезанного, Седмичника, Хвоща полевого и ряда других видов.
Специализированными органами вегетативного размножения являются столоны
. Это плагиотропные подземные или надземные побеги, несущие на конце почку, клубень или луковицу. В отличие от корневищ, столоны не являются запасающими органами. Они недолговечны и служат целям вегетативного размножения (Федоров и др., 1962). Размножение с помощью столонов свойственно Землянике, Лютику ползучему, Лапчатке гусиной, Адоксе и другим растениям.
1.2. Собственно бесполое размножение.
Бесполое размножение заключается в том, что происходит при помощи специализированных клеток – спор. В отличие от гамет, споры непосредственно, т.е. без слияния с другими клетками, прорастают в новое растение.
У низших растений споры часто снабжены жгутиками. Они называются зооспорами
. Если споры низших растений лишены жгутиков, они называются апланоспорами
. У высших растений споры никогда не имеют жгутиков.
Формируются споры в специальных вместилищах – спорангиях
. У низших растений спорангии одноклеточные, у высших – многоклеточные. Внутри спорангия формируется спорогенная ткань. Клетки этой ткани делятся путем митоза или мейоза и образуют споры.
По происхождению споры делятся на митоспоры
и мейоспоры
. Митоспоры образуются в результате митотического деления. Прорастают они во взрослую особь. Встречается подобный тип спор у некоторых низших растений. Мейоспоры образуются в результате редукционного деления. Прорастая, они развиваются в гаплоидные образования – гаметофиты. Характерен подобный тип спор для всех высших и некоторых низших растений.
Различают равноспоровые и разноспоровые растения. У равноспоровых растений все споры одного типа. При прорастании они формируют обоеполый гаметофит.
Разноспоровые растения имеют споры двух типов: микроспоры и мегаспоры. Микроспоры, прорастая, дают мужской гаметофит, а мегаспоры – женский гаметофит.
1.3. Половое размножение.
Это самый прогрессивный способ размножения. Он заключается в слиянии двух специализированных клеток – гамет, и образовании зиготы, из которой развивается новая особь. При половом размножении происходит соединение наследственных признаков двух организмов (материнского и отцовского). В результате полученное потомство более разнообразно генетически, что способствует более успешному выживанию вида.
Половое размножение встречается у всех групп растений. Различают две основные формы полового процесса: соматогамию и гаметогамию. Соматогамия – это половой процесс, который осуществляется путем слияния содержимого двух соматических клеток. Гаметогамия – это половой процесс, который происходит путем слияния гамет. В зависимости от относительной величины гамет различают три типа гаметогамии – изогамия, гетерогамия и оогамия.
При оогамном половом процессе различают женские и мужские гаметы. Женская гамета называется яйцеклеткой. Мужские гаметы бывают двух видов: сперматозоиды – половые клетки со жгутиками, и спермии – половые клетки без жгутиков.
Формируются гаметы в специальных органах – гаметангиях. У низших растений гаметангии, как правило, одноклеточные. У высших растений они многоклеточные. Мужские гаметангии носят название антеридиев. Женские гаметангии низших растений называются оогониями, у высших – архегониями. Архегоний – это многоклеточное образование, имеющее колбовидную форму. Состоит из расширенного брюшка и вытянутой шейки. По каналу шейки сперматозоиды попадают в брюшко, где сливаются с яйцеклеткой. Наиболее хорошо развит архегоний у мхов. В дальнейшем наблюдается его редукция.
Глава II.
Материал и методика исследований
Работа по выращиванию папоротников в оранжерейных условиях проводилась в период 1998-2000 гг. Объектом исследования послужили 7 видов тропических и субтропических папоротников: Адиант венерин волос (Adiantum capillus-veneris L.), Птерис критский (Pteris cretica L.), Костенец (Асплениум) луковиценосный (Asplenium bulbiferum G. Forst.), Листовик обыкновенный (Phyllitis scolopendrium (L.) Newm.), Циртомиум серповидный (Cyrtomium falcatum (L.) Presl.), Нефролепис возвышенный (Nephrolepis exaltata Schott.), Флебодиум (Многоножка) золотистый (Phlebodium (Polypodium) aureum (L.) G. Smith).
При выращивании папоротников из спор мы руководствовались методическими рекомендациями ряда авторов (Парамонова, 1956; Чичагова, 1958; Котухов, 1967; Арнаутова, 1987; Мак-Миллан Броуз, 1987; Хессайон, 1995). Для сбора спор срезали фертильную часть листа папоротника и помещали его в пакет из кальки. В течение нескольких дней пакет держали в теплом месте до раксрытия спорангиев и высыпания спор. Споры тонким слоем высевали на земляную смесь в чашки Петри. Земляную смесь готовили из 2 частей простерилизованной листовой земли и 1 части промытого речного песка. Закрытые чашки Петри ставили в затененное место. В чашках постоянно поддерживали высокий уровень влажности. После появления гаметофитов почву время от времени обильно поливали, что необходимо для успешного протекания полового процесса. После окончания полового процесса наблюдается появление проростков спорофита. Для закаливания и адаптации растений к более засушливым условиям в этот период стекло с чашки Петри ежедневно на некоторое время убирали. После появления 3-4-го листа у спорофита проводили пикировку растений. Горшки с распикированными растениями накрывали стеклом. Первые дни растения только опрыскивали, снимая стекло на 2-3 ч в день. После укоренения стекло снимали и полив проводили по мере необходимости.
Для изучения начальных этапов онтогенеза Циртомиума серповидного споры высевали в чашки Петри на жидкую среду Кнопа. Для этого в чашку Петри помещали фильтровальную бумагу, увлажненную раствором Кнопа следующего состава: Ca(NО3
)2
- 1 г, MgSO4
- 0,25 г, KСl - 0,125 г, KH2
PO4
- 0,25 г на 1 л воды.
Собранные споры тонким слоем распределяли на фильтровальной бумаге. Наблюдения за прорастанием спор, формированием гаметофитов и проростков проводили с помощью стереоскопического микроскопа МБС-9 и микроскопа “Биолам”. Наблюдения проводились ежедневно. Полученные данные фиксировались в виде рисунков.
При изучении развития гаметофита Циртомиума мы, следуя другим авторам (Nayar, Kaur, 1971; Арнаутова, 1988, 1989; Храпко, 1990а, 1990б, 1990в, 1991а, 1991б, 1996; Шорина, Ершова, 1990 и др.), выделяли 3 этапа: 1 - прорастание споры; 2 - образование нити гаметофита, 3 - формирование пластинки гаметофита. При изучении развития спорофита Циртомиума серповидного мы придерживались периодизации онтогенеза спорофитов папоротников, предложенной Г.П. Серой с соавторами (1980). В онтогенезе спорофита они выделяют 3 периода - препродуктивный, репродуктивный и пострепродуктивный. Препродуктивный период включает следующие возрастные состояния: проростки, юные спорофиты и молодые спорофиты. В пределах репродуктивного периода выделяют два возрастных состояния: зрелого и стареющего спорофита. В пострепродуктивном периоде авторы выделяют лишь одно возрастное состояние старого спорофита.
Морфологическое описание спорофитов и гаметофитов папоротников проводили на натуральных объектах, при этом использовали терминологию, предложенную А.А. Федоровым с соавторами (1956, 1962). Измерение мелких объектов проводилось с помощью мерной лупы. Данные количественных показателей обрабатывались методами вариационной статистики (Зайцев, 1990). При этом определяли такие показатели, как: среднее значение признака (М=SX/N), ошибка среднего значения (m=s/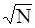 ), среднее квадратическое отклонение (s= ), среднее квадратическое отклонение (s=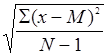 ), коэффициент вариации (V=s·100/M). ), коэффициент вариации (V=s·100/M).
Для оценки амплитуды изменчивости использовали унифицированную шкалу уровней изменчивости, разработанную С.А. Мамаевым (1969): V меньше 7% - очень низкий, V = 8-12% - низкий, V = 13-20% - средний, V = 21-30% - повышенный, V = 31-40% - высокий и V больше 40% - очень высокий уровень изменчивости.
Глава III.
Ботаническая характеристика объектов исследования
3.1. Общая характеристика отдела Папоротниковидные (Polypodiophyta).
Папоротниковидные относятся к числу наиболее древних групп высших растений. По своей древности они уступают только Риниофитам и Плауновидным и имеют приблизительно один возраст с Хвощевидными. Риниофиты – это вымершая группа растений; Плауновидные и Хвощевидные играют в современном растительном покрове Земли очень скромную роль; и лишь Папоротниковидные в настоящее время продолжают процветать. Современные ботаники насчитывают около 300 родов и более 10 000 видов папоротников (Тахтаджян, 1978а; Рейвн и др., 1990).
Папоротники распространены очень широко, фактически по всему земному шару, и встречаются в самых различных местообитаниях, начиная с пустынь и заканчивая болотами, озерами, рисовыми полями и солоноватыми водами. Но наибольшее их разнообразие наблюдается в тропических лесах (Ярошенко, 1975; Леме, 1976; Второв, Дроздов, 1978). Еще во второй половине XIX века Альфред Уоллес указывал на большое количество и разнообразие жизненных форм папоротников в экваториальной зоне. Так, только на одной из вулканических вершин на острове Яве насчитывается около 300 видов папоротников (Уоллес, 1975). Обилием папоротников удивляет флора Новой Зеландии, многие виды которых являются здесь эндемиками. Видовой эндемизм папоротников Новозеландской флористической области составляет 40%, а у родов Диксония (Dicksonia) и Циатея (Cyathea) – примерно 75% (Тахтаджян, 1978б). Очень часто встречаются здесь древовидные папоротники, которые придают лесу особый облик. Это прежде всего Циатея серебристая (C. dealbata), которая считается национальным растением Новой Зеландии (Петров, 1981). Богата видами птеридофлора Новогвинейской флористической области. Из 1000 видов папоротников, встречающихся здесь, около 600 являются эндемиками (Второв, Дроздов, 1978).
Полный жизненный цикл папоротников складывается из двух фаз – фазы гаметофита и фазы спорофита. Когда говорят о папоротниках, то имеют в виду прежде всего спорофит. Как и у большинства высших растений (за исключением Моховидных), спорофит папоротников является доминирующей фазой в их жизненном цикле.
Спорофит папоротников имеет корни, за исключением небольшого числа видов, что является результатом редукции. Корни у папоротников только придаточные, т.е. главный корень не получает здесь дальнейшего развития и вскоре отмирает. Впервые стержневая корневая система появляется лишь у голосеменных растений (Комаров, 1936).
Стебель у большинства современных папоротников представлен подземным корневищем, иногда мощным, иногда тонким, шнуровидным. У тропических древовидных форм он надземный, прямостоячий, одет массой отмерших листьев. Анатомическое строение стебля может быть весьма сложным и разнообразным. Проводящая система представлена протостелой, сифоностелой или диктиостелой. В эволюционном отношении диктиостела наиболее продвинутый тип и поэтому она характерна для наиболее прогрессивных семейств (Тахтаджян, 1978а).
Листья папоротников, как и большинства других высших растений, возникли в результате уплощения и срастания дихотомически ветвящихся теломов Риниофитов (Серебряков, 1952; Мейер, 1958; Ярошенко, 1963; Тахтаджян, 1978а; Письяукова, 1980). Убедительным доказательством веточной природы листьев является характерный для них верхушечный и длительный (иногда почти неограниченный) рост, большие размеры и сложно рассеченная листовая пластинка, крупные и сложные листовые следы и наличие листовых прорывов (лакун) в стеле. По морфологическому и анатомическому строению листья папоротников весьма разнообразны. Их размеры колеблются от нескольких миллиметров до 30 м в длину. В большинстве случаев листья папоротников совмещают функции фотосинтеза и спороношения. У некоторых папоротников (Страусник – Struthiopteris, Оноклея – Onoclea) листья дифференцированы на стерильные (фотосинтезирующие) и фертильные (несущие спорангии). Чаще всего листья папоротников перистые, хотя имеются виды (Пиррозия длинночерешковая – Pyrrosia petiolasa, Щиточешуйник уссурийский – Pleopteris ussuriensis, Кривокучник сибирский – Camptosorus sibiricus, Листовик обыкновенный – Phyllitis scolopendrium), листья которых цельные. Пластинка перистого листа имеет стержень (рахис). Если лист однажды перистый, то сегменты листа называются перьями. Сегменты второго, третьего и других порядков называются перышками.
Жилкование у примитивных форм папоротников дихотомическое. У более продвинутых форм дихотомическое жилкование сменяется сначала перистым, а затем сетчатым.
В репродуктивной фазе развития спорофита начинается его спороношение. Спороношение начинается с закладки инициальных клеток или групп клеток, из которых развивается спорангий – орган бесполого размножения растений. У самых примитивных из современных папоротников из семейств Ужовниковых (Ophioglossaceae) и Мараттиевых (Marattiaceae) спорангии возникают из групп клеток эпидермы и глубже лежащих слоев клеток. Их спорангии имеют многослойную стенку и не несут специальных приспособлений к раскрыванию. У более продвинутых папоротников спорангий развивается из одной клетки и имеет однослойную стенку. Постепенно вырабатываются приспособления к активному раскрыванию спорангиев в виде механического кольца той или иной формы. У древнейших папоротников спорангии сидели на верхушках конечных веточек дихотомического разветвленного листа, т.е. имели верхушечное (терминальное) расположение. Верхушечное расположение спорангиев в ходе эволюции листа переходит в краевое (маргинальное). Подобное расположение спорангиев наблюдается у многих современных папоротников. Значительным шагом вперед в приспособительной эволюции папоротников был переход спорангиев из краевого расположения в поверхностное (ламинальное). При этом спорангии перемещались на нижнюю (абаксиальную) сторону листовой пластинки. У исследуемых нами папоротников маргинальное расположение сорусов характерно для Птериса (Pteris critica), Асплениума (Asplenium bulbiferum) и Адиантума (Adiantum capillus-veneris), для других видов – Нефролеписа (Nephrolepis exaltata), Циртомиума (Cyrtomium falcatum), Флебодиума (Phlebodium aureum) и Листовика (Phyllitis scolopendrium) характерно ламинальное расположение сорусов.
В ходе эволюции в расположении спорангиев произошло и другое изменение. Сначала спорангии располагались одиночно. Подобное мы наблюдаем у представителей семейства Схизейных (Schizeaceae). Позднее, в ходе филогенеза, спорангии группировались в отдельные спорангиальные кучки – сорусы. Образование сорусов способствовало более эффективному снабжению спорангиев питательными веществами. Различают сорусы простые и смешанные. В простых сорусах, в отличие от смешанных, развитие и созревание спорангиев происходит одновременно. Преимущество смешанных сорусов заключается прежде всего в том, что развитие спорангиев в сорусе растягивается во времени. Поэтому резкое ухудшение условий произрастания не может привести к гибели одновременно все споры.
В процессе эволюции у спорангиев появились защитные приспособления. Сорусы у большинства папоротников покрыты различными типами индузия (покрывальца). Индузий – это вырост на поверхности листа папоротника, прикрывающий сорусы. Развивается преимущественно из эпидермиса листа, плаценты, или образован завернутым краем листа (Словарь..., 1984). В последнем случае, когда сорусы защищены загнутым на нижнюю сторону краем листовой пластинки, говорят о ложном покрывальце. Подобное покрывальце из исследуемых нами папоротников имеют Адиант венерин волос и Птерис критский.
При созревании спорангии папоротников раскрываются и споры рассеиваются. Приспособление, способствующее раскрыванию спорангиев, называют кольцом. Кольцо состоит из наружных клеток спорангия папоротника, имеющих сильно утолщенные радиальные стенки. Клетки кольца, разрастаясь в высоту, выступают над поверхностью спорангия в виде грубня. При дозревании спорангия кольцо от напряжения разрывается и способствует разбрасыванию спор. Различают три основных типа расположения кольца у спорангиев папоротников: поперечное, косое и продольное. Продольное кольцо возникало в процессе эволюции независимо несколько раз и в настоящее время является наиболее распространенным типом (Тахтаджян, 1978а).
Споры папоротников, как и других высших растений, образуются из клеток спорогенной ткани в результате редукционного деления. Подобные споры называются мейоспорами. Они состоят из гаплоидного ядра, цитоплазмы и двух оболочек – экзины и интины. Большинство современных папоротников – растения равноспоровые. Они образуют один вид спор, из которых вырастает обоеполый гаметофит. Представители трех семейств Папоротниковидных – Марсилеевые (Marsileaceae), Сальвиниевые (Salviniaceae), Азолловые (Azollaceae) – являются разноспоровыми растениями. Они образуют два типа спор – микроспоры и мегаспоры, которые при прорастании формируют соответственно мужской и женский гаметофиты. Все виды папоротников, с которыми работали мы, являются равноспоровыми. Попав в благоприятные условия, споры прорастают. Для прорастания спор необходимы влажность почвы и воздуха, тепло (от 15 до 30оС), а также свет. Споры большинства папоротников не прорастают в темноте.
Гаметофиты папоротников в целом отличаются значительным разнообразием. Они могут быть нитевидными, лентовидными, цилиндрическими, могут вести как подземный, так и наземный образ жизни, быть долговечными либо эфемерными. Но наиболее широко распространен сердцевидный гаметофит. Он плоский, нежный, быстро развивающийся и недолговечный. Единственная точка роста такого гаметофита расположена на верхушке между двумя его боковыми лопастями. Гаметофит состоит из одного слоя клеток, за исключением центральной, более массивной подушкообразной части. К почве гаметофит прикрепляется ризоидами, которые образуются на центральной подушкообразной возвышенности. На нижней, брюшной стороне подушки развиваются архегонии. Антеридии (обычно развиваются раньше) разбросаны на нижней поверхности или по краям базальной части подушки, а также на боковых лопастях. У папоротников можно наблюдать явление протандрии – когда антеридии развиваются раньше архегониев, и явление протогинии – когда раньше развиваются архегонии. Это способствует перекрестному межгаметофитному оплодотворению. Оплодотворение происходит только при наличии влаги. Сперматозоиды движутся в сторону архегония в результате хемотаксиса, вызванного выделением из архегония определенных химических веществ. На гаметофите может оплодотвориться несколько яйцеклеток, но обычно только одна из них завершает свое развитие и дает начало зародышу.
В заключение несколько слов нужно сказать о наших дикорастущих папоротниках. Птеридофлора Дальнего Востока освещена в работах, посвященных флоре региона и его отдельных районов (Комаров, Клобукова-Алисова, 1925, 1931; Воробьев и др., 1966; Воробьев, 1982; Ворошилов, 1982; Цвелев, 1991; Пешеходько, Храпко, 1994). На Дальнем Востоке произрастает 84 вида папоротников, относящихся к 17 семействам и 40 родам. Наиболее крупными являются семейства Кочедыжниковые (Athyriaceae) и Щитовниковые (Aspidiaceae). Папоротники играют заметную роль в сложении фитоценозов российского Дальнего Востока. Наиболее обильны и разнообразны они в травяном покрове хвойных и хвойно-широколиственных лесов, где нередко могут являться эдификаторами и доминантами.
3.2. Адиант венерин-волос (Adiantum capillus-veneris L.).
Относится к семейству Адиантовые (Adiantaceae), порядку Схизейных (Schizeales). Адиантовые – преимущественно тропическое семейство, представители которого являются лесными или петрофильными растениями. Включает 1 род и около 300 видов.
Адиант венерин-волос (Рис. 1) – наиболее популярный в культуре вид рода. Встречается в тропических и субтропических районах Европы, Азии, Африки. Растет в Крыму, на Кавказе, в горах Средней Азии. В своем распространении достигает умеренной зоны, встречаясь в Англии (Хессайон, 1995) и Ирландии (Вальтер, 1974). Растет на скалах, у водопадов, по каменистым берегам горных ручьев.
Адиант венерин-волос – это многолетнее травянистое растение с ползучим корневищем, покрытым темными чешуйками. Черешки листьев тонкие, темно-коричневые. Листья до 50 см длины, тонкие, нежные, дважды-трижды перистые с клиновидными сегментами, в свою очередь сидящих на волосовидных черешочках. Характерной особенностью листьев является их водоотталкивающее свойство. Капли воды стекают с них, на смачивая поверхность. Отсюда и название рода, Adiantum, от греч. а – отрицание, diaino – смачивать. Сорусы маргинальные, лишены настоящего покрывальца. Они л ежат вдоль концов жилок, идущих до самого края листового сегмента.Именно эта, покрытая сорусами, часть сегмента отгибается вниз, сегмент в верхней
Рис. 1. Адиантум венерин-волос (Adianthum capillus-veneris L.).
Рис. 2. Расположение сорусов у Адиантума
1 - часть листа; 2 - сегмент с сорусами; 3 - край сегмента с сорусами.
части как бы складывается пополам, и сорусы оказываются внутри особых карманчиков (Рис. 2). Темные сорусы просвечивают сквозь тонкую ткань листа, обрамляя его темным пунктиром.
Многие виды Адиантума широко вошли в культуру. В России большой интерес к ним возник еще в конце прошлого века. Журнал “Русский вестник садоводства” за 1890 г. рекомендовал виды Адиантума как одних из лучших растений для разведения в теплицах и комнатах (Гладкова, 1978а). Адиантумы легко размножаются спорами, часто дают самосев. Можно размножать делением куста. Оптимальная температура для выращивания – 16-20оС. Летом растения следует опрыскивать и притенять от яркого солнца (Агеева, Лаврова, 1992; Тавлинова, 1997). Растения весьма чувствительны к низкой влажности воздуха. Необходимо не допускать пересыхания земляного кома, что может привести к гибели надземной массы (Герасимов и др., 1997).
3.3. Птерис критский (Pteris cretica L.).
Русский синоним: крылатый папоротник. Относится к семейству Птерисовых (Pteriaceae), порядку Схизейных (Schizeales). Род насчитывает около 280 видов. Это наземные папоротники, распространенные в тропических и субтропических странах обоих полушарий, главным образом во влажных областях. Некоторые достигают в своем распространении Средиземноморья, Японии, США, а в южном полушарии – Новой Зеландии, Тасмании и Южной Африки. Растут на равнинах и невысоко в горах, в тени лесов или в условиях легкого затенения – на лесных опушках вблизи воды, в заболоченных местах, часто у скал ручьев и по берегам рек, иногда встречаются и в сухих местообитаниях. Наиболее часто из представителей рода в оранжерейной и комнатной культуре встречается Птерис критский (Рис. 3). Его ареал простирается от Средиземноморья до Китая и Японии. Растет он также в Африке, на Мадагаскаре, по склонам
Рис. 3. Птерис критский (Pteris cretica L.).
в горных лесах Закавказья, в Крыму. Широко распространен в тропиках и умеренных областях Южного полушария.
Птерис критский – многолетнее травянистое растение с ползучим корневищем. Листья кожистые, продолговатые, до 55 см длины, на длинных, голых черешках, оливково-зеленые, состоят из сегментов. Есть стерильные и фертильные сегменты. Первые – ланцетные, остропильчатые, вторые – более узкие, цельнокрайние. Сорус (ценосорус) маргинальный, расположен по краю листа и прикрыт ложным линейным покрывальцем, образованным из отвернутого края листа (Рис. 4). Ценосорус лежит на краевой жилке. Спорангии ценосоруса перемешаны с большим количеством парафиз. Спорангии имеют ножку из 3 рядов клеток и вертикальное кольцо для вскрытия.
Птерис критский – облигатный апомикт, что способствует легкой приспособляемости его к необычным условиям существования (Гладкова, 1978б). У папоротников может наблюдаться явление апоспории, когда гаметофит образуется из вегетативных клеток спорофита, а также и явление апогамии, когда спорофит образуется не из зиготы, а из соматических клеток гаметофита. В последнем случае спорогенез проходит без мейоза, и все деления в жизненном цикле митотические.
Большинство видов Птериса несложны для выращивания. Самые популярные из растений этой группы – разновидности Птериса критского. Наиболее распространены две разновидности: бело-пестрая с белыми полосками по всей длине листа; гребенчатая – отличается тем, что концы некоторых сегментов листа имеют бахромчатую рассеченность.
Птерис – теневыносливое и влаголюбивое растение. Температура для успешного развития в пределах 15-25оС. Почвы легкие, питательные.
3.4. Асплениум луковиценосный (Asplenium bulbiferum G. Forst.).
Русские синонимы: костенец, материнский папоротник. Относится к
Рис. 4. Ценосорус Птериса критского
1 - часть листа; 2 - участок сегмента с ценосорусом.
семейству Асплениевых (Aspleniaceae), порядку Циатейных (Cyatheales). В семейство входит около 4 000 видов наземных и эпифитных папоротников, населяющих преимущественно влажные местообитания. Характерными признаками семейства являются: диктиостелические корневища, покрытые чешуями, сорусы, снабженные настоящими покрывальцами.
Центральное место в семействе занимает род Асплениум, включающий около 700 видов. Представители рода распространены практически во всех областях земного шара, но наибольшее их разнообразие наблюдается в тропиках. Род характеризуется удлиненными, линейными или линейно-продолговатыми сорусами, расположенными обычно с одной стороны боковых жилок косо по отношению к средней жилке и прикрытыми узким покрывальцем. В культуре наиболее часто встречаются Асплениум гнездовой (A. nidus) и Асплениум луковиценосный.
Асплениум луковиценосный (Рис. 5) – широко распространенный в Новой Зеландии наземный, вечнозеленый папоротник (Петров, 1981). Это многолетнее травянистое растение с чешуйчатым цилиндрическим корневищем. Листья 80-90 см длины, темно-зеленые, трижды перистые. Сорусы маргинальные (Рис. 6). На верхней стороне листовой пластинки образуются выводковые (придаточные) почки, которые прорастают еще прикрепленными к материнскому растению (Рис. 7, 14). Эта интересная и практически важная особенность характерна для многих экзотических видов Асплениума (Гладкова, 1978в).
Папоротник нетребователен к почве, теневынослив, оптимальная температура – 16-18 оС . С марта по октябрь Асплениум опрыскивают и обильно поливают, не допуская высыхания земляного кома. В зимнее время поливают реже (Тавлинова, 1977). Хорошо размножается вегетативно и спорами.
Рис. 5. Асплениум луковиценосный (Asplenium bulbiferum G. Forst.).
Рис. 6. Расположение сорусов у Асплениума луковиценосного
Рис. 7. Вегетативное размножение Асплениума луковиценосного
3.5. Листовик обыкновенный (Phyllitis scolopendrium (L.) Newm.
Русский синоним: папоротник олений язык. Относится к семейству Асплениевых (Aspleniaceae), порядку Циатейных (Cyatheales).
Листовик обыкновенный (Рис. 8) распространен в Европе, Северной Америке, Восточной и Юго-Восточной Азии. Растет на влажных, затененных скалах, на сырых почвах, в лесах. Встречается на черноморском побережье Кавказа на высоте 1000 м над ур. моря (Карри-Линдал, 1981).
Растение до 50 см высоты, с коротким и толстым корневищем, несущим розетку вай. Листовая пластинка цельная, с немного волнистым краем, ланцетная, до 50 см длины и 3-6 см ширины, с сердцевидным основанием. Сорусы линейные, 5-30 мм длины, прямые, расположенные парами по обеим сторонам от средней жилки (Рис. 9). Индузии перепончатые. боковые, прикрывающие сорусы в начале их развития.
Декоративен. Особенно ценятся уродливые форму с многократно вильчато разделенной верхушкой листа, а также формы, образующие почки на листьях (Гладкова, 1978в).
На юге Дальнего Востока произрастает близкородственный вид – Листовик японский (Ph. japonica), который систематики нередко рассматривают в ранге подвида. Внешне не отличается от Ph. scolopendrium, но имеет иную скульптуру спор и другое число хромосом (Цвелев, 1991).
3.6. Циртомиум серповидный (Cyrtomium falcatum (L.) Presl.).
Латинский синоним: Aspidium falcatum. Относится к семейству Щитовниковых (Aspidiaceae), порядку Циатейных (Cyatheales).
Распространен в тропиках и субтропиках Японии, Кореи, Китая, Индии, Тайваня, Малайзии, Южной Африки, на Гавайах. Растет по склонам в прибрежной полосе. Циртомиум серповидный (Рис. 10)-многолетнее травянистое растение, с корневищем, от которого отходит пучок вай. Листья перистые, 35-50 см длины, 12-15 см ширины, блестящие, темно-зеленые.
Рис. 8. Листовик обыкновенный (Phyllitis scolopendrium (L.) Newm.
Рис. 9. Расположение сорусов у Листовика обыкновенного
1 - нижняя сторона листа с сорусами; 2 - часть листа с сорусами.
Рис. 10. Циртомиум серповидный (Cyrtomium falcatum (L.) Presl.).
Листочки очередные, саблевидные, загнутые кверху, с зубчатым краем. Сорусы округлые, латеральные, с нижней стороны пластинки, прикрыты покрывальцем (Рис. 11). Покрывальце округлое.
Папоротник переносит сухой воздух и сквозняки. Оптимальная температура 15-25 оС. Почвенная смесь: листовая земля, торф, перегной, песок в равных количествах. Размножается спорами.
3.7. Нефролепис возвышенный (Nephrolepis exaltata Schott.).
Русский синоним: мечевидный папоротник. Относится к семейству Даваллиевых (Davalliaceae), порядку Циатейных (Cyatheales).
Род включает 30 видов, распространенных в тропиках. За пределами тропической зоны встречается в Японии и Новой Зеландии. Это обитатели открытых мест. Нефролепис возвышенный – одно из первых высших растений, поселяющихся после извержения на склонах вулканов.
Нефролепис возвышенный (Рис. 12) – многолетнее травянистое растение. Сорусы латеральные, округлые, расположены на концах жилок. Покрывальце округлое. Спорангии на ножках, разновозрастные в пределах одного соруса.
Листья до 1 м длины, однажды перистые, мечевидной формы, вначале прямостоячие, затем веерно-наклоненные. Сегменты листа зубчатые. У садовых форм Нефролеписа может наблюдаться гетерофиллия, т.е. встречаются формы с разнообразно рассеченными листьями.
Около ста лет назад в Бостоне (США) была выведена форма с красиво поникающими листьями, названная N. exaltata f. bostoniensis. Эта форма быстро приобрела популярность по обе стороны Атлантики, так что в наши дни имеются уже десятки сортов бостонского папоротника – с волнистыми листочками, с закрученными краями листочков, с трижды и четырежды перисторассеченными листьями, так что все растение выглядит кружевным (Хессайон, 1995).
Рис. 11. Расположение сорусов у Циртомиума серповидного
1 - нижняя сторона листа с сорусами; 2 - участок сегмента с сорусами
Рис. 12. Нефролепис возвышенный (Nephrolepis exaltata Schott.).
Нефролепис легко размножается вегетативно (Рис. 14). На его корневище образуются наземные, безлистные, покрытые чешуйками укореняющиеся побеги (столоны). В течение одного года одно растение может образовать свыше ста новых (Сурова, 1978).
Оптимальная температура 15-20 оС. Светолюбив, хотя хорошо растет и в затенении. Летом желательно опрыскивание. Выращивают в горшках с хорошей аэрацией, в смеси листовой земли, песка и торфа. Размножается вегетативно и спорами.
3.8. Флебодиум золотистый (Phlebodium aureum (L.) G. Smith).
Синоним: Polypodium aureum. Относится к семейству Многоножковых (Polypodiaceae), порядку Полиподиевых (Polypodiales).
Флебодиум золотистый (Рис. 13) распространен в тропиках и субтропиках Америки. Это многолетнее травянистое растение с толстым, ползучим по поверхности почвы, ветвящимся корневищем, покрытым чешуями. Листья до 1 м длины, глубокораздельные, отходят от корневища рядами. Молодые листья покрыты восковым налетом, что обеспечивает водоотталкивающие свойства листа. Сорусы ламинальные, линейные, расположены посредине сегментов с нижней стороны листа. Размножается спорами и вегетативно участками корневища (Рис. 14).
Рис. 13. Флебодиум золотистый (Phlebodium aureum (L.) G. Smith).
Рис. 14. Вегетативное размножение папоротников1 - придаточными почками (Асплениум луковиценосный); 2 - наземными столонами (Нефролепис врзвышенный); 3 - участками корневища (Флебодиум золотистый).
Глава IV
Размножение папоротников
4.1. Вегетативное размножение папоротников.
Вегетативное размножение довольно широко распространено не только у покрытосеменных растений, но и у папоротников. Существуют самые разнообразные формы этого размножения. Оно может осуществляться с помощью длинных, разветвленных корневищ, характерных для целого ряда папоротников.
У представителей семейства Мараттиевых образуются придаточные почки, которые закладываются на прилистниках. Крупные листья Мараттиевых имеют у своего основания характерные прилистниковые придатки, которые сохраняются на стеблях после опадания листьев и служат не только запасающими органами (в них накапливается крахмал), но и органами вегетативного размножения. Покоящиеся на прилистниках почки при благоприятных условиях дают начало новым растениям.
У многих видов рода Ужовник (Ophioglossum) наблюдается способность к эффективному вегетативному размножению. Новые побеги у них появляются из придаточных почек, образующихся на корнях.
У некоторых папоротников придаточные почки образуются на листьях (Цератоптерис, Пузырник, Асплениум). Цератоптерис василисниковидный (Ceratopteris thalictroides) – известное аквариумное растение. В естественных условиях растет в мелководных водоемах, в канавах, является сорняком на рисовых полях. Молодые растения цератоптериса укоренены в почве. При затоплении поля они становятся свободноплавающими и активно размножаются вегетативным путем. В синусах листовых сегментов, на верхушках листьев, на рахисе появляются мелкие выводковые почки, развивающиеся в новые молодые растения, долго остающиеся связанными с материнским. Освобождаются они после загнивания или обламывания листа материнского растения.
У Пузырника луковиценосного (Cystopteris bulbifera), растущего во влажных лесах, по оврагам, на известковых скалах в Северной Америке, придаточные почки образуются на нижней стороне листьев. Почки имеют сферическую форму и напоминают луковички. Они отваливаются и дают начало молодым растениям. Благодаря вегетативному размножению этот папоротник не только образует большие заросли на одном месте, но и завоевывает новые территории.
Придаточные почки образуются также на листьях Асплениума луковиценосного и Асплениума живородящего. Они прорастают еще прикрепленными к материнскому растению. Отделившись от него и попав в благоприятные условия, они дают начало новым растениям. Форма вегетативного размножения, описанная у Цератоптериса, Цистоптериса и Асплениума, называется живорождением
, или вивипарией
. Иногда вивипария бывает очень интенсивной и подавляет развитие спорангиев (Левина, 1964).
Своеобразная форма вегетативного размножения свойственна дальневосточному папоротнику Кривокучнику сибирскому (Camptosorus sibiricus). Этот вид имеет цельные листья с сильно оттянутой верхушкой, переходящей в длинный жгут. Жгут заканчивается почкой, которая при соприкосновении с субстратом развивается в новое растение. Странствуя таким способом, вид завоевывает себе жизненное пространство.
Как уже отмечалось выше (см. главу III), легко размножаются вегетативно виды рода Нефролепис. Размножаются они с помощью наземных столонов. Некоторые виды этого рода размножаются при помощи клубней, образующихся на подземных столонах.
Эффективно размножаются вегетативным путем водные разноспоровые папоротники – виды родов Сальвиния (Salvinia) и Азолла (Azolla). У Сальвинии в узлах между листьями образуются почки, на которых развиваются ветви корневища. Они легко отламываются от основного стебля, и их обломки разносятся течением по водоему, где возникают новые заросли. В озере Кариба (Африка) Сальвиния ушковидная, завезенная сюда из Южной Америки, покрывает местами водную поверхность слоем до 25 см толщиной, препятствуя судоходству и мешая нормальной работе турбин гидроэлектростанции.
Азолла размножается вегетативно с помощью боковых ветвей, которые легко отламываются от главного стебля и течением разносятся в другие водоемы. Благодаря вегетативному размножению Азолла, как и Сальвиния, образзует в стоячих и слабо проточных водоемах значительные заросли.
Способность к вегетативному размножению широко используется человеком при размножении папоротников в условиях культуры. Для наших объектов подобная форма размножения характерна для Асплениума луковиценосного, Нефролеписа возвышенного и Флебодиума золотистого (Рис. 14). У этих видов имеются специализированные органы вегетативного размножения.
Технология искусственого вегетативного размножения папоротников
Для размножения Асплениума луковиценосного срезали лист папоротника с образовавшимися на нем молодыми растениями. Кювету наполняли смесью для черенкования и слегка ее уплотняли. Смесь для черенкования готовили из равных количеств листовой земли и хорошо промытого речного песка. Срезанный лист раскладывали на субстрате и прикрепляли небольшими проволочными скобками. Субстрат поливали. Кювету сверху закрывали полиэтиленовой пленкой и помещали в защищенное от прямых солнечных лучей место. Следили за тем, чтобы лист не подсыхал. Когда молодые растения укоренялись (примерно через 1,5-2 месяца), их разделяли и высаживали в горшки.
Более проста технология выращивания Нефролеписа возвышенного. Молодые растения, развивающиееся из почек на наземных столонах, легко укореняются. Для получения массового материала материнское растение следует выращивать в емкости с относительно большой площадью поверхности. Образующиеся наземные столоны в случае необходимости прикрепляли к субстрату проволочной скобой. Как только молодые растения укоренялись, связывающие их с материнским растением столоны обрезали. Молодые растения пересаживали в отдельные горшки.
Флебодиум золотистый размножали участками корневища. Это растение имеет толстое, длинное, стелющееся по субстрату корневище. Корневище ветвится, образуя боковые побеги. Боковые побеги (участок корневища с верхушечной почкой и листьями в количестве 1-2) обрезали и высаживали в отдельные горшки.
4.2. Размножение папоротников спорами.
Нами проведен эксперимент по выращиванию из спор таких папоротников, как Циртомиум серповидный, Листовик обыкновенный, Асплениум луковиценосный, Птерис критский и Флебодиум золотистый. Адиант венерин-волос и Нефролепис возвышенный из-за отсутствия спор нами не выращивались. Папоротники можно выращивать в горшках, прикрытых сверху стеклом для поддержания высокой постоянной влажности воздуха. Мы выращивали папоротники в чашках Петри по методике, описанной во 2-й главе настоящей работы.
Споры папоротников созревают в спорангиях. Спорангии изученных видов не имеют существенных морфологических отличий. Они округлой или слегка овальной формы, сидят на длинных ножках. На поверхности спорангия имеется продольное механическое кольцо, состоящее из одного ряда клеток. У большей части клеток кольца оболочки утолщены. Часть клеток кольца (примерно 1/3) имеет относительно тонкие оболочки. При созревании спор наступает момент, когда стенка спорангия разрывается. Разрыв происходит в месте, где клетки кольца имеют тонкие оболочки. Кольцо изгибается кнаружи, как бы выворачиваясь наизнанку, при этом споры высыпаются из спорангия. Этот процесс можно наблюдать под микроскопом МБС, при подсушивании спорангиев.
Спорангии папоротников собраны в сорусы. По положению сорусов на вайях у изучаемых видов встречаются 2 типа спорангиев. Для Адиантума, Птериса и Асплениума характерны маргинальные (краевые) спорангии. Сорусы у этих видов расположены по краю листовой пластинки. У Циртомиума, Нефролеписа, Листовика и Флебодиума спорангии ламинальные (поверхностные), сорусы у них расположены на нижней стороне листа.
Сорусы папоротников прикрыты индузием, лишь у Адиантума и Птериса покрывальца отсутствуют. Сорусы у этих видов защищены загнутым на нижнюю сторону краем листовой пластинки. Момент созревания спор мы определяли по изменению окраски соруса (зеленая окраска переходит в коричневую).
Собранные споры высевали на субстрат в чашки Петри. Выращивание проводили при комнатной температуре (18-25оС) в защищенном от прямых солнечных лучей месте. Через месяц (25-35 дней) на поверхности субстрата появлялся зеленый налет, состоящий из нитей и формирующихся пластинок гаметофита. Процесс от прорастания спор до формирования зрелого гаметофита в наших экспериментах продолжался около 2 месяцев. Сравнение гаметофитов исследованных видов не выявило существенных видовых отличий. В то же время форма и размеры гаметофитов одного и того же вида изменяются в зависимости от условий произрастания, в частности, от густоты посева. Подобная закономерность отмечалась в литературе и для дальневосточных видов папоротников (Храпко, 1989, 1996). Зрелые гаметофиты наших видов пластинчатые, сердцевидной формы. Пластинка состоит из одного слоя хлорофиллоносных клеток, за исключением многослойной срединной части. От нижней части гаметофита (от центральной многослойной части) отходят многочисленные ризоиды.
После оплодотворения появляются проростки спорофита. В нашем эксперименте первые проростки в зависимости от вида, условий выращивания и времени посева появлялись на 50-80 день после посева. Проростки представляют собой небольшие (1-2 см) растения. Они имеют 1-4 вайи и 2-5 коротких корешка. Проростки прикреплены к гаметофиту (Рис. 15, 16). Питание растения на этом этапе осуществляется как за счет гаметофита, так и за счет собственной ассимиляции. Листья проростков существенно отличаются от вай взрослых растений простотой своего строения. У проростков Циртомиума и Флебодиума листовая пластинка округлая, по краю цельная или слегка лопастная (Рис. 15). У проростков Асплениума и Птериса вайи характеризуются большей рассеченностью (Рис. 16).
Последующие возрастные состояния спорофита на данном этапе исследования отмечены нами только для Циртомиума серповидного.
4.3. Цикл развития Циртомиума серповидного.
Под циклом развития понимают совокупность всех фаз развития, пройдя которые организм достигает зрелости и становится способным давать начало следующему поколению (Биологический..., 1986). Для цикла развития папоротников характерна смена гаплоидной и диплоидной фаз. Гаплоидная фаза папоротников представлена покоящейся спорой и гаметофитом. Диплоидная фаза – спорофитом. Покоящиеся споры можно рассматривать в качестве латентного периода гаметофита (Шорина, 1987, цит. по: Храпко, 1996).
Споры Циртомиума находятся в спорангиях, которые собраны в сорусы на нижней стороне листа (латеральные сорусы) на концах жилок. Споры округлой или слегка бобовидной формы. Спорангии на длинных ножках. Ножка состоит из 3 слоев клеток. Спорангии округлые с продольным
Рис. 15. Гаметофиты и проростки Циртомиума серповидного (1) и Флебодиума золотистого (2).
Рис. 16. Гаметофиты и проростки Асплениума
луковиценосного (1) и Птериса критского (2).
механическим кольцом. Механическое кольцо имеет 20-25 клеток с утолщенными стенками. Сорусы округлые с покрывальцем (индузием). Индузии округлой формы, щитовидные (прикреплены в одной точке близ середины), по краю выгрызенно-зубчатые (Рис. 17).
Начальные стадии развития гаметофита Циртомиума мы наблюдали в экспериментальных условиях при посеве спор на фильтровальную бумагу, смоченную раствором Кнопа. Проращивание спор проходило при комнатной температуре (от 18 до 25оС). Первые проросшие споры отмечались на 7-8 день после посева. После прорастания споры образуется нить гаметофита. Формирование нити происходит в течение 15-20 дней. Нить состоит из 3-5 клеток. Клетки нити содержат хлоропласты. На этом же этапе начинают формироваться ризоиды. Через 25-30 дней после посева начинается формирование пластинки гаметофита. В нашем эксперименте пластинки гаметофитов на 35 день имели от 8 до 40 клеток. Период от прорастания спор до формирования зрелого гаметофита Циртомиума продолжается 60-70 дней. Этапы развития гаметофита отражены на рис. 18.
Гаметофит Циртомиума наземный, сердцевидной формы, состоит из слоя хлорофиллоносных клеток. На гаметофите имеются редкие папиллярные волоски, которые хорошо видны под микроскопом по краю пластинки. Папиллярные волоски представляют собой короткие, головчатые, сосочковидные выросты некоторых клеток гаметофита. Морфометрические данные гаметофитов и их изменчивость приведены в таблице 1.
Рис. 17. Сорусы Циртомиума серповидного.
Рис. 18. Этапы развития гаметофита Циртомиума серповидного1 -
прорастание спор; 2 - формирование нити; 3 - формирование пластинки
гаметофита; 4 - зрелый гаметофит.
Таблица 1
Изменчивость высоты и ширины гаметофитов Циртомиума серповидного
| Признаки
|
Лимиты
|
М
±
m
|
s
|
V, %
|
| Ширина, мм
|
4,1-5,6
|
5,1±0,08
|
0,49
|
9,6
|
| Высота, мм
|
3,0-4,5
|
3,85±0,08
|
0,48
|
12,4
|
Примечание: М - среднее значение признака, m - ошибка среднего значения, s - среднее квадратическое отклонение, V% - коэффициент вариации
Как видно из таблицы 1, амплитуда изменчивости размеров гаметофита, согласно шкалы С.А. Мамаева (1969), находится на низком или среднем уровне. Колебание параметров связано в основном с условиями выращивания и зависит, в частности, от густоты посева.
При хорошем уходе на 70-80 день после посева появляются первые проростки спорофита (Рис. 19). Проростки прикреплены к гаметофиту, за счет которого и осуществляется в основном их питание. Вайи проростков имеют совершенно иную форму, чем вайи зрелых растений. Листовая пластинка цельная, не разделенная на отдельные сегменты. Край листовой пластинки слегка выемчатый (Рис. 20). Характеристика некоторых признаков проростков Циртомиума дана в таблице 2.
Таблица 2
Изменчивость некоторых признаков проростков Циртомиума серповидного
| Признаки
|
Лимиты
|
М
±
m
|
s
|
V, %
|
| Количество листьев
|
2-4
|
2,9±0,12
|
0,7
|
24,0
|
| Высота растения, мм
|
19,0-31,0
|
26,4±0,73
|
5,5
|
15,3
|
| Ширина листовой пластинки, мм
|
4,1-10,2
|
5,82±0,4
|
2,24
|
38,4
|
Примечание: обозначения те же, что и в таблице 1.
Рис. 19. Гаметофиты и первые проростки Циртомиума серповидного, выращенные в чашке Петри.
Рис. 20. Проросток Циртомиума серповидного.
Уровень изменчивости признаков проростков выше уровня вариабельности гаметофитов и находятся на повышенном и высоком уровне.
Через 6-7 месяцев после посева наступает фаза молодого спорофита. Гаметофит к началу этой фазы отмирает и растение полностью переходит на самостоятельное питание. Усложняется строение листьев, которые становятся близки к вайям зрелых спорофитов. В отличие от листьев зрелых спорофитов, листья молодых растений имеют меньшее число сегментов (от 3 до 10). Количество сегментов у зрелых спорофитов варьирует от 8 до 25.
Через 15-20 месяцев растение переходит в фазу зрелого спорофита. Характерной чертой этого возрастного состояния является наличие спороношения.
Длительность периодов возрастного состояния Циртомиума серповидного весьма вариабельна. Она зависит от условий ухода, созданных при выращивании папоротника. Поэтому определять продолжительность той или иной фазы развития можно лишь приблизительно.
4.4. Использование полученных материалов в курсе ботаники средней и высшей школы.
Данные, полученные в ходе выполнения данной дипломной работы, могут найти широкое применение как на уроках биологии в средней школе, так и на занятиях по курсу общей ботаники в высшей школе. С успехом полученные материалы можно использовать при проведении факультативных занятий, а также занятий в кружках биологического профиля.
Прежде всего данный материал можно использовать при изучении темы «Отдел Папоротниковидные». Имея коллекцию оранжерейных папоротников, показать разнообразие представителей этой группы растений. Папоротниковидные очень разнообразны по внешнему виду. Кроме наземных корневищных паопротников в этой группе можно продемонстрировать такую жизненную форму, как эпифитные растения, например, Платицериум двувильчатый. Разнообразны и листья папоротников. Чаще всего встречаются перисторассеченные листья, но имеются виды с цельной листовой пластинкой - Костенец гнездовой, Листовик обыкновенный и другие. Обратить внимание, что молодые листья улиткообразно закручены. Черешки листьев покрыты характерными буроватыми чешуями. Кроме того листья папоротников растут верхушкой, как и побеги.
Для темы “Размножение папоротников” нами получен богатый материал. Здесь нужно познакомить учащихся со строением спорангиев. Показать процесс вскрытия спорангиев, что можно наблюдать под микроскопом при их подсушивании. Познакомить с разнообразием сорусов. Они различаются как расположением на листе, так и своей формой. Чаще встречаются поверхностные сорусы (Циртомиум), но могут быть и краевые (Адиантум). Сорусы по форме бывают округлые (Циртомиум), а бывают вытянутой, палочкообразной формы (Листовик). Могут иметь покрывальце разнообразной формы, а могут быть и без него (Птерис).
Изучая цикл развития папоротника, на живом материале можно продемонстрировать заростки (гаметофиты). Рассмотреть их строение под микроскопом и отметить пластинку, состоящую из слоя хлорофиллоносных клеток, и ризоиды, с помощью которых заросток прикрепляется к субстрату. Показать проростки папоротника, которые прикреплены к заростку и частично питаются за его счет. Отметить отличие листьев проростков от листьев взрослых растений.
В теме “Вегетативное размножение растений” целесообраззно познакомить с различными формами этого вида размножения у папоротников. Это прежде всего размножение корневищами (Флебодиум золотистый), а также с помощью выводковых почек (Асплениум живородящий, Цератоптерис василисниковый) и столонов (Нефролепис).
В теме “Побег” можно познакомить учащихся с видоизменением побега – корневищем, показать особенности его строения.
Выводы
1. Изученные нами виды папоротников можно с успехом выращивать из спор в оранжерейных условиях.
2. Некоторые виды папоротников хорошо размножаются вегетативно: Нефролепис возвышенный - столонами, Флебодиум золотистый - участками корневища, Асплениум луклвиценосный - придаточными почками, образующимися на листьях.
3. Спорангии изученных видов имеют однослойную стенку и продольное механическое кольцо. Ножка спорангия имеет 2 или 3 слоя клеток.
4. Сорусы у Птериса критского, Адиантума, Асплениума луковиценосного маргинальные (краевые); у Нефролеписа возвышенного, Циртомиума серповидного, Листовика обыкновенного и Флебодиума золотистого - ламинальные (поверхностные).
5. Сорусы Птериса критского и Адиантума имеют ложное покрывальце, представляющее собой отогнутый край листа, прикрывающий сорус. Сорусы других видов изученных нами напоротников покрыты настоящими покрывальцами.
6. Гаметофит выращенных папоротников автотрофные, дорзовентральные, имеют вид небольших сердцевидных пластинок. Существенных морфологических отличий между гаметофитами разных видов папоротников нет.
7. На развитие гаметофитов влияют условия среды - температура и влажность воздуха. Низкая температура и пересушивание почвы резко замедляют развитие гаметофитов.
8. Начало прорастания спор Циртомиума серповидного наблюдается на 7-9 день после посева. Период от прорастания спор до формирования зрелого гаметофита продолжается 60-70 дней. Полный цикл развития от посева спор до зрелого спорофита составляет около 20 месяцев.
9. Оптимальные сроки посева спор Циртомиума - с марта по август включительно.
Литература
Агеева Г.А., Лаврова К.Г. Цветы в вашем доме. Петрозаводск: Карелия, 1992 174 с.
Арнаутова Е.М. Выращивание папоротников из спор//Бюлл. Гл. ботан. сада. 1987. Вып. 144. С. 65-66.
Арнаутова Е.М. Гаметофиты некоторых видов рода Woodsia (Woodsiaceae)// Ботан. журн. 1988. Т. 73, N 2. С. 198-203.
Арнаутова Е.М. Развитие гаметофита у некоторых видов рода Gymnocarpium Newm.// Онтогенез высших цветковых растений. Киев, 1989. С. 6.
Биологический энциклопедический словарь/ Гл. ред. М.С. Гиляров. М.: Сов. Энциклопедия, 1986. 831 с.
Вальтер Г. Растительность Земного шара. М.: Прогресс, 1974. 422 с.
Васильев А.Е., Воронин Н.С., Еленевский А.Г., Серебрякова Т.И. Ботаника. Анатомия и морфология растений. М.: Просвещение, 1978. 478 с.
Верзилин Н.М. Путешествие с домашними растениями. Л.: Детгиз, 1954. 335 с.
Воробьев Д.П. Определитель сосудистых растений окрестностей Владивостока. Л.: Наука, 1982. 254 с.
Воробьев Д.П., Ворошилов В.Н., Горовой П.Г., Шретер А.И. Определитель растений Приморья и Приамурья. М.-Л.: Наука, 1966. 496 с.
Воронова Н.В. Комнатное цветоводство. Новосибирск: Новосибирское кн. изд-во, 1992. 80 с.
Ворошилов В.Н. Определитель растений советского Дальнего Востока. М.: Наука, 1982. 672 с.
Второв П.П., Дроздов Н.Н. Биогеография. М.: Просвещение, 1978. 271 с.
Гаммерман А.Ф., Шупинская М.Д.. Яценко-Хмелевский А.А. Растения-целители. М.: Высшая школа, 1963. 423 с.
Герасимов С.О., Журавлев И.М., Серяпин А.А. Редкие комнатные растения. М.: Аквариум, 1997. 224 с.
Гладкова В.Н. Семейство Адиантовые (Adiantaceae)// Жизнь растений. М.: Просвещение, 1978а. Т. 4. С. 189-190.
Гладкова В.Н. Подсемейство Петрисовые (Pteridoideae)// Жизнь растений. М.: Просвещение, 1978,. Т. 4. С. 195-197.
Гладкова В.Н. Семейство Асплениевые (Aspleniaceae)// Жизнь растений. М.: Просвещение, 1978в. Т. 4. С. 222-227.
Даддингтон К.Л. Эволюционная ботаника. М.: Мир, 1972. 307 с.
Жданов В.С. Аквариумные растения. М.: Лесная пром-сть, 1973. 160 с.
Зайцев Г.Н. Математика в экспериментальной ботанике. М.: Наука, 1990. 296 с.
Золотницкий Н.Ф. Аквариум любителя. М.: Терра, 1993. 784 с.
Карри-Линдал К. Европа. М.: Прогресс, 1981. 330 с.
Комаров В.Л., Клобукова-Алисова Е.Н. Малый определитель растений Дальневосточного края. Владивосток, 1925. 516 с.
Комаров В.Л., Клобукова-Алисова Е.Н. Определитель растений Дальневосточного края. Л.: Изд-во АН СССР, 1931. Т. 1. 622 с.
Комаров В.Л. Происхождение растений. М.-Л.: Изд-во АН СССР, 1936. 190 с.
Комнатный аквариум/ под ред. М.А. Пешкова. Алма-Ата: Изд-во сельскохозяйственной литературы, 1964. 238 с.
Котухов Ю.А. Выращивание папоротников из спор// Бюл. Гл. ботан. сада. 1968. Вып. 71. С. 85-89.
Кудрявец Д.Б. Как вырастить цветы. М.: Просвещение, 1993. 176 с.
Левина Р.Е. Многообразие и эволюция форм размножения растений. М.: Просвещение, 1964. 66 с.
Леме Ж. Основы биогеографии. М.: Прогресс, 1976. 309 с.
Мак-Миллан Броуз Ф. Размножение растений. М.: Мир, 1987. 192 с.
Мамаев С.А. О проблемах и методах внутривидовой систематики древесных растений. II. Амплитуда изменчивости// Закономерности формирования и дифферренциации вида у древесных растений/ Тр. ин-та экологии раст. и жив.. вып. 64. Свердловск: УНЦ АН СССР, 1969. С. 3-38.
Махлин М.Д. Занимательный аквариум. М.: Пищевая пром-сть, 1976. 288 с.
Махлин М.Д. Амурский аквариум. Хабаровск: Кн. изд-во, 1990. 320 с.
Машковский М.Д. Лекарственные средства. М.: Медицина, 1977. Т. 1. 623 с.; Т. 2. 560 с.
Мейер К.И. Морфогения высших растений. М.-Л.: Изд-во МГУ, 1957. 254 с.
Муравьева Д.А. Тропические и субтропические лекарственные растения. М.: Медицина, 1983. 336 с.
Никонов Н.Г. Созвездие кактусов. Свердловск: Средне-Уральское кн. изд-во, 1978. 136 с.
Пастушенков Л.В., Пастушенков А.Л., Пастушенков В.Л. Лекарственные растения. Л.: Лениздат, 1990. 384 с.
Пашина Г.В. Растения и косметика. Минск: Ураджай, 1993. 252 с.
Петров В.В. Четыре недели в Южном полушарии. М.: Мысль, 1981. 128 с.
Пешеходько В.М., Храпко О.В. Определитель папоротников Приморского края. Владивосток: Изд-во ДВГУ, 1994. 55 с.
Письяукова В.В. Элементы морфологической эволюции растений. Л.: ЛГПИ, 1980. 76 с.
Плонский В.Д. Аквариумные растения. М.: Аквариум, 1998. 144 с.
Рабинович М.И. Лекарственные растения в ветеринарной практике. М.: Агропромиздат, 1987. 288 с.
Рейвн П., Эверт Р., Айкхорн С. Современная ботаника. М.: Мир, 1990. Т. 1. 348 с.; Т. 2. 344 с.
Серая Г.П., Комов С.В., Безденежная Н.Л., Мыльников Н.М. Особенности морфогенеза и возрастные состояния спорофита некоторых видов крупнокорневищных лесных папоротников// Онтогенех травянистых поликарпических растений. Свердловск, 1980. С. 91-101.
Серебряков И.Г. Морфология вегетативных органов высших растений. М.: Советская наука, 1952. 391 с.
Словарь ботанических терминов/ под ред. И.А. Дудки. Киев: Наукова думка, 1984. 308 с.
Сурова Т.Д. Семейство Даваллиевые (Davalliaceae)// Жизнь растений. М.: Просвещение, 1978. Т. 4. С. 237-242.
Тавлинова Г.К. Цветы в комнате и на балконе. Л.: Колос, 1982. 192 с.
Тавлинова Г.К. Комнатное цветоводство. СПб.: Агропромиздат, 1997. 480 с.
Таршис Г.И. Подземные органы многолетних травянистых растений. Свердловск: СГПИ, 1975. 133 с.
Тахтаджян А.Л. Отдел папоротниковидные. Общая характеристика// Жизнь растений. М.: Просвещение, 1978а. Т. 4. С. 149-170.
Тахтаджян А.Л. Флористические области Земли. Л.: Наука, 1978б. 248 с.
Тахтаджян А.Л. Происхождение листовых органов// Жизнь растений. М.: Просвещение, 1978в. Т. 4. С. 12.
Уоллес А.Р. Тропическая пррирода. М.: Мысль, 1975. 223 с.
Федоров А.А., Кирпичников М.Э.. Артюшенко З.Т. Атлас по описательной морфологии высших растений: Лист. М.-Л.: Изд-во АН СССР, 1956. 302 с.
Федоров А.А., Кирпичников М.Э.. Артюшенко З.Т. Атлас по описательной морфологии высших растений: Стебель и корень. М.-Л.: Изд-во АН СССР, 1962. 350 с.
Хессайон Д.Г. Все о комнатных растениях. М.: Кладезь, 1995. 255 с.
Храпко О.В. Корноптерис городчато-пильчатый - Cornopteris czenulatoserrulata (Makino) Nakai// Биология редких сосудистых растений советского Дальнего Востока. Владивосток: ДВО АН СССР, 1990а. С. 6-15.
Храпко О.В. Лепторумора Микели - Leptorumohra miqueliana (Maxim. ex Franch. et Savat.) H. Ito// Биология редких сосудистых растений советского Дальнего Востока. Владивосток: ДВО АН СССР, 1990б. С. 15-19.
Храпко О.В. Пиррозия язычная - Pyrrosia lingua (Thunb.)Farw.// Биология редких сосудистых растений советского Дальнего Востока. Владивосток: ДВО АН СССР, 1990в. С. 19-30.
Храпко О.В. Кочедыжник красночерешковый - Athyrium rubripes (Kom.) Kom.// Биологические особенности сосудистых растений советского Дальнего Востока. Владивосток: ДВО РАН, 1991а. С. 4-13.
Храпко О.В. Леписорус уссурийский - Lepisorus ussuriensis (Regel et Maack) Ching// Биологические особенности сосудистых растений советского Дальнего Востока. Владивосток: ДВО РАН, 1991б. С. 13-22.
Храпко О.В. Папоротники юга Дальнего Востока России. Владивосток: Дальнаука, 1996. 200 с.
Цвелев Н.Н. Отдел Папоротниковидные Polypodiophyta// Сосудистые растения советского Дальнего Востока. СПб: Наука, 1991. Т. 5. С. 14-93.
Ценопопуляции растений/ Л.Б. Заугольнова, Л.А. Жукова, А.С. Комаров, О.В. Смирнова. М.: Наука, 1988. 184 с.
Чичагова М.С. Выращивание папоротников из спор в питательной среде// Бюл. Гл. ботан. сада. 1956. Вып. 24. С. 101-102.
Шорина Н.И., Ершова Э.А. Орляк обыкновенный// Биологическая флора Московской области. М.: Изд-во МГУ, 1990. С. 4-20.
Шретер А.И. Целебные растения Дальнего Востока. Владивосток: Дальневосточное кн. изд-во, 1992. 159 с.
Ярошенко П.Д. Теоретические вопросы в курсах ботаники для высшей школы. Владимир: ВГПИ, 1969. 217 с.
Ярошенко П.Д. Общая биогеография. М.: Мысль, 1975. 187 с.
Сatalogue of livung plant collections, Royal Botanic Gardens Kew, 1984, pt. l. 94 p.
Nayar B., Kaur S. Gametophytes of homosporous ferns// Bot. Rev. 1971. V. 37, N 3. P. 295-396.
|
