
ВЫСШЕЕ ОБРАЗОВАНИЕ
Г. М. АБДУРАХМАНОВ, И. К. ЛОПАТИН,
Ш. И. ИСМАИЛОВ
ОСНОВЫ
ЗООЛОГИИ
И
ЗООГЕОГРАФИИ
Рекомендовано
Министерством образования Российской Федерации
в качестве учебника для студентов высших педагогических
учебных заведений, обучающихся по специальностям «Биология»,
«География», «Педагогика и методика начального образования»

УДК 591.5
ББК 28.6я73
А 13
Рецензенты:
доктор биологических наук, заслуженный деятель
науки России О.Л.Крыжановский;
доктор биологических наук, профессор И.Х.Шарова
Абдурахманов Г. М. и др.
А 13 Основы зоологии и зоогеографии: Учебник для студ. высш пед. учеб. заведений / Г. М.
Абдурахманов, И. К Лопатин, Ш.И.Исмаилов. - М.: Издательский центр «Академия», 2001. -496с.
ISBN 5-7695-0625-3
В учебнике основное место уделяется систематике животных, рассматриваются происхождение, строение, функции, развитие животных, подробно описываются ареалы их распространения и фаунистические области земного шара, приводятся общие сведения о значении животных в биосфере и их роли в жизни и хозяйстве человека.
Учебник может быть использован также преподавателями школ и средних педагогических учебных заведений.
УДК 591.5
ББК 28.6я73
Учебное издание
Абдурахманов Гайирбег Магомедович,
Лопатин Игорь Константинович,
Исмаилов Шейх Ибрагимович
Основы зоологии и зоогеографии
Учебник
Редактор Е. А. Гороховская.
Технический редактор Я Ф. Коржуева.
Компьютерная верстка: Г. Ю. Никитина.
Корректоры В. И.Хомутова. Э. Г. Юрга
Подписано в печать 23.03.2001. Формат 60х90/16 Бумага тип. №
2 Печать офсетная. Гарнитура «Тайме». Усл. печ. л. 31,0. Тираж 30 000 экз. (1-и завод 1 -10 000 экз.).
Заказ №369.
Лицензия ИД № 02025 от 13.06.2000. Издательски._центр «Академия».
105043, Москва, ул. 8-я Парковая, 25. Тел./факс (095)165-4666, 367-0798, 305-2387.
Отпечатано на Саратовском полиграфическом комбинате.
410004, г. Саратов, ул. Чернышевского, 59.
ISBN
5-7695-0625-3
© Абдурахманов Г.М., Лопатин И.К., Исмаилов Ш.И., 2001
® Издательский центр «Академия», 2001
Зоология – это наука о животном мире. Хотя ее отдельные разделы касаются строения, жизненных функций, поведения и связей организмов как единого целого со средой, все же объектом зоологии оказываются не отдельные животные и даже не отдельные их типы; а все животное царство в целом.
Реклама

Зоология является составной частью биологии, изучающей живую природу. Живые организмы по своей структуре несравненно сложнее объектов неживой природы, соответственно этому и биология гораздо сложнее физики и химии. Все живые организмы относятся к нескольким царствам. Животное царство — это часть живого мира, представители которой характеризуются гетеротрофным питанием и подвижностью. На первый взгляд, различия между растениями и животными настолько очевидны, что не требуют обоснований. В действительности дело обстоит сложнее, и приведенное выше определение животного царства нуждается в дополнениях главным образом из-за ряда исключений и пограничных случаев.
Возьмем, к примеру, питание растений и животных. Первые из них — автотрофны. Они способны синтезировать питательные вещества из простых молекул в процессе фотосинтеза. Животные — гетеротрофны. Они добывают энергию путем поглощения питательного материала, синтезируемого растениями или другими живыми организмами. Короче, они нуждаются в готовых органических соединениях, поскольку сами синтезировать их не могут. Однако грибы и многие бактерии, относящиеся к другим царствам, также гетеротрофны.
Далее, отнесение живых организмов к царству животных только на основе их подвижности также недостаточно аргументирование. Среди животных немало сидячих, прикрепленных организмов, например губки, коралловые полипы, морские лилии или ряд моллюсков. С другой стороны, существуют подвижные растения, особенно из одноклеточных (зеленые жгутиконосцы). Не абсолютны и такие признаки, как наличие толстых целлюлозных оболочек в клетках растений и тонкой мембраны в животных клетках, ограниченный определенным периодом рост животных и продолжающийся всю жизнь рост растений и т. д. Среди животных оболочники имеют целлюлозные оболочки клеток, а крокодилы и черепахи растут в течение всей жизни. Поэтому правильнее будет характеризовать животных как организмы, обладающие комплексом следующих признаков. Большинство животных подвижно; их клетки покрыты тонкой мембраной; основные органы расположены внутри тела, имеющего довольно постоянную форму; рост обычно приурочен к определенному периоду развития; они гетеротрофны, а конечные продукты их обмена веществ — углекислый газ, вода и мочевина. Этот комплекс признаков в целом удовлетворительно характеризует сущность животного.
Реклама

Задачи зоологии весьма разнообразны. С одной стороны, это изучение строения (внешнего и внутреннего) и функционирования животных организмов, их индивидуального развития, их систематики, географического распространения, с другой — изучение взаимоотношений животных с другими организмами, в том числе с другими животными, выявление зависимости животных от условий среды обитания, изучение исторического развития различных групп и т. д. Так как среди животных немало видов, прямо или косвенно используемых человеком, а также вредителей культурных растений и переносчиков различных заболеваний, зоология интересуется и практическим значением животных, а полученные в результате зоологических исследований данные широко применяются в сельском и лесном хозяйстве, рыбоводстве, медицине и ветеринарии.
Разнообразие животного мира земного шара, разносторонность задач и многообразие методов исследования привели к выделению многих самостоятельных зоологических дисциплин.
Морфология животных
изучает внешнее (собственно морфология) и внутреннее (анатомия) строение. К ней же относятся гистология; изучающая микроскопическое строение тканей и органов; цитология, исследующая структурные особенности клеток животных; эмбриология — наука о закономерностях индивидуального развития. Большинство морфологических дисциплин основано на сравнительном изучении строения различных животных, что позволяет называть их сравнительной анатомией, сравнительной эмбриологией и т. д.
Физиология животных —
наука о жизненных процессах, проходящих в организме животных, таких, как питание и пищеварение, дыхание, кровообращение, выделение, нервная деятельность и т. д. В наше время все эти процессы изучаются также в сравнительном плане, и поэтому физиология стала сравнительной физиологией.
Поведение животных изучает этология —
относительно молодая зоологическая дисциплина, хотя в числе ее основоположни ков числится знаменитый исследователь поведения насекомых Анри Фабр, работавший в XIX в.
Экология животных
как часть общей экологии исследует взаимоотношение животных с окружающей средой и все разнообразие приспособлений животных к среде, обеспечивающих их выживание, расселение, размножение и развитие.
Генетика животных
изучает применительно к животным организмам закономерности наследственности и изменчивости.
Систематика животных
вместе с филогенетикой
— фундаментальный раздел зоологии, синтезирующий данные всех отраслей биологии, изучающих животных. Систематика отражает естественную систему организмов с их объективно существующими родственными взаимоотношениями. Исходный раздел систематики — фаунистика — изучает состав и особенности животного мира нашей планеты. С другой стороны, фаунистику можно считать частью зоогеографии — науки о географическом распространении животных.
Наряду с перечисленным существует и другое деление зоологии — по объектам исследования. Так выделяются науки о простейших — протозоология, о насекомых — энтомология, о рыбах — ихтиология, о птицах — орнитология и т. п.
Современная биология изучает живое на всех уровнях его организации — от молекулярного до биосферного. При этом мы наблюдаем постепенное усложнение уровней организации; на каждом уровне рождается новое качество, не свойственное предшествующему уровню. В то же время на каждом последующем уровне используются свойства простых компонентов, лежащих в основе более сложных систем.
Известный советский биофизик академик Г. Франк классифицирует организацию субстрата, из которого построено все живое, следующим образом.
Первый уровень— это макромолекулы белка и нуклеиновых кислот, слагающие любую живую систему. Они имеют определенные свойства, позволяющие живой системе функционировать. Называть подобные молекулы живыми не следует. Несмотря на то что макромолекулам присущи некоторые черты живого, они не обладают характерной для него совокупностью признаков.
Второй уровень организации — действующие химические системы — молекулы типа ферментов, которые видоизменяют другие молекулы. В настоящее время биохимики научились воспроизводить в условиях эксперимента цепочки ферментных систем, в которых уже проявляются самые элементарные принципы регуляции биохимических процессов. Более того, если иметь соответствующие ферменты и молекулы нуклеиновой кислоты, можно провести в искусственных условиях воспроизведение (репликацию) этой нуклеиновой кислоты. Тем не менее химические системы, работающие даже на очень сложном уровне, еще нельзя назвать живыми.
Третий уровень организации жизни — внутриклеточные органеллы и частицы, например ядра клеток, рибосомы, митохондрии, мембраны и т. д. Субклеточные органеллы имеют некоторые свойства и функции живого, но, строго говоря, они еще не живые. Только при переходе на клеточный уровень мы получаем настоящую живую систему — клетку.
Четвертый уровень— живая клетка в целом. Это единая функционирующая система со сложным молекулярным структурным аппаратом. Только клетке в целом присущ цикл развития и самовоспроизведения в ряду поколений. При этом используются свойства и макромолекул, и работающих химических систем, и субклеточных частиц в их единстве, устойчивости и организационном самоуправлении. Известен эксперимент по пересадке ядра, взятого из дифференцированной клетки эпителия кишечника лягушки, в ее яйцо. В определенных случаях из такого «составного» яйца развивался полноценный организм. Это значит, что специализированная клетка взрослого организма вопреки своей специфике способна в той или иной мере сохранять в ядре всю информацию, необходимую для развития целого организма.
Можно сделать вывод о том, что жизнь начинается с живой клетки, имеющей сложную внутреннюю молекулярную структуру, разнообразные мембраны и клеточное ядро, в котором содержится записанная в ДНК информация. Мембраны разделяют внутриклеточные вещества разного характера и отличаются специфической проницаемостью. Есть в клетке и механизмы передачи информации ферментным системам.
Химическая деятельность клетки крайне сложна. Для нее характерна координация скоростей многих параллельно идущих и переплетающихся «химических конвейеров». Ферментативные процессы в клетке представляют собой целый ансамбль. Все процессы, протекающие в клетке, возможны лишь при наличии четко организованного транспорта веществ, их передвижения, накопления и выделения. В целом это самостоятельная, очень устойчивая, саморегулирующаяся система, имеющая определенную цикличность развития, размножения и воспроизведения и управляющая теми субклеточными образованиями, которые в ней содержатся.
Пятый уровень— это многоклеточные организмы. Кстати, простейшие (одноклеточные) животные, хотя и состоят из единственной клетки, по своим функциям и принципу организации гораздо ближе к многоклеточным организмам, чем к клетке. Целому организму свойственна сложная и, вероятно, многократно дублируемая система саморегуляции, изучаемая физиологами. О высокой надежности ее говорят находки «живых ископаемых», сохранивших внешние формы и морфофункциональные признаки, несмотря на то, что как виды они существуют сотни миллионов лет. Характерным примером служит древнейшая рыба — целакант, известная по ископаемым остаткам давно, но неожиданно обнаруженная в Индийском океане всего каких-нибудь сорок лет назад. Ископаемый скелет ее, пролежавший в земле около 300 млн лет, ничем не отличается от скелета современного цела-канта. Неизменяемость в данном случае свидетельствует о высокой устойчивости и надежности системы.
Шестой уровень организации — экосистема в целом. Гигантской экосистемой является и биосфера нашей планеты. Для экосистемы характерна взаимосвязь и взаимозависимость различных организмов и их популяций. Легче всего проследить эту взаимосвязь на примере водной экосистемы, безразлично, пруд это или Мировой океан. В такой экосистеме осуществляется круговорот веществ, базирующийся на цепях и сетях питания.
Первым звеном пищевой (трофической) цепи являются одноклеточные водоросли (фитопланктон), использующие для питания растворенные неорганические вещества. Вторым звеном оказываются мельчайшие животные организмы (зоопланктон), питающиеся фитопланктоном. За счет этих звеньев живут планк-тоноядные («мирные») рыбы, служащие, в свою очередь, пищей хищным рыбам. Отмершими водными растениями и трупами животных питаются другие организмы — сапрофаги. В конце концов мертвая органика разлагается микроорганизмами-деструкторами и превращается в неорганические соединения, идущие на питание фитопланктона. В этом чрезвычайно сложном переплетении взаимоотношений проявляются сложные механизмы регуляции с изменяющимися количественными уровнями процессов.
Процессы, протекающие в экосистемах, являются выражением всеобщей связи и взаимодействия, существующих в природе. Эта всеобщность взаимодействия раскрывается как иерархия механизмов обратных связей. Благодаря им в любых живых системах наблюдается замечательная регуляция процессов и необычная их устойчивость.
Зоология тесно связана с практической деятельностью человека, и не только с теми ее видами, в которых прямо используются зоологические объекты (пчеловодство, рыборазведение, борьба с вредителями и т.д.), но и с делом охраны природы и восстановлением численности исчезающих видов животных. Без экологических знаний в этих видах своей деятельности человек был бы безоружен.
Живые организмы обитают в самых разнообразных условиях нашей планеты — на ее поверхности, в почве, в морях и океанах и даже в атмосфере. Совокупность всех живых организмов планеты — живое вещество — играет в ее развитии огромную роль. За 3 млрд лет, прошедших со времени возникновения жизни на Земле, живое вещество изменило и толщу горных пород, и водную оболочку, и атмосферу. Возникла биосфера.
Биосфера — это наружная оболочка земного шара, в которой развивалась жизнь в форме большого числа разнообразных организмов, населяющих поверхность суши, почву, нижние слои атмосферы и гидросферу, и которая по своему составу и своим свойствам является продуктом взаимодействия живой и неживой материи Земли (В. А. Ковда, 1971). Наибольший вклад в развитие представлений о биосфере и о протекающих в ней процессах внес выдающийся русский ученый академик В. И. Вернадский.
Живые организмы — самые активные участники таких процессов в биосфере, как поток энергии и круговорот веществ. Чтобы оценить их значение в качестве участников этих процессов, необходимо хотя бы в общих чертах рассмотреть, как они протекают.
Первичным источником внешней энергии, без которой невозможна жизнь на нашей планете, служит световое и тепловое излучение Солнца. Каждый живой организм получает эту энергию либо в прямом, либо в измененном виде, а затем передает ее другим организмам или выделяет во внешнюю среду. Можно сказать, что энергия проходит сквозь живую оболочку и излучается во внешнюю среду в виде тепла, которое живыми организмами уже не может быть усвоено. Поэтому мы говорим о потоке энергии, а не о ее круговороте. Конечно, проходя через биосферу, энергия трансформируется.
Первыми потребляют солнечную энергию зеленые растения. Они ее усваивают непосредственно путем фотосинтеза. Сосредоточенная в тканях растений энергия много раз преобразуется в глобальной экологической системе — биосфере, перед тем как выйти снова во внешнюю среду в «отработанной» форме теплового излучения. На каждом этапе превращения часть энергии теряется (уходит из общего потока). Уже растения теряют часть энергии на дыхание.
Вторыми на очереди потребителями энергии являются животные, поедающие растительную ткань или соки растений. Таких животных называют фитофагами, к ним относятся фитонемато-ды, насекомые, растительноядные моллюски, травоядные млекопитающие (например, грызуны, копытные). Из усвоенных веществ пищи (а усваивается сравнительно небольшая ее часть) фитофаги строят вещество своего тела, т. е. используют солнечную энергию в химическом, «законсервированном» виде. Они тоже в процессе жизнедеятельности теряют часть энергии на дыхание, но остальная ее часть усваивается другими потребителями, которые образуют третий уровень использования живой энергии.
Этот уровень — зоофаги, животные, питающиеся другими животными. К зоофагам относятся хищники и паразиты. Их можно разделить на несколько групп. К примеру, хищный жук, поедающий растительноядную гусеницу, может быть обозначен как хищник первого порядка. Насекомоядная птица, съедающая хищного жука, будет уже хищником второго порядка. Какой-нибудь сокол, нападающий на птицу и съедающий ее, окажется хищником третьего порядка. Между перечисленными организмами существуют тесные пищевые связи, по которым как по каналам растекается энергия, накопленная в зеленых растениях, теряется в процессах обмена веществ и дыхания и в конечном итоге возвращается (излучается) во внешнюю среду в форме тепловой энергии.
Но использование энергии, сконцентрированной в теле растений и животных, на этом не кончается. После их смерти за мертвое органическое вещество (а в нем еще достаточно энергетических запасов) принимаются животные сапрофаги и бактерии. Полуразложившиеся остатки растений в верхних частях почвы — детрит — используются беспозвоночными животными, грибами и почвенными бактериями. Трупы животных поедаются некрофагами (мухами, жуками-мертвоедами, стервятниками, гиенами и т. п.), а экскременты — копрофагами: жуками-навозниками, личинками мух и др. Итак, все живые и мертвые органические вещества находят своих потребителей, и это продолжается беспрерывно на протяжении миллиардов лет.
Доля энергии, усваиваемой живыми организмами, очень незначительна. Из общего ее количества, падающего на поверхность земли, растения поглощают не более 1 %. И на других уровнях (фитофаги, зоофаги и сапрофаги) используется примерно то же соотношение — около 1 % энергии предыдущего уровня. Остальное расходуется на процессы обмена веществ. Поэтому вдоль пищевой цепи происходит непрерывная и прогрессирующая убыль преобразуемой энергии.
Необходимо ясно представлять, что солнечная энергия непосредственно фиксируется и накопляется только зелеными растениями, поэтому их называют продуцентами. Все остальные живые организмы потребляют живое вещество, созданное продуцентами.
Эту совокупность живых организмов называют консументами (потребителями).
В отличие от энергетического потока, который, пронизывая биосферу, уходит в космическое пространство, вещество биосферы совершает циклические превращения, или круговорот. Примечательно, что количество вещества, принимающего участие в круговоротах, остается постоянным в течение целых геологических периодов, т. е. сотни миллионов лет. Это значит, что в биосфере совершается многократный круговорот веществ. Собственно, в круговоротах принимают участие не сами вещества, а слагающие их элементы — углерод, кислород, водород, азот и т. д. Живые организмы биосферы принимают участие в так называемом малом, или биологическом, круговороте. В самом обобщенном виде биологический круговорот можно описать следующим образом.
Растения, поглощая воду, а вместе с ней растворенные соли, а также углекислый газ из атмосферы, при помощи солнечной энергии создают органическое вещество. Животные, поедая растения, усваивают это органическое вещество, конечно, расщепляя его в процессе пищеварения и снова синтезируя «по-своему», превращая в свойственные животным организмам органические соединения. После смерти животных и отмирания растительной массы трупы и детрит потребляются сапрофагами, снова проходя этапы расщепления и нового синтеза. В конце концов органические соединения будут расщеплены бактериями и доведены до уровня неорганических веществ (солей), которые войдут в новый цикл, или круговорот, в качестве пищи новых растений. Таким образом, элементы многократно проходят через уровни консументов в одной и той же последовательности. В этом смысл круговорота веществ.
Имея представления о потоке энергии и круговороте веществ в биосфере, нетрудно определить место и роль животных в этих процессах. Животные как потребители готовых органических соединений принимают самое прямое участие в перераспределении энергии и вещества. Однако они не только потребляют и разрушают органическое вещество, но и способствуют перемещению различных химических элементов, которые в них накопляются. В этом их биосферная роль.
Животные зависят от окружающей среды, из которой они получают все, что им необходимо. Менее известна средообразую-щая роль животных, хотя, как оказалось, она весьма существенна. Достаточно сказать, что почва в значительной степени образуется при активном участии почвообитающих животных — роющих грызунов, многочисленных червей, личинок насекомых, клещей и разнообразных простейших. Относительно дождевых червей это было известно еще Ч. Дарвину. Животные принимали участие в формировании определенных ландшафтов, например степи. Доказано, что степь как тип растительности создавалась при участии больших стад копытных и других животных-«степняков».
Все это, вместе взятое, позволяет сделать вывод, что животные являются совершенно необходимым компонентом биосферы.
Животные играют значительную роль в жизни человека, прежде всего в его питании. На заре своего существования человек был простым собирателем, затем охотником, скотоводом, наконец, земледельцем. И в каждую из этих эпох он так или иначе использовал животных как источник высококалорийного белка. В различных климатических зонах процент животного белка в пище людей значительно колеблется. Различны и сами объекты.
Среди беспозвоночных издавна употребляются в пищу главным образом ракообразные (раки, крабы, креветки) и моллюски (устрицы, мидии, улитки и др.). Особенно интенсивно используются морские беспозвоночные жителями прибрежных районов Средиземного моря и Тихого океана, где в пищевой рацион входят даже морские ежи, голотурии и морские черви (палоло). Мировой промысел морских беспозвоночных уже превысил 10 % мировой добычи рыбы и продолжает расти. Осваиваются новые пищевые объекты, например криль (эвфаузиевые рачки, которыми питаются усатые киты). Проводятся исследования по использованию морского планктона.
Значительно большую роль в питании людей играют различные представители позвоночных животных. В настоящее время человечество получает до 10 % животных белков только за счет рыб.
Продуктивность Мирового океана наиболее высока в пределах так называемой шельфовой зоны с глубинами не более 200 м, но площадь шельфа невелика. В последние десятилетия на рыбопро-дуктивность океана сильно влияет деятельность людей: избыточный промысел, загрязнение воды нефтепродуктами, пестицидами и другими соединениями, уменьшение речного стока и т. п. Требуются серьезные меры по регулированию промысла и охране Мирового океана.
Основу морского промысла составляют такие рыбы, как тресковые, сельди, анчоусовые, камбаловые, скумбриевые и ряд других. Важную роль играют проходные рыбы, особенно тихоокеанские лососи — кета, горбуша, чавыча и др. В пресных водах добываются прежде всего карповые (сазан, лещ, плотва, карась, линь, толстолобик, белый амур).
Использование рыбных ресурсов в новых условиях требует их расширенного воспроизводства. Во многих странах и районах созданы специальные рыбоводные заводы и питомники. В них от идущих на нерест производителей получают половые продукты (икру и молоки), которые затем смешивают, что гарантирует оплодотворение. Развитие икры проходит в инкубационных аппаратах, а вылупившихся личинок — в специальных водоемах. Подросших мальков выпускают в естественные водоемы. В России работают рыбоводные осетровые и лососевые заводы. Молодь пресноводных рыб выращивают во многих районах России.
Большое развитие получило прудовое хозяйство. В прудах разводят не только карпа, но и форель, линя, сома и белого амура. В лучших прудовых хозяйствах получают до 4—5 т рыбы на 1 га водной площади.
В ряде стран в пищу используют некоторых земноводных — лягушек, саламандр. Крупных лягушек в США разводят на фермах. Употребляют в пищу также мясо крупных ящериц, змей и особенно черепах. Под влиянием неумеренной охоты исчезли гигантские сухопутные черепахи и резко сократилась численность морских видов.
Объектами промысловой или спортивной охоты служат многие птицы — гусеобразные, куриные, кулики и др. Все они имеют вкусное мясо. Во многих странах охотничье хозяйство ведется рационально, что наряду с разведением дичи дает добавочную продукцию с обширных территорий.
Немалый доход приносит и охотничий промысел млекопитающих. Дикие копытные — северные и благородные олени, лоси, сайгаки — служат важным источником мяса. Необходимо подчеркнуть, что численность всех этих животных у нас заметно увеличилась. В африканских государствах основную роль поставщиков мяса играют многочисленные антилопы, прекрасно использующие растительность саванны. Одно время большое значение имел китобойный промысел, но сейчас численность китов резко упала, и этот промысел запрещен, хотя некоторые страны нарушают запрет.
Вполне естественно, что в большинстве стран мира основную массу белка животного происхождения дает мясо животных — птиц и млекопитающих. Одомашнивание их началось еще в древности. Например, одомашнивание дикой банкивской курицы произошло в Индии за несколько тысячелетий до нашей эры. В результате селекции сейчас выведено около 100 пород домашних кур и достигнуто увеличение их массы примерно в 7, а яйценоскости в 40 раз. Это же можно сказать и о других домашних птицах — индюках, утках, гусях. Так, селезень крупной руанской породы весит 4—5 кг, а его дикий предок (кряква) — не более 1,5 кг.
Домашние млекопитающие составляют свыше 60% среди прирученных животных. В Европе и странах Северной Америки, так же как и в России, наибольшее количество мяса, а от рогатого скота — и молока получают от домашних животных. Ставится задача увеличения продуктивности домашних животных и выведения новых, еще более продуктивных пород.
Ценное пушно-меховое сырье поставляют многие виды зверей: соболь, куница, белка, бобр, песец, лисица, а также акклиматизированные нутрия и ондатра. Наша страна имеет богатый опыт восстановления численности видов, почти уничтоженных в результате хищнической эксплуатации в дореволюционной России. Так были восстановлены в своем прежнем ареале соболь, калан, котик. Этому способствовали организация заповедников и заказников, полный запрет добывания исчезающих животных.
В ряде стран, а также в южных районах России проводится заготовка яда змей для приготовления высокоэффективных средств (випратокс и др.), применяемых при лечении ревматизма, спазмов сосудов, бронхиальной астмы. Созданы специальные змеиные питомники, где змеи содержатся в подходящих условиях и от них периодически получают яд. Змеепитомники имеются в Узбекистане, Киргизии, для них ежегодно отлавливают сотни змей (гюрзы, эфы, гадюки, щитомордники). В связи с этим возникла проблема охраны и разумного использования последних.
Среди животных, продукты жизнедеятельности которых используются с глубокой древности, большое значение имеют медоносная пчела и тутовый шелкопряд.
Шелководство возникло около 5 тыс. лет назад в Китае и уже в средние века распространилось в Европе. Тутовый шелкопряд разводят для получения из его коконов натурального шелка. Поскольку гусеницы тутового шелкопряда питаются только листьями тутовника (шелковицы), шелководство возможно лишь в южных районах, где растет это дерево. Шелководством успешно занимаются в Средней Азии, Закавказье, на юге Украины и Молдавии.
Медоносных пчел разводят ради получения меда и воска, а в последнее время также пчелиного молочка (апилака) и прополиса — клеящего вещества. Широкое распространение получил также пчелиный яд, на основе которого готовят лекарства для лечения невралгических и сердечно-сосудистых заболеваний. Натуральный мед обладает очень ценными качествами. Это переработанный пчелами нектар цветов, обогащенный ферментами и другими полезными веществами. В его состав входят сахара (глюкоза, сахароза, фруктоза), органические кислоты, эфирные масла и витамины. Как источник углеводов мед стоит на одном из первых мест среди продуктов питания. Он обладает антибиотическими свойствами, полезен при ряде кишечных и простудных заболеваний и применяется также для лечения ран и язв. Воск употребляется в промышленности и медицине. Только в России заготовляется свыше 2 тыс. т воска в год.
Хотя экономическое значение продуктов пчеловодства очень велико, значение пчел как опылителей цветковых растений неизмеримо выше. В опылении растений принимают участие также дикие пчелы, шмели и в меньшей степени мухи и бабочки. Особая ценность шмелей как опылителей заключается в их способности посещать и опылять цветки клевера и люцерны — лучших кормовых трав, которые неохотно посещаются другими опылителями. Еще Дарвин писал о связи урожая семян клевера с наличием шмелей.
Для исследовательских и учебных целей во всех научных лабораториях разводят многих животных. Среди них есть и беспозвоночные (многие простейшие, черви-планарии, ракообразные и насекомые, особенно мушка дрозофила — излюбленный и очень удобный объект генетических исследований), и позвоночные — лягушки, голуби, крысы, золотистый хомячок, собаки, обезьяны. Известно, что большинство знаменитых опытов И. И. Павлова по изучению условных рефлексов было выполнено на собаках.
Кроме традиционных методов эксплуатации животных применяются и новые их формы. Примером этого является использование растительноядных рыб для борьбы с зарастанием водоемов и каналов. Построенный в Туркмении Каракумский канал, имеющий большое значение для орошения пустынных земель и водоснабжения населенных пунктов, очень быстро зарастал водной растительностью. Это препятствовало судоходству по каналу, уменьшало количество проходящей по каналу воды. Положение радикально изменилось после запуска в канал белого амура. Эта скороспелая и чисто растительноядная рыба взяла на себя роль природного мелиоратора и эффективно сдерживает зарастание канала, к тому же амур — еще и вкусная рыба.
Отрицательная роль животных в жизни и хозяйстве человека велика и разнообразна. К таким животным мы относим паразитов, переносчиков возбудителей болезней, ядовитых животных, вредителей сельскохозяйственных растений и продуктов хранения и т. д.
Целый ряд болезней вызывается простейшими. К ним относятся дизентерия, лямблиоз и многие другие. В большинстве случаев возбудители заболеваний попадают в организм человека через укус кровососа-переносчика. Одно из наиболее распространенных и тяжелых заболеваний человека — малярия — вызывается малярийными плазмодиями (одноклеточными из класса споровиков). Одну часть своего жизненного цикла плазмодии проходят в теле комаров. При укусе комара плазмодии попадают в кровь человека. Через укусы москитов человек заражается кожным лейшманиозом (пендинкой). Сонная болезнь вызывается паразитированием в человеке трипаносом (простейших из класса жгутиконосцев), а попадают они в кровь при укусах мухи цеце.
Хранителями и переносчиками возбудителей природно-очаго-вых болезней (вирусного клещевого энцефалита, клещевого возвратного тифа и др.) служат кровососущие, клещи — иксодовые и аргасовые. Возбудителей сыпного и возвратного тифа переносят вши, а основными переносчиками чумного микроба являются блохи.
Многие из перечисленных заболеваний имеют природно-оча-говый характер, т. е. приурочены к определенным районам или частям земного шара. Они передаются от больных животных (доноров) к здоровым (реципиентам) переносчиками — кровососущими насекомыми или клещами. Такие природные очаги существуют долгие годы. Учение о природной очаговости трансмиссивных заболеваний развил советский ученый академик Е. Н. Павловский.
Ощутимый вред здоровью человека (и домашних животных) наносят паразитические черви — гельминты. Паразиты истощают своих хозяев, отравляют их токсическими выделениями, что нередко приводит к тяжелым последствиям, а иногда и к смерти. Изучением биологии и жизненных циклов паразитических червей занимается наука гельминтология. В России крупнейшим гельминтологом был академик К. И. Скрябин. Он же предложил и основные методы профилактики борьбы с гельминтозами.
Среди животных немало ядовитых, укусы которых опасны для людей. Это ядовитые пауки — тарантул и каракурт, скорпионы, крупные сколопендры, жалящие осы и особенно ядовитые змеи. В нашей стране к ядовитым змеям относятся кобра, гюрза, эфа, несколько видов гадюк и щитомордники. Большинство из них встречаются только на юге России. В тропиках ядовитых змей гораздо больше, от их укусов погибают ежегодно несколько тысяч человек. Кожные выделения жаб также ядовиты и вызывают местное раздражение или общее отравление организма. Опасны и ожоги стрекательных батарей многих медуз и сифонофор.
Чрезвычайно существенный вред причиняют животные (насекомые, клещи, грызуны) сельскохозяйственным растениям. Практически каждая сельскохозяйственная культура имеет набор специфических вредителей. Многие из них вредят целому ряду растений, являясь многоядными. Такова, к примеру, саранча. Особые убытки причиняют вспышки массового размножения вредных насекомых. Только в США ущерб от вредных насекомых исчисляется ежегодно 4 млрд долларов. Изучением биологии, вредоносности насекомых и разработкой мер борьбы против них занимается специальная наука — сельскохозяйственная энтомология.
В наше время разработан ряд мероприятий по регуляции численности вредных животных. Широкое развитие получил химический метод борьбы, при котором применяются стойкие и высокотоксичные ядохимикаты. К сожалению, использование пестицидов зачастую приводит к нежелательным побочным результатам — истреблению наряду с вредителями и полезной фауны, а также к загрязнению среды обитания. Включаясь в круговорот веществ в биосфере, пестициды усваиваются растениями, накапливаются в тканях растительноядных животных и попадают в пищевые продукты человека. Поэтому ученые большинства развитых стран давно работают над проблемой ограничения химической борьбы с вредителями и замены ее биологическими методами. Биологическая борьба с вредителями предусматривает применение против них хищных, паразитических или патогенных организмов. Метод этот экологичен: он основан прежде всего на природных связях в биоценозе, таких как отношения хищника с жертвой или паразита с хозяином. Кроме того, такой метод не вносит изменений в круговорот веществ в биосфере, не загрязняет среду обитания человека.
Этот краткий обзор многообразных связей животных с человеком и его хозяйством показывает, что зоология является одной из самых актуальных биологических наук, так как ее данные и выводы широко применяются в медицине, ветеринарии, агрономии, животноводстве и другой хозяйственной деятельности человека.
Огромная роль животных в жизни биосферы заставляет обратить пристальное внимание на проблему охраны животного мира. Расширение масштабов использования природных ресурсов в эпоху научно-технической революции привело ко многим нежелательным последствиям. Самые тяжелые из них — разрушение природных экосистем (изменение среды обитания) и загрязнение биосферы — воздуха, воды и почвы. Все это нанесло большой, а в ряде случаев непоправимый ущерб животному миру. Ученых беспокоит исчезновение под влиянием человека некоторых видов животных, а также неуклонное падение численности и сокращение ареалов других видов. Подсчитано, что за последние 300 лет с лица Земли исчезло 63 вида млекопитающих и 94 вида птиц. О представителях других классов нам ничего неизвестно, так как их исчезновение прошло незаметно.
Международный союз охраны природы составил так называемую Красную книгу. В нее занесены виды животных, численность которых стала угрожающе низкой. Они обречены на гибель, если не будут приняты надлежащие меры охраны. Красная книга — это сигнал бедствия! Выпущены также Красные книги России и ряда регионов.
В России уделяется большое внимание сохранению животного мира. Уже первыми декретами в 1919 и 1922 гг. были взяты под полную охрану такие звери, как лось, зубр, сайгак, пятнистый олень, бобр и др. В настоящее время в России запрещена охота на многие виды зверей и птиц. Организованы многочисленные заповедники и заказники. Результаты охраны животных можно проиллюстрировать на примере соболя и сайгака, численность которых увеличилась соответственно в 12 и в 1000 раз. Тем не менее другие виды животных из-за сокращения их местообитаний или нарушения равновесия экосистем, в которые они входят, находятся под угрозой. Надо приложить еще немало усилий к охране животного мира нашей страны.
Зоология — одна из классических биологических наук. Ее зарождение, не считая первоначального накопления сведений о животных, связано с античными временами. Великий ученый и мыслитель Древней Греции Аристотель, считающийся родоначальником ряда наук, в IV в. до н. э. впервые систематизировал накопленные знания о животных и разделил все известные ему виды на две группы — животных, имеющих кровь, и животных без крови. К первой группе им были отнесены позвоночные животные (звери, птицы, земноводные, пресмыкающиеся, рыбы), ко второй — беспозвоночные (насекомые, пауки, раки, моллюски, черви). Аристотель впервые выдвинул идею о соподчинении частей организма, которая много позже будет воплощена в учении о корреляциях.
Эпоха Римской империи оставила нам многотомный труд Пли-ния Старшего (23—79 гг. н. э.) «Естественная история», в которой два тома посвящены живым организмам. Правда, большей частью это были сведения, почерпнутые из работ Аристотеля.
Падение Римской империи и установление господства христианской церкви привели к упадку наук. В эту эпоху, получившую название средневековья, занятие естественными науками не только не поощрялось, но прямо преследовалось. Признавались лишь библейские догмы о сотворении мира.
Накопление зоологических знаний возобновляется только в последовавшую за средневековьем эпоху Возрождения, с XV в. Ученых интересовало главным образом строение организма, поэтому наибольшие успехи были достигнуты в области анатомии. Знаменитый художник и ученый Леонардо да Винчи (1452—1519), изучая кости и суставы, установил сходство в строении костей ноги лошади и человека, несмотря на их внешнюю непохожесть. Тем самым он открыл явление гомологии, которое в дальнейшем объединило многих внешне различных животных и помогло заложить основу теории эволюции.
Своего расцвета природоведение эпохи Возрождения достигло в трудах швейцарца Конрада Геснера (1516—1565), сообщившего много сведений о животных, хотя зачастую не оригинальных, а почерпнутых из трудов древних ученых. В XVI—XVII вв. большой вклад в изучение анатомии животных, а также человека внесли врачи. Крупнейшим анатомом эпохи Возрождения был Андреас Везалий (1514—1564), опубликовавший первую наиболее точную работу по анатомии человека. Габриеле фаллопий (1523—1562) изучал органы размножения. Ему принадлежит описание труб, идущих от яичников к матке. Бартоломео Эустихио (1510—1574) открыл трубу, соединяющую ухо с горлом. Изучая кровообращение, Уильям Гарвей (1578—1657) обнаружил существование в сердце односторонне действующих клапанов и доказал, что кровь течет по венам в сердце и затем поступает в артерии, т.е. постоянно движется в одном направлении. Книга Гар-вея «Анатомическое исследование о движении сердца и крови у животных» (1628) вызвала полный переворот в зоологии.
Огромное значение для развития зоологии имело изобретение микроскопа. Голландец Антон Левенгук (1632—1723) при помощи изготовленного им микроскопа дал первое описание кровяных телец и капилляров, его помощник первым увидел сперматозоиды, но главным было открытие простейших, сделанное при рассматривании под микроскопом капли воды. В этот же период английский ученый Роберт Гук (1635—1703) выполнил ряд тонких микроскопических работ и в 1665 г. опубликовал книгу «Микро-графия», в которой впервые в истории биологии была изображена клетка. Это открытие имело важные последствия.
В конце XVII — первой половине XVIII в. были заложены основы систематики животного мира. Первую попытку в этом направлении сделал английский натуралист Джон Рей (1628—1705). В книге «Систематический обзор животных», вышедшей в 1693 г., Рей предложил классификацию животных, базирующуюся на совокупности внешних признаков, к примеру, по наличию когтей и зубов. Так, млекопитающих он разделил на две группы: животных с пальцами и животных с копытами. Последние, в свою очередь, были разделены на однокопытных (лошадь), двукопытных (корова) и трехкопытных (носорог). Были выделены и более дробные единицы.
Несмотря на несовершенство классификации Рея, принцип, положенный в ее основу, получил развитие в трудах знаменитого шведского ученого Карла Линнея (1707—1778). В 1735 г. Линней опубликовал книгу «Система природы», в которой изложил свою классификацию растений и животных. Он по праву считается основателем систематики, изучающей классификацию видов живых организмов. Близкие виды Линней группировал в роды, близкие роды — в отряды, а близкие отряды — в классы. Все известные виды животных были сгруппированы в 6 классов: млекопитающие, птицы, амфибии (объединивший пресмыкающихся и земноводных), рыбы, насекомые и черви. Каждый вид у Линнея имел двойное латинское название: первое слово в нем — название рода, второе — вида. Форма бинарной (двойной) номенклатуры сохранилась до сих пор. Линней стоял на позиции неизменяемости видов, хотя в конце концов был вынужден допустить возможность образования новых видов путем гибридизации.
В конце XVIII — начале XIX в. французский зоолог Жорж Кювье (1769—1832) разработал основы сравнительной анатомии животных и, в частности, учение о корреляциях. Кювье был основоположником палеонтологии. На основе этих работ в 1825 г. Анри Бленвиль ввел в систему понятие «тип» — высшую таксономическую единицу.
Французский биолог Жорж Бюффон (1707—1788) высказал идею изменяемости видов под влиянием окружающей среды. Бюффон — автор 44-томной энциклопедии «Естественная история»;
он установил наличие у животных рудиментарных органов, которые были когда-то нормально развитыми.
Другой французский естествоиспытатель, Жан Батист Ламарк (1744—1829), посвятил себя детальному изучению исторического развития живой природы. Он впервые ввел в употребление термины «беспозвоночные» и «позвоночные животные», много работал над систематизацией беспозвоночных, среди которых различал уже 10 классов, и в 1815—1822 гг. опубликовал большой труд «Естественная история беспозвоночных животных». В процессе таксономических работ ему неоднократно приходилось задумываться над возможностью эволюционного процесса. Его главный труд «Философия зоологии» (1809) посвящен изложению научной теории эволюции животного мира. Ламарк считал, что организмы меняются под прямым воздействием среды и приобретенные признаки наследуются, однако ему была чужда идея естественного отбора.
Против идеи неизменяемости видов в этот же период выступили русские ученые К. Ф. Рулье (1814-1858) и К.М.Бэр (1792-1876). Рулье призывал изучать животных в их естественном окружении и во взаимодействии со средой обитания. Его по праву можно считать провозвестником экологии. К. М. Бэр — автор выдающихся исследований в области эмбриологии животных, создатель учения о зародышевых листках.
Значительное влияние на развитие зоологии оказала сформировавшаяся в конце 30-х годов XIX в. клеточная теория. Ее создатели - М. Шлейден (1804-1881) и Т. Шванн (1810-1882). Эта теория убедительно показала единство живых организмов на клеточном уровне.
С выходом в свет знаменитого труда Чарлза Дарвина (1809— 1882) «Происхождение видов» (1859) начинается новый период в развитии биологии в целом и зоологии в частности. В книге Дарвина изложено эволюционное учение и определен важнейший фактор эволюции — естественный отбор.
Идеи Ч.Дарвина стали использоваться зоологами для разработки истории животного мира. Наибольший вклад в развитие филогении животных в XIX в. внесли такие ученые, как Э. Геккель (1834-1919) и Ф.Мюллер (1821-1897). Последний, будучи эмбриологом, установил закономерности во взаимоотношениях индивидуального развития (онтогенеза) и филогенеза животных. В 1866 г. Э. Геккель сформировал свой «биогенетический закон», согласно которому зародыши в процессе развития повторяют в сокращенном виде эволюционный путь, пройденный их предками («онтогенез повторяет филогенез»).
Доказательства эволюции, приведенные Ч.Дарвином, возбудили большой интерес к сравнительному изучению различных групп животных, в связи с чем возникают такие науки, как эволюционная сравнительная анатомия и эволюционная сравнительная эмбриология. В создании последней ведущая роль принадлежит русским зоологам И.И.Мечникову (1845—1916) и А.О.Ковалевскому (1840—1901). Выводы сравнительной эмбриологии, основанные на эволюционном учении, служили веским доказательством в пользу единства происхождения всех типов животного царства. Уже в начале XX в. было детально выяснено эмбриональное развитие большинства типов животных. В это же время В.О.Ковалевский (1842—1883) работами по ископаемым копытным заложил основы эволюционной палеозоологии. Чрезвычайно быстро развиваются систематика и зоогеография. Еще в додарви-новские времена Н. А. Северцов (1827—1885) установил связь между особенностями фауны и физико-географическими условиями, в которых развивается эта фауна. Тем самым была заложена основа экологической зоогеографии.
Вторая половина XIX в. отмечена появлением новой науки — экологии. Русские зоологи сформулировали многие главные положения и методические принципы теоретической экологии. Московский профессор К. Ф. Рулье одним из первых показал значение изучения животных в сообществе с другими организмами и фактически сформулировал понятие о популяции. В конце XIX — начале XX в. были проведены обширные исследования, в которых применялись экологические принципы при разработке проблем в области охотничьего хозяйства и борьбы с вредителями (М.Н.Богданов, Л.П.Сабанеев, А. А. Силантьев, Б.М.Житков и др.).
В XX в. зоология развивалась чрезвычайно активно. Здесь мы отметим кратко только вклад отечественных ученых. В XX в. проводились основные исследования фауны Мирового океана. Фундамент наших знаний о зоогеографии северных морей заложил К. М. Дерюгин, а картину состава и биоценотического распределения данной фауны Черного моря дал в классическом труде «К вопросу об изучении жизни Черного моря» (1913) С.А.Зер-нов. Экспедиционными судами «Витязь» (Россия) и «Галатея» (Дания) изучены глубины Мирового океана до 11 тыс. м и сделаны выдающиеся зоологические открытия. Эти работы продолжает научно-исследовательский флот Российской академии наук. К числу замечательных открытий следует отнести находку «живого ископаемого» — моллюска из класса моноплакофор, расшифровку систематического положения и установление нового типа морских животных — погонофор (А. В. Иванов) и многие другие.
Отечественные зоологи продолжали разработку проблем филогенеза животных на основе сравнительной анатомии и эмбриологии (В. Н. Беклемишев, П. П. Иванов, В. А. Догель, А. В. Иванов, Н.А.Ливанов, И. И. Шмальгаузен, Д. М. Федотов). Большое внимание уделялось изучению паразитических животных — простейших, червей и членистоногих. Научные школы В. А. Догеля, К. И. Скрябина и Е. Н. Павловского внесли большой вклад в разработку учения о природной очаговости трансмиссивных заболеваний, гельминтологии, экологической паразитологии.
Очень велик объем выполненных нашими учеными энтомологических работ. Насекомые — крупнейшая группа во всем животном царстве. Среди них много вредных видов, переносчиков заболеваний человека и домашних животных, но имеется немало и полезных — опылителей цветковых растений, производителей ценных продуктов (мед, шелк, воск). В области энтомологии велик вклад таких ученых, как А. А. Штакельберг, А. С. Мончадский, Г. Я. Бей-Биенко, С. И. Медведев, О. Л. Крыжановский, Г. С. Медведев. Большое значение имели почвенно-экологические исследования научной школы академика М. С. Гилярова.
При сравнении представителей разных систематических групп создается впечатление, что они необычайно разнообразны. Тем не менее различия животных не бесконечны.
Как было показано Ч. Дарвином, множество родственных групп животных произошло от одной предковой линии. «Спускаясь» от кончиков ветвей родословного древа животных к узлам ветвления и в конечном итоге к стволам, мы улавливаем общность многих организмов в их планах строения. Ученые установили несколько таких планов, в которые укладывается большое число вариантов. Следует помнить, что план строения — это то общее, что характерно для множества групп. Варианты же — это частности, детали, которые первыми бросаются в глаза и нередко маскируют принадлежность животного к определенному типу. Общность планов строения указывает на гомологию — сходство, основанное на родстве организмов.
За немногими исключениями, животные отличаются симметричным строением. Различают два типа симметрии — радиальную, или лучевую, и билатеральную, или двустороннюю. Оба этих типа одновременно встречаются только у беспозвоночных животных. Позвоночные — всегда билатеральны.
В теле радиально-симметрич-ного животного (рис. 1) можно различить главную продольную ось, вокруг которой в радиальном (лучевом) порядке расположены органы.
 Порядок радиальной симметрии зависит от числа повторяющихся органов. Если вокруг этой воображаемой главной оси расположено 5 одинаковых органов, то симметрия называется пятилучевой, если 4 — четырехлучевой и т. д. Вследствие этого через тело животного (его центр) можно провести строго определенное Порядок радиальной симметрии зависит от числа повторяющихся органов. Если вокруг этой воображаемой главной оси расположено 5 одинаковых органов, то симметрия называется пятилучевой, если 4 — четырехлучевой и т. д. Вследствие этого через тело животного (его центр) можно провести строго определенное
 число плоскостей симметрии, которыми тело делится на две половины, зеркально отображающие друг друга. Радиальная симметрия имеет две разновидности: радиально-лучевую и радиаль-но-осевую симметрии. число плоскостей симметрии, которыми тело делится на две половины, зеркально отображающие друг друга. Радиальная симметрия имеет две разновидности: радиально-лучевую и радиаль-но-осевую симметрии.
Радиально-лучевая симметрия наблюдается у многих организмов, взвешенных в воде (ряда одноклеточных, а также колониальных одноклеточных и некоторых колоний многоклеточных), у которых со всех сторон среда обитания одинакова.
Радиально-осевая симметрия наблюдается у нескольких групп беспозвоночных (кишечнополостных, иглокожих и др.), которые характеризуются тем, что ведут (или их предковые формы вели) прикрепленный образ жизни. Значит, сидячий образ жизни способствует развитию лучевой симметрии (Догель, 1981). Биологическое объяснение такого строения заключается в следующем. Сидячие животные одним полюсом (аборальным) прикрепляются к субстрату, другой же полюс (оральный), на котором находится ротовое отверстие, свободен. Этот полюс со всех сторон поставлен в идентичные условия по отношению к факторам окружающей среды. Поэтому различные органы одинаково развиваются на радиально расположенных частях тела, а главная ось соединяет оба полюса.
Билатеральная симметрия тела животного характерна тем, что через его тело можно провести только одну плоскость симметрии, делящую его на две равные (зеркально отражающие друг друга) половины — левую и правую. Двусторонняя симметрия возникла у животных при переходе их планктонных предков к жизни и передвижению на дне. При этом кроме переднего и заднего конца тела у них стали различаться спинная (дорсальная) и брюшная (вентральная) стороны. Примерами билатерально-симметричных животных могут служить черви, членистоногие, все хордовые животные, в том числе и человек.
Биологическое объяснение билатеральности заключается в следующем.
При переходе к ползающему (на дне) образу жизни две стороны животного — брюшная и спинная — попадают в разные условия по отношению к факторам среды. Один конец тела становится передним и к нему сдвигается ротовое отверстие, а также органы чувств. Это и понятно, поскольку при движении этот конец первым встречает источники раздражения. Главная ось тела проходит от переднего полюса, на котором оказывается рот, до заднего, где расположено анальное отверстие. Боковые части находятся в равном положении. Единственную плоскость симметрии можно провести, только «разрезав» животное на левую и правую половины вдоль главной оси тела.
В систематике животных используется целый ряд иерархически соподчиненных систематических категорий — таксонов. Основным таксоном является вид,
для обозначения которого используется латинское название из двух слов. Первое из них обозначает род,
к которому относится этот вид, а второе — название вида. Род объединяет группу родственных видов. Роды объединяются в семейства,
а далее по восходящей линии по такому же принципу следуют отряд, класс, тип.
Для обобщения сходств и различий между большими группами животных используются также таксоны более высокого уровня, чем тип, — раздел
и царство.
Кроме того, применяются дополнительные категории с приставками под- и над- (подвид, надкласс, надсемейство, надотряд и т. п.) и некоторые другие (например, инфракласс, триба и пр.).
Царство животных (Animalia) делится на два полцарства: одноклеточные (Protozoa) и многоклеточные (Metazoa).
Одноклеточные — это животные на клеточном уровне организации. В подавляющем большинстве они микроскопически малы, поэтому и стали известны только после изобретения микроскопа. Число современных видов свыше 40 тыс. Хотя одноклеточные состоят из одной клетки, они обладают всеми функциями организма. В теле их можно найти мембрану, ядро, цитоплазму с органоидами — митохондриями, рибосомами, — эндоплазматическую сеть, аппарат Гольджи и др., а также со специальными органеллами. К ним относятся вакуоли, фиблярные образования и др. Одноклеточным присущ жизненный цикл — совокупность всех стадий развития, которая в существовании каждого вида повторяется с определенной закономерностью. Обычно цикл начинается с зиготы, соответствующей оплодотворенному яйцу многоклеточных. За этой стадией следует однократное или несколько раз повторяющееся деление клетки — бесполое размножение. После этого образуются половые клетки — гаметы, которые сливаются попарно, снова дают зиготу, и цикл завершается. В жизненном цикле нередко наблюдается инцистирование — переход к покоящейся стадии.
Одноклеточные обитают во всех средах и имеют всесветное распространение. Среди них много паразитов животных и человека.
Подцарство разделяется на 7 типов (табл. 1).
Классификация простейших
Тип Саркомастигофоры —
Sarcomastigophora
(25000
видов)
Подтип Саркодовые — Sarcodina
Надкласс Корненожки — Rhizopoda
Надкласс Актиноподы — Actinopoda
Класс Акантарии — Acantharea
Класс Полицистинеи — Polycystinea
Класс Феодарии — Phaeodaria
Класс Солнечники — Heliozoa
Подтип Жгутиконосцы — Mastigophora
Класс Растительные жгутиконосцы — Phytomastigophorea
Класс Животные жгутиконосцы — Zoomastigophorea
Подтип Опалины — Opalinata
Класс Опалины — Opalinatea
Тип Апикомплексы —
Apicomplexa
(4800
видов)
Класс Перкинсеи — Perkinsea
Класс Споровики —Sporozoea
Подкласс Грегарины — Gregarinia
Подкласс Кокцидии — Coccidia
Подкласс Пироплазмы — Piroplasmia
Тип Миксоспоридии —
Myxozoa
(875 видов)
Класс Миксоспоридии — Myxosporea
Класс Актиноспоридии — Actinosporea
Тип Микроспоридии —
Microspora
(800
видов)
Тип Асцетоспоридии —
Ascetospora
(30
видов)
Тип Лабиринтулы —
Labirinthomorpha
(35 видов)
Тип Инфузории —
Ciliophora
(7500
видов)
Класс Ресничные — Ciliata
Класс Сосущие — Suctoria
Таблица 1.
Сравнительная характеристика типов простейших
| Типы
|
Органеллы
движения
|
Ядра
|
Половой процесс
|
Споры
|
Образ жизни
|
| I.Sarcomastigophora |
жгутики, псевдоподии |
одинаковые, редко дуализм |
копуляция |
нет |
свободно-ивущие, реже паразиты |
II. Apicomplexa
|
жгутиковые
гаметы
|
одинако
вые и много
ядерные
|
тоже
|
споры со спорозоитами, без стрекательных
капсул
|
паразиты
|
III. Myxozoa
|
нет
|
ядерный
дуализм,
много
ядерные
|
автогамия
|
многоклеточные
споры с амебоидными зародышами и стре
кательными капсулами
|
тоже
|
IV. Microspora
|
нет
|
одинаковые одноядерные
|
автогамия
|
одноклеточные споры с амебоид-ными зародышами и стрекательной
нитью
|
паразиты
|
V. Asceto-
spora
|
тоже
|
одинаковые много
ядерные
|
—
|
Споры многоклеточные, без стрекательных капсул |
тоже
|
Окончание табл.1
| Типы
|
Органеллы
движения
|
Ядра
|
Половой процесс
|
Споры
|
Образ жизни
|
VI. Labirin-thomorpha
|
зооспоры со жгутиками
|
одинаковые в многоклеточной
структуре
|
—
|
нет
|
свободно-живущие
|
VII. Cilio-phora
|
реснички
|
ядерный
дуализм, много
ядерные
|
конъюгация
|
тоже
|
свободноживущие, редко
паразиты
|
В данном учебнике рассматриваются подробно только три наиболее обширных типа простейших: саркомастигофоры, апикомп-лексы и инфузории.
(
SARCOMASTIGOPHORA)
К типу саркомастигофор относятся как свободноживущие, так и паразитические одноклеточные, двигающиеся при помощи непостоянных выростов цитоплазмы — псевдоподий или жгутиков. Иногда эти органеллы присутствуют одновременно. В типе три подтипа — саркодовые, жгутиконосцы и опалины.
К подтипу саркодовых принадлежат одно- или многоядерные одноклеточные, органеллами движения которых служат ложноножки, или псевдоподии. Форма тела меняющаяся, иногда постоянная. Многие обладают скелетом из минерального или органического вещества. Обитают в морях, пресных водах, почве, некоторые — паразиты (табл. 2). Общее число видов — около 10 тыс. В подтипе два надкласса: Корненожки и Актиноподы.
Форма тела корненожек разнообразна, псевдоподии подвижны, цитоплазма без дифференцировки на постоянные зоны. Над-класс объединяет 8 классов, в том числе амебы (Lobosea). Этот класс включает два подкласса: голые амебы (Gymnamoebia) и раковинные амебы (Testacealobosea).
У голых амеб тело голое, псевдоподии в виде лопастей (лободий), ядро одно или их несколько. Пища захватывается в любой части тела амебы (рис. 2).

Рис. 2. Амеба протей (Amoebaproteus):
1 — ядро; 2 —
сократительная вакуоль; 3 —
пищеварительная вакуоль
Бесполое размножение происходит делением надвое. Паразитические виды обитают в кишечнике хозяев. Наиболее патогенна дизентерийная амеба (Entamoebahystolytica), живущая в толстых кишках человека и вызывающая изъязвление слизистой и понос с выделением крови. Распространяется при помощи цист, выходящих с фекалиями наружу.
Раковинные амебы (рис. 3) имеют нередко инкрустированную песчинками наружную раковинку, сложенную органическими веществами, из устья раковинки выдвигаются лопаете- или нитевидные псевдоподии. Они способны инцистироваться и в таком виде распространяются ветром. Живут в пресной воде, нередко в торфяных болотах.
Корненожки фораминиферы (отряд Foramini-ferida), которых часто считают особым классом (Foraminifera), обладают одно- или многокамерной раковинкой из органического вещества — псевдохитина, выделяемого эктоплазмой. Раковинка часто пропитана углекислым кальцием или инкрустирована песчинками и кроме устья имеет множество пор. Псевдоподии в виде длинных переплетающихся нитей (ризоподии). Жизненный цикл сложный, с чередованием бесполой и половой форм размножения (рис. 4). Половое размножение с промежуточной редукцией, так что одно поколение гаплоидно, а другое — диплоидно (единственный случай в животном царстве). Фораминиферы играют важную роль в образовании земной коры, поскольку из их скелетов слагаются толщи осадочных пород.
Рис. 3. Разные виды раковинных корненожек:
А —
Arcella; Б —
Difflugia; В —
Euglypha — раковинка; Г —
Euglypha с псевдоподиями; 1 — псевдоподии; 2 —
ядро

Рис. 4. Жизненный цикл фораминиферы Elphidiumcrispa:
слева внизу — выход зародышей, образовавшихся в результате бесполого размножения; сверху — выход гамет и их копуляция
Актиноподы, за исключением большинства представителей класса солнечников, имеют минеральный внутренний скелет, что придает телу геометрически правильную форму. Пористая центральная капсула содержит ядро и внутрикапсулярную плазму. Псевдоподии радиальные, нитевидные, нередко переплетающиеся, а некоторые прямые, с осевой скелетной нитью (аксоподии). Ядро одно или их много. Размножение бесполое и половое. Встречаются колониальные формы. Надкласс делится на 4 класса: акантарии (Acantharea), полицистинеи (Polycyctinea), феодарии (Phaeodarea), солнечники (Heliozoa). Первые три класса ранее объединяли в класс радиолярии (Radiolaria). Полицистинеи и феодарии имеют большое значение в образовании осадочных пород земной коры, а также для палеонтологии (руководящие ископаемые). За исключением части солнечников живут только в морях. Солнечники — это небольшая группа пресноводных и морских актинопод с шаровидным телом. Морские виды имеют сферический скелет из кремнезема. Цитоплазма солнечников четко разделяется на зоны экто- и эндоплазму. Псевдоподии у них неветвящиеся, с осевыми скелетными нитями (аксоподии). В эндоплазме одно или несколько ядер. Размножение у солнечников бесполое и половое.
Таблица 2.
Сравнительная характеристика некоторых групп подтипа саркодовых
| Надкласс Rhizopoda |
Класс,
подкласс
|
Скелет |
Органеллы
движения
|
Размножение |
Местообитание |
Образ жизни |
| Класс Lobosea Подкласс Gymna-moebia |
нет |
лободии |
бесполое |
пресные воды или паразиты |
ползающие |
| Класс Lobosea Подкласс Testacea-lobosea |
раковина органическая и часто с инкрустацией |
тоже |
тоже |
пресные воды, почва |
тоже |
| Класс Foramini-fera |
раковина известковая (СаСОз) |
ризоподии, филоподии |
бесполое и половое |
моря |
ползающие, плавающие |
| Надкласс Actinopoda |
| Класс Acantharea |
10-20 игл (SrSO4
) |
аксоподии |
бесполое, редко половое |
моря |
плавающие |
| Класс Polycys-tinea |
ажурные сферы (SiO2
) |
аксоподии, фил-лоподии, ризоподии |
тоже |
тоже |
тоже |
| Класс Phaeodaria |
капсула с тремя отверстиями (SiO2
) |
тоже |
тоже |
тоже |
тоже |
| Класс Heliozoea |
редко внутренний (SiO2
) |
аксоподии |
тоже |
пресные воды и моря |
тоже |
Жгутиконосцы (рис. 5) характеризуются наличием органелл движения — жгутиков.

Жгутик — это вырост цитоплазмы, покрытый мембраной, внутри которого находятся трубчатые фибриллы. Последние продолжаются в глубь цитоплазмы и заканчиваются базальным телом, или кинетосомой. У некоторых жгутиконосцев в связи со жгутиком находится особая органелла — кинетопласт или блефаропласт, представляющая собой гигантскую митохондрию. Ядро одно или их несколько. Размножение бесполое или половое, либо только бесполое. Жгутиконосцы интересны тем, что в пределах подтипа есть автотрофы, осуществляющие на свету фотосинтез, и гетеротрофы. Подтип включает два класса: растительные жгутиконосцы (Phytomastigophorea) и животные жгутиконосцы (Zoomastigophorea).
(
PHYTOMASTIGOPHOREA)
Растительные жгутиконосцы обычно содержат хлорофилл и другие пигменты, способны к фотосинтезу и являются автотрофами. У некоторых видов хлорофилл утерян и они перешли к гетеротрофному питанию. Нередко встречаются колониальные виды (рис. 6).
Рис. 6. А —
колония Eudorinaelegans:
I
—
колония из 32 клеток; 2 —
бесполое размножение (каждая особь делится и дает начало дочерней колонии); Б —
колония Volvoxaureus с дочерними колониями внутри материнской
Ниже приводится краткая характеристика некоторых групп этого класса (табл. 3).
Таблица 3.
Сравнительная характеристика некоторых основных
отрядов растительных жгутиконосцев
| Класс
Phytomastigophorea
|
| Отряды (представители) |
Оболочка, скелет |
Жгутиковый аппарат |
Местообитание, образ жизни |
| Chrysomonadida (Mastigamoeba) |
пелликула, иногда образуют домики |
1—3 жгутика, иногда с псевдоподиями |
моря, пресные воды, автотрофы |
| Dinoflagellida (Peridinium, Notiluca) |
пелликула и панцирь из клетчатки |
2 жгутика |
моря, пресные воды, автотрофы, редко гетеротрофы |
| Prymnesiida (Coccolitho-phoros) |
пелликула и известковый панцирь |
2 жгутика и спиральный придаток — гаптонема |
моря, гетеротрофы, образуют меловые отложения |
| Euglenida (Euglena) |
пелликула |
1 жгутик |
пресные воды, автотрофы, миксотрофы |
| Volvocida (Chlamido-monas, Volvox) |
то же |
2—4 жгутика |
моря и пресные воды, автотрофы |
(
ZOOMASTIGOPHOREA)
Животные жгутиконосцы — свободноживущие или паразитические гетеротрофные формы, питающиеся сапрофитно (осмотическим путем) или заглатывая частицы пищи (анимальным способом). Подкласс делится на несколько отрядов, представители которых часто имеют практическое значение как паразиты растений, животных и человека. В таблице 4 приводится характеристика пяти важнейших отрядов.
Таблица 4.
Сравнительная характеристика некоторых отрядов
животных жгутиконосцев
| Класс
Zoomastigophorea
|
Отряды
(представители)
|
Жгутиковый аппарат |
Ядерный аппарат
(размножение)
|
Местообитание,
образ жизни
|
| Choanoflagellida (Codosida, Sphaeroeca) |
1 жгутик с воротничком из микроворсинок |
1 ядро, есть колониальные (бесполое) |
моря, гетеротрофы — голозои |
| Kinetoplastida (Bodo, Leptomonas, Trypanosoma, Crithidia, Leishmania) |
1 жгутик, редко 2, иногда с ундулирующей мембраной, развит кинетопласт |
1 ядро (бесполое) |
пресные воды, паразиты растений и животных |
| Diplomonadida (Lamblia) |
8 жгутиков, аксостиль |
2 ядра (бесполых) |
паразиты животных и человека |
| 35Trichomonadida (Trichomonas) |
4—6 жгутиков, из них 1 с ундули-рующей мембраной, аксостиль |
1 ядро (бесполое) |
паразиты человека и животных |
| Hypermastigida (Calonympha) |
много жгутиков и аксостилей |
тоже |
симбионты термитов, тараканов |
Свободноживущие, одноядерные организмы с одним жгутиком и воротничком вокруг него. Воротничок напоминает чашечку из плотно прилегающих протоплазматических тяжей. Полового процесса нет. Одиночные или колониальные формы.
Одноядерные, свободноживущие или паразитические виды. Жгутик один или два. Кинетопласт имеется. Нередко есть ундули-рующая мембрана — тонкая плазматическая перепонка, одним краем присоединяющая жгут. Половой процесс отсутствует.
У паразитических форм наблюдается смена стадий, связанная со сменой хозяев: промастиготная — форма с одним жгутом и неглубоким расположением кинетопласта; амастиготная (лейшмани-альная) — с округлым телом, без жгута, живет внутри клеток хозяина; эпимастиготная — жгут отходит от центра клетки вблизи ядра и соединен с телом ундулирующей мембраной, живет в полостях тела хозяина; трипомастиготная (трипаносомная) — жгут отходит вблизи заднего конца позади ядра и до переднего конца клетки соединен с ее телом ундулирующей мембраной, живет в полостях тела хозяина.
Среди кинетопластид есть опасные паразиты (табл. 5).
Таблица 5.
Кинетопластиды — паразиты человека и животных
| Виды
|
Заболевание
|
Хозяин
|
Переносчик
|
Регион
|
| Trypanosoma brucei rhodesiense |
сонная болезнь |
человек |
муха цеце |
Восточная Африка |
| Т. cruzi |
болезнь Чагаса |
человек, домашние животные |
клопы |
Южная Африка |
| T.brucei brucei |
нагана |
лошадиные, жвачные |
мухи |
Африка |
| Т. evansi |
сурра |
тоже |
слепни |
Азия, Африка |
| Т. equiperdum |
случная болезнь |
лошадиные |
без переносчика при случке от другой лошади |
Средиземноморье, Азия, Африка |
| Leishmania tropica |
кожный лейшманиоз |
человек |
москиты |
Средиземноморье |
| L. donovani |
кала-азар, висцеральный лейшманиоз |
тоже |
тоже |
Азия, Южная Америка |
Трипаносомы на эпимастиготной стадии живут в кишечнике кровососущих насекомых, откуда перебираются в слюнные железы и при сосании крови позвоночных (и человека) попадают в кровяное русло, где переходят в трипомастиготную стадию. Цикл замыкается при сосании крови больного, когда трипаносомы попадают в кишечник насекомого и там переходят в эпимастиготную стадию. У лейшманий в кишечнике насекомых живет промастиготная стадия, а в крови позвоночных животных — внутриклеточная амастиготная. Некоторые трипаносомы, живущие в Африке, вызывают сонную болезнь у людей и смертельную болезнь у скота — нагану. Переносчиком является муха цеце.
Лейшманий, встречающиеся в Средней Азии, вызывают лейш-маниозы. Один вид, паразитирующий в природе на грызунах и переносимый москитами, при заражении людей вызывает так называемую пендинскую язву — кожное заболевание. Другой вид поражает собак, лис и шакалов, а у людей вызывает висцеральный лейшманиоз — опасное внутреннее заболевание.
Содержит паразитов кишечника позвоночных. При делении пополам особи не расходятся, а живут парами клеток, соединенных между собой. Каждая особь одноядерная, с 3—4 жгутиками. Полового процесса нет. Представители отряда — лямблии — имеют некоторое медицинское значение как кишечные паразиты. Заражение происходит в инцистированном состоянии.
Состоит из паразитов мочеполовых путей позвоночных (и человека). Одно- или многоядерные виды с 4-6 жгутиками, иногда с ундулирующей мембраной и опорным тяжем — аксостилем. Размножение половое. Представитель — трихомонас.
Объединяет одноядерные, многожгутиковые виды без ундулирующей мембраны. Нередко они имеют сложные внутриклеточные опорные структуры. Половой процесс есть. Полезные сожители (симбионты) кишечника термитов и некоторых тараканов, помогающие им переваривать клетчатку растительной пищи.
Опалины — крупные, обычно многоядерные паразитические жгутиконосцы, живущие в задней кишке амфибий. Питание сапрофитное, главным образом путем пиноцитоза. Тело покрыто тысячами коротких жгутиков, напоминающих реснички инфузорий. Размножение половое и бесполое — делением пополам. Гаметы сливаются в кишке хозяина. Заражение опалинами происходит при заглатывании цист головастиками. Замечательно, что жизненный цикл опалин и их хозяев — амфибий — совпадает.
Споровики — паразиты животных и человека (табл. 6). Их жизненный цикл представляет собой чередование бесполого размножения (посредством простого или, чаще, множественного деления — шизогонии), полового процесса и спорогонии. У некоторых споровиков бесполое размножение может отсутствовать. Половой процесс происходит путем копуляции гамет, в результате чего образуется зигота. Первое деление зиготы — мейоз. Зигота выделяет оболочку и в таком виде называется ооцистой. При спорогонии внутри ооцисты формируются спорозоиты, лежащие в ней свободно либо находящиеся внутри спор. Спорой называется спорозоит, покрытый собственной оболочкой. Спорозоит является рас-селительной стадией, которая служит паразиту для заражения новых хозяев. В классе три подкласса: грегарины, кокцидии и пироплазмы.
Таблица 6.
Споровики — паразиты человека и животных
Возбудитель
|
Основной хозяин
|
Промежуточный
хозяин
|
Типы
размножения в
жизнен
ном цикле
|
Фаза распространения во внешней среде
|
Инвазия
|
Gregarina
cuneata, подкласс Gregarinia
|
хуки Tenebrio
|
—
|
гаметогония, спорогония |
циста с ооцистами
|
кишечная
|
Eimeria
magna, подкласс
Coccidia
|
кролики
|
—
|
шизогония,
гаметогония,
спорогония
|
ооциста с 4 спорами
|
тоже
|
Sarcospo-
ridia,
подкласс
Coceidia
|
собаки
|
коровы, овцы
|
шизогония, гаметогония, спорогония, эндодиогения |
ооциста с
2 спорами
|
тоже
|
Toxoplasma
gondii, подкласс
Coccidia
|
кошки, собаки
|
грызуны, человек
|
тоже
|
тоже
|
тоже
|
Plasmo-
dium vivax, подкласс
Coccidia
|
комар малярий
ный
|
человек
|
шизогония,
гаметогония,
спорогония
|
—
|
инокуляция
|
Babesia
bigemina, подкласс Piroplas-
mia
|
клещ иксодовый
|
коровы
|
простое деление
|
—
|
тоже
|
Грегарины паразитируют в кишечнике и различных полостях у беспозвоночных. Тело, покрытое пелликулой (оболочкой из наружного слоя эктоплазмы — наружного слоя цитоплазмы), червеобразное или сферическое, нередко с органоидом прикрепления на переднем конце — эпимеритом. В эктоплазме часто расположены сократительные волоконца — мионемы. Сократительной вакуоли нет. Кишечные формы трехчленисты. Они состоят из эпиме-рита, переднего членика — протомерита и заднего — дейтомерита, причем ядро находится в заднем членике (рис. 7).
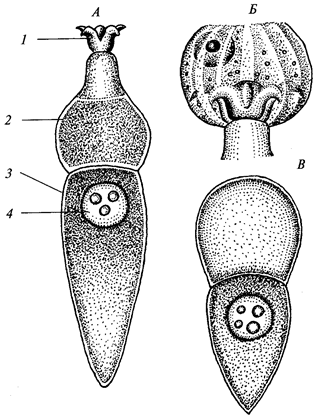
Рис. 7. Грегарина Corycellaarmata из кишечника насекомых:
А — взрослая грегарина: 1 — эпимерит; 2 —
протомерит; 3 —
дейтомерит;
4 —
ядро; Б —
эпимерит грерарины, внедрившийся в эпителиальную клетку кишечника; -8 — грегарина, сбросившая эпимерит
Половой процесс начинается соединением двух особей — гамонтов и сизигий, выделяющих общую оболочку. Под ее защитой идет образование гамет или гаметогония и их копуляция (рис. 8). У большинства грегарин шизогония отсутствует. Заражение происходит путем заглатывания ооцист.
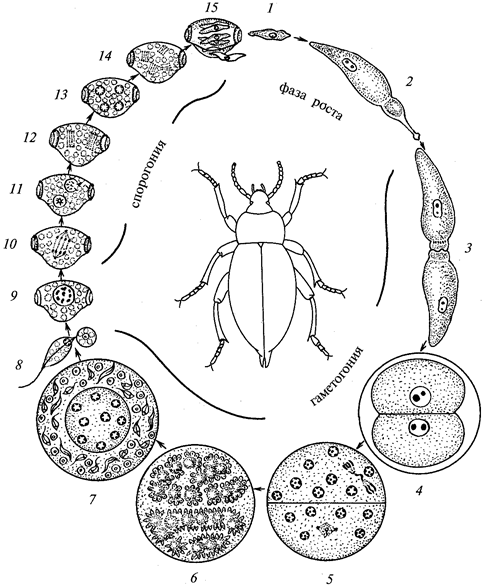
Рис. 8. Цикл развития грегарины Stylocephaluslongicollis:
1—4 —
в кишечнике жука Blapsmortisaga; / — спорозоит; 2 —
гамонт; 3 —
соединение двух особей, образование сизигия; 4 —
циста с двумя округлившимися гамонтами; 5—15 —
во внешней среде; 5, 6 —
деление ядер гамонтов, ведущее к образованию половых клеток (гамет); 7— сформированные гаметы; 8—
копуляция гамет; 9 —
зигота, образовавшая оболочку (ооциста); 10—14 —
развитие спорозоитов в ооцисте (спорогония); 15 — спорозоиты, выходящие из споры
Кокцидии — внутриклеточные паразиты позвоночных и человека. У большинства видов половое и бесполое размножение чередуется. Макрогамета образуется в результате роста гамонта без деления, микрогаметы — путем многочисленных делений гамонта (гаметоцита). Наибольшее значение имеют паразиты из отряда кокцидий (кровяных споровиков).
Включает внутриклеточных паразитов, живущих в эпителии кишечника, печени, почек и других органов. В жизненном цикле чередуются половой процесс и шизогония. У большинства видов хозяин один, и спорогония проходит во внешней среде. У некоторых кокцидий наблюдается смена хозяев, и тогда шизогония протекает в одном из них, а половой процесс и спорогония — в другом.
Примером может служить цикл развития видов рода эймерия (Eimeria) — паразитов кроликов, домашних птиц и др. В организм хозяина паразит попадает в стадии спорозоита и внедряется в клетки кишечника. Теперь их называют одноядерными ши-зонтами. Дальше происходит рост шизонта и умножение в нем числа ядер (скрытое размножение). Потом наступает деление путем шизогонии, вследствие чего тело шизонта распадается на множество одноядерных клеток — мерозоитов. Они подобны спорозои-там, но образуются в другой части цикла. Мерозоиты выходят из зараженной клетки и проникают в другие. Весь цикл шизогонии повторяется несколько раз. В результате возникает тяжелое кишечное заболевание животного.
После нескольких циклов шизогонии очередные мерозоиты образуют микро- и макрогаметоциты, которые вырастают и превращаются в макрогаметы (прямо) и микрогаметы (путем распада микрогаметоцита на большое количество подвижных гамет). Затем происходит копуляция, макрогамета становится диплоид-ной зиготой и покрывается двухслойной оболочкой, превращаясь в зигоцисту или ооцисту. На стадии зигоцисты паразиты выходят наружу и при проглатывании заражают новых хозяев.
Кокцидия токсоплазма (Toxoplasmagondii) вызывает опасное для человека заболевание — токсоплазмоз. Человек заражается при контакте с кошками, заглатывая цисты вместе с пищей или водой. В кишечнике человека, который является промежуточным хозяином, из цист выходят спорозоиты, способные затем локализоваться в любых тканях и органах, где они размножаются путем дополнительного бесполого способа — эндодиогении. В этом случае дочерние клетки образуются внутри материнской и лишь потом обособляются.
Отряд характерен тем, что в цикле развития его представителей есть смена хозяев, а стадий во внешней среде нет, и поэтому отсутствуют споры. Представители отряда — плазмодии (Plasmodium), вызывающие у человека тяжелое заболевание — малярию (табл. 7). Шизогония проходит в организме человека, гаметогония и спорогония — в комарах рода анофелес. В связи с этим комар является основным хозяином, а человек — промежуточным.
Таблица 7.
Характеристика видов рода
Plasmodium
— возбудителей малярии у человека
| Виды
|
Разновидность малярии
|
Инкубационный период
|
Приступы лихорадки
|
Симптомы
|
| Р. vivax |
трехдневная |
8-16 дней |
через 48 ч |
озноб, лихорадка, увеличение печени |
| Р. malariae |
четырехдневная |
20-35 дней |
через 72 ч |
поражение почек |
Р.
falciparum
|
тропическая |
7—12 дней |
нерегулярно |
непроходимость капилляров |
Заражение происходит при укусе комара. При этом спорозоит попадает в кровь человека и внедряется в стенки кровеносных сосудов и в клетки печени. Там он растет и размножается путем шизогонии. Образовавшиеся мерозоиты, вышедшие в кровь из клеток печени, внедряются в эритроциты, выедают их содержимое (гемоглобин) и проделывают цикл шизогонии. При выходе мерозоитов у больного человека начинается приступ лихорадки. По прошествии нескольких циклов шизогонии очередные мерозоиты превращаются в эритроцитах в макрогаметоциты. Дальнейшее развитие задерживается до тех пор, пока комар не напьется крови больного. В желудке комара, внутри сгустка крови, образуются гаметы, макрогамета оплодотворяется и возникает зигота. В отличие от других споровиков зигота плазмодия подвижна, за что и называется оокинетой («подвижное яйцо»). Оокинета выползает из кровяного сгустка, проникает через эпителиальную стенку желудка комара на ее противоположную сторону и покрывается оболочкой — ооцистой. Под этой оболочкой зигота делится — один раз путем мейоза (редукционно) и много раз митотически. В результате ооциста оказывается заполненной массой спорозоитов. Последние перебираются в слюнные железы комара и при сосании им крови очередной жертвы проникают в нее (рис. 9).
В России малярия как эпидемическое заболевание ликвидирована.
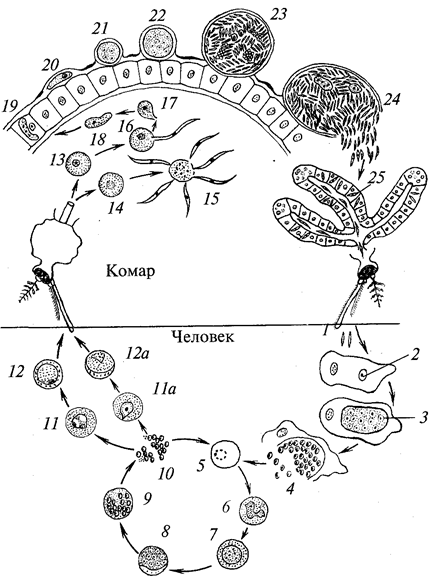
Рис. 9. Цикл развития кровяного споровика Plasmodiumvivax
(возбудителя малярии):
1 —
спорозоиты; 2—4—
бесполое размножение (шизогония) паразита в клетках печени: 2 — спорозит, внедрившийся в печеночную клетку; 3 —
растущий ши-зонт с многочисленными ядрами; 4 — шизонт, распадающийся на мерозоиты;
5-10 —
бесполое размножение (шизогония) в красных кровяных клетках (эритроцитах): 5 — молодой шизонт в форме кольца; 6 —
растущий шизонт с псевдоподиями; 7, 8 —
дальнейшие стадии роста шизонта, деление ядер; 9 —
распад шизонта на мерозоиты; 10 —
выход мерозоитов из эритроцита; 11 —
молодой макрогаметоцит; I
la
—
молодой микрогаметоцит; 12 —
зрелая макрогамета;
12а —
зрелый микрогаметоцит; 13 —
макрогамета; 14 —
микрогаметоцит; 15 —
образование микрогамет; 16 —
копуляция микро- и макрогаметы; 17 —
зиготы;
18 —
подвижная зигота (оокинета); 19 —
оокинета, проникающая через стенку кишечника комара; 20 —
оокинета, прикрепившаяся к наружной стенке кишечника комара и превращающаяся в ооцисту; 21, 22 —
растущая ооциста с делящимися ядрами; 23 —
зрелая ооциста со спорозоитами; 24 —
спорозоиты, покидающие оболочку ооцисты; 25 —
спорозоиты в слюнной железе комара
(
CILIOPHORA, или INFUSORIA)
Тип включает более 7 тыс. видов. Представители характеризуются органеллами движения — ресничками (хотя бы на некоторых стадиях развития), расположенными на поверхности клетки в определенном порядке, а также своеобразным ядерным аппаратом, состоящим из двух ядер — мелкого генеративного микронуклеуса и крупного вегетативного макронуклеуса (рис. 10). Бесполое размножение происходит путем поперечного деления и половое — конъюгацией.
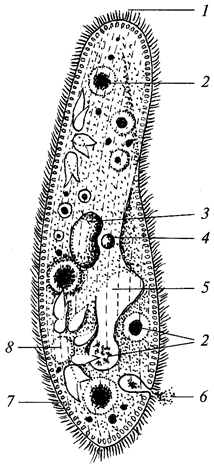
Рис. 10. Инфузория туфелька (Parameciumcaudatum):
1 —
реснички; 2 —
пищеварительные вакуоли; 3 —
большое ядро (макронуклеус); 4 —
малое ядро (микронуклеус); 5 —ротовое отверстие и глотка; 6
— непереваренные остатки пищи; 7 — трихоцисты; 8 —
сократительная вакуоль
Это наиболее сложно дифференцированные одноклеточные с постоянными цито-плазматическими структурами. Стенки клетки инфузорий имеют «альвеолярное строение» с четырьмя мембранами и полостями — «альвеолами» — посередине. Кроме того, стенка содержит трихоцисты. Это защитные приспособления в виде палочек, расположенных под пелликулой. При раздражении животного трихоцисты выстреливаются наружу.
Половой процесс сопровождается перестройкой ядерного аппарата. При конъюгации в клетках партнеров происходит распад макронуклеуса и редукционное деление микронуклеуса. Получаются четыре ядра, три из них отмирают. Оставшееся ядро делится на два пронуклеуса — мужской и женский. Между особями происходит обмен: одно ядро переходит в соседнюю клетку, другое — приходит из нее. Эти гаплоидные ядра сливаются, образуя диплоидное ядро — синкарион. Затем партнеры расходятся, а синкарионы в каждом из них путем деления создают новый ядерный аппарат. Половой процесс с такой же перестройкой ядерного аппарата может идти и без партнера. Слияние пронуклеусов в данном случае является самооплодотворением и носит название автогамии.
Свободноживущие инфузории живут в пресной и соленой воде, в толще прибрежного песка и почве. Питаются они бактериями, водорослями или одноклеточными животными. Нередки колониальные и сидячие формы. Многочисленные паразитические формы инфузорий обитают в желудке жвачных млекопитающих, на коже рыб (рис. 11), в толстых кишках человека. Инфузория балантидий (Balantidiumcoli) вызывает у людей тяжелые колиты.

Рис. 11. Жизненный цикл Ichthyophthiriusmultifiliis:
1 —
рыба, пораженная ихтиофтириазитом; 2 —
зрелый паразит из кожи рыбы; 3 —
паразит, покинувший рыбу; 4 —
циста размножения; 5 — выход «бродяжек» из цисты
В настоящее время предложено несколько вариантов классификации инфузорий. Согласно традиционной системе тип делится на два класса: ресничные инфузории и сосущие. Представители первого из них обладают ресничками в течение всей жизни, у них есть строго локализованное место приема пищи — клеточный рот. Сосущие инфузории — это сидячие формы, во взрослом состоянии не имеют рта, но зато снабжены щупальцами, служащими для ловли добычи и приема пищи. Подобно ресничным, у них два ядра (микро- и макронуклеус), и половой процесс идет по типу конъюгации. Ресничками обеспечены только стадии бесполого размножения — «бродяжки», которые отпочковываются от родительской особи и уплывают. Через некоторое время «бродяжки» садятся на субстрат, теряют реснички и образуют сосательные щупальца. Доказано, что сосущие инфузории происходят от ресничных.
В заключении раздела, посвященного простейшим, приводим характеристику важнейших заболеваний, вызываемых одноклеточными животными в России (табл. 8).
Таблица 8.
Протозойные заболевания человека в России
| Возбудитель
|
Заболевание
|
Основной хозяин
|
Промежуточный хозяин
|
Тип инвазии (инвазионная фаза)
|
| Entamoeba histolytica |
амебиаз |
человек |
— |
кишечная (циста) |
| Leishmania tropica |
кожный лейшманиоз |
человек, грызуны |
москиты |
инокуляция (клетки лейшмании) |
| Lamblia intestinalis |
лямблиоз |
человек |
— |
кишечная (циста) |
| Trichomonas vaginalis |
трихомоноз |
тоже |
— |
половым путем (жгутиковая фаза) |
| Plasmodium (vivax, falcipamm, malaria) |
малярия |
комар малярийный |
человек |
инокуляция (спорозоиты) |
| Toxoplasma gondii |
токсоплазмоз |
кошка, собака, лисица |
человек, грызуны |
кишечная (ооциста) |
| Balantidium coli |
балантидиаз |
человек, свинья |
— |
кишечная (циста) |
Таблица 9.
Сравнительная характеристика некоторых таксономических групп типа инфузорий
Особенности
|
Класс Ciliata |
Класс Suctoria
|
| Подкласс Holotricha |
Подкласс Peritricha |
Подкласс Spirotricha |
| Gymnosto-matida |
Trichosto-matida |
Hymenostomatida |
Peritri-chida |
Entodinio-morpha |
Heterotri-chida |
Hypotri-cha |
Oligotri-cha |
Представители
|
Didinium, Dileptus
|
Balantidium
|
Ichthyo-phthirius
|
Paramecium
|
Vorticella
|
Entodinium
|
Stentor, Spirostomum
|
Stylonichia
|
—
|
Dendrocometes Sphaerophraga |
Ресничный
аппарат
|
реснички
равной
длины
|
тоже
|
тоже
|
тоже
|
только
вокруг
воронки
|
реснички
разной
длины
|
тоже
|
тоже
|
тоже
|
только у «бродяжек»,
отсутствуют у взрослых
|
Околоротовые
мембра-неллы
|
нет
|
тоже
|
мембрана
и 3 мем-бранеллы
|
тоже
|
левозакрученная
спираль мембранелл
|
правозакрученная
спираль мембранелл
|
тоже
|
тоже
|
тоже
|
Образ жизни
|
пресноводные плавающие
хищники
|
паразиты
|
паразиты
|
пресноводные плавающие
|
пресноводные сидячие
|
симбионты
жвачных
|
пресноводные плавающие
|
пресноводные ползающие
|
морские плавающие
|
пресноводные сидячие
хищники
|
Подцарство характеризуется тем, что его представители во взрослом состоянии слагаются из множества клеток и их производных. Клетки дифференцированы как по строению, так и по функциям в разных направлениях и являются только частями тела. Жизненный цикл всегда имеет одноклеточные стадии (например, яйцо), в целом он отличается сложным индивидуальным развитием — онтогенезом, в процессе которого из яйца образуется взрослый организм.
В основе большинства представлений о возникновении многоклеточных лежит давнее убеждение, что они произошли от колоний Protozoa и что, следовательно, тело одноядерного простейшего в морфологическом отношении отвечает отдельной клетке многоклеточного животного. При этом полагают, что в процессе эволюции постепенно развилась новая индивидуальность многоклеточного организма, резкое подчинение ей и подавление индивидуальности отдельных клеток. Иными словами, метазоон по сравнению с простейшим признается индивидом высшего порядка. Колониальные гипотезы, таким образом, в полном соответствии с клеточной теорией рассматривают клетку как элементарную структурную единицу, позволяющую сравнивать и анализировать организацию всех Protozoa, Metazoa и Metaphyta (многоклеточных растений).
Гипотеза гастреи Э. Геккеля.
Первую гипотезу о колониальном происхождении многоклеточных — гипотезу «гастреи» — предложил Э. Геккель. В основу этой гипотезы, которую он разрабатывал с начала 70-х гг. XIX в., легла идея о гомологии зародышевых листков у всех многоклеточных животных, высказанная впервые Т. Гексли. Ко времени появления гипотезы гастреи учение о зародышевых листках сделало большие успехи благодаря работам Т. Гексли, К. Ф. Вольфа, К. Бэра и др. Геккель опирался на достижения эмбриологии своего времени и в частности на исследования А. О. Ковалевского.
Важнейшим «орудием» при создании теории гастреи был биогенетический закон, обоснованный почти одновременно Ф. Мюллером и Геккелем в 60-е гг. XIX в. Согласно Геккелю, «онтогения представляет собой краткое повторение филогении, механически обусловленное функциями наследственности и приспособляемости» (Haeckel, 1874). Он рассматривал так называемые первичные зародышевые листки — эктодерму и энтодерму гаструлы как проявление в онтогенезе соответственных примитивных органов первобытных предков. Всем прочим начальным стадиям онтогенеза Геккель также приписывал абсолютное рекапитуля-ционное значение. Все характерные стадии дробления '(рис. 12) отвечают, по мнению Геккеля, сходным стадиям филогенеза. Так, яйцу, или цистуле, соответствует одноклеточный предок Cytaea, стадии морулы — предковая форма «морея» и т. п. Особенно важной и широко распространенной в животном мире рекапитуляцией (т. е. повторением филогенеза в онтогенезе) Геккель считал двухслойную зародышевую стадию — гаструлу. Общего гипотетического прародителя всех Metazoa он создал по ее образу и подобию.
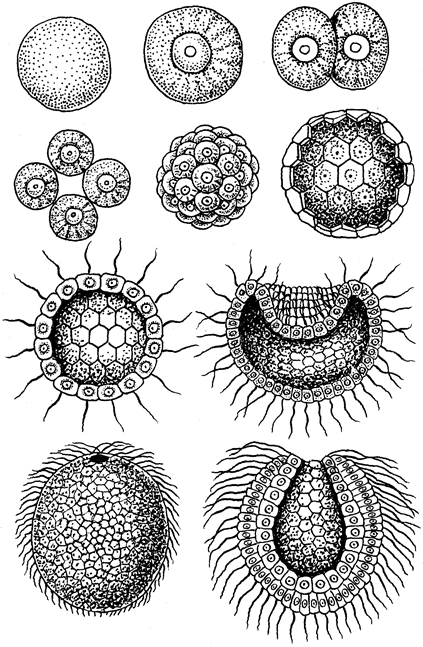
Рис.12. Стадии эмбрионального развития кораллового
полипа (по Геккелю)
Первой филогенетической стадией, по Геккелю, был одноклеточный амебообразный организм. От него произошли все ани-мально питающиеся организмы. Колония одинаковых амебоидных клеток дала затем начало «морее» — плотному шарообразному организму, рекапитуляцию которого в онтогенезе представляет морула. Путем накопления в центре морей жидкости или студенистого вещества, вытеснившего клетки на периферию, постепенно сформировалась свободноплавающая «бластея» (в онтогенезе ей отвечает бластула). Клетки бластеи сперва были покрыты псевдоподиями, которые позднее приобрели способность быстро двигаться и изгибаться и превратились в жгуты, служащие для плавания.
Следующей очень важной стадией была гастрея, образовавшаяся из бластеи путем выпячивания (инвагинации) стенки тела на переднем полюсе. Наружный клеточный слой гастреи был снабжен жгутиками и сохранил функцию движения, внутренний слой стал кишечным. В центральной, кишечной, полости, сообщавшейся ртом с наружной средой, происходило переваривание заглоченной добычи. Два эпителиальных слоя гастреи — эктодерма и энтодерма — представляли собой первичные органы, из которых у потомков гастреи возникли все их органы и ткани.
Современных кишечнополостных и губок Геккель рассматривал как мало изменившихся потомков гастреи, а стадию гаструлы — как рекапитуляцию гастреи.
Все многоклеточные, согласно Геккелю, в отличие от простейших имеют монофилетическое происхождение и развились от одной прародительской формы — гастреи, произошедшей, в свою очередь, от одноклеточных предков. От гастреи все Metazoa унаследовали первичный кишечник и два первичных зародышевых листка, их ткани представляют собой дериваты этих двух листков.
Свою гипотезу Геккель характеризовал как попытку улучшить филогенетические основы естественной классификации и выяснить пути развития главных систематических групп животного мира. Действительно, из гипотезы гастреи вытекал ряд существенных выводов для систематики, сравнительной анатомии, эмбриологии и гистологии. Однако самым важным следствием появления гипотезы гастреи было разрушение учения о типах Кювье, все еще господствовавшего в зоологии того времени.
Из гипотезы Геккеля следует, что истинные гомологии органов и их систем возможны у всех потомков гастреи, т. е. у представителей разных типов, тогда как теорией типов эта возможность отрицалась. Так как гаструла у всех Metazoa гомологична, то гомо-логичен всегда и кишечник. Гомологичны, далее, у всех Metazoa и кожные покровы, поскольку всегда имеется слой эпидермиса, служащий источником для развития других кожных слоев — кутикулы, железистых образований и пр. — и отвечающий эктодерме гастреи. Нервная система всегда развивается из эктодермы и гомологична во всех группах животных. Геккель видел также основания для гомологизации выделительных органов, целома и кровеносной системы у тех животных, у которых они имеются. Для органов чувств, скелета и сердца он считал общую гомологию неприемлемой и полагал, что все эти органы развились в разных группах независимо. Он признавал достоверным различное происхождение ротового отверстия в разных группах многоклеточных. Бластопор гаструлы, гомологичный рту гастреи, сохраняется во взрослом состоянии у кишечнополостных, у губок (в виде устья) и у низших червей. Рот иглокожих, членистоногих и позвоночных, по Геккелю, есть новообразование.
Таким образом, Геккель признавал широкие возможности конвергентного развития различных важных особенностей в строении животных.
Он считал первичной тканью однослойный жгутиковый эпителий, а все прочие ткани — вторичными производными эпителия. Эктодерму и энтодерму Геккель считал первичными зародышевыми листками. Мезодерма же, по его мнению, возникла в процессе эволюции гораздо позднее, так как в онтогенезе она всегда образуется из эктодермы и энтодермы и, по существу, даже не представляет единого листка, а имеет двойственную природу, слагаясь из пластинок, развившихся независимо из кожно-мус-кульной и кишечномускульной пластинок.
Так как мезодерма всегда развивается из парных зачатков, то, по Геккелю, у разных групп животных она имеет общее происхождение и может считаться гомологичной. Первичные зародышевые листки у низших — губок и кишечнополостных — в отличие от таковых у высших типов сохраняются как первичные органы, подобно тому, как это имело место у гипотетической гастреи.
Гипотеза Геккеля долгое время была господствующей, некоторые крупные зоологи придерживаются ее и теперь. Ее положительная роль в зоологии была чрезвычайно велика, так как она показала единство и общность происхождения всех многоклеточных и тем способствовала прогрессу дарвинизма.
Однако гипотеза гастреи страдает существенными недостатками, которые не укрылись уже от некоторых современников Геккеля и давали повод для ее резкой критики.
Действительно, гипотеза гастреи не согласуется со многими данными зоологии и должна уступить место более совершенной концепции. Впрочем, учение о протозойных колониальных предках Metazoa, лежащее в основе обобщений Геккеля, целиком сохраняет свое значение и поныне. Вторым «рациональным» зерном гипотезы гастреи следует считать учение о бластее, которое без особенных изменений было воспринято авторами некоторых других колониальных гипотез.
Известный русский эмбриолог В. В. Заленский (1874) подробно рассмотрел первые стадии эмбрионального развития различных животных с точки зрения соответствия их теории гастреи. Главнейшим моментом в онтогенезе животных он считал первую дифференциацию зародышевых листков. Общий ход рассуждении В. В. Заленского был таков. В типичных случаях у низших многоклеточных после дробления и стадии морулы формируется двуслойная бескишечная форма — планула. Если же образуется полая шаровидная бластула, то затем в ее полости появляются энтодермальные клетки и возникает стадия (дибластула), вполне сравнимая с планулой, так как она, по существу, тоже имеет два зародышевых листка и лишена эпителиальной кишки. В тех случаях, когда путем впячивания образуется гаструла с мешковидной кишкой и ртом, мы, на взгляд Заленского, имеем вторично измененное развитие, обеспечивающее очень раннее появление кишечника и характеризующееся выпадением стадии планулы. Поэтому Заленский думал, что общий предок Metazoa, скорее, должен был обладать признаками планулы, нежели гастреи. Заленский, фактически, был предшественником И. И. Мечникова, выдвинувшего известную гипотезу фагоцителлы.
Гипотеза фагоцителлы И.И.Мечникова.
Подобно Заленскому, И. И. Мечников подверг гипотезу гастреи резкой критике. В частности, он заметил, что идентичности первичной гаструлы у всех Metazoa, принимаемой Геккелем, в действительности не существует. У разных животных эта стадия имеет различные особенности и развивается по-разному, что далеко не всегда можно объяснить вторичными причинами. Истинные двуслойные, инвагинационные гаструлы, как этого требует теория гастреи, в действительности крайне редки. В законченном виде гипотеза фагоцителлы И. И. Мечникова изложена в заключительной главе его монографии «Эмбриологические исследования над медузами» (1886).
Будучи сторонником колониального происхождения, И. И. Мечников, как и Геккель, видел отдаленных предков многоклеточных в жгутиконосцах с животным питанием.
Мечников считал инвагинацию, путем которой образуется гаструла, вторичным способом образования энтодермы, возникшим в результате длительной и сложной эволюции.
Гипотеза И. И. Мечникова состоит в следующем. Первичный метазоон был шаровидным и имел сначала однослойное строение. Иными словами, признается бластея, и в этом — совпадение с гипотезой Геккеля.
Поскольку у Metazoa полость дробления обычно появляется очень рано и зародыш быстро превращается в бластулу, предком многоклеточных Мечников считал бластулообразную колонию жгутиконосцев. Он полагал, что тотальное дробление многоклеточных следует выводить из деления жгутиконосцев: первые меридиональные деления дробящегося яйца представляют собой наследие от флагелятных предков, поскольку для жгутиконосцев характерно именно продольное деление. Исходную шарообразную форму колонии Мечников пытается также объяснить исходя из продольного деления жгутиконосцев. Если деление клетки все время происходит продольно, то получается пластинка, но если третье деление изменится, станет поперечным, то в результате возникает шаровидная колония клеток. Именно такое изменение направления деления и произошло в филогенезе. Таким образом, предком Metazoa была колония, у которой направления деления чередовались в трех координатных плоскостях. Мечников думал, что образование двуслойной стадии произошло не путем впячива-ния, а путем иммиграции — внедрения отдельных клеток в полость бластулы, в результате чего образовался зачаток энтодермы. Эволюционные истоки такой иммиграции он видел в явлении фагоцитоза.
Питание первичного метазоона, по Мечникову, совершалось теми же клетками, которые служили для движения, т. е. жгутиковыми клетками путем внутриклеточного пищеварения (фагоцитоза). И. И. Мечников предположил, что перегруженные пищей клетки легко теряли жгутик и уходили в полость организма, затем они снова могли выходить на поверхность и формировать жгутик. Так получилась первая факультативная дифференциация на наружный слой клеток — «кинобласт» — и на внутреннюю клеточную массу — «фагоцитобласт». Эта дифференциация затем закрепилась в эволюции, и образовался компактный организм — паренхимелла, моделью которого он считал личинку губок — паренхимуллу. Позднее Мечников назвал этот организм фагоцител-лой. Это и был общий предок многоклеточных животных.
Дальнейшая судьба фагоцителлы такова. Некоторые ее потомки перешли к сидячему образу жизни и дали начало губкам. Другие стали ползать, приобрели билатеральную симметрию и ротовое отверстие. От
них произошли бескишечные плоские черви турбеллярии, у которых еще нет кишечника и пищеварение совершается в лакунах паренхимы и в блуждающих клетках — фагоцитах. Третьи, сохранив плавающий образ жизни, приобрели рот, испытали эпителизацию фагоцитобласта и превратились в первичных кишечнополостных — родоначальников сидячих полипов.
Таким образом, гипотеза И. И. Мечникова объясняла с эволюционной точки зрения все главные этапы онтогенеза Metazoa и предлагала новые филогенетически обоснованные представления о первичных зародышевых листках и их дальнейшей эволюции. На этой основе Мечников нарисовал вполне правдоподобную гипотетическую картину эволюционного становления Metazoa и первых этапов их филогенетического развития, картину, которая хорошо объясняет множество эмбриологических и сравнительно-анатомических факторов, непонятных с точки зрения других гипотез.
А. А. Захваткин в 1949 г. выдвинул гипотезу о происхождении многоклеточных от колониальных жгутиконосцев на основе па-линтомии — особой формы бесполого размножения путем последовательных делений клетки без стадий роста получающихся дочерних клеток. Такое деление является, по его мнению, прообразом дробления яйца у Metazoa.
Другой путь эволюционного формирования Metazoa предложил А. В. Иванов в конце 60-х гг., считавший, что гипотетические исходные колонии жгутиконосцев не были палинтомическими и вообще мало отличались от шаровидных колоний современных во-ротничковых жгутиконосцев.
Иванов принимает за основу теорию фагоцителлы Мечникова. Однако прообразом фагоцителлы он считает не личинку губок, а примитивное плоское многоклеточное трихоплакс, являющееся единственным представителем типа Пластинчатые (Placozoa). Схема возникновения многоклеточных, согласно Иванову, представлена на рис. 13.

Рис. 13. Главнейшие предполагаемые стадии филогенеза Metazoa
по А. В. Иванову:
1 — колония жгутиконосцев; Б —
миграция клеток жгутиконосцев внутрь; В —
ранняя фагоцителла; Г —
поздняя фагоцителла; Д —
первичная турбеллярия — появление рта и билатеральной симметрии; Е —
примитивная бескишечная турбеллярия — усиление дифференциации клеток, смещение рта на брюшную сторону; Ж —
примитивная губка — переход к сидячему образу жизни, замена локомоторной функции киноцитов гидрокинетической; 3 —
первичное кишеч-нополостное типа гастреи — образование рта, эпителизация фагоцитобласта
Поскольку в эмбриогенезе низших многоклеточных двуслойных зародыш образуется чаще путем иммиграции, большинство зоологов считают, что именно таким путем и шло преобразование шаровидной колонии жгутиконосцев в первый многоклеточный организм. При этом у предковых форм многоклеточных образование двух клеточных слоев сопровождалось специализацией клеток и колония жгутиконосцев превратилась в единый многоклеточный организм. Наружный слой сохранил двигательную и чувствительную функции, а внутренний — пищеварительную и половую.
Многоклеточных животных делят на три надраздела: фагоци-теллообразные
с единственным типом пластинчатых, паразои
с одним типом губок и эуметазои,
или настоящие многоклеточные.
Фагоцителлообразные, включающие только два вида мелких плоских морских животных, представляют собой наиболее примитивных многоклеточных. Их тело не имеет определенных очертаний, как у амебы, у них нет ни переднего, ни заднего конца и очень слаба дифференциация клеток. Многие зоологи считают их близкими по организации к первым многоклеточным животным.
В отличие от фагоцителлообразных, у паразоев существует ясная дифференциация клеток, но нет настоящих тканей и органов.
Дифференцированные ткани и органы появляются у эуметазоев, которые, в свою очередь, делятся на два раздела — лучистых (кишечнополостные и гребневики) с радиальной симметрией тела и билатеральных — двусторонне-симметричных животных. В таблице 10 приведена сравнительная характеристика многоклеточных, не относящихся к билатеральным.
Классификация билатеральных животных базируется на понятии о полости тела, представляющей собой пространство между стенкой тела и кишечником. У плоских червей это пространство занято паренхимой и полости тела, по существу, нет. У других же билатеральных эта полость есть. В одном случае она заполнена жидкостью и лишена собственной эпителиальной выстилки. Таковы, например, круглые черви, особый тип, относящийся к первич-нополостным животным. В другом случае полость тела имеет собственные клеточные стенки, которые формируются в онтогенезе из третьего зародышевого листка, или мезодермы. Эту полость называют вторичной, или целомом. Все высшие являются целомическими, или вторичнополостными, животными.
Таблица 10.
Сравнительная характеристика трех низших типов многоклеточных
| Признаки
|
Надраздел
Phagocytellozoa
Тип
Placozoa
|
Надраздел
Parazoa
Тип
Porifera
|
Надраздел
Eumetazoa
Тип
Coelenterata
|
| Зародышевые листки |
отсутствуют |
оформление зародышевых листков; инверсия листков в онтогенезе |
два зародышевых листка: эктодерма и энтодерма |
| Симметрия |
асимметрия |
осевая с гетеро-полярной осью и асимметрия |
радиальная симметрия с гетеро-полярной осью |
| Типы вегетативных клеток |
жгутиковые (кинобласт) и веретеновидные (фагоцитобласт) |
пинакоциты, хоаноциты, амебоидные, склеробласты, спонгиобласты |
эктодерма: эпителиально-мышечные, нервные, стрекательные; энтодерма: эпителиально-мышечные, железистые |
| Развитие личинки |
без выраженного метаморфоза; молодые стадии — «бродяжки» |
с метаморфозом; личинки: амфи-бластула, цело-бластула, паренхимула |
с метаморфозом, реже прямое; личинка: планула |
Билатеральных животных также обычно делят на два подраздела — первичноротых и вторичноротых, резко отличающихся эмбриональным развитием. У первичноротых бластопор остается ртом и у взрослого животного. Мезодерма формируется из пары клеток — телобластов зародыша.
У вторичноротых на месте бластопора образуется анальное отверстие взрослого животного, а постоянный рот закладывается независимо от бластопора и часто на противоположной стороне. Мезодерма же образуется путем выпячивания боковых стенок кишечника (энтероцельно). Ко вторичноротым принадлежат типы иглокожих, полухордовых и хордовых. Остальные типы билатеральных относят к первичноротым.
Классификация многоклеточных
Надраздел Фагоцителлообразные —
Phagocytellozoa
Тип Пластинчатые — Placozoa
Надраздел Паразои —
Parazoa
Тип Губки — Porifera, или Spongia
Надраздел Эуметазои —
Eumetazoa
Раздел Лучистые — Radialia
Тип Кишечнополостные — Coelenterata
Тип Гребневики — Ctenophora
Раздел Билатеральные — Bilateria
Подраздел Первичноротые — Protostomia
Тип Плоские черви — Plathelmintes
Тип Круглые черви — Nemathelminthes
Тип Головохоботные — Cephalorhyncha
Тип Коловратки — Rotifera
Тип Скребни — Acanthocephala
Тип Немертины — Nemertim
Тип Камптозои — Kamptozoa
Тип Кольчатые черви — Annelida
Тип Эхиуриды — Echiurida
Тип Сипункулиды — Sipunculida
Тип Членистоногие — Arthropoda
Тип Моллюски — Mollusca
Тип Онихофоры — Onichophora
Тип Язычковые, или Пятиустки — Linguatulida, или Pentastomida
Тип Тихоходки — Tardigrada
Тип Форониды — Phoronida
Тип Плеченогие — Brachiopoda
Тип Мшанки — Bryozoa
Тип Щетинкочелюстные — Chaetognatha
Тип Погонофоры — Pogonophora
Подраздел Вторичноротые ~ Deuterostomia
Тип Иглокожие — Echinodermata
Тип Полухордовые — Hemichordata
Тип Хордовые — Chordata
Следует сказать, что система типов беспозвоночных животных до сих пор окончательно не устоялась. Зоологи регулярно выделяют новые типы животных и среди уже известных групп, и в результате открытия новых животных, а также спорят о статусе прежних типов. В учебнике рассматриваются наиболее важные типы беспозвоночных.
Тип губки (
PORIFERA, ИЛИ SPONGIA)
Губки — водные сидячие многоклеточные животные. Настоящих тканей и органов нет. Нервная система у них отсутствует. Тело в виде мешка или бокала состоит из разнообразных клеток, выполняющих различные функции, и межклеточного вещества.
Стенка тела губок пронизана многочисленными порами и идущими от них каналами, сообщающимися с внутренней полостью. Полости и каналы выстланы жгутиковыми воротничковыми клетками. За немногими исключениями, губки имеют сложный минеральный или органический скелет. Ископаемые остатки губок известны уже из протерозойских пород.
Описано около 5 тыс. видов губок, большинство их обитает в морях (рис. 16). Тип делится на четыре класса: известковые губки (Calcarea), кремнероговые, или обыкновенные, губки (Demospon-gia), стеклянные, или шестилучевые, губки (Hexactinellida, или Hyalospongia) и коралловые губки (Sclerospongia). Последний класс включает небольшое число видов, которые обитают в гротах и туннелях среди коралловых рифов и имеют скелет, состоящий из массивного известкового основания из карбоната кальция и кремневых одноосных игл.
В качестве примера рассмотрим строение известковой губки. Тело ее мешковидное, основанием оно прикреплено к субстрату, а отверстием, или устьем, обращено кверху. Парагастральная область тела сообщается с наружной средой многочисленными каналами, начинающимися наружными порами.
В теле взрослой губки имеются два слоя клеток — экто- и энтодерма, между которыми залегает прослойка бесструктурного вещества — мезоглеи — с разбросанными в ней клетками. Мезоглея занимает большую часть тела, содержит скелет и кроме прочих — половые клетки. Наружный слой образован плоскими эктодермаль-ными клетками, внутренний — воротничковыми клетками — хо-аноцитами, из свободного конца которых торчит длинный жгутик. Клетки, свободно рассеянные в мезоглее, подразделяются на неподвижные звездчатые, выполняющие опорную функцию (колленциты), скелетные подвижные (склеробласты), занятые перевариванием пищи (амебоциты), резервные амебоидные, которые могут превращаться в любой из названных типов, и половые. Способность клеточных элементов переходить друг в друга говорит об отсутствии дифференцированных тканей.
По строению стенки тела и канальной системы, а также месту расположения участков жгутиконосного слоя различают три типа губок, наиболее простой из которых аскон и более сложные — сикон и лейкон (рис. 14).
Рис. 14. Различные типы строения губок и их канальной системы:
А —
аскон; Б —
сикон; В —
лейкон. Стрелки показывают ток воды в теле губки
Скелет губок образуется в мезоглее. Минеральный (известковый или кремневый) скелет состоит из отдельных или спаянных между собой игл (спикул), формирующихся внутри клеток-склеро-бластов. Органический (спонгиновый) скелет слагается из сети волокон, близких по химическому составу к шелку и образующихся межклеточно.
Губки относятся к организмам-фильтратам. Через их тело идет непрерывный ток воды, вызываемый действием воротничковых клеток, жгутики которых бьют в одном направлении — к парагастральной полости. Воротничковые клетки захватывают из проходящей мимо них воды пищевые частицы (бактерий, одноклеточных и т. д.) и заглатывают их. Часть пищи переваривается на месте, часть передается амебоцитам. Отфильтрованная вода выбрасывается из парагастральной полости через устье.
Размножаются губки как бесполым (почкованием), так и половым способом. Большинство губок — гермафродиты. Половые клетки залегают в мезоглее. Сперматозоиды выходят в каналы, выводятся через устье, проникают в другие особи губок и оплодотворяют их яйца. Зигота дробится, вследствие чего появляется бластула. Второй зародышевый слой (фагоцитобласт) образуется путем иммиграции либо инвагинации. У неизвестковых и некоторых известковых губок бластула состоит из более или менее одинаковых жгутиковых клеток (целобластула).
В дальнейшем часть клеток, теряя жгутики, погружается внутрь, заполняя полость бластулы, и в итоге возникает личинка-паренхимула.
Среди бластул губок встречаются так называемые амфибластулы, у которых анимальное полушарие состоит из мелких жгутиковых клеток, а вегетативное — из крупных клеток без жгутиков, но заполненных желтком. Амфибластулы проделывают гаструляцию в теле материнской губки: клетки вегетативного полушария впячиваются внутрь бластоцели. Однако при выходе личинки в воду энтодермальные клетки снова выворачиваются наружу (дега-струляция), возвращаясь к состоянию амфибластулы. После этого амфибластула оседает аборальным полюсом на дно, ее эктодермальные жгутиковые клетки выпячиваются внутрь, а энтодермальные остаются снаружи. Это явление называют извращением зародышевых листков. Оно наступает и в другом случае, когда личинка-паренхимула оседает на субстрат. Тогда ее эктодермальные клетки заползают внутрь, где и образуют воротничково-жгутиковые камеры. Энтодерма ложится поверх эктодермы. Устье формируется на вегетативном полюсе, который обращен вверх.
Чаще губки живут колониями, получающимися в результате недоведенного до конца почкования. Только немногие губки являются одиночными, встречаются также вторичноодиночные организмы (рис. 15). Значение их в жизни водоемов очень велико. Фильтруя через свое тело огромное количество воды, они способствуют ее очищению от загрязнений твердыми частицами.
Рис. 15. Колониальные и вторичноодиночные губки:
1 —
колония бокаловидных губок с хорошо обособленными зооидами (Sy-conciliatum); 2 —
аморфная многоустьевая губка (Mycaleochotensis); 3 — 5 —
кормусоподобные губки — промежуточная форма между многоустьевой особью и петинной колонией (Geodiaphlegraei, Chondrocladiagigantea, Phakelliacribrosa); 6, 7—
вторичноодиночные губки (Tentoriumsemisuberites, Polymastiahemisphaericum)
Таблица 11.
Сравнительная характеристика основных классов губок
Признаки
|
Классы
|
| Известковые
(Calcarea)
|
Стеклянные
(Hexactinellida)
|
Кремнероговые
(Demospongia)
|
| Скелет |
известковый |
кремниевый |
кремнероговой, роговой |
| Форма иголок |
трехосные, четырехосные, одноосные |
шестиосные и амфидиски |
четырехосные и одноосные, амфидиски |
| Образование иголок |
внеклеточное образование за счет выделения склероцитами |
внутриклеточное образование иголок (внутри склероцитов или в синцитии) |
внутриклеточное образование иголок и внеклеточное формирование спонгиновых нитей |
| Личинки |
амфибластула |
целобластула, паренхимула |
паренхимула |
| Морфологические типы |
аскон, сикон, лейкон |
лейкон |
лейкон |
| Представители |
Ascon, Sycon, Leucandra |
Euplectella, Hyalonema |
Geodia, Spongilla, Euspongia |

Рис. 16. Известковые и стеклянные губки:
1 — Polymastiacorticata; 2 —
губка морской каравай (Halichondriapanicea); 3 —
чаша Нептуна (Poterionneptuni); 4 —
байкальская губка (Lubomirskiabaikalensis);
5, 6 —
Clathrina primordialis; 7 — Pheronema giganteum; 8 —
Hyalonema sieboldi
К надразделу относятся многоклеточные, которые на всех стадиях развития сохраняют правильное положение зародышевых листков: эктодерма (кинобласт) находится снаружи, а энтодерма (фагоцитобласт) — внутри.
Кишечнополостные — это радиально-симметричные, водные (в большинстве морские), свободноплавающие или прикрепленные животные. Число видов около 10 тыс.
Представители типа — животные двухслойные. Стенка тела состоит из экто- и энтодермы, между которыми имеется прослойка мезоглеи. В простейшем случае (у одиночных полипов) тело напоминает мешок, прикрепленный одним полюсом к субстрату. На противоположном конце расположен рот, ведущий в полость кишки (гастральную полость). Вокруг рта развивается венчик щупалец, при помощи которого захватывается пища. Непереваренные остатки пищи выбрасываются через рот. Эта схема относится к сидячим формам, получившим общее название полипов.
Свободноплавающие кишечнополостные сильно уплощены в направлении главной оси тела и ротовым отверстием обращены книзу. Такие формы называются медузами.
Нередко одни и те же виды кишечнополостных на разных стадиях развития имеют строение то медузы, то полипов. Медузы всегда одиночны, полипы же часто образуют колонии, состоящие из множества особей. При этом особи могут быть одинаковыми по строению и выполнять идентичные функции (мономорфные колонии) или же сильно отличаться друг от друга этими признаками (полиморфные колонии).
Для представителей типа характерно наличие в покровах тела стрекательных клеток, служащих для защиты и нападения. Размножение половое и бесполое (чаще почкованием). У многих наблюдается чередование поколений. Тип делится на три класса — гидрозои, сцифоидные и коралловые полипы.
У гидроидных полипов тело цилиндрическое или яйцевидное, в нижней части которого обычно имеется ножка для прикрепления к субстрату или (у колониальных форм) для соединения с колонией. Гастральная полость у гидрозой без перегородок. Мезоглея с малым количеством клеточных элементов. Половые железы (гонады) формируются в эктодерме. В цикле развития наблюдается чередование полипоидного и медузоидного поколений (метагенез), реже медузоидное поколение отсутствует (гипогенез). Полипы представлены одиночными особями или колониями. Последние нередко полиморфны, и отдельные особи (гидранты) выполняют разные функции.
Ознакомиться со строением гидрозой можно на примерах пресноводной гидры и морских гидроидных полипов.
Гидра — одиночный, маленький (около 1 см) полип в форме вытянутого мешочка, прикрепленного к субстрату своей подошвой. На свободном конце тела расположен рот, ведущий в гастральную полость. Вокруг рта имеется венчик щупалец. Поверхность тела вплоть до краев рта покрыта эктодермой (рис. 17).
Рис. 17. Схема продольного разреза гидроидного полипа:
1 -
ротовое отверстие; 2 -
щупальце; 3 ~
гастральная полость; 4 —
эктодерма;
5 —
энтодерма; 6 —
мезоглея
В состав эктодермы входят эпителиально-мускульные клетки, концы которых вытянуты по продольной оси гидры в виде длинных сократимых волоконец. Между этими клетками разбросаны промежуточные клетки, за счет которых формируются половые и стрекательные клетки. Последние содержат стрекательную капсулу, заполненную жидкостью. Один конец капсулы ввернут внутрь наподобие полого отростка, закрученного в стрекательную нить. Наружная поверхность клетки несет тонкий чувствительный волосок — книдоциль. Прикосновение к нему возбуждает клетку, и стрекательная нить выворачивается наружу. Благодаря шипикам, находящимся на нити, а также ядовитости капсульной жидкости уколы нити ядовиты и могут убивать мелких животных.
В эктодерме располагаются также нервные (звездчатые) клетки, сообщающиеся своими отростками и образующие диффузное субэпителиальное сплетение — наиболее примитивную среди многоклеточных нервную систему. Единого нервного центра нет и к рефлексу способен каждый участок тела.
Энтодерма выстилает всю гастральную полость и продолжается до краев рта. В энтодерме есть эпителиально-мускульные пищеварительные клетки, железистые клетки, выделяющие пищеварительный сок. Эпителиальная часть энтодермальных клеток несет жгутики и способна образовывать псевдоподии для захвата пищевых частиц. Между экто- и энтодермой расположена мезоглея (ба-зальная мембрана).
Размножаются гидры бесполым (с помощью почкования) и половым способом. Половые клетки эктодермы превращаются непосредственно в яйца или многократным делением образуют множество сперматозоидов. Оплодотворение происходит в теле, затем гидра погибает, а из яиц развиваются новые гидры.
Морские гидроидные полипы в большинстве колониальны. Колония имеет форму деревца. На общем стволе колонии наподобие веточек образуются отдельные особи — гидранты, каждая из которых похожа на гидру. Гастральная полость у всех гидрантов и ствола колонии общая. Пища, захваченная одним гидрантом, распределяется по всей колонии. Эктодерма ствола колонии выделяет наружу органическую оболочку — теку, доходящую до гидрантов или охватывающую их в виде защитного колпачка. Полипы размножаются почкованием, что приводит к росту колонии. Половое размножение связано с образованием на стебле колонии половых особей — медуз, отрывающихся и переходящих к плавающему состоянию.
Медузы напоминают уплощенные полипы (рис. 18).
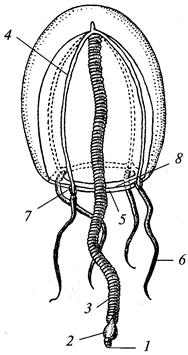
Рис. 18. Гидроидная медуза Sarta:
1 —
рот; 2 —
ротовой стебелек с расположенной на нем гонадой (3); 4—
радиальные каналы; 5— кольцевой канал; 6 —
щупальца; 7 — глазки; 8 —
парус
Они имеют форму зонтика или колокола. Вогнутая нижняя сторона несет посередине ротовой стебель со ртом на конце. Рот ведет в гастральную полость, которая состоит из центрального желудка и отходящих от него радиальных каналов, впадающих по краю зонтика в кольцевой канал. Желудок и каналы вместе образуют гастроваскулярную систему. По свободному краю зонтика прикреплена горизонтальная кольцевая мускулистая перепонка (парус), имеющаяся только у гидроидных медуз. От краев зонтика отходят щупальца. Мезоглея в теле медуз сильно развита и обладает студневидной консистенцией.
Нервная система медуз усложнена: кроме диффузного сплетения по краю зонтика образуется нервное кольцо, иннервирующее парус и органы чувств — глазки и органы равновесия — ста-тоцисты. Медузы — раздельнополы. Гонады находятся в эктодерме под радиальными каналами или на ротовом стебельке.
Половые органы выходят наружу через разрывы в стенке тела. Оплодотворение наружное. Дробление яйца полное, равномерное, приводящее к образованию бластулы. Двухслойный зародыш у подавляющего большинства кишечнополостных появляется путем иммиграции (внедрения части клеток в полость бластулы). Личинка — планула — снаружи покрыта ресничками и первое время ведет плавающий образ жизни. Впоследствии планула садится на дно передним концом (реже ложится на бок), в энтодерме формируется зачаток и вырастают щупальца. В конце концов планула превращается в маленького полипа, чем и заканчивается жизненный цикл.
Итак, в жизненном цикле гидроидных полипов наблюдается правильное чередование поколений — метагенез. При этом происходит смена гаплоидного и диплоидного поколений и гаметическая редукция. И медуза, и полип диплоидны, гаплоидны лишь гаметы.
Уклонение от общей схемы жизненного цикла заключается чаще всего в том, что образовавшиеся на стебле колонии медузы перестают отрываться и, оставаясь на месте, формируют половые клетки. У них обычно недоразвиты рот и органы чувств. Нередко эти медузы превращаются в мешки, набитые половыми клетками, т. е. подвижное самостоятельное поколение становится как бы половыми органами колонии.
Среди гидрозой выделяются плавающие полиморфные колонии отряда сифонофор, у которых отдельные особи, сидящие на общем стволе колонии, благодаря разделению функций превратились в органы. Среди них различают плавательный пузырь — пневматофор, плавательные колокола — нектофоры, кормящие полипы — гастрозоиды с ловчими арканчиками, а также половые полипы — гонофоры. У сифонофор стрекательные клетки объединены в стрекательные батареи, отчего сифонофоры очень сильно жалят.
Печальную известность приобрела ядовитая сифонофора «португальский кораблик» (Physalia).
Представители класса — крупные медузы без паруса. Желудок с четырьмя перегородками. Мезоглея с клеточными элементами. Гонады происходят из энтодермы. В цикле развития есть мета- и гипогенез.
Тело сцифоидных (рис. 19) имеет вид округлого зонтика. Посреди вогнутой нижней стороны на конце ротового стебелька помещается четырехугольный рот с ротовыми лопастями по краям. Это ловчий аппарат для захвата пищи. Рот ведет в энтодермальную глотку, переходящую в энтодермальный желудок. От желудка отходит сложная система радиальных каналов, которые впадают в кольцевой канал, окаймляющий края зонтика. По краю зонтика расположены щупальца и так называемые ропалии с органами зрения и равновесия. Усложнение нервной системы заключается в образовании ганглиев на протяжении нервного кольца.

Рис. 19. Схема строения сцифоидной медузы:
1 —
ротовые лопасти; 2 —
ротовое отверстие;
3 —
щупальца; 4 —
ропалии; 5 — кольцевой канал; 6 —
радиальный канал; 7 — гонада; 8 —
гастральные нити; 9 —
желудок; 10 —
эксумбрел-ла; 11 —
субумбрелла; 12 —
мезоглея. Эктодерма на поперечном срезе показана штриховкой, энтодерма — черной линией
Медузы раздельнополы. Гонады залегают в энтодерме, и половые продукты, прорывая стенку желудка, выводятся через рот наружу. Дробление яйца полное и равномерное. Из бластулы развивается планула, которая после недолгого периода плавания прикрепляется ко дну передним полюсом. На другом конце прорывается рот и образуются щупальца. В результате планула превращается в одиночного полипа — сцифостому. Впоследствии полип делится поперечными перетяжками, образуя подобие стопки молодых медуз (стробиляция), которые поочередно отрываются и начинают вести плавающий образ жизни. Молодые медузы — эфиры, сначала отличающиеся своим строением, постепенно вырастают во взрослых особей и переходят к половому размножению. В данном случае полипоидная стадия сцифостома — кратковременная, а медуза долгоживущая (рис. 20).
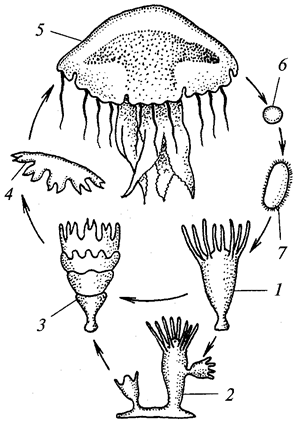
Рис. 20. Схема жизненного цикла сцифоидных (Chrysaora):
1 —
сцифистома; 2 — сцифистома, выпоч-ковывающая молодых сцифистом; 3 —
сцифистома в стадии стробилы; 4 —
эфира; 5 — медуза; 6 —
яйцо; 7 — планула
Коралловые полипы — колониальные или одиночные формы без медузоидной стадии. Гастральная полость радиальными перегородками — септами — поделена на камеры. Глотка эктодермальная. Гонады залегают в энтодерме. У большинства коралловых полипов есть известковый или органический скелет, образующийся за счет эктодермы или мезоглеи. К бесскелетным кораллам относятся прежде всего крупные полипы из отряда актиний (Actmaria), которые за свою форму, напоминающую цветок, называют также морскими анемонами.
Коралловые полипы устроены сложнее гидроидных (рис. 21). Тело у них цилиндрическое, нижний конец его наподобие плоской подошвы прикреплен к субстрату или соединен со стволом колонии. На противоположном конце, в центре ротового диска, окруженного щупальцами, помещается рот. Глотка напоминает трубку, свешивающуюся в гастральную полость и имеющую реснички для создания тока воды извне внутрь. Гастральная полость выстлана энтодермой. Септы состоят из мезоглеи, одетой энтодермой. Свободные края септ утолщены и превращены в мезентериальные нити, которые снабжены железистыми клетками, выделяющими пищеварительные ферменты.
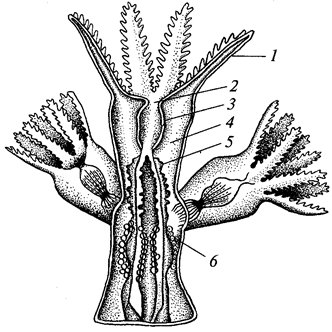
Рис. 21. Схематичное изображение части колонии
восьмилучевого коралла (Alcyonium):
7 — щупальца; 2 —
ротовое отверстие; 3 —
глотка; 4 —
септа; 5 — мезентериальные нити; 6 —
яйца
Под эктодермой находится слой обособившихся мускульных клеток (продольных и кольцевых). Такие же мускульные клетки развиты в энтодерме. Мезоглея у коралловых полипов образует толстый слой. У так называемых восьмилучевых кораллов в мезоглее залегают клетки — склеробласты с иголочками — спикулами внутри. Это скелетные элементы, за счет которых образуется внутренний скелет. У шестилучевых кораллов скелет наружный, так как выделяется эктодермой. Первоначально он имеет форму чашечки, в которой помещено тело полипа. Затем с внутренней стороны стенок этой чашечки внутрь тела вырастают скелетные перегородки (склеросепты), впячивающие стенку полипа в его гастральную полость.
Коралловые полипы размножаются бесполым и половым способами. Бесполое размножение обычно сводится к почкованию, но у мягких одиночных актиний наблюдается деление. Коралловые полипы раздельнополы. Гонады формируются в перегородках гастральной полости за счет энтодермы. При размножении сперматозоиды прорывают перегородки, попадают в гастральную полость и выводятся через рот наружу. Затем через рот женских особей они проникают в их гастральную полость и оплодотворяют яйца. Дробление яиц и формирование личинки-планулы происходит в теле материнского полипа. Планула выходит через его рот, плавает, а затем оседает на дно и превращается в молодого полипа.
Коралловые полипы распространены во всех морях Мирового океана. Так называемые рифообразующие, или мадрепоровые, кораллы живут в тропических частях Атлантического, Индийского и Тихого океанов, образуя подводные заросли — рифы, имеющие различную форму и протяжение. Коралловые рифы — место обитания множества организмов и являются одной из самых продуктивных систем океана.
Гребневики — морские свободноплавающие, реже ползающие по дну животные. Один вид ведет прикрепленный образ жизни. Как и у кишечнополостных, тело двухслойное с мезоглеей между экто- и энтодермой. Несмотря на некоторое, сходство с медузами, гребневики не идентичны ни медузам, ни полипам (табл. 12). Общее строение тела обнаруживает сочетание двух- и восьмилучевого типов симметрии. Двигаются гребневики при помощи видоизмененных ресничек. Характернейшая черта этих организмов — наличие клейких клеток (рис. 22). Насчитывается около 90 видов гребневиков из единственного класса с тем же названием.
Таблица 12.
Сравнительная характеристика кишечнополостных и гребневиков
| Типы, классы
|
Гастральная полость
|
Гонады
|
Жизненные формы
|
Развитие,
жизненный цикл
|
Тип
Coelenterata Класс
Hydrozoa
|
Растроваскулярная глотка отсутствует, у медуз с желудком и каналами |
в эктодерме
|
преобладает
полипоидная (бентосная)
|
прямое и с метаморфозом, метагенез |
| Тип Coelenterata Класс Scyphozoa |
эктодермальная глотка, желудок с карманами, каналы прямые и ветвящиеся |
в энтодерме в карманах желудка |
преобладает медузоидная (пелагическая) |
с метаморфозом и нередко с метагенезом |
| Тип Coelenterata Класс Anthozoa |
двухслойная глотка, выстланная эктодермой,
септы в желудке
|
в энтодерме на септах
|
только полипоидная (бентосная)
|
с метаморфозом, но без метагенеза
|
| Тип Ctenophora |
эктодермальная глотка, желудок, каналы ветвящиеся и неветвящиеся |
в энтодерме меридиональных каналов |
плавающие: овальные, ленточные; сидячие и ползающие |
прямое без метаморфоза |
Рис. 22. Строение клейкой клетки гребневика:
1 —
клейкие сосочки; 2 —
спиральная нить
Форма тела гребневиков чаще мешковидная (рис. 23).
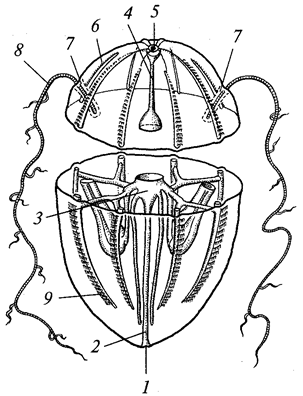
Рис. 23. Схема строения гребневика:
1 —
ротовое отверстие; 2 —
глотка; 3 —
желудок; 4 —
аборальный канал гастроваскулярной системы; 5 — аборальный орган; 6 —
меридиональный канал гастроваскулярной системы; 7—карман щупальца; 8—щупальце; 9 —
ряд гребных пластинок
На нижнем оральном полюсе расположен рот, противоположный полюс называют аборальным. Рот ведет в щелевидную энтодермальную глотку, имеющую форму сплющенной трубки, которая переходит в желудок. От желудка тремя ярусами отходят радиальные каналы. Часть из них впадает в меридиональные каналы, идущие параллельно поверхности тела от аборального полюса к оральному. Концы меридиональных каналов слепо замкнуты. Соответственно меридиональным каналам на поверхности тела проходит 8 ребер, на которых сидят гребневые пластинки — органы движения гребневиков. Каждая пластинка состоит из слившихся ресничек.
У большинства гребневиков есть пара щупалец в виде длинных арканчиков, служащих для захвата добычи. Эпителий щупалец содержит клетки, выделяющие клейкий секрет. Пойманная щупальцами добыча заглатывается краями широкого щелевидного рта.
Нервная система гребневиков состоит из поверхностного сплетения нервных клеток, из которых под рядами гребных пластинок образуются плотные тяжи, идущие к аборальному полюсу. Здесь имеются 4 ганглиозные массы, играющие роль координирующего центра. Над этим сплетением помещен аборальный орган, который представляет собой статолит, лежащий на четырех изогнутых дужках. Последние — образование из слившихся жгутиков рецепторных клеток, связанных с аборальным органом. Аборальный орган регулирует движение и выполняет функции органа равновесия.
Гребневики — гермафродиты, и их половые клетки происходят из энтодермы. Зрелые половые клетки сначала попадают в меридиональные каналы, а оттуда через рот наружу. Жизненный цикл протекает без метаморфоза. Дробление яйца полное и неравномерное. Гаструла образуется путем инвагинации.
Гребневики – елинственные первичнопланктонные кишечнополостные, что доказывается способом их движения и присутствием у них аборального органа. Число видов невелико. Заметного практического жначения для человека гребневики не имеют.
Животные, относящиеся к разделу билатеральных, отличаются двусторонней симметрией тела, при которой через него можно провести только одну плоскость симметрии.
Плоские черви - свободноживущие или паразитические формы, передвигающиеся при помощи ресничек или кожно-мускульного мешка. Пространство между внутренними органами заполнено паренхимой – рыхлой тканью мезодермального происхождения. Кишечник состоит из двух отдклов: эктодермальной передней кишки и анального отверстия нет. Кровеносная и дыхательная системы отсутствуют. Впервые появляется дыхательный аппарат протонефридиального типа и мозговых ганглий нервной системы. Половая система гермафродитна. Плоские черви насчитывают свыше 15 тыс. видов и 4 классов (табл. 13). Многие имеют важное практическое значение.
КЛАСС ТУРБЕЛЛЯРИИ, ИЛИ РЕСНИЧНЫЕ ЧЕРВИ (
TURBELLARIA
)
Ресничные черви – свободноживущие морские или пресноводные, редко наземные (в тропических лесах), хищные или паразитические формы (рис. 24). Под кожным покровом из ресничного или жгутикового эпителия с кожными железами расположены слои гладких мышц, образующих вместе с кожей кожно-мускульный мешок. Под ним находится паренхима, заполняющая пространство между внутренними органами. Кишечник у наиболее примитвных форм отсутсвует, и пища поступает через рот, распределяется в паренхиме, где и переваривается свободными фагоцитами. У высших форм имеется кишка различной формы — прямая или ветвящаяся. Выделительными органами являются протонефридии, первоначально выполняющие функцию осморегуляции.
Половой аппарат впервые возник в пределах этого класса и очень разнообразен. Турбеллярии, как правило, гермафродиты. У них имеются гонады и образуются половые протоки. Оплодотворение внутреннее.
Нервная система в примитивных случаях в виде рассеянного сплетения (как у кишечнополостных). У более высокоорганизованных форм — так называемый ортогон в виде отходящих от мозгового ганглия продольных стволов, соединенных поперечными перемычками.
Размножение большей частью половое, редко путем повторного поперечного деления, основанного на способности к регенерации. У морских турбеллярии развитие с метаморфозом и плавающей личинкой яйцевидной формы.
При классификации учитывается характер кожных покровов, строение кишечника и образ жизни.
Таблица 13.
Сравнительная характеристика классов плоских червей
Классы
|
Образ жизни
|
Кожно-мускуль-ный мешок
|
Кишечник
|
Органы чувств
|
Развитие
|
Основной хозяин
|
Промежуточный
хозяин
|
Turbellaria
|
свободножи-вущие (редко паразиты)
|
Ресничный эпителий, кольцевые, продольные, диагональные мышцы |
пищеварительная паренхима, трехветвистый, многоветвистый, прямокишечный |
глазки, статоцист, обонятельные ямки
|
прямое
|
—
|
—
|
Monogenea
|
эктопаразиты
|
тегумент, кольцевые, продольные
мышцы
|
тоже
|
глазки
|
с метаморфозом без смены хозяев, редко гетерогония |
рыбы, амфибии, рептилии
|
—
|
Trematoda
|
эндопаразиты
|
тегумент, кольцевые, продольные, диагональные мышцы |
двуветвистый
|
отсутствуют (глазки только у личинок)
|
с метаморфозом и сменой хозяев
|
позвоночные
|
мальки рыб, беспозвоноч
ные
|
Cestoda
|
тоже
|
тоже
|
нет
|
осязательные и
хеморецептор-
ные клетки
|
тоже
|
тоже
|
беспозвоночные и позвоночные |



Моногенеи — наружные паразиты рыб, реже паразитируют во внутренних органах амфибий и рептилий. Число видов около 2,5 тыс.
Близки к турбелляриям, но изменены паразитизмом (рис. 25). На заднем конце тела расположен особый прикрепительный диск с крючьями, присосками или двустворчатыми клапанами, между которыми защемляются участки тканей животного-хозяина. У личинок на этом же месте находится участок с эмбриональными крючьями — церкомер. Могут быть и ротовые присоски. Кожные покровы представлены тегументом — плотной оболочкой, образованной сильно видоизмененным эпителием, лишенным ресничек. Кишечник мешковидный или двуветвистый. Нервная система — ортогон.
Жизненный цикл моногеней проходит без смены хозяев и без чередования поколений. Развитие с метаморфозом. Цикл начинается с полового размножения. Из яйца выходит личинка, которая имеет кишку, глаза и плавает при помощи ресничек. Она активно ищет хозяина, прикрепляется к нему, теряет реснички и постепенно превращается во взрослого червя.
Сосальщики — внутренние паразиты позвоночных и беспозвоночных. Организация их похожа на таковую у турбеллярии. Приспособлениями к паразитизму являются следующие черты строения. У сосальщиков имеются органы прикрепления — присоски (ротовая и брюшная), церкомера с крючьями нет, покровы тела — тегумент. Кишечник часто разветвленный. Органы чувств развиты крайне слабо.
Оплодотворение внутреннее. Жизненный цикл сложный, со сменой нескольких партеногенетических поколений и одного обычного полового. Отмечается смена хозяев, среди которых обязательно находится моллюск и всегда есть стадии во внешней среде.
Среди сосальщиков много опасных паразитов позвоночных животных и человека (табл. 14).
Таблица 14.
Трематоды — паразиты человека и животных
Виды
|
Основной
хозяин
|
Заболевание и распростране
ние
|
Поражаемые органы
|
Промежуточный хозяин
|
Пути заражения основного хозяина
|
Печеночный сосальщик Fasciola hepatica
|
корова, редко человек
|
фасциолез; повсюду
|
печень
|
Пресноводный моллюск - малый прудовик |
проглатывание адолескариев при питье из водоемов |
Ланцетовидный
сосальщик
Dicrocoelium
dendriticum
|
овцы
|
дикроцелиоз;
повсюду
|
То же |
наземные
моллюски,
муравьи
|
Поедание вместе с травой муравьев с метацер-кариями |
Кошачья двуустка
Opistorchisfelineus
|
лиса, собака, человек |
Описторхоз,
повсюду
|
То же
|
пресноводные моллюски, рыбы |
Поедание рыбы с
метацеркариями
|
| Кровяная двуустка Schistosomahaematobium |
человек
|
шистосоматоз; Юго-Восточная Азия, Африка, Южная Америка |
крупные венозные сосуды брюшной полости |
пресноводные моллюски тропических регионов |
внедрение церкариев в кожу во время купания |
| ЛегочнаядвуусткаParagonimus nestermani |
То же |
парагонимоз; Япония, Корея |
легкие |
пресноводные моллюски, раки, крабы |
поедание раков с
метацеркариями
|
Рассмотрим в качестве примера образ жизни и развитие печеночной двуустки (рис. 26). Взрослый червь живет в печени и желчных протоках рогатого скота (очень редко у человека). Яйца, отложенные там же, должны попасть в воду, где из них выходит плавающая с помощью ресничек личинка — мирацидий. В его паренхиме находятся зародыши следующих поколений паразита. Мирацидий внедряется в другого хозяина (моллюска) и превращается в спороцисту, которая питается, всасывая пищу через свои покровы. В полости паренхимы спороцисты развивается следующее поколение.
Партеногенетические потомки спороцисты - редии - уже имеют рот глотку и кишку. В их полости тоже партеногенетически образуются церкарии, покидающие редию и выходящие из моллюска в воду. Через некоторое время они оседают на прибрежных травах, покрываются защитной оболочкой (инцистируются) и некоторое время сохраняют жизнеспособность. Скот вместе с травой заглатывает инцистированных церкариев, которых называют адолескарии, или метацеркарии.
Большое значение в медицине имеет кошачья двуустка, заражение которой происходит при поедании плохо прожаренной рыбы, где находятся метацеркарии. Необходимо отметить также кровянуто двуустку, живущую в крупных венах человека. Церкарии последней активно внедряются в кожу человека при купании.
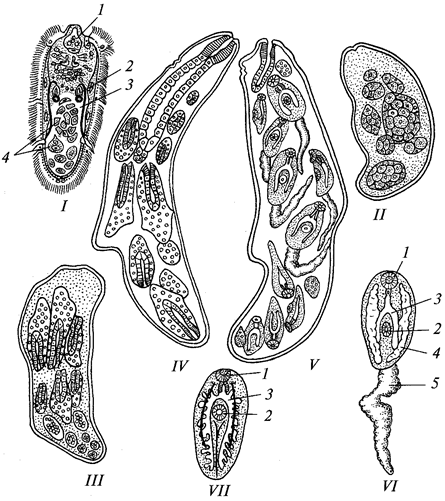
Рис. 26. Стадии развития фасциолы печеночной (Fasciolahepatica):
1 — мирацидий: 1 —
кишечник; 2 —
протонефридий; 3 —
выводной канал про-тонефридия; 4 —
зародышевые клетки; II—
Ш —
спороцисты, в которых развиваются редии; IV—
редия, в которой повторно развиваются редии; V—
редия, в которой развиваются церкарии; VI —
церкарий: 1 —
передняя, или ротовая, присоска; 2 —
брюшная присоска; 3 —
кишечник; 4 —
железы; 5 — хвост; VII —
адолескарий: 1, 2, 3 —
см. VI; 4 —
выделительная система
Представители класса - исключительно эндопаразитические плоские черви, на организацию которых паразитизм наложил наибольший отпечаток. Во взрослом состоянии живут в кишечникесвоих хозяев, молодые стадии - в полости тела и в органах как беспозвоночных, так и позвоночных (табл. 15).

Тело лентовидное, поделенное на членики (проглоттиды), редко нерасчлененное. Передний конец образует головку (сколекс) сприкрепительными органами в виде присасывательных ямочек и щелей (ботрий), присосок и крючьев. Кишечник отсутствует, и переваренная хозяином пища всасывается через микроворсинки тегумента, поступая затем в паренхиму червя. Нервная система-ортогон. Органы чувств развиты слабо. Гермафродитная половая система способна продуцировать огромное количество яиц. Оплодотворение внутреннее.
В жизненном цикле примитивных цестод есть личинки с ресничным эпителием и крючьями на оральном конце. Личинки на ходятся в воде, и для продолжения цикла их должен промежуточный хозяин, например рачок. В нем такая личинка (корацидий) сбрасывает реснички, превращается в шестикрючного зародыша-онкосферу), а затем в следующую личиночную стадию - червеобразный процеркоид с церкомером на заднем конце тела.
Когда рачок съедается вторым промежуточным хозяином - рыбой - процеркоид теряет церкомер, превращаясь в плероцеркоида. Последний уже имеет сколекс и является готовым, хотя и нерасчлененньш и неполовозрелым, червем. Завершение цикла (пост и созревание) проходит в окончательном хозяине - позвоночном, съевшем рыбу. Таков цикл развития у широкого лентеца.
У других цестод, например у свиного солитера (Taeniasolium) (рис. 27), цикл развития протекает на суше, личинки-корацидия нет, хозяев и стадий меньше, а вместо плероцеркоида развивается специализированная личинка — финна, находящаяся обычно в мышечной ткани промежуточного хозяина.
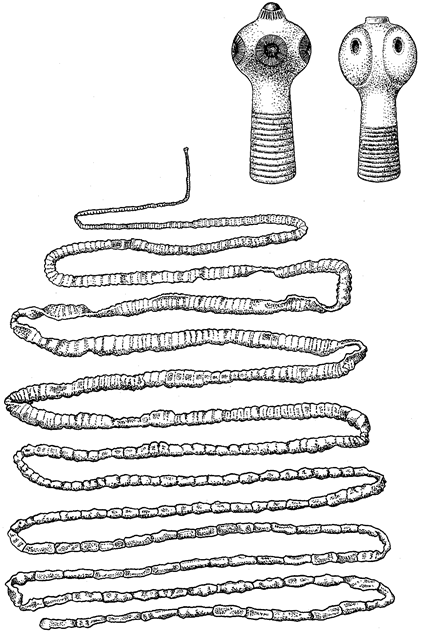
Рис. 27. Свиной солитер (Taeniasolium) — внизу. Вверху справа
головы свиного (слева) и бычьего (справа) солитеров
Регулярное употребление в пищу плохо обработанного (недоваренного) свиного мяса приводит к заражению человека, в кишечнике которого финна превращается в молодого солитера. На личиночной стадии некоторых цестод (например, эхинококка) происходит бесполое размножение путем выпочковывания ско-лексов, каждый из которых дает новую особь.
К числу цестод, имеющих наибольшее значение в медицине, относятся свиной и бычий солитеры, широкий лентец, эхинококк.
Тип круглых червей относится к так называемым первичнопо-лостным червям, у которых возникает схизоцель — полость тела, образующаяся в паренхиме за счет распада части ткани (сравнительную характеристику разных типов первичнополостных червей см. в табл. 16). Собственной эпителиальной выстилки схизоцель не имеет. Кроме того, появляется задняя кишка с анальным отверстием. Тело не сегментировано. Представители большей частью раздельнополы. Круглые черви живут в воде и на суше. В типе 2 класса: брюхоресничные (Gastrotricha) и нематоды, или собственно круглые черви (Nematoda). Здесь рассматривается только последний класс.
Нематоды — самый крупный класс среди червей. Распространены всесветно. Размеры от миллиметра до метра и более. Тело в поперечном сечении круглое, в конце суженное (рис. 28). Кожный покров состоит из кутикулы и гиподермы, имеющей клеточное строение или являющейся синцитием — продуктом слияния эпителиальных клеток. Вместе с гиподермой и продольными мышцами кутикула образует кожно-мускульный мешок. Кольцевой мускулатуры нет. Ресничные образования отсутствуют.
Кишечник нематод — в виде прямой трубки, заканчивающейся анальным отверстием. Редко (у нитчаток-филярий) кишечник замкнут на конце слепо. Выделительные органы — протонефридии или, чаще, кожные железы с выводными каналами. Нервная система — ортогон. Органы чувств развиты слабо и представлены осязательными бугорками или щетинками, обонятельными щелями (амфидами), а у морских нематод есть и светочувствительные органы типа глазного пятна или бокала.
Нематоды раздельнополы. Гонады трубчатого типа. Сперматозоиды самцов без жгутика на конце. Яйцекладущие или живородящие формы. Развиваются обычно без чередования поколений. Дробление яйца полное, детерминированное. Личинки похожи на взрослых, а их рост сопровождается несколькими линьками, при которых происходит смена кутикулы.
Таблица 16.
Сравнительная характеристика четырех типов первичнополостных червей
| Типы, классы |
Особенности организации |
| Покровы |
Мышцы |
Полость
тела
|
Ортогон |
Органы чувств |
Выделительная система |
Развитие |
Образ жизни |
| Тип Круглые черви (Nemathelminthes) |
| Класс Брюхорес-ничные Gastrotricha |
ресничный эпителий и кутикула |
пучки продольных мышц (гладкие) |
схизоцель с паренхимой |
2 боковых ствола |
хеморецеп-торы осязания |
2 прото-нефридия |
прямое |
свободно-живущие |
| Класс Нематоды Nematoda |
гиподерма и кутикула |
продольные (гладкие) |
тоже |
спинной и брюшной ствол |
хеморецеп-торы |
шейные железы |
тоже |
свободно-живущие и паразиты |
| Тип Коловратки Rotatoria |
гиподерма с псевдокутикулой + ресничный аппарат |
пучки продольных и кольцевых мышц (поперечнополосатые) |
схизоцель |
2 брюшных ствола |
глаза, щупальца |
2 прото-нефридия |
прямое (гетерогония) |
свободно-живущие |
| Класс Киноринхи Kinorhyncha |
гиподерма и кутикула |
пучки продольных мышц (поперечно-полосатые) |
схизоцель |
1 брюшной ствол |
глаза, осязательные волоски |
2 прото-нефридия |
с метаморфозом |
свободно-живущие |
| Класс Приапулиды Priapulida |
тоже |
кольцевые и продольные (гладкие) |
«вторичная» полость |
тоже |
— |
— |
тоже |
тоже |
| Класс Волосатики Nematomor-pha (рис. 24) |
тоже |
продольные (гладкие) |
схизоцель с паренхимой |
тоже |
— |
— |
с метаморфозом (со сменой хозяев) |
полупаразиты |
| Тип Скребни Acanthoce-phala |
тоже |
кольцевые и продольные |
схизоцель |
2 боковых ствола |
2 чувствительных сосочка |
2 прото-нефридия |
тоже |
паразиты |
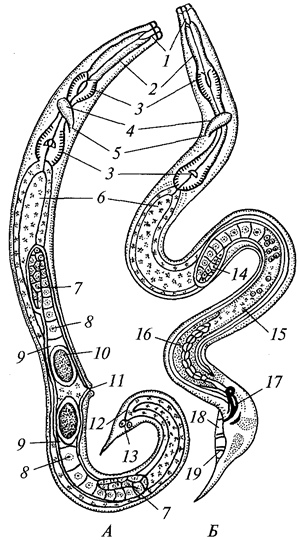
Рис. 28. Схема организации нематод. А —
самка; Б —
самец:
1 — ротовая полость; 2 —
пищевод; 3 —
бульбусы пищевода; 4 —
окологлоточное нервное кольцо; 5— выделительная пора;
6 —
средняя кишка; 7 — яичник; 8 —
яйцевод; 9 —
матка; 10 —
яйцо в матке; 11 —
женское половое отверстие; ]2 —
задняя кишка; 13 —
анальное отверстие; 14 —
семенник; 15 —
семяпровод; 16 —
семяизвергательный канал; 17 —
спику-лы; 18 —
бурсальные крылья; 19 —
ребро бурсы (органы осязания)
Характерная особенность нематод — постоянство клеточного состава (строго определенное число клеток, входящих в состав какой-нибудь ткани или органа). В связи с этим они не способны к регенерации.
Живут нематоды на дне морей, в пресной воде и почве. Свободноживущие формы играют важную роль в процессах разложения органических веществ в природе. Большое значение в медицине и ветеринарии имеют паразитические нематоды (табл. 17). Во многих случаях возникают сложные циклы со сменой хозяев и со свободными стадиями во внешней среде (или без этих стадий). Заражение происходит при заглатывании яиц паразита, лишь у трихины — при поедании одного хозяина другим.
Представители: детская острица, аскарида, ришта, трихина. Из паразитов растений — свекловичная нематода, стеблевая нематода картофеля, пшеничная угрица.
Таблица 17.
Нематоды — паразиты человека
Виды
|
Тип жизненного
цикла
|
Вызываемые заболевания
|
Основной хозяин
|
Промежуточный
хозяин
|
Яйцерождение или живорождение
личинок
|
Миграции личинок по
крови
|
Пути заражения
|
| Аскарида человеческая Ascarislumbri-coides |
геогельминт без смены хозяев |
аскари-диоз |
человек
(кишечник)
|
—
|
яйца |
есть |
прогла-
тывание
яиц с пищей и водой
|
Власоглав
Trichocephalus trichiurus
|
тоже
|
трихо-цефалез
|
человек (слепая кишка) |
—
|
то же
|
то же
|
То же |
Свайник
Ancylostoma
duodenale
|
то же |
анкилостомоз |
человек (12-перстная кишка) |
—
|
то же
|
тоже
|
то же
|
Острица
детская
Enterobius
vermicularis
|
тоже
|
энтеробиоз
|
человек
|
—
|
тоже
|
—
|
то же
|
Трихи-
нелла
Trichi-nella
spiralis
|
биогельминт
без смены
хозяев
|
трихинеллез
|
человек,
свинья,
крыса (кишка, мышцы)
|
—
|
личинки
|
есть
|
проглатывание капсул в свином мясе |
Нитчатка Банк-рофта Wuchereria
bancrofti
|
биогельминт со сменой хозяев
|
слоновая болезнь
|
человек
(кровь, лимфатические сосуды)
|
комар
|
тоже
|
тоже
|
при укусе комаров, зараженных
личинками
|
Ришта
Dracun-culus medinensis
|
геогельминт со сменой хозяев
|
дракунулез
|
человек
(подкожная клетчатка)
|
циклоп
|
личинки
|
мигрирует по лимфе
|
При питье
воды с циклопами, зараженными
личинками
|
Кольчатые черви — тип высших червей (около 17 тыс. видов), обладающих сложной организацией (рис. 29). Это хищники, сапрофаги или наружные паразиты.
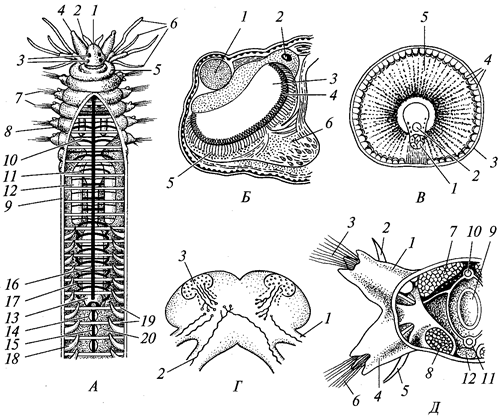
Рис. 29. Анатомия кольчецов:
А —
вскрытый Nereis: 7 — голова (простомиум); 2 —
головные тактильные усики; 3 — глаза; 4 —
пальпы (органы химического восприятия); 5 — ротовой сегмент (перистомиум); 6—
усики ротового сегмента; 7— параподии с пучками щетинок; 8 —
сегменты тела; 9 —
стенка тела; 10 —
глотка; 11 —
желудок; 12 —
боковые выросты кишечника; 13 —
средняя кишка; 14 —
брюшная нервная цепочка; 15 —
брюшной кровеносный сосуд; 16 —
спинной кровеносный сосуд; 17—
кольцевые кровеносные сосуды; 18 —
органы выделения (метанефридии); 19 —
перегородки вторичной полости тела; 20 —
вторичная полость тела (целом). Б —
глаз пелагической полихеты Vanadisramosa: I
—
линза; 2 —
клетка, производящая стекловидное тело; 3 —
стекловидное тело; 4 —
слой палочек ретины; 5 —
чувствительный слой ретины; 6—
оптический нерв. В —
поперечный разрез через жаберную нить и сложный глаз Branchiomma: / — хрящевой стержень жаберной нити; 2 —
кровеносный сосуд; 3 —
кроющий эпителий сложного глаза; 4 —
отдельные линзы глазков; 5— отдельные ретинальные клетки сложного глаза. Г—
поперечный разрез головного мозга Sthenelais (Aphroditidae): I
—
пальпарный нерв; 2 —
окологлоточный коннектив; 3 —
ассоциативные центры мозга (грибовидные тела, или глобулы). Д —
поперечный разрез сегмента тела Nereis: / — спинная ветвь параподии; 2 —
спинной усик; 3 —
пучок спинных щетинок; 4 —
брюшная ветвь параподии; 5 — брюшной усик; 6 —
пучок брюшных щетинок; 7 — спинная лента мышц;
8 —
брюшная лента мышц; 9 —
кишечник; 10 —
спинной кровеносный сосуд;
11 —
брюшной кровеносный сосуд; 12 —
брюшная нервная цепочка
Тело кольчатых червей состоит из головной лопасти (простомиума), сегментированного туловища и анальной лопасти (пигидиума). Количество туловищных сегментов варьирует от 5 до 800. Целом развит во всех сегментах, кроме головной и анальной лопастей. Примитивные формы имеют щетинконосные парные придатки тела — параподии.
Рот у кольчатых червей находится на брюшной стороне тела на границе головной лопасти и первого туловищного сегмента. Пищеварительная система обычно включает ротовую полость, глотку, среднюю и заднюю кишку. Кровеносная система, если есть, замкнутая. Сердце отсутствует. Выделительный аппарат — сегментарные органы (нефридии), которые могут соединяться с половыми протоками целомодуктами или срастаться с ними, образуя нефромиксии. Нервная система состоит из парного надглоточного ганглия и брюшной нервной лестницы либо цепочки. Половая система раздельнополая или гермафродитная.
Развитие у примитивных форм кольчатых червей с метаморфозом (типичная личинка — трохофора), у специализированных оно прямое.
Кольчатые черви живут в море, пресных водах или в почве. В типе 5 классов: первичные кольчецы (Archiannelida), много-
щетинковые, или полихеты, (Polychaeta), малощетинковые (Oli-qochaeta), мизостомиды (Myzostomida) и пиявки (Hirudinea) (табл. 18), из которых подробнее рассмотрим три наиболее важных.
Таблица 18.
Сравнительная характеристика классов кольчатых червей
| Классы
|
Тип движения
|
Наличие параподий
|
Органы чувств и нервная система
|
Органы выделения
|
Полость
тела
|
Кровеносная система
|
Половая система
|
Развитие
|
| Первичные кольчецы |
при помощи ресничек или парапо-дий и мышц |
отсутствуют или имеются |
лестничная, глаза, стато-цисты, щупальца |
протонефри-дии или метанефри-дии |
первичная или вторичная |
отсутствует или имеется |
раздельнополы |
с метаморфозом, личинка — трохофора |
| Многоще-тинковые |
при помощи параподий и мышц |
имеются или редуцированы |
брюшная нервная цепочка, глаза, щупальца, обонят. ямка |
метанефри-дии или нефромик-сии |
вторичная сегментированная |
имеется |
тоже |
тоже |
| Малоще-тинковые |
при помощи мускулатуры и гидравлики |
отсутствуют (рудименты) |
брюшная нервная цепочка |
метанефри-дии |
тоже |
тоже |
гермафродиты |
прямое |
| Пиявки |
при помощи мускулатуры и ползание с помощью присосок |
тоже |
брюшная нервная цепочка, глаза хемо-ренепторы |
тоже |
вторичная с паренхимой, сегментированная |
кровеносная или ложно-кровеносная |
тоже |
тоже |
| Мизосто-миды |
при помощи параподий и мышщ |
имеются |
брюшная нервная цепочка, щупальца, обонят. ямка |
тоже |
вторичная, несегментированная |
отсутствует |
тоже |
с метаморфозом, личинка — трохофора |
(
POLYCHAETA)
Полихеты (рис. 30) — морские черви с парными придатками — параподиями на туловищных сегментах. Тело покрыто тонкой кутикулой, без нервного сплетения в эктодерме. Головная лопасть несет глаза и различные придатки — пальпы и антенны, на которых размещаются органы химического чувства (см. рис. 29).
Рис. 30. Полихеты:
1 —
Typhloscolex; 2 —
Netochaeta; 3 —
Euniceviridis
В зависимости от количества сегментов различаются олигомер-ные (малосегментные) и полимерные (многосегментные) черви. По форме и характеру придатков сегменты могут быть одинаковыми (гомономными) или отличаться друг от друга. Гомономность является признаком примитивной организации и присуща сво-бодноподвижным формам.
Параподии — двуветвистый вырост боковой стенки сегмента, снабженный пучками щетинок и щупальцем — усиком на каждой или только на одной ветви. Это примитивные органы движения. У сидячих форм параподии нередко частично редуцируются.
Под кожей располагаются слои кольцевых и продольных мышц. Кожно-мускульный мешок подостлан изнутри эпителием, а под ним помещается полость тела — целом. В каждом сегменте целом состоит из двух мешочков, стенки которых сходятся над и под кишкой, создавая продольную перегородку. На границе между сегментами целомический эпителий образует двухслойную перегородку — септу, или диссепимент. В некоторых отделах септы могут редуцироваться. Целом выполняет опорную (за счет заполняющей его жидкости), распределительную, выделительную и половую функции.
Пищеварительная система начинается с ротовой полости, которая переходит в мускулистую глотку. За ней следует пищевод. У некоторых полихет есть маленький желудок. Средняя кишка представляет собой прямую трубку. Задняя кишка коротка, открывается анальным отверстием на дорсальной стороне анальной лопасти.
Дыхание у полихет осуществляется через поверхность тела, но у большинства возникают специальные участки, через которые идет газообмен. Обычно это спинной усик параподии, превратившийся в жабру.
Кровеносная система замкнутая, состоит из продольных главных сосудов (над и под кишкой), которые сообщаются благодаря системе кольцевых сосудов. Движение крови обусловливается пульсацией стенок спинного сосуда. При редукции системы ее функции принимает на себя целомическая жидкость.
Выделительная система представлена нефридиями разного строения. Как правило, каждый из них — это трубочка, один, расширенный, конец которой открывается в целом, другой — наружу. Так как нефридии имеются в каждом сегменте тела, их называют еще сегментарными органами. У некоторых низших форм эти органы представлены протонефридиями, у которых на внутреннем конце трубочки имеется пучок булавовидных клеток-соленоцитов. У других полихет этот пучок атрофируется, вместо него появляется отверстие, усаженное ресничками. Такие органы называются метанефридиями.
Нервная система в пределах класса изменяется от брюшной нервной лестницы до брюшной нервной цепочки. Кроме того, она уходит с периферии (из кожного эпителия) вглубь, иногда даже в полость тела. Отмечается концентрация нервных узлов. Органы чувств разнообразны и лучше выражены у свободноподвиж-ных форм. Это органы осязания, химического чувства (хеморе-цепторы) и зрения. Последние могут иметь вид глазного бокала или пузыря.
Половая система устроена просто. Многощетинковые кольчецы раздельнополы. Гонады формируются либо во всех (за исключением первого и последнего), либо только в особых, или плодущих, сегментах. Это кучка половых клеток, лежащих под целоми-ческим эпителием. Созревшие половые продукты через разрывы эпителия попадают в целом, а затем выводятся половыми воронками с выводным каналом наружу. В большинстве случаев эти воронки срастаются с нефридиями и тогда выполняют половую и выделительную функции.
Оплодотворение наружное. Дробление яиц полное, спиральное, детерминированное. После дробления образуется крайне типичная личинка — трохофора, а затем расчлененная на небольшое число сегментов метатрохофора. Новые сегменты (после-личиночные, или постларвальные) появляются всегда в одном месте — в зоне роста, которая находится между пигидиумом и предпоследним туловищным сегментом.
Полихеты играют важную роль в экосистемах морей, поскольку служат пищей рыб, крабов и других животных. В ряде случаев отдельные виды полихет даже переселяют в новые для них водоемы для улучшения пищевого рациона промысловых рыб.
Малощетинковые — пресноводные и почвенные полимерные формы, редко паразиты. Тело вытянутое, цилиндрическое, размером от миллиметра до 3 м (гигантские дождевые черви). Сегменты, за исключением переднего, снабжены мелкими щетинками — остатками параподий. Кожный эпителий, выделяющий тонкую эластичную кутикулу, богат слизистыми клетками. Последние особенно многочисленны в области пояска. Кожно-мускульный мешок и целом обычного для кольчатых червей типа.
Пищеварительная система специализирована лишь у дождевых червей (рис. 31). У них передняя кишка дифференцируется на мускулистую глотку, пищевод и мускульный желудок. Спинная сторона средней кишки образует продольное желобовидное впячивание внутрь — тифлозоль, что увеличивает всасывательную поверхность кишки.
Рис. 31. Анатомия дождевого червя Lumbricus:
1 —
простомиум; 2 —
церебральные ганглии; 3 —
глотка; 4 —
пищевод; 5 — боковые сердца; б — спинной кровеносный сосуд; 7 — семенные мешки; 8 —
семенники; 9 —
семенные воронки; 10
— семяпровод; 11 —
метанефридии; 12 —
средняя кишка; 13 —
мускулистый желудок; 14 —
зоб; 15
— яйцевод; 16 —
яйцевые воронки; 17—
яичники; 18 —
семяприемники. Римскими цифрами обозначены сегменты тела
Органы выделения — метанефридии. Нервный и кровеносный аппараты, как у полихет. Органы чувств развиты слабо. Это кожные органы чувств, органы обоняния, статоцисты и глаза, которые есть далеко не у всех представителей класса.
Половая система крайне своеобразна. Гонады находятся лишь в немногих сегментах тела. У почвенных форм семенники расположены в двух сегментах, имеются также резервуары (семенные мешки) и выводные протоки — целомодукты, открывающиеся на XV
сегменте. Яичники размещены только в одном сегменте, куда попадает сперма при перекрестном оплодотворении. Сперма хранится в них до момента откладки яиц. Яйца откладываются в слизистую муфту, которая образуется за счет выделений желез пояска. В этой же муфте происходит и оплодотворение, когда на яйца выдавливается сперма из семяприемников. Таким образом, оплодотворение у малощетинковых — наружное. Развитие прямое, в коконе, образующемся при смыкании концов муфты.
Есть и бесполое размножение — делением на две части, причем обе части впоследствии регенерируют недостающие участки. Этот способ размножения получил название архитомии.
Пресноводные формы ползают по дну или живут в норках. Наземные ведут роющий образ жизни и появляются на поверхности почвы только ночью или в дождливую погоду. Наибольшее практическое значение имеют дождевые черви, роющая деятельность которых способствует улучшению плодородия почвы. Пресноводные олигохеты являются хорошим кормом для донных рыб.
Пиявки — пресноводные, морские и изредка наземные формы. Большинство из них — хищники, питающиеся беспозвоночными или сосущие кровь позвоночных (рис. 32).
Рис. 32. Обыкновенные пиявки (Piscicolageometra),
паразитирующие на карпе
Тело сплющенное в спинно-брюшном направлении, состоит из 30—33 сегментов, различить которые можно при вскрытии. У примитивных пиявок остались щетинки, но у других они отсутствуют. Есть две присоски — передняя и задняя. Эпителий выделяет кнаружи плотную кутикулу и содержит массу слизистых и пигментных клеток. Число мышечных слоев велико, так как кроме кольцевых и продольных мышц имеются диагональные и дорсовентральные пучки.
Ротовая полость с хоботком или кутикулярными челюстями в виде зубчиков. Кишечник пиявок сложно расчленен, особенно у кровососущих видов. На начальном участке средней кишки развиты боковые слепые выросты — карманы, где насосанная кровь может сберегаться несколько недель не сворачиваясь. Консервирующим веществом служит гирудин, выделяемый слюнными железами.
Целом сохраняется у примитивных пиявок, но у большинства заполняется соединительной тканью — паренхимой. В связи с этим кровеносная система становится незамкнутой. Нефридии находятся лишь в нескольких сегментах. Гонады выглядят наподобие мешков, лежащих в паренхиме. Их протоки — видоизмененные цело-модукты. Оплодотворение внутреннее.
Пиявки имеют практическое значение как паразиты-кровососы. Раньше пиявки широко использовались для снижения кровяного давления. В последнее время вновь возросло использование медицинских пиявок (Hirudomedicinalis) при лечении различных заболеваний.
Членистоногие — самый крупный тип животного царства (свыше 1,5 млн видов). Водные и наземные формы имеют всесветное распространение.
Членистоногие — гетерономно расчлененные животные, причем группы сходных сегментов образуют три отдела: голову, грудь, брюшко. Заканчивается тело анальной лопастью — тельсоном. Голова состоит из головной лопасти, или акрона, и 4 сегментов. Ноги членистые. Кожные покровы представлены трехслойной кутикулой, разделенной в каждом сегменте (кроме головы) на щитки. Из-за твердости кутикулы рост сопровождается периодическими линьками. Кохно-мускульный мешок сохраняется лишь у самых примитивных форм. Движение происходит с опорой на наружный скелет, к которому прикрепляются пучки поперечно-полосатых мышц.
С помощью туловищных конечностей членистоногие захватывают и обрабатывают пищу. При питании используются придатки оснований конечностей.
Полость тела смешанная (миксоцель), так как стенки целоми-ческих мешков во время онтогенеза распадаются. Дыхание кожное, жаберное, легочное или трахейное. Кровеносный аппарат незамкнутый. В примитивных случаях органы выделения — цело-модукты, но у большинства появляются мальпигиевы сосуды. Кишечник состоит из трех отделов, средняя кишка у некоторых групп имеет печеночные выросты. Нервный аппарат — брюшная нервная цепочка, в которой различают надглоточный ганглий (головной мозг), окологлоточное кольцо и ганглии брюшной цепочки, соединенные коннективами друг с другом. Наблюдается тенденция к слиянию ганглиев и уменьшению числа сегментов тела. Органы чувств разнообразны и у высших представителей достигают значительного совершенства.
Размножение только половое, причем подавляющее большинство членистоногих раздельнополы. Нередки партеногенез и живорождение. Жизненные циклы разнообразны, однако всегда сопровождаются линьками.
Тип делится на четыре подтипа: трилобиты (вымершая группа), жабродышащие, хелицеровые и трахейные (табл. 19).
Таблица 19.
Сравнительная характеристика подтипов членистоногих
Подтипы
|
Среда обитания
|
Органы дыхания
|
Отделы тела
|
Сегментарный состав головы или головогруди
|
Придатки головы или головогруди
|
Органы выделения
|
Трилобиты Trilobitomor-
pha
|
море |
жабры
|
голова,
туловище
|
акрон + 4
сегмента, голова слитная
|
антенны + 4
пары конечностей
|
почки |
| Жабродышащие Branchi-ata |
море, пресные воды (редко суша) |
тоже |
голова, грудь, брюшко |
акрон + 4 сегмента (протоце-фалон и гнатоцефалон) |
2 пары антенн, мандибулы, 2 пары максилл |
тоже |
| Хелицеровые Chelicerata |
море, пресные воды, суша |
Жабры или легкие, трахеи |
ГОЛОВОГРУДЬ,
брюшко
|
акрон + 7 сегментов |
хелицеры,
педипаль-пы + 4 пары ходильных ног
|
почки +
мальпигиевы сосуды
|
Трахейные Tracheata
|
суша, вторично-волная среда
|
трахеи
|
голова,
туловище или голова, грудь, брюшко
|
акрон + 4 сегмента
|
1 пара антенн, мандибулы, 2 пары
максилл
|
мальпигиевы сосуды
|
Представители жабродышащих — первичные членистоногие, органами дыхания которых являются жабры. Подтип включает класс ракообразных, в котором свыше 20 тыс. видов.
Ракообразные — это водные членистоногие размером от миллиметра до метра (рис. 33).
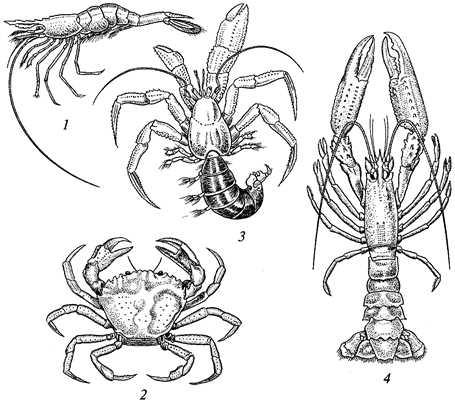
Рис. 33. Представители десятиногих ракообразных:
1 —
креветка Pandalusborealis; 2 —
краб Eriphiavemicosa; 3 —
рак-отшельник Pagurusbernhardus; 4 —
норвежский омар (Nephropsnorvegicus)
Сегментация разнообразна, у примитивных форм близка к гомономной. Число сегментов от 5 до 50. Обычно сегменты образуют отделы — голову, грудь и брюшко, наблюдается также различная степень слияния головных и грудных сегментов. Из придатков головы наиболее характерны две пары антенн (антеннулы, или антенны I, и антенны II) и антенны, гомологичные пальпам кольчецов (рис. 34).
Ножки во многих случаях двуветвисты, что служит примитивным признаком.
Покровы тела представлены хитиновой кутикулой, которая часто укрепляется отложениями солей углекислой извести, и лежащим под ней слоем гиподермального эпителия. Самый верхний слой кутикулы (эпикутикула), препятствующий у сухопутных членистоногих испарению воды из тела, у ракообразных отсутствует. Спинная сторона покровов нередко образует характерную складку (карапакс), которая иногда охватывает тело с боков, оставляя Щель, ведущую в жаберную полость.
Дыхательная система ракообразных представлена жабрами, связанными с конечностями. Кровеносная система включает сердце с отверстиями — остиями, а также сосуды разной степени развития. Последних может и не быть.
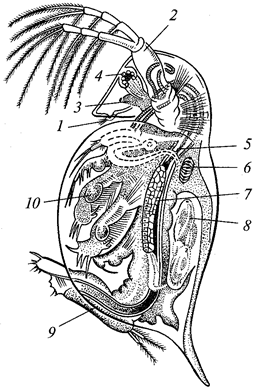
Рис. 34. Строение дафнии:
1 —
передняя антенна; 2 —
задняя антенна; 3
— науплиальный глазок; 4 —
фасеточный глаз; 5 —
кишечник; 6 —
сердце; 7 — яичник; 8 —
эмбрионы в выводковой сумке; 9 —
брюшко; 10 —
грудная ножка
Выделительные органы — измененные целомодукты, открывающиеся у основания антенн или максилл. Нервный аппарат — брюшная нервная цепочка различной сложности. Органы чувств разнообразны: органы осязания и химического чувства (чувствительные волоски), статоцисты и глаза, которые могут быть простыми (у личинок и примитивных форм) или сложными (фасеточными).
Громадное большинство раков раздельнополы, что часто сопровождается половым диморфизмом. Размножение половое. У низших раков имеют место партеногенез и чередование партеногенетических и обоеполых поколений (гетерогония). Осеменение сперматофорное или с копуляцией. Дробление яиц может быть полным, и тогда развитие идет с метаморфозом, причем первая личиночная стадия (науплиус) характерна для всех групп. У многих ракообразных развитие прямое.
Классификация ракообразных
Подкласс Жаброногие — Branchiopoda
Отряд Жаброноги — Anostraca
Отряд Листоноги — Phyllopoda
Подотряд Щитни — Notostraca
Подотряд Ветвистоусые — Cladocera
Подотряд Раковинные листоногие — Conchostraca
Подкласс Веслоногие — Copepoda
Подкласс Карповые вши, или Карпоеды — Branchiura
Подкласс Усоногие — Cirripedia
Подкласс Мистакокариды — Mystacocarida
Подкласс Цефалокариды — Cephalocarida
Подкласс Мешкогрудые — Ascothoracida
Подкласс Ракушковые — Ostracoda
Подкласс Высшие раки — Malacostraca
В классе девять подклассов с большим количеством отрядов (морфологическая характеристика основных подклассов приведена в табл. 20). Самые примитивные — жаброногие, у которых голова не срастается с грудью, а грудные ножки листовидные и выполняют функции движения, дыхания и подачи пищи ко рту. Представитель — жаброног (Branchipius).
Таблица 20.
Морфология некоторых подклассов ракообразных
| Подклассы
|
Расчленение тела
|
Глаза
|
Челюстной аппарат
|
Грудные
ноги и
|
Брюшко
и придатки
|
Жаброногие
Branchiopoda
|
Голова (расчлененная или слитная), грудь, с брюшко |
2 фасеточных и простой глазок |
3 пары челюстей: мандибу- лы, 2 пары максилл |
листовидные (дыхание, движение, фильтрация) |
8—19 сегм.+ тельсон, ног нет,
есть вилочка
|
Цефалокариды
Cephalocarida
|
голова слитная, грудь (10 сегм.),
брюшко
|
нет |
1 пара мандибул, 2 пары
ножек
|
двуветвистые,
плавательные, без жабр
|
9 сегм + тельсон,
ног нет, есть вилочка
|
Веслоногие
Copepoda
|
головогрудь
(голова + 1 гр. сегм.), грудь
(5 сегм.)
|
1 науп-лиальный
|
3 пары челюстей
|
1 пара — ногочелюсти, 4—5 пар — двуветвистые без жабр |
4 сегм.+ тельсон, ног нет, есть вилочка |
Усоногие
Cirri pedia
|
Отделы сдиты без сегментов |
нет |
нет |
4—6 пар - усовидные без жабр |
Брюшко недоразвито |
| Карповые вши Branchiura |
головогрудь (голова + 1 сегм.), грудь (4 сегм.), брюшко |
2 фасеточных, Знауплиальных |
1 пара — стилеты, 2 — присоски, 3— прикрепительные |
4 пары — двуветвистые плавательные без жабр |
брюшко слитное, ног нет |
Ракушковые Ostracoda
|
голова, грудь(2—3 сегм.), брюшко
слитное
|
фасеточ
ные, простой глазок, часто редуцированы
|
1 пара мандибул, 2 пары максилл |
2—3 пары ходильные, без жабр |
слитное, ног нет, есть вилочка
|
Высшие раки Malacostгаса
|
протоцефалон, челюстегрудь или головогрудь (голова + 1 гр. сегм.), грудь (7
сегм.), брюшко
(6—7 сегм.)
|
фасеточные
|
3 пары челюстей + 1—3 пары ногочелю-
стей
|
1—2 пары ногочелюстей, 5-7 пар ходиль-ных или плавательных ног
|
6—7 сегм.
+ тельсон, 3—6 пар ног, вилочки нет (у Nebalia брюшко иногда
рудуцировано)
|
У представителей подклассов веслоногих, усоногих и карповых вшей грудные ножки в дыхании не участвуют, брюшных ножек нет, отсутствуют также жабры, кровеносная система и фасеточные глаза. Метаморфоз у них сложный. Кроме свободноживущих форм есть сидячие и паразитические. Представители веслоногих — циклопы (Cyclops), карповых вшей — карпоеды (Arqulus), усоногих — морские желуди (Balanus). Мелкие пресноводные циклопы являются промежуточными хозяевами гельминтов.
Для подкласса ракушковых характерно наличие двустороннего головогрудного щита, полностью скрывающего тело животного. Организация упрощена.
Подкласс высшие раки включает наиболее высокоорганизованных ракообразных (табл. 21). У них определенное число сегментов тела, имеются брюшные ножки и сегментация более гетерономная, чем у других ракообразных. Последняя пара брюшных ножек — уроподы — у высших раков часто сильно видоизменена. Развитие протекает с метаморфозом или прямо. Представители — мизиды (отряд Mysidacea), мокрицы (отряд Oniscoidea), бокоплавы (отряд Amphipoda), речные раки (семейства Astacidae и Parastacidae), крабы (Brachiura — отдел Reptantia из отряда Decapoda). Многие виды имеют промысловое значение (например, камчатский краб, омар, речные раки) и служат пищей для рыб и беззубых китов.
Таблица 21.
Морфологическая характеристика некоторых отрядов высших раков
Отряды
|
Расчленение тела
|
Конечности головы
|
Конечности грудные
|
Конечности брюшные
|
Расположение жабр
|
Расположение сердца
|
Тонкопанцирные Leptostraca
|
Протоцефалон, челюстегрудь (3 гол., 8 гр., 7 брюш. сегм.) |
2 пары длинных антенн, 3 пары челюстей
|
8 пар — листовидные с жабрами
|
4 пары — плавательные, 2 пары рудимент. +
вилка
|
на 1-8 парах грудных
НОГ
|
в груди |
Ротоногие
Stomatopoda
|
протоцефалон,
челюстегрудь (3 гол. + 4 гр.
сегм.), грудь (4 сегм.), брюшко
(6 сегм.)
|
антеннулы длиннее антенн, 3 пары
челюстей
|
I пара — чувствующие, II—
V — хватательные, VI-VIII - ходильные (жабры на
1—5 парах)
|
I—V пары—
листовидные жаберные, VI — уроподы плавательные
|
HaI-Vпарах
Грудных ног и I-V парах
брюшных ног
|
в груди
и на брюшке
|
Равноногие Isopoda
|
лова + 1 гр. сегм.), грудь (6—7 сегм.), брюшко (6 сегм. или слит
ное)
|
2 пары антенн (вторая длиннее), 3 пары челюстей
|
челюстей, II-VIII пар — хо-дильные, II пара — хвататель
ные
|
пластинчатых дыхательных, VI — уроподы членистые
|
HaI-V парах брюшных ног
|
на брюшке
|
Окончание табл.21
Отряды
|
Расчленение тела
|
Конечности головы
|
Конечности грудные
|
Конечности брюшные
|
Расположение жабр
|
Расположение сердца
|
| Бокоплавы Amphipoda |
головогрудь
(голова + 1-2 гр. сегм.), грудь (6-7 сегм.), брюшко (6 сегм.)
|
2 пары антенн, 3 пары челюстей |
1—11 пары —
ногочелюсти, III—VIII - плавательные и ходильные, все пары с жабрами
|
I-III -лаватель
ные, IV— V — пры-гательные, VI —уроподы прыгательные
|
на III-VIII
парах грудных ног (6 пар)
|
в груди |
Десятиногие
Decapoda
|
протоцефалон,
челюстегрудь (3 гол. + 8 гр.
сегм.), брюшко
(6 сегм.)
|
2 пары антенн, 3 пары
челюстей
|
3 пары ногочелюстей, 5 пар
ходильных ног с жабрами
|
5 пар плаватель
ных ног + уроподы плавательные
|
на 8 парах
грудных ног
|
тоже
|
Тело разделяется на головогрудь и брюшко. Степень расчленения тела различная: иногда сегменты груди свободны, чаще слиты, брюшко расчлененное или слитное и т. д. Вторая пара конечностей — педипальпы — гомологична мандибулам ракообразных. Остальные 4 пары головогрудных конечностей представляют собой ходильные ноги. Брюшные конечности есть у водных хелицеровых (мечехвосты) и отсутствуют в типичном виде у наземных (паукообразные). Гомологами брюшных ног являются половые крышечки, легкие и паутинные бородавки.
Подтип включает 2 современных класса: меростомовые, или мечехвосты (Merostomata, или Xiphosura), и паукообразные (Arachnoidea) (табл. 22).
В отличие от рассматриваемого далее обширного класса паукообразных меростомовые представляют собой древнюю группу в основном вымерших морских хелицеровых, из которых в настоящее время сохранилось только пять видов.
Таблица 22.
Сравнительная характеристика классов хелицеровых
| Признаки
|
Классы
|
| Merostomata
|
Arachnoidea
|
| Отделы тела |
головогрудь + брюшко |
головогрудь + брюшко |
| Строение и функции хелицер |
3-члениковые |
2-члениковые клешневидные |
| Строение педипальп |
— |
строение и функции разнообразны |
| Ходильные ноги |
5 пар, оканчиваются клешнями, локомоторная функция, участвуют в питании |
4 пары, локомоторная функция, появляется членистая лапка |
Окончание табл. 22
| Признаки
|
Классы
|
| Merostomata
|
Arachnoidea
|
| Тип питания |
хищники |
хищники, фитофаги, паразитические |
| Органы выделения |
коксальные железы |
коксальные железы, мальпигиевы сосуды |
| Органы дыхания |
жабры |
легкие, трахеи |
| Органы кровообращения |
трубчатое сердце, 8 пар остий |
трубчатое сердце, 1—2—7 пар остий |
| Нервная система |
цепочная: головной мозг + 6 ганглиев брюшной нервной цепочки |
цепочная: головной мозг + 6—7 брюшных нервных ганглиев |
| Оплодотворение |
наружное |
наружно-внутреннее, внутреннее |
| Развитие |
прямое |
прямое, у клещей нередко с метаморфозом |
Класс паукообразных объединяет около 40 тыс. наземных хели-церовых, являющихся одними из наиболее древних обитателей суши (рис. 35).
Рис. 35. Выход хелицеровых на сушу (схема):
I —
первичноводные хелицеровые; 2 —
ночные бродячие хищники (скорпионы, жгутоногие, сольпуги, низшие пауки и др.); 3 —
устройство норок (норковые пауки и др.); 4 —
выход пауков из норки с паутинной выстилкой-трубкой; 5 — заселение растительности высшими тенетными пауками; 6 —
полет пауков на паутине и подъем клещей токами воздуха; 7— прямое освоение дневной поверхности суши (некоторые сенокосцы, пауки и др.); 8 —
переход из воды к жизни в скважинах почвы (кенении, низшие клещи); 9 —
заселение гниющих органических субстратов (многие клещи, ложноскорпионы и др.); 10—14 —
выход клещей из почвы и гниющих остатков в качестве паразитов животных и растений, пассивное расселение на хозяевах, по воздуху и иными путями; 75 — водяные клещи
Это хищники, паразиты (кровососы) и растительноядные животные.
Тело паукообразных чаще всего состоит из головогруди и брюшка. Головогрудь образована акроном и 7 сегментами, брюшко — 12, однако от этой схемы есть отклонения. Встречается более расчлененное тело, а у большинства пауков, наоборот, головогрудь и брюшко — нерасчлененные отделы. Головогрудь с 6 парами конечностей (хелицеры, педипальпы и 4 пары ходильных ног). Хелицеры расположены впереди рта, они похожи на короткие клещи (сольпуги, скорпионы, сенокосцы и др.), реже с коготком (пауки) или имеют вид стилетов для прокалывания кожи. У скорпионов педипальпы превращены в мощные клещи. Ходильные ноги состоят из 6-7 члеников и заканчиваются коготком (рис. 36).
Рис. 36. Строение скорпиона:
А —
со спинной стороны; Б —
с брюшной стороны; / — переднебрюшие; // — заднебрюшие; 1 —
хвостовой членик с ядовитой иглой; 2 —
головогрудь; 3 —
хелицеры; 4 —
педипальпы; 5 — ходильные ноги; 6 — дыхальца легких
Кожный покров — кутикула, под которой находятся гиподер-мальный эпителий и базальная мембрана. Кутикула трехслойная и хорошо защищает тело от испарения воды. Кожные железы включают ядовитые и паутинные.
Передний отдел кишечника (глотка) снабжен сильными мышцами и выполняет роль насоса. Средняя кишка обычно с боковыми выпячиваниями, увеличивающими ее вместимость. Печень парная. Секрет печени и слюнных желез энергично расщепляет белки. Он вводится в тело убитой добычи и превращает ее содержимое в полужидкое состояние, а затем уже происходит всасывание этой пищи при помощи глотки. Такое явление получило название вне-кишечного пищеварения.
Органы выделения у примитивных форм двоякой природы: есть целомодукты и мальпигиевы сосуды, которые открываются в кишечник (скорпионы). У большинства паукообразных (рис. 37—39) остаются только мальпигиевы сосуды.
Рис. 37. Книжный ложноскорпион Рис. 38. Домовой паук
(Chelifer cancroides) (Tegenaria domestica)

Рис. 39. Водянойпаук (Argyroneta aquatica)
Кровеносный аппарат незамкнутый и включает сердце, аорту и сосуды. У мелких форм может оставаться только сердце с парой остий, но может исчезать и оно.
Органы дыхания — легочные мешки и трахеи. Легкие представляют собой производное жабр водных предков, а трахеи являются новым приобретением, связанным с переходом к жизни на суше.
Нервная система происходит от брюшной нервной цепочки, обнаруживает тенденцию к концентрации. Ее метамерность лучше всего выражена у скорпионов и сольпуг. У клещей ганглии сливаются в общую массу. Органы чувств разнообразны. Органами механического чувства являются чувствительные волоски — трихо-ботрии, расположенные преимущественно на педипальпах. Они регистрируют колебание воздуха. Для обоняния служат так называемые лировидные органы — щели в кутикуле, ко дну которых подходят нервные окончания. Глаза паукообразных простые, обычно собранные в кучки, реже их всего два.
Паукообразные раздельнополы. Отмечается разная степень слияния гонад. Половое отверстие находится на первом сегменте брюшка. Оплодотворение внутреннее, причем осеменение может быть сперматоформным или наступает при копуляции. Представители класса в основном яйцекладущие, но отдельные скорпионы и клещи живородящие. Дробление яиц частичное, поверхностное. Развитие прямое, кроме клещей, у которых наблюдается метаморфоз.
Насчитывается свыше 10 отрядов паукообразных, из которых наиболее распространены и известны скорпионы, сольпуги, пауки, сенокосцы и ряд отрядов, объединяемых под общим названием «клещи» (рис. 40—43).
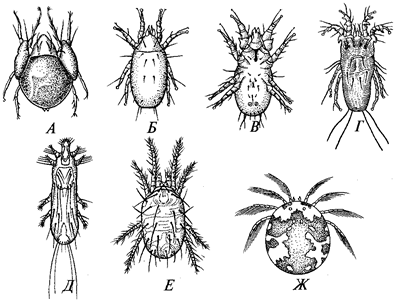
Рис. 40. Различные клещи:
А —
панцирный клещ Galumnamucronata; Б —
мучной клещ Tyroglyphusfarinae, самка; В —
то же с брюшной стороны; Г —
перьевой клещ с воробья Analgopsispasserinus, самка; Д—
паразитический клещ Syringophilusbipectinatus, живущий в очине перьев птиц; Е —
обыкновенный паутинный клещ Tetranychustelarius, самка; Ж —
водяной клещ Hydrarachnageographica

Рис. 41. Различные клещи:
1 —
четырехногий клещ Eriophyes; 2 —
водяной клещ Arrhenurusneumani; 3 —
чесоточный зудень (Sarcoptesscabiei), самка с брюшной стороны; 4 —
красно-телка Eutrombiculabatatas, хищная самка; 5 — вредитель злаков Siteroptegraminun
Рис. 42. Клещи — переносчики заболеваний:
А —
таежный клещ Ixodespersulcatus в позе ожидания животного-хозяина, самка; 5 — иксодовый клещ Dermacentorpictus, самка; В —
аргазовый клещ Omithodoruspapillipes, самка

Рис. 43. Схема жизненных циклов иксодовых клещей:
1 — треххозяинный цикл; 2 —
двуххозяинный цикл; 3 —
однохозяинный цикл; О —
яйцо; L
—
личинка; N —
нимфа; / — имаго
Практическое значение паукообразных очень велико. Настоящие пауки, питаясь насекомыми, уничтожают громадное количество вредителей. Некоторые клещи (иксодовые и аргасовые) переносят возбудителей тяжелых заболеваний человека и домашних животных (клещевой тиф, энцефалит, пироплазмоз рогатого скота и т. д.). Есть среди них вредители растений, особенно из группы паутинных клещей, а также запасов зерна (тироглифоидные клещи). Большинство почвенных клещей — полезные сапрофаги.
Таблица 23.
Сравнительная характеристика отрядов класса паукообразных
| Признаки
|
Отряды
|
| Скорпионы
|
Жгутоногие
|
Сольпуги
|
Лжескорпионы
|
Сенокосцы
|
Пауки
|
Клещи
|
| Число сегментов |
головогрудь слитная+ 12 брюшных |
головогрудь слитная+10 брюшных |
перед.отдел слитный + 2 грудинных + 10 брюшных |
головогрудь слитная +11 брюшных |
головогрудь слитная + 9 — 10 брюшных |
головогрудь и брюшко слитные +1 между ними |
слитные у большинства |
| Строение и функции хелицер |
клешневидные, схватывают добычу |
клешневидные |
3-членные двойные клешневидные |
клешневидные 3-членные |
клешневидные 2-членные |
клешневидные 2-член-ные |
клешневидные, входят в сосательный хоботок |
Окончание табл.23
| Признаки
|
Отряды
|
| Скорпионы
|
Жгутоногие
|
Сольпуги
|
Лжескорпионы
|
Сенокосцы
|
Пауки
|
Клещи
|
| Строение и функции педипальп |
то же |
клешневидные или хватательные |
как ходильные ноги, локомоторная функция |
клешневидные, схватывают добычу |
осязательная функция |
щупальцевидные, осязательная, копуляти-вная, жевательная функции |
щупальце-видные, входят в состав гипо-стома |
| Ходиль-ные ноги |
4 пары, локомоторная функция |
4 пары (1 — осязат. придатки) |
4 пары |
4 пары |
4 пары |
4 пары |
4 пары |
| Наличие желез |
ядовитая железа на тельсоне |
ядовитые |
нет |
паутинные |
нет |
ядовитые и паутинные |
паутинные у паутинных клещей |
| Дыхание |
легкие (4 пары) |
легкие (2 пары) |
трахеи |
трахеи |
трахеи |
легкие (1—2 пары) + трахеи |
трахеи или всей поверхностью тела |
| Развитие |
прямое, живорождение |
прямое |
прямое |
с метаморфозом, живорождение |
обычно прямое, иногда с элементами метаморфоза |
прямое |
с метаморфозом (личинка, нимфа) |
Представители подтипа - наземные членистоногие дышащие при помощи трахей (рис. 44, 45). Характеризуются тем что голова образована путем слияния головной лопасти (акрона) с усиками (антеннулами) и 3 или 4 первыми сегментами несущими ротовые придатки. Антенн нет, и несущий их сегмент (вставочный) может редуцироваться. Ротовые придатки представляют собой видоизмененные конечности (мандибулы, две пары мак-силл -максиллы I и максиллы II, нижняя губа - вторые мак-силлы). Сегментарный состав туловища разнообразен. Хорошо адаптированы к жизни на суше в условиях дефицита влажности к трахейным относятся 2 надкласса: многоножки (Myriapoda) и шестиногие (Hexapoda).


Рис. 44. Гигантская сколопендра Рис. 45. Костянка Lithobius
(Scolopendragigantea),
нападающая на ящерицу
К надклассу многоножек, насчитывающему свыше 53 тыс видов, относятся членистоногие, тело которых состоит из головы и Длинного сегментированного туловища со слабо выраженной дифференциацией. На всех или почти на всех сегментах имеются ноги.
В надкласс многоножек входят 4 класса: симфилы (Symphyla), пауроподы (Pauropoda), двупарноногие, или диплоподы (Diplo-poda), и губоногие, или хилоподы (Chilopoda), краткая характеристика которых приведена в таблице 24.
Надкласс шестиногих объединяет 2 класса: скрыточелюстные насекомые (Insecta-Entognatha) и открыточелюстные насекомые (Insecta-Ectognatha), которых ранее рассматривали как единый класс насекомых (Insecta).
Таблица 24.
Сравнительная характеристика классов многоножек
Классы,
образ жизни, число видов
|
Особенности
|
| Состав головы
|
Головные придатки, конечности
|
Глаза
|
Сегментный состав туловища
|
Туловищные конечности и придатки
|
Трахеи
|
Половые отверстия
|
Оплодотворение, развитие
|
Симфилы Symphyla, сапрофаги, 150 видов
|
акрон + 4 сегмента
|
усики, мандибулы, максиллы, гнатохиля-
рий
|
нет
|
15—22 сегм.+ тельсон
|
12 пар с кок-
сальными органами (III-XII пара), церки с паутинными железами
|
1 пара на голове
|
1 непарное у III пары ходильных ног
|
спермато-форное, анаморфоз
|
Пауроподы Pauropoda, сапрофаги,
350 видов
|
акрон + 4 сегм. (IV - свободный
шейный
|
усики ветвистые, мандибулы, гнатохиля-
рий
|
то же
|
10 сегм.+ тельсон
|
9 пар ног
|
нет
|
парные отверстия у II пары ходильных
ног
|
анаморфоз[1]
|
Диплоподы Diplopoda, сапрофаги, 12000
видов
|
то же
|
усики, мандибулы, гнатохиля-рий
|
простые, реже нет
|
30—75 сегм. + тельсон
|
на I—III сегм.
по 1 паре ног, с IV — по 2 пары ног (кроме тельсона), до
139 пар ног
|
на I—I1I сегм по паре, с ГУ-по 2 пары стигм
|
парные на II тулов.сегм.
|
спермато-форное, анаморфоз
|
Губоногие Chilopoda,
хищники 2800 видов
|
то же
|
усики, мандибулы, максиллы, максиллы
|
простые
|
до 181 сегм.+ тельсон
|
до 177 пар;
1 пара — ного-челюсти с ядом, поел. 1—2 пары —
половые ножки
|
по паре стигм на туловище через сегмент
|
1 непарное на последнем сегм.
|
спермато-форное, прямое или анаморфоз
|
(
INSECTA-ENTOGNATHA)
Класс включает 3 отряда: бессяжковые (Protura), ногохвостки (Collembola) и двухвостки (Diplura) (табл. 25). Как и остальные насекомые, скрыточелюстные обладают 3 парами ног, а их тело разделяется на голову, грудь и брюшко. Ротовые органы погружены в головную капсулу. Крылья отсутствуют. Скрыточелюстные — в основном мелкие насекомые, обитающие в почве и подстилке, а некоторые — на растениях. Многие участвуют в почвообразовании, а ряд видов ногохвосток — вредители овощных культур.
Таблица 25.
Сравнительная характеристика скрыточелюстных насекомых
Отряды
(представи
тели), число видов
|
Антенны
|
Глаза
|
Ротовой
аппарат
|
Грудные
ноги
|
Брюшко
|
Придатки
брюшка
|
Дыхание
|
Развитие
|
Бессяхко-
вые Protura (Eosentomon), 300
видов
|
нет
|
нет
|
внутренний
колюще-сосущий, стилето-
видные
мандибулы
|
1 пара
чувствующих, 2-3
пары —
ходиль-
ных
|
11 сегм.+ тельсон
|
на I-III
бр. сегм. рудименты
ног
|
кожное
или
трахейное, 2 пары стигм
на груди
|
анаморфоз
|
Ногохвост-
КиCollem-bola (Ony-
chiurus,
Podura),
200 видов
|
4-6-члени
ковые
|
скопления
простых глазков
или
их нет
|
внутренний
(грызущий), ротовые части
стилето-
видные
колюще-
сосущие
|
3 пары ходильных
|
6 сегм., часто слитное
|
прыга-
тельная
членистая вилочка и
брюшная
трубка
|
кожное
|
прямое
|
Двухвостки Diplura (Campodea, Japyx), 400
видов
|
много-члени-ковые
|
нет
|
внутренний
(грызущий), мандибулы саблевидные
|
3 пары ходиль-ных
|
10 сегм.+ тельсон
|
на пер
вых. бр.
сегм. рудименты ног, на заднем— церки членистые или клешневидные
|
кожное
или 3— 4 пары стигм на груди
|
с элемента
ми метаморфоза (протомор-
фоз)
|
(
INSECTA-ECTOGNATHA)
Открыточелюстные насекомые — самая большая группа животных (свыше 1 млн видов), распространенных по всему земному шару и живущих во всех средах, кроме морской.
Тело разделено на голову, грудь и брюшко (рис. 47). Голова одета общей хитиновой капсулой и несет на себе усики (гомологичные антеннулам других членистоногих, но в пределах класса именуемые антеннами); пару сложных глаз, между которыми могут быть несколько простых глазков; пару мандибул; пару мак-силл и нижнюю губу — результат слияния второй пары максилл (рис. 46). Строение ротового аппарата разнообразно и соответствует характеру питания. Грудной отдел несет 3 пары ног, также имеющих разное строение: ходильные, прыгательные, плавательные и пр. У всех крылатых на спинной стороне второго и третьего сегментов груди расположены 2 пары крыльев, представляющих собой выпячивания кожных покровов, а не настоящие конечности. На брюшке конечностей нет, однако у ряда насекомых встречаются их рудименты.
Рис. 46. Голова насекомого:
А —
вид спереди; Б —
вид сбоку (из Кенигсмена): 1 —
головная капсула; 2 —
простые глазки; 3 —
фасетированный глаз; 4 —
антенны (сяжки); 5 — мандибула; 6 —
верхняя губа; 7 — место прикрепления сяжка; 8 —
нижняя губа; 9 —
нижняя челюсть
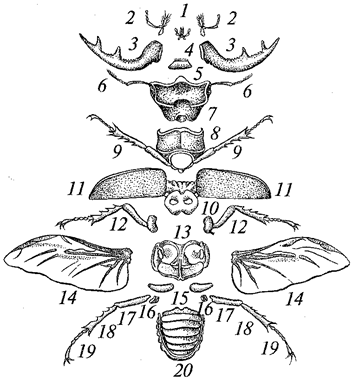
Рис. 47. Расчлененный самец жука-оленя (Lucanuscervus):
1 —
нижняя губа с щупиками; 2 —
нижние челюсти с щупиками; 3 —
жвалы (верхние челюсти); 4 —
верхняя губа; 5 — наличник; 6 —
усики; 7 — голова; 8 —
переднегрудь; 9 —
передние ноги; 10 —
среднегрудь; 11
— надкрылья; 12 —
средние ноги; 13 —
задне-грудь; 14 —
крылья; 15 —
тазики задних ног; 16—
вертлуги; 17—
бедра; 18—
голени; 19—
членистые лапки с коготками; 20 —
брюшко
Кожные покровы насекомых принципиально сходны с покровами паукообразных. В коже залегают пигменты — красящие соединения, обусловливающие окраску и рисунок насекомых. Иногда окраска зависит от структуры кожи (оптическая окраска). С кожными покровами связаны и различные железы — восковые, пахучие, прядильные, ядовитые и т. д., секреты которых играют важную роль в жизнедеятельности насекомых.
Хитиновая кутикула является наружным скелетом, к которому изнутри прикрепляется сложная мускулатура. Мышцы поперечно-полосатые. Особенного развития достигают грудные мышцы, приводящие в движение крылья.
Пищеварительная система образована кишечником, состоящим из 3 отделов, как у высших червей и других членистоногих. Печеночные выросты отсутствуют. Модификации кишечника, особенно переднего отдела, связаны с характером пищи. У насекомых, сосущих жидкую пищу, ротовые придатки видоизменены в хоботок и ротовая полость превращена в систему каналов. Слюнные железы нередко модифицированы в другие органы (прядильные железы гусениц бабочек) или содержат в составе секрета особые вещества (антикоагулянты у кровососущих форм и др.). Продукты пищеварения превращаются и всасываются в средней кишке, эпителий которой вырабатывает ферменты и бесструктурную перитро-фическую мембрану, облекающую в виде чехла пищевой комок. Задняя кишка имеет так называемые ректальные железы, с помощью которых происходит всасывание воды из переработанной пищи. На границе средней и задней кишок наподобие тонкостенных трубчатых выпячиваний отходят мальпигиевы сосуды — органы выделения насекомых. В качестве экскрементов через мальпигиевы сосуды в кишечник выделяются кристаллы мочевой кислоты.
Для дыхания служит сложная система трахей. Через отверстия на боку сегментов — стигмы — воздух проходит в трахеи и их разветвления — трахеолы, поступая к органам и тканям тела. Кровеносная система незамкнутая и состоит из сердечного сосуда и системы сосудов меньшего диаметра. У мелких форм, как обычно, наступает редукция сосудов. Гемолимфа, циркулирующая в сосудах и синусах, выполняет разнообразные функции: прежде всего снабжение тканей и органов питательными веществами и перенос продуктов обмена к органам выделения, при этом дыхательная функция ее развита меньше всего.
Нервная система, как у других членистоногих, исходно построена по типу брюшной нервной цепочки, но часто достигает высокого уровня развития. Центральная нервная система состоит из головного мозга, подглоточного ганглия и сегментарных ганглиев брюшной нервной цепочки. Кроме того, имеется симпатическая (регулирующая работу мышц внутренних органов) и периферическая нервная система. Число ганглиев брюшной нервной цепочки может уменьшаться за счет их слияния.
У насекомых есть органы осязания, слуха, обоняния, вкуса и зрения. Основу органов чувств составляют сенсиллы — нервно-чувствительные единицы, которые нередко собираются в комплексы, образуя орган. По типу простых сенсилл сформированы органы механического и химического чувства (осязания, обоняния и вкуса). Органы зрения у взрослых насекомых представлены сложными, или фасеточными, глазами, к которым иногда присоединяются немногочисленные простые глазки на лбу. Органы слуха имеются далеко не у всех насекомых. Особенно хорошо развиты они у кузнечиков, сверчков, саранчовых и цикад, т. е. у насекомых, которые обладают звуковыми органами и могут издавать звуки. Органы слуха подобных насекомых называются тимпанальными, они состоят из скопления сенсилл (сколпофоров), связанных с утонченными в виде барабанной перепонки участками кутикулы. Отверстия тимпанальных органов сообщаются с внешней средой.
В целом система воспринимающих рецепторов у насекомых очень сложна и достигает высокого совершенства. Этим обусловлено их сложное поведение.
Насекомые раздельнополы, при этом обычно существует резкий половой диморфизм. Половые железы парные, у самцов нередко имеется совокупительный орган. Осеменение у форм без совокупительного органа сперматофорное. Яйца богаты желтком, поэтому дробление частичное, поверхностное. Развитие делится на эмбриональное и постэмбриональное. Последнее проходит с метаморфозом, и лишь у одной группы примитивных насекомых есть анаморфоз.
Различают неполный метаморфоз (группа Hemimetabola) (табл. 26), при котором личинка похожа на взрослое насекомое (имаго) И метаморфоз происходит постепенно, и полный (группа Holome-tabola), при котором личинки червеобразные и существует куколочная стадия (табл. 27). У насекомых с полным превращением в зависимости от степени развития ног различают 4 типа личинок: протоподные (есть только зачатки грудных ног), олигоподные (3 пары нормально развитых ног), полиподные (3 пары грудных ног и несколько пар брюшных ложных ножек) и аподные (безногие). В регуляции жизненного цикла принимают участие нейросекреторные клетки мозга и эндокринные железы.
Таблица 26.
Обзор некоторых отрядов насекомых с неполным превращением
| Отряды
|
Ротовой аппарат
|
Крылья
|
Тип метаморфоза
|
Личинки
|
Важнейшие семейства
|
Прямокрылые Orthoptera
|
грызущий
|
I пара — кожистые, II — тонкие
|
Типичный неполный метаморфоз (личинка похожа на имаго) |
нимфа
|
саранчовые, сверчковые, медведковые, кузнечиковые |
Таракановые
Blattoptera
|
то же
|
I пара — кожистые, II — часто
редуцирована
|
то же
|
тоже
|
Блаттиды и др.
|
Окончание табл. 26
| Отряды
|
Ротовой аппарат
|
Крылья
|
Тип метаморфоза
|
Личинки
|
Важнейшие семейства
|
Полужесткокрылые
Hemiptera
|
колющесосущий (членис
тый хоботок)
|
I пара — полужесткие, II —
перепончатые
|
то же
|
то же
|
хищнецы, слепняки, щитники,
водомерки, гладыши, постельные
клопы
|
Равнокрылые Homoptera
|
колющесосущий (нечленистый хоботок)
|
2 пары
стекловидных со слабым жилкованием
|
тоже
|
тоже
|
тли, цикады, цикадки, листоблошки, червецы,
щитовки
|
Стрекозы Odonata
|
грызущий
|
стекловидных с сетчатым жил-
кованием
|
заметно отличается от имаго
|
наяда
|
красотки, настоящие стрекозы,
коромысла
|
| Вши Anoplura |
колющесосущий (втягивающийся хоботок) |
бескрылые |
Личинка почти не отличается от имаго (гипомор-фоз) |
имагообразная |
педикулиды |
Таблица 27.
Обзор некоторых отрядов насекомых с полным превращением
| Отряды
|
Ротовой аппарат
|
Крылья
|
Личинки
|
Куколки
|
Важнейшие семейства
|
Жесткокрылые Coieoptera
|
грызущий
|
надкрылья и крылья
|
олигоподные
|
открытые
|
жужелицы, плавунцы,
пластинчатоусые, щелкуны, листоеды, долгоносики, усачи,
короеды, божьи коровки
|
| Двукрылые Diptera |
лижущий, колюще-сосущий |
1 пара крыльев, II превращена в жужжальца |
аподные |
ложный кокон-пупарий |
комары, мухи, слепни, тахины, журчалки, оводы |
| Перепончатокрылые Hymeno-ptera |
грызущий |
2 пары перепончатых крыльев |
аподные, протопод-ные, полиподные |
открытые, в коконах |
муравьи, пилильщики, пчелы, осы, наездники, рогохвосты |
| Ручейники Trichoptera |
сосущий |
2 пары крыльев с волосками |
олигопод-ные |
открытые |
полицентро-пиды, гидропсихиды |
Окончание табл.27
| Отряды
|
Ротовой аппарат
|
Крылья
|
Личинки
|
Куколки
|
Важнейшие семейства
|
| Чешуекрылые Lepidoptera |
то же |
2 пары крыльев с чешуйками |
полиподные |
покрытые (в коконах) |
шелкопряды, совки, пяденицы, нимфалиды, белянки, голубянки |
| Блохи Aphaniptera |
колюще-сосущий |
бескрылые |
олигоподные |
открытые |
пулециды |
Кроме обоеполого размножения у насекомых известны случаи партеногенеза — обязательного, факультативного или циклического. При чередовании обоеполого и партеногенетического размножения имеет место правильная гетерогония. Размножение на стадии личинок (педогенез) зарегистрировано у двукрылых.
Значение насекомых в природе огромно, поскольку они принимают участие в различных биологических процессах — опылении цветковых растений, почкообразовании, разложении растительного опада, уничтожении трупов и экскрементов других животных и т. д. Сами насекомые служат пищей для птиц, амфибий, насекомоядных зверей и др.
Среди них немало вредных форм. Общеизвестно, что кровососущие насекомые переносят возбудителей серьезных заболеваний человека: комары — малярии, вши — сыпного тифа, блохи — чумы и т. д. (табл. 28). Среди растительноядных форм много вредителей сельскохозяйственных растений и древесных пород (колорадский жук, огородные блошки, короеды, сосновый шелкопряд, капустница, кровяная тля и сотни других видов), с которыми человеку приходится вести неустанную борьбу (табл. 29). Продукция ряда видов насекомых служит сырьем для промышленности или пищей для человека. Таковы медоносная пчела, дающая мед, воск и другие продукты, тутовый шелкопряд — производитель шелка, лаковые червецы, дающие лак и воск.
Таблица 28.
Насекомые — паразиты и переносчики заболеваний человека
| Паразиты и переносчики
|
Экологическая группа
|
Вызываемые или переносимые заболевания
|
Меры профилактики
|
Меры борьбы
|
Вши рода Pediculus
|
эктопаразиты (облигатные)
|
тифы (возвратный, сыпной)
|
личная гигиена
|
Уничтожение гнид, вшей, стрижка волос, термическая и химическая обработка одежды |
Блоха обыкновенная Pulex irritans
|
эктопаразит (факультативный)
|
чума
|
Чистота жилых помещений, обработка щелей в полах |
химическая борьба с блохами и с грызунами |
Окончание табл. 28
| Паразиты и переносчики
|
Экологическая группа
|
Вызываемые или переносимые заболевания
|
Меры профилактики
|
Меры борьбы
|
Клоп постельный Cimex lectularis
|
эктопаразит
(облигатный)
|
кожная аллергия
|
чистота помещений
|
Ремонт помещений,
обработка мебели ядохимикатами
|
Комары рода
Anopheles
|
кровососы
|
малярия, слоновая болезнь
|
изоляция и излечение больных, ащита от укусов комаров |
уничтожение комаров, лечение больных |
Москиты
рода Phlebotomus
|
кровососы
|
чума, лейшманиоз
|
защита от
укусов москитов
|
уничтожение
москитов и очагов зараженных зверьков
|
Мухи кровососу
щие (слепни, наст. мухи) Tabanidae, Muscidae
|
тоже
|
Сибирская язва
трипаносомозы, туляремия
|
защита от укусов мух
|
уничтожение москитов
|
Таблица 29.
Насекомые — вредители растений и технических материалов
| Повреждаемые растения и материалы
|
Систематический состав вредителей
|
Наиболее опасные вредители в России
|
| Полевые культуры |
саранчовые, равнокрылые, хоботные, полужесткокрылые, жесткокрылые, двукрылые, чешуекрылые |
азиатская саранча, клоп-черепашка, колорадский жук, свекловичный долгоносик, луковая муха, капустная белянка, луговой мотылек, хлебные жуки |
| Плодово-ягод-ные культуры |
чешуекрылые, жесткокрылые, пилильщики, равнокрылые, хоботные |
непарный шелкопряд, кольчатый шелкопряд, белая американская бабочка, плодожорки, цветоеды, филлоксера, яблонная моль |
| Лесные насаждения |
равнокрылые, хоботные, чешуекрылые, жесткокрылые, пилильщики, орехотворки, двукрылые |
короед-типограф, черный еловый усач, березовый заболонник, стеклянницы, сосновый побеговьюн, непарный шелкопряд, белая американская бабочка, сосновый пилильщик |
| Зерновые запасы |
чешуекрылые, жесткокрылые, перепончатокрылые |
жуки-хрущаки, зерновая моль |
| Технические материалы |
чешуекрылые, жесткокрылые |
шубная моль, кожееды, жуки-точильщики |
Классификация насекомых базируется на характере сегментации, строении ротовых конечностей и крыльев, а также характере метаморфоза. К открыточелюстным насекомым относится свыше 30 отрядов. Отряд щетинохвосток (Thysanura) часто относят к отдельному подклассу первичнобескрылых (Apterygota), объединяя остальные отряды в подкласс крылатых (Pterygota). Основные из них — стрекозы, таракановые, термиты, прямокрылые, вши, полужесткокрылые (клопы), жесткокрылые (жуки), сетча-токрылые, чешуекрылые (бабочки), перепончатокрылые (пчелы, осы, муравьи), блохи, двукрылые (мухи и комары).
Далее будут рассмотрены наиболее важные или широко распространенные отряды насекомых.
Крупные хищные насекомые с очень подвижной головой, несущей пару огромных глаз. Ротовые органы грызущие. Две пары крыльев имеют сходное сетчатое жилкование. У Крыловой мускулатуры отсутствуют аналоги среди других отрядов, так как она лишена продольных спинных мышц. Личинки водные с ректальными или хвостовыми жабрами. Около 4,5 тыс. видов.
Тело плоское, голова сверху прикрыта переднегрудным щитком. Ротовой аппарат грызущий. Передние крылья хитинизирова-ны и скрывают задние, которые складываются обычно веерообразно. Иногда крылья редуцированы. Всеядные, большей частью ночные насекомые, живущие под опавшей листвой или в почве. Некоторые виды синантропны и живут в домах. Около 3,5 тыс. видов, особенно широко распространены в тропиках. Представители — черный таракан (Blattaorientalis), прусак (Blattagermanica).
Общественные насекомые с резко выраженным полиморфизмом, близки с Таракановыми. Голова свободная, ротовые части грызущие. Крылья перепончатые, свойственные лишь самцам и самкам, способным к размножению. Живут семьями — колониями, состоящими из самцов, самок, рабочих (недоразвитых в половом отношении самок и самцов), среди которых выделяется специализированная для защиты от врагов каста — солдаты. Питаются древесиной или растительными остатками. Некоторые сильно повреждают деревянные постройки и сооружения. Около 2,5 тыс. видов, главным образом в тропиках.
Крупные или средних размеров насекомые с наклонной головой и большой переднеспинкой. Задние ноги чаще всего прыгательные. Крылья подразделены на узкие подкрылья и задние вееровидно складывающиеся крылья. Ротовые части грызущие. Есть специальные звуковые и слуховые органы. Самка с яйцекладом. Большинство растительноядны, реже хищники. Около 20 тыс. видов. Представители — кузнечики, сверчки, саранча. Многие являются вредителями сельскохозяйственных растений.
Ротовые органы колюще-сосущие (членистый хоботок). Крылья в покое складываются крышеобразно. Самки нередко бескрылые. Питаются клеточным соком растений. Представители — цикады, тли (рис. 48), щитовки и червецы. Большинство — вредители сельскохозяйственных культур и древесных пород, в том числе плодовых.
Рис. 48. Кровяная тля (Eriosomalanigerum):
А —
колония тлей на ветви яблони; Б —
крылатая полоноска; В —
бескрылая форма; Г
и Д —
личиночные стадии
Ротовые органы такие же, как у предыдущего отряда. Крылья в покое складываются плоско. Передняя пара превращена в полунадкрылья (основная часть плотная, кожистая). Заднегрудь часто с пахучими железами. Питаются клеточным соком растений либо являются хищниками или кровососами. Наземные и водные формы, среди которых немало вредителей сельскохозяйственных растений (клоп-черепашка, свекловичный и люцерновый клопы и др.).
Включает более 100 семейств и около 250 тыс. видов. Передние крылья лишены жилкования и превращены в твердые роговидные надкрылья. Ротовые органы грызущие. Задние крылья перепончатые, в покое складываются продольно и поперек, с видоизмененным жилкованием. Ноги бегательные, но нередко специализированные — гребные или прыгательные. Биологически очень разнообразны — хищники, растительноядные, сапрофаги, некрофаги. Живут в почве, на растениях, под корой, в воде и т. д. Много полезных форм (например, божья коровка, хищные жужелицы и др.), но немало и вредителей, таких как картофельный колорадский жук (рис. 49), короеды, зерновки, долгоносики и др.

Рис. 49. Колорадский жук (Leptinotarsadecemlinaeta), его личинка и кладка
Крылья с густой сеткой жилок, задняя пара иногда вытянута в виде узких лент. Усики прикреплены между глазами. Ротовые части грызущие. Личинки с характерно измененными ротовыми частями: мандибулы и максиллы длинные, серповидно изогнутые; мандибулы вдоль нижней стороны имеют желобок, прикрытый лопастью максилл, что приводит к образованию пары сосущих трубок, заостренных спереди. Сосущие трубки вонзаются в тело добычи, через их просвет в нее изливается пищеварительный сок, а затем всасывается разжиженное содержимое тела жертвы. Многие виды приносят большую пользу истреблением вредных насекомых. Таковы златоглазки и гемеробии, используемые в биологический борьбе с вредителями.
Объединяет более 100 тыс. видов. Крылья густо покрыты чешуйками. Ротовые органы без мандибул, обычно превращены в спирально закручивающийся хоботок, при помощи которого бабочки сосут нектар из цветков. Личинки (гусеницы) червеобразные, с брюшными ногами. Взрослые бабочки очень разнообразны по размерам и окраске, нередки крупные, ярко окрашенные формы, но очень много также мелких однотонных видов (например, моли). Гусеницы ряда видов (плодожорка, озимая совка, непарный шелкопряд, капустница и др.) вредят сельскохозяйственным растениям. Тутовый шелкопряд разводится для получения из коконов натурального шелка.
Один из самых крупных отрядов (свыше 150 тыс. видов). Крылья перепончатые, прозрачные, обе пары прочно сцеплены между собой. Ротовые органы грызущие или сосущие, но всегда с мандибулами. Брюшко самки с яйцекладом или жалом. Личинки безногие или гусеницеобразные. Размеры тела от 0,2 мм до нескольких сантиметров.
Грызущий ротовой аппарат характерен для наездников, некоторых ос и муравьев. У высших форм (пчелиных) нижние челюсти и нижняя губа удлинены и превращены в хоботок. В состав заднего отдела груди вошел первый сегмент брюшка, которое соединяется с грудью либо широким основанием, либо с помощью тонкого стебелька.
Образ жизни крайне разнообразен. Среди них есть фитофаги, личинки которых грызут листья, большое число видов питаются нектаром и цветочной пыльцой, не меньшее количество видов являются паразитами других насекомых (наездники, яйцееды). Многие ©сы — охотники за членистоногими. Весьма развиты инстинкт заботы о потомстве и различные формы сложного поведения (рис. 50).
Рис. 50. Перепончатокрылые:
/ — афелинус (Aphelinusmali), заражающий кровяную тлю; 2 —
талса (Thalessalunator), откладывающая яйцо в личинку рогохвоста, вредителя древесных пород; 3 —
пилюльная оса (Eumenes), несущая гусеницу в гнездо-кувшинчик на корм личинкам; 4 —
муравей-листорез (Acromyrmex), несущий кусок листа для устройства грибных садов; 5 — медоносная пчела (Apismeiifera), рабочая особь; 6 —
стеблевой рогохвост (Janusinteger), подрезающий яйцекладом побег смородины
Для человека перепончатокрылые имеют исключительно важное значение как опылители растений. Существенна роль медоносных пчел как производителей меда и воска. Паразитические Осы приносят пользу, истребляя вредных насекомых. Представители — пилильщики, наездники, осы, пчелы, муравьи.
Включает не менее 100 тыс. видов. Насекомые с одной парой Перепончатых передних крыльев. Вторая пара превращена в короткие булавовидные образования — жужжальца. Голова очень подвижна, ротовые придатки наподобие хоботка, приспособленного для сосания или слизывания. У комаров, мошек и слепней ротовой аппарат колюще-сосущий, сходный с хоботком клопов. 6 лижущем хоботке отсутствуют мандибулы, а иногда и максиллы, образован он нижней губой. Грудь очень большая и образована крупным вторым сегментом. Первый и третий сегменты мелкие и тесно слитые со вторым.
Личинки безногие, часть из них с редуцированной головой. Многие питаются разлагающимися веществами, нектаром или пыльцой растений либо являются хищниками. Самки комаров и ^лепней питаются кровью своих жертв, что необходимо для развития их гонад. Среди двукрылых есть и настоящие паразиты, личинки которых развиваются в теле животных и человека.
Практическое значение двукрылых очень велико. Кровососущие двукрылые (мошки, слепни, комары, мокрецы) изнуряют домашних животных и людей, они нередко служат переносчиками заболеваний — малярии, туляремии и др. Домашняя муха переносит возбудителей кишечных заболеваний (рис. 51). Велик ущерб, причиняемый зерновым злакам личинками гессенской и шведской мух. Пользу приносят паразитические мухи — та-хины, снижающие численность вредных насекомых.
Рис. 51. Комнатная муха (Muscadomestica) и ее превращение:
1 —
общий вид насекомого; 2 —
яйца; 3 —
личинка первого возраста; 4 —
взрослая личинка; 5 — пупарий; 6 —
куколка, вынутая из пупария
Моллюски — четко ограниченная группа беспозвоночных, насчитывающая свыше 130 тыс. видов. Обитают в морях, в пресных водах и на суше (табл. 30).
Таблица 30.
Сравнительная характеристика классов моллюсков
| Классы
|
Форма раковины
|
Голова,
придатки
|
Форма ноги
|
Органы
дыхания
|
Сердце
|
Нервная
система
|
Развитие (личинка)
|
Способ питания
|
| Полиплако-форы Polyplacophora |
панцирь из 8 пластинок |
слабо обособлена, придатков нет |
подошво-образная |
6—88 пар ктенидиев |
1 желудочек и 2 предсердия |
лестничного типа |
метаморфоз (тро-хофора) |
фитофаги |
| Аплакофоры Aplacophora |
кутикула со спикулами |
то же |
редуцирована |
нет или 1 пара |
редуцировано |
то же |
то же |
микрофаги, кораллобионты |
| Моноплако-форы Мопоplacophora |
конусовидная |
слабо обособлена, пара щупалец |
подошвообразная |
5 пар тенидиев |
2 желудочка и 2 предсердия |
то же |
то же |
фитофаги |
| Брюхоногие Gastropoda |
конусовидная или турбо-спиральная
|
хорошо обособлена, 2 пары щупалец |
подошво-образная, редко килеобразная или с лопастями |
1 —2 пары ктенидиев
или «легкое»
|
1 желудочек и 1 предсердие
|
разбросанно-узловая, 6 пар ганглиев
|
метаморфоз или прямое (велигер)
|
фитофаги, зоофаги, детрито-фаги
|
| Двустворчатые Bivalvia |
две створки |
головы нет |
клиновидная или редуцирована |
ктенидии или пластинчатые жабры (1 пара) |
1 желудочек и 2 предсердия |
разбросан-но-узловая, 3 пары ганглиев |
метамор-
фоз (велигер или глохидий), реже прямое
|
фильтра-торы |
Головоногие Cephalopoda
|
спиральная с камерами или редуцирована
|
хорошо обособлена, 8—10 щупалец или много |
модифицирована в воронку
|
1—2 пары ктенидиев
|
1 желудочек и 2—4 предсердия
|
головной
мозг (5 пар ганглиев + мантийный
ганглий)
|
прямое
|
зоофаги
|
План строения моллюсков своеобразен. Тело, как правило, состоит из 3 отделов: головы, туловища и ноги, представляющей собой мускулистый вырост брюшной стенки и служащей для движения. Туловище часто разрастается на спинную сторону, образуя внутренностный мешок. За некоторыми исключениями, тело прикрыто известковой раковиной. Кожные покровы содержат значительное количество слизистых и белковых желез. Под покровами много мускулатуры, так что у них, по существу, имеется кожно-мускульный мешок.
Полость тела заполнена рыхлой паренхимой с большим числом лакун. Целом очень мал и не играет роли общей полости тела. У более примитивных форм есть мантия — вырост спинной части кожи, спускающийся по бокам в виде складок. Между туловищем и мантией находится щелевидная мантийная полость. Наружная поверхность мантии выделяет раковину. У более эволюционно продвинутых форм мантийная полость превращается в легкое и исчезает. В мантийной полости помещаются перистые жабры (ктенидии), органы химического чувства, отверстия полового и выделительного аппаратов, анальное отверстие.
В глотке расположена радула, служащая для измельчения пищи, кроме того, у ряда моллюсков развиты ротовые челюсти. От желудка отходит печень, где пища переваривается внутриклеточно. Кровеносная система незамкнута. Сердце включает предсердия и желудочки. В остатки целома, окружающие сердце и половые железы, открываются целомодукты, выводящие экскреты и половые продукты.
Нервная система в пределах типа испытывает прогрессивную эволюцию от двойной нервной лестницы до разбросанно-узло-вого типа. Кроме того, имеется диффузное нервное сплетение в коже.
Среди моллюсков есть раздельнополые и гермафродитные группы. Развитие сходно с кольчецами. У примитивных форм из яйца выходит трохофорная личинка, у большинства остальных — видоизмененная личинка — парусник (велигер). Прямое развитие наблюдается у сухопутных легочных моллюсков.
Среди моллюсков есть как полезные (табл. 31), так и вредные (табл. 32) для человека.
Тип делится на 2 подтипа — боконервных и раковинных.
Таблица 31.
Промысловое значение моллюсков
| Классы
|
Исполь
зование в пищу
|
Исполь
зование
перламутра
|
Жемчуг
|
Другие промыслы
|
Образова
ние осадочных
пород
|
Брюхо-ногие Gastropoda
|
Patella, Buccinum, Haliotis, Strombus, Littorina, Achatina, Helix,
Aplysia
|
Haliotis, Trochus
|
—
|
Сургаеа —
деньги;
Cassis, Strombus — камеи; Buccinum, Murex -
краска
|
мрамор, известняк, ракушечник
|
Окончание табл 31
| Классы
|
Исполь
зование в пищу
|
Исполь
зование
перламутра
|
Жемчуг
|
Другие промыслы
|
Образова
ние осадочных
пород
|
Двустворчатые
Bivalvia
|
Mytilus, Ostrea, Pecten, Anodonta, Unio,
Area,Venus
|
Pteria,
Pinctada, Anodonta,
Unio,
Margari-
tana,
Cristaria
|
Пресноводные
жемчужницы: Margaritifera, Dahurmaia; морские жемчужницы: Pinctada,
Pteria
|
Биссус для веревок, тканей.
Тридакны — посуда
|
То же |
Головоногие
Cephalopoda
|
Loligo,
Octopus, Sepia
|
Nautilus |
— |
Sepia — краска из чернильного мешка |
тоже |
Таблица 32.
Моллюски, приносящие вред человеку
| Классы
|
Вредители сельского хозяйства
|
Промежуточные хозяева гельминтов
|
Вредители построек
|
Обрастание кораблей и других сооружений
|
| Брюхоногие Gastropoda |
улитки: Helix, Achatina; слизни: Arion, Umax |
Lymnaea, Truncatula, Bythinia Leachi, Zebrina идр. |
— |
Vermetidae, Patella, Littorina |
| Двустворчатые Bivalvia |
— |
— |
камнеточцы: Lithophagi-dae, Pholodidae, древоточцы: Teredinidae |
Dreissena, Ostrea,
Mytilus идр.
|
Классификация моллюсков
Подтип Боконервные —
Amphineura
Класс Панцирные — Polyplacophora
Класс Беспанцирные — Aplacophora
Подкласс Бороздчатобрюхие — Solenogastres
Подкласс Ямкохвостые — Caudofoveata
Подтип Раковинные —
Conchifera
Класс Моноплакофоры — Monoplacophora
Класс Брюхоногие — Gastropoda
Подкласс Переднежаберные — Prosobranchia
Отряд Кругожаберные — Cyclobranchia
Отряд Щитожаберные — Scutibranchia
Отряд Гребенчатожаберные — Pectinibranchia
Подкласс Заднежаберные — Opisthobranchia
Подкласс Легочные — Pulmonata
Надотряд Стебельчатоглазые — Styllommatophora
Надотряд Сидячеглазые — Basommatophora
Класс Двустворчатые — Bivalvia
Надотряд Первичножаберные — Protobranchia
Надотряд Жаберные — Autobranchia
Надотряд Перегородчатожаберные — Septibranchia
Класс Лопатоногие — Scaphopoda
Класс Головоногие — Cephalopoda
Подкласс Наутилиды — Nautiloidea
Подкласс Колеоидеи — Coleoidea
Отряд Каракатицы — Sepiida
Отряд Кальмары — Teuthida
Отряд Восьминогие — Octopoda
Отряд Вампироморфы — Vampyromorpha
Раковина у боконервных слагается из отдельных щитков или имеется только у личинок. Челюсти, головные глаза, щупальца и статоцисты отсутствуют. Нервная система состоит из 2 пар нервных стволов, соединенных многочисленными поперечными перемычками (двойная нервная лестница). Вокруг глотки расположено окологлоточное нервное кольцо, соединенное со стволами.
Класс объединяет около 1000 видов. Живут панцирные в морях, главным образом в полюсе прибоя. Ползают по камням или плотно присасываются к ним. Размеры их от нескольких миллиметров до 35 см. Характеризуются своеобразной раковиной, состоящей из весьма подвижно сочлененных пластинок. Внутренно-стного мешка нет. Мантия прикрывает не только ногу, но и голову, ее свободный край покрыт шиповатой кутикулой. Мантийная полость имеет вид круговой борозды, в которой расположены парные ктенидии (от нескольких до десятков пар). Панцирные раздельнополы. Личинка — трохофора.
Раковинные — это моллюски с конической или двустворчатой раковиной. Покровы без шиповатой кутикулы. Голова иногда может отсутствовать, так же как и нога. При наличии головы есть радула и челюсти. Имеются глаза, щупальца и статоцисты. Часто хорошо развит внутренностный мешок. Нервная система разбро-санно-узлового типа, боковые стволы соединяются ниже задней кишки. Подтип делится на 5 классов.
Раковина у моноплакофор цельная, в виде колпачка. Тело состоит из маленькой головы, высокого туловища и дисковидной ноги. На голове пара щупалец, но глаз нет. Мантия такого же типа, как у хитонов. В мантийной борозде находятся 5-6 пар перистых жабр, не образующих сплошного ряда. Мускулатура метамерного строения: имеется 8 пар мышц, идущих от ноги к спинной стороне раковины. Нога с широкой плоской подошвой.
Пищеварительная система включает радулу и двухлопастную печень, открывающуюся в желудок. Кровеносная система образована сердцем с парой желудочков, двумя парами предсердий и аортой. Кровоток осуществляется в системе лакун, расположенных вокруг внутренних органов. Желудочек сердца вместе с прилегающими предсердиями окружен тонкостенным целомическим мешком — перикардием. Спинные целомы сообщаются с внешней средой целомодуктами, которых 6 пар. Нервная система состоит из окологлоточного нервного кольца и 2 пар нервных стволов (педальных и плевровисцеральных), соединенных перемычками.
Моноплакофоры раздельнополы. Обитают в морях на больших глубинах. Это одни из древнейших моллюсков, дожившие до наших дней. Организация их очень примитивна, что выражается в метамерии многих органов — мышц, сердца, почек, жабр и т.д.
Класс брюхоногих, или улиток, включает свыше 90 тыс. видов. Размеры брюхоногих от 2—3 мм до 60 см. Раковина коническая или спиральная, у активных хищников и части других форм нередко отсутствует. Тело состоит из хорошо обособленной головы, несущей 1—2 пары щупалец и парные глаза, ноги (разной формы) и туловища, или внутренностного мешка. Туловище спирально закручено и асимметрично, как и раковина.
Большинство брюхоногих дышат жабрами. Но в некоторых случаях ктенидии исчезают и замещаются другими органами дыхания. В подклассе легочных жабры заменились легкими. Мантийная полость изменилась и стала легочной полостью (рис. 52). При этом свободный край мантии сросся со стенкой тела и мантийная (легочная) полость открывается наружу маленьким отверстием. К легочным относятся сухопутные и пресноводные улитки.
У некоторых переднежаберных сердце симметричное, включает желудочек и два предсердия. Однако у большинства брюхоногих правое предсердие нередко исчезает и сердце оказывается несимметричным.
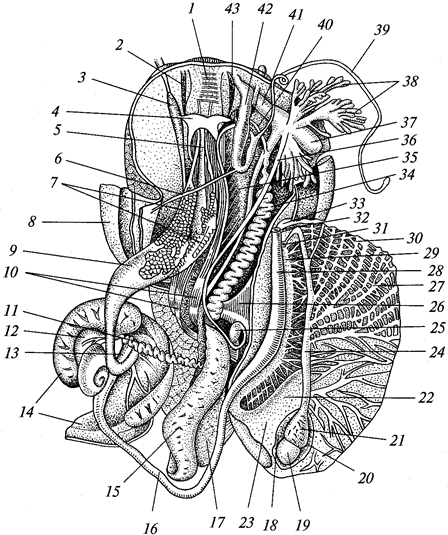
Рис. 52. Вскрытая виноградная улитка (Helixpomatia):
1 —
глотка; 2 —
впяченное внутрь переднее щупальце; 3 —
впяченное внутрь заднее щупальце; 4 —
церебральные ганглии; 5 —
пищевод; 6 —
ретрактор пениса; 7 — слюнные железы; 8 —
мантия; 9 —
зоб; 10 —
ретракторы головы, глотки и щупалец; 11 —
гермафродитный проток; 12 —
гермафродитная железа; 13 —
желудок; 14—
печень; 15 —
белковая железа; 16 —
тонкая кишка; 17—
задний конец ноги; 18 —
отверстие почки в перикардий; 19 —
желудочек сердца; 20 —
перикардий; 21 —
предсердие; 22, 23 —
почка; 24 —
легочная вена; 25 —
семяприемник; 26—
мускул раковины; 27 —
выносящий сосуд легкого; 28—
прямая кишка; 29—
мочеточник; 30 —
приносящий сосуд легкого; 31 —
наружное почечное отверстие; 32 —
анальное отверстие; 33 —
край дыхательного отверстия; 34 —
семеяйцевод; 35
— мускулатура ноги; 36 —
канал семяприемника; 37 —
мешок любовной стрелы; 38 —
пальцевидные железы; 39 —
жгут; 40 —
семяпровод; 41 —
влагалище; 42 —
мешок пениса; 43 —
половая клоака
Нервная система хорошо развита. У примитивных форм имеется 5 пар ганглиев, иннервирующих тело и органы чувств и как бы нанизанных на пару нервных стволов, которые иногда перекрещиваются (хиастоневрия). При переходе к высшим формам происходит переход к разбросанно-узловой нервной системе из нескольких ганглиев, соединенных волокнистыми перемычками. Органы чувств состоят из щупалец, осфрадиев — органов химического чувства, статоцистов и парных глаз.
Улитки могут быть раздельнополыми и гермафродитами. В любом случае половая железа одна. У гермафродитов оплодотворение перекрестное. Оплодотворение у большинства брюхоногих внутреннее. Развитие яйца очень напоминает таковое у многощетинковых кольчецов. Если есть метаморфоз, то из яйца выходит личинка — трохофора либо более сложная — парусник, или велигер.
Улитки обитают во всех средах. Питаются растительной или живой пищей, изредка паразитируют на морских звездах и ежах. Ряд видов улиток (слизни, виноградная улитка — Helixpoma-tia) — вредители полевых, огородных и садовых культур. В отдельных странах улиток употребляют в пищу. Раковины используют в качестве украшений. Некоторые виды легочных улиток являются промежуточными хозяевами гельминтов. В классе 3 подкласса (табл.33).
Таблица 33.
Характеристика подклассов брюхоногих
Подклассы
|
Дыхательная система
|
Мантийный
комплекс
|
Нервная
система
|
Сердце, почки
|
Развитие (личинки)
|
Переднежаберные Prosobran-chia
|
1—2 кте-нидия, направленные вперед |
впереди над головой
|
хиастонев-рия + асимметрия
|
1—2 предсердия, 1 желудочек, 1—2
почки
|
метаморфоз (трохофора, велигер), редко прямое |
Заднежаберные Opisthobranchia
|
1 ктенидий, направленный назад
|
смещен назад по правой стороне
|
хиастоневрии нет, редукция некоторых ганглиев справа |
1 предсердие, 1 желудочек, 1 1 почка
|
метаморфоз (велигер)
|
Легочные Pulmonata
|
легкое
|
впереди, смещен направо
|
хиастоневрии нет, концентрация ганглиев в голове |
тоже
|
прямое
|
В классе 3 надотряда (табл. 34). Двустворчатых узнают по раковине из двух пластинок, соединенных между собой на верхней стороне при помощи связки и зубцов (рис. 53). Последних может и не быть. Главная особенность представителей класса — отсутствие головного отдела, а следовательно, и глотки с радулой. Органы дыхания у большинства — пластинчатые жабры (видоизмененная пара ктенидиев). Внутренностного мешка нет.
Тело билатерально-симметричное, сплюснутое с боков, состоящее из туловища и ноги. Нога чаще всего имеет вид киля и приспособлена для рытья песка. С переходом к неподвижному образу жизни она редуцируется (например, у устриц). У многих форм на ноге есть особая железа, выделяющая быстро твердеющий в воде секрет — биссус. Нитями биссуса моллюск прикрепляется к субстрату.
Мантия выглядит наподобие двух складок, свешивающихся с боков. Нередко свободные края мантии местами срастаются, оставляя отверстия для выдвигания ноги, ввода и вывода воды из полости. Эпителий мантии выделяет раковину. Наружный ее слой эластичен и сложен из органического вещества. Этот же слой образует лигамент — эластичную поперечную ленту, которая растягивает створки раковины. Чтобы держать их закрытыми, моллюск непрерывно затрачивает мускульные усилия.
Таблица 34.
Характеристика надотрядов двустворчатых
Надотряд
|
Жабры
|
Замок
раковины
|
Нога
|
Ротовые
лопасти
|
Тип
питания
|
| Первичножаберные Protobranc-hia |
1 пара перистых ктенидиев |
таксодонтный |
с подошвой |
крупные с длинными придатками |
детритофаги |
Жаберные Autobranchia
|
1 пара
нитевидных или
пластинчатых жабр
|
таксодонт-
ный, гетеродонт-
ный или
отсутствует
|
Клиновидная без подошвы или отсутствует
(часто с биссусом)
|
Без придатков
|
фильтраторы
|
Перегородчатожабер-
ные Septibran-
chia
|
жабры
редуцированы,
имеются жаберные перегородки
|
отсутствует
|
клиновидная, узкая, без биссуса
|
редуцированы
|
зоофаги
|
По способу питания двустворчатые моллюски являются фильтраторами. По бокам рта есть две пары длинных ротовых лопастей, покрытых ресничками. Реснички подгоняют пищевые частицы ко рту. Поскольку редукция головы привела к утрате глотки с радулой и слюнными железами, рот открывается в пищевод, переходящий в мешковидный желудок. Задняя его стенка образует слепой вырост, эпителий которого выделяет студенистый «хрустальный столбик». Конец столбика растворяется желудочным соком и при этом освобождаются пищеварительные ферменты. Печень парная и хорошо развита, ее приток впадает в желудок снизу, как у хитонов. Средняя кишка очень длинная. Задняя кишка пронизывает желудочек сердца и заканчивается анальным отверстием.
Жабр одна пара, причем у примитивных форм они имеют вид перистых ктенидиев либо превращаются в крупные нитчатые или пластинчатые занавеси, занимающие большую часть мантийной полости. Изредка жабры редуцируются. Сильно увеличенные жабры наряду с дыхательной выполняют еще одну важную функцию.
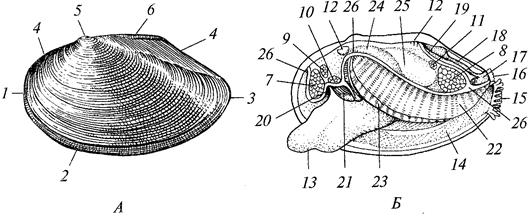
Рис. 53. Беззубка (Anodonta):
А —
внешний вид раковины; Б —
внутреннее строение, раковина и левая мантия удалены: 1 —
передний край раковины; 2 —
брюшной край; 3 —
задний край; 4 —
спинной (замковый) край; 5 — макушка; 6 —
наружный лигамент (связка); 7 — передний мускул-замыкатель; 8 —
задний мускул-замыкатель; 9 —
передний рет-рактор (втягиватель) ноги; 10 —
протракторы (вытягиватель) ноги; 11 —
задний ретрактор ноги; 12 —
элеваторы (подниматели) ноги; 13 —
нога; 14 —
мантийный листок; 15 —
вводный (дыхательный) сифон; 16 —
выводной (анальный) сифон; 17—
полость анального сифона; 18 —
спинной мантийный клапан; 19 —
спинное мантийное отверстие; 20 —
рот; 21 —
ротовая лопасть; 22 —
левая наружная полужабра; 23 —
левая внутренняя полужабра; 24 —
Кеберов орган; 25 —
околосердечная сумка; 26 —
линия, по которой отрезан левый мантийный листок
Они покрыты ресничным эпителием и являются вододвижущим приспособлением. Сердце расположено в перикардии и содержит два предсердия и желудочек. Кроме сосудов есть система лакун в соединительной ткани.
Органами выделения служит пара угловидно согнутых трубчатых почек, одним концом открывающихся в перикардий, а другим — в мантийную полость. Пластинчатожаберные в основном раздельнополы. Гонады парные. Их выводные протоки — яйцеводы или семяпроводы — открываются наружу по бокам основания ноги. Иногда гонады сообщаются с перикардием, и тогда роль выводных протоков берет на себя почка. Оплодотворение обычно наружное. Эмбриональное развитие часто приводит к образованию трохофоры, которая затем превращается в следующую личиночную стадию — парусник. У пресноводных представителей (беззубки и др.) из яиц появляются двустворчатые личинки — глохидии, которые временно становятся кожными паразитами рыб.
Нервная система состоит из 4 пар ганглиев, соединенных более или менее длинными коннективами. У большинства видов ганглии, иннервирующие переднюю часть тела (церебральные) и мантию (плевральные), сливаются вместе. Органы чувств развиты слабо, что связано с малоподвижным роющим образом жизни.
Есть 2 статоциста и органы химического чувства (осфрадии). Головных глаз нет, но нередко по краю мантии образуются сотни глазков.
Двустворчатые имеют большое практическое значение. Прежде всего необходимо отметить их положительную роль в водных экосистемах. Моллюски пропускают через себя огромное количество воды, отфильтровывая из нее взвешенные частицы детрита, бактерии и планктон. К примеру, сидячие моллюски — мидии (живущие и в наших морях), заселяющие 1 м2
дна, за сутки могут профильтровать до 280 м3
воды. Поэтому двустворчатые, подобно губкам, являются природными очистителями воды.
Ряд видов используется как источник перламутра и жемчуга. Перламутр — самый внутренний слой раковины, а жемчуг образуется вследствие реакции моллюска на попавшие между раковиной и мантийной складкой посторонние частицы. Перламутровые слои облекают такую частицу, формируя шарик — жемчужину.
Некоторые морские виды употребляются в пищу. Среди них общеизвестные устрицы и мидии. Моллюсками питаются многие рыбы. Отдельные виды приносят вред. Поселяясь на подводных частях судов, они иногда сильно утяжеляют их и мешают ходу, поэтому приходится периодически ставить суда в доки и счищать возникшие обрастания. Моллюски дрейсены (Dreissena) нередко поселяются в трубах водопроводов и закупоривают их. Несколько видов относятся к древоточцам. Наиболее вреден из них корабельный червь, или шашень (Teredo). Он имеет вид червя, достигающего в длину 10 см. Раковина шашеня редуцирована до степени 2 маленьких пластиночек, при помощи которых животное протачивает длинные ходы в деревянных днищах сосудов, подводных предметах, что ведет к их разрушению. У нас древоточцы встречаются в Черном и дальневосточных морях.
Класс включает около 700 видов крупных морских моллюсков, относящихся к 2 подклассам: наутилиды и колеоидеи. Среди них есть настоящие гиганты. Так, глубоководные кальмары достигают в длину (с вытянутыми щупальцами) 18м. Ведут подвижный образ жизни. Тело их билатерально-симметричное и четко подразделено на голову и туловище. Нога превращена в особые щупальца — «руки», вторично смещенные на голову и окружающие рот. Другой видоизмененной частью ноги является так называемая воронка, лежащая у входа в мантийную полость (рис. 54).
Рис. 54. Кальмар чудесная лампа (Lycoteuthisdiadema)
В мантийном комплексе 2 или 4 ктенидия. У четырехжаберных наутилид имеется наружная раковина, внутренность которой перегорожена поперечными перегородками. Только передняя камера раковины жилая, остальные заполнены газом. У двужаберных ко-лиоидей раковина рудиментарная, внутренняя. По этим признакам головоногих моллюсков и разделяют на указанные 2 подкласса.
Щупалец может быть много (у четырехжаберных корабликов) либо всего 8 или 10. Руки головоногих — широкие в основании и заостряющиеся к концам мускулистые выросты с продольным желобом на внутренней стороне. На этой же стороне находятся дисководные присоски, при помощи которых моллюск схватывает добычу и присасывается к субстрату. По бокам головы помещается пара очень больших глаз.
Туловище одето мантией. На брюшной стороне она отделена от туловища мантийной полостью. С внешней средой полость сообщается щелью. Для замыкания щели есть приспособление наподобие ямок и выступов — «запонок». Между ямками на брюшной стороне туловища имеется воронка в виде конической трубки, приросшей к туловищу. Расширенный задний конец воронки открыт в мантийную полость, а суженный передний торчит наружу. Воронка необходима для выбрасывания воды из мантийной полости и тем самым для движения. При раскрытой щели вода заполняет мантийную полость. Затем щель закрывается, мантия прижимается к туловищу, и вода из мантийной полости с силой выталкивается наружу через воронку, придавая животному обратный толчок. После этого щель раскрывается, мантийная полость заполняется водой и все повторяется сначала. Ритмические сокращения мантии и выталкивание воды служат не только для движения, но и для выполнения дыхательного акта — смены воды вокруг ктенидиев.
Кожа головоногих состоит из однослойного эпителия, подостланного соединительной тканью. В коже расположены многочисленные пигментные клетки или хроматофоры, контролируемые нервной системой. В связи с этим моллюски могут быстро и резко менять свою окраску.
У высших головоногих есть внутренний хрящевой скелет, развитый в «запонках» у основания рук, но главным образом вокруг головного скопления нервных ганглиев. Эта головная хрящевая капсула аналогично черепу позвоночных защищает мозг.
В устройстве пищеварительной системы обращает на себя внимание следующее. Глотка снабжена роговыми челюстями в виде клюва. Секрет слюнных желез ядовит. Желудок мешковидный, мускулистый. В него впадают протоки двулопастной или простой печени. В заднюю кишку перед анальным отверстием впадает проток так называемой чернильной железы, или чернильного мешка. Секрет железы, выброшенный через анальное отверстие в воду, образует стойкое темное облачко, скрывающее моллюска от врага.
Выделительная система представлена 2 или 4 почками, открывающимися в перикардий. Кровеносная система содержит сердце, в котором 1 желудочек и 2 или 4 предсердия. Количество предсердий соответствует числу жабр. Нервная система у двужаберных головоногих очень сложная. Все ганглии сливаются в общую окологлоточную массу, защищенную хрящевой капсулой. Это уже настоящий мозг. Интересно, что имеющийся у других моллюсков педальный ганглий, иннервирующий ногу, разделен у головоногих на 2 — ганглий щупалец, или рук, и ганглий воронки. Это еще одно доказательство того, что руки и воронка представляют собой видоизмененную ногу.
Органы чувств примитивны у четырехжаберных и сложны у двужаберных моллюсков. У первых это статоцисты, осфрадии — органы химического чувства — и глазные ямки. У двужаберных вместо осфрадиев есть обонятельные ямки, а пара крупных глаз не уступает по сложности строения глазам млекопитающих. Они обладают аккомодацией и, кроме того, могут приспосабливаться к освещению разной силы. У глубоководных представителей в коже имеются особые органы свечения.
Головоногие раздельнополы. Половая железа непарная. Оплодотворение сперматофорное, преимущественно в мантийной полости самки. Развитие прямое, происходит в яйце. Из яиц выходят уже сформированные головоногие моллюски.
Практическое значение головоногих велико. Многие из них, например каракатицы, кальмары, осьминоги, употребляются в пищу и являются промысловыми животными. Их добывают в Средиземном, Японском и других морях.
Головоногие моллюски — древняя группа беспозвоночных, большая часть представителей которой вымерли. В ископаемом состоянии обнаружено свыше 80 тыс. видов, что более чем в 10 раз превышает количество современных. Сохранение до наших дней Древних и в то же время высокоспециализированных головоногих можно объяснить их образом жизни и хорошими способами защиты от врагов.
Таблица 35.
Характеристика современных и двух вымерших подклассов головоногих
Под
классы
|
Раковина
|
Щупальца
|
Ктенидии
|
Сердце,
почки
|
Органы движения
|
Жизненная
форма
|
Наутилиды
ДЫ
Nautiloidea
|
наружная, спиральная, многокамерная |
много, гладкие
|
4
|
4 предсердия, 4 почки
|
воронка,
гидростатический
аппарат
|
бентопелагиче-ские
|
| Аммониты Ammoпеа |
наружная, перегородки камер волнистые |
5 пар, гладкие
|
2
|
2 предсердия, 2 почки
|
тоже
|
тоже
|
Окончание табл.35
| Подклассы
|
Раковина
|
Щупальца
|
Ктенидии
|
Сердце,
почки
|
Органы движения
|
Жизненная
форма
|
Белемншы
Belemnoidea
|
коническая,
внутренняя с
камерами
|
5 пар с
присосками
|
2
|
тоже
|
воронка,
плавники,
гидростатический
аппарат
|
нектонные
|
Колеоидеи Coleoidea
|
без раковины или с рудиментами
|
4—5 пар
с присосками
|
2
|
тоже
|
воронка,
плавники,
умбрелла,
мускулатура
|
бентосные,
бентопелагические, нектонные,
планктонные
|
Погонофоры — глубоководные морские животные, ведущие сидячий образ жизни. Живут в длинной и тонкой хитиновой трубке, выделяемой кожными железами. Из верхнего конца трубки выставляется передний участок тела со щупальцами. Тело состоит из 4 первичных сегментов, в каждом из которых есть свой целом. Третий сегмент длинный, в нем участки целома и многие другие органы расположены метамерно. Четвертый сегмент со вторичной сегментацией. Кишечник отсутствует. Пища собирается и всасывается через щупальца. Нервный тяж лежит на брюшной стороне тела. Кровеносная система хорошо развита. Сердце на спинной стороне в перикардии. Органы выделения — целомодукты. Погонофоры раздельнополы, но полового диморфизма нет. Целом в эмбриогенезе образуется энтероцельно. В типе 2 класса: френуляты (Frenulata) и вестиментиферы (Vestimentifera, или Afrenulata).
У погонофор длинное червеобразное цилиндрическое тело, обычно почти нитевидное, от нескольких сантиметров до 1,5 м длиной. На спинной стороне в области переднего целома находится от одного до нескольких сотен щупалец.
Брюшной нервный ствол имеет вид эпителиальной полости, расширяющейся впереди в мозг. Передний целом образует перикардий, выделительные целомодукты и заходит в щупальца. Второй целом в своей полости органов не образует. За счет третьего целома формируются гонады и половые воронки, в задней части — полоски нескольких метамерных щетинконосных сегментов. Кровеносная система замкнутая, состоит из спинного и брюшного сосудов. Брюшной сосуд расширяется у основания щупалец, образуя сердце.
Яйца откладываются самкой в переднюю часть собственной трубки.
Погонофоры впервые были открыты только в XX в. История их открытия и изучения очень интересна.
Погонофоры были открыты в 1914 г. во время экспедиции на голландском океанографическом судне «Сибога» в водах Индонезийского архипелага. Это были нитевидные червеобразные животные, живущие в хитиновых трубках правильной цилиндрической формы диаметром примерно 1 мм. Описавший червя французский зоолог Коллери, пораженный характерным видом его трубки, окрестил червя Siboglinum (Siboga — название судна, и linum — по-латыни нить) и не колеблясь отнес его к новому типу беспозвоночных. Коллери отметил полное отсутствие у червя пищеварительной системы и, пытаясь объяснить это, предположил, что Siboglinum — колониальное животное, у которого функции питания и размножения могут выполняться разными индивидуумами, как это часто имеет место у гидрозоев. Отсутствие в собранных материалах других индивидуумов, ответственных за питание колонии, было, однако, трудно понять.
Лет через 30 после их открытия швед Йохансон объединяет Siboglinumс другой, весьма отличной формой из Охотского моря, Lamellisabella, в новый класс морских беспозвоночных — погонофоры (от греческого «кто носит бороду», другими словами — бородатые), быстро возведенный зоологами в ранг подтипа. Открытия множились. Было замечено, что представители «бородатых», считавшиеся обитателями исключительно больших глубин, могут жить и на глубине нескольких десятков метров и встречаются гораздо чаще, чем предполагалось ранее. Но все эти погонофоры невелики по размерам и все лишены пищеварительной системы.
Первым попытался разрешить эту загадку советский ученый А. В. Иванов. Согласно его гипотезе эпидермальные клетки передних щупалец погонофор способны вылавливать мелкие органические частицы путем фагоцитоза. Но поскольку наблюдать фагоцитоз не удалось, Иванов позднее выдвинул другую гипотезу: собранные султаном щупалец частицы перевариваются вне тела животного ферментами, выделяемыми в наружную среду, а мелкие молекулы, образующиеся в ходе ферментативного разложения частиц, абсорбируются затем проницаемой клеточной мембраной. Второй гипотезе повезло не больше, чем первой, и она так и не была доказана.
В 1966 г. на глубине 1125 м у побережья Калифорнии были открыты погонофоры, которые зоолог Вебб назвал Lamellibrachiabarhami. Они достигают 0,5 м в длину при диаметре 6 мм и отличаются от всех других погонофор наличием особого вестиментального участка тела (вести-ментума), на котором на спинной стороне открываются парные половые отверстия. Этот участок расположен между щупальцевой короной и туловищем и снабжен 2 складками, или крыльями, благодаря которым между вестиментумом и стенкой трубки червя образуется дорсальная камера. Чтобы отметить эту особенность, а также другие не менее важные черты строения, в типе погонофор был создан новый отряд — вестиментиферы, входящий в новый класс — афренуляты. Характерная для типа «бородатых» сегментированная и снабженная хитиновыми щетинками задняя часть тела позволяла сблизить их с кольчатыми червями. Но по-прежнему было неизвестно, как и чем питаются эти странные животные.
В 1977 г. на гидротермали вблизи горячих подводных серных источников на Галапагосском рифте был открыт новый вид Riftiapachyptila(рис. 55).

Рис. 55. Скопление гигантских погонсфор Riftiapachyptila;
крабы, галатеиды и рыбы изобильны на границе поселений рифтий
Длина этого животного может превышать 1,5 м, а ярко-красный киноварный цвет его султана, контрастирующий с перламутрово-белой окраской трубки, придает подводным фотографиям с его изображением неоспоримую эстетичность. По биомассе этот вид во много раз превосходит все другие виды гидротермальных сообществ. Изучением первых собранных образцов занялся американский зоолог Джоунс. Менее чем через 4 года после первых находок было опубликовано описание вида. Это оказалась вестиментифера, но столь сильно отличающаяся от Lamellibrachia, что для нее создали особое семейство — Riftiidae. Одновременно Джоунс установил, что туловище; занимающее около 3/4 длины тела, заполнено тканью, образованной дольками, обильно снабженными кровеносными сосудами, — трофосомой. Эта ткань буквально насыщена округлыми бактериями диаметром около 3—5 мкм. Трофосома различима даже у самых молодых изученных особей (длиной 1,44 мм) и во множестве содержит кристаллы серы (рис. 56). Таким образом, эти животные находятся в симбиозе с сероокисляющими бактериями, осуществляющими хемосинтез, от которых они получают питательные вещества.

Рис. 56. Гигантская погонофора-ве-стиментифера Riftiapachyptila, извлеченная
из трубки, и фрагменты трубки рифтии и кольчатой трубки
вестиментиферы Tevniajerichonana
В 1985 г. на Международном совещании по гидротермальной фауне Джоунс сообщил, что число известных видов вестиментифер утроилось и они объединяются в 5 семейств, не считая некоторых видов, которые пока еще не определены. Шесть новых видов, чьи названия говорят о зонах, где они были собраны, были распределены в 3 новых семейства: Ridgeiidae (от английского ridge — хребет) с родом Ridgeia, содержащим 2 вида, собранных на гидротермальных излияниях подводных хребтов Хуан-де-Фука и Эксплорер, — Ridgeiapiscesae и R. phae-ophiale; Escarpiidae с родом Escarpia (от английского escarp — обрыв, склон, эскарп), включающим 2 вида, собранных в оазисах у подножия Флоридского эскарпа, — Escarpiaspicata и Е. laminata; Tevniiddae с двумя родами — Tevnia (поскольку название Ventia, производное от английского слова vent — выход, источник, уже использовалось в номенклатуре животных, то, чтобы сохранить упоминание о гидротермальной среде, Джоунс прибег к анаграмме) с единственным видом Т. jerichonana и Oasisia, род Tevnia, также включающий единственный вид О. alvinae.
Ученые продолжают обнаруживать и описывать все новых представителей вестиментифер. Создается впечатление, будто эти крупные животные, еще 20 лет назад не известные науке, способны эволюционировать с такой скоростью, что образуют четко различающиеся виды, роды и семейства, оставаясь строго приуроченными к определенным физико-химическим условиям среды, которые изменяются в узких пределах и, кроме того, значительно различаются в разных гидротермальных излияниях.
Джоунс предложил повысить старый подтип погонофор Obturata (иначе говоря, вестиментифер) до ранга типа и числить их под названием Vestimentifera, принимая во внимание следующие их анатомические отличия от собственно погонофор: различия в числе и размере внутренних полостей тела (целомических полостей), в конфигурации вентральной нервной цепочки и наличие особого тканевого листка, мезентерия, в терминальном районе тела. Таким образом, погонофоры и вестименти-феры были отнесены к 2 различным типам морских беспозвоночных, однако многие зоологи с этим не согласны и по-прежнему рассматривают их как один тип.
Гидротермальная фауна предстает все более и более как фауна древняя, содержащая высокий процент живых ископаемых, происхождение которых в зависимости от группы датируется временем от 150 до 250 млн лет назад. В изобилии обнаруживаются новые формы: к концу 1985 г. были описаны 15 новых семейств, более 70 новых видов (один из них — на рис. 57, а на рис. 58 изображены крабы, обитающие в этих сообществах беспозвоночных вместе с погонофорами).
Рис. 57. Помпейский червь Alvinellacaudata с многочисленными
сегментами и измененной задней частью тела


Рис. 58. Два характерных краба гидротермальных истоников: слева – крупный
Cyanagraeapraedator; справа – мелкий Bytograeathermidron, живущий
вблизи колоний погонофор
С каждым годом и с каждым открытием традиционная концепция практически пустынных океанских глубин, в которых жизнь в сильнейшей степени зависит от вертикального потока опускающихся с поверхности частиц, перестает быть господствующей вследствие признания той значительной роли, которую играет в этих местах хемосинтез бактерий-симбионтов.
Было выяснено, что целый ряд обитающих здесь животных питаются не экзогенно (органическим веществом внешнего происхождения), а эндогенно — органическим веществом внутреннего происхождения, получаемым ими от населяющих их тело хемосинтетических бактерий, и таким образом являются симбиотрофами.
(
DEUTEROSTOMIA)
Представители подраздела характеризуются тем, что в эмбриональном развитии бластопор превращается в анальное отверстие или анус образуется на месте замкнувшегося бластопора. Ротовое отверстие появляется вторично на новом месте. Целом происходит из выпячиваний на кишке, т. е. энтероцельно, а нервная система — из эпителиальных полосок, которые в ряде случаев погружаются и образуют нервные трубки. Скелет мезодермального происхождения.
Из беспозвоночных животных ко вторичноротым относятся типы иглокожих и погонофор.
Иглокожие — это морские, радиально-симметричные, донные животные с известковым мезодермальным скелетом из прилегающих друг к другу пластинок. Целом сложно дифференцирован, и за его счет образовался ряд систем, в том числе наиболее характерная для иглокожих амбулакральная. Кровеносная система примитивна и незамкнута. Органы дыхания развиты слабо или отсутствуют, органов выделения нет. Нервная система также примитивна и частично залегает в коже, а частично — в эпителии впятившихся внутрь участков стенки тела. Иглокожие раздельнополы. Развитие происходит с метаморфозом.
Среди современных иглокожих выделяют 5 классов (из них в учебнике рассматриваются 4), группирующихся в 3 подтипа: астерозои, эхинозои и кринозои (табл. 36).
Таблица 36.
Сравнительная характеристика классов иглокожих
Признаки
|
Подтип астерозои
|
Подтип эхинозои
|
Подтип кринозои |
| Классы |
Классы |
Класс |
| Морские звезды |
Змеехвостки |
Голотурии |
Морские ежи |
Морские
лилии
|
| Симметрия |
пятилучевая радиальная |
радиальная 5-лучевая |
билатериальная |
радиальная (у правильных) |
радиальная |
Скелет
|
известковыи цельный (сплошной) |
известковый цельный |
известковый, из отдельных пластинок |
известковый
сплошной, есть педицеллярии
|
известковый
|
Пищеварительная система
|
рот, желудок, задняя кишка, пищевод
|
РОТ, желудок
|
рот, желудок, кишечник, глотка, анус
|
рот, желудок (аристотелев фонарь), задняя
кишка, глотка,
печень
|
рот, пищевод, кишка (печень с выростами), анус
|
Выделительная
Система
|
амебоциты
|
амебоциты
|
амебоциты
|
амебоциты
|
амебоциты
|
| Органы дыхания |
кожные жабры |
—
|
водяные клетки |
5 пар кожных жабр |
—
|
| Амбулакральная система |
Астерозои — свободноподвижные иглокожие, в организации которых четко выражена лучевая (обычно пятилучевая) симметрия. Тело звездообразное или шаровидное, реже червеобразное. Скелет имеет вид панциря. Ротовое и анальное отверстия расположены на противоположных частях тела. Амбулакральная система служит для движения.
Известно около 1700 видов морских звезд. Живут они в морях нормальной солености. В теле различается оральная сторона, обращенная ко дну и несущая в центре ротовое отверстие, и противоположная — аборальная сторона, направленная к открытой воде и несущая анальное отверстие. Плоское тело состоит из центрального диска и отходящих от него широким основанием лучей, число которых часто равно 5, но бывает и большим. Скелет лучше развит на оральной стороне, где он сплошной. Вдоль лучей на этой же стороне проходят борозды. Среди известковых пластинок аборальной стороны выделяется мадрепоровая, пронизанная отверстиями. От поверхности пластинок отходят шипы и иглы.
Внутренние органы расположены в полости, образованной одним из участков целома. Кишечник идет насквозь от рта к анусу. У хищных звезд задняя кишка имеет особые ректальные железы. Желудок большой, мешко видный, от него в целом отходит 5 пар длинных слепых выпячиваний — печеночных мешков, выделяющих пищеварительный сок (рис. 59). Звезды — хищники, питающиеся моллюсками, червями и другими беспозвоночными.
Нервная система примитивна. Она может быть названа тройной, так как состоит из 1 поверхностной — эктоневральной и 2 глубоких — гипоневральной и аборальной систем. В каждую из них входят центральное кольцо и отходящие от него в лучи нервы. Органы чувств крайне просты: короткие щупальца на концах лучей и глазные ямки у основания щупалец.
Самая характерная система звезд и вообще иглокожих — амбулакральная (рис. 60, А).
Она представляет собой систему каналов, наполненных водянистой жидкостью. Амбулакральная система начинается на аборальной стороне мадрепоровой пластинкой, поры которой ведут в каменистый канал, подающий жидкость в околоротовое кольцо. От него берут начало радиальные каналы, проходящие в лучах и дающие боковые веточки. Каждая веточка заканчивается амбулакральной ножкой с ампулой. Ножки — очень растяжимые, полые мускулистые выросты, имеющие на концах присоски. На дне амбулакральной борозды каждого луча ножки расположены в 2 или 4 ряда.
Движение происходит вследствие растягивания ножек под напором вгоняемой из раздельного луча жидкости. Вытянувшиеся ножки присасываются к субстрату. Затем мускулатура их сокращается, жидкость уходит в ампулы, сами ножки укорачиваются, а животное подтягивается вперед. Далее ножки отцепляются от субстрата, в них снова вгоняется жидкость и т. д. Скорость движения, естественно, невелика — 5—10 см в минуту.
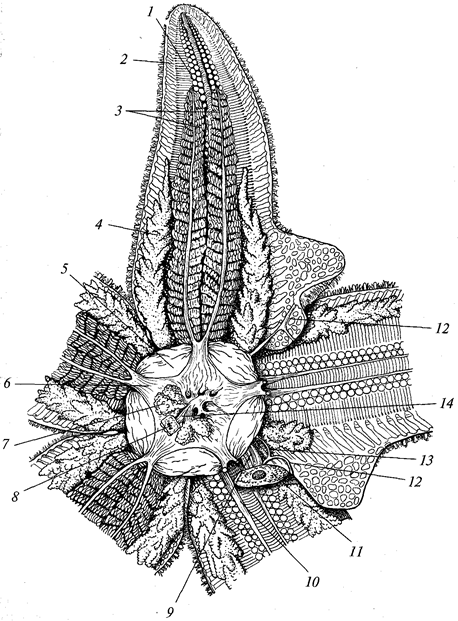
Рис. 59. Вскрытая морская звезда Astenasrubens:
1
~
амбулакральные пластинки; 2 —
маргинальные пластинки; 3 —
печеночные Мешки; 4 -
гонады; 5 - оральный отдел желудка; б -
аборальный отдел желудка; 7 -- ректальные железы; 8 —
кусочек спинной стенки тела с анальным отверстием; 9—
каменистый канал; 10 -
мускулы-ретракторы желудка; 11 -
участок кожи с мадрепоровой пластинкой; 12 -
половой столон (содержит развивающиеся половые клетки); 13 —
половой проток; 14 —
задняя кишка
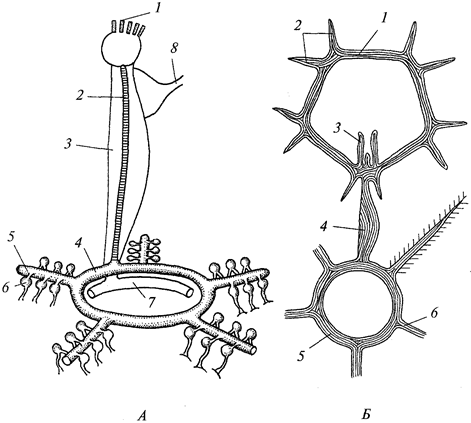
Рис. 60. Схемы амбулакральной системы (А)
и кровеносной
системы (Б)
морской звезды:
А: 1 —
мадрепоровая пластинка; 2 —
каменистый канал; 3 —
осевой синус (участок целома); 4 —
оральное кольцо амбулакральной системы; 5 — радиальный амбулакральный канал; 6 —
ампулы амбулакральных ножек; 7 — оральное перигемальное кольцо; 8 —
половой столон; Б'. 1 —
аборальное кольцо; 2 —
ветви к половым железам; 3 —
ветви к кишечнику; 4 —
осевой орган; 5 — оральное кольцо; 6 —
радиальные каналы
По такой же схеме построены и другие системы — перигемальная и кровеносная (рис. 60, Б).
Перигемальная система — это участок целома, состоящий из околоротового кольца и радиальных каналов. Она сопровождает нервную систему и служит для ее амортизации и питания: нервная система омывается перигемальной жидкостью. Каждый из радиальных каналов поделен вертикальной продольной перегородкой, внутри которой помещаются лакуны кровеносной системы. Они объединяются околоротовым кольцом. Есть и аборальное кровеносное кольцо. Оба кольца связаны осевым органом, который прилегает к каменистому каналу. Осевой орган состоит из губчатой ткани и продуцирует кровяные клетки. Вместе с каменистым каналом и участками целома он входит в осевой комплекс органов.
Половая система проста. В основании лучей залегают гонады, а между лучами расположены выводящие трубочки. Оплодотворение наружное. Дробление яйца полное, в результате получается покрытая жгутиками бластула. Гаструляция осуществляется путем впячивания. Мезодерма происходит от стенки первичной кишки. Бластопор превращается в анус или последний образуется после зарастания бластопора на его месте. Первичная личинка — диплеврула — билатерально-симметричная. В дальнейшем метаморфозе имеются и другие личиночные стадии.
У морских звезд хорошо развита способность к регенерации.
Класс морских ежей объединяет около 800 современных представителей, имеющих шаровидную (реже дисковидную) форму и покрытых многочисленными известковыми иглами, среди которых есть педицеллярии — иглы, видоизмененные в хватательные щипчики. Как и звезды, морские ежи ползают на оральной стороне, где размещен рот. Скелет развит сильнее, чем у других игло-кожих. По существу, это сплошная скорлупа, лишь вокруг рта и анального отверстия есть мягкие участки. Пластинки панциря расположены по меридианам тела. Таких меридиальных полос 10, и каждая слагается из 2 рядов пластинок — амбулакральных и интерамбулакральных (рис. 61). Амбулакральные пластинки с ножками. Скелет принимает участие в образовании жевательного аппарата — аристотелева фонаря, который состоит из многочисленных перекладин и пластинок, соединенных мышцами. 5 из них окружают рот в виде зубов.
Рис. 61. Правильный морской еж Echinocidaris с аборального
полюса, наполовину очищенный от игл:
1 —
интерамбулакральные; 2 —
амбулакральные ряды пластинок
Большинство систем органов ежей и звезд принципиально сходны. Нервная система погружена под панцирь. Среди органов чувств выделяются сферидии — булавовидные вздутые иглы, играющие роль статоцистов. Органы дыхания наподобие кожных жабр есть у многих ежей. Движение осуществляется при помощи амбулакральных ножек. Некоторые ежи могут передвигаться как на ходулях с помощью подвижных игл скелета, которые иногда достигают большой длины.
Класс включает около 600 видов. Характеризуются голотурии вытянутым червеобразным телом и сильно редуцированным скелетом. Размеры от нескольких миллиметров до 2 м. Двигаются на одной стороне тела, которую можно назвать брюшной. На переднем (оральном) конце расположен рот, окруженный венчиком щупалец; на заднем (аборальном) — анальное отверстие (рис. 62).
Рис. 62. Голотурия Cucumaria. Вид с правой стороны:
1 — щупальца; 2 — ампулы щупалец; 3
— верхние амбу-лакральные ножки; 4 —
клоакальное отверстие; 5—
нижние амбулакральные ножки
Амбулакральные ножки (3 ряда) нижней стороны хорошо развиты, с их помощью животные двигаются. Верхние 2 ряда ножек маленькие и лишены присосок. Щупальца также являются видоизмененными ножками, поскольку в них заходят выросты кольцевого амбулакрального канала. Скелет развит слабо и представлен разбросанными в соединительной ткани микроскопическими известковыми тельцами. Мускулатура стенок тела сильно развита. К ней относятся кольцевые мышцы, продольные мышечные ленты и мышцы-ретракторы, втягивающие внутрь передний конец тела.
Пищеварительная система — обычная для иглокожих, но задняя кишка образует объемистую мускулистую клоаку, в которую впадают многочисленные железистые трубочки, замкнутые на противоположном конце. Эти трубочки (кювьеровы органы), выполняющие функцию защиты, при раздражении животного выстреливаются через отверстие клоаки, вытягиваются и облепляют нападающего. С клоакой связаны и так называемые водяные легкие. Это обширные, сильно разветвленные тонкостенные мешки, впадающие в клоаку. Голотурии периодически втягивают воду через клоаку и с силой выбрасывают ее обратно. Вода из клоаки проходит в легкие, и кислород диффундирует через их стенки в полость тела. Кроме того, водяные легкие выполняют и выделительную функцию. Через их стенки из тела выходят амебоциты, нагруженные экскретами.
Половая система состоит из единственной гонады трубчатого типа. Все трубки сливаются в один проток, открывающийся близ переднего конца тела. Большинство голотурий раздельнополы, некоторые гермафродитны.
Для голотурий характерно явление автотомии, или самокалечения. При сильном раздражении в стенке клоаки образуется разрыв, через который выбрасываются кишечник, водяные легкие и гонады. Через короткий срок утраченные части регенерируют.
Среди голотурий есть промысловые виды. Это лишенные скелетных элементов трепанги, добываемые в прибрежных водах Тихого океана. Они употребляются в пищу. Промышляются и другие виды, например морские огурцы из Баренцева моря.
Кринозои — иглокожие, прикрепленные аборальной стороной или стебельком к субстрату. Рот и анальное отверстие расположены на оральной стороне. Амбулакральная система выполняет дыхательную функцию.
Подтип включает один современный и несколько вымерших классов.
Морские лилии — наиболее древний, вымирающий класс иглокожих. Стебельчатые лилии — сидячие донные животные. Они прикрепляются ко дну аборальной частью тела, из центра которой отходит стебелек. Их тело имеет вид чашечки, от которой отходит пять лучей — рук. В основании руки раздваиваются, иногда неоднократно. На разветвлениях рук расположены дополнительные веточки — пиннулы. Вдоль верхней оральной стороны рук проходит амбулакральная борозда со щупальцами. У бесстебельчатых лилий, способных к свободному передвижению, на аборальной стороне есть центральная пластина с усиками (циррами), которыми лилия цепляется за субстрат (рис. 63).

Рис. 63. Строение бесстебельчатой морской лилии Heliometraglacialis:
А —
внешний вид; Б —
детали чашечки и части лучей; В —
ротовой диск молодой лилии (увеличено); 1 —
центральный конус чашечки; 2 —
цирры; 3 —
места прикрепления цирр; 4 —
лучи; 5 —
первый членик (позвонок) лучей; 6 —
второй членик лучей; 7 — радиальная пластинка; 8 —
пиннулы; 9 —
ротовое отверстие; 10 —
амбулакральные бороздки; 11 —
анальное возвышение; 12 —
анус; 13 —
поры; 14 —
рудиментарные пластинки ротового скелета
Кишечник у морских лилий подковообразный, его начало и конец открываются на оральной стороне. Амбулакральная система обычная, но с околоротовым каналом связано 5 или более каменистых каналов, а поверхность чашечки пронизана сотнями пор. Из трех нервных систем сильнее развита аборальная.
Дыхательной и выделительной систем нет. Лилии раздельнополы. Гонады сложно устроены и продолжаются от аборального участка чашечки до пиннул. Половые продукты выходят через разрывы в стенках пиннул. Из яйца появляется личинка, которая после периода плавания прикрепляется ко дну передним концом. Свободный (бывший задний) конец образует чашечку. Постепенно сидячая личинка в ходе метаморфоза превращается во взрослый организм.
Полухордовые — двусторонне-симметричные целомические беспозвоночные, живущие в морях. Их тело состоит из трех отделов: хоботка, воротничка и туловища (рис. 64).
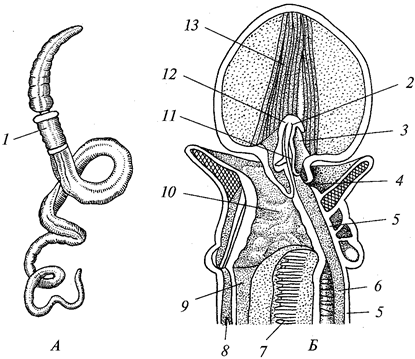
Рис. 64. Строение кишечнодышащих Enteropneusta:
А —
Saccoglossuskowalevskyi, видно разделение тела на хоботок, воротничок и туловище; Б —
продольный разрез переднего конца Ptychoderaminuta; / — жаберные щели; 2 —
сплетение кровеносных сосудов 3 —
перикардий; 4 —
хоботная пора; 5 — спинной нервный тяж; 6 —
спинной сосуд; 7 — жаберная кишка; 8 —
брюшной сосуд; 9 —
пищевод; 10 —
глотка; 11 —
сердечная лакуна; 12 —
нотохорд; 13 —
продольные мускулы хоботка
В каждом отделе есть свои целомические мешки — в хоботке непарные, а в других отделах — парные. Хоботок поддерживается небольшим слепым выростом глотки — нотохордом. В глотке имеются парные жаберные щели. Нервная система состоит из брюшного и спинного нервных стволов, связанных одним-двумя окологлоточными нервными кольцами. Незамкнутая кровеносная система включает два основных продольных сосуда — спинной и брюшной. Над нотохордом спинной сосуд, расширяясь, образует центральную лакуну, к которой примыкает мускулистый пульсирующий орган — перикардий. Органы выделения — целомодукты. Полухордовые раздельнополы, половые железы находятся в туловище. В типе 2 класса: кишечнодышащие и крыложаберные.
Кишечнодышащие — длинные (от нескольких сантиметров до 2,5 м) червеобразные животные, ведущие роющий образ жизни на дне моря. Размножаются половым и бесполым путем с помощью фрагментации. У большинства имеется плавающая личинка, у остальных развитие прямое.
Крыложаберные — сидячие, главным образом, колониальные животные, обитающие в разветвленных домиках или трубках. В отличие от кишечнодышащих обладают укороченным телом. Размножаются как половым, так и бесполым способом путем почкования. Бесполое размножение и приводит к образованию колоний.
Хордовые — высший тип вторичноротых животных. Включает около 43 тыс. современных видов, распространенных всесветно и во всех средах. Общие черты организации следующие. В течение хотя бы одной стадии онтогенеза имеется хорда, или спинная струна, играющая роль внутреннего осевого скелета. Центральная нервная система представляет собой трубку эктодермального происхождения, лежащую над хордой, т. е. дорзально. Передний отдел кишечной трубки — глотка — пронизан жаберными отверстиями и выполняет одновременно функции участка пищеварительного тракта и органа дыхания (последнее — у водных позвоночных, у наземных позвоночных жаберные щели только у зародышей, а потом зарастают). Сердце находится на брюшной стороне тела, под пищеварительной трубкой и хордой. Хордовые — билатеральные, целомические животные, различные по внешнему виду и деталям строения. В типе 3 подтипа.
Бесчерепные произошли от полухордовых из класса кишечнодышащих, способность которых к передвижению постепенно возрастала, что привело к усложнению нервной, двигательной и других систем. Современные бесчерепные включают небольшой класс (всего около 30 видов) головохордовых. Они широко распространены в умеренных и теплых водах Атлантического, Тихого и Индийского океанов и в связанных с ними морях, в частности в Черном море, где обитают на дне в прибрежной полосе. Известно несколько видов планктонных ланцетников, которые, по-видимому, являются личинками донных видов. Насчитывается 35 современных видов из единственного класса.
К этому классу относятся небольшие полупрозрачные животные, известные под названием ланцетники, которые достигают нескольких сантиметров в длину. Тело вытянутое (рис. 65), сдавленное с боков, спереди и сзади заостренное. Передняя часть тела шире задней, голова не выражена. Вдоль спины, вокруг хвостового отдела и вдоль задней части брюшной стороны тянется йлавниковая складка. Далее на брюшной стороне идут кпереди парные плавниковые складки. Задний конец тела, обрамленный плавниковой складкой, похож на лезвие хирургического ножа — ланцета, чем и объясняется название описываемого животного. По форме тела бесчерепные немного напоминают рыб, к которым их долгое время относили. Наружные покровы лишены пигментации и прозрачны. Эпидермис однослойный. Под эпидермисом — тонкий слой соединительной ткани.
Рис. 65. Продольный разрез ланцетника:
/ — предротовое отверстие, окруженное щупальцами; 2 —
хвостовой плавник; 3 —
спинной плавник; 4 —
метаплевральная складка; 5 — атриальное отверстие; б—
хорда; 7— миомер (показан только в хвостовой области); 8 —
миосепта; 9 —
Нервная трубка; 10 —
парус; 11 —
жаберные щели; 12 —
кишка; 13 —
печеночный вырост кишечника; 14 —
околожаберная полость; 15 —
эндостиль; 16 —
половые . железы
Центральная нервная система представлена трубкой, начинающейся немного отступя от переднего конца тела и идущей на фпинной стороне над осевым скелетом почти до самого заднего конца тела. Передний конец нервной трубки слегка расширен. В середине трубки проходит узкий продольный канал. От трубки отходит большое количество нервов. Из них первые две пары иннервируют переднюю часть тела и называются головными.
Органы чувств очень примитивные. Настоящих глаз нет, а вдоль Нервной трубки рассеяны многочисленные скопления черного пигмента, которые воспринимают световые раздражения, проникающие внутрь тела благодаря прозрачности наружных покровов. Бесчерепные избегают света. На переднем конце тела имеется ямка, которая, по-видимому, воспринимает химические раздражения. Осязательные чувствующие клетки рассеяны по всей поверхности тела.
Вдоль продольной оси тела, под нервной трубкой, проходит хорошо развитый упругий тяж — хорда, играющая роль осевого скелета. Хорда, окруженная несколькими прочными оболочками, образована вакуолизированными клетками и состоит из поперечных дисков. Она начинается впереди нервной трубки у самого переднего конца тела, благодаря этому указанный конец сильно укреплен, что облегчает зарывание животного в грунт. В связи с отсутствием головного мозга череп у ланцетника не развит, чем и объясняется название подтипа. Опорную роль выполняет также бесклеточная соединительная ткань в виде различных перекладин и перегородок, расположенных во всех частях тела, часть их отходит от хорды.
Мышечная система развита хорошо и в основном состоит из многочисленных сегментов, идущих косо по отношению к продольной оси тела. Мышечные элементы имеются и в других частях тела (на брюшной стороне, в основании плавниковых складок и т.д.). Благодаря хорошо развитому мышечному аппарату животные могут сильно изгибать тело и быстро зарываться в грунт. Они могут и плавать непродолжительное время на боку при помощи волнообразных сокращений тела. Плавание облегчается наличием плавниковых складок.
Передняя часть пищеварительной трубки (глотка), тянущаяся от рта до середины тела, пронизана длинными косыми жаберными щелями. Стенки, окружающие щели, обильно снабжены кровеносными сосудами. Жаберные щели открываются не прямо наружу, а в особую околожаберную (атриальную) полость, которая сообщается с наружной средой при помощи отверстия, находящегося посредине брюшной стороны тела. Околожаберная полость защищает дыхательный аппарат животных от повреждений и засорения при зарывании их в грунт.
Предротовое отверстие, окруженное щупальцами, которые предохраняют от попадания внутрь крупных частиц, ведет в предро-товую воронку, где находится ротовое отверстие, окруженное мускульной перегородкой — парусом. Вода загоняется в ротовую полость и далее в глотку движением многочисленных ресничек. На брюшной стороне глотки имеется эндостиль —
продольная борозда, клетки которой выделяют слизь. Благодаря мерцательному движению ресничек в глотку поступает вода вместе с органическими остатками и мелкими организмами. Слизь, выделяемая клетками эндостиля, склеивает пищевые частицы, и они препровождаются в среднюю кишку, где и перевариваются. От этого отдела кишечника отходит вперед отросток, который вполне гомологичен печени позвоночных. Экскременты удаляются через анальное отверстие, находящееся на брюшной стороне в задней части тела. Пищеварительный аппарат бесчерепных, несмотря на его значительные размеры, слабо дифференцирован на отделы, а поступление пищи обеспечивается весьма примитивным способом.
Кровеносная система замкнутая, состоит из двух главных сосудов — спинного и брюшного и их многочисленных разветвлений (рис. 66).
Рис. 66. Кровеносная система ланцетника:
1 —
приносящие жаберные артерии; 2 —
выносящие жаберные артерии; 3 —
сонные артерии; 4 —
спинная аорта; 5 — брюшная аорта; 6 —
кювьеровы протоки; 7 — корни спинной аорты; 8 —
хвостовая вена; 9 —
передние кардинальные вены; 10 —
задние кардинальные вены; 11 —
подкишечная вена; 12 —
воротная вена печени
Кровь со всего тела, богатая углекислым газом, собирается в брюшной сосуд и течет к переднему концу тела. От брюшного сосуда отходит множество сосудов к жаберному аппарату, в котором происходит обмен газов. Обогащенная кислородом кровь собирается в два сосуда, проходящих на спинной стороне тела и образующих один спинной кровеносный сосуд. Кровь в последнем течет к заднему концу тела. Следует отметить примитивные черты кровеносной системы: у бесчерепных нет сердца и кровь приводится в движение сокращением стенок ряда сосудов; кровь бесцветна, так как в ней нет гемоглобина — наиболее совершенного дыхательного пигмента.
Органы выделения, которых около 100 пар, размещенные вдоль жаберного аппарата, несколько напоминают по своему строению метанефридии. Каждый орган открывается несколькими отверстиями в полость тела и одним общим — в околожаберную полость. По
краям входных отверстий находится много специальных клеток — соленоцитов, тоже поглощающих продукты диссимиляции, что говорит о примитивности этой системы.
Бесчерепные — раздельнополые животные. Органы размножения устроены очень просто. Они представлены многими парными половыми железами, лежащими по бокам околожаберной полости. Созревшие половые клетки выходят через разрывы стенок желез в околожаберную полость, а оттуда — в воду, где происходит оплодотворение. Развитие зиготы совершается в воде.
Эмбриональное развитие бесчерепных (ланцетника) было впервые изучено А. О. Ковалевским и дало очень много для понимания положения этой группы в системе царства животных. Начальные стадии эмбрионального развития (рис. 67) — дробление яйца, образование бластулы и инвагинационной гаструлы — очень напоминают развитие иглокожих и других беспозвоночных. Рот, как у иглокожих, кишечнодышащих и у всех хордовых, вторичный.

А —
продольные разрезы: 7— бластула; II, III, IV —
гаструляция; V, VI —
образование мезодермы, хорды и нервной системы: 1 —
анемальный и 2 —
вегетативный полюса; 3 —
гастральная полость; 4 —
гастропор; 5 — нервный канал; 6 —
нервно-кишечный канал; 7 — невропор; 8 —
складка мезодермы; 9 —
целомиче-ские мешки; 10 —
хорда; 11 —
место будущего рта; 12 —
место будущего анального отверстия; Б —
поперечные разрезы четырех стадий развития (
I
—
IV
)'. 1 —
эктодерма; 2 —
энтодерма; 3 —
мезодерма; 4 —
полость первичного кишечника; 5 — нервная (медуллярная) пластинка; 6 —
нервная трубка; 7 — ее полость (невроцель); 8 —
хорда; 9 —
целом; 10 —
параллельный и 77 — висцеральный листки мезодермы
Полость тела (целом), так же как у названных животных, возникает путем отшнуровки от первичного кишечника, причем первоначально образуются три пары целомических пузырей. Таким образом, нет никаких сомнений в родственной связи бесчерепных с вторичноротыми, трехчленистыми беспозвоночными животными.
Дальнейшее развитие, а именно формирование нервной трубки, хорды, мышц и ряда других органов, происходит так же, как у позвоночных. На спинной стороне зародыша по средней продольной линии появляется борозда, которая замыкается и превращается в нервную трубку, лежащую под эпидермисом. От срединной части спинной стенки первичного кишечника отшнуро-вывается длинный тяж, превращающийся в хорду. Задние (т. е. третьи) целомические пузыри удлиняются и распадаются на большое количество мешков. Стенки последних представляют собой мезодерму, из которой образуются мышечные сегменты, кровеносные сосуды, органы выделения и др. Целом взрослого животного образуется из полости описываемых мешков.
До классической работы А. О. Ковалевского по развитию ланцетника бесчерепных считали низшими рыбами. Выдающийся русский эмбриолог показал, что этих животных следует рассматривать как промежуточный этап между двумя большими разделами животного мира — беспозвоночными и позвоночными.
(
UROCHORDATA, или TUNICATA)
Оболочники широко распространены в океанах и морях. Их насчитывается около 1100 видов, из которых около 1000 принадлежат к классу асцидий, ведущих прикрепленный образ жизни. Большинство асцидий — одиночные животные, остальные образуют колонии.
Тело покрыто толстой оболочкой — туникой (чем объясняется одно из названий подтипа), образующей мешок, сообщающийся с наружной средой двумя широкими трубками (сифонами). Через одну из них в организм попадает вода, через другую она выходит (рис. 68). Обычная величина тела — несколько сантиметров.
Нервная система развита слабо. Она представлена небольшим ганглием, лежащим над глоткой, и отходящими от него к разным органам нервами. Имеется тонкий кожно-мускульный мешок.
Пищеварительная система начинается ртом, сообщающимся с внешней средой через входной сифон, и состоит из глотки (на ее спинной стороне имеется эндостиль), желудка и подковообразной кишки, открывающейся анальным отверстием в выходной сифон. Глотка пронизана мелкими жаберными отверстиями, которые открываются в околожаберную полость. Получение пищи (мелких организмов и органических кусочков) и переваривание ее происходит, как у ланцетников.
Рис. 68. Асцидия:
/— внешний вид, //— внутреннее строение; 7 — ротовый сифон; 2—
клоакальный сифон; 3 —
туника (оболочка); 4, 5 —
мантия; 6 —
глотка; 7 — полость глотки; 8 —
жаберные отверстия; 9 —
эндостиль; 10, 11 —
околожаберная полость; 12 —
ее стенка; 13
— желудок; 14 —
печеночный вырост; 15 —
анальное отверстие; 16 —
семенник; 17 —
яичник; 18 —
протоки половых желез; 19 —
околосердечная сумка; 20 —
сердце; 21 —
нервный узел
Кровеносная система незамкнутая. Кровь приводится в движение сердцем, от которого к разным органам отходят сосуды, особенно сильно разветвленные в стенках жаберных щелей глотки. Последняя очень велика и выполняет, как и у ланцетников, роль органа дыхания, через который проходит вода, удаляющаяся после газообмена через выходной сифон.
Продукты диссимиляции накапливаются некоторыми клетками и остаются в организме.
Все оболочники гермафродиты; оплодотворение наружное и внутреннее. Многие виды размножаются также бесполым способом (почкованием).
Положение оболочников в системе животных длительное время оставалось невыясненным, пока А. О. Ковалевский обстоятельно не изучил развитие асцидий, показав, что оно очень сходно с развитием ланцетников и заканчивается образованием планктонной личинки, похожей по форме тела на головастиков и передвигающейся при помощи хвоста. Личинки имеют хорошо развитую нервную трубку и хорду. После короткого периода планктонной жизни личинки прикрепляются к твердому субстрату и их организация подвергается коренной перестройке, в основном регрессивной: хвост вместе с нервной трубкой (за исключением ее переднего конца, превращающегося в ганглий) и хордой редуцируются (как излишние при сидячем образе жизни), другие же органы, необходимые взрослым животным, развиваются. Оболочники благодаря прекрасно развитому фильтрационному аппарату стали многочисленной группой, добывающей себе пищу в любых местах океанов и морей. Подтип делится на 3 класса: асцидий, сальпы и аппендикулярии.
Класс включает около 1 тыс. видов. Это одиночные или колониальные сидячие животные.
Тело напоминает двугорлую банку, прикрепленную основанием к субстрату и имеющую два отверстия — ротовой и клоакальный сифоны. Снаружи тело покрыто туникой, содержащей клетчатко-подобное вещество (единственный в мире животных случай образования вещества, близкого к целлюлозе). Туника выделяется эпителием. Под ней лежит кожно-мускульный мешок, или мантия. Сокращение или расслабление мантийной мускулатуры вместе с биением ресничек эпителия внутренних стенок ротового сифона способствует нагнетанию воды в глотку.
Из ротового сифона вода попадает в глотку, которая занимает большую часть тела. Стенки глотки пронизаны множеством отверстий — стигм, открывающихся в атриальную полость. За глоткой идут пищевод, желудок и кишка, открывающаяся анальным отверстием в атриальную полость вблизи клоакального сифона. Питание пассивное (фильтрация).
Глотка служит и органом дыхания. Сердце сокращается то в одном, то в другом направлении одним и тем же сосудом. Как и все оболочники, асцидий гермафродиты, но половые железы развиваются неодновременно, и один и тот же организм функционирует то как самец, то как самка. Оплодотворение наружное, реже в клоакальной полости.
Развитие оплодотворенного яйца приводит к образованию хвостатой личинки.
Класс насчитывает 25 видов, обитающих в теплых морях. Представители внешне напоминают бочонок, в котором ротовой и клоакальный сифоны расположены на противоположных концах тела (рис. 69). Плавают за счет реактивного движения: вода с силой выталкивается из клоакального отверстия, благодаря чему животное движется толчками. Наблюдается чередование бесполого и полового поколений (метагенез). Из оплодотворенных яиц выходят бесполые сальпы, размножающиеся почкованием. Отпочковавшиеся особи формируют гонады и размножаются половым способом. Расселительной личинки, характерной для асцидий, нет.
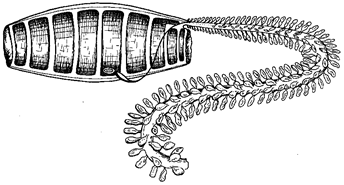
Рис. 69. Бочоночник (Doliolum)
(
APPENDICULARIAE, или LARVACEA)
Класс объединяет около 60 видов мелких оболочников. Внешне и строением напоминают личинок асцидий (рис. 70).
Рис. 70. Аппендикулярия (Oicopleura), животное в домике:
1 —
рот; 2
— анус; 3 — жаберное отверстие-стигма; 4 —
нервный спинной ствол:
5 —
хорда; 6 —
семенник; 7 — яичник; 8 —
домик; 9 —
его решетка; 10 —
ловчая сеть; 11 —
отверстие домика; сплошными стрелками обозначено направление тока воды; пунктирной стрелкой — направление движения домика
Хорда расположена в хвосте. Настоящая туника отсутствует. Живут в «домике» — образовании слизистого характера, который выделяет эктодерма мантии. Передняя часть домика имеет отверстие, закрытое решеткой из слизистых нитей. В домике есть ловчая сеть, к вершине которой обращен рот животного. Все это вместе образует ловчий цедильный аппарат. Цикл развития без чередования поколений и без четко выраженной стадии личинки.
По взглядам многих зоологов, аппендикулярии представляют собой потомков асцидий, утративших взрослую стадию и выработавших способность к половому размножению на стадии личинки (неотения).
(
VERTEBRATA, или CRANIATA)
Позвоночные, или черепные, — высшая группа хордовых, ведущая свое происхождение от бесчерепных. Позвоночные характеризуются активным питанием, что сопровождается усилением подвижности организма. Хорда замещается позвоночником, развивается череп, вооруженный челюстями. Появляются парные конечности и их пояса. Интенсивность всех функций организма повышается.
Общая характеристика
Тело позвоночных разделяется на голову, туловище (с конечностями, служащими для передвижения) и хвост. Кожа состоит из многослойного эпидермиса (эктодермального происхождения) и подстилающего его соединительнотканного слоя (мезодермального происхождения) и служит надежной защитой от механических и других вредных внешних воздействий, более частых при активном образе жизни. Кожа имеет разные придатки (костную или роговую чешую, перья, волосы и др.) и железы различного назначения, характерные для каждого класса подтипа. Размеры тела позвоночных, как правило, крупнее, чем беспозвоночных животных.
Нервная система.
Вся нервная система даже у низшего класса подтипа намного сложнее, чем у бесчерепных. Центральная нервная система состоит из головного мозга, развившегося из передней части нервной трубки бесчерепных, и спинного мозга, возникшего из остальной, более длинной части последней и простирающегося до заднего конца тела. Головной мозг разделяется На пять связанных между собой отделов: передний, промежуточный, средний, мозжечок, продолговатый. В каждом из этих отделов есть центры, управляющие определенными функциями жизнедеятельности животных (например, обоняния, зрения, дыхания, пищеварения и т.д.).
Органы чувств (зрения, слуха, обоняния и др.) весьма совершенны.
Эндокринная система.
К основным железам внутренней секреции относятся: гипофиз (мозговой придаток), надпочечники, вилочковая, щитовидная и околощитовидная железы, половые железы и др.
Скелет.
У позвоночных имеется скелет, который состоит из черепа, осевого скелета (хорды или позвоночного столба) и других частей. Череп развился для защиты головного мозга и наиболее сложных органов чувств (обоняния, зрения, равновесия и слуха), укрепления переднего отдела пищеварительного аппарата и связанного с ним у первичноводных позвоночных (бесчелюстных и рыб) жаберного аппарата. В черепе различают два отдела: мозговой
(связанный с головным мозгом и органами чувств, находящимися в голове) и висцеральный
(связанный с пищеварительным и жаберными аппаратами). В течение эволюции подтипа произошла замена хрящевого скелета костным.
Органы передвижения.
У первичноводных они представлены плавниками, у наземных — ногами и крыльями.
Мышечная система.
У первичноводных позвоночных (бесчелюстных и рыб) она в основном как и у бесчерепных, метамер-ного типа, но значительно более развитая. У наземных позвоночных (земноводных, пресмыкающихся, птиц и млекопитающих) она подвергалась все возрастающей дифференциации на разные группы мышц.
Полость тела
— целомическая.
Пищеварительная система.
По сравнению с таковой бесчерепных очень усложнилась. Это выразилось в образовании ротовой полости, служащей для активного захвата пищи; удлинении пищеварительной трубки и все возрастающей дифференциации ее на разные отделы (желудок, тонкий и толстый отделы кишечника); сильном развитии пищеварительных желез в стенках желудка и тонкой кишки; появлении поджелудочной железы, играющей важнейшую роль в процессах переваривания белков, углеводов, жиров и других сложных органических веществ; превращении печени из простого выроста средней части кишки у бесчерепных в мощный, паренхиматозный орган, который помимо необходимого участия в работе кишечника является местом, где совершаются сложнейшие биохимические процессы, имеющие исключительно большое значение для обеспечения нормального обмена веществ всего организма.
Дыхательная система. В
отличие от таковой бесчерепных она сильно усложнена. У первичноводных позвоночных около жаберных щелей имеются различные выросты (пластины, лепестки и др.) с густой сетью кровеносных сосудов, благодаря чему резко увеличивается поверхность жаберного аппарата, через который происходит газообмен. У наземных и вторичноводных позвоночных органами дыхания служат легкие, образовавшиеся из жаберных мешков. Внутренняя поверхность легких в процессе эволюции сильно увеличивается. У первичноводных позвоночных и у земноводных значительную роль в газообмене играет кожа.
Кровеносная система.
У всех позвоночных имеется сердце, развившееся из части брюшного сосуда бесчерепных, окруженное околосердечной сумкой (перикардием). Кровеносная система замкнутая. В крови имеются эритроциты, насыщенные гемоглобином, который поглощает в органах дыхания значительно больше кислорода, чем жидкая часть крови (плазма), и легко отдает его всем органам тела.
Лимфатическая система.
Эта система является посредником между кровеносными капиллярами и тканями тела. Она хорошо развита, ее протоки впадают в главные венозные сосуды. Цвет лимфы желтоватый.
Выделительная система.
Представлена двумя почками, образовавшимися в результате объединения многочисленных, сравнительно просто устроенных выделительных канальцев бесчерепных. Почки в процессе эволюции позвоночных усложнились, и их способность удалять продукты диссимиляции возросла. Выделение продуктов диссимиляции происходит также через органы дыхания и, за исключением пресмыкающихся и птиц, через кожу.
Размножение.
Позвоночные размножаются только половым путем, гермафродитизм — редчайшее исключение. Оплодотворение наружное у подавляющего большинства первичноводных (бесчелюстных и рыб) и земноводных, внутреннее — у ряда рыб и земноводных и у всех позвоночных, размножающихся на суше (пресмыкающихся, птиц и млекопитающих) и произошедших от них вторичноводных групп.
Развитие с превращением (т. е. с личинками) характерно для большинства бесчелюстных, рыб и земноводных. Прямое развитие наблюдается лишь у немногих первичноводных и земноводных и присуще всем настоящим наземным позвоночным (пресмыкающимся, птицам и млекопитающим) и произошедшим от них вторичноводным группам.
К подтипу относится около 42 тыс. видов. Подтип делится на два раздела: бесчелюстные и челюстные с двумя надклассами — рыб и четвероногих. Последние включают классы земноводных, пресмыкающихся, птиц и млекопитающих (рис. 71).
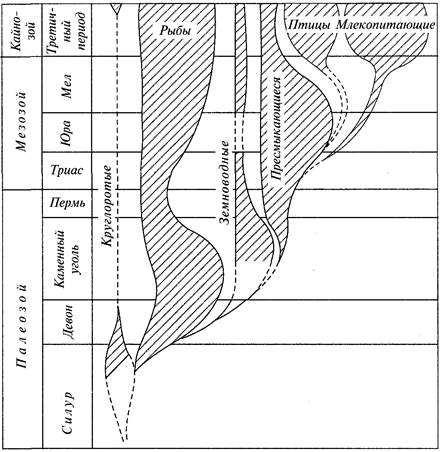
Рис. 71. Родословное древо позвоночных. Толщина ветвей дает
приблизительное представление об обилии или
малочисленности соответствующих групп
Четвероногих обычно подразделяют на две группы — анамний и аминот. Первые являются первичноводной группой, вторые наземные.
Бесчелюстные — самые древние позвоночные, возникшие от наиболее активных бесчерепных, родственных ланцетникам, в ор-довике. Они были многочисленны и широко распространены до середины девона, а потом почти целиком вымерли, не выдержав борьбы за существование с более активными их потомками — хрящевыми рыбами. В настоящее время существует малочисленная группа видов бесчелюстных, которые сохранились благодаря скрытому (в иле) или полупаразитическому образу жизни. К ним относятся два класса, ранее объединявшиеся в один класс кругло-ротые: цефалоспидоморфы (Cephalospidomorphi) с единственным современным отрядом миногообразных (Petromyzontiformes) (около 30 видов), обитающие в морях и пресных водах, и миксины (Myxini) с одним отрядом миксинообразных (Myxiniformes) (около 20 видов), обитающие в морях и океанах.
Тело у бесчелюстных вытянутое, червеобразное (рис. 72), слабее расчлененное, чем у других черепных, на голову, туловище и хвост.
Челюстей нет, рот круглый, способный крепко присасываться к разным субстратам. Кожа голая, очень богатая железами. Длина тела от нескольких десятков сантиметров до 1 м
Нервная система.
Головной мозг у бесчелюстных, как у всех позвоночных, состоит из пяти отделов (рис. 73). Передний мозг мал, нервные клетки сосредоточены на дне его (в так называемых полосатых телах)
и в сильно развитых обонятельных долях.
От них отходят вперед и разветвляются в обонятельной капсуле обонятельные нервы (I пара головных нервов). На утолщенных боковых стенках промежуточного мозга находятся габенулярные ганглии — первичные зрительные центры. От передней части дна промежуточного мозга отходят к сетчатке глаз зрительные нервы (II пара). От зрительных долей среднего мозга отходят к мышцам глаз глазодвига-тельные нервы (III пара головных нервов) и блоковые нервы (IV пара головных нервов). Мозжечок, в функции которого входят координация работы скелетных мышц и обеспечение равновесия тела, развит очень слабо в связи с простотой движения бесчелюстных. Продолговатый мозг развит лучше других отделов головного мозга, от него отходят: тройничные нервы (V пара головных нервов) — к коже передней части головы, слизистой оболочке ротовой полости, мышцам жаберных дуг; отводящие нервы (VI пара головных нервов) — к прямой мышце глаза; лицевые нервы (VII пара головных нервов) — к разным частям головы; слуховые нервы (VIII пара головных нервов) — к органу равновесия и слуха; языкоглоточные нервы (IX пара головных нервов) — к слизистой глотки и к другим органам; блуждающие нервы (X пара головных нервов) — к сейсмосен-сорным органам и к другим органам чувств кожи, к мышцам жаберного аппарата, к внутренним органам (сердцу, кишечнику и др.). I, II, VIII пары — чувствующие нервы; III, IV, VI пары — двигательные нервы; V, VII, IX, Х — смешанные нервы (одни ветви их — чувствующие, другие — двигательные).
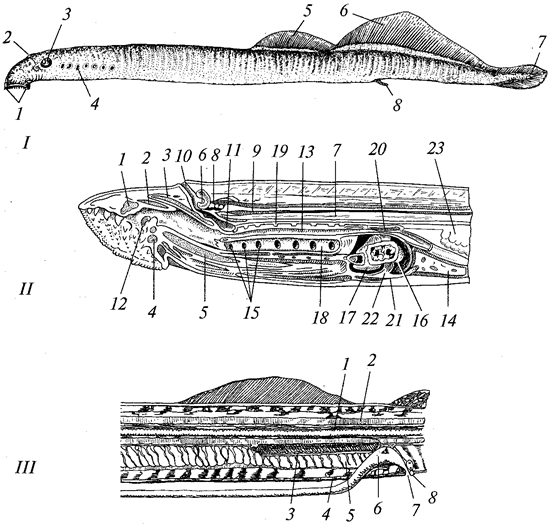
Рис. 72. Минога:
7 — внешний вид: 7 — ротовая присоска; 2 —
ноздря; 3 —
глаз; 4 —
жаберные щели; 5, 6 —
спинные плавники; 7 — хвостовой плавник; 8 —
мочеполовой сосочек; II
— продольный разрез через головной отдел самки: 7 — кольцевой хрящ; 2 —
передний верхний хрящ; 3 —
задний верхний хрящ; 4 —
кольцевой хрящ; 5 —
подъязычный хрящ; 6 —
обонятельная капсула; 7 — хорда; 8 —
головной мозг;
9 —
спинной мозг; 10 —
ноздря; 77 — гипофизарный вырост; 12 —
зубы язычковой пластинки; ]3 —
пищевод; 14 —
печень; 75 — три передние жаберные отверстия; 16 —
предсердие; 17 —
желудочек сердца; 18 —
дыхательная трубка; 19 —
спинная аорта; 20 —
место вхождения кардинальных вен в венозную пазуху; 21 —
печеночная вена; 22 —
венозная пазуха; 23 —
яичник;
III
—
продольный разрез задней части туловища самца: 7 — хорда; 2 —
спинной мозг; 3 —
туловищная почка; 4 —
мочеточник; 5 —
семенник; 6 —
задняя кишка;
7 — анус; 8 —
мочеполовой сосочек с мочеполовым отверстием на вершине
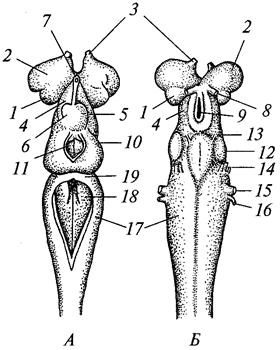
Рис. 73. Мозг миноги:
А —
сверху; Б —
снизу: / — передний мозг; 2 —
обонятельные доли; 3 —
обонятельный нерв; 4 —
промежуточный мозг; 5 и 6 —
правый и левый габенулярные ганглии; 7— пинеальный орган (эпифиз), прикрывающий париетальный (теменной) орган; 8 —
зрительный нерв; 9 —
воронка; 10 —
зрительные доли; 11 —
отверстие в крыше среднего мозга; 12 —
дно среднего мозга; 13 —
глазодвигательный нерв; 14 —
тройничный нерв; 15 —
лицевой нерв; 16 —
слуховой нерв; 17 —
продолговатый мозг; 18 —
ромбовидная ямка; 19 —
зачаточный мозжечок •
Спинной мозг тянется от продолговатого мозга до конца тела, форма его лентовидная. Нервные клетки сосредоточены внутри него, окружая узкую полость — невроцель,
а другие клетки (не нервные, служащие для опоры) — в наружном слое. От спинного мозга отходят спинномозговые нервы, число которых соответствует числу мышечных сегментов; каждый нерв отходит от мозга двумя корешками — спинным и брюшным. Нервы, связанные со спинными корешками, — чувствующие (передающие через спинной мозг в головной мозг раздражения из разных частей тела); нервы, связанные с брюшными корешками, — двигательные (передающие нервные импульсы от головного мозга через спинной мозг иннервируемым ими частям тела). Спинномозговые нервы иннервируют кожу, мышечную систему туловища и хвоста, внутренние органы.
Головные и спинномозговые нервы черепных возникли из нервов бесчерепных, связанных с нервной трубкой, которые ввиду меньшей дифференциации организма последних выполняют сходные функции.
Перечисленные десять пар головных нервов бесчелюстных имеются и у всех вышестоящих классов — хрящевых и костных рыб, земноводных, пресмыкающихся, птиц и млекопитающих, что свидетельствует о монофилетическом происхождении подтипа черепных. Однако в связи с появлением в процессе эволюции новых органов или редукцией органов, потерявших свое значение вследствие изменения образа жизни, функции нервов соответственно изменялись и могли развиваться новые ветви упомянутых нервов или исчезать ранее бывшие ветви. У трех высших классов позвоночных развились еще две пары головных нервов, связанных с продолговатым мозгом: добавочные (XI пара), ин-нервирующие определенные мышцы, и подъязычные (XII пара), особенно хорошо выраженные у млекопитающих.
Органы чувств.
У бесчелюстных органы чувств значительно сложнее аналогичных органов бесчерепных. Орган обоняния —
обширная камера со складчатой внутренней поверхностью, открывающаяся наружу одной ноздрей, а не двумя, как у остальных позвоночных. Однако у зародышей бесчелюстных закладываются две камеры. Следовательно, непарный орган обоняния возник у них по какой-то причине вторично. Обоняние в жизни этих животных играет первостепенную роль в отыскании жертвы (как правило, разных рыб), и обонятельные доли переднего мозга у бесчелюстных, как уже было отмечено, очень хорошо развиты.
Большое значение для ориентации во внешней среде имеют сейсмосенсорные органы
(называемые обычно органами боковой линии},
которые находятся в небольших углублениях кожи на голове, а также по бокам тела и состоят из чувствующих клеток со жгутиками, способных воспринимать токи воды, отходящие от плавающих животных, от других передвигающихся в воде объектов (например, судов), к которым бесчелюстные могут прикрепиться. Эти же органы воспринимают волны, вызываемые движением самих животных, а затем отраженные от упомянутых объектов, от участков берега и неподвижных подводных объектов.
Органы равновесия и слуха —
перепончатые лабиринты — расположены в задних отделах мозгового черепа. Каждый лабиринт состоит из двух мешочков: нижнего — круглого и верхнего — овального, от которого отходят два полукружных канала. Лабиринт наполнен жидкостью — эндолимфой,
в которой во взвешенном состоянии находятся мелкие известковые частицы — отоко-нии
(или отолиты).
Между стенками лабиринта и частями черепа, окружающими его, тоже имеется жидкость — экзолимфа.
Верхний мешочек с полукружными каналами выполняет функции органа равновесия: при изменении положения тела отоконии перемещаются и давят на определенные части стенок мешочка и полукружных каналов. Возникающие раздражения воспринимаются чувствующими клетками и по определенным ветвям VIII пары нервов передаются в мозжечок, откуда через спинной мозг идут импульсы к мышцам, которые обеспечивают нужное в данных условиях положение тела или быстро меняют его, если это необходимо. Нижний мешочек воспринимает раздражения, вызываемые звуковыми волнами, и передает их по слуховым ветвям VIII пары нервов в головной мозг. Слух у бесчелюстных, по-видимому, -развит слабо.
У миног два глаза, состоящих из тех же частей, что и у других позвоночных: белковой (с роговицей), сосудистой, радужной (со зрачком) и сетчатой оболочек, хрусталика и стекловидного тела. Следовательно, глаза бесчелюстных устроены довольно сложно, однако в связи с их образом жизни (закапывание в ил, внедрение в тело жертвы) эти органы упростились, а у миксин остались лишь рудименты глаз. У миног есть теменной глаз (теменной орган), развившийся из отростка промежуточного мозга и служащий для восприятия света, проникающего через пленчатую крышу черепа.
Механические, температурные, химические раздражения воспринимаются различными чувствующими клетками и окончаниями нервов, находящимися в коже.
Скелет.
Череп у бесчелюстных уже имеется, но строение его очень примитивное. Его мозговой отдел состоит из хрящевых стенок — боковых и нижней. Крыша мозга представляет собой соединитель-нотканную пленку. Затылочная часть черепа также не развита. С мозговым черепом соединены: спереди — обонятельная камера, а сзади с боков — правая и левая слуховые капсулы. Висцеральный отдел черепа состоит из губных хрящей, укрепляющих сосательный аппарат этих позвоночных, и хрящевого жаберного скелета, представляющего собой своеобразную решетку, образовавшуюся из слившихся дуг. К черепу сзади внизу примыкает хрящ, защищающий сердце. От черепа до заднего конца тела тянется хорда, значительно лучше развитая, чем у бесчерепных, и имеющая более прочную оболочку, спинная часть которой образует защитный футляр для спинного мозга. Вдоль верхней стороны хорды у миног расположены маленькие хрящевые дуги, по две в каждом сегменте, которые являются зачатками позвонков. У миксин названных дуг нет.
Мышечная система.
У бесчелюстных она состоит из большого числа сегментов, значительно лучше развитых, чем у бесчерепных, мышц, связанных с висцеральным отделом черепа, и мышц плавников.
Органы передвижения.
Они представлены только непарными плавниками, спинными и хвостовым, лопасти которых поддерживаются тонкими хрящевыми лучами. Бесчелюстные передвигаются, изгибая тело, плавники играют в их передвижении незначительную роль, помогая сохранять определенное направление. Бесчелюстные более сильные пловцы, чем бесчерепные, но они не способны, как рыбы, к длительному быстрому плаванию, часто присасываются к рыбам, судам и к другим плывущим предметам, много времени проводят на дне, в иле.
Пищеварительная система.
У бесчелюстных она начинается пред-ротовой воронкой, на внутренней поверхности которой образуются роговые зубчики и пластинки. Во рту помещается язык с роговыми зубцами на вершине и открываются протоки слюнных желез, выделяющих антикоагулирующие вещества. Бесчелюстные присасываются к добыче, продырявливают ее кожу и проникают в тело жертвы. Протеолитические ферменты, попадающие в рану, растворяют белки (внекишечное пищеварение), затем пища всасывается. Глотка разделена на два отдела: верхний, переходящий в пищевод, и нижний, переходящий в дыхательную трубку (слепо заканчивающуюся), в которую открываются внутренние жаберные щели. Пищевод переходит в кишку, заканчивающуюся анальным отверстием. Желудка у круглоротых нет. Кишечная трубка по сравнению с кишечником рыб короче и не образует петель. У миног на внутренней стенке кишки имеется спиральная складка, увеличивающая внутреннюю поверхность этой части пищеварительного аппарата. Печень представляет собой большой паренхиматозный орган. Имеется желчный пузырь, выводной проток которого открывается в начальную часть кишки. Поджелудочная железа представлена отдельными вкраплениями в стенках начала кишки и в печени. Многие виды миног и все мик-сины, как отмечалось выше, нападают на рыб и, вгрызаясь в их тело, выедают все внутренности. Мелкие виды миног питаются разными беспозвоночными.
Дыхательная система.
Главные органы дыхательной системы — жаберные мешки, образующиеся из выростов глотки, которые соединяются с выростами наружного покрова и открываются наружу жаберными щелями. Число последних чаще равно семи. Внутренняя поверхность мешков образует складки (лепестки) энтодермально-го происхождения, вследствие чего увеличивается поверхность газообмена. При вдохе в мешки из глотки поступает вода и значительная часть растворенного в ней кислорода поглощается жаберными лепестками. При выдохе вода с уменьшенным количеством кислорода и увеличенным количеством углекислого газа выходит через жаберные щели наружу. Если бесчелюстные присасываются на некоторое время к рыбам или к другим движущимся объектам, то вода поступает в жаберные мешки и выходит из них через жаберные щели. Газообмен частично совершается и через кожу.
Кровеносная система.
Бесчелюстные, как и все позвоночные, имеют сердце. Мышцы сердца поперечнополосатые, мышцы же сосудов гладкие. Благодаря работе сердца кровь движется значительно быстрее, чем у бесчерепных. Сердце состоит из двух камер — предсердия
и желудочка.
В сердце (сначала в предсердие, потом в желудочек) поступает по венам кровь со всего тела, насыщенная углекислым газом и бедная кислородом. От желудочка отходит артериальный сосуд — брюшная аорта, начало которой расширено и названо луковицей аорты.
От брюшной аорты берут начало несколько пар приносящих жаберных артерий, доставляющих кровь в жабры, где совершается отдача значительной части углекислого газа и насыщение крови кислородом, которая по выносящим жаберным артериям переходит в спинную аорту. Разветвления последней доставляют артериальную кровь во все части тела, снабжая их кислородом и поглощая образовавшийся в результате процессов тканевого дыхания углекислый газ, и по венам направляется в предсердие. Следовательно, у бесчелюстных один круг кровообращения, и через сердце проходит только венозная кровь. Образование форменных элементов крови (эритроцитов, лейкоцитов и тромбоцитов) происходит в тканях разных органов — печени, пищевода, кишки, почек и др. Селезенки у бесчелюстных нет.
Выделительная система.
Главные органы выделительной системы — две почки (лежащие на спинной стороне под хордой), образовавшиеся в результате объединения и усложнения выделительных органов бесчерепных. У зародышей всех бесчелюстных закладываются и функционируют предпочки (пронефрос),
называемые также головными почками,
так как они развиваются в передней части туловища. Основу их составляют почечные канальцы. Каждый канадец начинается открытой в целом воронкой, со стенкой которой соприкасается клубочек кровеносных капилляров (мальпигиево тельце).
Продукты диссимиляции, растворенные
плазме крови, попадают в канадец из сосудов клубочка, а из жидкости вторичной полости тела (целома) — через воронку. Все канальцы впадают в общий выводной проток (мочеточник), открывающийся наружу сзади анального отверстия.
Головные почки у взрослых особей большинства видов бесчелюстных заменяются более сложными туловищными почками (мезо-нефрос),
которые развиваются сзади первых и тянутся в виде лент, не доходя до конца туловища. Строение их канальцев, более сложное. Клубочек капилляров врастает в воронку канальца и вокруг него развивается боуменова капсула, вследствие чего улучшается передача продуктов диссимиляции из крови в канальцы. Последние более длинные, чем канальцы предпочек, и гуще оплетены капиллярами. В боуменовы капсулы из клубочков попадают не только продукты диссимиляции, но и полезные для организма растворимые органические вещества, которые во время прохождения по канальцу могут возвратиться в кровеносное русло, а также слишком много воды. Таким образом, туловищные почки лучше изымают из кровеносного русла продукты диссимиляции и в то же время способствуют сохранению полезных органических веществ и более экономно расходуют воду. Канальцы каждой туловищной почки впадают в общий выводной проток, который назван вольфовым каналом,
открывающийся наружу сзади анального отверстия.
Размножение.
Бесчелюстные раздельнополы. Половые железы (один семенник или один яичник) в период размножения занимают большую часть полости тела. Специальных выводных протоков эти железы не имеют, созревшие гаметы из желез выпадают в полость тела и через поры так называемого мочеполового синуса
(небольшой отграниченной части полости тела) выходят в воду, где происходит оплодотворение.
Многие морские миноги уходят для нереста в реки, некоторые — далеко вверх по течению. Такие виды называются проходными.
Небольшое количество видов миног постоянно обитает в реках. Развитие миног из яиц, бедных желтком, происходит с метаморфозом, у них развиваются личинки, имеющие эндостиль, схожий с таким же органом бесчерепных, который обеспечивает им питание мелкими организмами и органическими остатками. Личинки, как и ланцетники, обитают на дне и называются пескоройками.
Они живут несколько лет и после глубокой перестройки организма становятся взрослыми животными. Наличие стадии пескоройки подтверждает происхождение бесчелюстных от бесчерепных. У миксин развитие прямое.
Практическое значение.
Различные виды миног употребляют в пищу. Бесчелюстные, нападающие на промысловых рыб, причиняют вред, иногда очень значительный, в особенности рыбам, находящимся в сетях.
Челюстноротые — группа позвоночных, для которых характерно появление в висцеральном отделе черепа челюстей, служащих для захвата и измельчения добычи. У большинства их хорда хорошо развита лишь у личинок. Мозговой череп окружает мозг сверху. Ротовое отверстие выглядит наподобие щели. Обонятельные мешки и ноздри парные. У водных представителей (рыб) есть парные плавники, у наземных — парные конечности.
К этому надклассу относятся первичноводные челюстноро-тые позвоночные, обитающие только в воде. Движение рыб осуществляется с помощью плавников, среди которых выделяются парные — грудные и брюшные. Дышат жабрами. Имеют один круг кровообращения (рис. 74). В коже возникают защитные образования — чешуи. Хорошо развиты органы боковой линии.

Рис. 74. Схема кровообращения рыбы (Г)
и наземного позвоночного (II):
1 —
жаберные артерии; 2 —
сонная артерия; 3 —
спинная аорта; 4 —
брюшная аорта; 5—легочная артерия; 6—
кювьеров проток; 7— передняя кардинальная (яремная вена); 8 —
задняя кардинальная вена; 9 —
задняя полая вена; 10 —
печеночная вена; 11 —
легочная вена; 12 —
воротная вена печени; 13 —
подкишечная вена
Надкласс включает около 20 тыс. видов, распространенных в морях и пресных водах земного шара. Надкласс делится на 2 класса — хрящевые и костные рыбы (рис. 75, 76).

Рис. 75. Филогенетическое древо панцирных и хрящевых рыб
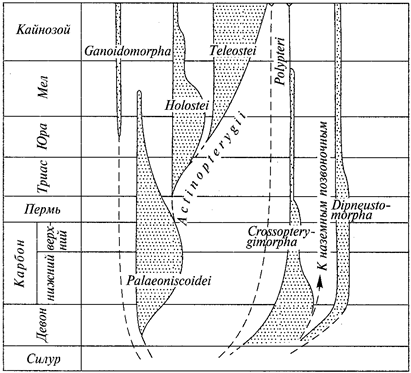
Рис. 76. Филогенетическое древо костных рыб
Хрящевые рыбы возникли в верхнем силуре от бесчелюстных, переходивших к более быстрому, длительному плаванию и более успешному захвату добычи вооруженным челюстями ртом. Они были первыми челюстноротыми позвоночными и господствовали, постепенно эволюционируя, до середины мезозойской эры, когда началось их вытеснение высшими костными рыбами. В настоящее время существует только одна небольшая группа хищных хрящевых рыб, названных пластиножаберными. Они широко распространены в морях. К пластиножаберным относятся акулы — прекрасные пловцы — и скаты, ведущие малоподвижный образ жизни на дне. Акул около 350 видов, скатов около 340 видов. Большинство хрящевых рыб имеют крупный размер. Длина самых больших акул достигает 15—20 м, скатов — 6—7 м. Мелких видов мало.
Форма тела акул (рис. 77) типично рыбообразная, приспособленная для быстрого плавания. У скатов форма тела уплощенная (позволяющая им плотно прижиматься к поверхности дна брюшной стороной и становиться незаметными для их жертв и врагов), хвост тонкий, не имеющий существенного значения для плавания.
Рис. 77. А —
акула нокотница (катран) (Squalusacanthias);
Б —
скат морской кот (Trygonpastinaca) из семейства хвостоколов
Покровы.
Тело хрящевых рыб в отличие от бесчелюстных не голое, а покрыто мелкими костными чешуйками, каждая из которых состоит из пластинки и зуба. Эти чешуйки называются плакоидны-ми,
они развиваются из мезенхимы, а эмаль, покрывающая зуб, — из эктодермы. Зубы чешуи вполне похожи по своему строению и развитию на зубы других позвоночных. Чешуя защищает тело рыб от механических повреждений. Как у всех рыб, кожа богата железами, выделения которых уменьшают трение, возникающее при плавании. Вещества, выделяемые железами некоторых видов этих рыб, могут быть ядовитыми для животных и человека.
Нервная система.
В связи с активизацией образа жизни нервная система хрящевых рыб развилась значительно лучше, чем у бесчелюстных. Размеры головного мозга и всех его отделов возросли, связи между отделами усилились, строение их усложнилось (рис. 78).
Рис. 78. Головной мозг акулы сбоку:
1 —
передний мозг; 2 —
обонятельные доли; 3 —
промежуточный мозг;
4 —
нижние доли; 5 — сосудистый мешок; 6 —
гипофиз; 7 — зрительный нерв; 8 —
эпифиз; 9 —
средний мозг; 10 —
мозжечок; 11 —
продолговатый мозг
Обонятельные доли переднего мозга очень велики, и главная его функция по-прежнему заключается в восприятии химических раздражении. Мозжечок в связи с приспособлением хрящевых рыб (это в особенности касается акул) к продолжительному и быстрому плаванию и необходимостью поддерживать равновесие тела сильно увеличился. Усложнилось и строение спинного мозга, форма его в поперечном сечении, как у всех вышестоящих позвоночных, округлая, а не лентовидная, как у бесчелюстных.
Органы чувств.
Все органы чувств усложнились. Обонятельные камеры парные. Сейсмосенсорные органы сосредоточены в каналах, идущих от головы к заднему концу тела. Размеры глаз увеличились, хрусталик шаровидный, но, как и у других рыб, дальность зрения незначительна; цвета хрящевые рыбы не различают. У лабиринта появился третий полукружный канал,
лежащий в другой плоскости, нежели полукружные каналы у бесчелюстных, что содействует более точному восприятию изменения положения тела в пространстве.
Скелет. В
связи с необходимостью иметь более надежную защиту для нервной системы и других органов и более прочную опору для сильно возросшей мышечной системы у рассматриваемых рыб развился хрящевой скелет, часто обызвествленный (рис. 79). Мозговой отдел черепа большей величины, чем у бесчелюстных, закрытый со всех сторон (с небольшим отверстием посредине крыши). В состав висцерального отдела черепа входят: два нёбноквадратных хряща
(правый и левый), выполняющих роль верхних челюстей; два меккелевых хряща
(правый и левый), служащих как нижние челюсти; подъязычная дуга,
состоящая из двух верхних хрящей (правого и левого), которые прикрепляются к мозговому черепу там, где находятся лабиринты, и двух нижних хрящей (подъязычных); членистые жаберные дуги
(правые и левые, число которых у большинства видов 5—7).
Рис. 79. Череп акулы сбоку:
/ — рострум; 2 — обонятельная капсула; 3 —
глазница; 4 —
слуховой отдел; 5 — затылочный отдел (1—5 —
мозговой череп); 6 —
нёбно-квадратный хрящ; 7— меккелев хрящ; 8—
подвесок (гиомандибуляре); 9 —
гиоиод; 10 —
копула подъязычной дуги; 11 —
жаберные дуги; 12 —
жаберные лучи; 13 —
губные хрящи (6—13 —
висцеральный скелет); 14 —
челюстной сустав; 15 —
связка; 16 —
зубы
На месте хорды развился позвоночный столб. Каждый позвонок состоит из тела с вогнутыми поверхностями (передней и задней). Такие позвонки называются двояковогнутыми
или амфицелъ-ными.
Хорда сохраняется в виде узкого стержня, проходящего через центры тел позвонков. От тел позвонков вверх отходят два отростка, которые образуют верхнюю дугу с остистым отростком.
От тел позвонков книзу отходят образующие нижнюю дугу поперечные отростки, а от них — короткие ребра. В канале, который образуется отверстиями в верхних дугах позвонков и вставочными пластинками между позвонками, находится спинной мозг.
Органы передвижения. У
хрящевых рыб кроме непарных плавников (спинных, анального и хвостового), которые имеются и у бесчелюстных, появились парные плавники —
грудные и брюшные, играющие немаловажную роль в поддержании тела в нормальном положении (спиной вверх, брюхом вниз) и выполняющие некоторые другие функции. Лопасти всех плавников имеют скелет из роговых лучей.
Опорой для спинных и анального плавников служат хрящи, расположенные в мышцах, для хвостового — задний конец позвоночника, для грудных — плечевой пояс, для брюшных — тазовый пояс.
Мышечная система.
В основном она состоит, как и у бесчелюстных, из большого количества боковых сегментов, масса и сила которых сильно возросли. Очень мощные мышцы находятся у акул в хвостовой части тела. Развиты мышцы, обеспечивающие движение плавников, жаберного аппарата и челюстей.
Пищеварительная система.
Подавляющее большинство пласти-ножаберных — хищники. Ротовая полость обширна, челюсти вооружены крепкими острыми зубами, возникшими из плакоидных чешуи, переместившихся в процессе эволюции в рот. Зубы расположены в несколько рядов. По мере снашивания зубы переднего ряда замещаются зубами следующего ряда. За ротовой полостью идут глотка, короткий пищевод, объемистый желудок, кишка и клоака (рис. 80).
Рис. 80. Вскрытая акула (самец):
1
- ноздря; 2 —
ротовая щель; 3 —
жабры: 4 —
наружные отверстия жаберных щелей; 5 — венозная пазуха; 6 —
предсердия;. 7 — желудочек, 8 —
артериальный конус; 9 —
брюшная аорта; 10 —
приносящие жаберные артерии; 11 — карди-альная часть желудка; 12 —
пилорическая часть желудка; 13 —
тонкая кишка; 14 —
вскрытая толстая кишка со спиральным клапаном; 75 — прямая кишка; 16 —
ректальная железа; 17 —
клоака; 18 —
печень; 19 —
желчный пузырь; 20 —
желчный проток; 21 —
поджелудочная железа; 22 —
селезенка; 23 —
почка; 24 —
семяпровод; 25 —
копулятивный отросток брюшного плавника; 26 —
щитовидная железа; 27 —
семенник
В начальную часть кишки открываются протоки печени и поджелудочной железы. Кишка короче, чем у костистых рыб, но ее внутренняя поверхность сильно увеличена благодаря наличию спиральной складки, число оборотов которой может достигать пятидесяти.
Печень, состоящая из трех лопастей, очень велика, у некоторых видов ее масса достигает 14—25 % всего тела. В ней может накапливаться много жира, который потребляется в периоды недостатка пищи или усиленной затраты энергии. Накопление жира уменьшает удельную массу тела, что способствует повышению плавучести этих рыб. Поджелудочная железа еще не имеет компактной формы и представлена, как у круглоротых, отдельными участками, прилегающими к стенкам начала кишки и печени.
Пищей для пластиножаберных служат различные рыбы. Многие из них поедают ракообразных, моллюсков, червей и других беспозвоночных. Интересно, что самые большие акулы — китовая (длина тела 15 м) и гигантская (длина тела 20 м) — питаются, подобно усатым китам, планктоном, процеживая через свою пасть воду.
Дыхательная система.
Основную часть дыхательной системы составляют большие жаберные пластины эктодермального Происхождения, прикрепленные одной из сторон к межжаберным перегородкам. Большая поверхность пластин обеспечивает достаточно интенсивный газообмен. При вдохе глотка расширяется, вода поступает через рот и омывает жаберные пластины. При выдохе объем глотки уменьшается и вода выходит наружу через жаберные щели, число которых у большинства видов 5, а у меньшинства видов 7. Жаберных крышек у хрящевых рыб нет. Кроме упомянутых жаберных щелей у большинства видов сзади глаз имеются рудиментарные жаберные щели — брызгальца,
открывающиеся в переднюю часть глотки, куда через них поступает вода при вдохе.
У скатов жаберные щели находятся на брюшной стороне; вода в жаберный аппарат попадает у них только через брызгальца. Газообмен частично совершается и через кожу.
Кровеносная система.
Сердце, как и у бесчелюстных, состоит из предсердия и желудочка, но величина и сила его мускулатуры возросли в связи с более активным образом жизни этих рыб (см. рис. 80). От желудочка отходит артериальный конус,
мышечная ткань которого такая же, как и сердца, поперечнополосатая, способная к сильным сокращениям. Внутри конуса имеется несколько пар клапанов, препятствующих возвращению крови обратно в желудочек. Из артериального конуса кровь переходит в брюшную аорту и далее по приносящим жаберным артериям в жабры, где насыщается кислородом и отдает углекислый газ, а затем через жаберные выносящие артерии, впадающие в спинную аорту, и многочисленные артериальные сосуды разносится по всему телу. После отдачи кислорода тканям и насыщения углекислым газом кровь по венам попадает в сердце. Следовательно, у хрящевых рыб, как и у бесчелюстных, один круг кровообращения и через сердце проходит только венозная кровь. Количество эритроцитов у этих рыб значительно больше, чем у бесчелюстных, что способствует лучшему снабжению кислородом тканей. Имеется селезенка,
играющая роль депо крови, откуда она может быстро поступать в те части тела, работа которых усилилась. В селезенке, а также в тканях почек образуются эритроциты, лейкоциты и тромбоциты.
Выделительная система.
Она представлена двумя длинными туловищными почками, строение которых по сравнению с теми же органами бесчелюстных несколько усложнилось: многие почечные канальцы лишены воронок и начинаются более крупными гломерулами (мальпигиевыми тельцами) — клубочками капилляров, окруженными боуменовыми капсулами. В результате этих изменений значительная часть продуктов диссимиляции поступает в канальцы не из целома, а из кровеносных сосудов. Канальцы удлинились, количество оплетающих их капилляров увеличилось. Благодаря этому в кровь возвращается больше растворенных органических веществ и воды, попавших из мальпигиевых телец в бо-уменовы капсулы. Моча удаляется через заднюю часть кишки — клоаку.
Размножение.
Пластиножаберные раздельнополы. Семенники (два вытянутых тела) лежат в брюшной полости на уровне пищевода. От них отходят тонкие семявыносящие каналы, впадающие в передние части почек, откуда сперматозоиды по вольфовым каналам попадают в клоаку. Таким образом, вольфовы каналы служат у самцов мочеточниками и семяпроводами. Яичники (их два) расположены в брюшной полости там, где у самцов находятся семенники. Созревшие яйцеклетки попадают в полость тела, а из нее — в воронки яйцеводов. Следовательно, прямого соединения яичников с яйцеводами нет. Яйцеводы образуются следующим образом: во время эмбрионального развития этих рыб зачаток канала туловищной почки расщепляется продольно на два канала — вольфов
и мюллеров.
Первый становится у обоих полов мочеточником, второй превращается у самок в яйцевод, а у самцов остается в виде рудимента или совсем исчезает. Яйцеводы открываются в клоаку.
Оплодотворение внутреннее, семя вводится самцами в клоаку самок при помощи совокупительных органов, образовавшихся из передних частей брюшных плавников, расположенных около клоаки. Сперматозоиды, попавшие из клоаки самок в яйцеводы, оплодотворяют яйцеклетки в верхней части этих трубок.
Развитие зигот совершается различными способами. У большинства видов яйцеклетки, содержащие много желтка, после оплодотворения спускаются по яйцеводу, в разных отделах его окружаются твердыми прочными оболочками и выходят наружу, где происходит развитие зародышей до взрослого состояния. У других видов зиготы, достаточно богатые желтком, не окружаются твердыми оболочками и развиваются в задних отделах яйцеводов. Такой способ развития называется яйцеживорождением.
Он встречается и у разных беспозвоночных и позвоночных. Наконец, у некоторых видов наблюдается настоящее живорождение:
яйцеклетки, не богатые запасными веществами, после оплодотворения спускаются по яйцеводам и прикрепляются в расширенных частях последних (в матках)
при помощи желточного мешка к их стенкам, откуда получают все нужные для развития вещества. Следовательно, у хрящевых рыб существуют разные способы защиты эмбрионов от неблагоприятных условий и от поедания их другими животными. Эти рыбы производят по сравнению с костными рыбами небольшое количество эмбрионов. Однако несмотря на эти приспособления и довольно хорошее развитие нервной и двигательной систем, число видов пластиножаберных во много раз меньше, чем ныне существующих костных рыб. Распространение их ограниченно: они обитают в океанах и связанных с ними морях преимущественно в субтропических и тропических районах, становятся редкими в районах умеренного климата, отсутствуют в приполярных районах. Все это свидетельствует о недостаточной приспособленности пластиножаберных к жизни в водной среде в условиях конкуренции их с современными костными рыбами.
Практическое значение.
В общем промысле рыб хрящевые рыбы составляют около 1 %, но в ближайшее время использование их в качестве источника пищевых продуктов и для ряда других целей, несомненно, возрастет. Одновременно нужно иметь в виду и отрицательное их значение: они поедают полезных рыб и в ряде районов опасны для людей.
Костные рыбы — самый большой класс позвоночных животных, насчитывающий около 20000 видов. Самые древние представители этого класса произошли от хрящевых рыб в конце силура. В настоящее время 99 % класса принадлежат к так называемым костистым рыбам,
которые впервые появились в середине триаса, но их эволюция долгое время шла медленно и только в конце мелового периода резко ускорилась и достигла удивительного расцвета в третичном периоде. Они населяют самые разнообразные водоемы (реки, моря и океаны вплоть до самой большой глубины, встречаются в арктических водах). Таким образом, костистые рыбы — наиболее приспособленные к обитанию в водной среде позвоночные. Кроме костистых рыб в состав класса входит еще несколько десятков видов древних костных рыб, сохранивших некоторые особенности хрящевых рыб.
Общая характеристика
Большинство видов этого класса приспособлено к быстрому плаванию, и форма их тела сходна с таковой акул. Менее быстро плавающие рыбы имеют более высокое тело (например, у многих видов карповых рыб). Виды, ведущие малоподвижный образ жизни на дне (например, камбалы), имеют такую же уплощенную форму тела, как скаты (рис. 81).

Рис. 81. Костистые рыбы:
1 —
сельдь (сем. Сельдевые); 2 —
лосось (сем. Лососевые); 3 —
карп (сем. Карповые); 4—
сом (сем. Сомовые); 5—
щука (сем. Щуковые); 6—
угорь (сем. Угревые);
7 — судак (сем. Окуневые); 8 —
бычок речной (сем. Бычковые); 9 —
камбала (сем. Камбаловые)
Покровы.
Длина тела рыб различна — от нескольких сантиметров до нескольких метров. В отличие от хрящевых и древних костных рыб среди костистых множество мелких видов, освоивших малые биотопы, недоступные для более крупных видов. Кожа подавляющего большинства костистых рыб покрыта небольшими костными, сравнительно тонкими чешуями, черепицеобразно налегающими друг на друга. Они хорошо защищают рыб от механических повреждений и обеспечивают достаточную гибкость тела. Различают циклоидную
чешую с закругленным верхним краем и ктеноидную
с мелкими зубчиками на верхнем крае. Количество чешуи в продольных и поперечных рядах для каждого вида более или менее постоянно и учитывается при определении видовой принадлежности рыб. В холодное время рост рыб и чешуи замедляется или прекращается, поэтому на чешуе образуются годовые кольца, подсчитывая которые можно определить возраст рыбы. У ряда видов кожа голая, лишенная чешуи. В коже множество желез, выделяемая ими слизь уменьшает, как и у других рыбообразных, трение при плавании, защищает от бактерий и т. д. В нижних слоях эпидермиса имеются различные пигментные клетки, благодаря которым рыбы малозаметны на фоне окружающей их среды. У некоторых видов окраска тела может меняться в соответствии с изменениями окраски субстрата. Подобные изменения осуществляются под влиянием нервных импульсов.
Нервная система.
Размеры головного мозга по отношению к величине тела несколько больше, чем у хрящевых рыб. Передний мозг относительно мал по сравнению с другими отделами, но полосатые тела его велики и посредством связей их с другими отделами Центральной нервной системы влияют на осуществление некоторых довольно сложных форм поведения. Нервные клетки в крыше переднего мозга отсутствуют. Промежуточный мозг и отделившиеся от него эпифиз и гипофиз хорошо развиты. Средний мозг крупнее других отделов головного мозга, в верхней его части имеются две хорошо развитые зрительные доли. Мозжечок у хорошо плавающих рыб велик. Возросли размеры и усложнилось строение продолговатого и спинного мозга. Подчинение последнего головному мозгу по сравнению с тем, что наблюдается у хрящевых рыб, усилилось (рис. 82).
Рис. 82. Головной мозг окуня:
1 —
обонятельная капсула; 2 —
обонятельные доли; 3 —
передний мозг; 4 —
средний мозг; 5 — мозжечок; 6 —
продолговатый мозг; 7 — спинной мозг; 8 —
глазничная ветвь тройничного нерва; 9 —
слуховой нерв; 10 —
блуждающий нерв
Органы чувств.
Произошли некоторые усложнения в органах чувств. Сейсмосенсорные органы расположены в каналах боковых стенок тела и образуют густую сеть на голове. Более развит круглый мешочек лабиринта, и вопреки долго существовавшему мнению, что костистые рыбы воспринимают лишь грубые сотрясения окружающей среды, многочисленными экспериментами доказано, что они реагируют на разнообразные звуковые колебания и используют их для общения внутри популяции. Способность к восприятию различных химических раздражении выражена очень хорошо. Многие виды воспринимают даже незначительные изменения температуры в окружающей их воде. Зрение костистых рыб рассчитано, как и у других рыб, на близкое расстояние; хрусталик шаровидный, неспособный менять свою кривизну, резкость изображения достигается перемещением его при помощи сокращения особой мышцы — серповидного отростка.
Скелет.
В течение эволюции рассматриваемого класса скелет постепенно окостенел. Хорда сохранилась лишь у низших представителей класса, число которых незначительно. При изучении скелета нужно иметь в виду, что одни кости возникают в результате замещения хрящей костной тканью, другие развиваются в соединительнотканном слое кожи. Первые называются основными,
вторые — покровными костями.
Мозговой отдел черепа представляет собой коробку, защищающую головной мозг и органы чувств: обоняния, зрения, равновесия и слуха (рис. 83).
Рис. 83. Схема расположения костей в черепе костистой рыбы. Висцеральный скелет отделен от мозгового черепа. Жаберная крышка не нарисована. Основные кости и хрящ покрыты точками, покровные кости — белые:
/ — угловая; 2 — сочленовная; 3 —
основная затылочная; 4 —
основная клиновидная; 5 — копула; 6 —
зубная; 7 — боковая обонятельная; 8 —
наружная крыловидная; 9 —
внутренняя крыловидная; 10 —
боковая затылочная; 11 —
лобная; 12 —
подвесок; 13 —
гиоид; 14 —
окостеневшая связка; 15 —
боковая клиновидная; 16 —
средняя обонятельная; 17—
задняя крыловидная; 18—
верхнечелюстная; 19 —
носовая; 20 —
глазоклиновидная; 21 —
теменная; 22 —
нёбная; 23 —
предчелюстная; 24 —
парасфеноид; 25 —
квадратная; 26 —
верхняя затылочная; 27 —
дополнительная; 28 —
сошник; 29—33 —
ушные кости; I— V —
жаберные дуги
Крыша черепа образована парными носовыми, лобными, теменными костями. Последние примыкают к верхней затылочной кости, которая вместе с парными боковыми затылочными костями и основной затылочной костью образует заднюю часть черепа. Низ черепа состоит (спереди назад) из сошника, парасфеноида (широкой длинной кости, очень характерной для черепа рыб) и основной кости. Передняя часть черепа занята капсулой, охраняющей органы обоняния; по бокам расположены кости, окружающие глаза, и ряд костей (обычно 5), защищающих органы слуха и равновесия.
Висцеральный отдел черепа состоит из ряда костных жаберных дуг, являющихся опорой и защитой жаберного аппарата и передней части пищеварительной системы. В состав каждой из упомянутых дуг входит несколько косточек. Дуг, к которым прикрепляются жабры, у большинства рыб 5 (с каждой стороны). Внизу жаберные дуги соединяются между собой, а передняя из них связана с подъязычной дугой, которая состоит из нескольких костей. Верхняя из этих косточек — подъязычно-челюстная (гиомандибуляре) прикрепляется к мозговому отделу черепа в области слухового отдела и связана через квадратную кость с костями, окружающими ротовую полость. Таким образом, подъязычная дуга служит для соединения жаберных дуг с остальными частями висцерального отдела, а ее верхняя кость — с мозговым отделом черепа.
Края рта и вся ротовая полость укреплены рядом костей. Верхнечелюстной ряд костей представлен (с каждой стороны) межчелюстной и верхнечелюстной костями. Далее идет ряд костей: нёбная, несколько крыловидных и квадратная. Квадратная кость вверху примыкает к подвеску (гиомандибуляре), а внизу — к нижней челюсти. Последняя состоит из нескольких костей: зубной (самой большой), угловой и сочленовной, соединяющейся с квадратной костью. У древних рыб (имевших еще хрящевой скелет) все дуги висцерального отдела черепа несли жабры, впоследствии же передние из этих дуг превратились в подъязычные дуги и челюстные ряды костей.
Позвоночный столб состоит из большого количества двояковогнутых (амфицельных) позвонков, в промежутках между которыми сохраняются остатки хорды. От каждого позвонка отходит вверх и несколько назад длинный остистый отросток. Основания этих отростков разделены, и они образуют канал, по которому проходит спинной мозг. От нижней стороны тел позвонков отходят два коротких поперечных отростка, к которым в туловищном отделе прикрепляются длинные изогнутые ребра. Они свободно оканчиваются в мышцах и образуют каркас боковых стенок тела. В хвостовой части тела от позвонков книзу отходят только нижние остистые отростки.
Органы передвижения.
У костных рыб, как и у хрящевых, они представлены непарными (спинные, анальный, хвостовой) и парными (грудные и брюшные) плавниками. Лопасти плавников поддерживаются костными лучами. Одни лучи — мягкие, состоящие из ряда костных участков, другие — твердые, цельные, концы которых у многих видов заострены. Лучи спинных и анального плавников опираются на косточки — базалии
(плавниковые подпорки), лежащие в мышцах у края тела. Парные плавники опираются на лежащие между мышцами пояса конечностей: грудные — на плечевой (или передний) пояс, состоящий из нескольких костей, верхняя из которых прикрепляется к черепу (рис. 84), брюшные — на тазовый (или задний) пояс, состоящий, как правило, из пары костей.
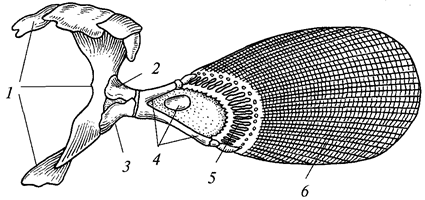
Рис. 84. Пояс передних конечностей и грудной плавник лучеперых рыб:
/ — первичный пояс; 2 —
лопатка; 3 —
коракоид; 4 —
базальные элементы; 5 — радиалии; 6 —
кожные лучи
Лучи хвостового плавника опираются на заднюю часть позвоночного столба. У низших костных рыб (как и у хрящевых) хвостовой плавник гетероцеркальный, верхняя лопасть которого значительно больше нижней лопасти. У костистых рыб хвостовой плавник — гомоцеркальный,
так как обе лопасти его приблизительно равны.
Постепенное окостенение скелета имело большое приспосо-бительное значение в эволюции рыб, поскольку оно способствовало развитию крепкой опоры для мышц и надежной защиты центральной нервной системы и внутренних органов. Так как в состав костного скелета может входить большое количество мелких косточек, это явилось важным условием для возникновения множества видов, имеющих малую величину тела.
Костный скелет имеет большую массу, чем хрящевой, что могло бы затруднить плавание рассматриваемых рыб. Поэтому у последних развился путем ответвления от передней части кишки плавательный пузырь,
лежащий над кишечником и заполненный смесью газов (азота, кислорода и углекислого газа), благодаря чему масса тела рыб значительно уменьшилась. У примитивных видов костных рыб (открытопузырных)
пузырь остается соединенным с кишечной трубкой в течение всей жизни. У большинства видов (закрытопузырных),
появившихся позднее, он полностью отделяется от кишки. В стенках пузыря имеются густые сплетения капилляров, которые обеспечивают наполнение его газами. Объем газов в закрытом пространстве, как известно, может при сжатии сильно уменьшаться и, наоборот, при уменьшении сжатия резко увеличиваться. Увеличение и уменьшение объема пузыря происходит вследствие работы мышц, окружающих брюшную полость. Поэтому плавательный пузырь не только уменьшает массу тела, но и выполняет также гидростатическую
роль, т. е. облегчает подъем рыбы вверх при его расширении и погружение — при его сжатии. Плавательный пузырь редуцировался у ряда видов рыб, ведущих малоподвижный образ жизни на дне, и у тех видов, которые, обладая сильной мускулатурой, способны быстро перемещаться вверх или вглубь. Наличие пузыря у последних могло бы вызывать при быстром подъеме сильное расширение его и выворачивание внутренностей, как это было доказано многими наблюдениями. У некоторых рыб плавательный пузырь, соединенный рядом косточек (веберов аппарат) с
лабиринтом, способствует передаче в последний некоторых звуковых волн.
Пищеварительная система.
У большинства костных рыб имеются некоторые особенности. Спиральной складки нет, увеличение поверхности кишечника происходит за счет его удлинения. Кроме того, от начальной части кишки у многих высших костистых рыб отходят пилорические отростки,
тоже увеличивающие поверхность кишки. Экскременты удаляются через анальное отверстие; клоаки нет. Строение рассматриваемой системы различно в зависимости от характера питания. У хищников, нападающих на других рыб и более крупных беспозвоночных, широкий рот, обычно усаженный большими острыми зубами; желудок большой, резко отделенный от пищевода и начала кишки, общая длина кишечника значительно короче, чем у растительноядных видов. У последних и в особенности у видов, питающихся мелкими беспозвоночными и органическими остатками, зубы малы или их нет; желудок почти не выражен или отсутствует. У карповых и некоторых других рыб в глотке имеются особые глоточные зубы
для механической обработки пищи. Печень хорошо развита, хотя не достигает такой величины, как у многих хрящевых рыб. Поджелудочная железа представлена отдельными дольками, находящимися в печени или в стенках начальной части кишечника, т. е. не имеет еще компактной формы, но развита лучше, чем у хрящевых рыб.
Изучение состава пищи разных рыб, особенно промысловых, имеет большое практическое значение, так как при наличии сведений о качественном и количественном составе органического мира определенных водоемов позволяет выяснить, какие виды и в каком количестве могут обитать в каждом из них, при каком сочетании видов кормовая база водоемов может быть всесторонне использована без ущерба для последующего восстановления ее и т. д. Эти вопросы специально изучаются в курсах по рыбоводству.
Дыхательная система.
Главными органами дыхательной системы являются жабры, состоящие из многих лепестков, прикрепленных проксимальными концами к жаберным дугам в отличие от жаберных пластин хрящевых рыб, прикрепленных одной стороной к межжаберным перегородкам. Следовательно, поверхность жабр костных рыб значительно больше, чем у хрящевых рыб. Более совершенен и механизм вдоха и выдоха. Довольно значительная часть газообмена (в среднем около 10%) совершается через кожу. В газообмене могут принимать участие плавательный пузырь и некоторые части кишечника.
Кровеносная система.
Сердце состоит из предсердия и желудочка, Круг кровообращения один. Артериального конуса у костистых рыб нет, и артериальный сосуд, отходящий от желудочка, начинается луковицей аорты. Количество эритроцитов значительно больше, чем у хрящевых рыб, что способствует усилению интенсивности процессов диссимиляции. Селезенка хорошо развита (рис. 85).
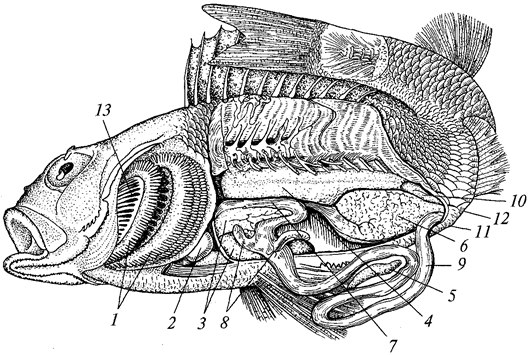
Рис. 85. Вскрытый окунь (самка):
/ — жабры; 2 —
сердце; 3 —
печень; 4 —
плавательный пузырь; 5 — селезенка; 6 —
яичник; 7 — желудок; 8 —
пилорические придатки; 9 —
кишка; 10 —
мочевой пузырь; 11 —
заднепроходное отверстие; 12 —
мочеполовое отверстие; 13 —
тычинки жаберной дуги
Выделительная система.
В выделительной системе особую роль играют туловищные почки, расположенные в виде двух длинных темно-красных узких лент вдоль позвоночного столба. Дистальные концы мочеточников (вольфовых каналов) соединяются и открываются наружу общим отверстием сзади анального отверстия. От дистальной части слившихся мочеточников отходит мочевой пузырь.
Размножение.
Половая система костистых рыб по сравнению с хрящевыми и древними костными рыбами упрощена, но способна производить большое число гамет. Два удлиненных семенника (их часто называют молоками) находятся под почками по бокам плавательного пузыря. Семя выходит не через вольфов канал, а через короткую трубку, образованную оболочкой семенника. Оба семяпровода открываются наружу общим половым отверстием сзади анального и мочевого отверстий или общим мочеполовым отверстием. Яичники (их обычно два) в период размножения представляют собой объемистые мешки, наполненные икрой, которая выходит наружу не через мюллеровы каналы (они редуцируются), а через короткие трубки, идущие от половых желез и открывающиеся наружу самостоятельным отверстием или в общее мочеполовое отверстие. Следовательно, созревшие яйцеклетки не попадают в полость тела, а быстро выходят из тела самок. Оплодотворение у подавляющего большинства видов костных рыб наружное. Внутреннее оплодотворение, а также живорождение свойственно сравнительно немногим представителям этого класса. Икра выметывается на водные растения и реже на другие подводные субстраты, а у меньшинства видов — в толщу воды. В связи с наружным оплодотворением масса гамет, зигот и мальков погибает. Поэтому плодовитость костных рыб очень велика. Например, карп производит более 1 млн икринок, щука — около 1 млн, палтус — 2—3,5 млн, треска — до 1 млн. Созревание половых желез зависит от разных факторов — внутренних и наружных. Ускоряющее воздействие на созревание половых желез оказывают гормоны гипофиза.
Оплодотворение икры можно осуществить в искусственных условиях, смешивая икру и семя рыб в присутствии небольшого количества воды. В настоящее время такая методика хорошо разработана и успешно применяется в широких масштабах на так называемых рыбоводных заводах. Оплодотворенная икра развивается в благоприятных условиях обычно до стадии мальков, способных добывать пищу в естественных водоемах, куда их выпускают. Такие мероприятия содействуют восстановлению ценных промысловых рыб, численность которых резко сократилась вследствие загрязнения водоемов, затруднения миграций рыб на места их размножения, их усиленной добычи.
На скорость роста рыб влияют различные условия жизни: питание, температура, состав растворенных в воде веществ и др. Рост в зависимости от изменений условий жизни идет неравномерно. Известно, например, что он замедляется в холодное время, что можно проследить на годовых кольцах чешуи рыб. Исследования роста рыб в разных водоемах имеют большое теоретическое и практическое значение, так как благодаря им можно выяснить, какие условия благоприятствуют ему, какие, наоборот, задерживают его, как восстанавливаются запасы промысловых рыб и т. д.
Продолжительность жизни рыб различна: одни живут менее года (некоторые виды хамсы и др.), другие — несколько лет (тихоокеанские лососевые — кета, горбуша и др.), третьи — несколько Десятков лет и даже около 100 лет (крупные осетровые, щука, сазан и др.).
Рыбообразные (бесчелюстные и рыбы) обладают приспособлениями (возникшими в течение длительной эволюции) для передвижения в воде, добывания пищи, спасения от хищников и т. п.
На основе этих приспособлений у них развились закономерные передвижения, характерные для разных периодов их жизни — миграции.
Самые короткие и частые из них — суточные
миграции. Зависят они от времени суток. Более длительные сезонные
миграции зависят от сезона года. К ним, в частности, относят зимовочные миграции, когда рыбы уходят на глубину и в малоподвижном состоянии, не питаясь, пережидают неблагоприятное для них время года. Например, хамса, которая питается и размножается в Азовском море, зимует в Черном море, так как вода в нем не подвергается такому сильному охлаждению, как в мелководном Азовском море.
Длительные, у ряда видов далекие миграции, называемые кормовыми,
совершают многие рыбы в места, где имеются в достаточном количестве организмы, которыми они питаются. Так, например, треска после окончания периода размножения в Атлантическом океане мигрирует в Баренцево море и некоторые другие северные моря.
Широко распространены у рыб нерестовые
миграции, обеспечивающие их размножение иногда в очень далеких местах от районов, где они долго живут, питаются и растут. К таким видам относятся проходные рыбы,
обитающие в морях, но размножающиеся в реках, впадающих в эти моря. Так, очень ценные осетровые рыбы (осетр, севрюга, белуга и др.) обитают в Каспийском, Азовском и Черном морях до наступления периода размножения, а для икрометания отправляются в верховья рек, впадающих в названные моря. После этого они возвращаются в моря, где живут до наступления следующего периода размножения. Мальки, развившиеся из отложенной и оплодотворенной в реках икры, сносятся течением в моря, где длительное время растут.
Такие рыбы размножаются несколько раз в течение жизни. Но есть рыбы, живущие несколько лет в океанах и морях, но размножающиеся только один раз в жизни. К ним относятся тихоокеанские лососи (кета, горбуша, чавыча и др.), которые для размножения идут в дальневосточные реки России, реки Северной Америки, Японии, Кореи, Китая, в которых происходит их нерест. Во время нерестовых миграций рыбы, как правило, не питаются и в результате погибают. В отличие от упомянутых рыб пресноводный угорь, обитающий в течение длительного времени (до 20 лет и более) в реках Европы, никогда там не размножается, а совершает далекие миграции (7000—8000 км) в Саргассово море, где нерестится и после этого погибает. Мальки угря подхватываются Гольфстримом и через 2 года попадают к берегам Европы и затем входят в реки.
Выделяют группу и полупроходных рыб,
живущих в опресненных частях морей и размножающихся в нижнем течении рек, впадающих в эти моря. К ним относятся популяции воблы, сазана, леща, обитающие на севере Каспийского моря.
Миграции известны и у многих беспозвоночных животных, обладающих достаточно эффективными способами передвижения. У позвоночных они наиболее сложны и продолжительны вследствие высокого развития их нервной, двигательной и других систем органов. Миграции осуществляются наследственно закрепленными инстинктами, развившимися в результате длительного естественного отбора. Благодаря миграциям животные всесторонне используют различные части своих ареалов. Исследования миграций имеют очень большое значение для охраны природы и целесообразного использования промысловых видов животных.
Практическое значение рыб.
Костные рыбы являются источниками белка и ряда других веществ, необходимых для полноценного питания людей. Население земного шара в течение XX в. и в особенности в последние десятилетия сильно увеличилось. В связи с этим добыча рыбы (а также водных моллюсков, ракообразных и других беспозвоночных) резко возросла. В дореволюционной России рыба добывалась в основном в реках, в других пресноводных водоемах и в южных морях страны.
В увеличении добычи рыбы значительную роль играет развитие прудового рыбоводства. В
прудах рыба питается личинками насекомых (в основном личинками хирономид), рачками, червями и т. д. Для того чтобы увеличить рыбопродуктивность прудов, рыб подкармливают жмыхами, бобовыми растениями и др. Чаще всего в прудах выращивают различные расы карпа. Совместно с карпом в прудах можно разводить карасей, линей и другие виды рыб, не являющихся конкурентами в питании. В правильно организованных прудовых хозяйствах существует несколько типов водоемов (для размножения, выращивания мальков, нагула товарной рыбы, зимовки и т. д.). В таких хозяйствах можно получать с 1 га 15—20 ц рыбы и более. Прудовое рыбоводство — одна из доходных отраслей животноводства.
Систематический обзор
Ныне существующие костные рыбы принадлежат к двум подклассам — лучеперым и лопастеперым рыбам.
Парные плавники опираются на костные образования — ради-алии,
расположенные лучеобразно (см. рис. 84). Дыхание жаберное, хоан нет. Плавательный пузырь — непарный, лежащий над Кишечником, возникший путем ответвления от него.
Эти рыбы были многочисленны в мезозойскую эру. Тела их были защищены костными чешуями, покрытыми тонким слоем дентиноподобного вещества ганоина.
Впоследствии, когда начался расцвет костистых рыб, подавляющее большинство их вымерло. Из четырех отрядов современных ганоидов здесь рассматривается один отряд, представители которого распространены в нашей стране.
Большинство видов крупных размеров: длина тела белуги до 4-5 м, русского и сибирского осетров — до 2-3 м (рис. 86) и т. д.; но стерлядь — небольшая рыба (длина тела — 40-60 см). Хвостовой плавник — гетероцеркальный. На теле пять рядов костных пластинок (жучек), образовавшихся из ганоидных чешуи. Череп хрящевой, но окружен покровными костями. Хорда хорошо развита. Тел позвонков нет. Сохранилось брызгальце. Имеются артериальный конус и спиральная складка в кишке. Таким образом, у этих рыб, как и у других ганоидов, сохранился ряд признаков хрящевых рыб.
Рис. 86. Сибирский осетр
Крупные осетровые — хищники, поедающие других рыб; остальные питаются мелкими беспозвоночными. Половозрелого состояния достигают поздно; самцы крупных видов в возрасте не ранее 10—12, самки — 12—15 лет; самцы стерляди — в 4—5 лет, самки — 7—8 лет. Большинство видов — проходные или полупроходные рыбы. Стерлядь обитает только в пресных водах, но совершает в реках довольно далекие нерестовые миграции. Всего известно 26 видов, из них 14 в России (главным образом в Каспийском и Азово-Черноморском бассейнах, в реках Сибири и Дальнего Востока). Издавна высоко ценились мясо и икра.
Современных лучеперых рыб, не входящих в над отряд ганоидных, объединяли ранее в группу костистых. Однако теперь эту группу не считают особым таксоном. Всех входящих в нее рыб распределяют по 8—10 отрядам. К общим признакам костистых рыб относятся следующие. Тело их покрыто костными чешуями, у некоторых видов вторично голое. Скелет в значительной степени или целиком костный. Остатки хорды сохраняются между телами позвонков. Вместо артериального конуса имеется луковица аорты. Спиральной складки в кишке нет, но у многих видов имеются пилорические отростки; кишечник удлинен.
В процессе эволюции костистых рыб происходили характерные, идущие в одном направлении изменения их организации: скелет постепенно окостеневал; в плавниках появились твердые заостренные лучи (у некоторых видов они превратились в колючки), брюшные плавники переместились вперед и расположились на уровне грудных плавников или впереди их; плавательный пузырь полностью отделился от передней части кишечника, и наполнение его смесью газов происходит только через кровеносную систему; совершился ряд изменений в других органах, которые способствовали более быстрому плаванию, защите от врагов и увеличили общую приспособленность рыб к обитанию в различных водоемах.
Эта огромная группа рыб разделяется на множество отрядов (30—40 и более). Здесь приводятся краткие сведения о некоторых из них в эволюционном порядке (т. е. от низших к высшим костистым рыбам).
Распространены во всех морях, чисто пресноводных мало; есть проходные виды. Многие имеют большое промысловое значение (разные виды сельдей, сардины, кильки, хамса и др.).
У многих видов на спине жировой плавник. Морские, проходные, некоторые чисто пресноводные. Промысловое значение очень велико (тихоокеанские лососи, семга, форель, нельма, омуль, хариус и др.). К этому отряду относится также обыкновенная щука, широко распространенная в пресных водах Северного полушария, которую добывают в некоторых районах в большом количестве. Она полезна как хищник, истребляющий малоценных сорных рыб.
Тело удлиненное, змееобразное, брюшных плавников нет. Подавляющее большинство видов обитает в морях, известен ряд проходных видов, в том числе европейский угорь, имеющий промысловое значение.
Пресноводные рыбы, число видов которых (свыше 2900) составляет 15 % всех видов класса. Многие имеют большое промысловое значение (сазан, лещ, карась, толстолобик, белый амур и др.). Выведены культурные породы сазана (разные породы карпа), карасей и др.
Обширный отряд, подавляющее большинство видов которого обитает в пресных водах, преимущественно в теплых. Тело, как правило, голое, около рта несколько пар усиков. Величина тела от гигантской (несколько метров) до карликовой (2 см). Многие сомообразные хищники. Широко известен европейский сом — хищник с широкой пастью, но мелкими зубами, длина тела которого иногда достигает 5 м, а масса — 90—300 кг.
Большинство этих рыб имеют два или три спинных плавника. Брюшные плавники расположены в передней части брюха под грудными. Подавляющее большинство видов обитает в северных морях. Ряд видов принадлежит к числу важных промысловых объектов — треска, пикша, сайда, а в последнее время особенно хек, минтай, путассу. Лишь один вид проник из северных морей в пресные воды — налим, особенно многочисленный в Сибири. В печени ряда тресковых рыб накапливается много витамина D.
Брюшные плавники находятся посредине брюха, два спинных плавника. Распространены преимущественно в морских тропических и умеренных водах, в нашей стране главным образом в Черном море. Имеют важное промысловое значение.
Самый большой отряд костистых рыб (свыше 6000 видов). Плавники с колючими лучами. Брюшные плавники находятся близко к переднему концу тела, под грудными плавниками или впереди их. Размеры тела от нескольких сантиметров до 4-5 м. Распространены на всем земном шаре в самых разнообразных водоемах. Ряд видов имеет большое промысловое значение: в пресноводных водоемах Северного полушария — судак, окунь и др., в морях — ставрида, скумбрия, тунец, нототения и др.
Хищные рыбы с уплощенным телом, лежащие во взрослом состоянии на дне на одном из боков. Брюшные плавники впереди грудных. Величина тела от 30—40 см до 5 м. Мальки имеют обычное рыбообразное строение тела, приспособленное к плаванию в толще воды. Ряд видов имеет важное промысловое значение.
(
SARCOPTERYGII)
Эти рыбы по ряду признаков (кости, прикрывающие череп;
жаберные крышки; костный скелет в парных плавниках; плавательный пузырь и др.) относятся к классу костных рыб. Но у них, как и у низших лучеперых рыб, сохранились примитивные признаки: развитая хорда, отсутствие тел позвонков, артериальный конус, спиральная складка в кишке и др. Сказанное свидетельствует о том, что лучеперые и лопастеперые рыбы произошли от древних костных рыб, живших в конце силура.
Однако пути эволюции обеих групп разошлись уже в начале девона. У лопастеперых рыб при сохранении указанных примитивных признаков сильно развились парные плавники с мощными опорными лопастями в их основании со скелетом из немногих подвижных удлиненных костей и появились парные плавательные пузыри, возникшие из задних жаберных мешков, лежащие на брюшной стороне. В пузыри поступал воздух через наружные ноздри и хоаны (внутренние ноздри). Эти древние лопастеперые рыбы обитали в небольших пресноводных водоемах и по разным причинам (высыхание воды, ухудшение ее качества, недостаток кислорода и кормовых объектов и т. п.) пытались выползать на берег, опираясь на плавники и используя для дыхания атмосферный воздух, и переходить в другие водоемы. Подкласс разделяется на два отряда: кистеперые рыбы и двоякодышащие рыбы.
Появились они в нижнем девоне. Обладали хорошо развитым опорным скелетом парных плавников. От них произошли в середине девона в результате длительного естественного отбора первые наземные позвоночные, дальнейшее приспособление которых к обитанию в прибрежных районах суши совершалось в каменноугольном периоде, и возникли примитивные земноводные — стегоцефалы. Последние широко распространились и начали вытеснять своих предков — кистеперых рыб. Одна из групп последних в триасе переселилась в море, где они обитали до конца мезозойской эры. В отложениях кайнозойской эры остатки их не встречались, и возникло мнение, что морские виды кистеперых давно вымерли. Однако в 1938 г. в Индийском океане у берегов Южной Африки была обнаружена кистеперая рыба, близкая к давно вымершим морским мезозойским видам, названная латимерией.
Впоследствии выяснилось, что эти рыбы постоянно обитают в районе Коморских островов (недалеко от Мадагаскара), где они изредка встречаются на довольно большой глубине. Длина их достигает 1— 1,8 м; чешуя крупная, массивная; парные плавники — мощные, массивные, очень подвижные; плавательный пузырь, т. е. бывшее легкое, для дыхания не приспособлен; хоан нет. Латимерии — хищные животные, питающиеся другими рыбами.
Эти рыбы произошли от кистеперых рыб и были широко распространены с девона до пермского периода, позднее становились все более редкими. В настоящее время существует только шесть видов: один австралийский, четыре африканских и один южноамериканский. За исключением одного африканского вида, это крупные животные, достигающие в длину 1,7—2 м. Они всеядные и даже хищные животные. Зубы у них срослись и превратились в массивные пластинки, способные дробить толстые раковины моллюсков. У австралийского вида одно легкое, у остальных — пара легких. Кровь после окисления в легких идет в сердце, в котором появилась неполная перегородка, разделяющая предсердие на две части — правую и левую, как у земноводных. Они обитают в заболоченных, пересыхающих водоемах и могут, окруженные грязевой камерой, довольно долго переносить отсутствие воды, используя для дыхания атмосферный воздух. Древние двоякодышащие рыбы, судя по их парным плавникам, могли переползать в другие водоемы. Но, вероятно, потомки кистеперых рыб оказались более приспособленными к освоению пограничных биотопов между водоемами и наземной средой и уже в девоне дали начало первым земноводным, с которыми двоякодышащие рыбы соревноваться не могли. Поэтому выжили те из них, которые могли длительное время переносить отсутствие воды в покоящемся состоянии.
Четвероногие — это позвоночные, перешедшие к жизни на суше, в воздушной среде. Некоторые из них связаны с водой в отдельные периоды жизни или вторично перешли к жизни в воде (китообразные), сохранив легочное дыхание. Передвижение по суше осуществляется с помощью парных членистых конечностей.
У взрослых дыхание легочное. Личинки земноводных дышат жабрами, у остальных классов жаберные щели закладываются на ранних стадиях эмбрионального развития, но вскоре зарастают. Имеются два круга кровообращения, однако полностью они обособляются лишь у птиц и млекопитающих. Увеличивается головной мозг и дифференцируются его отделы. Органы боковой линии исчезают, а другие органы чувств приспосабливаются к деятельности в воздушной среде.
Надкласс подразделяется на 4 класса: земноводные, или амфибии, пресмыкающиеся, или рептилии, птицы и млекопитающие. Последние 3 класса образуют группу амниот. Это — наземные животные. Оплодотворение у них внутреннее, а яйцо (у яйцекладущих форм) развивается только в воздушной среде. Личиночная стадия отсутствует, метаморфоза нет.
Особенности амниот заключаются как в характере эмбрионального развития, так и в строении взрослых особей. Яйца их содержат большое количество желтка, поэтому дробятся они частично, дискоидально. При развитии образуются зародышевые оболочки — наружная серозная и внутренняя амниотическая. В результате зародыш оказывается погруженным в амниотическую полость, заполненную жидкостью. Эта жидкость предохраняет его от высыхания и механических повреждений. Дыхание и удаление продуктов обмена у зародыша осуществляются при помощи специального зародышевого органа — аллантоиса, выполняющего функции мочевого пузыря.
У взрослых амниот кожа малопроницаема для газов и воды благодаря ее верхнему слою. Это предохраняет организм от высыхания. Скелет полностью окостеневает, а пояса конечностей усиливаются. Увеличивается поверхность легких, формируются мета-нефрические, или тазовые, почки, резко вырастают размеры мозга. Все больше обособляются большой и малый круги кровообращения.
Амфибии, как и рыбы, относятся к анамниям, и яйца их развиваются в воде. Личинка их водная и лишь в результате метаморфоза достигает взрослого состояния.
Земноводные — первые наземные позвоночные, еще сохраняющие связь с водой. Развитие у большинства происходит в воде. Взрослые амфибии имеют парные конечности с шарнирными суставами. Череп аутостиличный (нёбноквадратный хрящ срастается с мозговой коробкой), с шейным позвонком соединяется за счет образования двух затылочных мыщелков. Тазовый пояс причленя-ется к поперечным отросткам крестцового позвонка. Сердце со-
стоит из двух предсердий и желудочка. Значительно развит передний мозг, который делится на два полушария. В его крыше имеются нервные клетки. Глаза с подвижными веками. Голая кожа с большим количеством слизистых желез. Почки мезонефрические (туловищные). Температура тела непостоянная и зависит от температуры окружающей среды (пойкилотермия).
Класс включает 3 современных отряда: бесхвостые, хвостатые и безногие (рис. 87).

Рис. 87. Филогенетическое древо земноводных
Общая характеристика
Тело взрослых земноводных состоит из головы, туловища и двух пар ног. Хвост был у древних амфибий — стегоцефалов,
почти постоянно обитавших в воде, и сохранился у сравнительно небольшого числа видов, выделенных в отряд хвостатых (тритоны, саламандры), тоже проводящих в водоемах большую часть своей жизни во взрослом состоянии. У подавляющего большинства современных земноводных, выделяемых в отряд бесхвостых (лягушки, жабы и многие другие), хвост редуцировался в связи с тем, что они перешли к передвижению скачками с помощью задних ног. У немногих видов, ведущих полуподземный образ жизни (отряд безногие, или червяги), редуцировались ноги и хвост (рис. 88).

Рис. 88. Амфибии:
1 — кольчатая червяга (Siphonops); 2 —
тритон (Triturusvulgaris); 3 —
личинка амблистомы аксолотль; 4—
амблистома (Amblystoma); 5— пятнистая саламандра (Salamandrasalamandra); 6 —
протей (Proteus)
Тело у амфибий, как правило, сплющено в дорзовентральном направлении. Голова подвижно соединена с туловищем, что способствует лучшей ориентации взрослых земноводных в воздушной среде, особенно при лове пищевых объектов (главным образом летающих насекомых). Этим они отличаются от рыб, у которых тело сдавлено с боков и голова неподвижно соединена с туловищем, что необходимо при плавании.
Обычная длина тела амфибий несколько сантиметров, некоторые виды значительно крупнее, но в среднем представители этого класса имеют меньшую величину тела, чем остальные наземные позвоночные.
Покровы.
Кожа голая, костная чешуя была только у стегоцефалов. Как известно, масса тела при переходе из водной среды в воздушную возрастает (закон Архимеда). Поэтому редукция костной чешуи имела большое значение для приспособления первых наземных позвоночных к жизни на суше. В связи с редукцией чешуи и необходимостью защиты тела от механических повреждений поверхностные клетки эпидермиса претерпевают умеренное ороговение оболочек. Кожа весьма богата железами, выделения которых усиливают защиту от высыхания и у ряда земноводных содержат ядовитые вещества.
Окраска тела разнообразна и для каждого вида специфична. В большинстве случаев она имеет защитное значение, так как благодаря ей эти животные малозаметны в местах их обитания.
Кожа земноводных отделена от лежащей под ней мускулатуры обширными полостями с водянистой жидкостью, которых нет ни у рыб, ни у вышестоящих позвоночных — пресмыкающихся, птиц и млекопитающих, вполне приспособленных к жизни на суше. Эти полости, во-первых, уменьшают опасность высыхания тела, во-вторых, выполняют роль амортизаторов механических воздействий на поверхность тела, в-третьих, облегчают кожный газообмен, играющий у земноводных важную роль.
Нервная система. В
связи с более сложными и изменчивыми условиями жизни и коренным изменением характера движения взрослых, особенно на суше, нервная система амфибий довольно сильно отличается от аналогичной системы костных рыб (рис. 89).
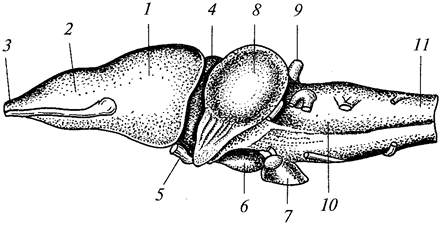
Рис. 89. Головной мозг лягушки, вид сбоку:
1 —
полушария переднего мозга; 2 —
обонятельная доля; 3 —
обонятельный нерв; 4 —
промежуточный мозг; 5 —
зрительная хиазма; 6 —
воронка; 7 — гипофиз; 8 —
средний мозг; 9 —
мозжечок; 10 —
продолговатый мозг; 11
~
спинной мозг
Передний мозг всегда крупнее среднего мозга и явственно разделяется на два полушария; в глубинных слоях его крыши появляются нервные клетки, которые можно рассматривать как слабые зачатки будущей коры головного мозга. Роль его в координации жизнедеятельности всего организма благодаря усилению его связей с другими отделами центральной нервной системы несколько возрастает. Мозжечок развит слабее, чем у рыб, так как во время движения положение тела земноводных вследствие опоры на ноги более устойчиво, чем у рыб. Участки спинного мозга в местах отхождения от него нервов к мышцам ног, которые выполняют значительно большую роль, чем мышцы парных плавников, заметно утолщены. Периферическая нервная система в связи с появлением длинных расчлененных ног, прогрессирующей дифференциацией мышечной системы и другими приспособлениями к наземной жизни подверглась существенным изменениям.
Органы чувств.
Из органов чувств наиболее существенным изменениям подвергся орган слуха. Передача звуковых волн из водной среды в ткани животного, которые тоже насыщены водой и имеют приблизительно такие же акустические свойства, как и вода, происходит значительно лучше, чем из воздуха. Звуковые волны, распространяющиеся в воздушной среде, почти целиком отражаются от поверхности животного, и только около 1 % энергии этих волн проникает в его тело. В связи с этим у земноводных в добавление к лабиринту, или внутреннему уху, возник новый отдел органа слуха — среднее ухо.
Оно представляет собой наполненную воздухом маленькую полость, сообщающуюся с ротовой полостью посредством евстахиевой трубы и закрытую снаружи тонкой, упругой барабанной перепонкой. В среднем ухе находится слуховая косточка (столбик), которая одним концом упирается в барабанную перепонку, а другим — в окошечко, затянутое пленкой и ведущее в полость черепа, где находится лабиринт, окруженный перилимфой. Давление внутри среднего уха равно атмосферному, и барабанная перепонка может вибрировать под влиянием звуковых воздушных волн, воздействие которых через слуховую косточку и перилимфу передается стенкам лабиринта и воспринимается окончаниями слухового нерва. Полость среднего уха образовалась из первой жаберной щели, а столбик — из расположенной около щели подъязычно-нижнечелюстной кости (ги-омандибуляре), которая подвешивала у рыб висцеральный отдел черепа к мозговому отделу там, где за ушными костями находился лабиринт.
Глаза прикрыты подвижными веками, которые защищают органы зрения от высыхания и засорения. Благодаря изменению формы роговицы и хрусталика земноводные видят дальше, чем рыбы. Амфибии хорошо воспринимают небольшие температурные изменения. Они чувствительны к воздействиям различных веществ, растворенных в воде. Их орган обоняния реагирует на раздражения, вызываемые газообразными веществами. Таким образом, органы чувств земноводных претерпели ряд изменений в связи с переходом к обитанию на суше. У личинок и взрослых животных, обитающих постоянно в воде, имеются, как и у рыб, сейсмосенсорные органы.
Земноводным свойственно довольно сложное инстинктивное поведение, особенно в период размножения. Например, самец жабы-повитухи, обитающей на западе Украины, наматывает на свои задние конечности шнуры икры и до развития головастиков прячется в укромных местах. Через 17—18 дней самец возвращается в воду, где происходит вылупление головастиков. Это своеобразный инстинкт защиты потомства. У ряда тропических бесхвостых амфибий известны еще более сложные инстинкты. Земноводным свойственны и условные рефлексы, однако они вырабатываются с большим трудом.
Скелет.
В связи с приспособлением амфибий к наземному образу жизни скелет сильно изменился во всех своих частях (рис. 90).
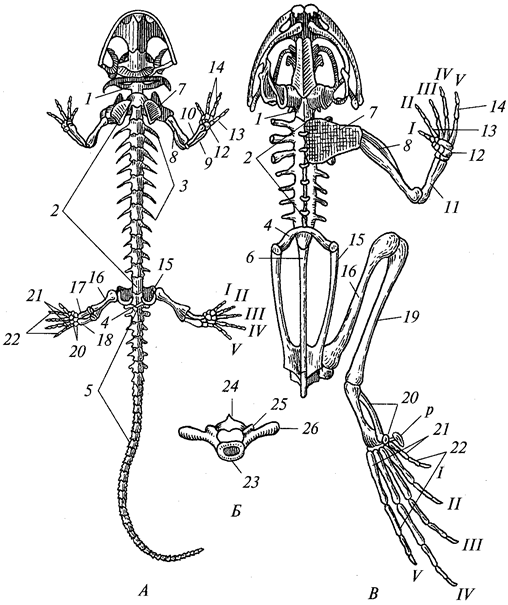
Рис. 90. Скелет саламандры (А)
и лягушки (В),
Б —
туловищный позвонок лягушки:
1 —
шейный позвонок; 2 —
туловищные позвонки; 3 —
ребра; 4 —
крестцовый позвонок; 5 — хвостовые позвонки; 6 —
уростиль; 7 — пояс передних конечностей; 8 —
плечо; 9 —
локтевая кость; 10 —
лучевая кость; 11
— сросшиеся лучевая и локтевая кости; 12 —
запястье; 13 —
пясть; 14 —
фаланги пальцев; 15
— подвздошные кости тазового пояса; 16 —
бедро; 17 —
большая берцовая кость; 18 —
малая берцовая кость; 19
— сросшиеся малая и большая берцовые кости; 20 —
предплюсна; 21 —
плюсна; 22 —
фаланги пальцев; 23 —
тело позвонка; 24 —
верхняя дуга с остистым отростком; 25 —
сочленовный отросток; 26 —
поперечный отросток; /— V —
нумерация пальцев, р —
рудимент предпальца
Череп амфибий имеет много хрящевых элементов, что, вероятно, объясняется необходимостью облегчить массу тела в связи с полуназемным образом жизни. В составе черепа много костей, перечисленных при описании черепа высших рыб, и в том числе характерный только для рыб и земноводных парасфеноид. Так как гиомандибуляре стала слуховой косточкой, то роль подвеска нижней челюсти выполняет квадратная кость. В связи с утратой во взрослом состоянии жаберного аппарата жаберные дуги редуцируются и сохраняются лишь их видоизмененные остатки. Подъязычная дуга сильно изменяется и частично редуцируется. Череп амфибий очень широкий, что отчасти связано с особенностями их дыхания. Нижняя челюсть, как и у костных рыб, состоит из нескольких костей.
Позвоночный столб у бесхвостых очень короткий и заканчивается длинной косточкой — уростилем,
образовавшимся из рудиментов хвостовых позвонков. У хвостатых земноводных этот отдел позвоночного столба состоит из ряда позвонков. У этих амфибий хвост играет существенную роль при передвижении: в воде он служит для плавания, на суше — для поддержания равновесия. Ребра слабо развиты (у хвостатых земноводных) или редуцированы, а их остатки слились с поперечными отростками позвонков (у остальных амфибий). У древних амфибий ребра имелись. Редукция их у современных форм объясняется необходимостью облегчить массу тела (сильно возросшую при переходе из водной среды в воздушную). Вследствие редукции ребер у земноводных нет грудной клетки. Первый позвонок устроен иначе, чем у рыб: он имеет две суставные впадины для сочленения с двумя затылочными мыщелками черепа, благодаря чему голова земноводных стала подвижной.
Скелет передней конечности состоит из плечевой кости, двух костей предплечья — лучевой и локтевой, костей запястья, пястных костей и фаланг пальцев. Скелет задней конечности состоит из бедра, двух костей голени — большой берцовой и малой берцовой, костей предплюсны, плюсневых костей и фаланг пальцев. Следовательно, сходство в строении обеих пар конечностей, несмотря на некоторые различия в их функциях, очень велико. Первоначально передние и задние ноги были пятипалые, у современных земноводных число пальцев может быть меньшим. Задние конечности у многих бесхвостых амфибий служат и для плавания, в связи с чем они удлинились, а пальцы соединились плавательными перепонками.
Пояса конечностей развиты значительно лучше, чем у рыб. Плечевой пояс состоит из костных и хрящевых элементов: лопатки надлопаточного хряща, ключицы, прокоракоидного хряща и коракоида, или вороньей кости. Ключицы и коракоиды соединены с грудиной, в состав которой тоже входят костные и хрящевые элементы. Головка плеча сочленяется с плечевым поясом. Задний пояс конечностей, или таз, состоит из трех костей: подвздошной, лобковой и седалищной. Образуемая этими костями большая вертлужная впадина служит для сочленения с головкой бедра. Таз соединен с одним позвонком — крестцовым, благодаря чему задние ноги в отличие от брюшных плавников рыб получили довольно прочную опору.
Мышечная система.
В связи с развитием у амфибий ног, состоящих из многих костей, усложнением поясов конечностей, возникновением подвижного соединения головы с туловищем, большим, чем у рыб, разделением на отделы позвоночного столба, изменениями способов дыхания, захвата пищи и другими приспособлениями к жизни на суше мышечная система подверглась значительной перестройке и стала намного разнообразнее по сравнению с той же системой рыб. У последних главную массу составляло множество сходных мышечных сегментов. У амфибии появились разные группы мышц: мышцы ног (например, бедренные, икроножная, малоберцовая, предплюсневые, сгибатели и разгибатели), мышцы, соединяющие ноги с поясами конечностей (например, грудная, дельтовидная, ягодичная и др.), мышцы, изменяющие объем ротовой полости (подчелюстная, подъязычная и др.), а также многие другие. Таким образом, у земноводных были заложены основы той мышечной системы, которая потом развивалась, усложнялась и подвергалась дальнейшей дифференциации у вышестоящих позвоночных — рептилий, птиц и млекопитающих. Однако у земноводных как низших наземных позвоночных сохранились следы метамерной мускулатуры, в частности довольно хорошо выраженные на брюшной стороне тела.
Пищеварительная система.
Ротовая полость очень широкая. Уряда видов (главным образом у хвостатых амфибии) имеется много мелких, однородных, примитивно устроенных зубов, которые сидят на челюстях, сошнике, нёбных и других костях и служат лишь для удержания добычи. У большинства видов (главным образом у бесхвостых амфибий) зубы частично или полностью редуцируются, зато у них сильно развивается язык. Последний у лягушек прикреплен к нижней челюсти передним концом и может для поимки добычи далеко выбрасываться задним концом вперед. Он очень липкий и хорошо приспособлен для выполнения указанной функции. У видов, постоянно живущих в воде, язык обычно редуцируется. Захват добычи у таких земноводных осуществляется челюстями.
Пищеварительная трубка сравнительно короткая и состоит из глотки, пищевода, желудка, тонкой кишки и совсем небольшой прямой (толстой) кишки. Задняя часть прямой кишки представляет собой клоаку; через нее выводятся кроме экскрементов моча и половые продукты. В ротовую полость впадают слюнные железы, отсутствующие у рыб. Секрет этих желез служит главным образом для смачивания пищи. Слюнные железы очень слабо развиты у видов, обитающих в воде, и значительно лучше — у наземных. Печень велика. Поджелудочная железа хорошо выражена. Пища взрослых земноводных в основном животная (насекомые, мелкие позвоночные и др.). Головастики же бесхвостых амфибий большей частью растительноядны.
Дыхательная система.
Почти у всех земноводных есть легкие (рис. 91). Эти органы имеют еще очень простое строение и представляют собой тонкостенные мешки, в стенках которых разветвляется довольно густая сеть кровеносных сосудов.
Рис. 91. Вскрытая самка лягушки:
1 —
сердце; 2 —
легкое; 3 —
левая лопасть печени; 4 —
правая лопасть печени; 5 — желчный пузырь в центральной лопасти печени; 6 —
желудок; 7 — поджелудочная железа; 8 -
двенадцатиперстная кишка; 9 -
тонкая кишка; 10 -
прямая кишка- 11 -
селезенка; 12 -
клоака (вскрыта); 13 -
мочевой пузырь; 14 -
отверстие мочевого пузыря; /5 - почка; 16 -
мочеточник: 17 -
парные отверстия мочеточников в клоаке; 18 -
правый яичник (левый удален); 19 -
жировое тело; 20 —
правый яйцевод; 21 —
левый яйцевод; 22 —
маточный отдел яйцевода; 23 —
отверстие яйцевода в клоаке; 24 —
спинная аорта; 25 —
задняя полая вена; 26 —
общая сонная артерия; 27 -
левая дуга аорты; 28 -
кожно-легочная артерия
Так как внутренняя стенка легких почти гладкая, то их поверхность относительно невелика. Трахея почти неразвита, и легкие связаны непосредственно с гортанью. Поскольку грудной клетки у земноводных нет, дыхание обеспечивается работой мышц ротовой полости. Вдох происходитследующим образом. При открытых ноздрях (которые в отличие от ноздрей рыб сквозные, т. е. кроме наружных ноздрей имеются и внутренние ноздри — хоаны) и закрытом рте оттягивается дно обширной ротовой полости и в нее поступает воздух. Затем ноздри закрываются особыми клапанами, дно ротовой полости поднимается, и воздух нагнетается в легкие. Выдох происходит в результате сокращения брюшной мускулатуры.
Значительное количество кислорода земноводные получают через кожу и слизистые оболочки ротовой полости. У некоторых видов саламандр легких совсем нет, и весь газообмен совершается через кожу. Однако кожа только в том случае может выполнять функции дыхания, если она влажная. Поэтому обитание земноводных в условиях недостаточной для них влажности воздуха невозможно. Личинки, живущие в воде, дышат жабрами (сначала — наружными, потом — внутренними) и кожей. У некоторых хвостатых земноводных, постоянно живущих в воде, жабры сохраняются всю жизнь. Таким образом, по способам дыхания амфибии еще близки к рыбам.
Кровеносная система.
Она тоже сильно изменилась по сравнению с той же системой рыб в связи с заменой жаберного дыхания на легочное, развитием из парных плавников расчлененных ног, преобразованием мышечной и других систем, вызванных приспособлением амфибий к наземному образу жизни (рис. 92).
Рис. 92. Кровеносная система лягушки:
А —
схема артериальной системы лягушки: 1 —
наружная сонная артерия; 2 —
внутренняя сонная артерия; 3 —
общая сонная артерия; 4 —
легочно-кожная артерия; 5 — артериальный конус; 6 —
правое предсердие; 7 — левое предсердие; 8 —
желудочек; 9 —
легочная артерия; 10 —
легкое; 11 —
кожная артерия; 12 —
печеночные артерии; 13 —
желудочная артерия; 14 —
кишечная артерия; 15 —
спинная аорта; 16 —
почечная артерия; 17—
почка; 18 —
левый семенник (правый удален); 19 —
подвздошная артерия; Б —
схема венозной системы лягушки: 1 —
бедренная вена; 2 —
седалищная вена; 3 —
подвздошная вена; 4 —
брюшная вена; 5 — воротная вена печени; 6 —
печеночная вена; 7 — левый яичник; 8 —
почка; 9 —
задняя полая вена; 10 —
большая кожная вена; 11 —
подключичная вена; 12 —
внутренняя яремная вена; 13 —
наружная яремная вена; 14 —
передняя полая вена; 15 —
легкое
Сердце намного крупнее и сложнее устроено, чем у рыб, и состоит из трех камер левого предсердия, правого предсердия и одного желудочка. В левое предсердие поступает по легочным венам кровь, насыщенная в легких кислородом и бедная углекислым газом. Такую кровь принято называть артериальной,
хотя ее приносят здесь вены. В правое предсердие поступает по венам кровь, отдавшая разным органам большую часть кислорода и насыщенная углекислым газом. Такую кровь принято называть венозной.
В результате сокращения предсердий кровь из них попадает в единственный желудочек, но там артериальная и венозная порции ее полностью не смешиваются, так как этому препятствуют многочисленные выросты на внутренней стороне желудочка. Поэтому после сокращения предсердий в правой части желудочка будет находиться венозная кровь, в левой — артериальная, а в средней — смешанная.
От правой части желудочка отходит артериальный конус (унаследованный амфибиями от кистеперых рыб), от которого берут начало четыре пары артерий (образовавшиеся из жаберных приносящих артерий рыб). Первая пара (самая передняя) — сонные артерии, несущие кровь к голове, вторая и третья пары — артерии, соединяющиеся в самый большой сосуд тела — аорту, разветвления которой направляются в разные части тела, четвертая пара — легочно-кожные артерии, которые потом разделяются на самостоятельные легочные артерии, несущие кровь в легкие, и кожные артерии, несущие кровь в кожу. В результате сокращения желудочка первая порция крови, попадающая в артериальный конус, будет венозной, и она направляется в самые близкие к правой части желудочка и широкие легочно-кожные артерии, вторая порция крови — смешанная — направ-
ляется в аорту, третья порция крови — артериальная — идет в сонные артерии. Следовательно, в отличие от жаберных приносящих артерий рыб, которые несут только венозную кровь, перечисленные главные артерии амфибий несут разную кровь, что обеспечивает наиболее целесообразное распределение ее в организме в связи с переходом к легочному способу дыхания. Кровь, насыщенная в коже кислородом, идет по венам в правое предсердие и несколько увеличивает содержание кислорода в венозной крови.
Таким образом, у амфибий в отличие от бесчелюстных и рыб не один, а два круга кровообращения: большой — от сердца по артериям и капиллярам во все части тела и из них по капиллярам и венам в правое предсердие, малый — от сердца по легочным артериям и капиллярам в легкие и из них по капиллярам и легочным венам в левое предсердие. Недостаток этой системы заключается в том, что большая часть тела снабжается смешанной, с пониженным содержанием кислорода кровью, не обеспечивающей высокую интенсивность обмена веществ.
В связи с развитием расчлененных ног, дифференциацией мышечной системы и другими изменениями тела у земноводных существенно изменилась сеть кровеносных сосудов.
Выделительная система.
Почки, как и у рыб, туловищные, но более компактной формы, расположенные в задней половине тела на спинной стороне. Моча удаляется через мочеточники (вольфовы каналы), впадающие, как и у кистеперых рыб, в клоаку, и накапливается в мочевом пузыре, который периодически опорожняется через клоакальное отверстие. Так как у амфибий кожа голая и они обитают во влажной или водной среде, то в их тело через кожные покровы может проникать значительное количество воды, избыток которой удаляется через почки.
Размножение.
Амфибии — раздельнополые животные. У самок половые железы представлены парными яичниками, а у самцов — парными семенниками. Оплодотворение в большинстве случаев происходит в воде. У бесхвостых амфибий этому предшествует сближение самца и самки: самец обхватывает сзади самку, надавливает передними конечностями на ее брюшную стенку и этим способствует выходу в воду икры, которую он сразу оплодотворяет. Таким образом, при наличии полового акта оплодотворение происходит вне организма самки. У большинства хвостатых амфибий (например, у тритонов) самец выделяет семя в особом мешочке (сперматофоре), который самка тут же захватывает краями клоаки. В данном случае полового акта нет, но оплодотворение — внутреннее. Наконец, у безногих амфибий самец вводит семя в клоаку самки при помощи своей способной выпячиваться клоаки.
У многих видов хорошо выражен половой диморфизм (в окраске, строении передних ног, которыми самцы удерживают самок, и по другим признакам). Самцы ряда видов могут издавать очень громкие звуки благодаря усилению этих звуков голосовыми мешочками — резонаторами.
Развитие.
Как правило, развитие происходит в воде. Из оплодотворенных яиц развиваются личинки (головастики), имеющие рыбообразную форму. Дышат они жабрами, и их внутреннее строение напоминает таковое рыб. По мере роста происходит превращение головастиков: сначала у них вырастают задние ноги, потом передние, жабры атрофируются, развиваются легкие, большие изменения происходят в кровеносной системе и т. д.
Практическое значение.
Многие амфибии полезны тем, что уничтожают большое количество насекомых — вредителей сельскохозяйственных растений, переносчиков, промежуточных и окончательных хозяев паразитов, вызывающих заболевания домашних животных и людей. Во многих биоценозах амфибии размножаются в большом количестве и играют существенную роль в круговороте веществ. Масса земноводных используются в разнообразных научных исследованиях и в учебных целях. В ряде стран амфибий (главным образом лягушек) употребляют в пищу. Вред же, причиняемый этими животными (в частности, в рыбоводном прудовом хозяйстве), сравнительно невелик. Между тем чрезмерный отлов этих позвоночных, уничтожение биотопов, где они водятся, широкое непродуманное использование ядовитых веществ для борьбы с разными вредными организмами губительно отражается на популяциях амфибий. Поэтому проведение разных мероприятий по их охране весьма необходимо.
Систематический обзор
Современные земноводные принадлежат к трем отрядам: хвостатые (около 300 видов), безногие (около 60 видов), бесхвостые (около 2000 видов). Среди наземных позвоночных класс амфибий занимает последнее место по количеству видов. В нашей фауне их около 30 видов.
Сравнительно небольшая группа животных, к которой относятся саламандры, тритоны, протеи (имеют жабры, живут в пещерных водоемах и лишены глаз), сирены (с жабрами и лишенные задних ног) и др. Большинство хвостатых амфибий распространено в Северной Америке. По внешнему облику, внутреннему строению и образу жизни они ближе к рыбам, чем другие земноводные. Большую часть жизни хвостатые амфибии проводят в воде, некоторые сохраняют жаберное дыхание; легкие очень примитивны. Есть основание предполагать, что водный образ жизни у этих амфибий — вторичный. Возможно, что после того как земноводные дали начало сухопутным позвоночным (пресмыкающимся), амфибиям стало трудно конкурировать со своими потомками. Нередко их жизнь была в меньшей опасности в мелких водоемах, где пресмыкающиеся жить не могут и почти не встречаются рыбы. Поэтому в процессе эволюции чаще выживали те амфибии, которые не заканчивали метаморфоза и оставались жить в воде. Явление выпадения взрослой стадии и достижения личинкой полово-зрелости называется неотенией.
Неотения имеет место и у животных других групп.
Маленькая группа червеобразных животных (червяги), сильно изменившихся в связи с роющим образом жизни и в то же время сохранивших ряд примитивных признаков. Оплодотворение внутреннее. Развитие личинок заканчивается в воде. Обитают преимущественно в Южной Америке, а также встречаются в тропической Африке и Южной Азии.
Самая многочисленная группа современных земноводных, хорошо приспособленных к полуназемному образу жизни. К этому отряду относятся лягушки, жабы, квакши, чесночницы, жерлянки и др. Особенно многочисленны в тропических странах. Многие виды живут на деревьях. Некоторые бесхвостые (например, жабы) лучше переносят условия наземной жизни и дальше уходят от водоемов, чем большинство видов этой группы. У таких земноводных кожа грубее, лучше развиты легкие и т. д. Однако и они обитают только во влажных местах.
Форма тела рептилий ящерообразная, змеевидная или плоская и широкая (черепахи). Кожные покровы сверху ороговевают, вследствие чего образуются щитки, чешуи, шиты и когти. Для воды и газов кожа непроницаема и в связи с этим почти лишена желез.
В позвоночнике формируется шейный отдел, обеспечивающий подвижность головы. Череп имеет затылочный мыщелок, височные ямы и ограничивающие их височные дуги. Конечности типичны для наземных позвоночных. Дыхание только легочное, причем есть трахея и бронхи. Сердце трехкамерное. Почки мета-нефрические (тазовые). Головной мозг увеличен за счет полушарий и мозжечка. Обитают в различных биотопах, главным образом в теплых и умеренных широтах. Температура тела непостоянная.
Современные пресмыкающиеся являются остатками господствовавших в прошлом классов. Ныне живущих отрядов — 4, ископаемых— 13 (рис. 93).
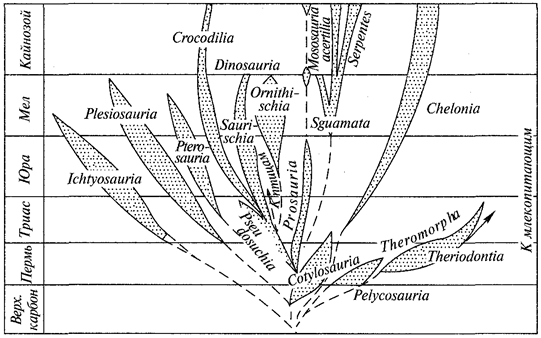
Рис. 93. Филогенетическое древо пресмыкающихся
Пресмыкающиеся произошли во второй половине каменноугольного периода от более сложных древних земноводных, близких к стегоцефалам. В процессе последующей длительной эволюции они стали настоящими наземными животными, приспособленными к более активному, чем амфибии, образу жизни в различных частях суши, часто в очень засушливых условиях, к размножению и эмбриональному развитию вне водоемов. К современным рептилиям относятся ящерицы, змеи, крокодилы и черепахи (рис. 94).

Рис. 94. Ящерицы:
1
— степная агама; 2 —
ушастая круглоголовка; 3 —
желтопузик; 4 —
степной геккон; 5 — серый варан; 6 —
хамелеон
Общая характеристика
Благодаря появлению шеи у рептилий голова подвижно сочленена с туловищем и может совершать движения не только, как у амфибий, в вертикальном направлении, но и в горизонтальном.
Туловище удлиненное (только у черепах широкое), довольно гибкое; хвост у подавляющего большинства видов длинный, способный к разнообразным движениям; ноги сильно развиты (у некоторых видов ящериц и у всех змей ноги редуцированы в связи с переходом к другому активному способу передвижения). Величина тела в среднем значительно больше, чем у амфибий, а некоторые виды достигают огромных размеров.
Покровы.
Кожа покрыта роговыми прочными и в то же время легкими чешуями эктодермального происхождения, которые защищают тело не только от механических повреждений, но и от высыхания. Ороговевший слой кожи периодически сбрасывается во время линьки. У черепах и крокодилов в мезенхимном слое кожи развиваются окостенения, у остальных рептилий они отсутствуют. Черепахи хорошо защищены от врагов панцирями. Кожа современных рептилий в отличие от земноводных сухая, лишенная желез.
Окраска наружных покровов разнообразна и у многих видов имеет защитное значение, так как благодаря ей животные малозаметны на фоне окружающей среды.
Нервная система.
В связи с более подвижным и сложным образом жизни нервная система рептилий развита лучше, чем у амфибий (рис. 95).
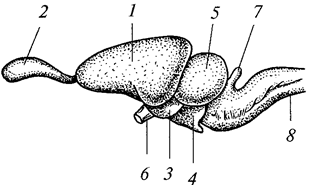
Рис. 95. Головной мозг ящерицы, вид сбоку:
1 —
полушарие переднего мозга; 2
— обонятельная доля; 3 —
промежуточный мозг; 4 —
гипофиз; 5 —
средний мозг; 6 —
перекрест зрительных нервов (хиазма); 7 — мозжечок; 8 —
спинной мозг
Передний мозг значительно больших размеров, чем У последних (в особенности за счет увеличения полосатых тел), на поверхности полушарий появляются нервные клетки — зачаток коры головного мозга. Поэтому у рептилий быстрее устанавлива-ются новые связи с внешней средой, чем у их предков. Возрастает роль этого отдела головного мозга как координатора жизнедеятельности всего организма. Усложняется строение промежуточного и среднего мозга, усиливается их связь с передним мозгом и другими отделами центральной нервной системы. Мозжечок в связи с усложнением способов передвижения и необходимостью сохранять равновесие тела развит заметно лучше, чем у земноводных. Увеличился и продолговатый мозг, который образует характерный для рептилий вертикальный изгиб. Спинной мозг длинный; строение его усложнилось. Это выразилось в более резком отделении наружного слоя (белого, т. е. проводящих путей) от внутреннего (серого, т. е. нервных клеток) слоя.
Из органов чувств у рептилий (за исключением змей и некоторых других видов) лучше, чем у амфибий, развиты глаза: они более подвижны. Приспособление их к видению на разных расстояниях (у многих видов на довольно больших) обеспечивается не только, как у рыб и амфибий, перемещением хрусталика, но и, как у высших позвоночных, путем изменения его кривизны. Кроме век имеется мигательная перепонка. У ряда видов есть теменной орган, связанный с промежуточным мозгом и способный к восприятию световых раздражении. Некоторые прогрессивные изменения произошли в органе слуха: немного увеличился нижний мешочек; слуховая косточка лучше передает колебания барабанной перепонки во внутреннее ухо; кроме овального окошечка появилось второе окошечко — круглое, что увеличивает подвижность экзолимфы, окружающей лабиринт; барабанная перепонка расположена не на поверхности головы, а в небольшом углублении. Органы обоняния развиты довольно хорошо, внутренняя поверхность их увеличилась. Органы вкуса у большинства видов развиты слабо. В осязании большую роль играет язык.
Скелет.
Он почти полностью окостенел благодаря замене грубой волокнистой костной ткани, характерной для земноводных, тонкой волокнистой, его прочность возросла, опорная и защитная функции улучшились (рис. 96).
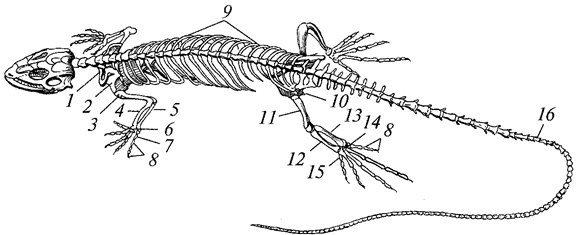
Рис. 96. Скелет ящерицы:
7 — ключица; 2 —
лопатка; 3 —
плечо; 4 —
лучевая кость; 5 —
локтевая кость; б — запястье; 7 — пястье; 8 —
фаланги пальцев; 9 —
ребра; 10 —
таз; 11 —
бедро; 12 —
большая берцовая кость; 13 —
малая берцовая кость; 14 —
предплюсна; 15 —
плюсна; 16 —
хвостовые позвонки
Череп увеличился; вместимость мозгового отдела стала большей, чем у амфибий. Боковая поверхность черепа у древних рептилий была сплошной, позднее в ней появились височные ямы — одна или две, что уменьшило его массу и увеличило поверхность для прикрепления жевательных мышц. Нижние челюсти, как и у амфибий, подвешены к мозговому отделу черепа при помощи квадратной кости и тоже состоят из нескольких костей (зубной, сочленовной и др.).
Тело первого позвонка (атланта)
приросло в виде зубца к телу второго позвонка (эпистрофея).
Кольцевидный атлант получил возможность вращаться вокруг зубца эпистрофея, благодаря чему стали возможны и повороты головы, а это имело большое значение для добывания пищи (в основном других животных) и для защиты от врагов. Атлант соединен с затылочным отделом черепа с помощью одного мыщелка, что также способствует увеличению подвижности головы. Позвоночный столб разделяется на пять отделов: шейный, грудной, поясничный, крестцовый и хвостовой. Позвонки грудного отдела соединяются с грудиной хорошо развитыми ребрами, вследствие чего образуется грудная клетка, отсутствующая у амфибий. От поясничных позвонков отходят укороченные ребра, не доходящие до грудины. Крестцовых позвонков два (у амфибий один). В хвостовом отделе (за исключением черепах) позвонков много.
Кости ног лучше развиты, и они более прочные, чем у земноводных. Между костями запястья и предплюсны имеются подвижные суставы. Поэтому предплечье и проксимальный ряд запястья, а также голень и проксимальный ряд предплюсны функционируют как целостные образования. Особо нужно отметить усиление поясов конечностей: плечевого — благодаря его соединениям с грудиной и грудным отделом позвоночника, тазового — вследствие того, что он опирается на два крестцовых позвонка и обе его половины образуют два сращения — седалищное и лонное.
Мышечная система.
Она подверглась дифференциации, развились новые группы мышц, от которых зависят повороты головы, движения возникшей у рептилий шеи, расширение и сужение грудной клетки и др. Значительно увеличилась мускулатура ног, развились сгибатели и разгибатели пальцев. Масса и сила всей системы возросли.
Пищеварительная система.
Она более дифференцирована, чем у земноводных (рис. 97). Зубы в основном сосредоточены на челюстях, количество их велико, но они однотипны и служат преимущественно для захвата добычи. Слюнные железы лучше развиты, чем у амфибий (у ряда видов выделения этих желез ядовиты). У рептилий начинается процесс образования твердого нёба, которое особенно развито у крокодилов. Благодаря образованию твердого нёба ротовая полость отделяется от носоглоточной, что облегчает как дыхание, так и поглощение пищи. Хорошо развиты (особенно у хищников, поедающих позвоночных животных) глотка, пищевод и желудок. Печень (с желчным пузырем) и поджелудочная железа более совершенны, чем у земноводных. Тонкая кишка значительно удлинена, толстая кишка короткая и заканчивается клоакой. Современные пресмыкающиеся питаются преимущественно животными, число растительноядных видов незначительно.
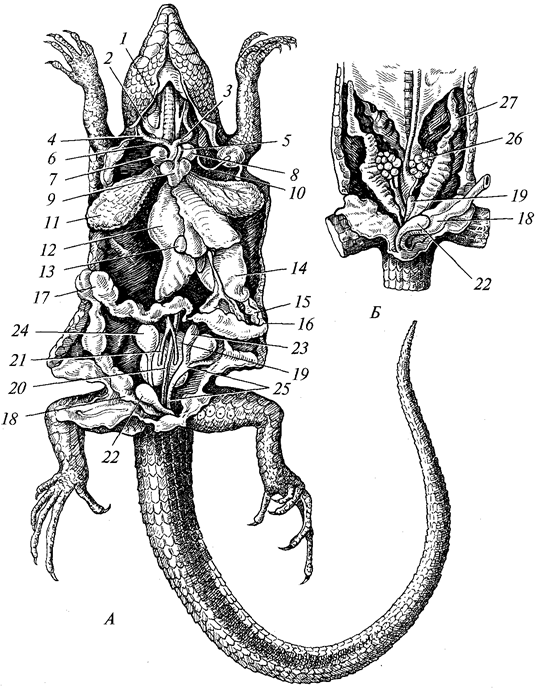
Рис. 97. Внутренние органы ящерицы; А —
самец; Б —
самка:
/ - внешняя и 2 -
внутренняя яремные вены; 3 -
левая и 4 -
правая сонные артерии; 5 - левая и 6 -
правая дуги аорты; 7 - правое и 8 -
левое предсердия; 9 -
желудочек сердца; 10 -
левая подключичная артерия- 11 -
легкое; 12 -
печень; 13 -
желчный пузырь; 14 -
желудок; /5 - поджелудочная железа; 16 —
двенадцатиперстная кишка; 17 —
толстая кишка- 18 —
прямая кишка; 19 -
почка; 20 -
спинная аорта; 21 -
выносящие вены почек; 22 -
мочевой пузырь; 23 -
придаток семенника; 24 -
семенник; 25 —
семяпровод; 26 —
яичник; 27 —
яйцепровод
Дыхательная система.
Органами дыхания у рептилий служат легкие (см. рис. 97), так как их кожа непригодна для газообмена; слизистая оболочка ротовой полости, играющая значительную роль в дыхании земноводных, используется для выполнения этой функции немногими видами пресмыкающихся. По этой причине и особенно в связи с возросшей активностью рептилий по сравнению с амфибиями строение легких и дыхательных путей усложнилось. Легкие представляют собой органы большей величины и глубже расположены в грудной полости, чем у земноводных. Внутренняя поверхность их значительно возросла благодаря развитию на ней множества перекладин с более густой сетью кровеносных сосудов. Воздух поступает в легкие, пройдя через хоаны, глотку, гортань, длинную трахею, просвет которой поддерживается хрящевыми кольцами. Во время прохождения через трахею воздух очищается от пыли, температура его приближается к температуре тела. У амфибий трахея была в зачаточном состоянии. Задний конец трахеи разделяется на два бронха, в стенках которых тоже имеются хрящевые кольца. Осуществление вдоха и выдоха происходит более эффективно, чем у амфибий, благодаря развитию у рептилий грудной клетки, объем которой увеличивается при вдохе и уменьшается при выдохе. Такой же механизм сохранился у высших позвоночных — птиц и млекопитающих.
Кровеносная система.
Сердце, как и у амфибий, трехкамерное, но в желудочке снизу появляется зачаток перегородки. У крокодилов она уже полная, и сердце у них становится четырехкамерным (два предсердия и два желудочка). Следовательно, у пресмыкающихся усиливается разделение венозного и артериального токов крови, но полной изоляции обоих токов еще нет даже у крокодилов, как это видно из нижеследующего описания (рис. 98).
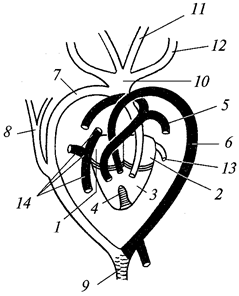
Рис. 98. Сердце и основные сосуды ящерицы (светлые — сосуды с артериальной кровью, штриховка — со смешанной кровью; черные — с венозной):
1 —
правое предсердие; 2 —
левое предсердие; 3 —
желудочек; 4 —
неполная перегородка, разделяющая желудочек на левую и правую половины; 5 — легочная артерия; 6 —
левая дуга аорты; 7 — правая дуга аорты; 8 —
подключичная артерия; 9 —
спинная аорта; 10 —
общий ствол сонных артерий; 11 —
левая внутренняя сонная артерия, 12 —
левая наружная сонная артерия; 13 —
легочная вена; 14 —
задняя полая вена и передние полые (яремные) вены
В левое предсердие поступает артериальная кровь — она идет из легких. В правое предсердие поступает венозная кровь — она собирается из всех частей тела. Далее кровь из обоих предсердий поступает в желудочек: в правую его половину — венозная, в левую — артериальная. Благодаря наличию перегородки, хотя и неполной, кровь в желудочке меньше смешивается, чем у земноводных. Артериального конуса нет, и главные артериальные стволы отходят непосредственно от сердца.
От правой стороны желудочка отходят легочные артерии (бывшая четвертая пара жаберных артерий), несущие в легкие венозную кровь. Бывшая третья пара жаберных сосудов атрофировалась. Вторая пара сосудов состоит из двух дуг аорты: правая дуга начинается от левой части желудочка и несет артериальную кровь, левая дуга начинается от правой части желудочка (но ближе к левой его части) и несет смешанную кровь. От правой дуги аорты отходят сонные артерии (бывшая первая пара жаберных сосудов), идущие к голове. Они снабжают мозг, особенно чувствительный к недостатку кислорода, артериальной кровью. Обе дуги аорты соединяются в аорту, которая разветвляется на множество артерий. Таким образом, организм пресмыкающегося в основном снабжается смешанной кровью, но она насыщена большим количеством кислорода, чем у земноводных, что способствует более интенсивному обмену веществ.
Выделительная система.
Она сильно изменилась в связи с усилением интенсивности обмена веществ рептилий, что привело к увеличению количества продуктов диссимиляции, и полным приспособлением их к наземному образу жизни, вследствие чего возникла необходимость более экономно использовать поступающую в организм воду. Поэтому у рептилий развились более сложные органы выделения, названные тазовыми
или вторичными почками {метаяефрос).
Все канальцы этих почек не имеют воронок и начинаются боуменовыми капсулами, окружающими мальпигие-вы тельца с увеличенным количеством капилляров. Канальцы тазовых почек очень длинные и густо оплетены капиллярами. В боу-меновых капсулах из просочившихся туда из мальпигиевых телец растворенных в воде продуктов диссимиляции и некоторых органических веществ (например, глюкозы) образуется первичная моча. Последняя во время прохождения через длинные почечные канальцы отдает обратно в кровь через оплетающие их стенки капилляры значительную часть воды и другие вещества, необходимые для организма. Образовавшаяся после этого вторичная моча выходит не через вольфовы каналы, а через заново развившиеся мочеточники в клоаку и накапливается в мочевом пузыре. Процессы образования первичной и вторичной мочи совершаются и в туловищных почках, но с возвратом меньшего количества воды в кровь. Первичноводные же позвоночные, в особенности земноводные, кожа которых голая, часто страдают от избытка воды, а не от ее недостатка. Следует также отметить, что у рептилий конечным продуктом разложения белков является мочевая кислота, на выведение которой из организма с мочой затрачивается мало воды. У земноводных и у ряда других первичноводных позвоночных разложение белков приводит к образованию мочевины, на удаление которой воды требуется много.
У эмбрионов рептилий есть зачаток головной почки и развитая туловищная почка, которая функционирует у взрослых особей (у ряда видов ящериц) до наступления половой зрелости. Эти факты подтверждают преемственность трех видов почек в течение эволюции позвоночных.
Размножение.
В состав мужской половой системы рептилий входят два семенника округлой формы, лежащие по бокам поясничного отдела позвоночника, два придатка семенников, образовавшиеся из передних участков туловищных почек, два семяпровода (вольфовы каналы) и совокупительные органы, развившиеся из стенки клоаки. Женская половая система состоит из двух яичников, сильно разрастающихся в период размножения, и двух яйцеводов (мюллеровых каналов), впадающих в клоаку. Оплодотворение только внутреннее. Семя вводится самцами в клоаку самки при помощи совокупительных органов. Созревшие яйцеклетки, содержащие много желтка, попадают в воронки яйцеводов. Соединение женских и мужских гамет происходит в верхней части яйцеводов, куда проникают после полового акта сперматозоиды. Зиготы, продвигаясь по яйцеводам, в средней части последних окружаются белковой оболочкой, а в задней покрываются скорлупой.
У некоторых видов ящериц и змей наблюдается партеногенетическое размножение самок. Самцов у таких видов, как правило, нет.
Развитие.
У рептилий развитие прямое; личинок у рептилий нет. Оплодотворенные яйца пресмыкающиеся обычно откладывают в почву, а некоторые виды — под гниющие растительные остатки. У ряда видов развитие зигот совершается в половых путях самки и сразу после выхода развившихся яиц наружу из них вылупляются сформированные животные, т. е. для таких видов характерно яйцеживорождение.
У немногих видов наблюдается настоящее живорождение: сосуды желточного мешка тесно соединяются с сосудами яйцевода, и эмбрион получает значительную часть питательных веществ из организма матери. Яйцеживорождение чаще наблюдается у видов, распространенных в северных и горных районах. Очевидно, яйцеживорождение является защитой развивающихся зародышей от охлаждения и других неблагоприятных условий. Нет ни одного вида рептилий, у которого яйца развиваются в воде. Даже у рептилий, проводящих большую часть жизни в воде (все крокодилы, многие черепахи, некоторые змеи), откладка яиц и их развитие совершаются только на суше. Морские змеи, никогда не покидающие воду, относятся к живородящим.
Яйца и зародыши рептилий обладают рядом приспособлений для развития на суше. Во-первых, яйца покрыты скорлупой
и другими оболочками, которые защищают их от высыхания и повреждений. Во-вторых, у зародышей развиваются особые зародышевые оболочки
(серозная, амнион и аллантоис), которые имеют большое значение для обеспечения эмбрионального развития животных на суше. Серозная оболочка и амнион развиваются из одной и той же двойной складки тела зародыша, происходящей из эктодермы и мезодермы. Эта складка появляется с двух противоположных сторон в виде зачатков, которые растут навстречу друг другу и смыкаются над зародышем. Наружная часть складки превращается в серозу (ее наружный слой эктодермальный, внутренний — мезодермальный), а внутренняя — в амнион (его наружный слой мезодермальный, внутренний — эктодермальный). Полость, образуемая амнионом, наполнена водянистой жидкостью, благодаря чему развитие происходит во влажной среде, и зародыш защищен от высыхания и последствий различных сотрясений, особенно опасных в условиях воздушной среды.
Происхождение.
Первые рептилии, возникшие во второй половине каменноугольного периода, обитали во влажных биотопах суши и, вероятно, размножались и развивались в воде. К этому времени суша уже была заселена низшими наземными растениями, различными беспозвоночными, начиналось завоевание воздуха насекомыми. Рептилии, которые могли удаляться на большие расстояния, чем амфибии, от водоемов, имели в изобилии пищу и не встречали серьезных врагов. В конце каменноугольного периода и неоднократно в течение пермского периода климат становился более засушливым, что вызывало гибель многих амфибий и, наоборот, содействовало прогрессивному развитию и более широкому распространению на суше рептилий. Спустя некоторое время они перешли к размножению и развитию вне водоемов, сначала в более влажных наземных биотопах, а потом в более сухих местах. В течение мезозойской эры они распространились по всему земному шару и возникло много различных групп. Наряду с хищниками 'было много растительноядных видов, появились рептилии, приспособленные к полету, и рептилии, обитающие в морях. Величина их тела была различна, но большинство были крупными животными, а некоторые имели гигантские размеры — до 20 м и более в длину. Климат на протяжении почти всей мезозойской эры был теплым, очень благоприятным для этих животных, не имеющих достаточно высокой постоянной температуры тела и теряющих свою активность при охлаждении окружающей среды.
В конце мезозойской эры на земном шаре происходили мощные горообразовательные процессы, сопровождающиеся резким изменением условий существования (температуры, влажности и др.). Это повлекло за собой массовое вымирание мезозойских рептилий, не выдержавших борьбу за существование в таких тяжелых для них абиотических условиях среды, осложненную соревнованием с млекопитающими и птицами. Тогда же возникли и стали многочисленными в течение кайнозойской эры новые группы пресмыкающихся — ящерицы и произошедшие от них змеи. Из старых же групп сохранились лишь один очень примитивный вид первоящеров, небольшое количество крокодилов и черепах. Современные рептилии распространены преимущественно в теплых странах, особенно в субтропических и тропических, в странах с умеренным климатом их мало, на далеком севере они отсутствуют.
Систематический обзор
В мировой фауне насчитывается около 6600 видов рептилий:
клювоголовых — 1, черепах — 230, крокодилов — 22, ящериц — около 3900, змей — около 2500. По численности видов рептилии стоят среди наземных позвоночных на втором месте, уступая только птицам. Сохранилось 3 современных подкласса и всего 4 отряда пресмыкающихся.
Отличаются от остальных рептилий прежде всего панцирем, состоящим из спинного и брюшного костных щитов, покрытых снаружи роговыми пластинками. Зубов нет, их заменяют заостренные роговые края челюстей. Скелет и другие системы органов довольно сильно изменены. Некоторые черепахи достигают огромной величины и имеют в длину до 2 м при массе 500—600 кг. Человек истребил большое количество черепах, так как мясо и яйца многих из них съедобны, а щиты идут на разные изделия. Большинство черепах временно или постоянно живут в воде. Интересно, что у последних панцирь сильно облегчен, а конечности превратились в ласты для плавания. Питаются черепахи преимущественно различными животными (беспозвоночными, рыбами и др.), но некоторые виды растительноядны.
Это единственный сохранившийся отряд в подклассе архозавров. Крупные хищные пресмыкающиеся (самые большие из них достигают 10—12 м), ведущие водный образ жизни. Организация крокодилов в некоторых отношениях более совершенна, чем других рептилий (лучше развита нервная система, сердце четырехкамерное, зубы сидят в ячейках, имеется развитое твердое нёбо и т. д.). В настоящее время их осталось небольшое число видов, и они все включены в Красную книгу. Крокодилы питаются крупными водными беспозвоночными, рыбами и другими позвоночными, в том числе млекопитающими. Некоторые виды нападают на человека. Из кожи крокодилов изготавливают различные изделия.
Единственный представитель этой очень древней группы — гаттерия (Sphenodonpunctatis) — сохранился только на островах Новой Зеландии. Строение характеризуется рядом примитивных признаков, к которым относятся двояковогнутые позвонки с остатками хорды между ними, наличие на сошнике зубов, отсутствие совокупительных органов и др. Длина тела этой рептилии достигает 60 см. После заселения Новой Зеландии людьми гаттерия была почти истреблена; сейчас от полного истребления ее защищает специальный закон об охране этого замечательного животного.
Отряд включает 4 современных подотряда, из которых здесь рассматриваются только 2.
Включает животных, как правило, небольших размеров. Обитают в самых разнообразных условиях: в степях, пустынях, лесах, горах и т. д. Мелкие виды питаются насекомыми и другими беспозвоночными, более крупные (вараны и др.) нападают на позвоночных; немногие виды растительноядны. Ящерицы истребляют вредных насекомых. Некоторых ящериц используют в пищу, но в общем практическое значение рассматриваемого подотряда невелико. Известна лишь одна ядовитая ящерица — ядозуб, живущая на юге Северной Америки.
Своеобразная группа позвоночных, произошедших от каких-то вымерших ящериц. Следует отметить, что среди современных ящериц имеются виды, у которых наблюдается редукция ног. Змеи отличаются от своих предков не только отсутствием ног, но и рядом других признаков. Тело очень вытянутое, количество ребер увеличилось (на них при ползании змеи опираются). У удавов сохранились рудименты таза и задних ног. Грудины у змей нет. Веки срослись и стали прозрачными, что защищает глаза от разных повреждений. Среднее ухо и барабанная перепонка редуцировались. Одни органы (почки, половые железы и др.) сильно удлинились, другие уменьшились или атрофировались (левое легкое и др.).
Почти все змеи — хищники, и лишь некоторые виды питаются насекомыми. Кости, образующие верхнюю челюсть, подвижно соединены между собой и с соседними костями, а левая и правая половины нижней челюсти соединены связкой. Поэтому змеи могут широко раскрывать рот и заглатывать добычу больших размеров; стенки пищеварительной трубки тоже способны к значительному растяжению. Среди рассматриваемого отряда довольно много ядовитых видов. Яд выделяется видоизмененными верхнегубными слюнными железами и попадает в тело жертвы через раны, нанесенные особыми удлиненными зубами, расположенными на верхней челюсти. Эффективным средством против змеиного яда являются специальные сыворотки.
В нашей стране мало ядовитых змей, среди которых наиболее распространена гадюка обыкновенная.
В степях южной части европейской территории обитает гадюка степная,
а также очень ядовитая гюрза.
На домашних млекопитающих яд змей действует нередко сильнее, чем на человека. Однако польза, приносимая змеями, вероятно, значительно превышает причиняемый ими вред, так как они истребляют большое количество вредных грызунов, а яд разных змей идет на приготовление ряда лекарственных препаратов, в связи с чем некоторые виды этих пресмыкающихся разводят в специальных питомниках (серпентариях).
Птицы появились в начале юрского периода (по новейшим данным — в триасе). Они произошли от пресмыкающихся, организация которых к этому времени уже сильно усложнилась в результате длительного эволюционного развития. Это сделало возможным переход древесных видов сначала к планирующему способу передвижения, а впоследствии к полету. Приспособление к полету наложило отпечаток на строение птиц и их жизнедеятельность. Отдел зоологии, посвященный всестороннему изучению птиц, называется орнитологией.
Птицы и млекопитающие имеют постоянную и притом высокую температуру тела, которая не зависит или слабо зависит от колебаний температуры внешней среды. У остальных животных температура тела меняется в зависимости от изменения температуры окружающей их среды. Животных с постоянной температурой тела называют гомойотермными,
а животных с переменной температурой — пойкилотермными.
У птиц температура тела очень высокая, в среднем выше, чем у млекопитающих. Даже у низших из современных птиц (бескрыла, или киви) она равна 37,8 °С. В среднем же температура тела разных видов птиц около 42 °С, а максимальная температура у ряда воробьиных достигает 45,5 "С. Столь высокая температура тела имеет огромное физиологическое значение, ибо она является одним из важнейших условий высокой интенсивности обмена веществ и всех физиологических процессов, происходящих в организме птиц.
Большое значение имеет не только высокая температура, но и ее постоянство. Тончайшие, наиболее совершенные физиологические процессы (в особенности процессы, протекающие в головном мозге) нуждаются в постоянстве условий внутри организма. Благодаря постоянству температуры
тела гомойотермные животные могут вести активную жизнь и при низкой температуре.
Общая характеристика
Тело птиц состоит из округлой головы, туловища, начинающегося очень подвижной шеей, передних конечностей, превратившихся в крылья, и хорошо развитых ног. Форма тела — обтекаемая, лишенная выступающих частей, компактная;
крылья у птиц, передвигающихся по земле или находящихся в покое, сложены на спине. Поверхность распростертых во время полета крыльев значительно превышает поверхность туловища. Отмеченные особенности имеют большое значение для обеспечения передвижения птиц в воздухе, а также по земле. Хвост отсутствует.
Покровы.
Кожа тонкая, что имеет существенное значение для уменьшения массы тела птиц и обеспечения подвижности перьев. В ней нет, как и у современных рептилий, желез, клетки наружных слоев ее эпидермиса сильно ороговевают, вследствие чего резко уменьшается испарение воды, столь необходимой для интенсивного обмена веществ, присущего птицам. Соедини-тельнотканный слой кожи подстилается рыхлой клетчаткой, в которой могут накапливаться запасы жира.
Тело покрыто перьями. Различают перья контурные и пуховые (рис. 99).
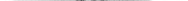
Рис. 99. Контурное перо (общий вид снизу):
1 —
ствол (стержень); 2 —
очин (на рисунке частично вскрыт); 3 —
отверстие очина; 4 —
душка (омертвевший сосочек); 5 — наружное опахало; 6 —
внутреннее опахало; 7 — пуховая часть опахала; 8 —
добавочный стержень
Контурное перо состоит из рогового стержня, который разделяется на очин
и ствол,
от ствола отходят длинные роговые пластинки — бородки первого порядка,
от них — более короткие бородки второго порядка,
сцепленные крючочками, в результате чего образуется прочная
пластинка — опахало.
В пуховом пере стержни слабые, бородки — мягкие, не сцепленные крючочками. Различают еще пух, в котором стержня нет, а имеются только мягкие бородки. Контурные перья прикреплены к коже не на всей поверхности, а только на определенных участках — птерилиях.
Остальные участки кожи — аптерии —
лишены контурных перьев, хотя они прикрыты ими. На этих участках у большинства пиц есть пух и пуховые перья. Последние могут быть и на птерилиях. Контурные перья названы так потому, что они в значительной степени определяют форму (контур) птиц. Они образуют прочную поверхность крыльев и на заднем конце тела — рулевую лопасть. Следовательно, контурные перья играют первостепенную роль в обеспечении полета. Весь перьевой покров предохраняет тело от излишней отдачи тепла и от механических повреждений, столь возможных во время полета, а также при передвижении по земле, на деревьях и т.д. Отмеченные положительные свойства перьевого покрова сочетаются с его малой массой, что так важно для летающих животных. Для сохранения эластичности контурных перьев птицы при помощи клюва смазывают их выделениями единственной кожной железы — копчиковой,
расположенной около клоаки.
Образование пера начинается с выпячивания эктодермального сосочка, схожего с первыми стадиями развития роговых чешуи рептилий. Внутри сосочка имеется вырост соединительнотканно-го слоя кожи с кровеносными сосудами, питающими развивающееся перо. Снаружи сосочек окружен эпидермальным чехликом, под покровом которого происходит ороговение групп эктодер-мальных клеток и превращение их в бородки. По окончании развития бородок чехлик сбрасывается, освобождая готовое перо в виде пуха. При формировании пуховых и контурных перьев образуются стержни, от которых отходят бородки. Следовательно, готовые перья представляют собой чисто эктодермальные образования, в развитии которых принимает участие мезодермальный слой кожи.
Несмотря на достаточную прочность перьев, они все-таки повреждаются в результате воздействия разных внешних факторов и изнашиваются. Поэтому у птиц происходит периодическая смена оперения — линька.
У одних птиц линьке одновременно подвергается весь перьевой покров. Такие птицы во время линьки не могут летать и прячутся в укромных местах. У других же птиц смена оперения совершается постепенно. Линька первого типа наблюдается у видов, которые добывают пищу не в полете; линька второго типа характерна для видов, которые настигают свою жертву на лету. Смена перьев необходима также в связи с изменениями условий среды в разные времена года, например, в зимнем оперении больше пуха и пуховых перьев, чем в летнем, и т. д.
Окраска перьев птиц разнообразна и характерна для каждого вида. Она обусловлена сочетанием разных пигментов: темных — меланинов,
желтых и красных — липохромов.
При отсутствии пигментов окраска перьев белая. Значение окраски различно: благодаря ей птицы менее заметны на фоне тех биотопов, где они обитают; она облегчает общение между особями одного и того же вида и в том числе встречу самцов и самок. Она меняется в зависимости от изменений абиотических условий среды, например, птицы, обитающие в более влажных и теплых местах их
ареала, имеют более темную окраску, а в более сухих и холодных местах — более светлую. У ряда видов окраска может меняться в связи с изменением сезонных климатических условий.
Нервная система.
В связи с очень энергичным образом жизни птиц, потребовавшим усложнения всех систем органов, нервная система развита значительно лучше, чем у рептилий. Головной мозг намного превосходит по размерам мозг рептилий (рис. 100).

Рис. 100. Головной мозг голубя сбоку:
1 —
большие полушария; 2 —
обонятельные доли; 3 —
эпифиз; 4 —
гипофиз; 5 — хиазма и зрительные нервы; 6—
зрительные доли среднего мозга; 7 — мозжечок;
8 —
продолговатый мозг
Передний мозг велик, полосатые тела его, в которых находятся центры регуляции зрения, движения и других функций, весьма развиты. Кора полушарий выражена значительно лучше, чем у пресмыкающихся. В результате этих и других прогрессивных изменений роль переднего мозга как координатора всей сложной жизнедеятельности птиц резко возросла, способность к установлению новых связей со средой путем более быстрого приобретения условных рефлексов значительно улучшилась. Обонятельные доли развиты меньше, чем у ранее рассмотренных классов позвоночных, так как у птиц в их отношениях со средой главную роль играют зрение и слух. Усложнились и другие отделы головного мозга, в особенности мозжечок, имеющий очень крупные размеры. Значение его чрезвычай-f но возросло в связи с приспособлением птиц к полету. Спинной ;
мозг ввиду редукции хвоста короткий, но строение его сильно , усложнилось. Головных нервов — 12 пар.
Органы чувств.
Глаза велики, их масса у многих видов превосходит массу головного мозга. Дальность зрения у многих видов очень велика, поле зрения весьма широко; аккомодация к видению на разных расстояниях совершенна и осуществляется разными способами — изменением кривизны хрусталика, а также роговицы, в меньшей степени более старым способом, т. е. перемещением хрусталика. Птицы хорошо различают цвета, их оттенки, особенности рисунка и окраски окружающих их предметов. Необходимость столь совершенного зрения птиц вполне понятна, если учесть быстроту их передвижения во время полета, да и во многих случаях по земле.
Орган слуха устроен более сложно, чем у рептилий; нижний мешочек, воспринимающий звуковые колебания, увеличен за счет отходящего от него отростка (его называют завитком улитки, сравнивая с гомологичным органом млекопитающих, у которых этот отросток имеет 2,5 оборота); слуховая косточка благодаря изменениям в ее строении лучше передает в лабиринт звуковые колебания барабанной перепонки, расположенной в более развитом, чем у рептилий, наружном слуховом проходе; обрамление отверстия последнего перьями (особенно у совиных) напоминает слуховую раковину млекопитающих, играющую роль рупора. Птицы, как известно, издают разнообразные звуки, объединяющиеся у многих видов в сложные мелодии. В этом отношении птицы превосходят млекопитающих, способных издавать преимущественно однообразные звуки. Звуковая деятельность играет в жизни птиц огромную роль: в общении родителей с птенцами, самцов с самками, с другими особями того же вида, а также с представителями других видов.
Обоняние, как отмечалось, развито слабее, чем у других позвоночных, но у видов, питающихся падалью, которую они находят по запаху, оно развито лучше. Вкусовые раздражения воспринимаются особыми скоплениями клеток, расположенными на слизистой оболочке ротовой полости и языке. Но эти органы играют в жизни птиц несравненно меньшую роль, чем у млекопитающих. В коже имеются свободные окончания нервов и скопления разных чувствующих клеток, функции которых различны: осязание, восприятие изменений температуры, положения перьев и др.
Для нервной деятельности птиц характерны разнообразные и очень сложные инстинкты.
Инстинктивная деятельность проявляется во взаимоотношениях самцов и самок, постройке гнезд, выкармливании и охране потомства, в способности многих птиц к дальним перелетам в связи со сменой времен года и т. д. Инстинкты проявляются на определенном этапе индивидуального развития птиц в определенных условиях внешней среды. Так, например, инстинкт ухаживания самцов за самками проявляется при достижении ими половой зрелости, наступление которой, в свою очередь, сильно зависит у ряда видов от удлинения светлого периода суток весной. Другие инстинкты свойственны птицам в течение всей жизни. Отсутствие сознания в инстинктивных действиях можно проиллюстрировать на следующем примере. Хорошо известно, что кукушки подбрасывают свои яйца в гнезда мелких воробьиных птиц. Вылупившийся кукушонок, еще слепой и глухой, выбрасывает яйца и птенцов хозяев гнезда, а те продолжают кормить быстро растущего, прожорливого нахлебника. Способность к образованию условных рефлексов у птиц развита во много раз лучше, чем у пресмыкающихся. Этой способностью люди пользуются при разведении домашних птиц. У кур, например, легко и быстро вырабатываются условные рефлексы: они сбегаются к месту кормления, вечером возвращаются на насесты и т. д.
Инстинктивная деятельность особенно ярко проявляется у птиц, совершающих дальние сезонные перелеты: весной — из южных стран в северные страны, где они размножаются и выкармливают своих птенцов, а осенью с приближением более холодного времени — обратно в южные страны. Инстинктивный характер перелетов подтверждается тем, что молодые птицы, появившиеся на севере, в ряде случаев летят на юг самостоятельно, а не с родителями, т. е. не используя опыта последних. Были поставлены опыты с перелетными птицами в условиях, которые оставались характерными для летнего времени года, тем не менее подопытные животные с приближением осени испытывали беспокойство и пытались улететь. Пути перелетов, места пребывания перелетных птиц в южных странах и места, где они выводят свое потомство в северных странах, почти всегда постоянны. Проблема перелетов до сих пор остается до конца не разрешенной, в частности, не выяснено с достаточной полнотой, чем руководствуются птицы, совершая столь далекие передвижения, хотя доказано, что они могут учитывать положение солнца, луны и звезд, обладают «компасным чувством» и т. д. Взрослые птицы, возвращающиеся в южные страны (причем в течение ряда лет), используют свой опыт в выборе наиболее удобных мест для миграций (вдоль рек, границ лесов и т.д.). Во время перелетов большое число птиц, конечно, погибают. Однако поскольку перелеты продолжаются в течение огромных промежутков времени, это означает, что они имеют большое приспособительное значение, так как обеспечивают размножение птиц в благоприятных условиях. Таким образом, способность к таким миграциям возникла у птиц в результате естественного отбора в течение длительного времени. Формирование перелетов начиналось с кочевок на небольшие расстояния. Постепенно расстояние увеличилось, когда это оказалось выгодным для сохранения вида. В заключение следует подчеркнуть, что известен ряд фактов, подтверждающих некоторую изменчивость миграций птиц, что является основой для возникновения новых путей перелетов. Понятно, что способность птиц к таким длительным миграциям стала возможной благодаря их совершенному приспособлению к полету и прогрессивным особенностям всех систем органов, в первую очередь нервной системы. Птицам свойственны и некоторые проявления рассудочной деятельности. Так, например, некоторые хищные птицы поджидают свою жертву, скрывшуюся в кусты, с противоположной стороны убежища.
Скелет.
Череп в связи с большим размером головного мозга относительно велик. Он соединен с позвоночным столбом, как у тех пресмыкающихся, от которых произошли птицы, одним мыщелком — затылочным. Кости черепа тонкие, что уменьшает массу головы и облегчает ее движения при захвате добычи и во время полета, а также понижает центр тяжести тела, способствуя большей его устойчивости. Защита мозга усиливается сращением у взрослых птиц костей черепа, хотя у зародышей швы, разделяющие кости, имеются. В состав черепа входят в основном те же кости, что и у пресмыкающихся. Квадратные кости подвешивают, как и у земноводных и пресмыкающихся, нижнюю челюсть (образованную несколькими костями) к мозговому отделу черепа. Нёбо птиц образовано отростками верхнечелюстных костей, нёбными костями, сошником и крыловидными костями (рис. 101).
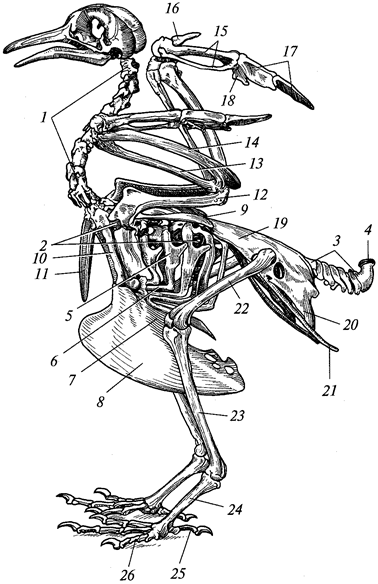
Рис. 101. Скелет голубя:
1
— шейные позвонки; 2 —
грудные позвонки; 3 —
хвостовые позвонки; 4 —
копчиковая кость; 5 —
спинная часть ребра с крючковидным отростком; 6 —
брюшная часть ребра; 7 — грудина; 8 —
киль грудины; 9 —
лопатка; 10 —
коракоид; 11 —
ключица; 12 —
плечо; 13 —
лучевая кость; 14 —
локтевая кость; 15 —
пясть; 16 —
I палец; 77—11 палец; 18 —
III палец; 19 —
подвздошная кость; 20 —
седалищная кость; 21 —
лобковая кость; 22 —
бедро; 23 —
голень; 24 —
цевка; 25 —
I палец; 26 —
IV палец
Для всех отделов позвоночника, кроме шейного, характерно сращение многих позвонков между собой. Эта особенность позвоночного столба обусловливает неподвижность туловища птиц, что имеет большое значение при полете. Шейный отдел позвоночника очень длинный. Строение первых двух шейных позвонков (атланта и эпистрофея) и седлообразные суставные поверхности остальных шейных позвонков обеспечивают чрезвычайную гибкость шеи. Поэтому голова птиц очень подвижна, что облегчает поиски и хватание пищи, чистку перьев, смазывание их выделениями копчиковой железы и т. д.
Ребра состоят из двух частей — позвоночной и грудинной. Такое строение ребер облегчает изменение объема грудной клетки, что очень важно при вдохе и выдохе. Соседние ребра соединены между собой маленькими клювовидными отростками, благодаря чему усиливается прочность грудной клетки. Грудина у летающих птиц и у пингвинов, плавающих при помощи ластов, т. е. видоизмененных крыльев, имеет киль,
к которому прикрепляются сильно развитые большие грудные и другие мышцы. У бегающих птиц (страусов и др.) киля нет.
Передние конечности у птиц превратились в крылья. Основу крыла составляет измененный скелет передних конечностей наземных рептилий, который состоит из хорошо развитой плечевой кости, прямой и более тонкой лучевой кости, изогнутой и более толстой локтевой кости и кисти, в которой произошли редукция ряда костей и слияние некоторых костей между собой. Из костей запястья остались лишь две косточки. Пястный отдел кисти представляет собой две кости, соединенные своими концами, образующие «пряжку». Сохранились рудименты трех первых пальцев, из которых второй развит лучше других. В результате этих изменений передняя конечность потеряла способность к передвижению по земле и не может выполнять хватательные функции, но ее кости образуют удлиненное и надежное основание для прикрепления мышц и перьев. Плечевой пояс состоит из узких лопаток, вороньих костей и ключиц. Последние срастаются нижними концами и образуют «вилочку».
Вследствие превращения передних конечностей в крылья движение по земле осуществляется только при помощи задних конечностей, причем вся тяжесть падает на них. В связи с этим скелет таза испытал ряд существенных изменений. Таз птиц очень велик. У рептилий он был соединен с позвоночником в области двух (крестцовых) позвонков. У птиц сращение таза с позвоночным столбом, кроме крестцовых позвонков (их тоже обычно два), распространяется на оставшиеся хвостовые, все поясничные и даже задние грудные позвонки. Эти позвонки срастаются между собой и образуют сложный крестец, в состав которого может входить до 24 позвонков. Таким образом, ноги птиц имеют очень прочную опору. Седалищные кости срастаются на всем протяжении с подвздошными. Лобкового соединения нет и таз спереди широко открыт, благодаря чему птицы могут нести яйца большой величины.
С тазом сочленяется бедро. После бедра идет очень длинная голень, состоящая из большой берцовой кости, к которой в виде тонкого рудимента прирастает малая берцовая кость. Как показали эмбриологические исследования, с дистальным концом большой берцовой кости срастаются слившиеся кости первого (про-ксимального) ряда — предплюсны. Второй (дистальный) ряд костей предплюсны срастается с плюсневыми костями и образуется довольно длинная кость — цевка.
В результате описанных изменений скелет ноги птицы стал более прочным, а общая длина этой конечности значительно удлинилась, что обеспечило более быстрое передвижение и облегчило добывание пищи при помощи клюва. Пальцев у большинства птиц четыре, причем первый направлен назад.
Скелет птиц в связи с приспособлением их к полету довольно сильно облегчен, что достигается разными способами: многие кости очень тонкие, в них мало костного мозга, в большие кости заходят ответвления воздушных мешков и т. д. Кости, наполненные воздухом, называются пневматическими.
Мышечная система.
Она значительно дифференцированна и массивнее, чем у рептилий. Мышцы в основном сосредоточены на туловище и проксимальных частях конечностей, что способствует усилению компактности тела, важной для полета. Мышцы, выполняющие наибольшую нагрузку, содержат белок миоглобин
(мышечный гемоглобин, близкий по составу к гемоглобину эритроцитов), благодаря которому в их волокнах накапливается резерв кислорода. Многочисленные мышцы шеи обеспечивают разнообразные и быстрые движения головы. Наибольшего развития достигает большая грудная мышца, прикрепляющаяся к килю грудины.
В функции этой мышцы входит опускание крыла, для чего необходимо преодолевать большое сопротивление воздуха. Масса ее может доходить до 1/5—1/4 массы тела. Подъем крыла обеспечивается малой грудной (подключичной) мышцей, которая прикреплена к вороньей кости (коракоиду) и грудине, а ее сухожилие — к головке плечевой кости. Расширение грудной клетки происходит благодаря сокращению межреберных и некоторых других мышц. Очень хорошо развиты мускулы ног, из которых самые большие прикреплены начальными концами к разным участкам таза, а более мелкие находятся на бедре и голени. Весьма совершенны разгибатели и сгибатели пальцев. У птиц развился мышечный механизм, обеспечивающий им прикрепление при помощи пальцев к веткам деревьев во время сна. Имеется подкожная мускулатура, от которой зависит изменение положения перьев на больших участках поверхности тела и отдельных перьев.
Различают два главных вида полета — парящий
и машущий.
Первый присущ многим птицам: обладая прекрасно развитой нервной системой, они хорошо ориентируются в воздушных течениях и используют восходящие токи воздуха для парения при помощи распростертых крыльев, редко взмахивая ими. Второй возможен только при непрерывной работе крыльев, число взмахов которых у самых мелких птиц — колибри доходит до 50—80 в секунду. Скорость полета различна: у голубей — 30—60 км/ч, у ласточек — 40— 60, у стрижей — до 100—120 км/ч. Скорость полета возрастает при попутном ветре и замедляется при встречном. Перелетные птицы, хорошо приспособленные к длительному нахождению в воздухе, преодолевают в сутки в среднем 200—400 км. Полет происходит, как правило, на небольшой высоте (100—300 м), но птицы могут подниматься и на несколько тысяч метров. Длина пролетных путей некоторых видов птиц достигает 12000 км.
Пищеварительная система.
Все современные птицы не имеют зубов. Края челюстей — роговые, образуют клюв, служащий для захвата пищи и у некоторых видов — для размельчения ее. Поскольку в ротовой полости птиц не происходит механической обработки пищи, как это имеет место у млекопитающих, эта обработка почти целиком совершается в других частях пищеварительной трубки. В ротовую полость открываются протоки слюнных желез, секрет которых смачивает пищу. Из ротовой полости пища попадает в глотку и далее в пищевод, где у многих птиц она задерживается в особом расширении — зобе.
В нем смоченная пища размягчается. Из зоба через нижний отдел пищевода пища проходит в железистый желудок,
где она обволакивается пищеварительными соками, а затем переходит в мышечный желудок.
У хищных птиц этот отдел выражен слабо, но он прекрасно развит у растительноядных видов. Мощная мускулатура желудка, его жесткая внутренняя поверхность, а также находящиеся в его полости проглоченные камешки обеспечивают перемалывание различных пищевых продуктов, заключенных в крепкие оболочки. Здесь же, поскольку пища в предыдущем отделе была смешана с пищеварительными соками, продолжается ее химическое разложение, но в основном оно происходит в тонком отделе кишечника.
Тонкая кишка развита лучше, чем у пресмыкающихся, и в 2— 4 раза превосходит длину тела, но она значительно короче, чем у многих млекопитающих. Очень длинная тонкая кишка и содержащаяся в ней пища чрезмерно увеличивали бы массу тела. Сравнительно небольшая длина тонкого кишечника возмещается сильным действием пищеварительных соков. Эти соки выделяются железами кишечной стенки и хорошо развитой поджелудочной железы. Важную роль в пищеварении играет печень,
состоящая у птиц из двух долей. Желчного пузыря у некоторых птиц (например, у голубя) нет. Всасывание переваренной пищи происходит очень быстро благодаря наличию на внутренней поверхности тонкой кишки большого количества ворсинок.
Толстая кишка очень короткая, она заканчивается клоакой, в которую впадают, как и у пресмыкающихся, выводные протоки мочевых и половых органов. В короткой толстой кишке не могут задерживаться и формироваться экскременты, и птицы испражняются очень часто. Быстрое опорожнение кишечника, возможное благодаря интенсивным процессам пищеварения у птиц, выгодно для облегчения их массы.
Состав пищи различен не только у разных видов, но и у птенцов и взрослых особей одного и того же вида. Так, например, куриные зерноядны, но их птенцы питаются насекомыми. Очевидно, для роста и развития молодых птиц большое значение имеет животный корм.
Дыхательная система.
Она устроена значительно сложнее, чем у рептилий. Бронх,
ответвляющийся от заднего конца трахеи, входит в легкое, пронизывает его насквозь, и его полость переходит за пределами легкого в полость большого воздушного мешка. От центрального бронха отходят вторичные бронхи,
из которых несколько бронхов (чаще четыре) тоже пронизывают легкое и соединяются с другими воздушными мешками. Остальные вторичные бронхи (10—14) заканчиваются слепо, т.е. не соединены с воздушным мешком. Вторичные бронхи соединены между собой многочисленными парабронхами,
от которых отходит множество каналов — бронхиол,
густо оплетенных капиллярами. Поверхность бронхиол очень велика и обеспечивает интенсивный газообмен, столь необходимый птицам.
Воздушные мешки — тонкостенные образования — отходят от упомянутых выше сквозных бронхов (рис. 102). Имеются следующие воздушные мешки: парные — шейные, подмышечные, переднегрудные, заднегрудные и брюшные, непарный — межключичный.
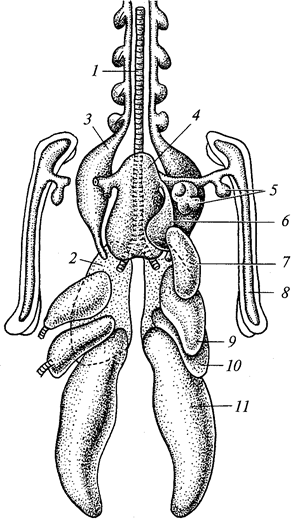
Рис. 102. Схема воздушных мешков птиц; вид с брюшной стороны:
1 —
трахея; 2 —
легкое; 3 —
шейный мешок; 4 —
межключичный мешок; 5—8 —
выросты межключичного мешка; 9 —
переднегрудной мешок; 10 —
заднегрудной мешок; 11 —
брюшной воздушный мешок
У птицы, не находящейся в полете, при вдохе вследствие сокращения определенных мышц расширяется грудная клетка (при этом поднимается грудная кость), что влечет за собой расширение воздушных мешков и приток в них через легкие воздуха. Сами легкие малоэластичны и почти не расширяются. При выдохе, происходящем при сжатии грудной клетки и опускании грудины, воздушные мешки сжимаются и значительная часть наполняющего их воздуха попадает в легкие. Так как в воздушных мешках кислород не потребляется, то капилляры бронхиол насыщаются кислородом не только при вдохе, но и при выдохе. Поэтому принято говорить, что у птиц двойное дыхание.
У летящих птиц грудина остается неподвижной (ибо она является опорой для главных летательных мышц), расширения и сжатия грудной клетки не происходит и вентиляция легких в таком состоянии совершается в результате того, что при подъеме крыльев воздушные мешки расширяются, а при опускании крыльев сжимаются.
Интенсивность газообмена усиливается у птиц благодаря очень высокой частоте вдохов и выдохов (до 90-100 раз в минуту у небольших птиц, т.е. значительно больше, чем у млекопитающих).
Кроме того, гемоглобин птиц быстрее отдает тканям кислород, чем гемоглобин млекопитающих.
Воздушные мешки помимо их важнейшей роли в процессах дыхания выполняют и другие функции: заполняя брюшную полость тела и проникая в некоторые длинные кости, они уменьшают приблизительно на 10 % массу тела, являются изолирующей прокладкой между органами, уменьшают теплоотдачу, облегчают процессы дефекации и откладки яиц.
У птиц очень длинная трахея. Просвет ее поддерживается хрящевыми кольцами, которые в некоторых случаях окостеневают. Гортань, от которой начинается трахея, у птиц не приспособлена к издаванию звуков, а певчей, или нижней, гортанью является орган, возникший в результате преобразования нижней части трахеи и начальных частей главных бронхов, имеющий внутри перепонки и сложно-дифференцированную мускулатуру, способствующую вибрации последних. Трахея помимо очищения воздуха от пыли и согревания его имеет значение для усиления издаваемых нижней гортанью разнообразных звуков.
Кровеносная система.
Сердце птиц четырехкамерное и состоит из двух предсердий и двух желудочков (рис. 103). Правый желудочек, получающий венозную кровь из правого предсердия, полностью отделен от левого желудочка, получающего артериальную кровь из левого предсердия. Левая дуга аорты атрофировалась, а правая дуга аорты, отходящая от левого желудочка, несет артериальную кровь, попадающую по разветвлениям аорты, во все части тела. Следовательно, у птиц полностью отделился артериальный ток крови от венозного, и их организм снабжается только артериальной кровью. Это способствовало резкому усилению обмена веществ у птиц по сравнению с их предками.
Рис. 103. Кровеносная система птиц (светлое — сосуды с
артериальной кровью; черное — с венозной):
1 —
правое предсердие; 2 —
правый желудочек; 3 —
легочная артерия; 4 —
легочные вены; 5 — левое предсердие; 6 —
левый желудочек; 7 — правая дуга аорты; 8 —
спинная аорта; 9 —
безымянная артерия; 10 —
сонная артерия; 11 —
подключичная артерия; 12 —
плечевая артерия; 13 —
грудная артерия; 14 —
внутренностная артерия; 15 —
брыжеечная артерия; 16 —
бедренная артерия; 17
— седалищная артерия; 18 —
яремная вена; 19 —
плечевая вена; 20 —
грудная вена; 21 —
передняя полая вена; 22 —
копчиково-брыжеечная вена; 23 —
воротная вена почки; 24 —
общая подвздошная вена; 25 —
седалищная вена; 26 —
бедренная вена; 27—
почечная вена; 28 —
задняя полая вена; 29 —
печеночная вена; 30 —
воротная вена печени; 31 —
поджелудочная вена; 32 —
брыжеечная вена
Сердце птиц очень часто сокращается: у мелких видов в состоянии покоя — 400—600 раз в минуту, в полете — до 1000, у видов средней величины в состоянии покоя — 200-300, в полете — до 600 раз в минуту; у более крупных видов частота сокращений меньше, но она превосходит аналогичные показатели рептилий и даже млекопитающих. Следовательно, кровь движется в теле птиц с очень большой скоростью, что способствует быстрому удалению продуктов диссимиляции и насыщению тела кислородом и питательными веществами. Сердце птиц, выполняющее такую большую работу, относительно велико: масса его достигает у мелких видов 2,4—2,85 % всей массы тела; у более крупных видов относительная масса сердца меньше, но она значительно больше, чем у рептилий (0,2—0,3 %) и даже у млекопитающих (0,2—1,0 %). Количество крови, число эритроцитов и содержание гемоглобина у птиц значительно больше, чем у рептилий. Отмеченные особенности кровеносной системы, а также ряд других, не упомянутых здесь, обеспечивают весьма энергичный образ жизни птиц.
Выделительная система. В
результате интенсивного обмена веществ у птиц выделяется большое количество продуктов диссимиляции, которые как можно скорее должны быть удалены из организма. Эту функцию выполняют тазовые почки. Последние, унаследованные птицами от рептилий, усложнились, относительная масса их сильно увеличилась, достигнув 1-2,6 % массы всего тела. Почки у большинства видов птиц внешне подразделяются на три доли. Моча удаляется через мочеточники в клоаку, она, как и у пресмыкающихся, богата мочевой кислотой и консистенция ее кашицеобразная. Мочевого пузыря у птиц нет, и моча часто удаляется из клоаки. Накопление ее в мочевыносящих путях увеличивало бы массу тела птиц.
Размножение.
Мужские половые органы сходны с теми же органами пресмыкающихся. Семенники лежат около почек. Левый семенник обычно больше развит, чем правый, но семя вырабатывается обеими железами. Семяпроводы, образовавшиеся, как и у пресмыкающихся, из вольфовых каналов, открываются в клоаку.
Оплодотворение внутреннее, хотя специальных совокупительных органов у большинства птиц нет и передача семени в половые пути происходит при половом акте в результате прижимания клоаки самца к клоаке самки.
Женская половая система почти у всех птиц состоит только из одного левого яичника и левого яйцевода (бывшего мюллерова канала). Правый яичник развивается в очень редких случаях, а производимые им яйцеклетки выходят через левый яйцевод.
Редукция на одной стороне тела женских половых органов произошла, вероятно, в связи с необходимостью облегчения массы тела, меньшее же количество откладываемых птицами по сравнению с пресмыкающимися яиц компенсируется хорошо развитой заботой о потомстве. Созревшие яйцеклетки, окруженные желтком, попадают в воронку яйцевода. Яйцевод состоит из нескольких отделов: трубки, внутренняя поверхность которой покрыта мерцательным эпителием и имеет железы невыясненного значения; широкой части, стенки которой выделяют белок, и хлгазы
(спиральные завитки на обоих полюсах яйца); узкой части, в которой выделяется тонкая белковая оболочка яйца; хорошо развитой, очень мускулистой матки, где яйцо окружается скорлупой; мускулистого влагалища, выталкивающего готовое яйцо в клоаку. Яйцеклетки оплодотворяются в верхней части яйцевода.
Яйца птиц разнообразны по размерам, окраске и толщине скорлупы. Обычно величина яиц пропорциональна размерам тела птиц, но отступления от этого правила нередки. Яйца, откладываемые на землю, камни и т. д., имеют более толстую скорлупу, чем яйца, откладываемые в гнезде. Яйца многих птиц имеют защитную окраску. Количество яиц в кладке у разных видов варьирует.
Половой диморфизм у ряда видов проявляется слабо (совы, голуби и др.), но у большинства птиц самцы отличаются от самок яркостью оперения, размерами, голосом, наличием хохла, воротника, различных роговых выростов. Самки же этих видов имеют неяркую окраску и лишены различных придатков. Способность к пению тоже присуща, как правило, самцам. В период спаривания половое возбуждение самцов может проявляться кроме пения различного рода звуками, телодвижениями и т. п.
Развитие оплодотворенных яиц начинается еще в половом аппарате самки, однако вскоре оно приостанавливается и продолжается уже после откладки яиц. Почти все птицы насиживают яйца, т. е. обогревают их своим телом. Обычно насиживают самки, но у ряда видов эту функцию выполняют оба родителя или только самцы. Продолжительность эмбрионального развития у разных птиц различна и характерна для каждого вида: у кур — 21 день, у самых больших из современных птиц, африканских страусов, — 55—60 дней, а у самых маленьких, колибри, — 10—12 дней.
Птицы относятся к амниотам, и их зародыши имеют те же оболочки, что и пресмыкающиеся, т. е. амнион, серозу и аллан-тоис. Когда у эмбриона развиваются легкие, аллантоис становится ненужным и ссыхается. Развившийся птенец пробивает скорлупу при помощи особого выроста на клюве — яйцевого зуба.
У одних птиц вылупившиеся птенцы вполне развиты и почти сразу начинают самостоятельно двигаться и добывать себе корм. Такие птицы называются выводковыми.
К ним относятся, как правило, птицы, устраивающие свои гнезда на земле, у воды и т. д. (например, куриные, гусиные и др.). У других птенцы вылупляются недоразвитыми: слепые, голые или покрытые редким пухом, беспомощные — некоторое время их выкармливают родители. Такие птицы называются птенцовыми.
К ним относятся, как правило, птицы, устраивающие свои гнезда на деревьях, вообще высоко над землей (например, голубиные, дятлы, воробьиные и др.). Между обеими группами нет резкой границы.
Происхождение.
Птицы, как отмечалось выше, произошли от пресмыкающихся. Наиболее убедительным доказательством этой научной гипотезы в течение долгого времени считался отпечаток животного (обнаруженный в юрских отложениях), названного археоптериксом
(Archaeopterixlithographica). В организации этой древней птицы сочетались признаки рептилий (длинный хвост, пальцы с когтями на передней конечности, наличие самостоятельной малой берцовой кости, зубы на челюстях и др.) и птиц (перьевой покров, примитивные крылья, цевка, большие глазницы и др.). Археоптериксы были величиной с сороку, они вели древесный образ жизни, передвигались, цепляясь когтями за ветки и, вероятно, планируя при помощи крыльев, усаженных большими перьями, могли перебираться с одного дерева на другое.
В последнее время появились сведения о нахождении остатков еще более древнего предка птиц, у которого признаки рептилий (в особенности сильные задние ноги) были выражены сильнее, чем у археоптерикса. Этим подтверждается гипотеза о происхождении птиц от небольших пресмыкающихся — динозавров, которые передвигались при помощи прыжков, опираясь на задние ноги. В меловом периоде мезозойской эры появились птицы, вполне похожие на современных, но у которых еще сохранились зубы. В начале третичного периода кайнозойской эры зубатых птиц уже не было. Таким образом, класс птиц подразделяется на два подкласса: ящерохвостые, или древние птицы (Archae-ornithes), к которым относятся археоптерикс и веерохвостые, или настоящие птицы (Neornithes) (к последним принадлежат и зубатые птицы, хотя некоторые орнитологи выделяли их в отдельный подкласс).
Систематический обзор
Современных птиц обычно подразделяют на два надотряда — плавающие и типичные, или новонёбные птицы. Всего в мировой фауне насчитывается около 8600 видов птиц.
Надотряд содержит единственный отряд пингвинообразные (Spheniciformes). Это прекрасно плавающие птицы, обитающие в Антарктиде. Некоторые виды, следуя холодному течению, могут мигрировать вдоль западных берегов Южной Америки далеко в направлении к экватору. Размеры до 1 м. Плавают при помощи крыльев, превратившихся в ласты. Киль имеется. Стопа широкая, что позволяет пингвинам «ходить» в вертикальном положении по земле, снегу и льдам. Оперение сильноизмененное, напоминающее мех, несмачиваемое. Питаются рыбой. Живут колониями. Забота о потомстве чрезвычайно хорошо выражена. Шкурки идут для изготовления разных изделий. Всего известно 16 видов.
Объединяет всех остальных современных птиц и включает в себя 33 отряда. Краткие характеристики некоторых из них здесь приводятся.
Включает один вид — африканского страуса (Struthiocamelus), самую крупную из современных птиц массой до 136 кг и высотой до 244 см. Страус не летает, но способен быстро бегать, развивая скорость до 50 км/ч. Грудина у него без киля. Откладывает яйца массой около 1,5 кг. Распространен в степях и полупустынях Африки. Питаются грубой растительной пищей. В настоящее время в ряде стран страусов разводят на фермах ради перьев и яиц.
В отряде два вида нелетающих страусоподобных птиц рода нанду (Rhea). Киля у нанду нет. Они меньше страуса, высота до 170 см и масса до 25 кг. Обитают в пампасах и полупустынях Южной Америки. Питаются листьями, побегами и мелкими беспозвоночными.
Мелкие нелетающие бескилевые птицы массой 2—3 кг. В отряде три вида из рода киви (Apteryx), обитающие в лесах и кустарниковых зарослях Новой Зеландии. Питаются беспозвоночными. У киви длинный тонкий клюв с ноздрями на его конце. Ведут ночной образ жизни. Киви являются национальной эмблемой Новой Зеландии и строго охраняются.
Птицы преимущественно средней величины. Кормятся, как Ц правило, на земле. Пища главным образом растительная, у птен-|||; цов — преимущественно животная. Добывают пищу, разгребая землю ногами. Гнездятся на земле. Выводковые. Курообразные (за исключением перепела) оседлы или кочуют на недалекое расстояние. Практическое значение очень велико. Ряд видов — важные промысловые птицы. Человек вывел домашние породы кур, цесарок, индеек и др. Предком домашних кур является банкивская курица (Gallusgallus), до сих пор обитающая в Южной Азии. Она мала (масса самца 900-1250 г, самки — 500—700 г) и несет около 4—13 яиц в год. Путем систематического отбора в разных направлениях были выведены разнообразные породы кур. Масса петухов некоторых пород достигает 4 кг, а кур — 3 кг. Куры наиболее яйценоских пород дают свыше 200 яиц в год.
Предок домашних индеек — обыкновенная индейка (Meleagrisgallopavo). Этот вид до сих пор распространен в диком состоянии в США и является предметом охоты. Домашние цесарки происходят от обыкновенной цесарки (Numidameleagris). Этот вид в настоящее время встречается в Африке.
Небольшие птицы, обитающие преимущественно на деревьях, в скалах. Прекрасно летают. Пища главным образом растительная (семена, плоды), редко животная (беспозвоночные). Птенцовые. Оба родителя выкармливают потомство выделениями зоба — «молочком». Многочисленные породы домашних голубей происходят, как это показал Чарлз Дарвин в «Происхождении видов» и других трудах, от сизого голубя (Columbalivia), распространенного и в настоящее время в южных частях Европы и Азии и в Северной Африке.
Включает птиц разного внешнего облика и образа жизни (журавли, дрофы, коростель, трехперстка и др.), обитающих в открытых местах - степях, лугах, болотах, у воды. Иногда отдельные подотряды рассматриваются как особые отряды. Пища растительная (ягоды и др.) и животная (черви, насекомые, лягушки, мелкие грызуны). Гнезда на земле. Выводковые.
Птицы, населяющие в основном околоводные или водные биотопы (кулики, чайки, чистики). Перелетные.
Кулики — птицы средней и небольшой величины, живущие вблизи воды или в открытых местах. Клюв и ноги длинные. Пища главным образом животная (насекомые, моллюски, черви и др.), которую кулики добывают с помощью длинного клюва. Гнезда на земле. Выводковые. Перелетные; зимовки очень далеки. Некоторые виды имеют большое промысловое значение.
Чистики — птицы средней величины, обитающие на северных морях (до 89° с. ш.). Хорошо ныряют. Голова большая, ноги далеко отставлены назад. Передние пальцы соединены перепонками, заднего пальца нет. Пища животная (у более крупных чистиков — рыба, у других — беспозвоночные). Летом живут колониями, образуя так называемые птичьи базары.
Выводковые.
Птицы (гуси, лебеди, настоящие и нырковые утки и др.) средней или значительной величины, жизнь которых связана с водоемами. Хорошо плавают и летают. Ноги короткие с четырьмя пальцами; передние пальцы соединены перепонкой. Клюв широкий, снабженный роговыми пластинками или зубцами. Гуси питаются растениями на сырых лугах, а иногда на полях. Пища настоящих уток смешанная, у многих видов преимущественно растительная, добываемая ими в воде на мелких местах, иногда и на полях. Нырковые утки могут добывать пищу на большой глубине. Гнездятся около воды (в траве, прибрежных зарослях и т. д.), на водоемах, иногда довольно далеко от них. Выводковые. Большинство — перелетные. Практическое значение гусиных очень велико, многие из них являются важнейшими промысловыми птицами. Домашние гуси произошли от дикого серого гуся (Anseranser). Домашние утки были выведены человеком из обыкновенной кряквы (Anasplatyrhynchos). Этот вид широко распространен в Европе, Азии и Северной Америке.
Птицы (цапли, аисты и др.) крупных и средних размеров, живут главным образом около водоемов или во влажных местах. Похожи на журавлей, но отличаются от них внутренним строением и некоторыми внешними признаками. Хорошо приспособлены к передвижению по земле и к полету. Пища преимущественно животная (позвоночные и беспозвоночные). Гнездятся на деревьях, зданиях, а некоторые на земле. Птенцовые. Перелетные (северные формы) и оседлые (южные формы).
птицы (
FALCONIFORMES)
Птицы (орлы, ястребы, соколы и др.) средних или больших размеров. Прекрасно летают. Глаза большие. Оперение твердое. Клюв загнут, края его — острые, режущие. Четырехпалые ноги с сильными когтями служат для захвата и умерщвления жертвы. Большинство питается живыми млекопитающими или птицами, некоторые потребляют падаль, а мелкие виды — также и насекомых. Непереварившиеся части пищи (шерсть, кости, хитин и др.) отрыгиваются в виде комков — так называемых «погадок».
Гнезда на деревьях, скалах, земле. Птенцовые. Одни виды оседлы, другие — кочуют, третьи — перелетные. Большинство хищных птиц полезны, так как они истребляют грызунов, а мелкие — соколы — и вредных насекомых.
Ночные хищные птицы разной величины. Внешний вид очень характерен: глаза большие; оперение мягкое; когти на ногах и клюв загнутые, острые; края клюва режущие. Летают бесшумно. Едят мышевидных грызунов и мелких млекопитающих, птиц, пресмыкающихся, рыб. Некоторые мелкие виды (например, сыч) питаются также насекомыми. Отрыгивают погадки. Гнезд не строят; откладывают яйца в дупла, на земле, в гнезда других птиц. Птенцовые. Почти все оседлы. Большинство сов очень полезны, так как истребляют много вредных животных (в особенности мышевидных грызунов).
Птицы средней величины, обитатели лесов. Крылья и рулевые перья — длинные. Питаются насекомыми. Ряд видов отряда подбрасывают свои яйца в гнезда мелких воробьиных птиц. Перелетные; места зимовки очень далеки. Птенцовые. Приносят пользу истреблением большого количества вредных насекомых.
Птицы (щурка золотистая, зимородок и др.) небольшой и средней величины, ярко окрашенные, с твердым оперением и сильным слегка изогнутым клювом. Питаются различными животными (насекомыми, рыбой и т. д.). Яйца откладывают в дуплах, норах, расщелинах и т. д. Птенцовые. Перелетные (за исключением зимородка).
Птицы средней величины, приспособленные к лазанью по деревьям. Имеют жесткое оперение, цепкие ноги и долотообразный клюв. Питаются насекомыми, которых достают из-под коры деревьев, а также семенами хвойных деревьев. Яйца откладывают в Дуплах. Птенцовые. Оседлые или кочуют на небольшие расстояния. Истреблением насекомых-вредителей леса приносят пользу.
Птицы (скворцы, воробьи, синицы, дрозды, соловьи, ласточки, вороновые и др.) малой и средней величины. Ноги четырехпалые, сравнительно короткие, первый палец направлен назад. Клюв, как правило, прямой и довольно короткий. Места обитания главным образом связаны с древесной и кустарниковой растительностью. Большая часть воробьиных питаются насекомыми, многие зерноядны. Птенцовые. Многие перелетные. Воробьиные — самый большой отряд птиц, включает около 3/5 всех видов данного класса. В России составляет около половины видов всех птиц. Большинство воробьиных, истребляя вредных насекомых, приносят большую пользу. Однако воробьиные причиняют и некоторый вред, иногда существенный. Ряд видов этого отряда вредит культурным растениям.
Воробьиные и другие птицы могут быть переносчиками блох, клопов, различных клещей (куриных, пастбищных, амбарных и др.), паразитических червей. Установлено, что некоторые птицы (в особенности воробьи) распространяют куриную холеру, оспу и дифтерит домашних птиц. Возможно участие некоторых птиц в распространении ящура и сибирской язвы. Однако следует подчеркнуть, что польза, приносимая птицами, во много раз превосходит причиняемый ими вред, и необходимо всячески способствовать размножению полезных птиц и их охране.
звери (
MAMMALIA, или THERIA)
Млекопитающие, как и птицы, — гомойотермные амниоты, т. е. животные с постоянной температурой тела. Температура тела млекопитающих в среднем ниже, чем у птиц, но в общем высока: у лошади — 38,5 °С, у коровы — 38,5, у свиньи — 39 °С. Их тело покрыто шерстью. Живородящих детенышей выкармливают молоком. Млекопитающие разнообразны по внешнему виду, размеру и массе (от 1,5 г до 150 т). Их объединяют следующие черты организации.
Кожа несет роговые образования: волосы, когти и т. д. Эпидермис многослойный. Окраска обусловлена пигментами. В кори-уме из волокнистой соединительной ткани проходят кровеносные сосуды. В подкожной клетчатке откладывается жир. Волосяной покров играет термоизолирующую роль. Кожные железы многочисленны и разделяются на сальные, потовые, пахучие и млечные.
Позвонки платицельные, между ними имеются хрящевые диски. Череп с крупной мозговой коробкой и уменьшенным количеством костей. У всех млекопитающих есть костное нёбо. Плечевой пояс соединен с осевым скелетом только мышцами и связками. При общем типе строения скелета парных конечностей длина и конфигурация костей меняется в зависимости от типа движения. Высокого развития достигает жевательная мускулатура. Подкожная мускулатура участвует в терморегуляции и общении животных, так как обеспечивает мимику, особенно у хищников и приматов. Грудная и брюшная полости разделены диафрагмой.
Пищеварительный тракт млекопитающих отличается значительной длиной и дифференцировкой отделов. Челюсти с зубами различной формы и назначения. Легкие имеют альвеолярную структуру. Сердце четырехкамерное. Почки метанефрические. Развитие внутриутробное, зародыш питается через плаценту (исключение представляют клоачные). Полушария головного мозга имеют «новую кору» — неопаллиум — из серого мозгового вещества. Новая кора больших полушарий является центром высшей нервной деятельности, координирующим работу других отделов мозга. Органы чувств достаточно высоко развиты, причем обоняние более эффективно, чем у других наземных позвоночных, и развито также у вторичноводных форм. Слуховые органы состоят из трех отделов, включая наружное ухо. Звуки производятся вследствие колебания голосовых связок верхней гортани. Органы зрения (парные глаза) у многих млекопитающих различают цвета. Высокий уровень развития нервной системы и разрешающие способности органов чувств обусловливают сложное поведение, основанное на инстинктивной деятельности и накоплении индивидуального опыта.
Класс млекопитающих делится на 2 подкласса и включает в себя 19 современных отрядов. Общее число видов достигает 4 тыс (рис.104).
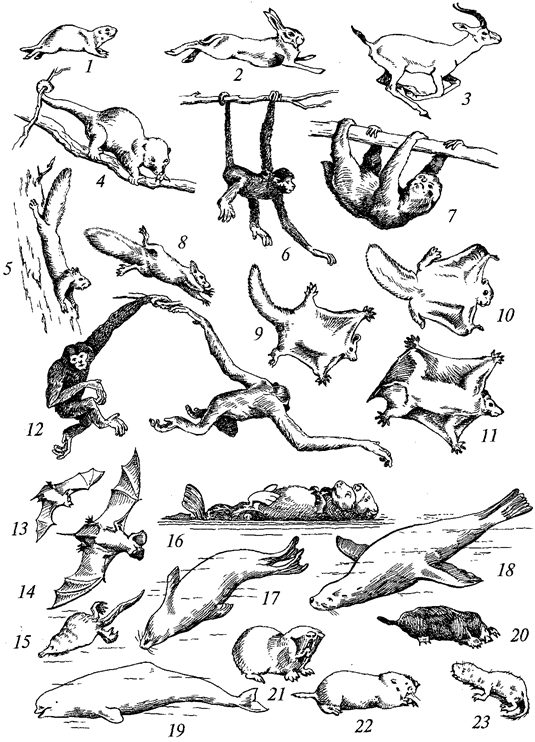
Рис. 104. Форма тела и характер передвижения у разных млекопитающих:
Б е г 1-малый суслик; 2
-заяц русак; 3-
джейран. Лазанье по деревьям- 4 -
кускус (сумчатые); 5 -
белка; 6 -
паукообразная обезьяна; 7 - трехпалый ленивец Прыжки в к р о н ах: 8 -
белка; 9 -
летяга; 10
- сумчатая летяга; // - шерстокрыл; 12 -
гиббон. Полет рукокрылых: 13 - нетопырь карлик; 14 -
широкоухий складчатогуб. Плавание: 15-
выхухоль, /О самка калана с детенышем; 77 - обыкновенный тюлень; 18 -
сивуч; 19 -
белуха. П о д з е м н ы е м л е к о п и т а ю щ и е: 20 -
крот; 21 -
слепыш; 22 -
цокор; 23 —
африканский голый землекоп
Млекопитающие — высший класс позвоночных и всего царства животных. Все системы их органов, в особенности нервная система, достигли наибольшего совершенства. От высших млекопитающих произошел человек. Огромное практическое значение этих животных общеизвестно, одомашненные виды их принадлежат к наиболее важным объектам деятельности работников зоотехнии и ветеринарии. Отдел зоологии, изучающий млекопитающих, называется териологией,
иногда маммологией.
Общая характеристика
Внешний облик млекопитающих говорит об их способности к сложной нервной деятельности, к быстрым и совершенным движениям (см. рис. 104). Тело состоит из головы, туловища, двух пар конечностей (у китообразных и сиреновых задняя пара ног атрофируется) и хвоста (у некоторых видов он отсутствует). Характерны большая голова, подвижное, гибкое туловище и сравнительно длинные ноги.
Покровы.
Кожа хорошо развита и у многих видов достигает большой толщины и прочности, чем и объясняется широкое использование ее для изготовления различных изделий. Кожа состоит, как и у других черепных, из двух слоев — эпидермального и соединительнотканного. Верхние ряды клеток эпидермиса по мере их ороговения постоянно слущиваются.
Для наружных покровов млекопитающих характерно наличие волос, которые у ряда видов (например, у китов) вторично исчезли. Волосы, как и чешуи рептилий и перьев птиц, развиваются из ороговевших клеток эпидермиса. Однако в развитии чешуи и перьев есть много общего, и можно утверждать, что перья произошли из чешуи, волосы же по своему происхождению не имеют отношения к чешуям пресмыкающихся и возникли заново, а не в результате видоизменения чешуи. Волосяной покров отличается рядом превосходных свойств: он легкий, прочный, не ограничивает свободы движения тела, плохо проводит тепло. У животных, обитающих в холодном климате, волосяной покров развит лучше зимой, чем летом. У водных млекопитающих волосы могут совсем редуцироваться, а их роль выполняет толстый слой жира, который хорошо защищает от излишней теплоотдачи и, кроме того, облегчает массу тела. Основания волос связаны с нервными окончаниями, что увеличивает чувствительность млекопитающих к прикосновениям, болевым раздражениям и т. д. У многих млекопитающих на верхней губе развиваются длинные, упругие и особенно чувствительные волосы (вибриссы),
благодаря которым животные могут передвигаться в темноте, не натыкаясь на различные предметы. Волосяной покров защищает кожу от повреждений. У некоторых млекопитающих (например, ежей) волосы превратились в иглы. В этих случаях защитная функция волос выражена особенно сильно.
На конечностях млекопитающих развиваются роговые образования: когти, ногти и копыта. Они служат для защиты от механических повреждений, используются для обороны, нападения и т. д.
На голове у многих млекопитающих имеются рога. У одних видов (например, у быков) они представляют собой полые роговые придатки, сидящие на особых костных выступах черепа. Эти рога, как правило, не сбрасываются. У других видов (например, у оленей) рога — костные образования, покрытые кожей, которая впоследствии может слущиваться. Такие рога обычно развиты у самцов, но иногда бывают и у самок, и ежегодно сбрасываются.
У большинства млекопитающих кожа очень богата железами. Этим они отличаются от современных пресмыкающихся. Различают, как известно, две группы кожных желез млекопитающих — сальные и потовые. Сальные железы имеют гроздевидную форму.
Выделения этих желез возникают в результате перерождения клеток и служат для смазывания волос, придания им прочности и гибкости. На губах, на конъюнктиве (внутренняя поверхность век), около анального отверстия, наружных половых органов и в других местах могут находиться видоизмененные сальные железы, выполняющие другие функции (смазка кожи в местах, подверженных трению; выделение пахучих веществ, служащих для привлечения особей другого пола, и т.д.).
Потовые железы имеют вид закрученных трубочек и выделяют водянистую (иногда густую и окрашенную) жидкость, в которой растворено небольшое количество солей и других веществ. Значение потоотделения для предохранения организма от перегрева общеизвестно. У ряда млекопитающих (слоны, собаки и др.) потовые железы слабо развиты или даже совсем отсутствуют, и защита от перегрева достигается у них другими способами (ночной образ жизни, обливание тела водой, испарение слюны с языка и т.д.). Вместе с потом выделяется и некоторое количество продуктов диссимиляции. Поэтому усиленная работа потовых желез облегчает работу основных органов выделения — почек. К видоизмененным потовым железам относятся железы наружного слухового прохода, выделяющие серу, которая препятствует проникновению насекомых и других посторонних тел.
Млечные железы, столь характерные для всех представителей рассматриваемого класса, тоже представляют собой видоизмененные потовые железы. Молоко содержит все необходимые для нормального развития и роста новорожденных (до перехода их на самостоятельное питание) вещества: белки, углеводы, жиры, витамины, соли и др. Состав молока у различных млекопитающих различен. Особенно велико содержание жира в молоке китообразных — до 45 %. Чем жирнее молоко, тем лучше оно возмещает затраты энергии, значительно возрастающие в условиях холодного климата и особенно в воде. Почти у всех млекопитающих молоко выделяется через соски, число которых различно и в известной степени связано с количеством детенышей в одном помете.
Нервная система.
У млекопитающих по сравнению с другими позвоночными она достигает совершенства: обеспечивает тонкое приспособление к постоянно меняющимся условиям среды, точное согласование работы всех органов, очень интенсивный обмен веществ, разнообразные и очень быстрые движения, отыскание и преследование жертвы, защиту от врагов, сложные взаимоотношения между самцами и самками, совершенную заботу о потомстве.
Сложность строения и величина различных отделов центральной нервной системы, особенно головного мозга млекопитающих, сильно возросли по сравнению с тем, что наблюдалось у высших пресмыкающихся и птиц (рис. 105). Мозжечок в связи с усовершенствованием работы мышечной системы и необходимостью поддержания равновесия тела при быстрых и разнообразных движениях разросся; обычно он состоит из трех частей — червячка и лежащих по бокам его полушарий.
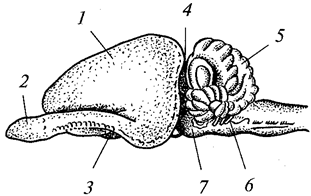
Рис. 105. Мозг кролика сбоку:
1 —
большие полушария; 2 —
обонятельные доли; 3 —
зрительный нерв; 4 —
средний мозг — четверохолмие; 5 —
мозжечок; 6 —
продолговатый мозг; 7 — варолиев мост
Наибольшие изменения произошли в переднем мозге. Он превосходит по величине все остальные отделы головного мозга, вместе взятые, и закрывает сверху и с боков средний мозг. У высших млекопитающих передний мозг настолько разрастается, что сверху закрывает и мозжечок. Характерным отличием переднего мозга млекопитающих является значительное развитие коры,
в которой сосредоточены центры, воспринимающие зрительные, слуховые, осязательные и другие раздражения, двигательные и ассоциативные центры (центры самых сложных проявлений нервной деятельности). Увеличению размеров коры способствует наличие борозд, особенно сильно развитых у высших представителей класса.
Передний мозг лучше, чем у других позвоночных, связанный с различными частями нервной системы, оказывает огромное влияние на работу всего организма. Ни у кого из животных централизация всей нервной деятельности (т. е. подчинение ее переднему мозгу) не выражена так сильно, как у млекопитающих. Именно с корой связана способность животных к приобретению условных рефлексов, установлению новых связей с окружающей средой.
Спинной мозг сильно утолщен; количество нервных клеток и проводящих путей в нем значительно увеличилось. Эти прогрессивные изменения произошли в связи с усовершенствованием мышечной и других систем органов. Возрастает также роль спинного мозга как проводника многочисленных раздражении, идущих от кожи и других органов в головной мозг и, наоборот, от центров мозга — к мышцам и к другим органам.
Органы чувств.
У млекопитающих они сильно развиты. Органы, воспринимающие разнообразные осязательные раздражения, находятся в коже и очень многочисленны (в коже рассеяны и органы, воспринимающие температурные и болевые раздражения). Органы вкуса расположены на языке и в различных частях ротовой полости.
У млекопитающих пища остается во рту значительно дольше, чем у птиц и других позвоночных, и вкусовые раздражения могут быть лучше восприняты. Специализация органов вкуса хорошо выражена: одни воспринимают сладкое, другие — горькое и т. д.
Обоняние у большинства видов очень обострено. Известно, что хищники могут напасть на след жертвы, обоняя ничтожные количества газообразных веществ, исходящих от ее следов.
Зрение млекопитающих уступает в некоторых отношениях (дальность видения, широта зрительного поля) зрению птиц, но превосходит его (в особенности у высших форм) по точности восприятия особенностей предметов (форма, окраска и т.д.).
Органы равновесия, находящиеся, как и у других позвоночных, в лабиринте, хорошо развиты. Для млекопитающих при их быстрых и сложных передвижениях восприятие любых изменений положения тела в пространстве исключительно важно.
Орган слуха имеет чрезвычайно сложное строение и состоит из трех отделов — внутреннего, среднего и наружного. Во всех этих отделах произошли большие изменения. В лабиринте от нижнего мешочка отходит длинная, спирально закрученная улитка, из рептилий наметившаяся только у крокодилов. В улитке находится кор-тиев орган,
напоминающий по своему строению многострунный инструмент: в нем натянуты волоконца различной длины, воспринимающие звуки различной высоты. В среднем отделе уха имеются три слуховые косточки: стремечко, наковальня
и молоточек.
Следует напомнить, что у земноводных (у которых впервые возникло среднее ухо), пресмыкающихся и птиц имеется лишь одна слуховая косточка-столбик, соответствующая стремечку и образовавшаяся из подвеска (гиомандибуляре) рыб. Эмбриологические, сравнительно-анатомические и палеонтологические исследования показали, что наковальня происходит из квадратной кости, которая у земноводных, пресмыкающихся и птиц выполняет роль подвеска, а молоточек — из сочленовной кости. Наличие трех слуховых косточек в среднем ухе млекопитающих является характерным признаком этого класса. Эти косточки, определенным образом сочлененные, значительно улучшают передачу звуков. Для млекопитающих характерно развитие наружного отдела органа слуха, который у птиц слабо развит, у рептилий имеется только в зачаточном состоянии, а у земноводных совсем отсутствует. Этот отдел состоит из длинного наружного слухового прохода, отделенного от среднего уха барабанной перепонкой, и ушной раковины. Последняя служит своеобразным рупором и облегчает животному нахождение источника звука.
Млекопитающим, как и всем животным, свойственны разнообразные врожденные (безусловные) рефлексы. Эти рефлексы обеспечивают глотание, чихание, выделение пищеварительных соков, расширение и сужение кровеносных сосудов, отдергивание частей тела при болевых раздражениях и др. В нервной деятельности млекопитающих большую роль играет сложное врожденное поведение — инстинкты. Одни инстинкты в своей видоспецифичной форме проявляются у всех млекопитающих. Таковы, например, инстинкты, связанные с контактом между полами, инстинкты заботы о потомстве и т. д. Другие инстинкты свойственны лишь определенным видам и возникли в связи с условиями существования, характерными для этих животных. Такова, например, строительная деятельность бобров, сооружающих плотины на ручьях и реках. Строительное искусство бобров инстинктивно, животные ему не обучаются и сознание при этом не проявляется. Если бобра содержать в клетке, то он при наличии хотя бы в какой-то степени подходящих материалов пытается построить плотину в условиях, где она совершенно бесполезна. На ход инстинктивных действий влияет способность к различным действиям, приобретенная в результате развития условных рефлексов.
Однако млекопитающим свойственны и более сложные проявления нервной деятельности, т. е. рассудочной, когда животному «приходится решать задачи» без предварительного обучения (путем приобретения условных рефлексов) на основании установления связей между различными явлениями. Так, например, в помещении, где находилась обезьяна — шимпанзе, высоко подвешивали хорошо известный ей фрукт. Обезьяна прыгала вверх, но достать его не могла. Здесь же были разбросаны ящики, на которые животное, склонное к быстрым прыжкам, раньше могло становиться. Взгромоздившись на ящик, обезьяна добраться до лакомства вновь не могла. И все же задача ею была решена: поставив один ящик на другой, она доставала фрукт. Многие другие опыты подтверждают, что способность к простой рассудочной деятельности проявляется и у других млекопитающих.
Большинству млекопитающих присуща способность издавать различные звуки. У них развился в гортани сложный голосовой аппарат. Способность издавать звуки проявляется у обоих полов и играет большую роль в жизни животных. С помощью звуков они сигнализируют друг другу об опасности, нахождении корма и т. д. Это показатели сложной нервной деятельности.
В нервной деятельности млекопитающих, как и в других физиологических процессах, огромную роль играют различные органы внутренней секреции.
Скелет.
Он претерпел ряд изменений, которые связаны с усложнением нервной, мышечной и других систем органов. Череп млекопитающих отличается прежде всего большими размерами Мозгового отдела, в связи с чем основание его широкое (рис. 106). В черепе ярко выражена тенденция к срастанию отдельных костей.
Рис. 106. Схема черепа млекопитающих (темные — замещающие кости, светлые — покровные кости):
1 — решетчатая; 2 —
глазнично-клиновид-ная; 3 —
крыло-клиновидная; 4 —
передняя клиновидная; 5 — основная клиновидная; 6 — каменистая; 7 — верхняя затылочная; 8 —
боковая затылочная; 9 —
основная затылочная; 10 —
остаток хрящевого черепа (носовая перегородка); 11 —
носовая; 12 —
слезная; 13 —
лобная; 14 —
теменная; 15 —
межтеменная; 16 —
чешуйчатая; 17 —
сошник; 18 —
нёбная; 19 —
крыловидная; 20 —
межчелюстная; 21 —
верхнечелюстная; 22 —
скуловая; 23 —
зубная; 24 —
стремечко; 25 —
наковальня; 26 —
молоточек; 27 —
барабанная кость; 28 —
остатки гиоида; 29 —
подъязычная кость; 30—32 —
хрящи гортани: 30 —
щитовидный, 31 —
черпало-видный, 32 —
перстеневидный хрящ; 33 —
шиловидный отросток — остаток гиоида, прирастающий к височной кости; I
—
IV
—
висцеральные дуги
Четыре затылочные кости почти у всех млекопитающих срастаются в одну. Вокруг лабиринта несколько ушных костей срастаются, образуя одну каменистую кость. Последняя, в свою очередь, соединяется с барабанной костью (развившейся из угловой кости нижней челюсти предков млекопитающих) и большой чешуйчатой костью. У человека в результате слияния этих костей возникает сложная височная кость. В основании черепа лежат две клиновидные кости — основная и передняя. У человека они сливаются в одну. В связи с усложнением органа обоняния развивается решетчатая кость. Интересно, что у зародышей есть зачаток. парасфеноида — кости, развитой только у костных рыб и земноводных. Верх черепа состоит из двух теменных костей (срастающихся у копытных в одну кость) и лобных (у насекомоядных, рукокрылых, обезьян и человека срастающихся в одну). Межтеменная кость срастается обычно с теменными костями или с затылочной. Впереди лобных костей идут носовые кости. Объем черепной коробки в пределах класса сильно меняется в зависимости от величины головного мозга; у низших форм он значительно меньше, чем у высших. Соответственно меняется соотношение размеров мозгового и лицевого отделов черепа.
Рот сверху ограничен верхнечелюстными и межчелюстными костями, которые у обезьян и человека срастаются. Верхнечелюстная кость с помощью скулового отростка сочленяется со скуловой костью, а последняя соединяется со скуловым отростком чешуйчатой кости, в результате чего образуется скуловая дуга млекопитающих.
Твердое нёбо хорошо развито. Оно образовано отростками межчелюстных, верхнечелюстных и нёбных костей. Следовательно, у млекопитающих носовая полость вполне отделена от ротовой, что для этих животных имеет особенно большое значение, так как у них в ротовой полости происходит совершенная механическая обработка пищи. Крыловидные кости у обезьян и человека срастаются с клиновидной.
Нижняя челюсть состоит только из зубной кости, так как сочленовная кость превратилась в молоточек, а угловая — в барабанную. Квадратная кость, подвешивавшая у пресмыкающихся (а также у земноводных) нижнюю челюсть к мозговому отделу черепа, преобразовалась в наковальню. Нижняя челюсть соединяется у млекопитающих с мозговым отделом черепа (с чешуйчатой костью) с помощью сочленовного и венечного отростков. Все кости, ограничивающие ротовое отверстие, хорошо развиты и являются прочной опорой для достигшей большого совершенства зубной системы млекопитающих.
Череп сочленяется с атлантом с помощью двух мыщелков затылочной кости. Этим млекопитающие отличаются от современных пресмыкающихся и птиц, у которых имеется только один мыщелок. Атлант свободно вращается вокруг зубца эпистрофея. Шея у млекопитающих подвижнее, чем у пресмыкающихся. Длина ее различна: у одних (например, у мышей) она очень короткая, у других (например, у верблюдов) очень длинная. Тем не менее почти у всех млекопитающих шейных позвонков семь, что свидетельствует о единстве происхождения этих животных.
Грудная клетка хорошо развита (рис. 107). Она надежно защищает сердце и легкие, в то же время очень подвижна, что важно для обеспечения дыхательных движений. Ребра подвижно соединены с одной стороны с грудными позвонками, с другой — с помощью хрящей — с грудиной. Число ребер и грудных позвонков различно (12—18). Поясничные позвонки массивны, в особенности у крупных животных; число их у разных видов колеблется от 5 до 7. Крестцовые позвонки (их чаще всего 2—4, но у некоторых видов число их доходит до девяти) срастаются в одну кость, к которой прирастают и некоторые хвостовые позвонки. Число хвостовых позвонков весьма различно и определяется длиной хвоста.
Укрепились и подверглись существенным изменениям пояса конечностей. В состав плечевого пояса входят лопатки и ключицы.
Рис. 107. Скелет кролика:
I —
шейные позвонки; 2 —
грудные позвонки; 3 —
поясничные позвонки; 4 —
крестец; 5 —
хвостовые позвонки; 6
— ребра; 7 — рукоятка грудины; 8 —
лопатка; 9 —
акромиальный отросток лопатки; 10 —
коракоидный отросток лопатки; II —
подвздошный отдел безымянной кости; 12 —
седалищный отдел той же кости; 13 —
лобковый отдел той же кости; 14 —
запирательное отверстие; 15 —
плечо; 16 —
локтевая кость; 17 —
лучевая кость; 18 —
запястье; 19 —
пястье; 20 —
бедро; 21 —
коленная чашечка; 22 —
большая берцовая кость; 23 —
малая берцовая кость; 24 —
пяточная кость; 25 —
таранная кость; 26 —
плюсна
Вороньи кости сохраняют самостоятельность только у самых низших форм, а у остальных прирастают к лопаткам и становятся их отростками. Лопатка широкая, на ней развивается гребень, заканчивающийся акромиальным отростком. Ключицы у большинства видов очень хорошо развиты, но у копытных, сирен, ряда хищников и китообразных они отсутствуют.
Таз (за исключением тех млекопитающих, у которых атрофировались задние конечности) хорошо развит. Подвздошные кости удлинились и направлены вперед, седалищные разрослись назад, а лобковые малы. В результате этих изменений таз млекопитающих имеет наклонное положение и задняя часть его направлена вниз и назад. Вертлуг находится сзади крестца, что связано с переходом от ползания к хождению и бегу. У взрослых животных кости таза обычно срастаются в одну безымянную кость.
Проксимальные кости ног (плечевая, лучевая, локтевая, бедренная, большая и малая берцовые), как правило, длинные. У ряда видов локтевая и малая берцовая кости могут подвергаться редукции. Очень характерно присутствие коленной чашечки. В запястье — девять костей (четвертая и пятая дистальные кости срастаются). В предплюсне хорошо развиты две. Проксимальные кости — пяточная и таранная. В голеностопном сочленении участвуют большая берцовая и таранная кости. Количество пальцев у большинства млекопитающих полное — по 5 на каждой ноге. Однако у копытных млекопитающих число пальцев редуцируется до трех, двух и одного. Скелеты конечностей претерпевают сильные изменения в связи с особенностями образа жизни.
Мышечная система.
Она более развита и дифференцированна, чем у пресмыкающихся. Особенно сильно развиты мышцы, связанные с конечностями: двуглавая, гребенчатая, лобково-бедрен-ная, грушевидная, ягодичные, многочисленные разгибатели и сгибатели пальцев и т. д. Характерно также развитие широкой мышцы спины, трапециевидной, большой и малой грудных мышц и зубчатых, мышц шеи и т. д.
В связи с усовершенствованием механизма дыхания у млекопитающих развилась мускульная диафрагма, отделяющая грудную полость от брюшной. Она вдается в грудную полость в виде купола. При вдохе уплощается, в результате чего объем грудной клетки увеличивается за счет брюшной полости. Мышечная система претерпевает существенные изменения в зависимости от образа жизни различных видов млекопитающих.
Пищеварительная система.
Она претерпела также ряд изменений (рис. 108).
Рот у всех млекопитающих, за исключением самых низших и китообразных, окружен мягкими губами, которые играют существенную роль при захвате пищи. Зубы очень прочные, строение их сложное. Они получают питание от густой сети кровеносных сосудов, связаны с нервами и крепко сидят в ячейках челюстей — альвеолах. Такие зубы заменяют большое количество примитивных зубов, свойственных рептилиям и другим позвоночным, число их характерно для каждого вида. Так, у собаки 42 зуба, у лошади — 40, у свиньи — 44, у коровы — 32. У рептилий и ниже их стоящих позвоночных зубы служат в основном для захвата и удержания добычи, и строение их сходно. У млекопитающих зубы подразделяются на резцы, клыки, малые и большие коренные зубы. Их строение и форма зависят от выполняемой ими работы (функции). Резцы,
служащие для откусывания пищи, плоские, с острым режущим верхним краем; клыки,
служащие для разрывания добычи, конические, с заостренным концом; коренные зубы,
служащие для размалывания и перетирания пищи, имеют широкую верхнюю поверхность с бугорками.
У млекопитающих лучше, чем у других наземных позвоночных, развиты железы ротовой полости, к которым относятся множество мелких слизистых желез, расположенных на языке и внутренней поверхности щек, нёба, губ, и крупные слюнные — подъязычная, заднеязычная, подчелюстная и околоушная. Первые три развились из подъязычной железы пресмыкающихся, а последняя присуща только млекопитающим и развилась из щечных желез.
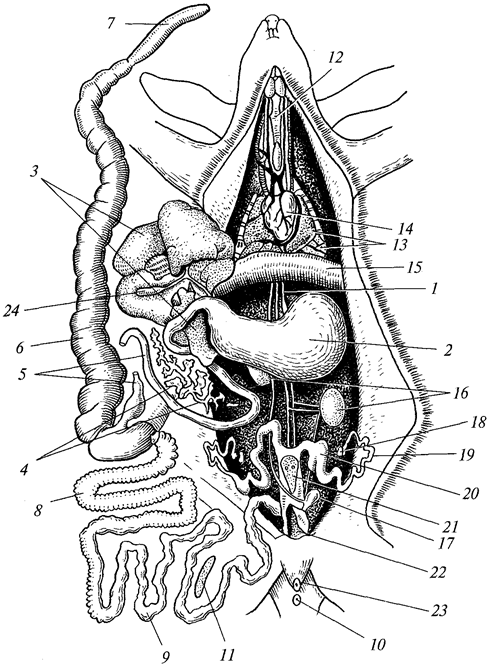
Рис. 108. Вскрытый кролик:
1 -
пищевод; 2 -
желудок; 3 -
печень; 4 -
поджелудочная железа; 5 - тонкая кишка; б-
слепая кишка; 7- червеобразный отросток; 8-
толстая кишка; 9-
прямая кишка; 10 —
заднепроходное отверстие; 11 —
селезенка; 12 —
трахея: 13 -
легкие; 14 -
сердце; 15 —
диафрагма; 16 -
почки; 17 -
мочевой пузырь; 18 -
яичник; 19 -
фаллопиева труба; 20 -
матка; 21 -
влагалище; 22 -
мочеполовой синус; 23 —
мочеполовое отверстие; 24 —
желчный пузырь
Хорошее развитие ротовой полости, полная изоляция ее от носовой, совершенная зубная система, мощные жевательные мышцы, большой мускулистый язык, слюнные и другие ротовые железы делают возможным тщательную механическую обработку пищи, а также отчасти химическую.
Пища, раздробленная, разжеванная и обильно смоченная слюной, проходит через глотку и пищевод в желудок. Последний хорошо развит у всех млекопитающих и представляет собой объемистый мешок с мускулистыми стенками. В желудке завершается механическая обработка пищи и начинается (как и у других позвоночных) под влиянием сока желез слизистой оболочки (в нем содержатся фермент пепсин и соляная кислота) распад белков на более простые азотистые соединения. Особенно сложно устроен желудок жвачных, в котором происходит обработка жестких растительных кормов.
Из всех позвоночных млекопитающие имеют самый длинный кишечник, что облегчает переваривание разнообразной пищи, особенно растительной. Даже у тех млекопитающих, которые питаются животной пищей, длина кишечника значительно превосходит общую длину тела, а у растительноядных она огромна (например, у лошади отношение длины кишечника к длине тела 12:1, у коровы — 20:1). Печень велика. Ее многообразные функции были указаны раньше. Желчный пузырь, хорошо развитый у большинства млекопитающих, у некоторых видов (например, у мышей, крыс и др.) отсутствует. Поджелудочная железа сильно развита и играет важнейшую роль в пищеварении (как отмечалось выше, в ее соке имеются ферменты, способствующие перевариванию белков, жиров и углеводов). Тонкий кишечник очень длинный. Он начинается двенадцатиперстной кишкой и разделяется на несколько отделов. Внутренняя поверхность тонких кишок имеет многочисленные выросты — ворсинки, внутри которых проходят кровеносные и лимфатические сосуды. Благодаря ворсинкам поверхность тонкого кишечника увеличивается, что способствует выделению большего количества пищеварительных соков и ускоряет всасывание переваренной пищи.
Дыхательная система.
У млекопитающих она сильно усложнилась по сравнению с их предками — пресмыкающимися. Воздух, проходя через носовую полость, которая имеет чрезвычайно сложное строение, нагревается и в значительной степени очищается от различных твердых примесей. Благодаря развитию твердого и мягкого нёба воздухоносные пути отделены полностью от ротовой полости и воздух все время беспрепятственно поступает в организм. Длинная трахея разделяется на два бронха, которые входят в легкие и распадаются на множество все более тонких трубочек — бронхов и бронхиол. Последние заканчиваются тонкостенными воздушными мешочками, на стенках которых имеются многочисленные легочные пузырьки — альвеолы.
Эти пузырьки густо оплетены кровеносными капиллярами. Количество альвеол велико и измеряется миллионами. Следовательно, поверхность соприкосновения их с капиллярами, через которую происходит обмен газами, весьма обширна. Дыхательная поверхность легких пресмыкающихся в несколько раз меньше.
Легкие млекопитающих по размерам превосходят те же органы птиц и в отличие от них не прирастают к внутренним стенкам грудной клетки. Покрывающая их плевра только соприкасается с плеврой, покрывающей изнутри грудную клетку. Между обоими листками плевры воздуха нет;
поэтому при вдохе легкие, давление воздуха внутри которых равно атмосферному, увеличиваются в объеме. При выдохе легкие сжимаются. Изменение объема грудной клетки осуществляется быстро благодаря хорошо развитым мышцам, поднимающим ребра, и диафрагме. Кожное дыхание у млекопитающих вследствие ороговения верхнего слоя кожи существенной роли не играет; через нее поглощается около 1 % кислорода, потребляемого этими животными. Незначительное количество кислорода поступает в организм млекопитающих и через слизистую оболочку верхних дыхательных путей.
Кровеносная система.
У представителей этого класса сердце четырехкамерное (рис. 109).
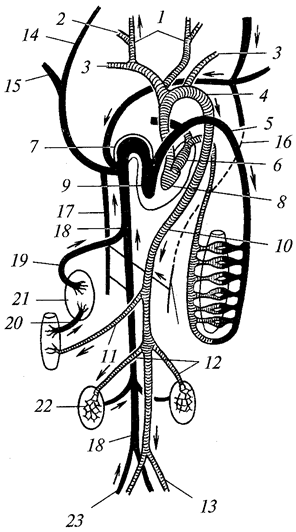
Рис. 109. Схема кровеносной системы млекопитающих:
/ и 2 —
наружная и внутренняя сонные артерии; 3 —
подключичная артерия; 4 —
левая дуга аорты; 5 — легочная артерия; 6 —
левое предсердие; 7 — правое предсердие; 8 —
левый желудочек; 9 —
правый желудочек; 10 —
спинная аорта; 11 —
внутренностная артерия; 12 —
почечная артерия; 13 —
подвздошная артерия; 14 —
яремная вена; 75 — подключичная вена; 16 —
левая непарная вена; 17 —
правая непарная вена; 18 —
задняя полая вена; 19 —
печеночная вена; 20 —
воротная вена печени; 21 —
печень; 22
~
почка; 23 —
подвздошная вена
Из двух дуг аорты у млекопитающих осталась только одна, но не правая, как у птиц, а левая. Отходит она от левого желудочка и несет чисто артериальную кровь. Следовательно, у млекопитающих, как и у птиц, артериальный ток крови полностью отделен от венозного, и все части тела получают кровь, богатую кислородом. Размеры сердца по отношению к величине тела больше, чем у пресмыкающихся. Частота сокращений сердца хотя и меньше, чем у птиц, но значительно больше, чем у пресмыкающихся. В связи с большой работой, выполняемой сердцем, сосуды (венечные), питающие его, сильно развиты. Нервная регуляция сердечной деятельности, как и всей кровеносной системы, очень совершенна. Сеть кровеносных сосудов, особенно капилляров, у млекопитающих более разветвленная, чем у их предков.
Очень характерно строение эритроцитов. Они, как правило, очень малы (у человека диаметр эритроцита 7 мкм), лишены ядер (хотя образуются из клеток, имеющих ядро) и имеют двояковогнутую форму. Количество их в 1 мм3
крови измеряется миллионами. Благодаря этому поверхность эритроцитов велика и они поглощают огромное количество кислорода.
Кровь млекопитающих быстрее свертывается, чем кровь пой-килотермных животных, — это имеет важное значение при различных ранениях, которым часто подвергаются млекопитающие при их активном образе жизни.
Кровеносная система тесно связана с хорошо развитой лимфатической. Лимфа постепенно собирается в лимфатические сосуды, а самый крупный сосуд этой системы впадает в нижнюю полую вену.
Выделительная система.
В связи с высокой интенсивностью обмена веществ (в результате чего образуется большое количество продуктов диссимиляции) выделительная система значительно сложнее, чем у рептилий (см. рис. 108). Почки (большие тела, лежащие у позвоночного столба в его поясничной части), как и у пресмыкающихся, тазовые, но строение их отличается рядом прогрессивных особенностей. Количество кровеносных клубочков увеличилось, почечные канальцы значительно удлинились, поверхность соприкосновения их с кровеносными сосудами сильно возросла. Отмеченные особенности почек обусловливают более совершенное удаление из организма продуктов диссимиляции и в то же время сохранение в организме воды и других необходимых организму веществ.
Моча по мочеточникам стекает в мочевой пузырь, а оттуда периодически удаляется из организма через мочеиспускательный канал. У самцов последний пронизывает совокупительный орган. У самок мочеиспускательный канал короткий и открывается около отверстия влагалища. У рептилий удаление мочи происходило через заднюю часть кишечника — клоаку; у млекопитающих совершилось полное отделение мочевыводящих путей от кишки.
Часть продуктов диссимиляции выделяется с потом через кожу. Поэтому закупорка всех или значительной части выводных протоков потовых желез может привести к отравлению организма.
Размножение.
Семенники у многих видов временно (в период размножения) или навсегда выходят из брюшной полости и находятся в особом мешке — мошонке. Вследствие этого семяпроводы (бывшие вольфовы каналы) петлеобразно изогнуты. Они открываются в основание совокупительного органа (мужского члена), через единственный канал которого выводятся моча и семя (у пресмыкающихся совокупительные органы, находящиеся в клоаке, служат только для выделения семени). В совокупительном органе ряда млекопитающих (например, у многих хищных) имеются косточки, форма которых даже у близких видов может быть различна. Хорошо развиты добавочные железы: семенные пузырьки, куперовы железы, предстательная железа, окружающая начало мочеиспускательного канала. Выделения этих желез имеют большое значение для поддержания нормального состояния семени и его способности к оплодотворению.
В женской половой системе млекопитающих произошли большие изменения в связи с тем, что самки млекопитающих рождают вполне сформированных детенышей. Яичники лежат в брюшной полости недалеко от воронок яйцеводов (мюллеровых каналов). Средние отделы женских половых труб превратились в матки, в которых происходит развитие зародышей. Для обеспечения питания зародышей стенки матки обильно снабжаются кровью. Конечные части женских половых путей отделены от задней кишки и мочеиспускательного канала.
У живородящих млекопитающих, за исключением сумчатых, дистальные части женских половых труб полностью сливаются и образуют влагалище. Процесс слияния постепенно распространяется и на средние отделы женских половых труб — матки. У одних млекопитающих (большинства грызунов, слонов и др.) обе матки еще вполне самостоятельны (двойная матка),
у других (некоторых грызунов, свиней и др.) происходит слияние дистальных частей и образуется двураздельная матка,
у третьих (насекомоядных, большинства хищных, китов, большинства копытных и др.) сливаются средние части и образуется двурогая матка.
Наконец, у многих летучих мышей, обезьян и человека слияние маток происходит полностью и образуется простая матка.
Яйцеводы (или фаллопиевы трубы) остаются самостоятельными.
Одни млекопитающие полигамны:
единственный самец, являющийся вожаком стада, оплодотворяет всех самок стада; другие млекопитающие моногамны,
они живут парами, которые существуют обычно в течение только одного сезона, а у некоторых видов — в течение большей части жизни.
Современные млекопитающие (за исключением клоачных — они откладывают яйца) — живородящие. Низшие представители класса млекопитающих (сумчатые) рождают слаборазвитых детенышей. Остальные же, очень многочисленные млекопитающие (плацентарные), рождаются во вполне сформированном состоянии, что обеспечивается существенными изменениями зародышевых оболочек, а также полового аппарата и всего материнского организма.
Одни млекопитающие сразу после рождения способны к самостоятельному передвижению, другие рождаются слепыми и беспомощными, но во всех случаях детеныши выкармливаются молоком и находятся под защитой родителей. В связи с высоким развитием нервной системы забота о потомстве у млекопитающих очень совершенна. Поэтому представители этого класса дают меньше потомства, чем пресмыкающиеся, а тем более земноводные и рыбы. Крупные млекопитающие рождают по одному детенышу.
Происхождение.
Самые примитивные млекопитающие появились в начале мезозойской эры — в триасе. Их предками были хищные пресмыкающиеся — циноодонты
(рис. 110).
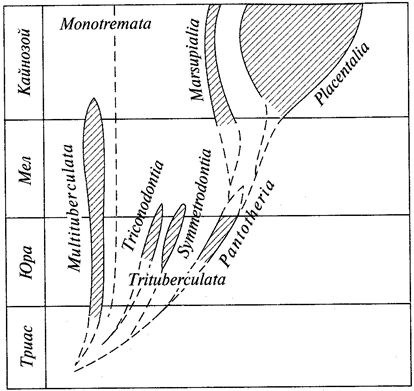
Рис. 110. Филогенетическое древо млекопитающих
Останки этих рептилий были обнаружены в разных местах земного шара. В их изучение большой вклад внесли советские ученые П.П.Сушкин, Л. П. Татаринов и др. У териодонтов был ряд особенностей, приближающих их к млекопитающим: увеличенный мозговой отдел черепа, что свидетельствовало о прогрессивном развитии головного мозга; сравнительно длинные ноги, сочленяющиеся с туловищем таким образом, что это содействовало более быстрому передвижению, чем это было свойственно другим рептилиям; нижняя челюсть состояла из большой зубной кости, которая у некоторых териодонтов сочленялась непосредственно с мозговым отделом черепа при сохранении обычного сочленения этой челюсти при помощи сочленовной и квадратной костей; уменьшение последних двух костей в связи с началом превращения их в слуховые косточки (молоточек и наковальню); наметившаяся дифференциация зубов на резцы, клыки и коренные, которые сидели в ячейках, или альвеолах, и др.
У териодонтов, вероятно, сохранились некоторые признаки земноводных (два затылочных мыщелка, кожа, богатая железами, и др.), которые характерны и для млекопитающих. У некоторых териодонтов имелись вибриссы, т. е. зачатки волосяного покрова.
Систематический обзор
Современные млекопитающие принадлежат к двум подклассам:
клоачные, или первозвери, и живородящие млекопитающие, или настоящие звери.
У первозверей наряду с характерными для млекопитающих признаками (наружный покров волосяной или иглы, млечные железы, нижняя челюсть, состоящая только из одной кости — зубной, два затылочных мыщелка, твердое нёбо и др.) имеются признаки, свойственные рептилиям (клоака, куда открываются мочеточники, яйцеводы и семяпроводы, размножение путем от-кладки яиц, ряд особенностей скелета — отсутствие срастания вороньей и других костей с лопаткой и т. п.). Кроме того, имеющиеся у клоачных признаки млекопитающих выражены слабее, чем у двух других подклассов: более примитивный мозг; менее развитая двигательная система, низкая средняя температура тела (32 °С) и сильные колебания ее (от 26 до 35 °С), что свидетельствует о недостаточном развитии всех систем, обеспечивающих высокую постоянную температуру тела; отсутствие сосков и т. д. В подкласс входит один современный отряд — однопроходные (Monotremata).
К клоачным относятся только четыре вида: один вид утконоса (Австралия, о. Тасмания) и три вида ехидн (Австралия, о. Тасмания, о. Новая Гвинея). Утконос
хорошо приспособлен к обитанию в воде. Его тело покрыто густой несмачиваемой шерстью, между пальцами имеются плавательные перепонки, голова заканчивается широким роговым клювом, напоминающим утиный, через который животное процеживает воду при добывании разных беспозвоночных. Зубы (схожие с зубами млекопитающих) имеются только у молодых животных и потом рассасываются и заменяются роговыми пластинками. Ехидны
ведут наземный образ жизни, тело их покрыто иглами. Они беззубы, имеют вытянутое рыло и длинный червеобразный язык. Питаются термитами и другими насекомыми, пристающими к липкому языку, который ехидна просовывает в их гнезда.
Клоачные откладывают яйца, окруженные пергаментообразной скорлупой. Они богаты питательными веществами и поэтому достигают большой величины (1—1,5 см) в отличие от мелких зигот живородящих млекопитающих. У ехидн яйца развиваются в особой сумке на брюхе самки, утконосы насиживают яйца в гнездах. Вылупившихся из яиц детенышей клоачные выкармливают молоком, которое выделяется через поры на брюшной стенке.
Клоачные — самые древние млекопитающие, первые представители которых появились еще в триасе и были потом вытеснены более развитыми представителями этого класса. Ехидны и утконос сохранились в Австралии и на указанных островах потому, что на этих территориях в результате давней изоляции их от главных материков никогда не было настоящих зверей. Кроме того, их выживанию способствовали приспособления к специальным видам питания, описанным выше.
Включает один отряд Сумчатые (Marsupialia). Эти млекопитающие по своей организации стоят выше клоачных. Нервная, мышечная, кровеносная и другие системы у них развиты лучше. Температура тела более высокая (до 37 °С) и подвержена меньшим колебаниям, чем у клоачных. Половая система в связи с живорождением (хотя и примитивным) тоже усложнилась, и мочеполовые пути отделены от заднего отдела кишечника, т. е. клоака у них отсутствует. Молоко выделяется через соски.
Мужская половая система близка к таковой высших млекопитающих. Совокупительный орган служит не только для выведения семени, но и мочи. Средние части женских половых труб превратились в матки, где совершается развитие зародышей, а задние части стали влагалищами. Следовательно, у низших зверей в отличие от высших млекопитающих два влагалища (у некоторых видов образуется даже третье). Соответственно этому совокупительный орган самцов бывает раздвоенным на конце.
У большинства сумчатых плацента не развивается, а у остальных видов она очень примитивна. Поэтому зародыши не получают в матке достаточного питания и детеныши рождаются малоразвитыми и очень маленькими. Они самостоятельно выбираются из матки и прикрепляются к соскам, но некоторое время не могут самостоятельно сосать, молоко поступает в их кишечник благодаря сокращению мышц сосков. Подросших детенышей мать носит некоторое время на спине. Однако у большинства видов самки имеют на брюхе сумку, куда новорожденные перебираются самостоятельно и в которой их постепенное развитие до вполне оформленного состояния длится значительно дольше, чем в матке. Так, например, у гигантского кенгуру, высота которого около 2 м, зародыш находится в матке 39 дней, а в сумке — более 7 месяцев. Длина зародыша при выходе его из матки не более 25 мм. Число детенышей у разных видов колеблется от одного до 7—8 и более, но количество сосков у многоплодных видов меньше, чем новорожденных, и те из них, которые не успели прикрепиться к соскам, погибают.
Сумчатых нельзя рассматривать как непосредственных предков плацентарных, но по своей организации они близко стоят к тем вымершим первичным живородящим млекопитающим, которые дали начало плацентарным.
Плацентарные распространены во всех частях света, за исключением Австралии. К плацентарным относятся все домашние млекопитающие. От высшей группы плацентарных — обезьян — произошел человек. В инфраклассе 17—18 современных отрядов.
У высших млекопитающих по сравнению с двумя ранее рассмотренными группами — клоачными и сумчатыми — усложнилась вся организация. Именно у плацентарных животных достигла наивысшего развития нервная система, особенно кора переднего мозга и связанная с этим высокая способность адаптации к меняющимся условиям внешней среды и зачатки рассудочной нервной деятельности. То же самое следует сказать о системе органов передвижения, о дыхательной, кровеносной, пищеварительной и других системах. Температура тела плацентарных значительно выше, чем у клоачных и сумчатых. Она поддерживается, как правило, на одном уровне, что говорит о высокой интенсивности обмена веществ и сложной терморегуляции. Характерная для плацентарных наиболее совершенная форма живорождения обеспечивает их эмбриональное развитие в наиболее благоприятных условиях. Забота о потомстве (выкармливание молоком, защита от врагов и т. д.) развита у высших млекопитающих лучше, чем у кого-либо из животных.
Предками плацентарных млекопитающих были так называемые пантотерии —
мелкие животные с трехбугорчатыми зубами, существовавшие в триасе и юрском периоде. Первые плацентарные млекопитающие появились в конце или, возможно, в середине мелового периода. Это были примитивные насекомоядные животные. От них в начале третичного периода произошли различные отряды рассматриваемого подкласса, которые быстро эволюционировали в течение всего названного периода в разных направлениях, осваивая различные места обитания. Во время эволюции отдельные из появившихся групп млекопитающих вымирали и им на смену приходили новые. Ниже приведены краткие характеристики большинства современных отрядов плацентарных млекопитающих.
К этому отряду относятся низшие из современных плацентарных млекопитающих (кроты, ежи, землеройки, выхухоли и др.). Это небольшие или мелкие животные с характерным вытянутым хоботком. Передний мозг небольшой, с развитыми обонятельными долями, не имеет извилин, его полушария не закрывают мозжечка, а у некоторых даже виден средний мозг. Соответственно и черепная коробка мала. Зубы слабо дифферен-цированны. Многие питаются насекомыми, но ряд видов может нападать на мелких позвоночных. Некоторые виды имеют промысловое значение. Ряд видов приносит пользу, уничтожая вредных насекомых. Могут причинять и вред (например, крот поедает много таких полезных животных, как дождевые черви). Количество видов — около 370.
Млекопитающие, приспособленные к планированию на небольшое расстояние с помощью покрытой шерстью перепонки, натянутой между шеей, боком туловища, конечностями и хвостом. Сохранилось только два вида, распространенных во влажных лесах Юго-Восточной Азии. Шерстокрылы интересны в том отношении, что от подобных животных, вероятно, произошли настоящие летающие млекопитающие — рукокрылые.
Млекопитающие, приспособленные к длительному полету. Пальцы передних конечностей сильно удлинены и между ними (за исключением первого), боком тела, частью задних конечностей и хвостом натянута летательная перепонка. В связи с приспособлением к полету у летучих мышей, как и у птиц, весь скелет облегчен, кости черепа срослись, а на грудной кости развился киль, к которому прикрепляются сильные грудные мышцы. Ушные раковины хорошо развиты. Обонятельные доли велики, но передний мозг небольшой и не прикрывает мозжечка. Зубы мелкие, всех родов, верхние резцы иногда редуцируются. Многие питаются насекомыми. Значительное количество видов питаются плодами или всеядны. Несколько видов летучих мышей (в Южной Америке) прокалывают резцами кожу млекопитающих (главным образом копытных) и сосут кровь. Истреблением вредных насекомых летучие мыши приносят большую пользу. Известно около 850 видов, большинство которых обитает в теплых странах.
Малочисленный отряд (около 60 видов), который до недавнего времени рассматривался как подотряд грызунов. Животные небольшой (зайцы, кролики) и малой величины (пищухи, или сеноставки), как правило, с сильно развитыми ушами, длинными задними ногами и очень коротким хвостом, растительноядны. На верхней челюсти две пары резцов — очень длинные средние и короткие боковые, на нижней челюсти — только два длинных средних резца. Клыков нет, между резцами и коренными зубами большой промежуток — диастема. Резцы и остальные зубы без замкнутых корней и поэтому могут непрерывно расти, возмещая быстро стачиваемые коронки. Костное нёбо в виде узкого мостика между обоими рядами зубов. В передней части желудка происходит лишь брожение корма, в задней — начинается его переваривание. Кишечник очень длинный, со слепой кишкой. Очень плодовиты. Зайцы и кролики широко распространены на земном шаре.
Большое значение кроликов и зайцев как мясных и пушных объектов общеизвестно. Однако при сильном размножении они могут причинять большой вред полезным растениям, а также быть источником некоторых серьезных инфекционных заболеваний. Разнообразные породы домашних кроликов происходят от дикого кролика (Oryctolaguscuniculus), который распространен в Северной Африке, Западной и Средней Европе и завезен в прошлом столетии на юг Украины.
Самый большой отряд плацентарных, насчитывающий около 2500 видов (мыши, крысы, белки, тушканчики, летяги, песчанки, хомяки, полевки, дикобразы, водосвинки, нутрии, бобры, слепыши и др.). Обитают в самых разных условиях, некоторые хорошо приспособлены к древесному образу жизни, к планирующему полету, к жизни в воде, под землей и т.д. Большинство видов — мелкие животные, но есть и довольно крупные. Очень многие живут в норах. Как правило, растительноядные, но есть и всеядные. Самый характерный признак — чрезвычайно развитые резцы (два на верхней челюсти и два на нижней), отделенные большой диастемой от коренных зубов. Клыков нет. Корни резцов, а у большинства видов и корни остальных зубов незамкнутые, благодаря чему зубы могут постоянно расти. Костное нёбо в виде широкой пластинки, расположенной между обоими рядами зубов в верхней челюсти. Кишечник очень длинный. Очень плодовиты. Шкурки и мясо ряда видов используются человеком. Есть виды (мышовки и др.), поедающие много насекомых. Некоторые виды причиняют большой вред, поедая полезные растения и разные пищевые продукты. Ряд видов — переносчики опаснейших заболеваний (туляремии, чумы и т.д.).
Сильные животные, преимущественно средних и больших размеров, питающиеся, как правило, позвоночными животными. К этому отряду относятся семейства: собачьи, медвежьи, енотовые, куньи, виверровые, гиеновые, кошачьи. Передний мозг сильно развит и у многих видов покрыт бороздами. Крепкие ноги вооружены острыми когтями. Почти у всех видов семейства кошачьих когти втяжные. Клыки очень велики. Последний ложнокоренной зуб на верхней челюсти и первый коренной на нижней челюсти у большинства хищников сильно выдаются и имеют высокие и острые бугры; эти зубы, называемые хищными,
служат для разгрызания костей, сухожилий и т. д. Жевательная мускулатура очень хорошо развита, в связи с чем на черепе образовались гребни, служащие для ее прикрепления. Питаются в основном животной пищей, но даже типичные хищники едят и растения, а некоторые виды (медведи, барсуки) живут главным образом за счет растительных продуктов. Польза, приносимая хищниками, значительна: они истребляют вредных грызунов, многие виды имеют большое промысловое значение.
Человек приручил некоторых хищников. Первым домашним животным была собака. Собаки, по-видимому, возникли от скрещивания волков (Canislupus) и шакалов (Canisaureus) и последующего искусственного отбора в разных направлениях. В настоящее время существует около 350 пород и отродий (видоизменений пород) собак. Собаки легко скрещиваются с волками и шакалами и дают плодовитое потомство.
Домашняя кошка происходит от дикой буланой (степной) кош-| ки (Felislybica), обитающей в Африке, Передней, Средней и Цент-Ц ральной Азии. Впервые кошка была приручена в Египте.
Хищники причиняют и вред. Крупные хищники могут нападать на домашний скот и человека. Мелкие представители этого отряда могут истреблять домашних птиц. Некоторые хищники семейства собачьих являются хозяевами паразитических червей (ленточных, круглых и др.), которые живут в организме домашних животных и человека. Волки и другие хищники при укусах могут передать вирус бешенства человеку и животным. Однако поскольку вредные хищники приносят и значительную пользу, то не может быть и речи об их полном истреблении, наоборот, во многих районах они нуждаются в охране.
Современные хищники произошли от древних хищных млекопитающих — креодонтов,
впоследствии вымерших. Предками же креодонтов были примитивные насекомоядные. В отряде около 240 видов.
Крупные млекопитающие, приспособленные к длительному пребыванию в воде (в морях и некоторых больших озерах) и плохо передвигающиеся на суше. Отряд включает 31 вид. К ним относятся моржи, ушастые тюлени (котики и др.) и настоящие тюлени. Ластоногие произошли от сухопутных хищников, с которыми их до сих пор иногда объединяют в один отряд. В связи с приспособлением к жизни в воде форма тела ластоногих обтекаемая, голова округлая, лишенная ушных раковин или с зачатками их, шерсть короткая, жесткая (у котиков — с мягким подшерстком), у взрослых моржей — совсем рудиментарная. Конечности превратились в ласты, пальцы соединены плавательной перепонкой. У моржей и ушастых тюленей задние конечности еще могут подгибаться, у тюленей же они постоянно обращены назад и расположены вдоль хвоста (короткого у ластоногих). Подкожный жировой слой очень толстый, защищает животное от воздействия низких температур (сохраняет тепло) и облегчает массу тела. У некоторых видов имеются воздушные мешки, ответвляющиеся от трахеи или даже от пищевода; они дают возможность иметь запас воздуха при нырянии и уменьшают массу тела. Передний мозг хорошо развит. Глаза большие, с шаровидным хрусталиком, что дает возможность видеть в воде. Зубная система в связи с тем, что в воде жевать нельзя, упростилась: зубы однородные, число их увеличилось, они служат только для захва-тывания добычи. У моржей верхние клыки превратились в мощные бивни, при помощи которых эти животные выкапывают со дна моллюсков. Питаются ластоногие рыбой и различными беспозвоночными (моллюсками и др.). Размножение происходит на суше. Ластоногие — большей частью стадные животные.
Ластоногих добывают из-за мяса, жира, шкур и меха. Очень ценный мех дают котики. Промысел ластоногих везде ограничивается, однако их численность сокращается. Ряд видов занесены в Красные книги.
Крупные и гигантской величины млекопитающие, которые всю жизнь проводят в воде. В связи с этим они отличаются от сухопутных млекопитающих еще больше, чем ластоногие. Китообразные произошли от древних наземных хищных млекопитающих, близких к креодонтам. Насчитывается 80 видов. Форма тела — рыбообразная, шея не выражена, голова очень велика (у больших китов длина ее превышает 1/3 длины всего тела). Передние конечности превратились в огромные ласты, в скелете которых увеличилось количество фаланг пальцев. Задние конечности, бесполезные при плавании, редуцировались — сохранились только остатки таза. Тело заканчивается мощным хвостом вторичного происхождения, который не имеет позвонков, а состоит только из мышц, соединительной ткани и жира. У многих видов на спине развивается непарный плавник. Кожа лишена волосяного покрова, за исключением отдельных волос на челюстях. Подкожный жировой слой достигает 50 см в толщину и играет ту же роль, что и у ластоногих. Скелет имеет губчатое строение, в костях много жира, что сильно уменьшает массу тела. Глаза малы, слезные железы и слезный канал отсутствуют. Ушных раковин нет, орган слуха сильно редуцирован. Носовые раковины тоже редуцированы. Головной мозг по абсолютной массе велик (у гигантских форм до 7 кг), но по отношению к массе тела мал (у синего кита это отношение равно 1:14000). Обонятельные доли отсутствуют. Носовые отверстия находятся вверху и соединяются особым каналом с гортанью, так что вода во время захвата пищи проникнуть в легкие не может.
Фонтаны, поднимающиеся из носовых отверстий у полярных китов, образуются струёй выдыхаемого влажного воздуха, которая может подхватывать и брызги воды. У подотряда зубатых китов (кашалоты, дельфины, касатки и др.) зубы или упрощены и однотипны, или же, как у подотряда беззубых, или усатых, китов (серые киты, синие киты, финвалы, сейвалы и др.), совсем редуцированы. Вместо зубов у последних в гигантской ротовой полости свешиваются вниз многочисленные (300—400 шт.) роговые пластины (китовый ус) до 5 м в длину. Нижние края их образуют сито, задерживающее массу мелких водных беспозвоночных (моллюсков, раков и др.). Переход таких исполинских животных к питанию мелкими организмами понятен: большая добыча в океанах встречается сравнительно редко, планктонных же организмов, в особенности в полярных широтах, очень много, а китам нужно огромное количество пищи. Зубатые киты питаются преимущественно рыбой. Массовое истребление китов привело к тому, что эта интереснейшая группа оказалась на грани полного уничтожения. Поэтому в последнее время достигнуты международные соглашения о запрещении охоты на китов.
Небольшие животные величиной с зайца, обитающие главным образом в скалистых местностях на Аравийском полуострове и в Африке. Ноги (передние — четырехпалые, задние — трехпалые) имеют на последних фалангах пальцев примитивные копыта. Зубная система обладает чертами, характерными для копытных: клыков нет, верхние резцы растут непрерывно. В желудке два отдела: передний является «складом» для пищи, в заднем, железистом, происходят процессы пищеварения. Даманы — боковая ветвь копытных, но они ближе к предкам этой группы, чем другие представители. Изучая их, можно составить представление о некоторых особенностях примитивных копытных (малая величина, многопалые конечности и т.д.). Известно 10 видов.
Хоботные когда-то были многочисленной, широко распространенной группой животных. В настоящее время существуют только два вида: индийский слон (Elephasmaximus), обитающий в Южной Азии, и африканский слон (Loxodontaafricanus), сохранившийся только в национальных парках, организованных в некоторых странах Африки (южнее Сахары). Это самые большие наземные млекопитающие. Индийский слон предпочитает лесистые места, африканский — саванны. Колоннообразные ноги заканчиваются пятью более или менее сращенными пальцами, одетыми небольшими копытами. Нос и верхняя губа вытянуты в хобот, мускулатура которого очень развита и сложна. Хобот может поднимать и большие тяжести, и совсем крохотные предметы; он служит также для приема воды. Череп кажется громадным вследствие больших выростов для прикрепления мощных мышц, но объем его мозговой части по сравнению с телом невелик. Масса мозга 5 кг, т. е. около 1/500 массы тела. Зубная система сильно изменена: два резца верхней челюсти превратились в мощные бивни (неправильно называемые клыками) исключительной крепости, растущие в течение всей жизни. В каждой половине челюсти имеется один огромный коренной зуб со сложной поверхностью, служащий для перетирания веток и другой растительной пищи. Каждый такой зуб функционирует 10—15 лет, потом замещается выступающим сзади и передвигающимся вперед очередным зубом. Всего в течение жизни слона сменяется 24 зуба (по шести в каждой половине челюсти).
Индийские слоны используются как домашние животные. Обладая колоссальной силой, они выполняют очень тяжелые работы.
Слоны приручаются, но в неволе, как правило, не размножаются. Поэтому настоящими домашними животными их считать нельзя.
Хоботные, так сильно отличающиеся от обычных млекопитающих, произошли от небольших представителей этого класса. Характерные признаки (большая величина тела, хобот, бивни и т. д.) развились постепенно в длинном ряду предков. Из современных млекопитающих довольно близки к вымершим небольшим предкам хоботных даманы.
Крупные копытные, обитающие в прибрежной зоне Атлантического и Индийского океанов, заходят в некоторые реки и озера. Животные находятся на пути к полному вымиранию. В настоящее время сохранился дюгонь (Индийский океан) и несколько видов ламантинов (Атлантический океан, реки Южной Америки). До середины XVIII в. у Командорских островов жила огромная морская корова (длина около 10 м), истребленная в результате хищнического промысла.
Тело у сирен обтекаемой формы, торпедообразное, заканчивается хвостом. Передние конечности превратились в ласты, задние полностью атрофировались, имеются рудименты таза. Растительноядны. Зубная система имеет некоторое сходство с той же системой слонов. Сирены произошли от общих предков с дамана-ми и хоботными.
Эти копытные животные еще сравнительно недавно были широко распространены и многочисленны. В настоящее время остались лишь тапиры, носороги и лошади. Число пальцев различно: у тапиров передние ноги четырехпалые, задние — трехпалые, у носорогов все ноги трехпалые, у лошадей — однопалые. Характерно для всех непарнокопытных сильное развитие третьего пальца, через который проходит ось конечности, у лошадей он остался единственным. Ключиц нет. Все непарнокопытные ~ крупные животные.
Семейство тапиры (Tapiridae) — самые примитивные животные из современных непарнокопытных. Подобно вымершим древним представителям отряда, они имеют многопалые конечности, живут в лесах и питаются мягкой растительностью. Когда-то тапиры были широко распространены в разных частях земного шара, но потом были вытеснены более совершенными непарнокопытными и теперь сохранились в болотистых лесных чащах Центральной и Южной Америки и Юго-Восточной Азии. Такое «разорванное» распространение характерно для многих вымирающих форм.
Семейство носороги (Rhinocerotidae) — огромные, сильные животные (длиной до 5 м и массой более 2 т) с очень толстой кожей, лишенной волос. На носовых и лобных костях у них сидят один или два рога. Эти непарнокопытные обитают в Южной Азии и в Африке. Они были широко распространены в течение большей части третичного периода, а некоторые из них жили еще в четвертичном периоде. Сейчас внесены в Красную книгу.
Семейство лошадиные (Equidae) — наиболее прогрессивная группа непарнокопытных, приспособленных к жизни на открытых пространствах — в степях и полупустынях. Животные сильные, быстро бегают, их органы чувств (в особенности слух и обоняние) прекрасно развиты. Современные лошадиные относятся к одному роду — лошадь (Equus), в состав которого входят четыре вида (другие авторы выделяют их в особые подроды). В особый подрод выделяют, например, зебр. Эти животные с характерным полосатым рисунком обитают на открытых местах разных частей Африки. Одни зоологи всех зебр относят к одному виду, распадающемуся на несколько подвидов, другие различают три самостоятельных вида. Дикие ослы (Equusasinus) сохранились на севере Эфиопии, в Сомали и Эритрее. От них произошли домашние ослы. В полупустынных районах Азии (Туркмения, Иран, Афганистан, Монголия, Китай, Тибет, Западная Индия и т. д.) обитают куланы (Equushemionus). Этих животных часто называют полуослами, но по мнению других авторов, куланов следует считать примитивными лошадьми. Дикие лошади Пржевальского (Equusprzewalskii), названные в честь великого русского путешественника Н. М. Пржевальского, открывшего ее, в диком состоянии сохранились, возможно, только в Центральной Азии, хотя еще недавно были распространены в степях Европы и Северной Азии. В Европе последние дикие лошади, близкие к лошади Пржевальского, — тарпаны (Equusgmelini) — были истреблены в середине прошлого столетия. От кого именно произошли домашние лошади, точно не установлено, возможно, что от нескольких диких видов, близких друг к другу.
К этому отряду, который до последнего времени рассматривали в составе отряда парнокопытных, относятся верблюды и ламы. Крупные животные с длинной шеей, хорошо развитым шерстным покровом. Ноги двупалые (как у парнокопытных, сохранились только третий и четвертый пальцы, рудиментов остальных пальцев нет) с искривленными тупыми когтями и мозолистыми подушкообразными утолщениями. Желудок состоит из трех камер — рубца, сетки и сычуга. Строение рубца и сычуга сильно отличается от строения тех же камер у парнокопытных.
Слепая кишка короткая. Существенно отличаются от парнокопытных строением половых органов, плаценты и других органов.
Верблюды прекрасно приспособлены к жизни в пустынях, очень выносливы и подолгу обходятся без пищи и воды. Для них характерна наклонность к образованию жировых горбов, содержимое которых потребляется во время голодовки и отсутствия воды. В настоящее время известны только два вида верблюдов: одногорбый,
или дромадер
(Camelusdromaderius), сохранившийся лишь в одомашненном состоянии, и двугорбый,
или бактриан
(С. bactrianus), оставшийся в небольшом количестве в западных пустынях Монголии и, возможно, Китая. Верблюды до сих пор широко используются как вьючные животные в пустынях, а также для других хозяйственных целей.
Ламы, или безгорбые верблюды, обитают только в Южной Америке, высоко в Андах. Известны два диких вида — гуанако
и викунъя.
От гуанако человек вывел домашние формы: алъпаку
для получения шерсти и ламу
для использования как вьючное животное.
Большой и разнообразный отряд растительноядных млекопитающих, приспособленных, как правило, к быстрому передвижению, в связи с чем у них на каждой ноге развиты только два пальца — третий и четвертый, между которыми проходит ось ноги, чем и объясняется название отряда. Второй и пятый пальцы сохранились в виде рудиментов, лучше развитых у низших парнокопытных; первый палец отсутствует. Ключиц нет. Отряд делится на два подотряда: нежвачные, или свинообразные (с двумя семействами — свиньи и бегемоты), и жвачные (с несколькими семействами, из которых здесь будут рассмотрены только три — олени, жирафы и полорогие). Количество видов — около 170.
(
NONRUMINANTIA, или SUIFORMES)
Млекопитающие средней или довольно значительной величины. Ноги короткие (второй и пятый пальцы хотя и малы, но развиты лучше, чем у других парнокопытных). Кожа голая или покрыта редкой жесткой шерстью. Подкожный жировой слой хорошо развит. Рогов нет. Зубы всех родов. Желудок простой. Нежвачные более примитивны и менее широко распространены, чем жвачные. К ним относится сравнительно небольшое количество видов.
К семейству бегемотов (Hippopotamidae) относится всего два вида (обитающих в Африке), из которых настоящий бегемот, или гиппопотам, достигает огромных размеров, проводит значительную часть жизни в воде, а другой — карликовый бегемот — обитает около рек в тропических лесах и похож по образу жизни на свиней. Оба растительноядны. Гиппопотам может сильно вредить полям.
К семейству свиных (Suidae) относится довольно значительное количество видов. Они распространены главным образом в Азии, Африке и Европе. В Южной Америке обитают наиболее примитивные представители свиней — пекари. Свиньи имеют удлиненное рыло с хрящевым «пятачком» на конце; тело их покрыто редкими волосами (щетиной). Клыки сильно развиты. Питаются преимущественно растениями, но охотно потребляют и корма животного происхождения (червей, насекомых, мелких грызунов, падаль и т. д.). Рождают много детенышей (до 12). В нашей фауне — всего лишь один вид диких свиней — кабан (Susscrofa). Он был сильно истреблен, но в последние десятилетия численность его благодаря различным охранным и акклиматизационным мероприятиям очень выросла. В некоторых районах разрешен отстрел кабанов. Средняя масса кабанов 80—150 кг, в редких случаях — до 250 кг. Дикие кабаны подвижны, тело их более стройное, ноги длиннее, шерсть лучше развита, чем у домашних свиней. Наши домашние свиньи происходят от кабана. В результате длительного и систематического отбора человек вывел породы домашних свиней, которые отличаются от диких предков быстрым ростом, большими размерами, высокой плодовитостью, усиленной способностью откладывать жир.
Стройные млекопитающие средних и больших размеров, как правило, приспособленные к быстрому передвижению. Ноги длинные, второй и пятый пальцы рудиментарны. Пястные и плюсневые кости сливаются в столбчатую кость, которая важна как опора при быстром передвижении. Кожа покрыта густой шерстью. Подкожный жировой слой почти не развит. Большинство видов (у оленей, как правило, самцы) вооружены рогами. Питание исключительно растительное: травы, кора и другие части деревьев (преимущественно молодых), ягоды, лишайники. На верхней челюсти отсутствуют резцы, а у большинства и клыки. Клыки нижней челюсти имеют такую же форму, как и резцы, и служат для откусывания травы. Коренные зубы имеют сложную складчатую поверхность и хорошо приспособлены для перетирания жесткой растительной пищи. Желудок сложный и состоит из четырех отделов — рубца, сетки, книжки и сычуга. Кишечник очень длинный, хорошо развита слепая кишка. Беременность продолжается долго. Большинство жвачных рождают раз в год одного-двух детенышей.
Жвачные прекрасно приспособлены к питанию трудноперева-римыми растительными кормами и обитают в разнообразных условиях (в степях, лесах, горах, тундре и т.д.). Благодаря приспособительным особенностям жвачные, несмотря на истребление ряда видов, остаются еще достаточно многочисленными.
Семейство оленевые (Cervidae) — жвачные животные с костными ветвистыми рогами. Рога, за исключением северного оленя, свойственные только самцам, ежегодно сбрасываются. Рудименты второго и пятого пальцев довольно значительны. Клыки на верхней челюсти сохраняются. Олени обитают в Азии, Европе и Америке. Многие из них сильно истреблены. Молодые рога (панты) марала, изюбря и пятнистого оленя представляют ценность, так как из них получают лекарственные вещества, стимулирующие обмен веществ. Ради пантов этих оленей приручают и разводят в полудомашнем состоянии. Северный олень (Rangifertarandus) как домашнее животное широко используется в районах Крайнего Севера России.
Семейство жирафы (Giraffidae) — жвачные животные с маленькой головой на длинной шее. Живут в саваннах Африки, но заходят и в горы. Жирафы достигают огромного роста — до 5—6 м. К тому же семейству принадлежит животное средних размеров — окапи, живущее в тропических лесах Африки и близкое к предкам жирафов.
Семейство полорогие (Bovidae, или Cavicornia) — одно из самых больших и разнообразных по условиям обитания семейств парнокопытных. Жвачные с неветвистыми полыми рогами, сидящими на отростках лобных костей. Рога развиваются из эпидермиса и состоят из рогового вещества; они никогда не сменяются (за исключением американского вилорога). Самки большинства видов рогов не имеют или их рога меньшей величины, чем у самцов. От
диких представителей этого семейства произошли различные породы мелкого и крупного рогатого скота.
Сайгак, или сайга (Saigatatarica), раньше добывался из-за рогов. В XIX и XX вв. этот вид, широко распространенный в степях низовьев р. Урала, в Казахстане, Средней Азии, был почти истреблен. В настоящее время в результате принятых мер по охране сайгака численность его резко увеличилась.
Относящиеся к роду Оvis бараны — горные животные, живут на высоте от 150 до 6000 м.
Козлы, относящиеся к роду Сарга, еще больше, чем бараны, приспособлены к жизни в горных условиях и доходят до полосы вечных снегов. Из диких козлов в России на Кавказе обитают дагестанский тур, тур западнокавказский, безоаровый козел, а в Саянах и на Алтае — сибирский козерог. Домашние козы произошли Д от упомянутых выше безоарового (Сарга hircus, или С. aegargrus) и винторогого козла (Сарга falconeri).
Буйволы (Bubalus) включают одомашненные (индийский буйвол, филиппинский буйвол) и не одомашненные виды (африканский буйвол). Буйволы живут около водоемов, много времени проводят в воде и питаются разнообразной водной растительностью. Одомашненные буйволы используются как тягловая сила и как источник пищевых продуктов.
К роду быков (Bos) относятся бизоны (Bosbison) — огромные животные, еще в XIX в. населявшие в большом количестве степи Северной Америки. Они были быстро истреблены и теперь разводятся в специальных заповедниках. В Европе, в Беловежской Пуще, в небольшом количестве сохранились близкие к бизонам зубры (В. bonasus). В настоящее время принимают различные меры, способствующие размножению зубров в Беловежской Пуще, в Крыму и на Кавказе. Бизоны не были одомашнены.
Относящиеся к этому же роду приземистые, мохнатые яки (В. mutus) в диком состоянии живут в Центральной Азии. Они прекрасно приспособлены к суровым условиям высокогорья. Яки одомашнены и разводятся в Киргизии и в некоторых других горных районах Средней Азии и на Алтае.
В Южной Азии обитают лобастые быки. К ним относятся гаур (В. gaurus), бантенг (В. avanicus) и камбоджийский бык, или куп-рей (В. sauveli). Одомашненная форма гаура — гаял. Бантенг является родоначальником домашнего балийского скота, разводимого на Малайском архипелаге. Камбоджийский бык открыт совсем недавно в лесах Кампучии.
В средние века в Восточной и Средней Европе был широко распространен дикий бык — тур (Bosprimigenius), на которого раньше усиленно охотились. Последний дикий тур погиб в 1627 г. От тура происходят разнообразные породы крупного рогатого скота, а также горбатый скот — зебу, распространенный главным образом в сухих степях, преимущественно в Южной Азии и Африке.
Один из древних отрядов млекопитающих, объединяющий более 200 видов разнообразной величины. У приматов сохраняются многие примитивные черты строения. Они обладают головным мозгом больших размеров и характеризуются весьма сложным поведением. Большинство видов ведут древесный образ жизни в субтропических и тропических областях Азии, Африки и Америки. В отряде два подотряда — полуобезьяны и человекоподобные приматы, или обезьяны.
Небольшие древесные животные. Лицевая часть черепа сильно вытянута, мозговая коробка относительно меньше, чем у настоящих обезьян. Полушария переднего мозга не закрывают мозжечка и лишены извилин. Обонятельные доли, как и у насекомоядных, хорошо развиты. Глазницы у большинства направлены скорее в стороны, чем вперед. Ключицы имеются, конечности пятипалые. Одни пальцы заканчиваются когтями, а другие — ногтями. Первый палец задней конечности способен противопоставляться остальным пальцам. Матка двурогая. Как правило, самка рождает одного детеныша. Сумеречные или ночные животные, раститель-ноядные, насекомоядные или всеядные. Обитают в Южной Азии и Африке (главным образом на о. Мадагаскар). Произошли от примитивных насекомоядных.
Древесные или наземные животные, средней или небольшой величины. Лицевая часть черепа меньше вытянута, чем у полуобезьян, и мозговая коробка развита значительно лучше. Полушария переднего мозга велики, имеют на поверхности много извилин и закрывают сверху мозжечок. Обонятельные доли малы. Глазницы направлены вперед. Ключицы имеются, конечности пятипалые. Лапы хватательные — первый палец противопоставляется остальным. Матка простая. Почти все — дневные животные. Пища обезьян в основном растительная, но многие из них могут питаться и мелкими животными (беспозвоночными и позвоночными). Подотряд делят на 2 секции.
Секция широконосые обезьяны (Platyrrhina) — маленькие и средней величины древесные животные: игрунки, капуцины, ревуны и др., как правило, с цепким хвостом. Ноздри направлены в стороны, так как перегородка между ними широкая. Седалищных мозолей и защечных мешков нет. Обитают в Центральной и Южной Америке.
Секция узконосые обезьяны (Catarrhina) — древесные или наземные животные разной величины. Цепкого хвоста не имеют и цепляются за ветки только с помощью конечностей. Ноздри направлены вперед, так как перегородка между ними узкая. Обитают в Африке и Южной Азии. Секция разбивается на четыре семейства: мартышкообразных (мартышки, павианы, макаки и др.), гиббоновых, понгид (орангутаны, гориллы, шимпанзе) и гоминид (человек и его вымершие предшественники).
Мартышкообразные обезьяны (Cercopithecidae) имеют защечные мешки, большей частью длинный хвост и почти все — седалищные мозоли. Большинство ведет древесный образ жизни. Гиббоновые (Hylobatidae) — древесные обезьяны с чрезвычайно длинными передними конечностями. Хвоста и защечных мешков нет. Есть небольшие седалищные мозоли. Обитают в Индокитае и на островах Суматра, Калимантан и Ява. Понгиды (Pongidae) не имеют хвоста, защечных мешков и седалищных мозолей. Они отличаются от остальных обезьян и всех млекопитающих сильно развитой корой переднего мозга и очень сложной нервной деятельностью. Орангутан обитает на островах Калимантан и Суматра горилла и шимпанзе - в тропических частях Африки.
Приматы - достаточно многочисленная и процветающая группа что объясняется их основным преимуществом по сравнению с другими млекопитающими: сильным развитием переднего мозга его коры, благодаря чему их поведение очень усложнено, они быстро приобретают новые навыки, несомненно, проявляют элементы рассудочной деятельности, более развитые у высших обезьян. Большую роль в их жизни играют пятипалые конечности хорошо приспособленные к хватательным движениям. Строение и физиология высших обезьян и человека во многом очень сходны, но все же между ними существуют очень серьезные различия в основном в степени развития высшей нервной деятельности и в умении использовать различные предметы для воздействия на окружающую среду. Поэтому филогенетическое развитие от высших человекоподобных обезьян до человека разумного было довольно продолжительным.
Несомненными предками человека, но очень примитивными, были австралопитеки, жившие около 3 млн лет тому назад. Их останки были найдены в разных местах Африки. Становление человека т е. усложнение его мозга и нервной деятельности, все большее использование им разных предметов, ставших орудиями труда выработка речи, осуществлялось в длинном ряду предков (питекантропы, синантропы, неандертальцы и др.), начиная с конца третичного периода и в течение почти всего четвертичного периода (продолжительность которого около 1 млн лет). Человек разумный (Homosapiens), по своим физическим и психическим возможностям не отличавшийся от современного человека, появился не позднее 40-50 тыс. лет тому назад. С тех пор человеческое общество прошло через ряд социально-экономических формаций, изучением которых заняты не биологические науки, а науки общественного цикла.
Зоогеография —
наука, изучающая распространение животных на земном шаре и устанавливающая общие закономерности этого распространения. Основным объектом зоогеографии служит фауна, иначе говоря, исторически сложившаяся совокупность видов животных, населяющих какую-нибудь область (будь то целый материк, остров или любой географический район) и входящих во все ее биоценозы. Кроме того, зоогеография изучает и ареалы, т. е. области распространения отдельных видов, родов или других систематических групп животных.
Общеизвестно, что животный мир тундры резко отличается от животного мира широколиственного леса, пустыню населяют одни виды, а тайгу — другие, наконец, в океане мы встречаем совершенно особые формы, не живущие на суше. Это объясняется разными условиями среды обитания, в первую очередь климатом и характером растительности.
Однако есть ряд отличий в характере фаун, объяснить которые только особенностями среды обитания нельзя. К примеру, области влажных тропических лесов Южной Америки, Африки и Азии с одинаковым климатом и очень сходным типом растительного покрова населяют разные фауны. Причины их кроются в истории формирования фаун названных материков: длительное развитие фаун в условиях изоляции приводит к появлению коренных различий между ними. Если же между двумя участками земного шара существует связь и возможен обмен животными, то на громадном протяжении возникает сходная фауна.
Отсюда следует вывод, что распространение животных на земном шаре можно объяснить, учитывая не только ныне действующие условия среды, но и условия прошлых эпох, т. е. историю земной коры.
Зоогеографию принято делить на регистрационную (описательную), сравнительную и каузальную (причинную).
Приступая к изучению любого фаунистического комплекса, первоначально регистрируют, по возможности, все виды, входящие в него. Затем видовую структуру фауны одного района сравнивают с таковой соседних и более отдаленных фаун. Это дает возможность выяснить место, занимаемое той или иной фауной в ряду других, ее характерные особенности, «собственные черты», т. е. степень оригинальности. В дальнейшем это позволяет судить о возрасте фауны, а также о ее былых и современных связях с другими фаунами. Однако регистрация и даже сравнение — не самоцель, а путь к выяснению причин, определяющих состав и характер фауны. Поэтому подразделения зоогеографии правильнее было бы рассматривать как этапы исследования.
Причины заселения участка территории или акватории земного шара тем или иным фаунистическим комплексом можно свести к факторам среды, действующим в настоящее время. Это могут быть условия климата, характер поверхности или тип растительности, с которым взаимодействуют животные организмы. Так как подобные взаимодействия изучает экология, соответствующий раздел каузальной зоогеографии, рассматривающий влияние среды на распространение современных животных, называется экологической зоогеографией.
Выше указывалось, что не все особенности географического распространения животных объясняются влиянием современных условий. Фаунистические комплексы развиваются во времени, и тот раздел каузальной зоогеографии, который пытается объяснить современное распространение животных причинами, действовавшими в прошлом, называется исторической,
или генетической, зоогеографией.
К числу таких причин относятся бывшие материковые соединения, изменения климата и растительного покрова в минувшие геологические эпохи и т. д.
Все разделы зоогеографии представляют собой единое целое и отличаются только подходом к толкованию фактического материала, предоставляемого описательной и сравнительной зоогеографией.
Для выполнения задач, стоящих перед различными направлениями, зоогеография пользуется собственными методами исследования, применяя вместе с тем достижения и методы смежных наук. Число этих наук велико. Прежде всего это зоология, а также ряд наук о Земле.
Зоогеография теснейшим образом связана с зоологической систематикой. Перед тем как приступить к зоогеографическим построениям, нужно точно выяснить систематическую принадлежность исходного материала, отнести его к определенным видам.
Изучение микроэволюционных процессов дает неоценимый материал для понимания зоогеографических тенденций. Поэтому зоогеография использует идеи и данные теории эволюции. В свою очередь, результаты исследований зоогеографии очень полезны для систематики и эволюционного учения.
Очевидна связь зоогеографии с палеозоологией. Ископаемые дают самую лучшую основу для выяснения истории распространения животных. Без палеонтологического материала в большинстве случаев нельзя обосновать ни одну теорию расселения из центров происхождения тех или иных форм.
Зоогеография настолько тесно связана с географией растений (фитогеографией), что их нужно рассматривать, как уже говорилось, в качестве частей единой науки — биогеографии.
Обширный комплекс наук о Земле (климатология, ландшафто-ведение, геология), а также их разделы, изучающие прошлое нашей планеты (палеогеография, историческая геология, палеокли-матология), в той или иной степени используются зоогеографией. Не зная геологического прошлого страны, нельзя объяснить современное распространение животных, особенно тех групп, ареалы которых размещены на разных материках. Без изучения климатов прошлого, особенно периодов, предшествовавших современной эпохе, трудно установить причины сохранения в современных условиях ряда видов, экологические требования которых находятся как бы в противоречии с современной обстановкой, например холодолюбивых видов в Центральной Европе или влаголюбивых форм в среднеазиатских пустынях.
Основными направлениями исследований в зоогеографии являются, во-первых, выяснение современного распространения животных на планете, во-вторых, выявление причин различий между фаунами разных частей земного шара и, в-третьих, обнаружение закономерностей, которые регулируют или регулировали в прошлом расселение животных из центров их происхождения. Одновременно зоогеография освещает географическую точку зрения на процесс эволюции животных.
Вполне правомерно говорить и о практическом значении зоогеографических работ, особенно сегодня, когда человек с его технологическими средствами воздействия на биосферу выходит на один уровень с природными силами, изменяющими облик нашей планеты. Развитие средств и путей сообщения приводит к тому, что антропический фактор[2]
становится мощной силой в расселении животных. Напомним в этой связи историю расселения колорадского жука в Европе, европейских видов животных в Новой Зеландии и Австралии, малярийного комара в Бразилии, не знавшей малярии до завоза этого комара из Африки.
Откликаясь на запросы народного хозяйства, зоогеография призвана прогнозировать все возможные перемены в животном мире, чтобы предупредить исчезновение редких видов, прежде всего ценных. Велико ресурсоведческое значение зоогеографии, особенно для пушного и морского промыслов, оценки охотничьих территорий, для медицины.
Важна роль животных и как индикаторов изменений окружающей среды, обусловленных, в частности, ее загрязнением.
Одной из центральных проблем современной зоогеографии следует считать изучение всевозможных причин и путей изменения фауны — от природных до антропических. В связи с этим зоогеография ставит перед собой следующие задачи.
1. Изучение ареалов — областей земного шара, населенных популяциями определенных видов.
2. Выявление причин, определяющих характер географического распространения животных организмов.
3. Изучение закономерностей формирования фаун под влиянием природных и антропогенных факторов.
4. Прогнозирование изменений животного мира в обозримом будущем для предотвращения обеднения его видового состава или сдвига последнего в нежелательную для человека сторону.
В настоящее время зоогеография, объединяясь со многими естественными науками, успешно решает эти задачи.
Зоогеография как наука в строгом смысле слова является детищем XIX в. Становление и развитие ее обусловлено накоплением фактов прежде всего в области биологии и географии и их акамезон.
Крупный биолог и зоогеограф М.А.Мензбир первым в отечественной литературе (1882) предложил выделить четыре этапа развития зоогеографии, отражавшие отношение ученых к вопросам о происхождении и развитии органического мира Земли. Его периодизация, по существу, относилась к истории биологии в целом. Мензбир различал периоды, связанные: 1) со слепой верой в истину библейской догмы о сотворении мира, 2) с господством теории катастроф, 3) с отказом от теории катастроф и распространением принципа актуализма, 4) с победой дарвиновского учения об эволюции.
Американские биогеографы, в частности Л. Стюарт (1957), выделяют в истории зоогеографии три периода.
Первый из них связан с работами К.Линнея, заложившего основы систематики животных. Протяженность периода — почти столетие. Для него характерны описания распределения на Земле различных групп животного царства. К этому же периоду относились и первые попытки районирования, т. е. выделения зоогеогра-фических областей.
Второй период начался с появлением эволюционного учения Ч.Дарвина. В это время зоогеографы, продолжая изучение и описание географического распространения животных, уже пытаются объяснить возникновение характерных сочетаний видов (фаун), основываясь на эволюционной теории.
Наконец, третий период, начавшийся в XX в. и продолжающийся в настоящее время, характеризуется прежде всего «эколо-гизацией» зоогеографии, т. е. изучением среды обитания животных и учетом взаимодействий между организмом и средой. К этому же периоду относится и влияние на биогеографию генетических принципов, предложенных Г.Менделем.
Уточнение истории зоогеографии и ее периодизации принадлежит советским биогеографам А. Г. Воронову (1963) и Г.В.Наумову (1969), которые придерживались точки зрения Мензбира. Именно эта усовершенствованная периодизация служит основой представленного здесь исторического очерка.
Но сначала обратимся к предыстории зоогеографии.
В трудах Аристотеля (384-322 гг. до н.э.), описавшего около 500 видов животных, большой интерес вызывают высказывания о медленных изменениях географической среды, поднятии отдельных участков суши при опускании других, аналогию с которыми можно найти в современных взглядах на вековые колебания земной поверхности и связанные с ними климатические изменения.
Развитие географии, связанное с именами Эратосфена (275— 194 гг. до н. э.) и особенно Страбона (63-20 гг. до н. э.), потребовало описания животных и растений, характеризующих определенный участок земной поверхности.
Эпоха Римской империи оставила нам многотомный труд Плиния Старшего (23-79 гг. н.э.) «Естественную историю», два тома которой посвящены живым организмам. Нового, впрочем, у Плиния оказалось мало. В основном это были сведения, почерпнутые из работ Аристотеля, с добавлением фантастических известий о несуществующих организмах.
Установление господства христианской церкви и падение Римской империи в V в. н. э. привело к упадку наук и забвению наследства античных ученых.
Лишь на Востоке арабы, позаимствовав многие знания у разоренной ими античной цивилизации и став ее преемниками, вносили в науку и свои собственные наблюдения. Большой интерес представляют труды таджикского ученого Абу-Али Ибн-Сины (980—1037), который собирал в своем «Каноне» практически все естественно-научные знания того времени и в том числе привел массу ценных сведений о животных Средней Азии.
В эпоху Возрождения (XIII—XVI вв.) произошел культурный переворот во всех областях жизни и деятельности людей, чему в немалой степени способствовало стремление ряда стран к расширению рынков. В этот период пробуждается интерес к географическому знанию. Итальянец Марко Поло, совершивший путешествие в Монгольское ханство через территорию современной Средней Азии, оставил описание трех народов и стран, которые ему довелось посетить за 24 года странствий (1271—1295), где также содержится немало достоверных сведений о природе и животных Азии.
Необходимо отметить и русских землепроходцев. Им мы обязаны первыми знаниями о животном мире Сибири, главным образом о пушных зверях. Сейчас мы судим о былом распространении соболя по «отпискам» так называемых служилых людей. Первые сведения о фауне Байкала, в частности о нерпе, доставил Василий Власьев, а Савва Есипов привел целый список сибирских животных, куда входили олень, лось, козел, лисица, соболь, росомаха, бобр и др. На северо-востоке Сибири первооткрывателями были обнаружены моржовые лежбища. В 1665 г. Юрий Селиверстов описал миграцию кеты в реке Анадырь, причем подметил, что идущая из моря на нерест кета погибает в верховьях реки. А Владимир Атласов связал питание соболя, лисицы и выдры с обилием в реках кеты. Это были настоящие фаунистические заметки. Пробывший 15 лет в Сибири (1661—1676) Юрий Крижанич написал «Историю Сибири», где дал характеристику ее трем зонам («трем климатам») — тундре, тайге, степи, описав климат, растительность и животный мир каждой из них. Это была первая биогеографическая работа по Сибири.
XVII в. для естественных наук был эпохой главным образом описательной. Естествоиспытатели собирали фактический материал, коллекционировали, совершенствовали методы исследований. Однако попыток систематизировать данные о географическом распространении животных почти не было.
В 1605 г. Виртсген произвел анализ фауны Британских островов и европейского континента, сделав правильный вывод о тождестве животных Англии, Северной и Западной Европы. Сходство Виртсген объяснял недавней связью этих континентов, датируя ее временем «после потопа», в который верил тогда не только он.
Подобную работу в 1660 г. провел английский капитан Симпсон. Он исследовал Фолклендские острова и сравнил распространенных там лисиц с южноамериканскими. Они оказались сходными. Симпсон делает аналогичный предыдущему вывод — Фолклендские (Мальвинские) острова соединялись некогда с Южной Америкой. Но оба вывода остались в то время практически незамеченными. Даже в XVIII в. факты географического распространения организмов все еще объяснялись исходя из религиозной догмы о сотворении мира.
Начало XVIII в. ознаменовалось трудами нескольких научных экспедиций. Особое место среди них занимает вторая камчатская экспедиция (1733—1743) Витуса Беринга. Ее участник Г.Стеллер исследовал фауну и флору Сибири. Он, в частности, открыл и описал морскую корову — млекопитающее семейства дюгоне-вых, обитавшее на мелководьях у Командорских островов. С. П. Крашенинников, также участвовавший в этой экспедиции, дал характеристику природы Камчатки, прежде всего ее животного населения.
Натуралист И. Гмелин, член Петербургской академии наук, в 1733—1743 гг. исследовал области к востоку от Байкала и бассейн Лены. Он первым обратил внимание на то, что Западная и Восточная части Сибири отличаются одна от другой, и границу между ними провел по Енисею.
В первый период развития зоогеографии,
охватывающий вторую половину XVIII в., появились труды многочисленных исследователей-натуралистов. Они были обобщены работами знаменитого шведского ученого Карла Линнея (1707—1778) — основателя научной систематики растений и животных. Однако будучи приверженцем библейской догмы о сотворении животных в раю, расположенном в междуречье Тигра и Евфрата, Линней предложил гипотезу о том, что местом творения была высокая гора на острове, расположенном в тропиках. У ее подножия, согласно Линнею, были сотворены тропические виды животных, выше, соответственно изменению климата в верхних поясах гор, — виды умеренной зоны, еще выше — полярные и т. д. Когда остров соединился с материком, все виды расселились по «своим», уже широтным, климатическим зонам Земли.
М. В.Ломоносов одним из первых высказал идею исторического развития Земли, распространив ее как на земную кору, так и на органический мир. К примеру, он считал, что обнаружение ископаемых морских моллюсков на вершинах гор служит доказательством того, что в этих районах когда-то располагались морские бассейны.
В этот период развития зоогеографии появились фаунистические описания крупных районов земного шара, что породило вопрос о причинах различия фаун. Высказывались предположения, что изменения животного мира Земли шли параллельно геологическим изменениям. Это был период накопления материалов, позволивших высказать гипотезу не только о смене фаун, но и об эволюции видов.
Второй период в развитии зоогеографии
датируется концом XVIII — первой половиной XIX в. Его обычно называют периодом господства теории катастроф.
Совсем отрицать изменения растительного и животного мира Земли при обилии имевшихся палеонтологических фактов было уже невозможно. Для согласования с библейской догмой об актах творения возникла компромиссная теория катастроф, согласно которой творец создает органический мир каждой геологической эпохи заново, специальным актом творения, затем все живое гибнет в результате грандиозной мировой катастрофы, после чего наступает новый акт творения.
Блестящий представитель науки конца XVIII в. Жорж Бюффон (1707—1788), автор многотомной «Естественной истории», признавал быстрые изменения поверхности земли, зависимость распределения животных и растений от размещения морей и суши, значение горных хребтов и больших водных пространств как преград к расселению животных и высказал ряд важных биогеографических положений. В то же время он придерживался теории катастроф, говорил о существовании семи геологических эпох, которые разделялись гигантскими катастрофами, уничтожавшими органический мир предыдущей эпохи. Правда, Бюффон отрицал полное уничтожение видов, допускал выживание части их и последующее пополнение за счет вновь созданных.
В законченном виде теория катастроф была сформулирована в начале XIX в. Жоржем Кювье (1769—1832), а в наиболее крайней форме представлена в работах его ученика д'Орбиньи.
В первом периоде развития зоогеографии трудами Ж. Бюффона, Е. Циммермана и П. С. Палласа было заложено направление, которое потом назовут фаунистическим или региональным.
В 1777 г. немецкий ученый Е.Циммерман опубликовал книгу «Опыт зоологической географии», где основательно изложил сведения о миграциях животных и ввел термины «зоогеография» и «географическая зоология». Первая, по Циммерману, изучает различные части земного шара с точки зрения их животного населения, а вторая занимается установлением причин распространения животных. Задачами зоогеографии Циммерман считал объяснение современного и прошлого распространения животных, определение центров расселения. В отличие от Линнея он полагал, что каждый вид имеет свой центр творения. По сути, мы вправе считать Циммермана основателем зоогеографии как науки.
Огромный вклад в развитие зоогеографии внес российский академик П.С.Паллас (1741—1811), который вместе со своими учениками совершил ряд путешествий и собрал огромный фактический материал по фауне и флоре востока европейской части России, а также Сибири и других территорий. Большое внимание Паллас уделял особенностям обитания и распределения животных. Он является одним из первых биогеографов, применявших экологические исследования.
На этот же период приходятся и первые попытки установления естественных фаунистических областей, предпринятые И.Мин-дингом (1829), А.Вагнером (1844) и др.
Итогом зоогеографических исследований данного периода явился труд Л. Шмарды «Географическое распространение животных» (1853). Это была сводка всех известных тогда факторов распределения фауны. Шмарда заложил фундамент экологического направления в зоогеографии, поскольку полагал, что распределение животных обусловливают факторы среды — свет, температура, пища и т. п. По теоретическим взглядам он был катастрофистом и не признавал идеи эволюции даже после выхода трудов Дарвина.
К началу третьего периода
(середина XIX в.) ученые стали отказываться от теории катастроф. Во многих работах уже появляются мысли о постепенной эволюции облика Земли. В 1832 г. увидела свет книга английского геолога Ч.Лайеля (1797—1875) «Основы геологии». Она нанесла серьезный удар по теории катастроф и вместе с тем подготовила почву Дарвину для разработки научной теории эволюции. Лайель утверждал, что для изменения поверхности земного шара не требуется гигантских катастроф или иного вмешательства высшей силы и, принимая во внимание возраст нашей планеты, мы должны допустить, что постепенные, незаметные процессы, такие, как образование оврагов, размыв береговой линии, разрушение каменных пород под действием солнца, воды, ветра и т. п., за тысячи и миллионы лет могут привести к появлению иных форм рельефа, смене суши морем и т. п. «Настоящее есть ключ к пониманию прошедшего», — вот убеждение Лайеля. Изучение процессов, протекающих на Земле в настоящее время, достаточно для суждения о тех же процессах и условиях среды прошлых геологических эпох. Поэтому причинами резких изменений в составе ископаемых фаун, которые объяснялись губительными катастрофами, Лайель считал вековые колебания суши и моря и вызванные ими переселения животных. Он утверждал, что фауны прошлых периодов генетически связаны друг с другом. Эта точка зрения получила наименование «принципа актуализма». Лайел, хоть и не сразу, стал убежденным сторонником Дарвина.
Идеи, подобные лайелевским, возникали и раньше. Так, еще в 1822 г. русский академик К.М.Бэр в докладе «Как развивалась жизнь на Земле» выдвинул гипотезу о постепенном развитии природы от низших ее проявлений до человека. Бэр утверждал, что история животного мира древнее истории человечества, а история Земли древнее, чем история животного мира. Позже, в 1834 г., с эволюцией животных Бэр уже связывает их географическое распространение.
Из работ, где нашел воплощение принцип актуализма, нужно назвать книгу английского биогеографа Э. Форбса «О связи между распределением существующей фауны и флоры Британских островов и геологическими изменениями в эпоху делювия» (1846). Форбс на много десятилетий определил развитие биогеографии, применив почти современные методы исследования. Для выяснения генезиса фаун он рекомендовал изучать геологические данные, начиная от современных и переходя к предшествующим эпохам, а не наоборот, как это делали до него. Форбс доказал, что в видовом составе организмов, населяющих Британию, можно распознать следы пяти типичных флор и фаун различной древности, которые последовательно сменяли друг друга, начиная от миоцена. Появление ряда видов растений и животных он объяснил бывшими материковыми связями Британии с Францией, Испанией и Скандинавией. Форбс был одним из первых биогеографов, исследовавших фауну и флору окружающих Британию морей.
Блестящим представителем биогеографии данного периода был швейцарский ботаник Альфонс Декандоль. Хотя он не касается проблем зоогеографии, его методология представляет огромный интерес как для ботаников, так и для зоологов. Опираясь на методику Форбса, Декандоль изучал распределение и распространение растений, учитывая условия окружающей среды, и предложил новый метод исследования. Он считал, что к помощи геологических данных нужно обращаться только тогда, когда современные физические условия не дают ответа для истолкования тех или иных явлений. Декандоль указывал на необходимость тщательного изучения видовых ареалов — основного и надежного материала для установления регионов флоры. Результаты своих наблюдений Декандоль опубликовал в «Географии растений» (1855) — самом выдающемся произведении додарвиновского периода развития биогеографии.
К четвертому периоду
(60-е гг. XIX в.) биогеография подошла к научному объяснению причин распространения организмов. Тем не менее представление о непрерывности эволюционного процесса, о происхождении современных фаун от давно исчезнувших для подавляющего большинства ученых первой половины XIX в. оставалось чуждым, несмотря на то, что оно было вполне убедительно аргументировано Ламарком в «Философии природы» (1809).
После выхода в свет труда Дарвина «Происхождение видов» (1859) стало ясно, что только эволюционное учение дает возможность понять причины, обусловившие современное распространение животных и растений, поскольку организмы в такой же степени подвержены медленным и постоянным изменениям, как и физико-географические условия, в которых они находятся. Из этого следует, что изменение видов тесно связано с изменением среды их обитания.
Новые идеи привели к бурному развитию биологических наук, в том числе и биогеографии, факты которой были использованы Дарвином в качестве доказательств эволюции.
Изучая характер распространения животных на островах вблизи Южной Америки, Дарвин пришел к выводу, что его можно объяснить лишь изменением видов во времени, так как разница между видами, населяющими острова, тем больше, чем значительнее и длительнее их географическое разъединение. Дарвин внес большой вклад в развитие биогеографии, показав роль географических преград для формирования эндемичных видов и своеобразных фаун, примером которых является фауна Галапагосских островов. Он доказал возможность заселения территорий организмами за счет случайных заносов — ветром, морскими течениями, перелетными птицами и другими способами. Дарвин также подтвердил и четко сформулировал закон о едином центре происхождения каждого вида. Разрывы между частями ареала одного вида он объяснил вымиранием его представителей в промежуточных районах. Только после Дарвина стало возможным развитие направления, которое изучает эволюцию фаун и называется исторической (генетической) зоогеографией. Историческая зоогеография получила мощное развитие в трудах К. Рютимейера и А. Уоллеса.
В работе «О происхождении животного мира» Рютимейер (1867) на основании палеонтологических данных и фактов современного распространения животных сделал попытку выделить на Земле фаунистические слои различной древности. Наиболее древней он считал фауну Мирового океана — колыбели жизни, утверждая, что от нее произошли обитатели пресных вод и суши. Из сухопутных фаун к самой древней он относил австралийскую с ее однопроходными и сумчатыми, являющимися потомками мезозойской фауны. К более молодым Рютимейер причислял африканскую и индийскую фауны (третичный возраст), а к самой молодой — мало изменившуюся ледниковую фауну Северной Америки и севера Старого Света.
Знаменитый сподвижник Дарвина Уоллес, самостоятельно пришедший к идее естественного отбора как главного фактора эволюции организмов, опубликовал двухтомный труд «Географическое распространение животных» (1876), в котором обобщил огромный зоогеографический материал, в том числе и данные палеонтологии о прошлом распространении видов. Уоллес использовал метод, предложенный Декандолем, тщательно изучив ареалы видов и более высоких таксономических единиц (родов, семейств), и, кроме того, учел историю происхождения фаун различных частей Земли.
В 1875 г. П.Склэтер статистическим методом установил естественные зоогеографические области Земли. Вместо бессистемного выделения мелких зоологических областей он обосновал существование небольшого числа крупных областей, характеризующихся эколого-фаунистической общностью.
Видоизменяя зоогеографические области Склэтера, Уоллес выделил б основных фаунистических: Палеарктическую, Эфиопскую, Восточную (Индо-Малайскую), Австралийскую, Неотропическую и Неарктическую. Каждая из них членилась на несколько подобластей. Уоллеса считают основоположником исторической зоогеографии, хотя по справедливости эту честь он должен делить с Рютимейером. Однако Уоллес мало внимания уделял экологическим факторам, воздействующим на распространение организмов.
Основоположником экологической зоогеографии по праву надлежит считать нашего соотечественника Н.А. Северцова (1827— 1885). Еще в додарвиновские времена в монографии «Периодические явления в жизни зверей, птиц и гад Воронежской губернии» (1855) Северцов, применив оригинальный метод, установил связь между особенностями фауны и теми физико-географическими условиями (климат, почва и т.п.), в которых она живет и развивается. Распространение и миграция животных объяснялись им исключительно на основе экологических факторов современности. В следующей крупной работе «Вертикальное и горизонтальное распространение туркестанских животных» (1873) Северцов обобщил материалы своих среднеазиатских экспедиций и пришел к выводу, что нынешнее распространение животных объясняется не современными, а давно прошедшими географическими и физическими условиями, которые открывает геология. Он выделил группировки животных по районам их первоначального возникновения и распространения: среднеазиатские, евросибирские, южноазиатские и т.д. Так в труде H-А.Северцова объединились экологическое и историческое направления в зоогеографии. На экологических принципах им было построено зоогеографическое районирование Палеарктики (1877), где за основу приняты ландшафтные зоны тундры, тайги, переходной, пустынной и прибрежной зон.
И Уоллеса, и Северцова можно считать основателями эволюционной зоогеографии. Северцовское направление развивали его ученики М.А.Мензбир и П.П.Сушкин.
Мензбир (1855—1935) разработал зоогеографический метод, согласно которому зоогеограф должен начинать с изучения видового состава, переходя от настоящего к прошлому. При районировании следует руководствоваться наличием или отсутствием видов, типичных для областей. Выделенные по фаунам зоогеогра-фические единицы должны иметь разное значение, так как фауны разных стран весьма отличаются друг от друга. На основе принципов зоогеографического районирования Мензбир выделил следующие зоогеографические единицы: область, подобласть, провинция, округ, участок, местоположение. При выделении областей он принимал во внимание богатство фауны и ее историческое прошлое. Прочие более мелкие единицы ученый выделял на основании современного состояния фауны, поскольку они в большинстве одинакового возраста и в их развитии огромную роль играет различие стаций. Разница в составе фаун устанавливалась при сравнении центров единицы, а не переходных полос.
П.П.Сушкин (1868—1928), гармонично объединяя экологическое и историческое направления в зоогеографии, разработал картину эволюции фаун Сибири и Центральной Азии. Он предложил гипотезу о былом существовании мощного центра формирования сухопутной фауны, располагающегося на северо-востоке Азии, — Берингии, которая соединяла северо-восток Азии и северо-запад Северной Америки. Этим он и объяснял большое сходство фаун севера данных материков.
Гипотеза Сушкина была в дальнейшем разработана Б. К. Штег-маном. Его работа «Основы орнитологического разделения Палеарктической области» (1938) замечательна попыткой разрешить трудности детального зоогеографического районирования суши путем проведения линейных границ. В качестве основных структур зоогеографии он рассматривал не региональные единицы, границы между которыми проводились относительно произвольно и статично, а изменяющиеся во времени и пространстве сосуществующие или антагонистические «типы фаун». При этом области взаимопроникновения двух фаун он обозначал на картах отдельными мазками соответствующих цветов. Почти одновременно с подобным предложением выступил зоогеограф В. Рейниг, который под «типом фауны» (или «кругом фауны», по терминологии многих зарубежных зоогеографов) подразумевал комплекс видов, принадлежащих к одному общему центру распространения.
Северцовское направление успешно развивал и крупный энтомолог А. П. Семенов-Тян-Шанский. Ему принадлежит основательно разработанная схема зоогеографических подразделений Палеарктики вплоть до провинций.
Л. С. Берг продолжил и развил намеченное Северцовым ландшафтное расчленение Палеарктики, связав ландшафты с определенными комплексами животных и растений. На основе распространения пресноводных рыб он первым произвел районирование Палеарктики. Им объяснен ряд зоогеографических парадоксов. К примеру, в статье «Биполярное распространение организмов и ледниковая эпоха» (1920) он выдвинул учение о биполярности, развивая его в биогеографическом и палеогеографическом направлениях. Известно, что ареалы ряда морских животных располагаются в умеренных широтах Северного и Южного полушарий с перерывом в тропиках. Такую разобщенность Берг объяснял событиями, происходившими во время ледникового периода. Аналогичным было и его учение об амфибореальности, т. е. о нахождении тех или иных видов и родов «на западе или на востоке умеренных широт и отсутствии посередине». Причина подобного распространения кроется также в палеогеографических условиях прошлых геологических эпох. Интересные зоогеографические проблемы решались Бергом при объяснении фаунистических загадок Каспия и Байкала. Вполне современно звучат высказывания Л. С. Берга о необходимости различать два принципа районирования моря: зонально-географическое и собственно зоогеографическое. В первом случае основой является зона, во втором — иерархические единицы: области, подобласти, провинции, которые выделяются по степени сходства (видового, родового и т. д.) их фауны.
Итак, во второй половине XIX в. и в первые десятилетия XX в. в науке преобладающее значение получило историческое направление. Наряду с этим первая половина XX
в. ознаменовалась усилением связи биогеографии с экологией.
Учет экологических факторов с целью объяснения закономерностей распространения видов в какой-то степени был характерен для многих работ и в прошлом. Это было особенно типично для России с ее громадными пространствами и сочетанием разнообразнейших условий среды. Впоследствии необходимость связи биогеографии с экологией диктовалась практическими соображениями. Исследование географического распространения видов показывало, что внутри ареала существуют формы со своеобразными местными особенностями, зависящими от сочетаний факторов географической обстановки. В результате тесных взаимоотношений биогеографии с экологией появились экологическая зоо- и фитогеография. Примером слияния зоогеографических идей с экологическими может служить книга А. Н. Формозова «Снежный покров как фактор среды, его значение в жизни млекопитающих и птиц СССР» (1946). Тесное переплетение двух наук приводит к тому, что отдельные зоогеографы, как, впрочем, и геоботаники, не видят разницы между экологией и биогеографией. Такое положение весьма характерно для американской биогеографии, хотя необоснованность его давно подмечена самими американскими учеными.
Другая крайность заключается в стремлении географов превратить биогеографию в часть ландшафтоведения, т. е. сделать ее обслуживающим разделом физической географии. Пограничное положение зоогеографии и фитогеографии на стыке биологических и географических наук бесспорно. Однако объект исследования — виды животных и растений — заставляет отнести биогеографию к биологическим наукам. Для характеристики ландшафта нужно знать не столько видовой состав, т. е. фауну, сколько типичное для него животное население. Обычно это многочисленные, широко распространенные, так называемые фоновые, или ландшафтные, виды. Они и служат объектом изучения геозоологии.
В XX
в. в результате обобщения огромного фаунистического материала появляются сводные монографические работы и ряд учебных руководств. Упомянем наиболее важные из них. Экологическое направление отражено в таких трудах, как «Основы экологической зоогеографии» Ф.Даля (1921—1923), «Зоогеография на экологической основе» Р.Гессе (1924), «Экологическая география животных» Р.Гессе, В.Элли и К.Шмидта (1951). Зоогеографии Мирового океана была посвящена работа С. Экмана «Зоогеография моря» (1935). Особое значение для развития зоогеографии в России и подготовки специалистов-зоогеографов имело руководство «Общая зоогеография» В.Г.Гептнера (1936). Большую ценность в нем представляет раздел, посвященный расселению животных и учению об ареале. Все последующие работы советских зоогеографов испытали на себе влияние этого труда. В выходивших впоследствии руководствах — «Зоогеография» И.И.Пузанова (1938), «География животных» Н.А.Бобринского, Л. А. Зенкевича, Я.А.Бирштейна (1946), «География животных» Н.А.Бобринского (1951) и др. — центр тяжести изложения переносился то на экологическую, то на историческую часть.
Ф.Дарлингтон (1957) на основе предложенных им методов и принципов описал характер распространения и вероятный ход расселения всех классов позвоночных животных, как водных, так и наземных. Большое внимание в ней уделяется закономерностям островного и материкового распространения животных. Дарлингтон является противником гипотез и теорий, часто упоминаемых в исторической зоогеографии (например, теорий перемещения материков и т. п.). Он считает, что незначительные изменения в очертаниях материков вместе с более широкими, чем это обычно признается, возможностями расселения сухопутных животных через морские преграды вполне объясняют современную картину распространения основных групп животных на земном шаре.
В 1967 г. была издана большая работа Г.Деллатина «Очерк зоогеографии». Касаясь лишь попутно общих вопросов, таких как теория ареала или экологические основы зоогеографии, главное внимание он уделил морской, пресноводной и сухопутной фаунам. Весьма ценен в книге раздел, посвященный фаунистическим элементам и их значению для выяснения истории фаун.
До недавнего времени в мировой зоогеографической литературе не было руководств по зоологическому картографированию. Большинство зоогеографических работ довольствовалось давно известными методами картирования ареалов животных. Методические трудности зоологического картографирования вытекают из специфики самих животных (подвижность, скрытый образ жизни, способность выбирать и создавать среду обитания, сезонные и годичные колебания численности и др.). Однако картирование животного населения особенно важно как для геозоологических целей, так и для проведения региональных зоогеографических работ. Практическая сторона изучения распространения животных, особенно таких, как охотничье-промысловые звери и птицы, вредители сельского и лесного хозяйства, переносчики и хранители инфекционных заболеваний человека, также настоятельно требует создания специальных карт зоологического содержания. Очевидна необходимость создания зоологических карт территорий, где проводятся крупные хозяйственные мероприятия — освоение целинных земель, мелиорация ландшафтов, прокладка магистралей, размещение новых населенных пунктов.
В связи с этим следует отметить книгу Н. В. Тупиковой «Зоологическое картографирование» (1969), в которой обосновывается целое направление в науке — зоогеографическое картографирование и рассматривается методика составления зоологических карт. В книге также содержится обзор основных типов зоологических карт: ареалов и изменений их границ, населения отдельных видов, биотопической приуроченности животных, фаунистиче-ских комплексов и животного населения.
Современный этап развития зоогеографии
характеризуется возникновением нескольких ее направлений. Часть из них лежит в русле классических традиций, другие же являются принципиально новыми. Прежде всего заслуживают внимания попытки обоснования и развития комплексной науки — биогеографии.
Наиболее последовательную позицию в этом вопросе занимает школа московских биогеографов, возглавляемая профессором А. Г. Вороновым. В подготовленных ими учебниках излагаются общие проблемы биогеографии и развернутые характеристики биомов земного шара. А. Г. Воронов определяет биогеографию как науку о географическом распространении и размещении сообществ организмов и их компонентов. Основную задачу этой науки он видит в установлении географической специфики причинных связей между средой обитания в целом и ее факторами, с одной стороны, и сообществами и их компонентами — с другой, из чего вытекает теснейшая взаимозависимость биогеографии, экологии и биоценологии, а также известное перекрывание всех трех наук. Особенностью биогеографии как самостоятельной комплексной науки он считает нацеленность ее на изучение таких проблем, как география продукции и массы живого вещества планеты, причины современного распространения сообществ организмов, географические особенности биотических отношений и антропическое влияние человека на биомы[3]
и биоты[4]
в разных географических условиях.
Наряду с этим направлением продолжает развиваться классическое фауно-генетическое направление. В последние десятилетия большой вклад в развитие этого направления внесли О.Л.Крыжа-новскийиЯ.И.Старобогатов. О.Л.Крыжановский (1965) детально проанализировал наземную фауну и ее генезис такого интересного и сложного региона, как Средняя Азия, и предложил схему ее зоогеографического районирования. Я. И. Старобогатов (1970) разработал принципы и методы зоогеографического районирования применительно к пресноводным животным и уточнил ряд общих закономерностей распространения фауны внутренних водоемов.
Особое направление, выделившееся еще в 60-е гг., — островная биогеография — заложено в работах Престона (1962) и Мак-Артура и Уилсона (1967). Они предложили новые подходы к изучению динамики островных фаун и обратили внимание на связи между числом видов и площадью острова, на соотношения процессов колонизации и вымирания (оборот видов), а также на фаунистический коллапс. Практическое развитие это направление получило только в последнее время, главным образом в охране природы. В эпоху антропического преобразования естественных ландшафтов остатки их, часто в виде заповедных территорий, становятся изолятами — островами в море сельскохозяйственных или урбанизированных территорий. Поэтому Престон считает, что к ним приложимы положения, применимые к настоящим островным популяциям организмов. Таким образом, в частности, можно решить вопрос о «минимальной динамичной площади» заповедника, которая обеспечит охрану видов, а также создать резерваты, состоящие из множества отдельных участков, своеобразных «архипелагов», совокупная фауна которых, как показали наблюдения, богаче, чем на единой большой площади.
В исторической зоогеографии выделилось направление, получившее название викариантной зоогеографии. Она рассматривает распространение географических изолированных таксонов как результат действия преграды, разделившей ранее единый ареал предкового таксона. Частным случаем викариантной зоогеографии является «мобилистская зоогеография», базирующаяся на представлениях о континентальном дрейфе. Ее сторонники пытаются согласовать схему филогенетического ветвления (кладограмму) с последовательностью расхождения фрагментов единого мезозойского континента Пангеи. Материал для суждения о хронологии геологических событий дает современная геофизика.
Многие вопросы и проблемы зоогеографии до настоящего времени остаются дискуссионными и оживленно обсуждаются на зоогеографических конференциях, которые регулярно проводятся в нашей стране. В настоящее время зоогеография из классической описательной науки переросла в науку остро проблемную, решающую практические задачи рационального использования животного мира.
Наша планета имеет несколько различных по плотности оболочек, окружающих ее ядро. Выделяют твердую оболочку — литосферу, водную — гидросферу, воздушную оболочку — атмосферу. Особую важность для человека представляет биосфера — живая оболочка Земли со всей совокупностью населяющих ее организмов. В пределы биосферы входит верхняя часть литосферы (кора выветривания) до глубины 30—60, редко до 200 м, вся гидросфера и нижний слой атмосферы до высоты 10—15 км (тропосфера). Кора выветривания и вся гидросфера постоянно заселены организмами, в тропосфере же животные и растения пребывают временно, поскольку здесь нет условий для такого важного процесса, как размножение.
Живые организмы занимают все пригодные для существования участки планеты. Даже поверхности ледниковых щитов, центральные районы жарких пустынь и горячие источники населены растениями и животными. На льду обитают микроорганизмы, водоросли и низшие насекомые — ногохвостки. Пауки, клещи и насекомые обнаружены на снегу в Гималаях на высоте 7 тыс. м. В безжизненных на первый взгляд местах пустынь Аравии и Южной Африки существуют бедные видами сообщества животных. По-настоящему лишены жизни лишь кратеры действующих вулканов и участки морей с чрезмерно высоким содержанием солей (Мертвое море) или отравленные сероводородом (придонные слои Черного моря).
Как уже говорилось, наибольшей плотностью обладает живой покров у самой поверхности Земли. Здесь и происходят основные взаимодействия живой и неживой природы. Эту тонкую пленку часто называют биогеоценотическим покровом, ландшафтной оболочкой или биогеосферой.
Биогеосфера неоднородна. Самыми крупными ее представителями являются биоциклы, их три: моря (океаны), пресные воды и суша. Наименьшим ее подразделением является биогеоценоз, т. е. совокупность живых и неживых компонентов наиболее однородного участка. Термин «биогеоценоз» предложен академиком В.Н.Сукачевым и определяется им как «совокупность на известном протяжении земной поверхности однородных природных явлений (атмосферы, горной породы, растительности, животного мира и микроорганизмов, почвы и гидрологических условий), имеющая свою общую специфику взаимодействия этих слагающих ее компонентов и определенный тип обмена веществом и энергией их между собой и другими явлениями природы и представляющая собой внутреннее противоречивое диалектическое единство, находящееся в постоянном движении, развитии». Все организмы биогеоценоза составляют биоценоз данного участка, или биотопа. Таким образом, биогеоценоз состоит из двух компонентов — биоценоза и биотопа. Понятию «биогеоценоз» приблизительно соответствует понятие «экосистема».
В биосфере не существует организмов, абсолютно независимых от своего окружения. В среде обитания животные подвергаются комплексному воздействию разнообразнейших экологических факторов. Эти факторы влияют прежде всего на их географическое распространение, обусловливают миграции и устранение отдельных видов из тех или иных биотопов; изменяя плодовитость и смертность, они определяют плотность популяций. Наконец, факторы среды обеспечивают выработку приспособлений к ним — адаптивные модификации в виде спячки, диапаузы, фотопериодические реакции и т. п.
Экологические факторы принято делить на абиотические, биотические и антропические. В категорию абиотических входят климатические (свет, температура и влажность воздуха, осадки, ветер) и эдафические (физические и химические свойства почвы) факторы. Под биотическими факторами подразумевается взаимодействие и взаимовлияние живых организмов. Антропические факторы — это сумма влияний, исходящих от человека.
Факторы среды изменчивы. Примером могут служить колебания температуры воздуха в течение суток и года или изменения ее при перемещении от экватора к полюсам. Нормальное существование животных наблюдается только при тех изменениях экологического фактора, которые находятся в пределах его минимального и максимального значений. Эти значения представляют собой границы толерантности (пределы выносливости) относительно данного фактора, его экологический диапазон (рис. 111).
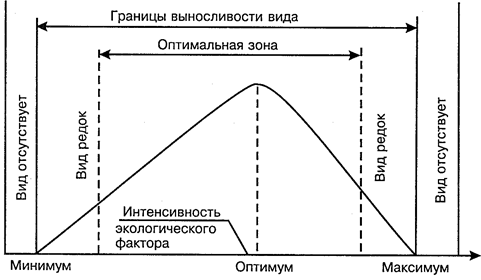
Рис. 111. Границы толерантности организмов в зависимости от
интенсивности экологического фактора (из Р.Дажо, 1975)
Между минимумом и максимумом, ограничивающими возможность существования животного, находится относительно узкая зона оптимума, наиболее для него благоприятная. Между зоной оптимума и границей, определяющей возможность существования вида, лежит зона пессимума, где организм оказывается в плохих условиях. Зона пессимума, как и зона оптимума, устанавливается по плотности популяций вида, т. е. по числу особей, приходящихся на единицу площади. Конечно, в зоне оптимума вид будет представлен более плотными популяциями.
По аналогии с химической валентностью экологи ввели понятие «экологическая валентность». Экологическая валентность вида — это его способность заселять различную среду, характеризующуюся определенными изменениями экологических факторов. Вид с низкой экологической валентностью, способный переносить лишь ограниченные колебания факторов среды, называют стенобионтным. Примерами стенобионтных видов сухопутных животных являются человекообразная обезьяна шимпанзе, обитающая в тропических влажных лесах, или наш глухарь — житель хвойного леса умеренной зоны. В противоположность этому вид, способный заселять различные местообитания или местообитания с изменчивыми условиями, называют эврибионтным. Примером эврибионтного вида может служить лисица, встречающаяся на равнине и в горах, в лесу и в степи, в пределах нескольких ландшафтно-климатических зон. К типичным стенобионтам, обитающим в водной среде, относятся рифообразующие кораллы, требующие строго определенных (высокой) температуры, солености и степени освещенности морской воды. Они распространены в пределах тропической зоны Мирового океана. Киты же, будучи эврибионтами, при наличии пищи живут в различных широтах.
Для выражения относительной степени толерантности употребляют соответствующие данным факторам термины с добавлением приставки «стено» (узкий) или «эври» (широкий). Так, относительно температуры животные бывают стенотермными или эвритермными, солености водной среды — стеногалинными или эвригалинными, пищевого рациона — стенофагами или эврифагами и т. д. Наконец, в отношении местообитания их можно разделить на стеноэков и эвриэков.
Фактор среды, ограничивающий возможности существования и распространения организмов, называется лимитирующим. Для распознавания лимитирующих факторов необходимо изучать виды на границах их ареалов.
Океаны и моря представляют собой самый крупный биоцикл Земли. На их долю выпадает 71 % поверхности планеты. В то же время они включают и наиболее богатую фауну, которая составляет 64 % видов животных, тогда как на сушу приходится только 36 %. Это и понятно, ведь жизнь зародилась в море, до настоящего времени здесь обитают представители очень многих классов животных, за исключением огромного большинства насекомых, многоножек и амфибий. Многие классы животных обитают только в море. К ним относятся коралловые полипы, плеченогие, боко-нервные и головоногие моллюски, бесчерепные, оболочники, губки, многощетинковые кольчецы, немертины и др. Вместе с тем следует отметить, что в океане и поныне есть представители очень древних групп животных, которые за миллионы лет изменились сравнительно мало. Это свидетельствует о замедленности темпов эволюции морских животных по сравнению с наземными.
Между наземными и водными средами наблюдаются существенные различия в их физико-химических свойствах. Особое экологическое значение для морских организмов имеют плотность, давление, глубина проникновения солнечной радиации, распределение тепла, содержание газов и растворенных солей, течения.
Среди морских животных по их отношению к давлению различают эврибатные и стенобатные виды.
На распределение животных в море сильное влияние оказывает свет, вернее, степень проникновения солнечной радиации, которая зависит от веществ, растворенных и взвешенных в воде. По мере увеличения глубины степень проникновения солнечной радиации уменьшается, причем довольно быстро. На глубине 1 м инфракрасное излучение полностью поглощается, а видимый свет наполовину слабее, чем на поверхности. На глубине 200-400 м света уже не хватает для существования растений. Большие глубины практически лишены освещения, и животные там обитают в темноте. Толщу воды в море принято делить на зоны: хорошо освещенную эвфотическую (от 0 до 30 м), дисфотическую (30— 200 м) и лишенную света афотическую (ниже 200 м).
Важную роль в жизни водных животных играет распределение в морях тепла. Источником его служит энергия солнечной радиации, поэтому распределение температур на поверхности и в верхнем слое воды зависит от климата соответствующей части земного шара, где располагается данный водный бассейн. В морях наблюдается также вертикальное зональное распределение температур. Однако эта зональность нарушается течениями.
Охлаждается вода из-за собственного излучения и испарения с поверхности моря. Благодаря постоянному перемешиванию слоев (из-за течений, ветра, конвекционных токов) изменения температур затрагивают значительную толщу воды. Что же касается больших глубин, то там имеется собственный температурный режим.
Содержание кислорода в морской воде колеблется незначительно. Насыщение им осуществляется в верхних слоях, где живут растения и наблюдается волнение и перемещение вод.
Большое значение имеет соленость морской воды. В открытом океане средняя концентрация растворенных солей равна 3,5 г/л (35 %о), в тропических морях, где сильное испарение, она бывает выше, а в полярных водах ниже, особенно летом (из-за тающих льдов). Соленость морской воды подвержена значительным пространственным и сезонным колебаниям. Существенные изменения ее отражаются на распространении стеногалинных организмов и определяют состав морской фауны. Так, рифообразующие кораллы — типичные стеногалинные формы — крайне чувствительны и к небольшому опреснению воды. Поэтому коралловые рифы прерываются против устьев даже небольших рек. Эврига-линные организмы распространены более широко, чем стеногалинные. В таких морях, как Балтийское, наблюдается закономерное изменение фауны по градиенту солености: от пролива Каттегат до Ботнического залива соленость падает с 32 до 3 %о, и параллельно этому уменьшается число морских видов рыб, моллюсков, раков и др.
Важнейшим фактором существования и распространения морских организмов являются течения. Они влияют на распределение температуры в море, смещая его температурные зоны, а также на соленость отдельных участков. Основные океанические течения описывают гигантские круговороты. Различают течения теплые и холодные. Первые возникают в тропической зоне, вторые приносят воду из приполярных областей. Некоторые течения проходят в определенном направлении и постепенно сходят на нет (Гольф-стрим), другие образуют замкнутый круг (Экваториальное проти-вотечение в тропической части Атлантики).
Особое значение течения имеют для пассивно плавающих форм, например мелких ракообразных. Благодаря течениям в центр Атлантики регулярно приносятся и крупные водоросли саргассы с располагающимися на них мшанками, червями, гидроидами, асцидиями и другими животными, обитающими у берегов Южной и Центральной Америки. Крупные морские животные — рыбы, водяные змеи и др. — также регулярно заносятся в новые для них районы океана.
В море можно выделить две основные группы биотопов в зависимости от того, имеется у них твердый субстрат в виде дна (берега) или нет. Огромную толщу морской воды, где жизнь не связана с твердым субстратом, называют пелагиалью; вся же поверхность дна, включая берег, именуется бенталью (рис. 112).
Обитатели пелагиали в течение всей жизни пассивно или активно плавают. Группа животных, активно передвигающихся благодаря специальным органам, составляет нектон. В нее входят рыбы, китообразные, крупные головоногие моллюски (например, кальмары). Распространены они, как правило, весьма широко, нередко по всему свету. В океанах к таким видам относятся: из млекопитающих касатка, из рыб тунец и некоторые акулы и целый ряд других животных. Организмы, пассивно перемещающиеся, представляют собой планктон. Большинство из них совершает плавательные движения, но они обеспечивают лишь незначительные, в основном вертикальные перемещения, не позволяющие противостоять течениям. Планктонные животные имеют специальные приспособления, уменьшающие массу тела или увеличивающие его поверхность.
Вертикальное расчленение бентали гораздо сложнее. Здесь различают литоральную и абиссальную зоны. Первая охватывает поверхность континентального шельфа (материковой отмели) от зоны заплеска (участков, не заливающихся водой, но постоянно увлажненных брызгами морской воды) до начала материкового ската. Абиссальная зона включает морское дно и прилегающий к нему слой воды от края материкового ската. Резкой границы между литоралью и абиссалью нет, их разделяет переходная батиальная зона.
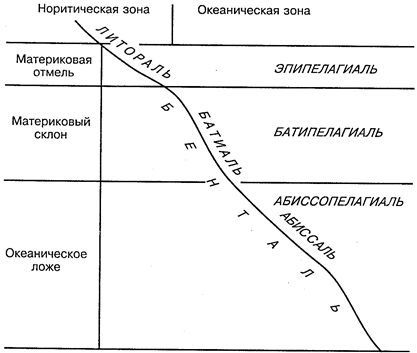
Рис. 112. Вертикальное расчленение океана
(по А.В.Константинову, из В.А.Радкевича, 1977)
Область континентального шельфа невелика (лишь 7,5 % площади дна Мирового океана), но чрезвычайно богата животным населением. Объясняется это разнообразием биотопов (наличие различных грунтов и зарослей подводных растений, опресненность прилегающих к устьям рек участков и т. д.) и обилием пищи. Пищей служат заросли водорослей и морских трав на мелководьях, где для них имеются хорошие условия освещенности, а также органические вещества, приносимые материковыми водами.
Абиссальная зона — глубоководная часть моря. По площади она во много раз превышает другие зоны, поскольку к ней относится около 80 % морского дна. В отличие от пелагиали здесь постоянно царит тьма, держится ровная температура (в Северной Атлантике 3 "С, в Антарктике 0,5 °С), течения весьма слабые, соленость высокая, грунт очень мягкий, илистый, а давление чрезвычайно велико. Такой комплекс условий почти не меняется на всем протяжении абиссали Мирового океана. Поэтому она считается одной из самых однообразных сред нашей планеты. Пищевые условия абиссали тяжелые. Ресурсы пищи представлены в основном детритом, падающим сверху. Погружаясь, он разлагается бактериями, и до дна доходит лишь небольшая его часть. Вот почему в глубоких водах не бывает изобилия животных. Главными обитателями здесь являются иглокожие. Они питаются илом. Есть на дне и фильтраторы — актинии, черви, моллюски, стеклянные губки, а также крабы и морские пауки-пантоподы, часть из которых де-тритоядные, другие хищники.
Постоянство условий существования в абиссали привело к тому, что здесь сохранилось много реликтов — древних животных, которые в других участках Мирового океана давно вымерли. Это стебельчатые морские лилии, рыбы-химеры и ряд других. В фауне абиссали есть и представители молодых групп животных, что свидетельствует о заселении абиссали выходцами из пелагиали и ли-торали.
Пресные воды представляют собой второй, хорошо отличающийся в зоогеографическом отношении, биоцикл. Сюда относятся озера, реки и ручьи, образующиеся за счет атмосферных осадков. Площадь пресных водоемов сравнительно невелика — около 4 млн км2
. Условия среды в них резко отличаются от морских и характеризуются прежде всего пестротой и большим размахом колебаний. Сроки существования пресных водоемов по сравнению с морскими очень коротки, кроме того, очертания их довольно быстро меняются. Все материковые водоемы неглубоки, в среднем менее 100 м, за исключением озер типа Байкала (около 1600 м) или Танганьики (1435 м). Реки вообще во всех отношениях не могут сравниться с морями.
В отличие от Мирового океана, все части которого сообщаются между собой и в целом представляют единый бассейн, пресные водоемы разобщены. Прямых связей между животными, населяющими их, может вообще не быть, как это наблюдается, к примеру, между бассейнами рек, изолированными водоразделами. Среди населения пресных вод совершенно отсутствуют иглокожие, головоногие моллюски, оболочники, очень мало видов из класса немертин, многощетинковых кольчецов. Вместе с тем здесь встречаются единичные представители губок и кишечнополостных. Только с пресными водами связаны земноводные. Тем не менее пресноводная фауна происходит от морской, и вселение животных из моря в реки и озера продолжается и сейчас. Это можно наблюдать в тропическом поясе.
При всем различии пресноводных бассейнов им свойственны общие факторы среды, влияющие на распространение в них живых организмов. Таковыми прежде всего являются химизм, температура, наличие или отсутствие движения воды. В отличие от моря в пресных водоемах давление практически не играет большой роли.
Химизм воды — фактор изменчивый. Главная составная часть солей морской воды — хлористый натрий — в пресноводных бассейнах отсутствует. Только некоторые озера содержат солоноватую или соленую воду, однако колебания солености в них весьма значительны — от 0,3—0,5 %о (совершенно пресные водоемы) до 12—17%о (Каспий) и даже 230 %о (Мертвое море). Некоторые животные приспособлены к колебаниям солености. Это эвригалин-ные организмы. Другие же — стеногалинные — строго избирательны к степени солености.
Существенным фактором для пресных водоемов служит жесткость воды (содержание извести). Так, губки, мшанки и определенные раки живут только в мягкой воде.
Важное значение для пресноводных животных имеет и содержание гумуса, а также количество растворенного в воде кислорода. Гумусные водоемы (лесные реки, болота) обычно бедны жизнью, в них мало не только рыб и моллюсков, но и личинок комаров. Разложение органических веществ здесь приводит к обеднению воды кислородом и в результате — к массовой гибели рыб, к так называемым заморам. Это особенно опасно для стеноксибионтов, требующих много кислорода. Обеднение водоемов кислородом вызывается также сбросом в них промышленных и бытовых отходов.
Немалое значение в жизни пресных вод имеет температура. Температурный режим континентальных водоемов определяется широтой местности и ее климатом. Кроме того, резкие температурные скачки могут наблюдаться в одном и том же водоеме. Поэтому большинство обитателей внутренних вод относится к эвритермным организмам.
Свет в пресных водоемах благодаря их преимущественной мел-ководности не играет особой роли. Обычно он проникает до дна и обусловливает развитие богатой водной растительности. Последняя поставляет большое количество кислорода и служит кормовой базой для растительноядных животных. Только в глубоких озерах типа Байкала существует настоящая афотическая зона.
Из механических факторов в пресноводных бассейнах при отсутствии приливов и отливов и ослаблении волнения весьма существенны течения. Амплитуда колебания скорости движения пресной воды огромна — есть стоячие водоемы (озера) и в противоположность им известны горные ручьи с бешено мчащимся и прыгающим по камням потоком. Пресноводные животные весьма чувствительны к скорости движения воды и в связи с этим разделяются на обитателей текучих вод — реофилов и любителей стоячей воды — лимнофилов.
В целом обитателей пресных вод следует отнести к эврибионтам. Однако различие условий жизни в стоячей и текучей воде обусловливает и разный характер распространения в них животных.
Условия существования животных организмов, обитающих в биоцикле «суша», резко отличаются от предыдущих. На суше наблюдаются гораздо более широкие изменения всех факторов среды обитания, чем в море или пресных водоемах.
Особое значение здесь приобретает климат и прежде всего одна из его составляющих — влажность воздуха, под влиянием которой и формировалась сухопутная фауна.
Главнейшими факторами, определяющими существование и распространение сухопутных животных, наряду с влажностью, служат температура и движение воздуха, солнечный свет, растительный покров. Пища здесь играет не меньшую роль, чем в других биоциклах, химизм же среды практически не имеет значения, поскольку атмосфера всюду одинакова, если не считать местных отклонений, обусловленных промышленными выбросами в атмосферу, о чем будет сказано далее.
Влажность воздуха в различных регионах Земли неодинакова. Изменение ее может вызвать разные реакции у животных. Если исключить организмы, нормальное существование которых не зависит от влажности, остальные животные будут либо влаголюбивыми — гигрофилы, либо сухолюбивыми — ксерофилы.
Влажность воздуха и почвы зависит от количества осадков. Следовательно, осадки оказывают на живые организмы опосредованное влияние. Вместе с тем осадки могут быть и самостоятельным фактором. Определенную роль играет, к примеру, форма осадков. Так, снежный покров зачастую ограничивает распространение видов, добывающих корм на земле. Например, хохластый жаворонок зимой не встречается севернее границы района относительно малоснежной и короткой зимы. С другой стороны, глубокий снег позволяет отдельным видам (сибирский лемминг и другие мелкие зверьки) перезимовывать и даже размножаться в зимний период. В снежных пещерах и тоннелях укрываются от холода нерпы и их враги — белые медведи.
Температура играет огромную роль в жизни обитателей суши, гораздо большую, чем в океане. Это объясняется большей амплитудой ее колебания на суше. Температура — прекрасный показатель климатических условий. Она часто более показательна, чем другие факторы (влажность, осадки). Средняя температура июля характеризует лето, января — зиму. Напомним, что воздействие температуры на организмы на суше более опосредовано другими климатическими факторами, чем в море.
Каждый вид имеет собственный диапазон наиболее благоприятных для него температур, который называется температурным оптимумом вида. Разница диапазонов предпочитаемых температур у разных видов очень велика. Если пределы температурного оптимума вида широки, он считается эвритермным. Если же этот оптимум узок и выход за пределы температурного лимита вызывает нарушение нормальной жизнедеятельности вида, последний будет стенотермным. Сухопутные животные более эвритермны, чем морские. Большая часть эвритермных видов населяет умеренные климатические зоны. Среди стенотермных могут быть термофильные, или политермные (теплолюбивые), и термофобные, или олиготермные (холодолюбивые) виды. Примерами последних служат белый медведь, мускусный овцебык, моллюски рода Vitrina, многие насекомые тундры и Альпийского пояса гор. В общем количество их сравнительно невелико хотя бы потому, что фауна холодных зон гораздо беднее по сравнению с другими. Стенотермных теплолюбивых видов значительно больше. Практически вся фауна тропиков земли, а это самая большая по числу видов фауна, состоит именно из них. Сюда входят целые классы, отряды, семейства. Типичными стенотермными теплолюбивыми животными являются скорпионы, термиты, рептилии, из птиц — попугаи, туканы, колибри, из млекопитающих — жирафы, человекообразные обезьяны и многие другие.
Кроме того, на суше существует немало эвритермных форм. Это изобилие обусловлено чрезвычайной изменчивостью температуры на суше. К эвритермным животным относятся многие насекомые с полным превращением, серая жаба Bufobufo, а из млекопитающих — лисица, волк, пума и др. Животные, хорошо переносящие значительные колебания температур, распространены гораздо шире, чем стенотермные. Нередко ареалы эвритерм-ных видов простираются с юга на север через несколько климатических зон. К примеру, серая жаба населяет пространство от Северной Африки до Швеции.
Кроме указанных факторов среды в жизни наземных животных важную роль играет свет. Однако непосредственная зависимость, как это наблюдается у растений, здесь отсутствует. Тем не менее она есть. Это выражается хотя бы в существовании дневных и ночных форм. Следует отметить, что играет роль не само по себе освещение, а сумма света. В тропическом поясе этот фактор особого значения не имеет в силу своего постоянства, но в умеренных широтах положение меняется. Как известно, длина светового дня там зависит от времени года. Только длинным полярным днем (продолжительность его несколько недель) можно объяснить тот факт, что перелетные птицы Крайнего Севера успевают за короткое время вывести и выкормить птенцов, поскольку пищей им служат насекомые, а они деятельны круглые сутки. Изобилие света продвигает на север границы жизни многих видов. Короткий же зимний день не позволяет даже холодолюбивым птицам получить достаточное количество корма для компенсации энергетических затрат, и они вынуждены откочевывать на юг.
Мощным фактором, регулирующим жизненный цикл ряда животных, служит длина светового дня. Явление фотопериодизма, в объяснение которого существенный вклад внес советский зоолог А.С.Данилевский, обусловливает развитие определенного количества поколений у насекомых в течение года, а также возможность расширения ареалов животных в другие широтные пояса.
К существенным климатическим факторам нужно отнести и ветер. На земном шаре есть места, где он дует постоянно и с большой силой. Это особенно характерно для морских побережий и островов. Здесь, как правило, нет летающих насекомых — бабочек, мух, мелких пчел, ос, в то время как на близко расположенном материке они обитают. Отсутствие этих насекомых влечет за собой и отсутствие летучих мышей, питающихся ими. Для океанических островов типичны бескрылые насекомые, что уменьшает для них степень риска оказаться в море. Таким образом, ветер в известной мере определяет состав фауны.
Характер субстрата, т. е. грунта, также играет важную роль в жизни сухопутных животных. При этом имеет значение не только химизм почвы, но и ее физические свойства. Наблюдается зависимость распространения животных от наличия солей в почве. Наиболее чувствительны к засолению почв членистоногие. К примеру, жуки рода Bledius, как и многие жужелицы, обычно встречаются только на засоленных почвах. Такие животные относятся к галофильным. Многие животные чувствительны и к типу горных пород. На известковых скалах, например, обитают моллюски, раковина которых построена из извести.
Однако чаще химизм почвы оказывает на животных опосредованное влияние, в частности, через кормовые растения. Роль пищевого фактора в жизни животных общеизвестна. Как уже говорилось, животные, будучи гетеротрофами, в целом существуют за счет растений, используя только готовые органические соединения. Следует отметить, что видовое разнообразие растений и животных на суше создает ряд особенностей, характерных именно для наземных экосистем.
Во-первых, в наземных экосистемах преобладают раститель-ноядные животные, т. е. фитофагов больше, чем зоофагов. Во-вторых, животные здесь связаны с высшими растениями, а не с водорослями, как это наблюдается в море. В-третьих, фитофаги наземных экосистем отличаются большой избирательностью по отношению к виду кормового растения. Это так называемые стенофаги. Животные, проявляющие малую избирательность к виду корма, относятся к эврифагам. Крайняя стенофагия, т. е. когда животное питается только одним видом растения, называется монофагией. Она наблюдается, например, у гусеницы тутового шелкопряда. Стенофагия имеет большое значение, так как от распространения кормового растения зависит и распространение его потребителя. С другой стороны, истребление этого растения вызывает исчезновение животного, связанного с ним.
Немало среди животных и зоофагов. Это хищники и паразиты. Среди них также встречаются стенофаги, но в целом для зоофагов, и прежде всего для хищников, характерна смена кормовых объектов.
Особенности питания животных влияют не только на их распространение, но и на биологию, на сезонные передвижения, или миграции.
К числу важнейших факторов среды, от которых зависит существование и распространение животных, относится растительный покров, определяемый, в свою очередь, особенностями климата и почв. Растительный покров обусловливает характер биогеоценоза и является его индикатором. У каждой растительной формации свой набор видов животных. Так, в хвойных лесах нашего Севера, где растут брусника, багульник, зеленые мхи и другие растения, свойственные тайге, мы обязательно найдем глухаря, синицу-московку, кедровку, клестов, бурундука, соболя, рысь. К лиственным европейским лесам, состоящим из дуба, бука, липы, ясеня, приурочены сони, кроты, землеройки, ежи, благородный олень, лань, лесной кот, барсук, орлы (змееяд, карлик), дикий голубь, сова-сплюшка, дубонос, иволга, болотная черепаха, квакша. Степным и пустынным формациям также свойствен специфический комплекс видов. Отсюда следует, что распределение биоценозов на земном шаре подчиняется определенным законам, зависит главным образом от климата и имеет зональный характер.
На земле различают тропическую зону, две полярные и две переходные умеренные зоны. Для каждой из них характерны свои растительные формации и связанные с ними группировки животных.
Наиболее типичным биотопом тропической зоны является гилея, или влажный тропический лес. Для произрастания такого леса необходима высокая температура и достаточное увлажнение в течение круглого года, незначительные сезонные колебания температур, не превышающие 8 °С, и средние годовые их значения не ниже 20 °С, чаще 25-26 °С. Максимальная температура в этих лесах у экватора доходит до 35 °С, а суточные колебания ее составляют 3—15 °С. Осадков в гилее обычно выпадает не менее 2000 мм в год. Высокая и постоянная влажность, стабильно высокая температура, отсутствие ветра создают уникальный комплекс условий, в первую очередь, для растительности. Растения здесь плодоносят весь год. В экваториальных лесах обращает на себя внимание многоярусность, громадное видовое разнообразие деревьев и по-лидоминантность, т. е. на значительном пространстве не отмечается преобладание того или иного вида.
Необычная тропическая среда населена своеобразными животными. По количеству видов и жизненных форм при немногочисленности особей биоценозы гилей не знают себе равных. Этот биотоп, кроме всего прочего, предоставляет животным огромное количество убежищ и экологических ниш, больше, чем другие наземные биотопы. Естественно, что обитатели влажного тропического леса термо- и гигрофильны.
Между тропическими лесами, с одной стороны, и пустынями — с другой, простирается саванна. Саванные растительные формации возникают в районах с жарким климатом, где наблюдается периодичность выпадения осадков, т. е. сезон дождей сменяется сухим сезоном. Запасов воды в саванне недостаточно для произрастания лесов. Вместо них формируются разреженные древостой, иногда — отдельные группы деревьев. Это придает саванне вид парка. Необозримые пространства здесь заняты высокими травянистыми растениями — так называемой слоновой травой и др.
В сезон дождей саванна напоминает зеленое море: осадков выпадает много, температура высокая, бурно развивается растительность. В сухой же сезон влаги поступает меньше, чем испаряется, вегетация растений прекращается, травы засыхают, а деревья сбрасывают листья. В это время в саванне часты пожары, вызываемые иногда естественными причинами, но обычно траву выжигают местные жители.
Наиболее типичны саванны для Африки. Они занимают огромные территории к югу от Сахары, за исключением гор и влажных тропических лесов бассейна Конго. Саванны располагаются также на полуострове Индостан в Азии и в Южной Америке к северу и югу от района влажных тропических лесов. Здесь их называют парамо.
Фауна саванн богата и разнообразна и отличается особыми жизненными формами, принадлежащими к специфическим для нее систематическим группам животных. Это обусловлено двумя причинами: периодичностью выпадения осадков и отсутствием лесов. Животные здесь теплолюбивы, но не стенотермны. Кроме того, они приспособлены к периодическим засухам. В связи с этим у них наблюдается сезонная ритмика жизненных проявлений, в частности летняя спячка.
Среди животных саванн преобладают бегающие и роющие формы. К первым кроме копытных относятся хищники. Хищных млекопитающих в саванне вообще много. Львы и леопарды охотятся за копытными, кошки и виверры — за мелкими антилопами, грызунами и птицами, гиены и шакалы нападают на ослабленных и больных животных, не брезгуя и падалью. К типичным бегающим формам из птиц, встречающихся в саванне, относятся страусы, птицы-секретари, марабу, дрофы, рябки. На деревьях колониями гнездятся ткачики. Роющие животные представлены в основном грызунами из семейств мышиных и беличьих. Они питаются семенами, плодами и луковицами растений. Любопытно, что там, где много копытных, мало грызунов, и наоборот. В саваннах обитают многочисленные термиты, строящие большие гнезда, так называемые термитники, которые иногда достигают высоты 2 м и более.
На границах тропической и умеренных зон, большей частью в центре материков, располагаются пустыни, занимающие 23% площади континентов. В Восточном полушарии пояс пустынь тянется от Северной Африки (Сахара) через Аравию, Западный Индостан, Среднюю Азию, Казахстан до Центральной Азии. Центральные части Австралии также представляют собой пустыни. В Южной Африке простираются пустыни Калахари и Намиб, в Западном полушарии — пустыни Атакама (в Чили), Мохаве и ряд других.
Пустыни характеризуются комплексом признаков, главными из которых являются сухой климат (незначительное количество осадков при сильном испарении влаги), высокие температуры воздуха летом и низкие — зимой (в пустыне Гоби амплитуда колебаний их достигает 80-90 °С), недостаточное увлажнение верхних слоев почвы и глубокое залегание грунтовых вод, перегрев поверхности почвы, подвижность субстрата и нередкая его засоленность. Режим увлажнения в пустынях различного типа разный. В некоторых пустынях осадки выпадают летом, а засуха наступает зимой. В других, наоборот, осадки характерны для зимы, а засуха — для лета. В отдельных пустынях выраженного сезона дождей может не быть. Наконец, в так называемых пустынях туманов осадков вообще нет, а наблюдаются частые туманы. Однако при всем разнообразии режимов влажности в пустынях годовое количество осадков там обычно не превышает 100—200 мм. В пустынях Средней Азии и Казахстана, к примеру, в разных районах оно колеблется от 55 до 180 мм.
По характеру субстрата пустыни бывают песчаные, глинистые, щебнистые (гаммады), лёссовые, солончаковые и т.д.
Условия существования организмов в пустынях очень суровы. Растения здесь встречаются редко и не образуют сомкнутого покрова. Это либо сухие и колючие травы, либо полукустарнички и кустарники с мелкими кожистыми листьями и часто с колючками, либо, наконец, растения-суккуленты с сочной мякотью (кактусы, опунции, молочаи, солянки). В пустынях, где есть сезон дождей, появляются однолетники-эфемеры, успевающие за очень короткое время прорасти, созреть и дать семена.
Большинство животных пустынь ксерофилы и эвритермы, но у них есть пределы температурной выносливости. Насекомые, например, погибают при 50—55 "С, ящурки не могут находиться на раскаленном песке дольше 4 мин, тушканчики гибнут при 34 °С. Чтобы уберечься от перегрева, одни животные закапываются в грунт или сидят весь день в норах, другие забираются на веточки кустарников.
С другой стороны, малое количество убежищ, редкая тень кустарников, раскаленный субстрат вынуждают животных искать спасения в быстром беге. К таким животным относятся, например, некоторые грызуны (кенгуровые крысы), а из насекомоядных — прыгунчики. Классическим примером стремительных бегунов служат тушканчики. Их задние лапы удлинены, передние укорочены. Длинный хвост выполняет роль балансира и руля при быстром прыгающем беге, представляющем собой серию прыжков на задних ногах. Тушканчики идеально приспособлены к жизни в пустынях и прекрасно переносят недостаток воды. Почки их вырабатывают очень концентрированную мочу. Кал у них полусухой, а потовые железы отсутствуют. Кроме того, тушканчики вообще не пьют, они довольствуются метаболической водой.
В целом сухость воздуха и отсутствие водоемов (или они встречаются очень редко) обусловливают выработку у пустынных организмов целого ряда приспособлений, позволяющих им долгое время обходиться без воды. Многие животные, особенно насекомые, могут вообще не пить. Влагу они получают от растительного или животного корма. Физиологические процессы у них направлены на экономию воды, в частности, для них характерна способность использовать метаболическую воду, образующуюся при окислении пищи. Ряд животных запасают воду в организме. Те же виды, которым питьевая вода нужна, совершают переходы или перелеты к источникам или водоемам, располагающимся порой на расстоянии 200—300 км (например, рябки).
В самое жаркое время года некоторые животные пустынь впадают в спячку, которая, к примеру, у степной черепахи или у желтого суслика продолжается без перерыва 8—9 месяцев, захватывая и зимнее время.
Хищников в пустыне немного, и они обычно не бывают крупных размеров. Это лисицы фенек и корсак, барханная кошка и манул, рысь каракал, хорь-перевязка.
В целом пустынные биоценозы отличаются бедностью видового состава и простотой структуры. В то же время они очень чувствительны к антропическим воздействиям. В пустынях добываются нефть и газ, строятся оросительные каналы, создаются водохранилища, прокладываются дороги. Издавна здесь пасут овец, охотятся и заготавливают топливо. Эта деятельность в какой-то степени нарушает природный комплекс, а если она осуществляется не на научной основе, то это приводит к исчезновению многих видов растений и животных. В результате разрушаются целые экосистемы. Охрана животного мира пустыни, сохранение ее неповторимого фаунистического комплекса — важная задача современности.
В условиях континентального климата при годовой сумме осадков 200—500 мм на границах пустынь, с одной стороны, и лесов — с другой, образуются степи. Полоса степей в Евразии тянется от Восточной Венгрии до Забайкалья. В Северной Америке они называются прериями, в умеренной зоне Южной Америки — пампасами. Климат степей континентальный, контрастный. Летом здесь жарко и сухо. В наших степях среднеиюльские температуры составляют 20—23,5 °С, зима малоснежная и довольно холодная, нередки морозы до 40 °С, а в Зауралье — до 50 °С.
Степи характеризуются густым травяным покровом, развивающимся на черноземах или каштановых почвах. Древесная растительность отсутствует на плоских водоразделах и ютится в долинах рек, пересекающих степную зону. Летом, когда вегетация многих растений прекращается, наступает период полупокоя. Полный покой наблюдается зимой. В наших степях растительный покров составляют ковыли, типчак, корневищные злаки, севернее — разнотравье. В американских прериях растут те же ковыли, мятлик, бородач, но разнотравья здесь больше, чем в Евразии.
Животный мир степей характеризуется обилием фитофагов, особенно грызунов, живущих в норах. Это многочисленные суслики, сурки, полевки, а в Северной Америке — луговые собачки и суслики. Когда-то у нас в степях бродили стада копытных: дикие лошади-тарпаны, а также туры и сайгаки. Из них только сайгаки дожили до наших дней, но они вытеснены человеком в полупустыни Прикаспия. В американских прериях водились бизоны, однако увидеть их сегодня можно лишь в национальных парках.
Обилие грызунов создает хорошую кормовую базу для хищников. В степях обычны лисица, степной хорек, нередки волки. На грызунов охотятся и хищные птицы — орлы-могильники, луни, мелкие соколы. Наряду с грызунами степные хищники поедают большое количество насекомых, которых в степи очень много. Это разнообразные саранчовые, муравьи, листогрызущие жуки и т. д. Среди них есть массовые и вредные виды, размножающиеся периодически в огромном количестве и уничтожающие растительность.
На юге степи переходят в полупустыни, а на севере проявляется промежуточная зона — лесостепь. Разнообразие растений и животных в лесостепи возрастает, так как здесь есть все условия для существования животных, связанных с древесными породами. Некоторые ученые считают лесостепь самостоятельной ландшафтной зоной.
Леса развиваются в тех областях умеренных зон, где годичная сумма осадков превышает 300 мм. В южных частях своего пояса леса произрастают благодаря осадкам, в северных же, не страдающих от засухи, — только благодаря температуре и продолжительности вегетационного периода. В связи с этим тайга опоясывает север земного шара сплошным кольцом, а лиственные леса приобретают вид больших прерывистых массивов. Различают три основных типа лесов умеренной зоны: субтропические вечнозеленые, широколиственные листопадные и хвойные (тайга).
Широколиственные листопадные леса развиваются в областях с умеренной температурой, без летней жары и сильных зимних морозов, где количество осадков составляет не менее 500 мм в год, причем распределяется оно по сезонам сравнительно равномерно. Эти леса произрастают главным образом в странах с морским климатом. В центре материков они исчезают. В Евразии широколиственные леса тянутся постепенно суживающейся полосой от атлантического побережья Западной Европы до Урала, а затем после большого перерыва снова появляются в Приморском крае России, в Японии, Корее, Китае. В Западном полушарии они располагаются на востоке США. Породообразующие деревья этих лесов — дуб, липа, клен, ясень, бук, а также дикие плодовые.
Относительно мягкий климат, разнообразие растительности умеренных широт — основные условия существования животных. Но зима в этих областях довольно холодная, и это заставляет животных мигрировать к югу либо впадать в состояние спячки или диапаузы.
Состав и экологический облик фауны данной зоны всюду сходен, причем в разных частях ее сохраняется определенное соотношение древесных и наземных видов, фитофагов и хищников. Из копытных здесь встречаются благородный олень и изюбр, серна, дикий кабан. Зубр дожил до наших дней только в заповедниках. В кронах деревьев обычны белки и сони, с водоемами связаны бобры. Птицы разнообразны, особенно многочисленны певчие дрозды, соловьи, зарянки, иволги, из голубей обычен вяхирь. Среди насекомых масса фитофагов, питающихся листьями, побегами, корнями и плодами деревьев. Здесь часто наблюдаются вспышки массового размножения насекомых, но они редко происходят в естественных лесах, где равновесие между компонентами не нарушено. Однако лиственные леса Европы и востока США так давно используются человеком, что это равновесие практически нарушено повсюду. Созданные же искусственные леса отличаются бедностью породного состава и требуют постоянного ухода, в частности борьбы с вредителями.
Самый крупный тип леса — тайга, или хвойный лес, состоит из зарослей ели, пихты, кедра, лиственницы и сосны и имеет вид сплошной зоны, распространяющейся от Атлантики до Тихого океана. Большая часть хвойных лесов сосредоточена в Евразии (55 %), немало их и в Северной Америке. В Южном полушарии аналога тайги не существует.
Климат тайги суровый. Среднегодовые температуры тут колеблются от 5 до О "С. Лето короткое и относительно жаркое (средне-июльская температура 13—14°С), зима продолжительная, многоснежная, холодная, морозы нередко достигают 30 и даже 40 "С. Важным фактором является вечная мерзлота, что, в частности, типично для тайги Восточной Сибири.
Суровые климатические условия тайги — причина бедности видового состава как растений, так и животных. Последним свойственна долгая зимняя спячка (у зимоспящих видов), способность создавать зимние запасы корма, ряд морфологических приспособлений (густое оперение или шерстный покров, белая окраска зимой и т. д.). Наиболее характерными обитателями тайги являются рябчик, глухарь, бородатая и ястребиная совы, кукша, кедровка, клест и черный дятел желна. Из зверей только в тайге обитают соболь, лесной лемминг, красная полевка. Бурундук и белка-летяга также живут в этой зоне.
Для питания животных тайги особенно большое значение имеют семена хвойных деревьев, в том числе кедровые орешки. В основном ими питаются кедровка, дятел, белка, бурундук. В рационе соболя и медведя орешки тоже занимают важное место. У птиц — потребителей семян хвойных — строение клюва приспособлено для добывания корма из шишек. К примеру, клюв у клеста пинце-тообразный, у щура крючковидный, у кедровки долотообразный. Подобная специализация ведет к постоянным перекочевкам в поисках шишек. Периодичность урожаев обусловливает колебание численности птиц, дальние миграции последних и инвазии (вселение) в новые места. Много в тайге и потребителей ягод и грибов. Это медведь, олени, белки, куриные птицы.
Летом в тайге выплаживаются бесчисленные количества кровососущих насекомых — мошек и комаров. Ими питаются насекомоядные птицы. Однако обилие этих насекомых сильно затрудняет жизнь крупных млекопитающих, не говоря уже о человеке.
К северу от тайги лежит переходная полоса лесотундры и кустарниковой тундры. Побережье Северного Ледовитого океана и его острова занимает типичная тундра, простирающаяся через весь север Евразии и Северной Америки.
Полярные зоны, ограниченные на севере и на юге Полярным кругом, характеризуются астрономически непрерывным летним днем и такой же непрерывной зимней ночью. Это самые холодные области земного шара.
Растительный покров тундры включает ряд многолетних видов — полярную иву и березу, имеющих вид мелких кустарников, бруснику, веронику, дриаду. Но более всего развиты мхи и лишайники. Очень типичны подушки растительности. Из-за вечной мерзлоты корневая система растений поверхностная. Вообще, жизнь здесь сосредоточена в тонком слое, залегающем между почвой и атмосферой. Это хорошо прослеживается при анализе населения моховых дернин или «корочки» прогретого голого грунта, где обнаруживается масса беспозвоночных, особенно низших насекомых, личинок комаров-долгоножек, панцирных клещей, нематод и т. д. В водоемах летом выплаживается громадное количество кровососов (гнус). Беспозвоночные составляют 90 % зоомассы тундр.
Летом тундра оживает главным образом за счет появления большого количества птиц, особенно водоплавающих — гусей, уток, лебедей, многочисленных куликов. Много и хищников, таких как белые совы, кречеты, мохноногий канюк. Пищей соколам и кречетам служат птицы, канюкам и совам — лемминги и полевки. Лемминги наиболее многочисленны в фауне млекопитающих, особенно в годы массового размножения. Летом они находят вдоволь пищи, зимой же скрываются под толщей снега, где прокладывают ходы. На них охотится песец. Из крупных зверей в тундре живут северные олени, а в Северной Америке — овцебыки. Рептилии и амфибии практически не играют никакой роли в жизни тундры, так как за Полярным кругом изредка встречаются лишь живородящая ящерица, сибирский четырехпалый тритон и 2 вида жаб.
Зимой жизнь в тундре надолго замирает. Остаются зимовать только песец, белый медведь, овцебык, заяц-беляк, волк, горностай и лемминги. Даже белая сова и большинство оленей откочевывают к югу.
Благодаря бедности видового состава и простоте структуры экосистемы тундры сравнительно легко установить связи и взаимодействия между отдельными ее компонентами. В таком простом сообществе сильно возрастает роль каждого его звена. Массовое размножение леммингов вызывает увеличение численности их потребителей — песцов и белых сов, что, в свою очередь, влечет за собой уменьшение количества грызунов, за которым следует и снижение численности хищников. В этих процессах наблюдается определенная периодичность.
Освоение Крайнего Севера ставит перед человеком серьезные проблемы охраны природы, поскольку некоторые виды животных становятся все более редкими (белый медведь, краснозобая казарка), а экосистемы в целом оказываются легкоранимыми. Даже передвижение по тундре вездеходов и тракторов приводит к уничтожению растительности, протаиванию грунтов и эрозии почвы.
Высокогорья также отличаются специфическими условиями среды. Здесь отмечаются недостаток кислорода, низкая температура с резкими колебаниями даже в течение суток, интенсивная солнечная радиация с обилием ультрафиолетовых лучей, сильные ветры. Такая обстановка складывается в верхних поясах гор, выше лесной зоны. В зависимости от географического положения горного хребта и от местных условий границы высокогорья находятся на разном уровне, закономерно понижаясь от экватора к полюсам. Под экватором верхняя граница леса проходит на высоте 3800 м, в Гималаях — 3600, в Альпах — около 2000, а на Полярном Урале — на уровне 300 м. Имеет значение и экспозиция склона: на северных склонах Кавказа верхняя граница леса находится примерно на высоте 1800 м, на южных — 2500 м.
Из многих характерных черт высокогорья следует выделить температурный режим. При подъеме с равнины отмечается постепенное убывание температуры (на 1 °С через каждые 140—195 м в зависимости от широты местности). Поэтому в альпийской зоне лето короткое, а зима длинная и суровая.
Высокогорья покрыты низкорослыми коврами многолетних трав (альпийские луга) или подушковидными колючими кустарниками и полукустарниками (нагорные ксерофиты) либо заняты нагорными степями и пустынями. Растительный покров несомкнутый: отдельные площадки, покрытые зеленью, перемежаются со скалами и каменистыми осыпями. Для альпийского разнотравья характерны ярко цветущие растения на коротких стеблях. Это горечавка, примула, крокус, эдельвейс. Многочисленны особые виды злаков, осок, лапчаток. В субальпийской зоне отмечаются стланиковые и криволесные формации из рододендронов, можжевельников, а на юге Азии даже бамбуков.
Животный мир высокогорий также своеобразен, хотя и не богат видами. Жизнь в верхних поясах гор ограничена жесткими рамками. Резкие колебания температур обусловливают обитание здесь только эвритермных форм. Млекопитающие покрыты длинной и густой шерстью, а птицы — плотным оперением. Высокогорные животные крупные (проявление правила Бергманна[5]
), размножаются в сжатые сроки. Адаптация к недостатку кислорода выражается у них в увеличении числа эритроцитов в крови и размеров сердца. Пойкилотермные животные часто обнаруживают склонность к меланизму: рептилии, бабочки и жуки, живущие в горах, темнее, чем на равнине. Многие животные высокогорий ведут только дневной образ жизни. Темная окраска покровов, возможно, с одной стороны, полезна как экран, предохраняющий от ультрафиолетового излучения, а с другой — как поглотитель солнечной энергии. Сильными ветрами объясняется бескрылость многих насекомых, встречающихся здесь. Копытные звери — горные козлы, бараны — имеют узкое твердое копыто «стаканчиком» и великолепно прыгают. В высокогорьях преобладают потребители зеленой и подземной массы растений и сапрофаги. Многие, однако, всеядны. Насекомоядные птицы появляются тут только летом. Несмотря на приспособленность к суровым условиям высокогорий, птицы и крупные млекопитающие вынуждены зимой совершать вертикальные миграции в расположенные ниже пояса в поисках пищи.
Следует отметить, что, поскольку горные хребты не представляют собой единой зоны и разобщены равнинами и межгорными впадинами, распространение животных здесь носит островной характер. Этим, в частности, объясняются обилие в горных странах своеобразных эндемичных животных и низкая их численность. Вот почему высокогорные биоценозы особенно чувствительны к ант-ропическим воздействиям.
Ареал — часть территории или акватории земного шара, на которой постоянно встречаются популяции определенного вида или другого таксона животных.
В зоогеографии главным объектом изучения является ареал вида, поскольку вид — основная и исходная таксономическая категория живых организмов. Наряду с этим полноправно изучение как надвидовых (родовых, семейственных и т. д.), так и подвидовых ареалов. В отдельных случаях, особенно при исследовании процесса географического видообразования, именно подвидовые ареалы дают ценный научный материал. Аналогично иерархии категорий в систематике (вид, род, семейство, отряд, класс, тип) существует и иерархия ареалов: ареал вида слагается из подвидовых ареалов, рода — из видовых, семейства — из родовых и т. д.
Однако рассматривать ареал лишь как площадь не совсем правильно. Точнее представлять его как трехмерную структуру, поскольку водные биотопы имеют не только горизонтальное, но и вертикальное протяжение, да и наземные животные, распространенные и в горах, и на равнине (арктоальпийские, бореомонтанные), располагаются не только на площади, но и в некоем объеме.
Ареалы формировались в результате взаимодействия процессов эволюции животных и изменений среды их обитания в различные геологические эпохи. Поэтому изучение ареалов играет важную роль при выяснении их происхождения и палеогеографи-ческой эволюции. Кроме того, при сравнении ареалов одних и тех же видов через небольшие отрезки времени (10—20 лет) выявляется современная тенденция к расширению или сокращению площади, занятой популяциями вида. А изменения численности и занимаемой площади — четкие показатели состояния вида. Следовательно, по ареалу нетрудно судить о том, что происходит с интересующим нас животным в условиях антропического воздействия на природную среду.
Таким образом, карты распространения определенных таксо-нов остаются важнейшими орудиями труда зоогеографов.
Правильное представление об ареале (а также сравнение ареалов нескольких видов) становится возможным только после его картографического изображения. Существуют различные методы картирования ареалов.
Одним из наиболее распространенных и в то же время конкретных является точечный метод: местонахождение каждого обнаруженного вида обозначается точкой на контурной карте (рис. 113).

Рис. 113. Ареал дубового коконопряда в Великобритании, нанесенный
методом точечного картирования (по Д.Хису, 1973; с изменениями)
С помощью данного метода в отличие от других можно точнее определить пункт сбора, поимки особей того или иного вида либо его регистрации. Он оказывается единственно приемлемым методом, когда картируется распространение редкого или малоизученного вида. Поэтому точечный метод хорош также в начале исследования географического распространения животного. При всех своих преимуществах этот метод имеет и недостатки. Он малопоказателен, так как требует известной доли воображения, чтобы представить себе форму и площадь ареала. Кроме того, животные, как известно, встречаются не в точке, а на более или менее широком пространстве.
Эти недостатки можно компенсировать с помощью контурного метода, дающего возможность картировать границы ареала (рис. 114). С этой целью на контурной карте крайние, т. е. расположенные на границах ареала, точки соединяют сплошной линией, придавая изображению замкнутую форму.
Рис. 114. Контурное изображение ареала саксаульной сойки
(из И.К.Лопатина, 1980)
В последнее время во многих странах применяется сеточный метод, который называют еще растровым, или методом формальных квадратов (рис. 115). При этом поле контурной карты разбивают на квадраты со стороной 50 км (для специальных целей масштаб может быть другим). Квадратом может служить и поле градусной сетки со стороной 1/4, 1/16 и т. д. градуса. Если в пределах квадрата обнаружены особи изучаемого вида, он затушевывается или обозначается условным знаком. Прочие квадраты остаются без обозначений.
На растровой карте разными условными знаками могут быть выделены местонахождения, известные по старым литературным данным и по новым находкам, что позволяет судить, меняются ли площадь и форма ареала со временем.
Рис. 115. Ареал грызуна Malacothristypica в Южной Америке,
нанесенный методом растрового картирования (из М.Удвари, 1978)
Каждый вид имеет свой собственный ареал, и в природе практически невозможно найти два абсолютно одинаковых по площади, расположению или по форме ареала, за исключением мелких островов, где различные виды животных могут населить всю его территорию, и тогда их ареалы будут изображаться на карте одинаково. Тем не менее все разнообразие ареалов можно свести к какому-то ограниченному числу типов.
Для номенклатуры ареалов важно использовать 3 оси координат, или 3 составляющие ареала: широтную (распространение с севера на юг), долготную (с запада на восток) и высотную (вертикальную, сверху вниз).
Широтная составляющая ареала (зональное распространение) является его основным географическим признаком. Она определяется в первую очередь климатическими, особенно температурными, факторами, прямо или косвенно влияющими на распространение большинства видов. Границы ареалов таких видов на определенном протяжении совпадают с границами ландшафтной зоны, хотя зональная приуроченность каждого из них может значительно меняться на всем протяжении ареала.
Высотная составляющая ареала (вертикальная поясность) в определенной степени аналогична широтной, поскольку также определяется главным образом температурными градиентами. По характеру вертикального распространения можно различить альпийские и монтанные (горные) виды. Последние заселяют среднегорья. Выделяют и промежуточные между ними — субальпийские формы. Часто приходится сталкиваться с равнинно-горным расселением. Примером может служить арктоальпийское или бореомонтанное распространение видов (рис. 116).
Рис. 116. Основные типы равнинно-горного
распространения животных (по К.Б.Городкову, 1984)
Долготная составляющая ареала определяется степенью удаленности от океана, обусловливающей характер климата. Границы ареалов могут быть связаны с континентальным или морским, аридным или гумидным климатом. В пределах гумидной части Евразии, к примеру, наиболее существенны следующие границы:
между Западной и Восточной Европой, по Уралу, по западному краю Среднесибирского плоскогорья («линия Иогансена»), в районе Байкала и по водоразделу бассейна Охотского моря.
Комбинация составляющих ареала позволяет охарактеризовать тип распространения и создать описательные классификации ареалов.
В основу классификаций должно быть положено одно из основных свойств ареалов — их размеры. Можно построить иерархическую систему ареалов — от узколокальных до глобальных (рис. 117). Наряду с очень узкими, занимающими территории в несколько квадратных километров и меньше, встречаются широкие ареалы, которые охватывают целые регионы, материки и даже весь земной шар. Как правило, наземные животные чаще имеют узкие ареалы, морские же — широкие. Иногда вид ограничен в своем распространении локальным районом, естественной географической (или фаунистической) областью. Такие виды называются эндемичными для данной области или просто эндемиками.
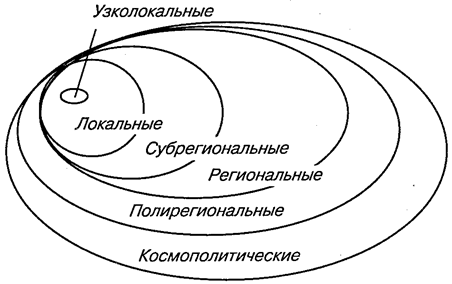
Рис. 117. Схема пространственного совмещения ареалов
различных размерных групп (по К. Б. Городкову, 1983)
Особенно узкие ареалы типичны для видов, распространение которых ограничено какими-нибудь непреодолимыми преградами. Это прежде всего островные либо пещерные формы, обитатели горных долин или, наоборот, верхних зон горных хребтов. Нередки весьма узкие ареалы у нелетающих видов насекомых. Например, жужелицы-брызгуны обитают на Кавказе в пределах одного-двух хребтов: Carabuspolychrous населяют Бзыбский хребет в Абхазии, С. komarovi — Сванетию, С. cordicollis встречаются только на Эльбрусе. Аналогичные ареалы у бескрылых жуков-листоедов рода Oreomela в горах Тянь-Шаня и Памиро-Алтая. Так, О. medvedevi живет только в одном районе Шугнанского хребта на Памире, О. transalaica — на Заалайском хребте, О. bergi — в альпийской зоне Туркестанского хребта в верховьях реки Исфары. Все без исключения виды этого рода, а их больше 50, имеют очень узкие эндемичные ареалы. Исключительно узкие ареалы у трогло-бионтов, постоянно живущих в пещерах, причем это касается как наземных, так и пресноводных видов. Итак, все узкие ареалы свойственны эндемикам.
Локальные ареалы, постепенно расширяясь, переходят в субрегиональные и региональные. Если вид распространен на всей площади региона, то его ареал называют трансрегиональным (рис. 118).
Еще более широкие ареалы, так называемые поли- или мультирегиональные, включают в себя несколько (не менее 2) фаунистических областей, часто располагающихся на разных материках (рис. 119, 120). Наконец, различают космополитные, или всесветные, ареалы (рис. 121). Они характерны чаще всего для морских животных, таких как кашалот (Physeter), дельфин (Grampusgri-seus), касатка (Orcinusorca). Из наземных животных космополитами стали определенные насекомые-синантропы. С некоторой натяжкой к космополитам можно отнести сокола-сапсана.
Рис. 118. Трансрегиональный ареал бабочки-боярышницы
Рис. 119. Полирегиональный ареал пластинчатоусых жуков
рода Trichius (из С.И.Медведева, 1960)
Рис. 120. Полирегиональный ареал жуков-навозников рода Onitis
(из В.Балтазари, 1963)
Рис. 121. Космополитный ареал жуков-навозников
рода Onthophagus(из В.Балтазари, 1963)
Между систематическим рангом группы и размером ареала существует корреляция: чем выше ранг группы, тем большую площадь она занимает. Следовательно, ареал рода больше ареала вида, ареал семейства больше ареала рода и т. д. Классы, как правило, распространены всесветно. Главными причинами, обусловливающими специфичность ареалов, являются экологическая валентность видов, их способность к расселению (вагильность), филогенетический возраст и скорость видообразования. Совокупность этих признаков и определяет величину ареала вида или их группы.
Формы ареалов, т. е. их очертания, столь же разнообразны, как и величина. Факторы, определяющие форму и величину ареала, чаще всего одни и те же. Очертания многих ареалов совпадают с ландшафтными или вообще с физико-географическими рубежами. В умеренных областях Северного полушария, а также в высоких широтах ареалы ряда видов вытянуты с запада на восток и значительно меньше — с севера на юг, т. е. носят зональный характер. Это объясняется особенностями распределения климатических условий, которые быстрее изменяются при продвижении с севера на юг, чем с запада на восток. В местах, далеко отстоящих друг от друга, но расположенных на одной широте, условия среды часто сходны.
Как показал Ю.И.Чернов (1975), наиболее точное совпадение ареала с конфигурацией зоны наблюдается в тех случаях, когда вид связан с зональным типом растительности. Зональные ареалы у «степняков» — джунгарского хомячка, кузнечика эверс-манна, степного таракана и др. Ареал лапландского подорожника Calcariuslapponicus совпадает с зоной тундры, мохноногого сыча — с таежной зоной; с широколиственными лесами связаны ареалы сонь — орешниковой, полчка и в некоторой степени лесной и садовой. Если вид и расселяется за пределы ландшафтной зоны, то он встречается лишь в экстразональных биотопах, не типичных для нового ландшафта.
В горных странах ареалы многих видов вытянуты вдоль хребтов и тоже укладываются в одну из вертикальных зон — лесную, субальпийскую и т. д. Объясняется это тем, что вдоль определенного склона хребта наблюдаются одинаковые условия, чаще климатические.
Вытягивание ареала, обусловленное экологическими факторами, может привести к образованию так называемых ленточных, или линейных, типов ареалов. Они свойственны многим околоводным или водным животным, особенно обитателям литорали. Аналогичную форму имеют ареалы пресноводных рыб. Хорошим примером такого ареала служит распространение среднеазиатских осетровых рыб из рода Pseudoscaphirhynchus, обитающих в Аму-дарье и Сырдарье.
Нередко встречаются и сопряженные ареалы. Их очертания определяются границами ареала другого вида. Это бывает в том случае, когда между видами существуют экологические взаимоотношения, которые обычно носят характер пищевой связи. Как правило, она устанавливается между паразитами и хозяевами и сожителями, хищниками и жертвами, животными-фитофагами и их кормовыми растениями: тигр связан с кабаном, куница — с белкой, соболь — с бурундуком (хищник — жертва), бабочка олеандровый бражник — с олеандром, жуки-нарывники Mylabriselegantissima — с песчаной акацией, африканский пальмовый орлан — с масличной пальмой (потребитель — кормовое растение). Монофаги встречаются только там, где есть кормовое растение. Однако ареалы последних больше, поскольку распространение животных зависит не только от наличия корма, но и от других факторов. Значительно реже наблюдается сопряжение ареалов паразита и хозяина. Паразиту часто требуется промежуточный хозяин, а то и два, и отсутствие одного из них, даже при наличии основного хозяина, препятствует распространению этого паразита.
Все рассмотренные выше ареалы, от узколокальных эндемичных до громадных, занимающих весь материк, относятся к сплошным ареалам.
Вместе с тем постоянно встречаются ареалы, разделенные на части, т. е. прерывистые, или дизъюнктивные. О них следует говорить в том случае, когда отдельные части ареала обособлены друг от друга в такой степени, что никакая связь между заселяющими их популяциями вида невозможна.
Ареал может прерываться по-разному. Бывает так, что в основном он сплошной, но встречаются обособленные маленькие участки его, которые принято называть островными местонахождениями. Ареалы подобного типа считаются расчлененными (фрагментированными). Они возникают и в силу естественных причин, но часто своим происхождением обязаны деятельности человека. Например, вырубка лесов приводит к исчезновению истинно лесных видов, сохраняющихся лишь в отдельных массивах.
Части дизъюнктивного ареала могут располагаться на одном материке, типы их разнообразны, но общим является то, что в этом случае между частями ареала нет преград в виде морских проливов или участков океана. На разделяющей территории природные условия тоже часто не отличаются от таковых в местах обитания видов, поэтому дать удовлетворительное объяснение причинам подобных разрывов довольно трудно. Типологические материковые дизъюнктивные ареалы могут различаться в зависимости от места расположения их частей. Выделяют, к примеру, евро-азиатские дизъюнктивные ареалы, когда одна их часть находится в Европе, другая — в Азии.
Как правило, части дизъюнктивного ареала связаны с одной ландшафтной зоной. Широко известными примерами такого рода являются амфиевро-азиатские неморальные ареалы амфибий — жерлянок рода Bombina, рыб — горчака и вьюна, птиц — голубой сороки и др. В широколиственных лесах Европы встречаются В. bombina и В. variegata, в Приморье их замещают В. orientalis, а в Китае — В. maxima (пример викариата на уровне видов). Горчак и голубая сорока в различных частях ареала представлены разными подвидами: в Европе живет горчак обыкновенный (Rhodeussericeussericeus), в бассейне Амура — горчак амурский (R. s. amurensis), в Испании — голубая сорока Cyanopicacyanacooki, а в Приморье — С. с. cyanus (рис. 122).
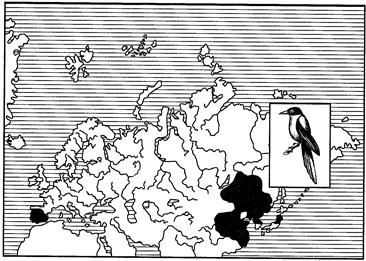
Рис. 122. Амфиевро-азиатский неморальный ареал голубой сороки
Различают также арктоальпийский и бореомонтанный типы дизъюнктивного ареала. В первом случае основная часть ареала (зональная) находится в тундре, другая — в альпийском поясе гор Европы (рис. 123).
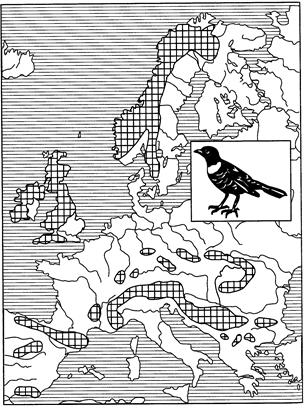
Рис. 123. Арктоальпийский ареал белозобого дрозда (из Г.Делаттина, 1967)
Особый и нередко встречающийся тип дизъюнкций представляют горные разрывы ареалов, когда отдельные их части находятся в верхних поясах горных хребтов и отделены друг от друга более или менее широкими долинами и межгорными понижениями. К примеру, серна — высокогорный копытный зверь — в каждом горном массиве имеет изолированный ареал. Популяции ее в Пиренеях, Альпах, на Балканах, горах Тавр в Малой Азии и на Кавказе не связаны друг с другом (рис. 124).
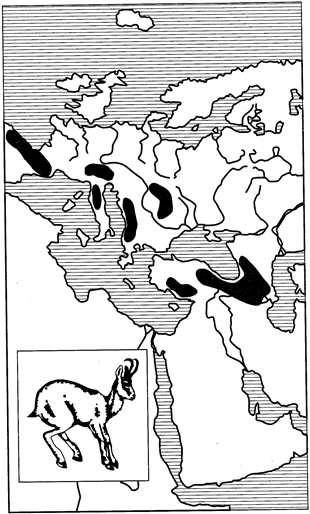
Рис. 124. Горный разрыв ареала серны (из И.К.Лопатина, 1980) 336
Аналогичный горный разрыв характерен для ареалов видов одного рода, например горных козлов Сарга. Последние заселяют горы Южной Европы, Малой и Средней Азии, Гималаев, Южной Сибири. Подобные ареалы характерны для высокогорных насекомых, например для представителей уже упоминавшегося рода жуков-листоедов Oreomela, объединяющего более 50 видов, причем только на Тянь-Шане их 28, в Джунгарии и Синьцзяне — 11, в Тибете и Цинхае (КНР) — 9. Каждому хребту свойственны свои виды, а межгорные долины и ущелья являются непреодолимыми преградами для их расселения, поскольку жуки не опускаются ниже пояса криволесья и чаще населяют только альпийский пояс хребтов.
Причины материковых разрывов ареалов разнообразны. Чаще разъединение объясняется фрагментацией сплошного ареала, вызванной изменением климата, конкуренцией более приспособленных форм, геоморфологическими перестройками или, наконец, деятельностью человека (вырубка лесов, мелиорация болот, распашка целины и т.п.). В любом из этих случаев можно допустить либо вымирание отдельных популяций, либо вытеснение их другими, более приспособленными к новым условиям видами, что иногда подтверждается палеонтологическими находками в области разрыва.
В Северном полушарии ряд дизъюнкций ареалов обусловлен событиями плейстоцена, в связи с чем они получили название ледниковых. Допускается, что некогда сплошные зональные ареалы Северного полушария были разорваны наступившими льдами. Там же, где покровного обледенения не было, климат настолько ухудшился, что некоторые популяции вымерли, а сохранившиеся в наиболее благоприятных районах не смогли расселиться и восстановить свой прежний ареал. Эти климатические изменения не коснулись теплолюбивых неморальных видов, уцелевших в убежищах, находившихся в юго-западной Европе и на юге Дальнего Востока (они там живут и в современную эпоху). Климат же Сибири изменился настолько, что расселение через нее стало невозможным. События ледникового периода вызвали также разрывы ареалов арктоальпийского и бореомонтанного типов. Во время четвертичного покровного оледенения климат Северной Европы был холодным, и характерная для него арктическая (тундровая), а также бореальная (таежная) фауна широко распространилась к югу. Достаточно сказать, что, по данным палеонтологии, заяц-беляк, песец и рысь жили в то время даже в Крыму. В альпийской зоне южных гор, в свою очередь, развивалось местное оледенение и ледниковые языки опускались далеко в долины. После таяния и отступания материковых льдов холодолюбивые виды, обитавшие на окраинах ледника, мигрировали на север и заняли здесь освободившиеся ото льда пространства. Популяции некоторых видов при сокращении горного оледенения отступали вместе с ледниками в самые верхние пояса хребтов, где климат оставался холодным. Между горами Центральной и Южной Европы, с одной стороны, и тайгой или тундрой — с другой, образовались разрывы ареалов. Северная часть нынешнего ареала рассматривается как новый (по времени) ареал, возникший в результате активного расселения животных, а южная (горная) — как реликтовая. Таким образом, заяц-беляк и тундряная куропатка в Альпах являются ледниковыми реликтами.
Горные разрывы ареалов возникли в результате сложных геоморфологических процессов, происходивших в определенных частях земного шара.
Особый тип дизъюнктивных ареалов составляют ареалы, у которых между их частями находятся морские пространства. Подобные океанические дизъюнкции ареалов характерны для надвидо-вых таксонов — родов, семейств, отрядов и т. д. В пределах Северного полушария к данному типу ареалов относятся циркумполизональ-ные, циркумполярные, циркумбореальные и циркумтемператные (расположены по всей умеренной зоне), а также амфиатланти-ческие ареалы (рис. 125, 126). Части последних располагаются на материках Евразии и Северной Америки в соответствующих зонах, а циркумполизональные и панголарктические охватывают все зоны материков.
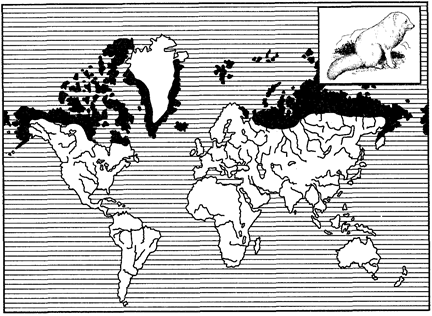
Рис. 125. Циркумполярный ареал песца (из В. Г. Гептнера, 1936)
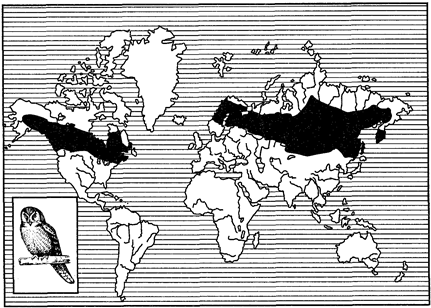
Рис. 126. Циркумбореальный ареал ястребиной совы
(из В. Г. Гептнера, 1936)
Очень обычны формы европейско-североамериканского и ази-атско-американского разрывов. В этих случаях между ареалами родственных наземных и пресноводных таксонов находятся водные преграды — Атлантический океан, Берингов пролив или даже северная часть Тихого океана.
Примерами океанических разрывов ареалов может служить распространение тапиров (рис. 127) в тропических частях Южной Америки и юго-восточной Азии, сумчатых млекопитающих (рис. 128), жаб-свистунов (рис. 129) в Австралии и Южной Америке и др.
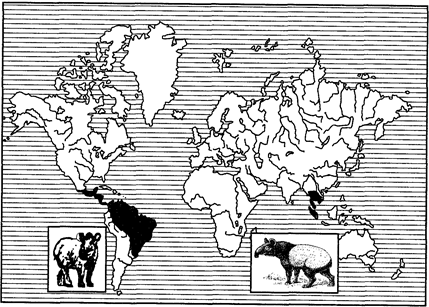
Рис. 127. Дизъюнктивный ареал тапиров (из И.К.Лопатина, 1980)
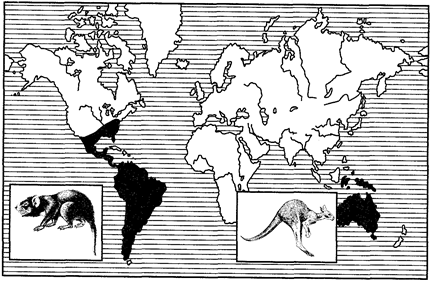
Рис. 128. Дизъюнктивный ареал сумчатых млекопитающих
Рис. 129. Дизъюнктивный ареал жаб-свистунов (из И.К.Лопатина, 1980)
Среди океанических разрывов замечательны случаи биополярного и амфибореального распространения (рис. 130): соответствующие группы животных встречаются в приполярных или умеренно холодных частях Земли и отсутствуют в тропиках и субтропиках.
Рис. 130. Биополярное распространение червей-приапулид
(обозначено точками) и амфибореальное распространение
сардин (обозначено штрихом)
Причины многих океанических разрывов ареалов не установлены, особенно когда нет палеонтологических находок на территориях, где те или иные группы животных не встречаются. Предложено несколько теорий, объясняющих происхождение подобных ареалов (теории мостов суши, оттесненных реликтов и др.).
Ареалы близких видов располагаются рядом, а иногда могут накладываться друг на друга (перекрывание ареалов). Так, ареалы зайца-беляка и зайца-русака в Северной Европе на большом протяжении расположены на одной и той же территории. Бывает, что ареал одного вида, как правило, небольшой величины, охвачен гораздо большим ареалом другого вида. Из-за различных требований, предъявляемых видами к окружающей среде, перекрывание их ареалов — явление нередкое. Оно получило название симпатрии.
Иная картина наблюдается при изучении ареалов подвидов. В данном случае ареалы очень часто прилегают друг к другу, но никогда не перекрываются. Каждый подвид населяет определенную область видового ареала (подвиды аллопатричны), и подвиде -вые ареалы замещают друг друга, или викарируют. Примером географического викариата служит распространение подвидов колхидского фазана на территории от Кавказа до Тихого океана. Викарируют не только подвиды, но и близкие виды. Общеизвестно, что европейскому зубру (Bisonbonasus) соответствует в Северной Америке викарный вид бизон (Bisonbison). В рассмотренных случаях замещающие формы не только близки по происхождению, но и занимают одинаковые биотопы. Такой тип викариата назван систематическим. Если же одинаковые биотопы в разных странах населены различными по систематическому положению и происхождению видами, конвергентно сходными между собой из-за одинакового образа жизни, то пользуются термином «экологический викариат». Примером экологического викариата является замещение европейского крота в Африке златокротом (другое семейство), а в Австралии — сумчатым кротом (другой подкласс).
Величина ареалов и их границы находятся в тесной взаимосвязи. Если размеры ареалов очень часто зависят от экологической валентности населяющих их видов, то значение экологических факторов, определяющих границы ареалов, еще более велико. Эти границы нередко обусловлены определенной комбинацией факторов среды или даже их одним градиентом. Особое значение имеют температура, освещение, питание, характер субстрата. Для наземных животных существенную роль, кроме того, играет влажность воздуха, для водных — химизм воды, содержание кислорода и т. д.
Существует разделение границ на климатические, ландшафтные, биоценотические. Их можно назвать экологическими границами, поскольку они определяются целой совокупностью факторов. С другой стороны, известно немало случаев, когда граница остается стабильной, и расселения не происходит, несмотря на то что условия среды внутри ареала и за его пределами однородны и вполне благоприятны для вида. Подобные явления объясняются историческими причинами и указывают на то, что в прошлом условия среды по обе стороны границ были различными. Границы ареалов, происхождение которых обусловлено минувшими событиями и не оправдывается экологической обстановкой настоящего времени, называются историческими или реликтовыми.
Биоценотические границы определяются взаимоотношениями разных видов в сообществе. Эти взаимоотношения носят разный характер (симбиоз, комменсализм, конкуренция и т. п.). Биологическое соперничество — одна из причин стабильности границ ареалов двух конкурирующих видов. Так, например, обстоит дело с ареалами двух близких видов — лесной куницы и соболя на Урале и в Западной Сибири.
Однако стабильность границ — явление относительное и временное, тем более в настоящий период существования животного мира, когда в процесс расширения и сокращения ареалов вмешивается человек.
Согласно дарвиновской теории эволюции, каждый вид возникает в определенной части земной поверхности и его первоначальный ареал в зависимости от характера формообразовательного процесса может быть большим или меньшим. Но со временем он расширяется, что происходит в результате расселения организмов. Под расселением, по определению А. И.Толмачаева (1974), следует понимать динамический процесс распространения видов за пределы занимаемой ими территории.
Изменения ареалов прослеживаются по изменениям очертаний их границ. Как правило, эти процессы протекают постепенно и ускользают от прямого наблюдения. Лишь редко очертания границ меняются в течение одного или двух столетий, поэтому изменения устанавливаются с достоверностью.
Предпосылками расселения животных являются колебания численности особей в популяциях. Эти колебания происходят вследствие изменений соотношения между рождаемостью и смертностью особей. При увеличении плотности популяции у ее членов появляется тенденция к расселению, при уменьшении — к потере части занимаемых участков.
Темпы расширения ареалов различны у разных видов, и это связано как с особенностями среды обитания, так и с характером самих видов. Способность видов к расселению зависит от их структурных, физиологических, поведенческих, популяционных и других свойств и носит название вагильности. Вагильность вида осуществляется в определенных условиях среды, влияющих на плодовитость, дальность разноса зачатков (икра, личинки) и на другие свойства организмов, обусловливающие эту способность. Расселение животных — многофазный процесс, предусматривающий преодоление различных преград и препятствий. Различают вагильность активную (организмы расселяются благодаря самостоятельному передвижению — полет, плавание, бег), пассивную (расстояние преодолевается за счет переноса ветром, течениями, человеком и т. д.) и смешанную (сочетание активной и пассивной вагильности).
Преграды к расселению можно условно свести к физическим и экологическим. К первым для сухопутных животных относятся прежде всего водные пространства, для водных организмов — большие участки суши, разделяющие водные бассейны. Мировой океан — среда сравнительно однородная, поэтому ареалы его обитателей, как правило, очень большие по площади. Это касается не только таких отличных пловцов, как китообразные, но и сидячих форм из мира беспозвоночных. Наиболее серьезной преградой для расселения морских животных оказывается суша, в том случае, когда обойти ее мешают не соответствующие требованиям вида соленость, давление, температура воды и т. п.
В отличие от Мирового океана пресноводные бассейны не представляют единого целого, что сильно затрудняет расселение животных этого биоцикла. Основными преградами здесь следует считать море (соленую воду) и водоразделы (участки суши между водоемами). К числу местных препятствий относятся также водопады и пороги на реках. Изоляция отдельных бассейнов, например озер, не соединенных речными системами, чрезвычайно велика. Это приводит к появлению узких ареалов очень многих видов, что нетипично для обитателей других биоциклов.
Преградами, препятствующими распространению сухопутных животных, в первую очередь являются границы других биоциклов — морей и пресных вод. Существование громадного количества узких ареалов на суше свидетельствует о том, что число преград к расселению здесь больше, чем где бы то ни было. Морские проливы могут представлять собой непреодолимый барьер для нелетающих видов наземных животных. Так, Мозамбикский пролив, ширина которого 400 км, разделяет Африку и Мадагаскар, населенных резко различающейся фауной.
Морская вода неблагоприятна для тех пресноводных животных и наземных обитателей, которые не имеют непроницаемых защитных покровов. Поэтому активное расселение их через морскую воду практически невозможно. Этим объясняется отсутствие на островах большинства амфибий, типичных для материка. Рептилии благодаря особенностям строения кожи могут переплывать сравнительно узкие проливы. Таким образом, островные фауны всегда более бедны, а видовой состав их несбалансирован. Фауна островов тем беднее, чем шире водная преграда, отделяющая их от ближайшего материка.
Как указывает Ф.Дарлингтон (1966), главную роль при расселении животных играет расстояние.
В ряде случаев роль серьезной преграды к расселению животных выполняют реки. Крупные реки нередко разделяют соседние территории, населенные различной фауной. Амазонка, например, служит границей распространения 50 видов птиц, сотен видов насекомых и других животных. Волга в низовьях, с одной стороны, представляет собой западную границу распространения многих степных и пустынных видов млекопитающих (желтого суслика, хомячка эверсманна, пегой землеройки), а с другой — восточную границу ареала слепыша и крапчатого суслика. Такой же фаунистической границей служит Днепр в нижнем течении. На правобережье его встречаются крот, крапчатый суслик, зеленая ящерица, из насекомых — светлячок, в левобережных степях эти животные отсутствуют.
Существенную преграду для расселения наземных животных представляют горы. Однако рассматривать их как чисто топографические препятствия нельзя. Дело в том, что склоны гор разной экспозиции часто покрыты различной растительностью и отличаются в климатическом отношении. Таким образом, горные хребты во многих случаях служат экологическими барьерами. Общеизвестна роль Гималаев, разделяющих две фаунисти-ческие области — Палеарктическую и Индо-Малайскую.
О роли горных хребтов как преград к расселению говорит и наличие огромного количества узких ареалов видов одного рода в странах с сильно расчлененным рельефом. Так, в горах Средней Азии встречается около 100 видов жуков-жужелиц рода Carabus, aна равнинах европейской части России — не более 30.
Не только высокогорный пояс служит преградой для расселения равнинных животных. Настоящие «альпийцы» не могут расселяться через долины с не подходящими для них условиями среды. Это также свидетельствует об экологической природе большинства физических барьеров.
Различные типы пустынь — холодные или жаркие, песчаные, глинистые, щебнистые — для большинства не адаптированных к ним животных представляют одну из существенных преград к расселению. Особенно важна в этом отношении протяженность пустынь. Пустыня Сахара, к примеру, отделяет эфиопскую фауну большей части материка от палеарктической фауны севера. Индо-малайская фауна юга Азии не распространяется к северу, так как этому препятствуют пустыни Передней Азии и Гималаи.
Наконец, следует сказать о преградах антропического происхождения. С точки зрения экологии расселения окультуренные земли сравнимы с пустынями. Для ряда видов расселение через них исключено. Судьба многих вымирающих под воздействием человека видов животных — доказательство этому. Меньше всего приспособлены к расселению через «культурную пустыню» лесные и степные виды.
Так называемые биологические преграды обусловлены экологическими связями между видами животных и животных и растений. Отсутствие необходимого кормового объекта для монофага, хозяина для паразита, подходящей жертвы для хищника создает препятствия для расселения или вообще делает его невозможным. Расселение ограничивается и конкуренцией видов, занимающих одну и ту же экологическую нишу. На этом, в частности, основано явление викариата (замещения), характерное для подвидов одного вида, совершенно сходных по своим требованиям. Граница между ареалами западного и восточного соловья, соболя и куницы, серого и крапчатого суслика служит примером в данном случае. Иногда расселение вида сдерживается хищником. Так, молодь ондатры, расселяющаяся за пределы болот (на луга, поля и другие «чужие» местообитания), регулярно уничтожается норкой. В лесостепных и степных районах юга европейской части России наблюдается рост численности и расселение копытных — кабана, косули, оленей, что в известной мере обусловлено регулированием численности волка.
Пространство и время также выполняют функцию преград. Расселяясь из центра происхождения по территории, где нет преград и условия однообразны, вид тем не менее занимает не все пригодное для жизни пространство. Это означает, что расселение не завершилось и само расстояние служит своеобразной преградой. Примером, иллюстрирующим значение времени и пространства для расселения, являются наблюдаемые в природе случаи продвижения акклиматизированных или случайно занесенных видов. Показателен в этом отношении факт расселения в Западной Европе колорадского картофельного жука (рис. 131). Начиная с 1918 г. и до настоящего времени он продолжает расселяться на восток. Понадобилось свыше 50 лет, чтобы жук заселил территорию протяженностью с запада на восток около 4000 км. Вместе с тем остаются незаселенными пригодные для его обитания большие районы европейской части России, и если бы не вмешательство человека, проникновение в них колорадского жука было бы только вопросом времени.
Рис. 131. Первоначальное расселение колорадского жука в Европе
(из Н.В.Тимофеева-Ресовского, А.В.Яблокова, Н.В.Глотова, 1973)
В ряде случаев отсутствие вида в той или иной местности объясняется существованием там преград в прошлом. И хотя этих преград теперь уже нет, для заселения лакун требуется длительное время. Такими лакунами могли быть районы последнего материкового оледенения. Это доказывает временность преград. Последние могут возникать, а затем исчезать, в результате чего для животных открываются новые, ранее недоступные пути расселения. Вопрос о возникновении и уничтожении преград или, иначе говоря, об изменении условий расселения животных — один из основных вопросов зоогеографии.
Активное расселение животных особенно характерно для обитателей Мирового океана и суши. Среди жителей внутренних водоемов в силу того, что последние изолированы друг от друга, оно наблюдается редко и свойственно в основном летающим насекомым, личинки которых ведут строго водный образ жизни (водяные жуки, стрекозы, ручейники и др.).
Активное расселение основано на различных формах самостоятельного движения (полет, бег, ползание, плавание и т.д.).
Поразительны темпы расселения кольчатой горлицы Strepto-peliadecaocto. До 1925 г. она не регистрировалась севернее Болгарии, однако к 1955 г. расселилась по всей Средней Европе, достигла Франции, Бельгии, Голландии, Дании, а в 1957—1958 гг. отмечалась на гнездовании в Шотландии, Южной Швеции, Южной Норвегии и в Прибалтике (рис. 132). Причины столь необычной экспансии неизвестны.
Рис. 132. Расселение кольчатой горлицы в Европе (из У.Зедлага, 1972)
Нередко сухопутные звери переплывают водные преграды, заселяя острова, расположенные вблизи материка. Хорошо плавают олени, лоси и косули. Плавание — обычный способ передвижения белого медведя. Гиппопотамы достигают острова Занзибар, переплывая морской пролив шириной 30 км. Даже такие мелкие зверьки, как ласка, могут проплывать до 1,5 км. Рекордная дальность плавания была отмечена для крокодилов — 1000 км. Плавают и змеи, а расселение амфибий возможно только в пресной воде, так как соленая для них губительна.
Пассивное расселение животных или их яиц и личинок осуществляется при помощи разнообразных факторов переноса.
Для обитателей Мирового океана наиболее реальным является перенос морскими течениями. Ими могут воспользоваться планктонные формы, почти лишенные приспособлений для активного передвижения, но обладающие хорошо развитыми органами парения (разнообразные приспособления, увеличивающие поверхность тела и плавучесть: выросты, шипы, иголочки, жировые капельки, пузырьки воздуха и т. п.). Держась в толще воды, эти организмы могут преодолевать значительные расстояния. Бентосные, особенно сидячие организмы океана, часто имеют планктонных или нейстонных личинок, так что и для них возможно использование морских течений при расселении.
Учитывая скорость течения и продолжительность существования личиночной стадии, легко рассчитать расстояние, на которое могут разноситься личинки сидячих или малоподвижных бентосных видов. Так, проплывая с помощью Гольфстрима (его скорость не менее 5 км/ч) в среднем 120 км в сутки, личинки кольчатых червей за 20 дней существования покрывают расстояние в 2400 км, личинки иглокожих за 40 дней — 4800 км. Используют морские течения медузы, сифонофоры, многие раки и даже рыбы, т. е. формы, обладающие активными средствами к расселению. При этом литоральные животные, для которых морские глубины являются серьезной преградой, легко преодолевают ее в стадии плавающей, или точнее, дрейфующей личинки. Этим объясняется сходство литоральной фауны отдаленных районов Мирового океана, расположенных на одних широтах. Такие малоподвижные или сидячие животные, как губки, кораллы, усоногие раки, моллюски, могут переноситься на колоссальные расстояния, используя предметы, дрейфующие в океане, чаще всего бревна или стволы деревьев.
Весьма характерна пассивная вагильность для пресноводных животных, особенно беспозвоночных. По всей видимости, вся пресноводная фауна в определенной мере имеет приспособления к пассивному переселению. Иначе невозможно объяснить поразительное сходство видового состава разобщенных водоемов, поскольку водные животные не имеют специальных органов для передвижения на суше. Способы пассивного расселения здесь весьма разнообразны: передвижение при помощи речных течений, транспортировка на плавающих предметах, разнос ветром, перенос мелких организмов на лапах водоплавающих птиц и т. д. Расселение проточными водами носит название гидрохории, а ветром — анемохории.
Расселение при помощи других животных — биохория — бывает случайным и закономерным. Особый ее случай — форезия — прикрепление на длительное время одного организма к другому для смены местообитания. Расселение паразитов основано на активном передвижении их хозяев.
Особый вид пассивного расселения — антропохорное, в котором участвует человек. Развитие средств сообщения между отдаленными странами, часто расположенными на разных материках, перевозка различных грузов и товаров делают неизбежным непроизвольный завоз самых разнообразных животных. Кроме того, интродукция и акклиматизация животных проводятся в плановом порядке.
Человек в роли фактора пассивного расселения различных видов животных выступает уже с давних времен. Еще древние полинезийцы, совершавшие плавания в океане на примитивных челнах, развезли по островам Тихого океана свиней и собак, которые затем одичали и вошли в состав местных фаун.
С каждым столетием завоз животных увеличивался. Однако это способствовало распространению вредителей и возбудителей болезней, и в середине XX в. в ряде стран создаются карантинные службы. Антропическое влияние на ландшафты, а через них и на животный мир, осуществляемое прямо или косвенно, можно оценить как важнейший зоогеографический фактор настоящего времени.
Несмотря на то что многие виды животных обладают разнообразными способностями к расселению, этого все же недостаточно для успешного преодоления преград и последующей колонизации нового района. Переселившимся организмам нужен биотоп или экологическая ниша, сходная с той, которую они занимали прежде. Если такую нишу вид не найдет, результат вселения может быть двояким: вид либо погибнет, либо изменит экологические потребности. Последнее наступает тем легче, чем большим полиморфизмом обладает переселяющаяся популяция.
Выживание переселенцев зависит как от внешних факторов, так и от их собственных свойств. Если параметры новой среды мало чем отличаются от прежних, колонизация может быть успешной. Этим объясняются многочисленные случаи укоренения европейских животных в Новой Зеландии, являющейся климатическим аналогом Великобритании.
Однако проблема колонизации решается не только физическими факторами окружающей среды, чрезвычайно важно и биоценологическое воздействие. Конкуренция видов создает замкнутость биоценозов, и это, по-видимому, основная причина, препятствующая вселению видов в насыщенные материковые биоценозы. Однако островные фауны, за некоторым исключением, бедны видами и имеют много свободных экологических ниш. Поэтому колонизация их переселенцами осуществляется практически беспрепятственно. Основная задача, стоящая перед расселяющимся видом, заключается в том, чтобы «вписаться» в новую экосистему.
В ряде случаев вселение и колонизация нового вида, отличающегося большой конкурентоспособностью и широкими экологическими потенциями, приводят к вытеснению аборигенных видов. Так, в Новой Зеландии европейские виды птиц почти повсеместно вытесняют местных. Акклиматизированный в ряде рек Северной Америки и Австралии карп успешно вытесняет местные породы рыб. Узкопалый рак, пересаженный в некоторые озера Белоруссии и Германии, за короткий срок сменил широкопалого.
Следует добавить, что хозяйственная деятельность человека, связанная обычно с изменением ландшафта, нередко способствует вытеснению аборигенных форм и создает благоприятные условия для новых колонистов.
Расселение животных предполагает наличие первичного ареала — сравнительно небольшого района, где происходит видообразование, в ходе которого новая форма, дивергировавшая от родительской, приобретает видовую самостоятельность. Впоследствии таким же образом возникают другие близкие формы, объединяемые в роды, последние, в свою очередь, составляют таксономические единицы большего ранга — трибы, семейства и т. д. Каждая из этих единиц имеет свой ареал, слагающийся из видовых ареалов. Ареал рода (трибы, семейства) не просто сумма видовых ареалов: ареалы отдельных видов каждого рода не только дополняют друг друга, но и в определенной степени перекрываются (напомним, что видовые ареалы могут быть как аллопатричны-ми, так и симпатричными). В таком случае общие очертания ареала рода определяют территорию, на которой встречается хотя бы один из его представителей. Поэтому при изучении родовых ареалов устанавливают не только их пределы, но и размещение в них отдельных видов. Чаще всего распределение видов в границах родового ареала оказывается неравномерным, и даже есть участки, где встречается всего лишь один вид. Места концентрации большого количества видов получили название очагов видового разнообразия или центров распространения (рис. 133, а, 6).
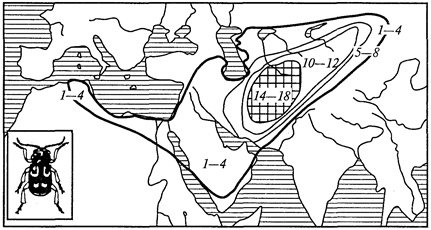
Рис. 133, а.
Ареал рода Thelyterotarsus (жуки-листоеды); цифрами обозначено
число видов в отдельных частях ареала; очаг видового разнообразия заштрихован
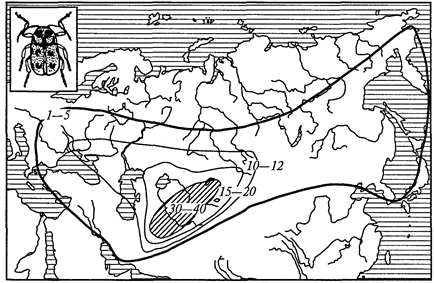
Рис. 133, б.
Ареал подрода Asionus (скрытоглавы азиатские) рода Cryptocephalus
(жуки-листоеды); цифрами обозначено число видов в отдельных частях ареала;
очаг видового разнообразия заштрихован
Выявить центры видового разнообразия несложно. Однако эти центры не могут одновременно считаться и центрами их происхождения, т. е. родиной данных групп. Обнаруживая скопление большинства видов рода, можно лишь предположить, что здесь его родина и что отсюда началось расселение принадлежащих ему видов. Однако, учитывая, что ареалы динамичны и что первичный ареал (т. е. центр возникновения) мог располагаться на территории, где уже не существует ни одного вида рассматриваемого рода, проблема однозначно решается только при наличии соответствующего палеонтологического материала. Но так как ископаемые данные могут быть скудными или могут отсутствовать, то приходится пользоваться косвенными сведениями.
Фауна —
исторически сложившаяся совокупность видов животных, обитающих в данной области и входящих во все ее биогеоценозы.
Фауна объединяет все виды животных той или иной области (района, местности), хотя они и входят в различные биоценозы. Так, фауна европейской части России на равных основаниях включает северного оленя, степного орла, щуку, бабочку-махаона, губку бадягу, в то время как местообитания их совершенно разные. Вместе с тем нельзя причислять к местной фауне виды, сознательно завезенные человеком и содержащиеся в зоопарках, аквариумах и т.д. Виды же, завезенные случайно, а также преднамеренно, но одичавшие или существующие без помощи человека, необходимо учитывать в составе местной фауны. К таковым в первую очередь относятся вредители, попавшие из других стран или районов (колорадский жук в европейской части России, гессенская мушка в Северной Америке). Подобные виды выделяют в категорию заносных.
Каждая фауна характеризуется определенными, свойственными только ей признаками, позволяющими сравнивать ее с другими фаунами. Самым важным признаком любой фауны является ее видовой состав. Фаунистическое исследование начинается с учета видов, обитающих в пределах изучаемого района, т. е. с инвентаризации фауны. Количество видов, входящих в состав фауны, отражает ее богатство.
Существенным признаком любой фауны оказывается экологическая природа составляющих ее видов. Для фауны тропического леса характерно наличие большого количества видов, приуроченных в своем обитании к деревьям. К ним относятся лазающие формы млекопитающих, птиц, рептилий и амфибий, многих насекомых и других животных, а также виды, связанные с деревьями трофическими отношениями: плодоядные формы, потребители листвы, потребители и разрушители древесины. Большинству животных тропической фауны свойственна круглогодичная активность. Фауне степей присущи другие экологические типы — животные бегающие и роющие, проводящие зиму в состоянии спячки, питающиеся жесткой травой, семенами злаков и др.
Главнейшей особенностью фауны являются ее связи с соседними, а также с более отдаленными фаунами. Эти связи можно охарактеризовать показателями общности систематического состава фауны, обычно выражаемой в процентах.
Наряду с фаунистическими связями не менее существен учет еще одного важнейшего признака — степени самобытности фауны, выражающейся в наличии эндемичных видов или родов. Чем выше систематический ранг эндемиков, тем самобытнее фауна. Так, присутствие в составе фаун эндемичных отрядов и семейств отражает длительность развития фауны в условиях изоляции. В то же время наличие только эндемичных видов, а тем более подвидов, свидетельствует об относительной молодости фауны и прочных связях ее с другими фаунами. Примером древней и самобытной фауны служит фауна Австралийской области, в составе которой насчитывается 8 эндемичных семейств млекопитающих (сумчатые) и не менее 3 эндемичных семейств птиц, не считая большого количества эндемичных родов всех классов позвоночных животных. Самобытна и фауна Неотропической области. На территории Южной Америки обитают представители многих эндемичных семейств (12 из них млекопитающие, 26 — птицы), множество родов рептилий, амфибий, насекомых и т. д. Голарктическая фауна, куда относится и фауна России, значительно беднее и менее оригинальна, чем упомянутые выше, и насчитывает всего несколько эндемичных семейств позвоночных животных, хотя эндемичных родов немало.
Каждая фауна обладает определенной систематической структурой, иначе говоря, специфичным распределением видов между родами, семействами и более высокими систематическими единицами. Структура фауны познается не только через количественное соотношение различных систематических единиц. Серьезное значение имеет также географический анализ ее, т. е. установление сходства и различий в распределении входящих в нее видов. Виды, отличающиеся сходным распространением, представляют географические элементы фауны. Соответственно характеру распространения эти элементы носят определенные названия: северные, южные, западные, восточные. Такие названия пригодны для географического анализа фаун лишь ограниченных территорий. Если же рассматриваются ареалы видов той или иной фауны в целом, то наименования географических элементов будут другими. В зоогео-графической литературе широко применяются термины: европей-ско-сибирский, восточносибирский (ангарский), центральноази-атский, бореальный и др. Термин «заносные», или «адентивные», элементы означает, что данные виды натурализовались благодаря заносу извне и не являются единой географической группой.
Таким образом, географический анализ фауны дает представление о типе распространения входящих в нее видов. Но для познания фауны этого мало. Необходимо выяснить вероятное происхождение видов, как они попали в состав фауны и как происхождение каждого из них сказывается на распространении. Ответы на данные вопросы дает исторический (или генетический) анализ фауны. Он базируется на изучении ареалов не только видов, но и родов. При проведении его требуется прежде всего решить вопрос, какие элементы фауны возникли в пределах изучаемой территории и какие попали в результате расселения из других центров. Первые получили название автохтонных элементов, вторые — аллохтонных.
Изучение структуры фауны является первым этапом ее исследования. Но этого недостаточно, чтобы понять ее особенности и в дальнейшем судить о ее происхождении. Специфика одной фауны, ее отличия от других становятся очевидными только при сравнении их структур. Для этого обычно берутся две соседние фауны, в отдельных же случаях требуется сравнение и отдаленных фаун. Необходимым условием такого рода исследований является примерно равная степень изученности сравниваемых фаун, хотя это не всегда соблюдается из-за неполноты фаунистических сведений.
Сравнительный анализ начинается с изучения списков видов локальных фаун. При этом обращается внимание на качественные отличия, т.е. на таксоны (виды, роды и т.д.), которые присутствуют лишь в одной из сравниваемых фаун. Количество видов (или других таксонов), характерных только для одной локальной фауны, служит показателем ее специфики.
При сравнительном анализе большое значение имеют определение степени общности и различия между двумя фаунами. Более точно это можно установить с помощью количественных методов. Для математического выражения степени сходства фаун двух районов предложен ряд формул. Наиболее часто применяется формула Жаккара, по которой определяют коэффициент общности (К),
выражающийся в процентах:
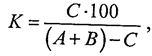
где С —
число видов, общих для обеих фаун;
А, В —
число видов соответственно в первой и второй фауне.
Используется также формула Сьеренсена:
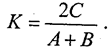
Оба коэффициента в общем однотипно отражают степень сходства фаун сравниваемых районов. Применяют и другие показатели, например индекс различия фаун.
Одной из важнейших особенностей фауны является эндемизм. Эндемичными систематическими единицами называются виды, роды и другие таксономические единицы, которые встречаются только в какой-то определенной области и нигде более. В силу этого эндемики представляют специфический компонент любой фауны и абсолютно отличают ее от других фаун. Если более широко распространенные виды указывают на связи между фаунами, то эндемики резко отличают их друг от друга.
Количество эндемиков в различных фаунах неодинаково. Наиболее высок процент эндемизма в островных фаунах, а на континентах — в районах с сильно расчлененным рельефом, т. е. в горных странах, поскольку географическая изоляция — необходимое условие процесса видообразования.
Для образования эндемичных видов требуется не только географическая изоляция, но и продолжительное время. Длительное и непрерывное развитие фауны благоприятствует возникновению новых видов, в то время как быстрые изменения условий существования, наоборот, ведут к вымиранию одних и расселению других. Обилие эндемиков отражает длительность развития фаун.
Различия между эндемичными и неэндемичными компонентами фауны не абсолютны. В большинстве фаун есть так называемые субэндемики, или почти эндемичные таксоны (роды, виды).
В историческом смысле можно говорить о прогрессивном и реликтовом эндемизме. Прогрессивными эндемиками (неоэндеми-ками) считаются виды или роды, возникшие в фауне недавно и еще не распространившиеся за пределы ее территории. Это молодые новообразования, которые незначительно отличаются от остальных видов этого же рода. Между ними и другими видами, обитающими на одной территории, существуют легко различимые связи. Часто прогрессивные эндемики образуют группы близкородственных видов. Их экологические требования соответствуют окружающим условиям среды.
К реликтовым эндемикам (палеоэндемикам) относятся животные, ареал которых в прошлом был гораздо больше, а в настоящее время они сохранились лишь на отдельной территории. Реликтовые эндемики (их иногда называют консервативными) занимают в фауне, как правило, изолированное положение. Близкородственных видов у них нет, они систематически обособлены. Реликтовые эндемичные роды часто включают лишь несколько или всего-навсего один вид (монотипичные роды). Экологическая обстановка их «устраивает» относительно. Конечно, если бы она полностью не отвечала их потребностям, они бы давно вымерли. Однако распространение этих видов в ареале носит пятнистый характер, они узколокальны и связаны с ограниченными биотопами. Наличие в фауне реликтовых эндемиков говорит о том, что благоприятная для них обстановка сохраняется длительное (в геологическом смысле) время, пусть даже на небольшом пространстве. Замечательным примером реликтового эндемизма служит гаттерия Sphenodonpunctatus — единственный представитель семейства клинозубых Sphenodontidae, отряда клювоголовых Rhyn-chocephalia рептилий, живущая в Новой Зеландии. Для консервативных эндемиков высокого ранга нередко характерно дизъюнктивное распространение.
Каждая фауна имеет свою историю. Формирование фауны, или фауногенез, продолжается очень длительное время, измеряемое иногда целыми геологическими периодами. Те виды и группы животных, которые живут в настоящее время на той или иной территории, могут быть весьма отдаленными потомками других животных, давно вымерших либо больше не встречающихся в каких-то частях Земли. Описанием процесса зарождения и смены фаун занимается генетическая, или историческая, зоогеография. Для того чтобы ее выводы и заключения были достоверными, необходимо привлечь целый арсенал прямых и косвенных доказательств смены комплекса видов животных во времени и пространстве. Лучшую опору для познания прежнего облика фауны дают ископаемые находки. Они и являются прямыми доказательствами пребывания какого-нибудь вида в составе изучаемой фауны в определенный период ее развития. Примерами могут служить находки скелетов мамонта в Сибири или окаменевших костей динозавров в Монголии.
Но такие находки редки, а многие группы животных не сохраняются в ископаемом состоянии вообще. При отсутствии палеонтологических материалов характер фауногенеза устанавливается по косвенным свидетельствам — данным исторической геологии, палеогеографии, выводам на основе теории эволюции. Большую пользу приносят также и собственные зоогеографические сведения о дизъюнкциях ареалов, эндемиках и реликтах.
Своеобразие фауны островов интересовало ученых с момента зарождения зоогеографии как науки. В ряде случаев изучение особенностей островной биоты приводило к появлению крупных обобщений и биологических теорий. Так, знакомство Ч. Дарвина с фауной Галапагосских островов навело его на мысль о дивергенции близких видов, происходящих от общего предка. Специально фауне островов А. Уоллес посвятил книгу «Островная жизнь» (1880). Острова интересуют биологов, во-первых, из-за относительной простоты структуры их биоценозов и трофических связей между видами, что облегчает изучение сообщества, а во-вторых, из-за необычных, порой удивительных форм жизни.
Все разнообразие островов Мирового океана можно свести к нескольким основным типам. Главным критерием выделения типов является происхождение островов. Чаще всего они бывают океаническими и материковыми. Океанические острова никогда не были частями материков, а образовались в результате поднятия дна океана, либо извержения подводных вулканов и застывания лавы, либо, наконец, вследствие жизнедеятельности рифообразующих кораллов. Примерами подобных островов являются архипелаги Микронезии и Полинезии в Тихом океане, остров Св. Елены и др.
Материковые, или континентальные, острова представляют собой части материков, отделившиеся от них в определенную геологическую эпоху. К ним относятся Британские, Большие Зонд-ские (Ява, Суматра, Калимантан и др.), Тайвань, Новая Гвинея, Тасмания и многие другие.
Остров материкового происхождения при отделении от материка наследует тот или иной (но не полный) набор местообитаний и связанных с ним видов. С одной стороны, этим определяется умеренное зоогеографическое своеобразие таких островов: с момента их отделения от материка начинается дифференцировка некоторых видов, образуются местные формы (подвиды, виды, реже роды). С другой стороны, происходит объединение первоначальной фауны за счет исчезновения ряда видов, причины которого будут рассмотрены ниже. Наконец, на материковых островах отсутствуют виды или роды, возникшие на материках или проникшие на них после отделения острова. Островная фауна в любом случае будет беднее предковой материковой.
На острове океанического происхождения первое время после его образования вообще нет никаких организмов. Современный органический мир таких островов формируется за счет видов, попавших на них путем залетов или заносов, т. е. активным или пассивным путем. В некоторых же случаях в фауну входят обитатели океана, приспособившиеся к жизни на суше: некоторые рыбы, раки (например, пальмовый вор), пресноводные моллюски и др. Фауна океанических островов от начала своего образования и до достижения устойчивого состояния непрерывно обогащается.
Независимо от происхождения фауна острова несет «островной» отпечаток, степень которого зависит от площади острова, расстояния его до ближайшей суши, от климатической зоны, в которой он располагается, и т.д. Иначе говоря, существуют конкретные признаки островных фаун, 4 из них являются основными: бедность и дефектность, выраженный эндемизм, ограниченное количество архаических форм и, наконец, адаптивная радиация в рамках небольшого количества эндемичных групп.
Относительная бедность и дефектность (систематическая неполнота) островных фаун объясняется тем, что площадь острова всегда меньше площади ближайшего материка и, стало быть, часть биотопов на нем отсутствует или занимает по сравнению с материком меньшую территорию. К примеру, на острове может не быть крупных рек, а саванны или леса могут иметь меньшую протяженность. На континентальном острове должны жить все группы животных, характерные для материка, от которого он отделился. Отсутствие на острове каких-то групп или видов объясняется либо тем, что они приурочены к биотопам, не имеющимся на острове, либо их вымиранием вследствие недостатка условий для поддержания необходимой численности популяций. К факторам, вызывающим островное обеднение, относится, естественно, и антропический.
На океанических островах могли укорениться лишь виды, способные активным или пассивным путем пересекать водные преграды. А поскольку такой способностью обладают далеко не все животные, бедность и дефектность фауны островов можно предсказать заранее. На океанических островах обитают летающие животные — птицы, летучие мыши, многие насекомые. Из нелетающих форм раньше других здесь появляются грызуны, из рептилий — гекконы. И те и другие могут заноситься на «плотах», т. е. плавающих стволах деревьев, вырванных или сломанных тропическими ураганами и выброшенных в море.
Даже крупные древние континентальные острова имеют относительно бедную и дефектную фауну. Так, на Мадагаскаре обитают только 2 семейства амфибий, но нет строго пресноводных рыб, сухопутных черепах из семейств трехкоготные и пресноводные, из змей отсутствуют аспидовые и гадюки и такие характерные для Африки семейства птиц, как страусы, птицы-секретари, цесарки, журавли, птицы-носороги, дятлы, вьюрковые и др. Из млекопитающих на Мадагаскаре нет и типично африканских семейств — слонов, носорогов, обезьян, кошек, гиен, антилоп, трубкозубов и т. д. Особенно следует подчеркнуть отсутствие копытных, кроме речной свиньи, и обилие хорошо летающих рукокрылых, родственных как африканским, так и индо-малай-ским формам.
Следовательно, бедность и дефектность фауны — общий и характерный признак всех островов, проявляющийся в тем большей степени, чем остров меньше по площади и дальше находится от материка или другого острова.
Эндемизм — второй характерный признак островной фауны — является результатом изоляции острова от других участков суши. Степень эндемизма тем больше, чем древнее остров и чем он более удален от материка. Таким образом, изоляция является причиной не только бедности и дефектности фауны, но и ярко выраженного островного эндемизма. Так, на Мадагаскаре, отдаленном от Африки морским проливом шириной 400 км и ставшем островом еще в мезозое, живут эндемичные семейства млекопитающих (тенреки из насекомоядных, 3 семейства лемуров) и птиц (нелетающие пастушковые куропатки, филепитты, ванги, а в прошлом и эпиорнисы), многочисленные эндемичные роды и огромное количество видов всех классов животных. Фауна птиц Новой Каледонии содержит более 20 % эндемичных видов и 1 эндемичное семейство.
Геологически молодые, расположенные близко от материка острова (например, Британские, Сахалин) эндемиков высокого ранга не имеют, хотя эндемичные подвиды или даже виды здесь обитают. Следовательно, процент эндемизма и систематический ранг эндемиков могут служить показателем геологического возраста острова. К примеру, согласно зоогеографическим данным, и Британия, и Сахалин стали островами только в четвертичном периоде, что подтверждается геологическими исследованиями.
Богатство фауны острова, степень ее эндемизма могут зависеть, кроме того, от его площади, разнообразия рельефа и других природных условий, а также от климатической зоны, в которой он находится. Все это создает большее или меньшее разнообразие экологических ниш.
Третья особенность островных фаун заключается в том, что они нередко являются последними убежищами архаичных видов животных. Такие виды немногочисленны и встречаются на Мадагаскаре, в Новой Зеландии, на Сулавеси, а также на Антильских островах. В одной из оригинальнейших наземных фаун — новозеландской — вообще отсутствуют аборигенные млекопитающие, но имеется древнейший вид рептилий — гаттерия, или туатара, а также гладконогие лягушки лиопельмы (2—3 вида) — древние и архаичные представители отряда бесхвостых. В состав современной мадагаскарской фауны входят древние тенреки (30 видов), настоящие лемуры, индри и руконожки (из примитивных приматов), легуаны (из игуановых рептилий). Интересно общее распространение последних. Легуаны представлены многочисленными видами в фаунах Северной и Южной Америки, а в Восточном полушарии встречаются на островах Фиджи и Тонга. На островах Куба и Гаити до нашего времени дожили архаичные щелезубы (2 вида), представители насекомоядных млекопитающих. В ископаемом состоянии близкие роды обнаружены в Северной Америке.
Исследователей фауны островов часто поражают ее необычные представители: например, с одной стороны, гиганты — гигантский варан на острове Комодо, черепахи на острове Маврикий, истребленные человеком птицы эпиорникс на Мадагаскаре и дронты на Реюньоне (также достигали гигантских размеров), с другой стороны — карлики, такие как мелкая форма тигра на Зондских островах, мелкие барсуки и лисицы на острове Сардиния и т. д. (ископаемые слоны на острове Мальта достигали размеров кабана). Островная популяция лошадей, предки которых были завезены с материка, выродились в мелкую породу — пони. Причины как одного, так и другого явления до настоящего времени не выяснены.
Немногочисленные, а то и одиночные виды животных, попав на острова, оказываются родоначальниками процветающих и достаточно многочисленных эндемичных групп. Этот феномен адаптивной радиации в рамках небольшого количества эндемичных групп представляет собой четвертую особенность островных фаун. Адаптивная радиация обусловливается наличием на островах разнообразных незанятых экологических ниш. Поэтому некоторые виды, попав на острова, настолько изменяются, что иногда уже мало чем напоминают предковую форму. Классическим примером адаптивной радиации являются гавайские птицы-цветочницы (семейство Drepanididae). Их предок, американский вьюрок, залетев на Гавайские острова, не нашел здесь конкурентов и дал начало нескольким эволюционным ветвям, нынешние представители которых по внешнему виду и образу жизни напоминают зябликов, медососов, пищух, дубоносов и дятлов.
Важнейшей задачей систематической зоогеографии является характеристика регионов суши и океана по составу их фаун. Вопрос о принципах их выделения относится к центральным вопросам современной зоогеографии, так как разработаны они до настоящего времени недостаточно. Издавна схемы зоогеографического районирования базируются на изучении ареалов видов и целых фаунистических комплексов, а различия между фаунами объясняются историческими условиями их формирования.
При зоогеографическом районировании могут быть два подхода: фауногенетический и ландшафтно-зональный (зонально-климатический). Первый, примененный еще П.Склэтером (1875) и А.Уоллесом (1876), а затем широко использованный М.А.Мензбиром (1934), А. П. Семеновым-Тян-Шанским (1936), И.И.Пузановым (1938) и другими учеными, основан на детальном исследовании ареалов видов и фаунистических комплексов и на выяснении исторически обусловленных различий между фаунами. Главную роль при выделении зоогеографических регионов (зоо-хоронов) играют виды с узкими ареалами, т. е. эндемики и реликты. Именно они подчеркивают самобытность и древность определенной фауны.
Ландшафтно-зональный подход базируется на изучении животного населения, т. е. совокупности животных, связанных с общностью местообитаний и разнообразными отношениями друг с другом и компонентами растительного сообщества. Здесь используются типологические критерии (сообщество, животное население), в случае же фауногенетического подхода зоогеограф имеет дело с уникальными категориями (фаунистический комплекс, фауна).
При зоогеографическом районировании стоит задача отразить сходство и различия сравниваемых фаун, оценить их в количественных показателях и определить, где проходят границы между территориями, занятыми различными фаунами.
Большинство зоогеографов считают, что при выделении зоо-хоронов высокого ранга, например царств, полцарств и областей, нужно обязательно учитывать исторический фактор (главные этапы эволюции фаун), иначе не могут быть поняты пути их становления. В противоположность этому выделение округов и районов требует в основном учета ландшафтно-зональных особенностей территории.
Для ограничения территории зоохоронов используется и так называемый метод синперат. Он заключается в том, что на карту линейным способом наносят границы ареалов возможно большего числа видов или родов. Многие границы совпадут или окажутся близко друг к другу. Полосы совпадения или сближения (так называемые сгущения границ ареалов) именуют синператами. Они и служат границами выделов. Как правило, синператы совпадают с различного рода преградами к расселению видов.
Что касается списков таксонов (семейств, родов и видов), используемых для районирования, они должны быть подтверждены фактическими находками, а линейные границы ареалов должны отражать пределы действительного распространения таксонов, а не являться экстраполяцией. В сравниваемых списках фаун должны фигурировать таксоны равноценного ранга (нельзя сравнивать, к примеру, списки, один из которых представляет собой перечень видов всей фауны птиц района, а другой — только воробьиных). Наконец, из сравниваемых списков необходимо исключить те виды животных, которые были случайно или преднамеренно завезены человеком (например, колорадский жук или енот-полоскун в фауне России). Это требуется для правильного понимания географической истории группы.
Определенному центру эволюции фауны соответствует зоогеографическая область и тем более царство. И. И. Пузанов (1949) справедливо считал, что и при установлении подобластей решающим критерием должен быть не тип современного распределения биоценозов, который зависит от современных климатических особенностей, а история фауны, т. е. ее принадлежность к тому или иному центру эволюции. Подобласти как центры эволюции фаун более низкого порядка могут включать в себя несколько экологических зон с характерными для них биотопами.
При выделении же провинций и районов, которые трудно понять исторически, главным критерием может быть статистика видов, т.е. распределение их по климатическим зонам и ландшафтам. Границы этих зон большей частью и будут служить границами зоогеографических регионов ранга провинций и районов. Таким образом, разномасштабное зоогеографическое районирование производится на компромиссной основе: зоохороны высокого ранга (царство, область, иногда и подобласть) определяются с учетом истории фауны, а низкого (провинция, район, участок) — главным образом экологически.
Таким образом, как было отмечено выше, зоогеографическое районирование является завершающим, синтетическим этапом фаунистического исследования достаточно большой территории или акватории. До сих пор для решения этой задачи исследователь должен был обобщить первичные данные о находках отдельных таксонов в конкретных точках и составить списки фаун неких оперативных территориальных единиц, обычно называемых выделами. Именно на этом этапе в сугубо объективную картину первичных данных вторгается элемент субъективности, ибо от выбранных исследователем размеров и конкретных границ выделов в значительной мере зависит результат районирования изученного региона. Дальнейший процесс районирования обычно заключается в количественной оценке сходства фаун выделов с помощью одного из многочисленных индексов, а затем — в кластерном анализе выделов на основе данных об их сходстве. Уязвимость субъективного подхода к выбору рабочих выделов хорошо осознавалась многими исследователями. В некоторых случаях выбор выделов может быть облегчен наличием естественных границ между ними, например, ими могут быть высокие горные хребты, но такие случаи являются, скорее, исключениями.
Попытки прямого использования данных по фаунам конкретных точек в зоогеографическом районировании нам вообще неизвестны. Причина этого очевидна — изученность фаун отдельных точек крайне неравномерна. Это хорошо иллюстрируют наши данные (Г.М.Абдурахманов и др., 1995) по географическому распространению видов жуков-чернотелок (Coleoptera, Tenebrionidae) на Кавказе и в Монголии. Бросается в глаза большое количество точек, где были зарегистрированы 1 или 2 вида, хотя минимальная локальная фауна чернотелок должна включать, по нашим представлениям, не менее 3—4 видов. Довольно часто этим единственным видом является фоновый широко распространенный вид. При любом известном способе вычисления сходства фаун такие точки имеют максимальное сходство, и их объединение на первых же шагах кластерного анализа приводит к абсурдным результатам.
Невозможность прямого использования в зоогеографическом районировании данных по фаунам точек особенно досадна сейчас, когда компьютеризация зоологических исследований и распространение технологий машинных баз данных приводят к накоплению в ЭВМ больших массивов истинно первичных фау-нистических данных — регистрации таксонов в конкретных точках. Только такие данные являются вечными и не потеряют ценности при дальнейшем развитии техники и повышении точности исследований в отличие от данных, обобщенных в виде описаний ареалов или в виде указаний на квадраты фиксированной сети, подобной той, которая принята для картирования ареалов беспозвоночных Европы.
Предлагаемый новый метод опирается на следующие ключевые положения.
1. Кластерному анализу для целей районирования подвергаются не фауны субъективно выделенных исследователем выделов, а объективно регистрируемые фауны отдельных точек.
2. Собственно сравнению фаун отдельных точек предшествует всесторонний компьютерный анализ имеющихся данных, в ходе которого можно провести селекцию данных, чтобы повысить их значимость и достоверность.
3. На каждом шаге кластеризации пересчет матрицы сходства точек и кластеров производится не на основе вычисления их усредненного сходства, а на основе объединения фаун сливающихся точек или кластеров и нового вычисления их сходства.
4. В кластерном анализе учитывается не только сходство фаун точек, но и физическое расстояние между самими точками, рассчитываемое по их точным географическим координатам. Это позволяет сгладить случайное сходство далеко отстоящих точек, возникающее зачастую из-за неполной изученности их фаун и случайных совпадений.
5. Процесс кластерного анализа фаун точек протекает параллельно как в памяти компьютера (в виде преобразований матриц сходства), так и на экране дисплея (в виде динамического преобразования закрашенных в разные цвета областей на карте). Это дает исследователю уникальную возможность визуально контролировать каждый шаг процесса кластеризации точек и получать при необходимости дополнительную информацию о каждом шаге.
Принятая в учебнике система фаунистических регионов суши представляет собой модернизированную схему Склэтера—Уоллеса. Схема районирования морского и пресноводного биоциклов принята соответственно по схемам Г.Деллатина (1967) и Я.И.Старобогатова (1970).
Перечень причин, по которым надо изучать океаны, не может быть исчерпывающим, поскольку объем имеющихся знаний об океанах неуклонно возрастает. Сегодня в такой перечень следует включить следующее.
Океаны существуют, и наше любопытство перевешивает все остальные причины для их изучения.
Океаны дают нам пищу, поэтому нужно принять как факт нашу давнюю зависимость от океанов и использование их в качестве источника пропитания для человечества.
Способы транспортировки, которые можно применять в океанах и атмосфере благодаря текучести среды, во многих отношениях превосходят наземный транспорт, но для их эффективного использования необходимы крупномасштабные исследования течений и ветров.
Океаны — важный источник минеральных ресурсов: от соли до таких экзотических элементов, как магний, и от фосфатных удобрений до прозаического песка.
Морская вода во всех ее фазах — жидкой, твердой и парообразной — служит основной средой, через которую тепловая энергия распространяется по всей планете. Поэтому исследования погоды и климата теснейшим образом связаны с изучением океанов.
Морская вода благодаря ее способности разлагать сложные молекулярные структуры содержит практически все известные элементы. Однако она сохраняет свою химическую стабильность, так что никогда не бывает ни слишком кислой, ни слишком щелочной. Эта «автоподстройка» играет важнейшую роль в способности морской воды поддерживать жизнь. Действительно, только в океанах, как принято считать, возможно было развитие «живых» молекул на Земле.
Морская вода в силу своих абсорбционных свойств поглощает и выделяет газы, обмениваясь ими с атмосферой; тем самым она косвенно включена в процесс переноса лучистой энергии, происходящей между Землей и космическим пространством.
Океаны занимают более 70 % земной поверхности (рис. 134), и испарение воды из них превышает поступление ее с атмосферными осадками. Поэтому именно океаны приводят в действие гидрологический цикл — круговорот воды в природе, от которого полностью зависит вся наземная жизнь. Океан — и в тропиках, и вблизи полюсов — нагревается и остывает сверху вниз; его тепловой баланс почти полностью определяется процессами, происходящими только на его поверхности. Атмосферная же циркуляция, наоборот, приводится в действие снизу вверх, так как испаряющаяся морская вода поступает в атмосферу в основании воздушного столба.
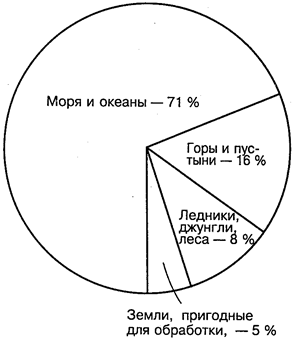
Рис. 134. Соотношение океанов и суши на поверхности Земли
Океаны в любой момент содержат значительную долю всей кинетической энергии, получаемой Землей от Солнца. Иными словами, количество солнечной энергии, запасенной водным столбом с единичной площадью сечения, значительно превосходит величину этой энергии, содержащейся в равном по площади сечения столбе горных пород суши или атмосферного воздуха. Поэтому, пытаясь найти источники энергии, альтернативные минеральному топливу, мы должны основное внимание направить на океаны.
Океаны и суша распределены по земной поверхности несимметрично. Это обстоятельство, будучи результатом сложной геологической истории Земли, имеет важнейшее значение для динамики как океана, так и атмосферы; оно же решительно повлияло на развитие человечества.
Океаны, если говорить о способности поддерживать живые сообщества, предоставляют почти в 80 раз больше пространства для жизни, чем наземный мир. Однако из-за того, что жидкость, заполняющая океанические бассейны, способна легко перемещаться во времени и в пространстве, число различных видов организмов в океанах гораздо меньше, чем на суше.
Морская вода благодаря ее высокой удельной теплоемкости сохраняет сравнительно постоянную температуру, несмотря на то, что оказывается в весьма широком диапазоне условий — от тропических зон с их избыточным солнечным нагревом до полярных зон с избыточным охлаждением, происходящим также путем излучения. Постоянство температуры оказывает огромное влияние на образ жизни морских организмов, делая его совершенно отличным от способа существования наземных видов.
Морская вода в тысячу раз плотнее воздуха, в котором обитает большинство наземных организмов, и поэтому формы жизни, существующие в океане, в среднем имеют значительно меньшие размеры, чем те, что встречаются на суше. Популярное изречение о том, что «при такой жизни лучше быть маленьким», особенно подходит для условий жизни в море. Однако в океане живут и крупнейшие из животных, когда-либо обитавших на Земле, — синие киты.
Окраины океанических бассейнов, где суша встречается с морем, относятся к областям Земли с самой высокой продуктивностью органического вещества. Их продуктивность обусловлена тем, что это зоны сближения энергии и массы: океаны несуг к своим берегам энергию волн, собираемую с обширных просторов водной поверхности, подверженной действию ветра, а реки несут химическое сырье, без которого невозможна жизнь.
Люди также устремляются к окраинам океанов, не только создавая на побережье множество поселений, но и перенося в заселенные прибрежные районы большую часть всего органического материала, производимого сельским хозяйством, горными работами и промышленностью во внутренних областях материков.
Полярные океаны — «последний рубеж» — быть может, являются важнейшими областями, обеспечивающими продолжение нашего существования. Постоянство климата на Земле зависит от энергии перехода между жидкой и твердой фазами воды и от альбедо (способности отражать солнечные лучи) покрытых льдом частей океана.
Исследование океана связано с изучением великого множества сложных процессов: физических, биологических, химических, геологических, метеорологических и др. В ткань перечисленных процессов вплетается и человеческая деятельность. Наша задача — «распустить» эту ткань на отдельные нити, описать каждую нить качественно и количественно, а затем снова соединить их, пытаясь тем самым понять изменчивость океана. Однако в практическом смысле изучение океанов определяется необходимостью лучше знать, как воздействуют океаны на нас, и наоборот.
Фаунистическое расчленение литорали (рис. 135,1) проявляется очень четко, так как отдельные области этого биохора достаточно сильно изолированы как сушей и климатическими зонами, так и широкими отрезами открытого моря.
Выделяют центральный Тропический регион и расположенные к северу от него Бореальный, а к югу — Антибореальный регионы. В каждом из них выделяют разное количество областей. Последние, в свою очередь, подразделяются на подобласти.
Рис. 135. Фаунистическое расчленение литорали (1) и пелагиали (2)
Мирового океана (из Г.Делаттина, 1967):
А —
Тропический регион: А, —
Индо-Пацифическая область; А. —
Тропико-Атлантическая область; В —
Бореальный регион: В. —
Арктическая область; В1
—
Борео-Пацифическая область; В2
—
Борео-Атлантическая область; С —
Антибореальный регион: С — Антарктическая область; С — Антибореальная область
Этот регион характеризуется наиболее благоприятными условиями существования, что обусловило формирование здесь наиболее полной, гармонически развитой фауны, не знавшей перерывов в эволюции. Подавляющее большинство классов морских животных имеют в регионе своих представителей. Тропическая зона по характеру фауны четко расчленяется на две области — Индо-Пацифическую и Тропико-Атлантическую.
Индо-Паорфическая область.
Данная область охватывает громадное пространство Индийского и Тихого океанов между 40° с. ш. и 40° ю. ш., и только у западного побережья Южной Америки южная граница ее резко сдвинута к северу под влиянием молодого течения. Сюда же относятся Красное море и Персидский залив, а также бесчисленные проливы между островами Малайского архипелага и Тихого океана. Благоприятный температурный режим, обусловленный большой площадью мелководий, и стабильность среды на протяжении многих геологических периодов привели к развитию здесь исключительно богатой фауны.
Млекопитающие представлены дюгонями (род Dugong) из отряда сирен, один вид которых живет в Красном море, другой — в Атлантике, третий — в Тихом океане. Эти крупные животные (3-5 м в длину) обитают в мелких заливах, обильно заросших водорослями, и изредка заходят в устья тропических рек.
Из морских птиц, связанных с побережьями, для Индо-Пацифической области характерны мелкие буревестники и гигантский альбатрос (Diomedeaexulans).
Морские змеи (Hydrophiidae) представлены большим количеством (до 50) характерных видов. Все они ядовиты, многие имеют приспособления для плавания.
Рыбы морской фауны чрезвычайно разнообразны. Они чаще всего ярко окрашены, покрыты разноцветными пятнами, полосами и т. п. Из них следует упомянуть сростночелюстных рыб — диодонов, тетрадонов и кузовков, рыб-попугаев (Scaridae), у которых зубы образуют сплошную пластинку и служат для откусы-вания и дробления кораллов и водорослей, а также рыб-хирургов, вооруженных ядовитыми колючками.
Громадного развития в море достигают коралловые рифы, состоящие из зарослей шестилучевых (Madrepora, Fungia и др.) и восьмилучевых (Tubipora) кораллов. Коралловые рифы следует считать наиболее типичным биоценозом Индо-Пацифической литорали. С ним связаны многочисленные моллюски (Pteroceras и Strombus), отличающиеся ярко разрисованными и разнообразными по форме раковинами, гигантские тридакны массой до 250 кг, а также голотурии, служащие предметом промысла (употребляй ются в пищу в Китае и Японии под названием трепанга).
Из морских кольчатых червей отметим знаменитого пололо. Большие массы его в период размножения поднимаются на поверхность океана. Эти беспозвоночные употребляются в пищу полинезийцами.
Местные отличия фауны Индо-Пацифической области позволили выделить в ней Индийско-Западнопацифическую, Восточнопацифическую, Западноатлантическую и Восточноатлантиче-скую подобласти.
Тропико-Атлантическая область.
Эта область гораздо меньше по протяженности, чем Индо-Пацифическая. Она охватывает литораль западного и восточного (в пределах тропической Атлантики) побережья Америки, воды Вест-Индского архипелага, а также западное побережье Африки в границах тропической зоны.
Животный мир этой области значительно беднее предыдущей, лишь вест-индские моря с их коралловыми рифами содержат богатую и разнообразную фауну.
Морские звери здесь представлены ламантинами (из тех же сирен), способными заходить далеко в реки тропической Америки и Африки. Из ластоногих встречаются белобрюхие тюлени, морские львы и галапагосский котик. Морских змей практически нет.
Фауна рыб разнообразна. Она включает гигантских скатов-мант (до 6 м в поперечнике) и крупного тарпона (до 2 м в длину), являющегося объектом спортивного лова.
Коралловые рифы достигают пышного развития лишь в Вест-Индии, но вместо тихоокеанских мадрепор здесь распространены виды рода Асгорога, а также гидроидные кораллы МШерога. Чрезвычайно обильны и разнообразны крабы.
Литораль западного побережья Африки отличается самой бедной фауной, почти лишенной коралловых рифов и связанных с ними коралловых рыб.
Область разделяется на 2 подобласти — Западноатлантическую и Восточноатлантическую.
Регион расположен к северу от Тропического региона и охватывает северные части Атлантического и Тихого океанов. Он разделяется на 3 области: Арктическую, Борео-Пацифическую и Борео-Атлантическую.
Арктическая область.
К этой области относятся северные побережья Америки, Гренландии, Азии и Европы, расположенные за пределами влияния теплых течений (вне области остаются северные берега Скандинавии и Кольского полуострова, обогреваемые Гольфстримом). Охотское и Берингово моря по температурным условиям и составу фауны также относятся к Арктической области. Последняя соответствует экологической зоне, где температура воды держится на уровне 3—4 °С, а нередко и ниже. Большую часть года здесь сохраняется ледовый покров, даже летом на поверхности моря плавают льдины. Соленость Арктического бассейна относительно низкая из-за массы пресной воды, приносимой реками. Ледяной припай, характерный для данной области, препятствует развитию литорали на мелководьях.
Животный мир беден и однообразен. Наиболее типичными млекопитающими являются моржи, тюлени-хохлачи, полярный, или гренландский, кит, нарвал (дельфин с гипертрофированным в виде прямого рога левым клыком) и белый медведь, основное местообитание которого — плавающие льды. Птицы представлены чайками (прежде всего розовой и полярной), а также чистиками. Фауна рыб бедная: обычна сайка из тресковых, навага и полярная камбала.
Беспозвоночные более разнообразны и многочисленны. Малое количество видов крабов компенсируется богатством бокоплавов, морских тараканов и других ракообразных. Из моллюсков для арктических вод типична иольдия (Yoldiaarctica), очень много актиний и иглокожих. Особенностью арктических вод является то, что здесь на мелководьях живут морские звезды, ежи и офиуры, которые в других зонах ведут глубоководный образ жизни. В ряде районов фауна литорали более чем наполовину состоит из сидящих в известковых трубочках кольчатых червей.
Однообразие фауны данной области на всем ее протяжении делает излишним выделение в ней подобластей.
Борео-Пацифическая область. В
пределы области входят прибрежные воды и мелководья Японского моря и омывающие с востока Камчатку, Сахалин и северные Японские острова части Тихого океана, а кроме того, литораль его восточной части — побережья Алеутских островов и Северной Америки от полуострова Аляска до Северной Калифорнии.
Экологические условия в этой области определяются более высокими температурами и их колебаниями в зависимости от времени года. Температурных зон несколько: северная — 5—10 "С (на поверхности), средняя — 10-15 "С, южная — 15-20 °С.
Для Борео-Пацифической области характерны морская выдра, или калан, ушастые тюлени — морской котик, сивуч и морской лев, сравнительно недавно водилась стеллерова морская корова (Hydrodamalisgigas), полностью уничтоженная человеком. Из рыб типичны минтай, терпуг и тихоокеанские лососи — кета, горбуша, чавыча.
Разнообразны и обильны беспозвоночные литорали. Они нередко достигают очень крупных размеров (например, гигантские устрицы, мидии, камчатский краб).
Многие виды и роды животных Борео-Пацифической области сходны с представителями Борео-Атлантической области либо идентичны им. Это так называемое явление амфибореальности. Данный термин обозначает тип распространения организмов: они встречаются на западе и востоке умеренных широт, но отсутствуют между ними.
Таким образом, амфибореальность представляет собой один из видов разрыва ареалов морских животных. Объясняет этот вид разрыва теория, предложенная Л.С.Бергом (1920). Согласно данной теории, расселение животных бореальных вод через арктический бассейн происходило как из Тихого океана в Атлантику, так и наоборот в эпохи, когда климат был теплее современного, а выход из морей дальнего севера через пролив между Азией и Америкой осуществлялся беспрепятственно. Такие условия существовали в конце третичного периода, а именно в плиоцене. В четвертичном периоде резкое похолодание привело к исчезновению бореальных видов в высоких широтах, установилась зональность Мирового океана и сплошные ареалы превратились в разорванные, поскольку связь жителей умеренно теплых вод через полярный бассейн стала невозможной.
Амфибореальное распространение имеют чистиковые птицы, обыкновенный тюлень, или ларга (Phocavitulina), многие рыбы — корюшка, песчанка, треска, некоторые камбалы. Оно свойственно также ряду беспозвоночных — некоторым моллюскам, червям, иглокожим и ракообразным.
Борео-Атлантическая область.
Эта область включает большую часть Баренцева моря, Норвежское, Северное и Балтийское моря, литораль восточного побережья Гренландии и, наконец, северо-восток Атлантического океана к югу до 36° с. ш. Вся область находится под влиянием теплого течения Гольфстрим, поэтому ее фауна носит смешанный характер, и наряду с северными сюда входят субтропические формы.
Эндемичен гренландский тюлень-лысун. Морские птицы — кайры, гагарки, топорики — образуют гигантские гнездовья (птичьи базары). Из рыб обычны тресковые, среди которых встречается эндемичная пикша. Многочисленны также камбаловые, зубатки, скорпены, морские петухи.
Среди разнообразных беспозвоночных выделяются ракообразные -~ омар, различные крабы, раки-отшельники; иглокожие — красная морская звезда, красивая офиура «голова медузы»; из двустворчатых моллюсков широко распространены мидии и сердце-вишки. Кораллов много, но рифы они не образуют.
Борео-Атлантическую область обычно разделяют на 4 подобласти: Средиземноморско-Атлантическую, Сарматскую, Атлан-то-Бореальную и Балтийскую. К первым 3 относятся моря — Баренцево, Черное и Азовское.
Баренцево море располагается на стыке теплых атлантических и холодных арктических вод. В связи с этим фауна его носит смешанный характер и отличается богатством. Благодаря Гольфстриму в Баренцевом море почти океаническая соленость и благоприятный климатический режим.
Литоральное население его разнообразно. Из моллюсков здесь обитают съедобные мидии, крупные хитоны, гребешки; из иглокожих — красная морская звезда и еж Echinusesculentus; из ки-шечнополостных — многочисленные актинии и сидячие медузы Lucernaria; типичны также гидроиды. Колоссальные скопления образуют асцидии Rhailusiaobliqua.
Баренцево море относится к высококормным морям. Здесь широко развит промысел многочисленных рыб — трески, морского окуня, палтуса, пинагора. Из непромысловых рыб обитают колючие бычки, морской черт и др.
Балтийское море из-за мелководности, ограниченной связи с Северным морем, а также из-за впадающих в него рек сильно опреснено. Северная часть его зимой замерзает. Фауна моря бедная и смешанная по происхождению, так как к бореоатлантическим присоединяются арктические и даже пресноводные виды. К первым относятся треска, сельдь, салака, килька и морская игла. Из арктических видов можно назвать бычка-рогатку и рачка морского таракана. Из пресноводных рыб обитают судак, щука, хариус и ряпушка. Интересно отметить практически полное отсутствие здесь типично морских беспозвоночных — иглокожих, крабов и головоногих моллюсков. Гидроиды представлены Cordylophoralacustris, морские моллюски — морским желудем Balanusimprovisus, мидией и съедобной сердцевидкой. Встречаются и пресноводные беззубки, а также перловицы.
Черное и Азовское моря по своей фауне относятся к Сарматской подобласти. Это типичные внутренние водоемы, поскольку связь их со Средиземным морем осуществляется лишь через неглубокий пролив Босфор. На глубинах ниже 180 м вода в Черном море отравлена сероводородом и лишена органической жизни.
Фауна Черного моря исключительно бедна. Литоральная зона населена моллюсками. Здесь встречаются блюдечко Patellapontica, черная мидия, гребешки, сердцевидка и устрица; мелкие гидроиды, актинии (из кишечнополостных) и губки. Эндемичен ланцетник Amphioxuslanceolatus. Из рыб обычны губаны (Labridae), морские собачки (Blennius), скорпены, бычки, султанки, морские коньки и даже 2 вида скатов. У берегов держатся дельфины — пыхтун и афалина.
Смешанность фауны Черного моря выражается присутствием определенного количества средиземноморских видов наряду с чер-номорско-каспийскими реликтами и видами пресноводного происхождения. Средиземноморские иммигранты здесь явно преобладают, и «медитерранизация» Черного моря продолжается.
К югу от Тропического региона, аналогично Бореальному на севере, располагается Антибореальный регион. К нему относится литораль Антарктиды и субантарктических островов и архипелагов: Южно-Шетландских, Оркнейских, Южной Георгии и др., а также прибрежные воды Новой Зеландии, Южной Америки, юга Австралии и Африки. Именно вдоль тихоокеанского побережья Южной Америки из-за холодного течения граница Антибореаль-ного региона продвинута далеко на север, до 6° ю. ш.
На основании разобщенности участков литорали региона в нем выделяют две области — Антарктическую и Антибореальную.
Антарктическая область.
Область охватывает воды трех океанов, омывающих берега Антарктиды, и расположенные вблизи архипелаги. Условия здесь близки к арктическим, но еще более суровы. Граница плавающих льдов проходит примерно между 60—50° ю. ш., иногда несколько севернее.
Фауна области характеризуется наличием ряда морских млекопитающих: гривистого сивуча, южного котика, настоящих тюленей (морской леопард, тюлень Уэделла, морской слон). В отличие от фауны Бореального региона здесь совершенно отсутствуют моржи. Из птиц прибрежных вод следует назвать в первую очередь пингвинов, обитающих громадными колониями по берегам всех материков и архипелагов Антарктической области и питающихся рыбой и ракообразными. Особенно известны императорский пингвин, пингвин Aptenodytesforsteri и пингвин Аде-ли (Pygoscelisadeliae).
Антарктическая литораль весьма своеобразна из-за большого количества эндемичных видов и родов животных. Как это часто наблюдается в экстремальных условиях, сравнительно малому видовому разнообразию соответствует огромная плотность популяций отдельных видов. Так, подводные камни здесь бывают сплошь покрыты скоплениями сидячего червя, в большом количестве можно обнаружить ползающих по дну морских ежей, звезд и голотурий, а также скопления губок. Рачки-афиподы весьма разнообразны, и около 75% их эндемичны. Вообще антарктическая литораль, по данным российских антарктических экспедиций, оказалась намного богаче, чем можно было предположить, судя по суровым температурным условиям.
И среди литоральных, и среди пелагических животных Антарктической области встречаются виды, которые обитают и в Арктике. Такое распространение получило название биполярного. Под бипо-лярностью, как уже отмечалось, подразумевается особый тип дизъюнктивного расселения животных, при котором ареалы сходных или близких видов располагаются в полярных или чаще в умеренно холодных водах Северного и Южного полушарий с перерывом в тропических и субтропических водах. При исследовании глубоководной фауны Мирового океана обнаружено, что для организмов, считавшихся ранее биполярными, характерно сплошное распространение. Только в пределах тропической зоны они встречаются на больших глубинах, а в умеренно холодных водах — в литоральной зоне. Тем не менее случаи истинной биполярности не так уж редки.
Для объяснения причин, вызвавших биполярное распространение, были предложены две гипотезы — реликтовая и миграционная. Согласно первой биполярные ареалы были некогда сплошными и охватывали также тропическую зону, в которой произошло вымирание популяций определенных видов. Вторая гипотеза была сформулирована еще Ч. Дарвином и развита Л. С. Бергом. По этой гипотезе биполярность — результат событий ледникового периода, когда охлаждение коснулось не только арктических и умеренно холодных вод, но и тропиков, что дало возможность северным формам распространиться до экватора и дальше к югу. Окончание ледникового периода и новое потепление вод тропической зоны вынудило многих животных выселиться за ее пределы к северу и к югу или вымереть. Таким путем образовались разрывы. За время существования в изоляции северные и южные популяции успели преобразоваться в самостоятельные подвиды или даже близкие, но викарирующие виды.
Антибореальная область.
Собственно Антибореальная область охватывает побережья южных материков, расположенных в переходной зоне между Антарктической областью и Тропическим регионом. Положение ее аналогично таковому Борео-Атлантической и Борео-Пацифической областей в Северном полушарии.
Условия обитания животных этой области по сравнению с условиями других областей значительно лучше, фауна ее достаточно богата. К тому же она постоянно пополняется за счет выходцев из прилежащих частей Тропического региона.
Наиболее типична и богата антибореальная фауна Южноавстралийской подобласти. Морские звери здесь представлены южными котиками (род Arctocephalus), морским слоном, тюленем-крабоедом и морским леопардом; птицы — несколькими видами пингвинов из родов Eudyptes (хохлатым и малым) и Pygoscelis (P. papua). Из беспозвоночных следует назвать эндемичных плеченогих (6 родов), червей Terrebellidae и Arenicola, крабов рода Cancer, которые встречаются и в Борео-Атлантической подобласти Северного полушария.
Южноамериканская подобласть характеризуется тем, что литоральная антибореальная фауна ее распространена вдоль берегов Южной Америки далеко на север. Один из видов котиков Arctocephalussustralis и пингвины Гумбольдта доходят до Галапагосских островов. Продвижение этих и многих других морских животных на север вдоль восточного побережья материка облегчается Перуанским холодным течением и поднятием на поверхность придонных вод. Перемешивание слоев воды обусловливает развитие богатого животного населения. Одних только десятиногих раков здесь свыше 150 видов, и половина из них эндемична. Случаи биполярности известны и в этой подобласти.
Южноафриканская подобласть по площади невелика. Она охватывает атлантическое и индоокеанское побережья Южной Африки.
Открытые части Мирового океана, где жизнь протекает вне связи с субстратом, называются пелагиалью. Выделяют верхнюю зону пелагиали (эпипелагиаль) и глубоководную (батипелагиаль).
Эпипелагиальная зона расчленяется по своеобразию фауны на Тропический, Бореальный и Антибореальный регионы, которые, в свою очередь, подразделяются на ряд областей.
Регион характеризуется стабильно высокой температурой верхних слоев воды. Годовые амплитуды ее колебаний в среднем не превышают 2 °С. Температура слоев, располагающихся глубже, гораздо более низкая. В водах региона наблюдается довольно значительное видовое разнообразие животных, но огромных скоплений особей одного вида почти не бывает (рис. 136). Многие виды медуз, моллюсков (крылоногих и других пелагических форм), почти все аппендикулярии и сальпы встречаются только в пределах Тропического региона.

Рис. 136. Фауна теплых морей (из А.В.Воронова, 1963):
1 —
фаэтон; 2 —
рыба диодон; 3 —
мечехвост; 4 —
каури; 5 — морская черепаха каретта; 6 —
морской еж; 7— сростночелюстная рыба-балистес; 8 —
электрический скат; 9 —
летучая рыба; 10 —
жемчужница; 11 —
меч-рыба; 12 —
червь-палоло; 13 —
сифонофора; 14 —
морская змея; 15 —
кашалот; 16 —
коралл-фунгия; 17—
тунец; 18—
парусник
Атлантическая область.
Отличается эта область следующими характерными чертами фауны. Китообразные представлены полосатиком Брайда, а из рыб типичны макрели, угри, летучие рыбы, акулы. Из животных плейстона встречаются ярко окрашенная сифонофора — сильно жалящая физалия, или португальский кораблик.
Участок тропической Атлантики, названный Саргассовым морем, населен особым сообществом пелагических животных. Кроме уже упоминавшихся при общей характеристике моря обитателей нейстона на свободно плавающих саргассовых водорослях находят приют своеобразные морские коньки Hippocampusramu-losus и рыба-игла, причудливая рыба-антеннарий (Antennariusmar-moratus), множество червей и моллюсков. Примечательно, что биоценоз Саргассова моря представляет собой, в сущности, литоральное сообщество, размещенное в пелагической зоне.
Индо-Пацифическая область.
Для пелагической фауны этой области свойствен вид кита индийский полосатик Balaenopteraindica. Вместе с тем здесь есть и другие более широко распространенные китообразные. Среди рыб обращает на себя внимание парусник Istiophorusplatypterus, отличающийся громадным спинным плавником и способностью развивать скорость до 100—130 км/ч; обитает также его родственник меч-рыба (Xiphiasgladius) с мечевидной верхней челюстью, которая встречается и в тропических водах Атлантики.
Данный регион объединяет холодные и умеренно холодные воды Северного полушария. На Крайнем Севере большая часть их зимой покрывается льдами и даже летом всюду видны отдельные льдины. Соленость из-за огромных масс пресной воды, приносимой реками, относительно низка. Животный мир беден и однообразен. Южнее, примерно до 40° с. ш., простирается полоса вод, где их температура сильно колеблется и животный мир сравнительно богаче. Основной район добычи промысловых рыб располагается именно здесь.
Акваторию региона можно разделить на 2 области — Арктическую и Эвбореальную.
Арктическая область.
Фауна пелагиали данной области бедна, но весьма выразительна. В ней выделяются китообразные: гренландский кит (Balaenamysticetus), финвал (Balaenopteraphysalus) и дельфин-единорог, или нарвал (Monodonmonocerus). Рыбы представлены полярной акулой (Somniosusmicrocephalus), мойвой (Mallotusvillosus), которой питаются чайки, треска и даже киты, и несколькими формами восточной сельди (Clupeapallasi). Мол-люски-клионы и рачки-каланусы, размножающиеся громадными массами, составляют обычную пищу беззубых китов (рис. 137).

Рис. 137. Фауна холодных морей (из А.Г.Воронова, 1963):
1 - северный клион; 2 - хохлач; 3 —
топорик; 4 -
сивуч; 5 - нерпа; 6 -
котик-7 - лимацина; 8 -
нарвал; 9 -
белуха; 10 -
кайра; 11 -
гренландский киг 12 -
большой полосатик; 13
- чистик; 14 -
морж; 75 - палтус; 16 -
липарис-17 -
четырехрогий бычок; 18 -
треска; 19 -
гага; 20
~
ликод сетчатый- 21 —
живородящая бельдюга
Эвбореальная область.
Пелагиаль области охватывает северные части Атлантики и Тихого океана к югу от Арктической области и к северу от тропиков. Колебания температуры в водах этой области весьма значительны, что отличает их от арктических и тропических вод. В видовом составе фауны бореальных частей Атлантического и Тихого океанов наблюдаются различия, но число общих видов велико (амфибореальность).
Фауна пелагиали Атлантики включает несколько видов китов (бискайский, горбач, бутылконос) и дельфинов (гринда и афа-лина). Из пелагических рыб обычны атлантическая сельдь Clupeaharengus, скумбрия, или макрель, тунец Thynnusthunnus, нередкий и в других частях Мирового океана, меч-рыба, треска, пикша, морской окунь, шпрот, а на юге — сардина и анчоус.
Здесь же встречается гигантская акула Cetorhinusmaximus, питающаяся планктоном, подобно усатым китам. Из позвоночных пе-лагиали отметим медуз — сердцевидку и корнерота.
В пелагиали бореальной части Тихого океана кроме амфиборе-альных видов обитают киты — японский и серый, а также многие рыбы — дальневосточная сельдь Clupeapallasi, сардины (дальневосточный Sardinopssagax и калифорнийский S. s. coerulea виды), обычны японская скумбрия (Scomberjaponicus) и королевские макрели (Scomberomorus), из дальневосточных лососей — кета, горбуша, чавыча, нерка. Среди беспозвоночных широко распространены медузы Chrysaora и Суапеа, сифонофоры, сальпы.
К югу от Тропического региона расположен пояс Мирового океана, который выделяется в Антибореальный регион. Подобно своему аналогу на севере, он также характеризуется суровыми условиями среды.
Пелагическая зона этого региона населена единой фауной, так как между водами океанов здесь нет преград. Китообразные представлены южным (Eubalaenaaustralis) и карликовым (Сарегеа marginata) китами, горбачом (Megapteranovaeangliae), кашалотом (Physetercatodon) и полосатиками, которые, как и многие другие киты, широко мигрируют по всем океанам. Среди рыб необходимо назвать биполярных — анчоуса, сардину особого подвида (Sardinopssagaxneopilchardus), а также присущих только анти-бореальной фауне нототений — Nototheniarossi, N. squamifrons, N. larseni, имеющих важное промысловое значение.
Как и в литоральной зоне, здесь можно выделить Антибореаль-ную и Антарктическую области, но рассматривать их мы не будем, поскольку фаунистические отличия между ними невелики.
Среди схем фаунистического расчленения континентальных водоемов наибольшего внимания заслуживают две. Одна их них была предложена Л. С. Бергом (1934, 1949), исследовавшим географическое распространение пресноводных рыб, другая — Я. И. Старобогатовым (1970, 1986), изучавшим распространение моллюсков внутриматериковых вод. При некоторых различиях в оценке ранга соответствующих выделов (подобластей или провинций) схемы, разработанные Бергом и Старобогатовым, во многом сходны и выгодно отличаются от других схем фаунистического расчленения континентальных водоемов тем, что в них учитывается фауна гигантских озер и выделяются соответствующие самостоятельные области.
Согласно схеме Старобогатова, континентальные воды земного шара группируются в 9 областей: Палеарктическую, Понто-Каспийскую, Байкальскую, Сино-Индийскую, Эфиопскую, Танганьикскую, Неарктическую, Неотропическую, Австралийскую (рис.138).
Рис. 138. Зоогеографические области Земли (фауна континентальных водоемов):
1 —
Палеарктическая область, подобласти: 1а —
Европейско-Сибирская; 16 —
Охридская; 1в —
Нагорно-Азиатская; 7г — Восточно-Сибирская; 2 — Понто-Каспийская солоноватоводная область; 3 —
Байкальская область; 4 —
Сино-Индийская область, подобласти: 4а —
Амуро-Японская; 46 —
Китайская; 4в —
Индо-Малайская; 5— Эфиопская область; 6—
Танганьикская область; 7— Неарктическая область, подобласти: 7а —
Тихоокеанская; 76—
Атлантическая; 8 —
Неотропическая область, подобласти: 8а —
Центрально-Американская; 86 —
Южно-Американская; 9 —
Австралийская область (по Я. И. Старобогатову, 1969)
Палеарктическая область.
Эта область занимает всю Европу, запад Северной Африки, Переднюю Азию (кроме юго-востока), Центральную Азию и весь север Азии, за исключением бассейна Амура и рек, находящихся южнее хребта Джугджур. В состав области не входят Байкал и Каспий, а также лиманы и приустьевые части крупных рек, впадающих в Черное и Азовское моря. В области обитают 2 эндемичных рода моллюсков — Bythinella и Marstoniopsis из семейства Bythyniidae, много эндемичных родов из семейств Pyrgulidae и Lithoglyphidae, очень характерны представители семейства Melanopsidae, а из легочных моллюсков — семейства Acroloxidae. Что касается двустворчатых моллюсков — наяд, то большинство из них встречается лишь на юге области.
Для фауны области характерен ряд эндемичных родов рыб, таких, например, как карась, горчак, ерш и др.
В связи с неоднородностью фауны Палеарктической области в ней выделяются 4 подобласти: Европейско-Сибирская, Охридская, Нагорно-Азиатская и Восточно-Сибирская. В последнее же время ее предлагается делить на 3 подобласти — Европейско-Централь-ноазиатскую, Охридскую и Сибирскую.
Понто-Каспийская солоноватоводная область.
Данная область включает Каспий (кроме залива Кара-Богаз-Гол), лиманы и приустьевые части рек, впадающих в Черное и Азовское моря. Для области характерны такие эндемичные подсемейства двустворчатых моллюсков, как Didacninae и Hypaninae, много эндемиков более низкого ранга (из дрейссен — подрод Pontodreissena, из пиргулид — роды Tarricaspia, Caspia, Caspiohydrobia). Благодаря солоноватой воде и связи в прошлом с морем в области обитают некоторые морские моллюски, отличающиеся своей эвригалин-ностью (например, Mytilasterlineatus, Abraovata) и занесенные человеком.
Следует отметить, что в четвертичном периоде Черное и Каспийское моря неоднократно соединялись, образуя единый бассейн. Это способствовало обмену их фаун и развитию общих групп животных.
Байкальская область.
Область включает озеро Байкал и верхний участок реки Ангары. Фауна Байкала богата и крайне своеобразна, что говорит о ее длительной и сложной истории. Только здесь мы встречаем представителей 2 эндемичных семейств рыб — голомянковых (Comephoridae) с 2 видами и широколобкоподобных, или байкальских, бычков (Cottocomephoridae) с 24 видами. Первые живут на глубине 100—250 м, вторые заселяют все озеро — от мелководий до больших глубин.
Много в области эндемичных ракообразных (бокоплавов), ресничных червей (трикладид), а также моллюсков. Среди последних выделяются эндемичные семейства Baicaliidae и Benedictiidae, aтакже множество эндемичных родов и видов.
Сино-Индийская область.
Область занимает водоемы Южной и Восточной Азии, а также островов Зондского архипелага. В нее не входят только высокогорные верховья крупных рек Инда, Ганга, Брахмапутры, Меконга, Янцзы, Хуанхэ. Фаунистически она одна из самых богатых областей, причем фауна (в частности, моллюски) имеет тропический облик. Здесь очень разнообразны двустворчатые моллюски. Среди них масса эндемиков. Из семейства Unionidae встречается 5 подсемейств из 6, причем 2 являются эндемиками. Особый облик фауне придают пресноводные и эстуарные (приустьевые) митилиды, которых нет в других областях, за исключением Австралийской.
Своеобразны в данной области пресноводные рыбы. Из древних аравоновых рыб, распространенных в тропиках, обитают скле-ропагесы, инкубирующие свою икру во рту. В реках Индии, Бирмы, в горных ручьях Индонезии обычны разнообразные представители семейства плоскоперых. Необычны стеклянные сомы из рода Cryptopterus. К примеру, тело индийского стеклянного сома прозрачно и в отраженном свете переливается, как стенка мыльного пузыря. Замечательны своим поведением рыбы-брызгуны, живущие не только в пресной, но и в соленой воде. Они охотятся за летающими насекомыми, выбрызгивая капли воды изо рта и сбивая мух и бабочек с надводных растений или прямо в воздухе.
Сино-Индийская область подразделяется на 4 подобласти: Амурскую, Японскую, Китайскую, Индо-Малайскую.
Эфиопская область.
Данная область объединяет водоемы Африки (южнее Сахары), за исключением озера Танганьика, Мадагаскара и крайнего юго-запада Аравийского полуострова. Пресноводная фауна области целиком тропическая и несколько напоминает фауну южных частей Сино-Индийской области. Здесь среди моллюсков встречается эндемичное семейство двустворчатых — Etheriidae, множество эндемичных родов — Saulea, Afropomus из пилид, Soapitia и других из битиний, Lentorbis, Acutordia из пла-норбид и др.
Из рыб в области много цихловых — только в Малави более 200 видов из 20 родов, причем 180 из них эндемичны. В озере Виктория 170 эндемичных видов цихловых. Среди африканских цихловых есть много видов, широко известных любителям аквариумных рыб, например цихласомы, а тилапий разводят теперь в ряде стран ради вкусного мяса. Замечателен у мозамбикской тилапий способ охраны потомства: самка вынашивает его во рту и выпускает лишь подросших рыбок.
Из двоякодышащих рыб в Эфиопской области обитают 4 вида чешуйчатников — протоптеров, которых иногда выделяют в особое эндемичное семейство Protepteridae. Один из видов — бурый протоптер — живет во временных водоемах. В дождливый сезон он активно питается, растет, размножается, а в сухой — вырывает ртом гнездо сначала в илистом, затем в глинистом дне, складывается пополам, окружается коконом из затвердевшей слизи и впадает в спячку. Как только водоем высыхает, протоптер начинает дышать атмосферным воздухом, который поступает в камеру через мелкие трещины и отверстия в пробке, закрывающей камеру.
С наступлением очередного сезона дождей протоптерусы возвращаются к активной жизни.
Разнообразны в области и сомовые, включающие несколько семейств — клариевых, перистоусых, электрических. Обитающий в Ниле электрический сом способен вырабатывать импульсы тока, достигающие 360 В.
Танганьикская область.
В состав области входит только одно озеро Танганьика глубиной до 1470 м (второе по глубине после Байкала), расположенное в экваториальной Африке. Оно отличается богатой ихтиофауной. Среди рыб в области многочисленны эндемичные цихлиды — 40 родов и свыше 140 видов встречаются только здесь.
Моллюски, живущие в озере, представлены эндемичными семействами Syrnolopsidae, эндемичным подсемейством Tiphobiinaeиз палюдомид, многочисленными эндемичными родами и видами. Богатство фауны и высокий процент эндемизма отличают Танганьику от других озер Африки и объясняются древностью озера. По мнению многих ученых, его фауна развивалась без перерывов с начала третичного периода.
Неарктическая область.
Эта область занимает Северную Америку от арктического побережья на севере до 26—27-й параллели на юге. Фауна области по ряду признаков напоминает палеарктическую. Из моллюсков здесь обычны вивипариды, среди которых есть как широко распространенные (Viviparus), так и эндемичные роды (из подсемейства Campelominae). Очень много в области битиний, но большинство из них относятся к эндемичным подсемействам Nymphophilinae и Fonligeninae. Обитает также эн-демичное семейство Lepyriidae. Легочных моллюсков в Неарктической области много, причем планорбиды представлены эндемичным подсемейством Planorbulinae. Перловицы и беззубки (из двустворчатых моллюсков) также многочисленны.
Среди рыб прежде всего нужно отметить эндемичное семейство центрарховых (из окуневидных), состоящее из 12 родов, включающих 30 видов. Эндемичные семейства перкопсовые (лососе-окуни) и афредодеровые (окуни-пираты) содержат всего 3 вида. Щуковые и умбровые, обитающие также в Палеарктике, в Неарктической области представлены эндемичными видами.
Область отчетливо делится на две подобласти — Тихоокеанскую и Атлантическую. Фауна второй богата и разнообразна.
Неотропическая область.
Область охватывает водоемы Южной и Центральной Америки, а также Антильских островов. Отличительными чертами ее фауны является отсутствие среди моллюсков таких семейств, как вальватиды и вивипариды. Зато богато представлены здесь литторидиниды (15 эндемичных родов). Очень разнообразна и многочисленна группа двустворчатых моллюсков — наяд, насчитывающая среди прочих и 2 эндемичных семейства — Mycetopodidae и Mulleriidae. Много эндемичных родов из более широко распространенных семейств. Обращает на себя внимание тот факт, что весь состав моллюсков носит четко выраженный тропических характер.
Пресноводные рыбы области весьма разнообразны. Среди них следует отметить эндемичное семейство речных скатов-хвостоколов, 2 рода которых живут в бассейне Амазонки и других реках, впадающих в Атлантический океан. Есть также эндемичный представитель двоякодышащих рыб — лепидосирен, сходный по экологии со своим африканским сородичем протоптером. Из тропико-политных очень древних аравановых рыб в области живут эндемичные роды аравана и арапаима. Последняя относится к числу самых крупных пресноводных рыб в мире — до 3 м в длину при массе 150—180 кг. Наибольшей численности и разнообразия в Неотропической области достигают харациновые рыбы (5 семейств, 100 родов и более чем 800 видов). К ним относятся знаменитая пиранья и множество красивых ярких рыбок, которых охотно содержат в аквариумах: неоны, лебиасы, клинобрюшки и др. Пираньи являются прожорливыми хищниками. Они охотятся за всем, что движется в воде, нападают стаями, в том числе и на купающихся людей, особенно если на теле у них есть кровоточащие ранки.
Неглубокие реки северо-восточной части области населяют широко известные электрические угри, относящиеся к эндемич-ному семейству Electrophoridae. Эта крупная (до 1,5 м) рыба имеет в ротовой полости особые участки сосудистой ткани, которая позволяет ей усваивать кислород непосредственно из атмосферного воздуха. Для захвата новой порции воздуха угорь поднимается к поверхности воды не реже одного раза в 15 минут, а то и чаще. Наиболее интересная особенность электрических угрей — громадные электрические органы, занимающие около 4/5 длины тела. Напряжение разряда у рыб метровой длины обычно достигает 350 В. Указанные органы используются угрем для защиты от врагов и парализации добычи — небольших рыб.
Половину пресноводных рыб Неотропической области составляют различные сомы. Их около 1000 видов из 12 эндемичных семейств. Однако карповые рыбы в области отсутствуют.
Неотропическая область делится на две подобласти — Центрально-Американскую и Южно-Американскую.
Австралийская область.
Область включает водоемы Австралии, Тасмании, Новой Гвинеи с прилегающими к ней островами, а также водоемы Новой Зеландии, Новой Каледонии и островов Фиджи. Пресноводные моллюски представлены здесь эндемичными семействами брюхоногих Tateidae и Batiidae. Из двустворчатых — наяд — обитают только виды семейства Hyriidae, в частности, 2 эндемичных подсемейства. Среди других, неэндемичных семейств в области много эндемичных родов.
Пресноводные рыбы Австралийской области весьма оригинальны. В первую очередь следует отметить рогозуба — двоякодыша-щую рыбу эндемичного семейства рогозубовых, или однолегочных (Ceratodidae). Рогозуб (Neoceratodusforsteri) — единственный современный представитель семейства. Встречается он лишь в Северо-Восточной Австралии, где населяет бассейны рек Бернетг и Мэри. Это крупная рыба, достигающая в длину 175 см, массой выше 10 кг. Как и все рыбы, она дышит жабрами, но, кроме того, каждые 40—50 минут поднимается к поверхности, чтобы захватить атмосферный воздух. Вдох и выдох рогозуб производит через ноздри. Легочное дыхание для него обязательно, особенно когда в перегретой стоячей воде развиваются гнилостные процессы и исчезает весь кислород. Рогозуба употребляют в пищу, мясо его очень ценится.
Среди круглоротых и рыб Австралийской области есть эндемичные представители миног, сомов, бычков и атеринок.
С зоогеографической стороны интересно наличие в области со-левыносливых рыб семейства галаксиевых. Они распространены только в Южном полушарии. Всего их насчитывают 46—48 видов, из которых более 30 населяют водоемы Австралийской области. Большинство галаксиевых — мелкие рыбы, имеющие длину 10— 15 см, только новозеландская гигантская галаксия достигает 30 см. Для объяснения замечательного характера распространения галаксиевых предлагались разные гипотезы. Присутствие их в пресных водоемах широко разобщенных океанами материков Южного полушария — Австралии, Новой Зеландии, Южной Америки, Южной Африки — можно объяснить наличием в прошлом соединения этих материков. Но некоторые зоогеографы считают, что солевы-носливые рыбы могли быть разнесены морскими течениями из района своего происхождения. Вопрос этот остается дискуссионным.
Особой чертой фауны водоемов Австралийской области следует назвать отсутствие карповых рыб.
Районирование суши имеет долгую историю. В 1858 г. П. Склэ-тер предложил шесть основных фаунистических областей: Палеарктическую, Эфиопскую, Индийскую, Австралийскую, Неарктическую и Неотропическую. В 1876 г. А. Уоллес, приняв фаунистическое районирование Склэтера, стал называть Индийскую область Восточной. Склэтеровская классификация областей многими признается и сейчас. Однако она неоднократно подвергалась корректировке.
Были попытки создать единицы более высокого ранга, отражающие исторические связи фаун. Так, в 1890 г. А-Бленфорд объединил склэтеровские области в три группы: Арктогейскую (с областями Эфиопской, Восточной, Палеарктической и Неарктической), Южно-Американскую и Австралийскую. Этим группам Р. Лидеккер присвоил ранг царств. Два последних царства были впоследствии названы соответственно Неогейским и Нотогейским. Ф.Дарлинггон в 1957 г. предложил для Арктогеи название Магагея. Такой классификацией зоогеографических областей пользуются чаще всего. При этом системы фаунистических и флористических областей земного шара во многом не совпадают, хотя общее районирование весьма сходно.
В свете новейших фаунистических данных и при использовании сведений о распространении насекомых, моллюсков и других групп беспозвоночных имелись все основания пересмотреть существовавшие системы фаунистических областей. Это и было сделано в 1976 г. О.Л.Крыжановским. Но наиболее приемлемо следующее районирование (рис.139):
Царство Палеогея
Эфиопская область
Индо-Малайская область
Мадагаскарская область
Полинезийская область
Царство Арктогея[6]
Подцарство Палеарктическое
Европейско-Сибирская область
Область Древнего Средиземья
Восточно-Азиатская область
Подцарство Неарктическое
Канадская область
Сонорская область
Царство Неогея
Неотропическая область
Карибская область
Царство Нотогея
Австралийская область
Новозеландская область
Патагонская область
Рис. 139. Фаунистические области суши:
I— царство Палеогея, области: I1
— Эфиопская; I2
— Индо-Малайская; I3
— Мадагаскарская; I4
— Полинезийская; II — царство Арктогея, области: II1
— Европейско-Сибирская; II2
—
Древнего Средиземья; II3
— Восточно-Азиатская; II4
—
Канадская; II5
— Сонорская; III— царство Неогея, области: III1
— Неотропическая; III2
— Карибская; IV —
царство Нотогея, области: IV1 —
Австралийская; IV2
— Новозеландская; IV3
—
Патагонская
Это царство включает тропики и частично субтропики Старого Света: материк Африки к югу от Сахары, крайний юг Аравии, остров Мадагаскар, юг и юго-восток Азиатского материка — полуострова Индостан и Индокитай, Зондские острова, остров Новая Гвинея и архипелаги Полинезии. Северо-западная граница его проходит по югу Сахары, юго-восточная — через архипелаги тропической зоны Тихого океана. Эти границы представляют собой переходные территории и отличаются смешанной фауной.
Область охватывает материк Африки к югу от пустыни Сахары, южную (гористую) часть Аравийского полуострова и островов Сокотру в Индийском океане.
Природные условия области определяются рельефом и географическим положением Африки, большая часть которой находится в пределах тропической зоны. По характеру поверхности Африка представляет собой гигантское плоскогорье. На востоке над ним возвышаются Эфиопское нагорье и цепь вулканических гор Центральной Африки, на крайнем юге — Драконовы горы, а на западе — высоты Камеруна и Гвинеи. Эти горы, расположенные на периферии материка, служат преградой для влажных морских ветров, так что в центральных районах Африки господствует засушливый тропический климат. Крайний юг отличается умеренно теплым климатом средиземноморского типа,
Большая часть Эфиопской области занята саваннами различного типа — от редколесий и парковых лесов до высокотравных и опустыненных саванн. Саванны развиваются в условиях периодического увлажнения при высоких круглогодовых температурах. В сухой сезон влаги здесь испаряется больше, чем поступает, во влажный — наоборот. Древесные породы в сухой сезон теряют листву, они устойчивы к пожарам. Безлесные пространства саванн покрыты зарослями злаков, в основном так называемой слоновой травой. Из деревьев наиболее распространены баобаб, акации с зонтиковидными кронами, масличная пальма и древовидные молочаи.
Влажные экваториальные, или дождевые, леса занимают в Африке западную и центральную части экваториальной области в бассейне Конго и на побережье Гвинейского залива. В горах произрастают дождевые леса, требующие высокой стабильной температуры и хорошего увлажнения в течение всего года (не менее 2000 м), по долинам рек располагаются галерейные леса. Растительный мир экваториальных лесов чрезвычайно богат и разнообразен. На площади в 1 га можно насчитывать до 50 видов деревьев, а в среднем примерно 600 видов. Общее богатство флоры цветковых африканских дождевых лесов оценивается более чем в 11 тыс. видов.
Фауна Эфиопской области (рис. 140, 141) также очень богата и разнообразна. Это прежде всего касается тропических, особенно лесных, районов.
Из млекопитающих здесь обитают эндемичные отряды даманов (Hyracoidae) и трубкозубов с единственным семейством Orycteropodidae, представленным одним видом — африканским трубкозубом (Orycteropusafer), питающимся термитами и муравьями. Даманы включают как лесных, так и горных животных размером с зайца. Это растительноядные звери с оригинальными присосками на подошвах, позволяющими им карабкаться по стволам деревьев и отвесным скалам. Встречаются также эндемичные семейства выдровых землероек, златокротов, беге мотов, жирафов, иглохвостых белок, долгоногов и некоторых других.

Рис. 140. Характерные представители фауны Эфиопской области
(млекопитающие):
1— жираф; 2 —
карликовый бегемот; 3 —
кистеухая свинья; 4 —
даман; 5 — трубкозуб; 6 —
пятнистая гиена; 7 — антилопа куду; 8 —
лев; 9 —
слоновая землеройка; 10 —
окапи; 11 —
зебра; 12 —
мартышка; 13 —
африканский слон; 14 —
черный колобус; 75 — шимпанзе; 16 —
горилла

Рис. 141. Характерные представители фауны Эфиопской области
(от птиц до насекомых):
1 —
двупалый страус; 2 —
птица-секретарь; 3 —
турако; 4 —
нектарница; 5 — рогатый ворон; 6 —
китоглав; 7 — яичная змея; 8 —
зеленая мамба; 9 —
красно-клювый ткачик; 10 —
поясная ящерица; 11 —
многопер; 12 —
веслоногая лягушка;
13 —
жук-голиаф; 14 —
сколопендра
Для этой области характерно обилие копытных. Жвачные включают примерно 40 родов антилоп — от мелких дукеров (голубой дукер высотой 30 см) до крупных: канн, куду и гну размером с лошадь. Большинство из них насчитывает много видов. Быки представлены кафрским буйволом, распространенным по всему материку, за исключением пустынь севера. Оленьки и жирафы — еще два типичных семейства африканских жвачных. Оленьки — самые мелкие из копытных (размером с зайца). Самцы у них не имеют рогов, но зато обладают развитыми верхними клыками. Обитают оленьки в дождевых лесах. Жирафы являются эндемичным семейством, насчитывающим всего 2 вида, относящихся к 2 родам: окапи (Okapiajonstoni) высотой до 2 м, живущий в густых лесах Конго, и жираф (Giraffacameleopardais) до 6 м в высоту, обитающий в саваннах и редколесьях.
Из нежвачных в Африке обитают бегемоты (гиппопотамы) и свиньи. Бегемоты (Hippopotamidae) представлены 2 видами: огромным бегемотом (Hippopotamusamphibius), самцы которого достигают массы 3 т при длине тела 400—420 см, и карликовым бегемотом (Choeropsisliberiensis) длиной всего около 150 см и массой 250—260 кг. Близкие к бегемотам свиньи в Эфиопской области обычны. Это бородавочники (Phacochoerus) с большой плоской мордой, покрытой выростами в виде бородавок, и громадными изогнутыми клыками, а также кистеухие свиньи (Potamo-schoerus) — ярко-рыжие животные с кисточками белых волос на ушах. Настоящих свиней (род Sus) в Африке нет.
Из непарнокопытных в Эфиопской области обитают 2 вида носорогов, которые отличаются отсутствием передних зубов и наличием двух рогов, а также зебры, многочисленные стада которых еще и сегодня бродят в саванне. Всего в Африке 3 вида зебр.
Из отряда хоботных встречается только африканский слон (в отличие от индийского бивни у него имеют и самец, и самка). Стада живут в саванне и лесах, нередко поднимаются в горы.
Хищники данной области не менее разнообразны и характерны, особенно виверровые (Viverridae). Их около 10 родов. Гиены распространены всюду и выполняют роль санитаров, поедая трупы. Кроме гиен (3 вида) здесь обитает их родственник гривистый волк (Protelescristatus). Впрочем, он близок и к виверрам. Некоторые же систематики выделяют его в отдельное семейство. Волков в Африке нет. Их заменяют стаи эндемичных для области гиеновых собак (Lycaonpictus). Лисицы встречаются повсеместно. Кошачьих мало, и они в своем распространении, как правило, выходят за пределы Африки. Это лев, живущий в саванне или даже в полупустыне (во многих районах уже истреблен), леопард, гепард, сервал и мелкие виды кошек. Африканская дикая кошка была одним из прародителей домашних пород.
Среди грызунов в этой области выделяются дикобразы (Hist-ricidae) (распространены в Азии) и представители следующих эндемичных семейств: шипохвостые белки (Anomaluridae), долгоноги (Pedetidae), тростниковые крысы (Thryonomyidae), горные мыши (Petromyidae), гребнепалые крысы (Ctenodactylidae) и пескорои (Bathyergidae). Последние ведут подземный образ жизни.
Отряд насекомоядных представлен здесь 3 эндемичными семействами. Особого внимания заслуживает семейство златокротов (Chrysochloridae), получивших свое название за металлический отлив меха. В семействе около 30 видов. Оно распространено главным образом в Южной Африке вплоть до Уганды. Сходство златокротов с настоящими кротами чисто конвергентное. Живут златокроты в песчаных местах, где роют поверхностные ходы. Эндемичны также выдровые землеройки (Potamogalidae), обитающие в Западной Африке у воды. Они хорошо плавают и ныряют в поисках рыбы и ракообразных. Последние из эндемичных семейств — слоновые землеройки, или хоботные прыгунчики (Масго-scelididae). Они насчитывают до 40 видов, похожи на тушканчиков, но отличаются вытянутой в хоботок мордочкой, питаются насекомыми.
Полуобезьяны в Эфиопской области относятся к семейству лори (Lorisidae). Встречаются они и в Индо-Малайской области. Замечательно, что африканские полуобезьяны весьма далеки от мадагаскарских лемуров и близки к индийским. Всего здесь 2 эндемичных рода — потто и галаго, включающих незначительное количество видов. Обезьяны же в этой области многочисленны и разнообразны. Все они относятся к 2 семействам — мартышкооб-разные (Cercopithecidae) и понгиды (Pongidae). Последние представлены родами горилла и шимпанзе, в каждом из них по 1 виду с подвидами. Гориллы и шимпанзе — лесные обезьяны, причем первая ведет семейный, а вторая — стадный образ жизни. Из мар-тышкообразных в Африке обитают мартышки (около 100 видов), павианы, мандрилы и колобусы. Образ жизни у них весьма различный. Мартышки живут в лесах большими стаями и чрезвычайно подвижны. Павианы обитают в горах и лесах. Они также ведут стадный образ жизни. В стаде обычно существует строгая иерархия, и возглавляет его сильный старый самец.
В целом фауна млекопитающих Эфиопской области представляет собой смесь эндемичных семейств, семейств, общих с Индо-Малайской областью, с которой она тесно связана, и более или менее широко распространенных семейств. Однако следует отметить, что здесь нет семейств настоящих кротов, медведей, енотов, бобров, оленей и верблюдов. Одногорбый верблюд — домашнее животное в Северной Африке. В диком состоянии он, возможно, встречался в доисторические времена.
В отличие от млекопитающих птицы Эфиопской области, хотя и чрезвычайно разнообразны и многочисленны, но относятся в основном к широко распространенным семействам. Эндемичных родов и видов много, но ареалы их ограниченны, и для характеристики области в целом они малопригодны. К чисто африканским нужно отнести страусов (Struthionidae), представленных 1 видом — двупалым страусом; птиц-секретарей (Sagittariidae) — тоже 1 вид; турако (Musophagidae), близких к кукушкам; птиц-мышей (Coliidae), оригинальных китоглавов (Balaenicipitidae), мо-лотоглавов (Scopidae). Более широко распространены цесарки (свыше 20 видов), ткачики, нектарницы, медоуказчики, крупные птицы-носороги. Чаще всего ареал этих групп включает Африку и Индию. Немало здесь и тропикополитных птиц — попугаев, трогонов, бородаток, питт, а также космополитных — кукушек, дятлов, сизоворонок, зимородков и др. В зимнее время орнитофауна Африки обогащается за счет многочисленных зимующих птиц, прилетающих из Европы, например аистов, журавлей, ласточек и т. д.
Рептилии Эфиопской области весьма многочисленны, но эндемиков высокого ранга мало. Из отряда чешуйчатых чрезвычайно характерны хамелеоны. Однако они встречаются также на Мадагаскаре (здесь обитает самый крупный вид), в Южной Испании, в Аравии, Индии, на Цейлоне (Шри-Ланка).
Много в этой области ящериц, гекконов, эндемичных поясо-хвостов (Cordylidae) и безногих червеобразных ящериц (слепые сцинки Feylinidae), агам и громадных варанов (нильский достигает 2 м длины), а также змей. В Эфиопской области обитает крупный вид тропикополитного семейства — слепозмейки Typhlopshumbo (длина до 80 см) и ряд более мелких видов. Питоны (Pythoninae), живущие в тропиках Старого Света и Австралии, представлены несколькими видами рода Python, начиная с мелких (длина 1—1,5м) — королевского и карликового и кончая крупным иероглифовым питоном, достигающим 7—9 м. В лесах Западной Африки живет род питонов калабария. Единственный вид его Calabariareinhardti приспособлен к роющему образу жизни.
Своеобразны яичные змеи (Dasypeltidae) — 2 рода их встречаются в Африке, 1 — в Индии. Эти змеи питаются птичьими яйцами, причем скорлупу они разрушают так называемой яичной пилой — отростками передних позвонков, прободающих сверху стенку пищевода. В Африке многочисленны и ядовитые аспидовые змеи (более 20 видов из 10 родов). К ним относятся настоящие кобры (черношейная и ошейниковая — так называемые плюющие кобры), водяные крабы, мамбы и пестрые аспиды Elaps. Настоящие гадюки (Viperinae), обитающие только в Старом Свете, имеют в Эфиопской области центр распространения, но отсутствуют на Мадагаскаре. Для Африки характерны виды родов Causus (жабьи гадюки), Atractaspis (земляные гадюки), Bitis (африканские гадюки), среди которых ярко окрашенная габонская гадюка достигает 2 м. Типичны здесь и эфы, а также рогатые (Cerastes) и древесные (Athens) гадюки, обитающие на деревьях.
В Африке живут сухопутные черепахи (Testudinidae), относящиеся к 4 родам с несколькими видами, и трехкоготные черепахи (Trionychidae) из 2 эндемичных родов, а также черепахи пеломе-дузы (Pelomedusinae). Последние распространены в Африке, на Мадагаскаре и в Южной Америке.
Все еще многочисленны в этой области и крокодилы. Африканские крокодилы относятся к 2 родам и ведут преимущественно водный образ жизни, изредка заходя и в соленые воды.
Из амфибий, обитающих в Эфиопской области, следует назвать эндемичных Phrynomeridae (род шпорцевых лягушек Xenopusиз липовых), своеобразных жаб (например, лазающая ночная жаба Nectophryne с липкими дисками на пальцах и живородящая жаба Nectophrynoides), многочисленных настоящих лягушек, рако-форид, или веслоногих лягушек, встречающихся также в Азии, узкоротов (Breviceps) и крайне необычных волосатых лягушек (Astylosternus), у самцов которых в брачный период по бокам и на бедрах появляются длинные волосовидные выросты кожи. Хвостатых амфибий здесь нет, их заменяют червяги, характерные для тропиков всего мира, кроме Австралии.
Удивительны пресноводные рыбы Эфиопской области. Это представители древних групп — кистеперые (Polypteridae), эндемич-ное семейство с родами многопер и каламоихт. Своеобразие этих рыб так велико, что специалисты относят их к особому отряду или даже к подклассу. Живут в водоемах Африки и двоякодыша-шие рыбы рода Protopterus. В засушливый период, когда вода во временных водоемах пересыхает, протоптеры, зарываясь в ил, впадают в спячку, которая продолжается 6—9 месяцев. Протоптеры относятся к семейству чешуйчатниковых (Lepidosirenide), один род которого Lepidosiren живет в пресных водах Южной Америки.
Немало в Эфиопской области и эндемичных семейств рыб, принадлежащих к примитивному подотряду Isospondyli (кнериевые, кромериевые, фрактолемовые и др.). Из костноязыких рыб в Африке живут рыбы-бабочки (Pantodontidae), узкокрылые (Mormy-ridae), гимнарховые (Gymnarchidae). Гимнархи могут испускать электрические импульсы. Такой же способностью обладают электрические сомы (Malapteruridae), единственный представитель которых живет в Ниле и в водоемах Конго. Сомы вообще чрезвычайно обильно представлены в Африке, их не менее 6 семейств и около 250 видов. Многочисленные семейства пресноводных рыб Эфиопской области указывают на ее фаунистические связи с Южной Америкой (Characidae, Cichlidae) или с Индо-Малайской областью (Anabantidae — лабиринтовые). Как и везде в тропиках, в области нет лососевых и щуковых.
Беспозвоночные Эфиопской области настолько разнообразны, что охарактеризовать их можно только в общих чертах. Много здесь скорпионов. Они относятся к древнему подсемейству Isometrinae(представитель его встречается и в Южной Америке) семейства Scorpionidae, преимущественно африканскому. Один из скорпионов пандинус-император достигает длины 16 см. По всей Африке распространены сольпуги (на Мадагаскаре их нет). Разнообразны пауки: крупные ядовитые птицеяды (Theraphosa), различные кру-гопряды, плетущие огромные тенета в лесах, и пр. Встречаются и громадные многоножки — кивсяки, употребляемые местным населением в пищу. Большой численности достигают ядовитые сколопендры. Иногда они весьма ярко окрашены или имеют измененные в виде пластинок задние конечности — своеобразный звуковой орган.
Из насекомых обращают на себя внимание термиты. В дождевых лесах Африки их не менее 500 видов. Гнезда их напоминают большой гриб. Термиты — главные разрушители растительных остатков. Если бы они вдруг исчезли, деревья быстро бы исчерпали запасы минеральных веществ. В результате накопилось бы много неразложившегося листового опада и лес прекратил бы свое существование. В саваннах термиты не менее многочисленны, но их гнезда сделаны из глины в виде высоких конусов (выше 2 м). С этими гнездами связана жизнь целого ряда животных, использующих термитники как убежища или поедающих их хозяев (трубкозуб).
Исключительным богатством отличается фауна саранчовых. Среди них есть стадные виды (шистоцерка, расселяющаяся на тысячи километров и съедающая на своем пути всю растительность). Из жуков в аридных районах Африки преобладают разнообразные чернотелки (Tenebrionidae), в лесах — громадные жуки-голиафы длиной 10—11 см из подсемейства бронзовок, жуки-олени, усачи и др. Распространена здесь и широко известная муха цеце — переносчик возбудителя сонной болезни. Дневных бабочек в области сравнительно немного, и им не свойственна роскошная окраска, как это наблюдается у бабочек в тропиках Азии или Южной Америки.
Среди наземных моллюсков выделяются своей величиной (с крупное яблоко) растительноядные виды рода ахатина.
Эфиопская область по характеру ландшафтов, особенностям растительного покрова и, конечно, дифференциации фауны может быть разделена на 3 подобласти: Восточно-Африканскую, Западно-Африканскую и Южно-Африканскую, или Капскую.
К данной области относятся остров Мадагаскар, а также ряд архипелагов и отдельных островов Индийского океана: Коморские, Сейшельские, Маскаренские и другие более мелкие острова.
Громадный остров Мадагаскар (587 тыс. км2
) достаточно разнообразен по природным условиям. Центральные части его гористы, прибрежные — равнинны. Плоскогорья в центре острова высотой в среднем 1000—1500 м (отдельные вершины до 2600 м) лишены леса, восточные же равнины покрыты густой тропической растительностью, а южная оконечность острова имеет пустынный облик. Мозамбикский пролив (ширина 420 км) отделяет Мадагаскар от Африки, а Индийский океан — от Индо-Малайской области (расстояние 3700 км).
Флора Мадагаскарской области очень богата. В ней насчитывается около 6000 видов растений. Приблизительно 1500 из них встречаются и в Африке. Но в отличие от последней на Мадагаскаре хорошо представлены пальмы, бамбуки и орхидеи. Крайне необычно так называемое дерево путешественников — эндемик острова. В углублениях у черешков крупных листьев его удерживается дождевая вода в таком количестве, что ею можно напиться. В этих своего рода водоемах обитают даже древесные лягушки.
Фауна Мадагаскара типично островная со значительным количеством эндемиков и наряду с этим с большими дефектами в составе, что весьма заметно при сравнении ее с эфиопской материковой фауной. Животное население Мадагаскара очень своеобразно (рис. 142): среди фитофагов абсолютно преобладают плодоядные древесные формы, травоядных почти нет, вообще отсутствуют крупные виды хищников, нет также грифов — потребителей трупов. Напомним, что для эфиопской фауны характерно обратное соотношение жизненных форм.

Рис. 142. Характерные представители фауны Мадагаскарской области:
1 – тенрек; 2 – лемур вари; 3 – лемур ката; 4 – руконожка; 5 – ванга; 6 – Фосса; 7 – хамелеон; 8 – павлиноглазка аргема; 9 – гигантская черепаха
Специфика мадагаскарской фауны четко выявляется при обзоре аборигенных нелетающих зверей. Они представлены только 4 отрядами, причем все виды эндемичны.
Насекомоядные представлены 1 семейством щетинистых ежей, или тенреков (Tenrecidae), состоящее из 13 родов с 30 видами. Экологически тенреки вследствие их адаптивной радиации весьма разнообразны. Есть тенреки типа ежей, кротов, землероек, встречаются даже плавающие тенреки — аналоги выдр.
Отряд приматов на Мадагаскаре состоит из 3 семейств — настоящие лемуры (Lemuridae), индри (Indridae) и руконожковые (Daubentoniidae). Настоящих лемуров 15 видов из 6 родов, среди них следует отметить крупных вари и катга. Семейство индри включает 4 вида, а в семействе руконожковых всего 1 вид — мадагаскарская руконожка, или айе-айе — крошечная полуобезьяна с пышным хвостом. Лемуры и индри известны на Мадагаскаре и в ископаемом состоянии по крайней мере с плейстоцена.
Из отряда грызунов типично эндемичное подсемейство хомяков — мадагаскарские хомяки (Nesomyinae), представленное 12 видами.
Все хищники области — из семейства виверр (7 эндемичных родов). Самым интересным из них, безусловно, является фосса (Cryptoproctaferox) — зверь настолько загадочный, что некоторые специалисты относят его к кошачьим. Это крайне примитивное животное напоминает нижнетретичных ископаемых хищников — предков виверр и кошек.
Другие млекопитающие представляют собой переселенцев из Африки (вероятно, результат завоза или случайного заноса) и мало изменились с тех времен. Это мыши, крысы и один вид ки-стеухой свиньи. Летучих мышей в Мадагаскарской области много, но среди них нет специфичных именно для этой области. В прошлом жил и бегемот. Остатки его найдены в плейстоценовых отложениях. Следует сказать, что здесь совершенно отсутствуют слоны, носороги, обезьяны, кошки, собаки, гиены, трубкозубы, антилопы.
Птицы Мадагаскара на 50 % состоят из эндемичных видов, остальные — в основном те же, что и в Эфиопской области (молотоглавы, рябки, ткачики, нектарницы и т.д.) или в Индо-Малайской (белоглазки, стрижи-саланганы и др.). Но дефектность орнитофауны на Мадагаскаре еще более выражена. Здесь отсутствуют страусы, секретари, турако, птицы-носороги, бородатки, медоуказчики и многие другие. Типичными же обитателями являются 3 эндемичных семейства: мадагаскарские питты (Philepitidae), пастушковые куропатки (Mesoenatidae) и ванги (Vangidae), последние включают 11 видов из 9 родов — случай островной дифференциации эндемичных форм. Вымершие в плейстоцене гигантские нелетающие птицы — эпиорнисы и додо (последние жили на Маскаренских островах) — тоже были энде-миками области. Как и везде, здесь распространены виды космополитных семейств — поганок, бакланов, ястребиных (но орлов нет), цапель, аистов, кукушек, голубей, сов, зимородков и ласточек.
Фауна рептилий на Мадагаскаре довольно богата. Много хамелеонов, гекконов, черепах (сухопутных и бокошейных). Крокодилы, так же как и в Африке, представлены нильским крокодилом, причем он встречается и на Сейшельских и Коморских островах, что неудивительно, поскольку крокодилы отлично плавают и не избегают соленой воды. Настоящие ящерицы, как и ядовитые змеи, в области отсутствуют. Нет на Мадагаскаре и настоящих удавов (Constrictor), а также питонов. Из удавовых (Boidae) обитают только мадагаскарские удавы (подсемейство Sanciniinae) — эндемичная группа, включающая 4 рода, 2 из которых живут на самом Мадагаскаре, остальные — на Реюньоне и острове Раунд близ Маврикия. Сведения о присутствии в Мадагаскарской области южноамериканских родов Costrictor и Corallus ошибочны.
Земноводных мало, и все они относятся к лягушкам (около 150 видов, большинство эндемики). Это веслоногие лягушки, или ракофориды, а также узкоротые (Brevicipitidae). Последние более разнообразны.
Настоящих пресноводных рыб на Мадагаскаре нет, но встречаются несколько видов хромисов и карпозубых, а также различные солевыносливые формы.
Беспозвоночные животные Мадагаскара в большинстве своем явно эфиопского происхождения, в том числе и скорпионы, отдельные роды которых (Crosphus, Heteroscorpius) эндемичны. Сольпут на Мадагаскаре нет совсем. В энтомофауне Мадагаскара нередки и индо-малайские фаунистические связи. Многие представители имеют причудливую форму или великолепную окраску. Богомол Brancsikiaseroplana отличается необычайным листовидным расширением переднеспинки и напоминает фантастический летательный аппарат. Весьма типична здесь крупнейшая из всех павлиноглазок мира — ярко-желтая Argemamittrei (длина бабочки около 26 см). Большинство же бабочек относится к семействам и родам, распространенным на материке.
В эту область входит тропическая и субтропическая части Азии, а также ряд архипелагов и островов Индийского и Тихого океанов. Западная граница ее — широкая, проходящая по пустыне Тар полоса, где индийская фауна смешивается с переднеазиатской. Северная граница четка лишь на западе, где она идет по гребню Гималаев. На востоке же выраженность ее теряется, а на территории Китая она приобретает характер широкой полосы. Большинство зоогеографов считают, что тут граница проходит между бассейнами рек Хуанхэ и Янцзы, совпадая с северной границей субтропических лесов. Особенно спорна юго-восточная граница области. Со времен Уоллеса ее проводили по проливу, разделяющему острова Бали и Ломбок, Калимантан и Сулавеси. Позднее Гексли назвал эту границу «линией Уоллеса», утверждая, что она должна разделять Филиппины, оставляя многие острова архипелага Австралийской области. По мнению других, границу следует проводить значительно восточнее, включая в Индо-Малайскую область как Сулавеси, так и Малые Зондские и Молуккские острова. Это так называемая «линия Вебера». Она определена статистически и делит территорию на части с примерно равным соотношением азиатских и австралийских элементов фауны. Ботани-ко-географы к Палеотропическому доминиону относят также Новую Гвинею и острова и архипелаги Тихого океана, лежащие в пределах тропической зоны, кроме Полинезии. Серьезным аргументом для этого служит сходство их флористического состава, в связи с чем и зоологи начинают пересматривать свои взгляды.
Ясно, что фауна Новой Гвинеи, Молуккских и других островов (до Фиджи на востоке) носит переходный характер. Но изучение распространения сравнительно большого количества групп животных, в том числе рептилий и насекомых, дает основания для включения этого района в состав Индо-Малайской, а не Австралийской области. Оказалось, что многочисленные эндемики Папуасской подобласти по происхождению и родственным связям являются палеотропическими, а не австралийскими. С нашей точки зрения, Индо-Малайской области принадлежат Филиппины, Зондские острова и Новая Гвинея. В таком виде граница совпадает с линией, отделяющей Индо-Малазийское подцарство флористов от Австралии. В восточной части области размещаются огромные массивы экваториальных лесов, на востоке Гималаев и в Индо-Китае находятся высокие горы, покрытые богатой лесной растительностью. В Индостане они сменяются сухими и открытыми пространствами и даже пустынями. Такому разнообразию ландшафтов соответствует богатая фауна, одна из самых гармоничных на земном шаре (рис. 143, 144).
Млекопитающие Индо-Малайской области включают 46 семейств (по классификации Симпсона, 1945), 5 из них — сумчатые и однопроходные — встречаются только в Папуасской подобласти, представляя здесь австралийский элемент. Эндемичных же семейств всего 4. Остальные — общие для Эфиопской области, Голарктики и даже Неотропического царства.
Из отряда однопроходных, или яйцекладущих, на юго-востоке области встречаются новогвинейские проехидны (Proechidna) и ехидны (Tachyglossus), из сумчатых — семейства сумчатые мыши (Dasyuridae), бандикуты (Peramelidae), фалангисты (Phalangeridae) и кенгуру (Macropodidae), причем часть родов последнего семейства более характерна для Новой Гвинеи, чем для Австралии.
Из насекомоядных в области водятся ежи (подсемейство щетинистых ежей (Echinosoricidae) — эндемично), кроты, шерстокрылы (эндемичны), плодоядные летучие мыши (Pteropidae) и некоторые другие. Из отряда приматов особенно интересно эндемичное семейство тупайи (Tupaiidae) — мелкие, похожие на белку животные, близкие к лемурам. Полуобезьяны родственны африканским, а не мадагаскарским, и включают роды Loris и Nycticebus(тонкий и толстый лори). Представители семейства долгопятов водятся только на Малайском архипелаге и Филиппинах. Настоящих обезьян очень много. Это макаки, тонкотелые обезьяны и, наконец, человекообразные — гиббоны и орангутаны. Следует отметить, что ни один род обезьян тропической Азии не встречается в Африке, и наоборот.

Рис. 143. Характерные представители фауны Индо-Малайской
области (млекопитающие):
1 — проехидна; 2 —
тупайя; 3 —
шерстокрыл; 4 —
гиббон; 5 — орангутан; 6 —
полуобезьяна лори; 7 — долгопят; 8 —
тигр; 9 —
носач; 10 —
мангуст; 11 —
дымчатый леопард; 12 —
бабирусса; 13 —
тапир; 14 —
индийский слон; 15 —
малая панда; 16 —
носорог

Рис. 144. Характерные представители фауны Индо-Малайской области
(от птиц до насекомых):
1 — райская птица; 2 —
калао; 3 —
питта; 4 —
ласточка салангана; 5 — дронго; 6 —
алмазный фазан; 7— летающий дракон; 8 —
гарпия-обезьяноед; 9 —
гавиал; 10 —
казуар; 11 — комодский варан; 12 —
очковая змея; 13 —
крайт; 14 —
бабочка орнитоптера
Ящеры, или панголины (Manidae), подчеркивают афроазиат-ские фаунистические связи. Грызунов много, особенно белок, но большинство родов в Азии и Африке разные. Распространены также дикобразы, песчанки, летяги, многочисленные мыши и крысы, встречающиеся даже на мелких островах.
Богато представлены хищные. Среди них выделяются волки, и прежде всего эндемичный род Cuon (красный волк), куньи, виверровые (к ним относится и индийский мангуст — истребитель змей), кошки, из которых наиболее известен тигр. Нередки здесь леопард, гепард, а также более мелкие виды. На западе области еще сохранился лев.
Слоны представлены индийским видом, тапиры — 1 видом, обитающим на Суматре и полуострове Малакка (3 вида живут в Южной Америке), носороги — 3 видами (все они с резцами в отличие от африканских сородичей). Свиньи встречаются везде, но на юго-восточные острова, видимо, завезены человеком. Из быков характерны буйвол, бантенг, гайял и мелкие островные виды. Антилоп, в противоположность Африке, мало. Настоящие козлы и бараны — голарктические вселенцы с севера.
Фауна птиц богата и сложна по составу. Эндемизм в ней проявляется умеренно. Из «бескилевых» на юго-востоке области встречается несколько видов казуаров (Casuariidae), которые вместе с сорными курами, или мегаподами, возможно, являются выходцами из Австралийской области. Фазаны, павлины и настоящие куры достигают в Индо-Малайской области подлинного разнообразия и богатства. Авдотки (Burhinidae) и рябки (Pteroclidae) — обитатели открытых пространств, попугаи водятся в лесах. Последних, впрочем, немного. Заслуживают упоминания рыбные совы (Ketupa) с неоперенными пальцами, питающиеся рыбой. Нередки в области птицы-носороги (Bucerotidae), достигающие крупных размеров: длина индийского калао, например, 120 см, а размах крыльев больше метра. Многочисленны и разнообразны здесь бородатки и дятлы. К эндемичному семейству относятся синие птицы (Irenidae), живущие в лесах. Большинство из них хорошие певцы. Очень много ярких полностью или почти тропикополит-ных птиц (трогоны, нектарницы, зимородки, цветососы, ткачики и др.). На северо-востоке четко выявляются фаунистические связи области с Голарктикой.
Богатая фауна рептилий Индо-Малайской области представлена черепахами, ящерицами, змеями и крокодилами. Черепахи включают 2 эндемичных семейства: большеголовых черепах (Platysternidae) с несоразмерной огромной невтяжной головой, единственный вид которых живет на Индокитайском полуострове, и двукоготных (Carettochelyidae), тоже 1 вид их обитает на Новой Гвинее. Остальные распространены значительно шире. Водные трех-коготные (Trionychidae) и сухопутные (Testudinidae) насчитывают массу видов.
Изобилие здесь и ящериц, но эндемичных семейств только 2. Эти оригинальные червеобразные ящерицы Dibamiidae (3 вида), роющиеся в земле, и распространенные лишь на Калимантане безухие вараны (Lanthanotidae), которые представлены единственным видом — калимантанским безухим вараном. Особенно обильны в области агамы, гекконы, сцинки. Среди агамовых ящериц выделяются своеобразные летучие драконы (Dracovolans), широкие кожистые складки на боках тела позволяют им планировать при прыжках с деревьев на расстояние до 25—30 м. Обращают на себя внимание вараны (комодский достигает длины 3 м), ведущие наземный, древесный и полуводный образ жизни.
Из змей многочисленны слепозмейки (Typhlopidae), роющие вальковатые змеи (Aniliidae), 1 род которых — коралловая свер-ташка — населяет Южную Америку, а 2 других — цилиндрическая змея и аммохилус — живут в Индо-Малайской области, близкие к ним щитохвостые змеи (Uropeltidae), которые многими зоологами рассматриваются как подсемейство предыдущих. Из всех змей 45 видов эндемичны для области. Разнообразны здесь и питоны. Крупный сетчатый питон (до 10м) — одна из двух гигантских змей мира — великолепно плавает и лазает по деревьям. Немало и ядовитых змей из семейства ужовых (Colubridae), аспид овых (Е1а-pidae), к которым относятся королевская и настоящая кобра, а также чрезвычайно ядовитые крайты (Bungarus) из семейства га-дюковых (Viperidae) и род Trimeresurus — куфии, или копьеголо-вые змеи, из семейства ямкоголовых (Crotalidae).
Крокодилы, встречающиеся в области, относятся к настоящим крокодилам, узкорылым рыбоядным гавиалам и аллигаторам, 1 вид которых живет в низовьях Янцзы в Китае. Из амфибий в области обитают все 3 отряда: безногие, хвостатые и бесхвостые. Эндемичных семейств среди них нет. Червяги представлены рыбозмеями (род Ichtyophis). Живут они в земле по берегам рек, а отнюдь не в воде. Хвостатые амфибии имеют интересные фаунистические связи: семейство скрытожаберников распространено в Восточном Китае и в Северной Америке, а саламандры — в Европе, Азии, Северной Африке и в Северной Америке. Бесхвостые амфибии очень разнообразны и многочисленны. Это настоящие (Rana) и веслоногие (Rhacophoridae) лягушки, представленные 150 видами, замещающими квакш по образу жизни, а также узкоротые лягушки и чесночницы.
Пресноводные рыбы в Индо-Малайской области в общих чертах близки к эфиопским, но архаические группы здесь отсутствуют, 3 семейства эндемичны. Главная черта ихтиофауны — необычное развитие карповых (Cyprinidae) и отсутствие лососевых.
Индо-Малайская область имеет красочную и пышную фауну насекомых. Среди огромного количества бабочек особенно красиво семейство кавалеров (Papilionidae). К ним в первую очередь относятся громадные птицекрылы (Troides, или Ornithoptera), живущие на Зондских островах и Новой Гвинее. Окраска крыльев у самцов поразительна — бархатисто-черная с изумрудно-зеленым и золотисто-желтым, красно-черная и т. д. В Индии обитает знаменитая бабочка-каллима, или так называемый сухой лист, обладающая миметической окраской и формой. Несъедобные данаиды, собравшись роями, совершают миграции за сотни километров. В Индии и Индокитае живет гигантская павлиноглазка атлас (Attacusatlas) с прозрачными «окошками» на крыльях.
Среди жуков типичны ярко окрашенные листоеды, златки, усачи, снабженные «рогами» жуки-олени, жуки-носороги. У многих насекомых тело сверху сплющено в виде сухого или зеленого листа. Такая покровительственная форма и окраска наблюдается у палочников, богомолов и даже у жуков. Жужелицы эндемичного рода Mormolyca совершенно плоские и, несмотря на крупные размеры, прекрасно имитируют сухой лист, лежащий на земле.
Так же, как и в Африке, в этой области обитает масса термитов и муравьев. Среди последних немало видов, вступающих в сим-биотические отношения с растениями. Муравьи-портные славятся способностью «сшивать» из листьев большие гнезда.
Из других членистоногих заслуживают упоминания первично-трахейные (Peripatidae), распространенные в Южном полушарии, скорпионы из эндемичных семейств Charilinidae и др. Своеобразны телифоны, сольпуги и громадные пауки-птицееды (яванский мохнатый птицеед достигает 9 см).
Очень интересно присутствие в Индии и на Цейлоне (Шри-Ланка) гигантских дождевых червей, встречающихся, подобно первично-трахейным, на южных материках.
Индо-Малайская область по составу фауны подразделяется на Индийскую, Малайскую, Индокитайскую и Папуасскую подобласти. Типичные черты фауны этой области ярче всего выражены в Малайской подобласти, Индийская сравнительно обеднена, а две последние представляют собой переходные зоны со смешанной фауной, но с явным преобладанием индо-малайских элементов.
К Полинезийской области относят громадное количество островов Тихого океана, исключая Новую Зеландию и острова Кар-мадек. В западном направлении область простирается до островов Палау, архипелага Бисмарка и Соломоновых островов, в восточном — до островов Пасхи и Сала-и-Гомес, в северном — до Гавайских островов, а в южном — до островов Норфолк и Лорд-Хау.
Все острова Океании можно подразделить на вулканические (высокие), коралловые (низкие) и известняковые.
К первым относятся Фиджи, Самоа, острова Общества (Товарищества) и Маркизские. Они гористы, достигают высоты 1,5— 2 тыс. м, иногда и выше. На склонах гор можно увидеть потоки застывшей лавы. Берега островов обрывисты, либо имеют вид песчаных пляжей, либо низменны и их окаймляет полоска мангровых зарослей. Обычно берега вулканических островов на большом протяжении сопровождаются коралловыми рифами (береговыми или барьерными). Наветренные склоны таких островов благодаря пассатам, несущим влагу, покрыты густой тропической растительностью. Растительность подветренных склонов гораздо беднее и состоит лишь из зарослей кустарников или трав.
Коралловые острова, как говорит само название, представляют собой продукт деятельности рифообразующих кораллов. Они либо узки и вытянуты в длину, либо имеют вид кольца (атоллы) с лагуной посередине. Растительность здесь скудная — встречаются только кокосовые пальмы, панданусы и галофильные кустарники.
Известняковые острова возникают в результате поднятия дна океана, поверхность их изрезана крупными оврагами и покрыта скалами, а берега крутые. Как правило, здесь развиваются густые древесно-кустарниковые заросли.
Географически острова Океании делят на 3 большие группы: Полинезию на востоке, Меланезию на западе и Микронезию на севере.
Фауна островов Океании имеет ярко выраженный иммиграционный характер и крайне дефектна. Из млекопитающих встречаются летучие мыши и немногочисленные и большей частью мелкие грызуны. Нелетающие наземные млекопитающие достигают только Соломоновых островов. Нет здесь крокодилов и сухопутных черепах. Змеи представлены очень немногими группами (на Фиджи обитает эндемичный аспид, а на Тонга и Самоа — небольшой удав рода Ganodoia), да и те к востоку от Соломоновых островов становятся все более редкими. Квакши и лягушки достигают архипелага Бисмарка, а на Фиджи есть 2 вида эндемичных лягушек. Жабы обитают только на Гавайских островах. Местные эндемичные ящерицы нередки на Новой Каледонии, Фиджи, Тонга и др. Большинство из них относятся к гекконам и сцинкам. Лишь на Фиджи и Тонга встречается реликтовый эндемичный род игуан.
В направлении от Новой Гвинеи на восток четко прослеживается тенденция обеднения фауны. Это одно из доказательств расселения животных с Азиатского материка через водные преграды. Ширина преград и различные способности к их преодолению — основные условия успешного расселения.
Фауна млекопитающих в Полинезийской области, если не принимать во внимание мышей и крыс, связанных с человеком, ограничивается плодоядными рукокрылыми, доходящими до Самоа. Это представители индийского рода Pteropus и Emballonura, палеарктического Myotis, а также тропикополитного длиннокрыла Miniopterus.
Основное ядро фауны позвоночных составляют птицы. Около 100 родов птиц Полинезийской области подразделяются на несколько групп. Не менее 35 из них эндемичны, 40 родов общи с малайскими, остальные близки или тождественны папуасским. Доминируют различные плодоядные голуби из подсемейства Treroninae и Gourinae. На острове Самоа живет Didunculus — единственный в подсемействе Didunculinae монотипичный род. Кроме этого есть стрижи-саланганы, белоглазки, зимородки и небольшое количество других семейств. Присутствуют и представители очень широко распространенных родов — ласточек (Hirundo), сов (Asio), кукушек (Cuculus) и др. На Новой Каледонии обитает кагу (Rhinochetesjubatus), относящийся к эндемичному отряду Rhino-cheti, близкому к южноамериканским солнечным цаплям. Численность этой лесной птицы неуклонно уменьшается в связи с вырубкой лесов и неумеренной охотой.
Совершенно своеобразны птицы Гавайских островов. Это прежде всего эндемичное семейство гавайских цветочниц (Drepanididae). Считается, что все 22 вида его являются потомками американских пищух — медососов. Находясь в условиях изоляции, семейство образовало несколько экологических типов. Гавайские цветочницы — ярко окрашенные птички с разнообразными клювами. У одних он короткий и толстый, у других — длинный изогнутый, у третьих — надклювье вдвое длиннее подклювья и т. д. Некоторые виды из-за красоты оперения сильно истреблены человеком.
Для беспозвоночных островов Океании характерна та же тенденция: они богато представлены в западной части, но к востоку разнообразие их убывает. На Фиджи и в Новой Каледонии (до 80 % беспозвоночных Новой Каледонии — эндемики) есть бабочки кавалеры, они доходят и до островов Самоа. Жуки на 30—75 % эндемичны, и среди них много нелетающих форм. Моллюски Гавайских островов чаще всего принадлежат к эндемичному семейству Achatinellidae с огромным количеством видов и еще большим — географических рас, строго локализованных по отдельным горным хребтам и долинам.
Арктогейское, или Голарктическое, царство — огромная территория, охватывающая всю северную внетропическую часть земного шара, т.е. Европу, Северную Африку, большую часть Азии и Северную Америку. Сюда же относятся все острова полярного бассейна, Азорские, Мадейра, Зеленого Мыса, а также Японские, кроме самого южного из них. Южная граница царства соответствует границе, разделяющей пояса умеренного и жаркого климатов.
Ландшафты Арктогеи разнообразны. Вдоль северных побережий Евразии и Северной Америки протянулась зона тундры, на юге она сменяется зоной тайги, или хвойных лесов, а еще южнее расположены зоны лиственных лесов, степей и пустынь. Местами зональный характер имеют заросли ксерофитов. В пределах Арктогеи расположены грандиозные горные системы с вертикальной зональностью климатов и растительности.
Фауна Арктогеи, несмотря на громадность территории, сравнительно бедна, что объясняется однородностью основных ландшафтов и исторической молодостью самой фауны.
Млекопитающие здесь насчитывают 7 эндемичных семейств. Это выхухоли (Desmanidae), бобры (Castoridae), тушканчики (Dipodi-dae), селевинии (Seleviniidae), пищухи, или сеноставки (Lagomy-idae), аплодонтовые (Aplodontidae), вилороги (Antilocapridae). Есть здесь и субэндемичные семейства — кроты (Talpidae), гоферовые (Geomyidae) и мешотчатопрыгуновые (Heteromyidae), входящие в пределы Индо-Малайской и Неотропической областей.
Эндемичных семейств птиц еще меньше. К ним относятся тетеревиные (Tetraonidae), гагары (Gaviidae), образующие отдельный отряд, и чистиковые (Alcidae), также составляющие самостоятельный эндемичный отряд. Эндемичных же родов немало (сорока, турухтан, подорожник, пуночка и др.).
Рептилии крайне малочисленны на севере, но к югу их количество и разнообразие возрастают. Есть среди них и одно эндемичное семейство ядозубов, число же эндемичных родов довольно велико.
Амфибии представлены эндемичными семействами углозубов (Hynobiidae), скрытожаберных гигантских саламандр (Cryptobran-chidae), амбистом (Ambystomatidae), саламандр (Salamandridae), амфиум (Amphiumidae), протеев (Proteidae) и сиреневых (Sirenidae).
Все они относятся к хвостатым, но среди бесхвостых эндемиков высокого ранга нет.
Пресноводные рыбы, хотя и уступают по разнообразию тропическим, включают эндемичные семейства осетровых (Acipenseridae), веслоносов (Polyodontidae), панцирных щук (Lepidosteidae), ильных рыб (Amiidae), лососевых (Salmonidae), сиговых (Coregonidae), хариусов (Thymallidae), чукучановых (Catostomidae), умбровых (Umbridae), далий (Dalliidae), щук (Esocidae), пещерных рыб (Amblyopsidae), перкопсид (Percopsidae), афредодерусов (Aphredoderidae), ушастых окуней (Centrarchidae), окуневых (Регcidae), байкальских голомянок (Comephoridae) и широколобок (Cottocomephoridae).
Беспозвоночные Арктогеи далеко не так разнообразны и красочны, как в тропиках. Мало тут и эндемиков высокого ранга. Из эндемичных насекомых здесь обитают отряд гриллоблаттид (Grylloblattidae), реликтовая группа, встречающаяся в Северной Америке, Японии и Российском Приморье, и ряд семейств насекомых, известных лишь энтомологам. Из эндемичных или почти эндемичных следует назвать виды бабочек аполлонов (род Parnassius), жужелиц-брызгунов (Carabus). Встречаются также некоторые эндемичные пауки.
В целом для фауны Арктогеи характерно отсутствие тропических групп, а если они и представлены, то лишь единичными видами широко распространенных родов.
Царство Арктогея состоит из Палеарктического и Неарктического подцарств, расположенных соответственно в Восточном и Западном полушариях.
Подцарство охватывает умеренные и холодные части Евразии и Северной Африки, лежащие к северу от Палеогейского царства, а также ряд архипелагов и островов Атлантического и Тихого океанов (острова Зеленого Мыса, Канарские, Азорские, Исландию — в Атлантике, Командорские, Курильские и Японские — в Тихом океане).
Природные условия Палеарктики достаточно разнообразны. Более детально они будут охарактеризованы при описании областей. Отметим только четко проявляющуюся зональность в распределении климатов и растительности. С севера на юг простираются зоны тундры, тайги, лиственных лесов, степей, субтропических лесных и жестколиственных формаций, пустынь и полупустынь. Значительные пространства занимают горные хребты с присущей им поясной зональностью, во многом аналогичной широтной зональности.
Фауну Палеарктического полцарства можно считать древней тропической фауной Евразии, сильно измененной в результате аридизации конца третичного периода и особенно четвертичного оледенения. Поэтому она, во-первых, сильно обеднена, во-вторых, ей свойственно множество филогенетических молодых групп, обладающих сухо- и холодоустойчивостью. Наличие сравнительно небольшого количества реликтов указывает на бывшие фаунисти-ческие связи этого полцарства с районами Палеогеи.
Поскольку большая часть Палеарктики входит в состав России, изучение ее представляет практический интерес.
Среди млекопитающих Палеарктики есть 2 эндемичных семейства — выхухоли и селевинии. Эндемичных родов много. Из копытных это серна (Rupicapra), косуля (Capreolus), верблюд (Camelus), хомяк (Cricetus) и др.
Насекомоядные кроме названных выхухолей включают кротов, ежей и землероек. Среди последних самыми обычными являются бурозубки (Sorex). Летучих мышей немного, и основная их масса сосредоточена на юге и юго-западе. Из грызунов наиболее типичны беличьи (белки, суслики, сурки, бурундуки), сони, мыши, слепыши и тушканчики. Из зайцеобразных распространены зайцы и пищухи.
Из непарнокопытных в Палеарктике встречаются только виды семейства лошадей — лошадь Пржевальского и кулан, который в числе нескольких подвидов населяет пространство от Ирана до Монголии и Тибета. Парнокопытных здесь тоже сравнительно немного. Это дикий кабан, верблюды, овцы (род Ovis) и козы, зубр и разнообразные олени.
Хищные не очень характерны для данного полцарства. Гиены и виверры заходят лишь в южные его районы. Центр тяжести падает на собак, кошек, куниц и медведей. Последних особенно много.
В Палеарктике отсутствуют хоботные, носороги, жирафы, человекообразные обезьяны и некоторые другие группы, свойственные фауне Эфиопской и Индо-Малайской областей, с которыми она граничит.
Фауна птиц Палеарктического полцарства значительно уступает орнитофауне тропических стран. Тем не менее отдельные группы их эволюционировали и достигали расцвета в последние геологические эпохи именно здесь. Таковы, например, воробьиные — семейства славковых (Sylviidae), свыше 300 видов которых населяет древесные и кустарниковые насаждения, овсянковые (Emberizidae), вьюрковые (Fringillidae). Часто встречаются здесь и вороновые. Среди них есть эндемичные роды — сорока (Pica) и др. Обычны также тетеревиные и куриные, к примеру фазаны. Зато кукушки, дятлы, голуби, сизоворонки распространены значительно меньше, чем в тропиках. А тропикополитные попугаи, нектарницы, птицы-носороги и многие другие семейства либо отсутствуют вообще, либо представлены единичными видами.
Фауна рептилий и амфибий в Палеарктике несравненно беднее, чем в тропических районах Африки или Азии, но в южных частях полцарства достаточно разнообразна и самобытна. Из чешуйчатых характерны ящерицы — настоящие (Lacertidae) и агамы (Agamidae). На крайнем юге встречаются отдельные виды хамелеонов. Ядовитые змеи относятся главным образом к гадюковым. К
ним присоединяются единичные виды ямкоголовых (щитомордники) и аспидовых (кобры). Удавы, столь широко распространенные в тропиках, в Палеарктике представлены лишь мелкими удав-чиками (Егух). Крокодилы, слепозмейки и кожистые черепахи встречаются на юге и то единично.
Амфибии в Палеарктике сравнительно оригинальны. Это выражается в наличии эндемичного семейства углозубов (Hynobiidae). Но отряд хвостатых населяет всю Арктогею. Лягушки и жабы здесь обычны, а из квакш есть только 2 вида.
Пресноводные рыбы в большинстве состоят из видов семейств карповых и лососевых, причем первые преобладают на юге, а вторые — на севере. Два эндемичных семейства — голомянковых и широколобок — населяют озеро Байкал. Из эндемичных родов, которых в Палеарктике немало, заслуживают упоминания караси (Carassius), горчаки (Rhodeus), ерши (Acerina).
Беспозвоночные включают массу эндемичных родов и видов, но они не столь разнообразны и красочны, как в тропиках. Почти не выходят за пределы области хищные жуки-брызгуны рода Сагаbus. To же можно сказать и о красивых бабочках аполлонах, летающих в горах. Термиты и палочники встречаются на юге и являются тропическим элементом фауны, равно как и скорпионы. Более подробно беспозвоночные будут охарактеризованы при описании областей.
Районирование Палеарктики (какой бы ранг ни придавался этому региону) различные зоогеографы проводят по-разному. Схемы Склэтера и Уоллеса базируются на зоогеографических критериях (распространение животных с учетом уровня эндемизма). Н.А.Северцов же в 1877 г. предложил другой принцип районирования — зонально-климатический. Однако при выделении фауни-стических областей необходимо основываться на характеристике фауны, а не климата. Поэтому для зоогеографических целей принцип, предложенный Северцовым, не подходит.
М.А.Мензбир (1934) разделял Палеарктику на три подобласти: Сибирско-Европейскую, Средиземноморскую и Среднеазиатскую. А. П. Семенов-Тян-Шанский (1936) на востоке региона выделил еще одну подобласть — Палеарктическую. Подобного деления придерживался и В.Г.Гептнер (1936), назвав подобласти Европейско-Сибирской, Средиземноморской, Центрально-Азиатской и Маньчжурско-Китайской. Для районирования Палеарктики по Мензбиру—Семенову—Гептнеру характерно то, что весь север Евразии — от побережья полярного бассейна до южных степей — относится к единой подобласти.
Между тем еще Уоллес подметил факт фаунистической неоднородности западной и восточной частей Сибири. Учитывая это обстоятельство, И. И. Пузанов (1949) разделил Европейско-Сибирскую подобласть на Европейско-Обскую и Ангарскую, или Восточно-Сибирскую, проводя границу между ними по Енисею. Енисейский фаунистический рубеж признан в настоящее время не только у нас в стране, но и за рубежом.
Кроме того, следует отметить, что Средиземноморская и Среднеазиатская (или Центрально-Азиатская, по другим авторам) подобласти представляют собой части единой громадной территории, где флора и фауна формировались с верхнемелового или, возможно, палеогенового времени. Первым, кто подчеркнул значение этого генетического центра для развития современной флоры, был М. Г. Попов (1927). Он предложил называть его областью Древнего Средиземья, которая вскоре получила «права гражданства» и в фаунистическом делении Палеарктики.
Вместе с тем выделение особой Евроазиатской степной области неправомочно. Как показал Ю. И. Чернов (1975), степной фаунистический комплекс представляет конгломерат видов — выходцев из нескольких региональных генетических очагов — и не имеет целостности.
Таким образом, фаунистическое подразделение Палеарктиче-ского полцарства можно представить следующим образом:
Подцарство Палеарктика
Европейско-Сибирская область
Европейско-Обская подобласть
Ангарская подобласть Область Древнего Средиземья
Средиземноморская подобласть
Сахаро-Гобийская подобласть Восточно-Азиатская область
В эту область входит весь север Евроазиатского материка — от Британских островов и Западной Европы до Чукотки и Камчатки, где господствует умеренно холодный и арктический климат. Данная область больше, чем любая другая часть Северного полушария, испытала влияние четвертичного оледенения. Это привело к обеднению и видоизменению древней третичной фауны Евразии. Южная граница области проходит по северу Пиренейского полуострова, вдоль Альп и Балкан, через горы Крыма и Главный Кавказский хребет и, не достигая западного побережья Каспия, поворачивает к северу. Далее она идет по полупустыням и степям Поволжья и Северного Казахстана к верховьям Иртыша и Енисея, через Северную Монголию до северо-западных частей хребта Большой Хинган и долины Амура.
Природные условия такой громадной территории чрезвычайно разнообразны. Они будут описаны при характеристике подобластей.
Фауна области (рис. 145, 146) крайне бедна на севере и значительно обогащается к югу. В ней четко проявляется дифференциация по широте (зональная) и несколько слабее по долготе. В Субарктике ареалы животных нередко носят кругоарктический характер, в зоне тайги это проявляется реже, а еще южнее практически отсутствуют.
Европейско-Обская подобласть.
Она объединяет западную половину Европейско-Сибирской области до Енисея. Природные условия и фаунистические особенности ее опишем по ландшафтным зонам. Однако следует подчеркнуть, что в подобласти строгого совпадения границ ареалов и зон почти не бывает.
1.Зона тундры
включает типичную тундру, занимающую побережье Северного Ледовитого океана и некоторые острова, а также лесотундру, которую можно рассматривать как переходную полосу к следующей, таежной, зоне. Условия обитания животных в тундре уже рассматривались, поэтому остановимся на них кратко.
В растительном покрове тундры мхи и лишайники (около 800 видов) преобладают над цветковыми (200—300 видов). На западе зоны развита ягельная тундра, где господствуют лишайники кладонии. Многие растения образуют подушки. Таковы, например, камнеломка и крупка. На болотистых участках растут осоки и пушица. Корневая система растений в данной зоне располагается в верхних ярусах почвы, а надземные части их прижаты к земле. Это говорит о том, что наиболее благоприятным для жизни является ярус у «линии раздела» двух сред — атмосферы и почвы. Здесь же концентрируется большинство педобионтов (обитателей почвы) — червей, личинок насекомых и т. д.
Млекопитающие тундры в пределах подобласти немногочисленны. Из копытных здесь распространен северный олень, встречающийся и на островах Новой Земли. Среди хищников в первую очередь следует отметить песца Alopexlagopus — единственного представителя собачьих, которому свойствен сезонный диморфизм окраски. Летом он грязно-бурый, а зимой снежно-белый, причем у так называемых голубых песцов зимний наряд может быть темно-серым с голубым отливом или коричневым с серебром. Эта форма встречается вместе с белыми и не имеет значения в системагическом отношении. Во время зимних перекочевок песцы доходят до островов Северного Ледовитого океана. Песцы населяют тундру с холмистым рельефом, так как норки роют летом на возвышенных местах и береговых террасах, зимой же прямо в снегу. Питаются песцы в основном леммингами, но не брезгуют и другой как животной, так и растительной пищей.
Кроме песца в тундре можно встретить ведущего полуводный образ жизни белого медведя, где он охотится на леммингов, добывает яйца птиц и птенцов, поедает растительные корма. И все же белый медведь — житель дрейфующих льдов. Только беременные самки долгое время придерживаются безлюдных островов, рождая там в середине зимы медвежат, а в апреле пускаются в странствие по льдам.

Рис. 145. Характерные представители фауны Европейско-Сибирской
области (млекопитающие):
1- белый медведь; 2 – лемминг; 3 – песец; 4 – бурый медведь; 5 – соболь; 6 – косуля; 7 – благородный олень; 8 – летяга;9 – перевязка; 10 – зубр; 11 – выхухоль; 12 – кабарга; 13 – слепыш; 14 – суслик; 15 - бобр

Рис. 146. Характерные представители фауны Европейско-Сибирской
области (от птиц до насекомых):
/ — глухарь; 2 — краснозобая казарка; 3 —
удод; 4 —
сипуха; 5 — орел-могильник; 6 —
дрофа; 7 — огненная саламандра; 8 —
кречет; 9 —
большая синица;
10 —
зеленый дятел; 11 —
стерлядь; 12 —
жужелица; 13 —
жук-олень
Еще несколько видов хищников живут в тундре. Это горностай, ласка, росомаха (Gulogulo). Последняя выполняет здесь важную санитарную роль, поедая трупы северного оленя либо нападая на больных или раненых животных.
Из грызунов в тундре Европейско-Обской подобласти наиболее типичны лемминги (род Lemmus): норвежский (L. lemmus) и обский (L. obensis). Эти зверьки известны тем, что через определенные промежутки времени размножаются в огромном количестве и сразу же предпринимают далекие миграции, переплывая при этом реки и даже узкие морские проливы. Встречаются в подобласти и несколько видов полевок, например, красная (Clethrio-nomusrutilus) и красно-серая (С. rufocanus).
Главную массу позвоночных тундры составляют птицы. Большинство из них посещают тундру только в теплое время года для гнездования и вывода птенцов. Особенно это касается птиц, связанных с водоемами. Летом в тундре обычны казарки (Branta) — краснозобая и белощекая, гусь гуменник (Anserfabalis), малый лебедь (Cygnusbewickii), различные ржанки — тулесы (Squatarolasqua-tarola), галстучники (Charadrius) и прежде всего хрустан (CM. mori-nellus), плавунчики (Phaloropus) и др. Из птиц, не связанных с водоемами, в тундре обитают куропатки — белая (Lagopuslagopus) и тундряная (L. mutus), белая, или полярная, сова (Nycteascandi-аса), кречет (Faicogyrfaico), некоторые овсянковые — подорожник лапландский (Calcariuslapponicus), пуночка (Plectophenaxnivalis), aтакже трясогузковые, особенно краснозобый конек (Anthuscervina).
Пресноводные рыбы представлены лососевыми. Из них типичны семга (Saimosalar), гольцы (Salvelinus) — арктический и другие, несколько видов сигов (Coregonus) и хариусы (Thymallus). Нередки также щука, окунь, карась.
Беспозвоночные тундры однообразны, фауна их бедна. По биомассе первое место занимает дождевой червь эйзения норденшельда (Eisenianordenskioldi). Особи этого широко распространенного не только в тундре, но и в тайге вида на севере довольно мелки, но численность их велика. Лучше других представлены в тундре членистоногие — мелкие панцирные клещи, первичнобескрылые насекомые, комары-кровососы, мухи и шмели, опыляющие цветы. Тундровые шмели отличаются густым опущением. Дневные бабочки немногочисленны. Чаще встречаются перламутровки Clossianafria, С. polaris, С. frigga, сатиры Oeneisnorna, Erebiaempla, E. disa и желтушки (Colias). Следует отметить, что для ряда бабочек тундры характерно арктоальпийское распространение.
Особую группировку животного мира арктических побережий составляют морские птицы и звери. На гнездовьях в массе встречаются кайры, чайки-моевки, тупики, чистики, поморники, образующие птичьи базары.
2.Зона тайги
охватывает хвойные и хвойно-мелколиственные леса, простирающиеся широкой сплошной полосой от Атлантики до Тихого океана. Европейская тайга образована главным образом темнохвойными лесами (ельник, пихтарник). На местах бывших пожарищ или вырубок к ним присоединяются береза, ольха и осина. Часто встречаются моховые болота. Из-за вечной мерзлоты корневая система деревьев расположена в поверхностном слое земли.
Животный мир тайги несравненно богаче, чем в тундре, и не только за счет улучшения климатических условий, но и благодаря обилию кормов, а также многоярусности.
Тайга Европейско-Обской подобласти характеризуется наличием нескольких видов зверей, не распространяющихся на восток, в заенисейскую часть. Это лесная куница, норка и черный хорек. Очень характерны также соболь (Marteszibellina), летяга (Pteromysvolans), лесной лемминг (Myopusschisticolor) — потребитель мхов и лишайников, что очень редко встречается среди зверей, бурундук (Eutamias), запасающий на зиму семена хвойных. Остальные звери менее типичны, поскольку встречаются либо в широколиственных лесах (бурый медведь, косуля, бобр, белка обыкновенная), либо заходят в тундру (заяц-беляк, росомаха, полевки).
Орнитофауна тайги связана с лесными и болотными формациями, где птицы находят кров или пищу. К таковым относятся прежде всего глухарь (Tetraourogallus). Этот самый крупный представитель европейских куриных (размером с индюка) отличается резким половым диморфизмом: самец темный, издали кажется черным, брюхо и хвост с белыми пятнами, самка желтовато-рыжая с белым брюхом. Живет глухарь в крупных массивах старых лесов, гнездится на земле, токует, как известно, на одних и тех же местах из года в год. Глухарь — оседлая птица. Зимой она может зарываться в снег на ночь, а то и днем. Нередок здесь и рябчик (Tetrastesbonasia). В отличие от глухаря он хорошо летает, лавируя между стволами и ветками. Селится в еловых лесах с примесью березы, ивы и осины, особенно охотно — во влажных захламленных лесах с ягодниками. Гнездо, как и глухарь, вьет на земле. Самцы рябчиков токуют и дерутся из-за самок в марте-апреле.
Типично таежными птицами являются клесты. В европейской тайге их 2 вида — еловик (Loxiacurvirostra) и сосновик (L. pityopsittacus). Клесты интересны тем, что время их гнездования может приходиться и на зиму, если достаточно пищи. Вместе с клестами в тайге гнездится щур (Pinicolaenucleator). Наличие клестов и щуров зависит от урожая семян хвойных пород. В неурожайные годы они предпринимают более или менее далекие перекочевки в поисках пищи. Это касается и кедровки (Nucifragacaiyocatactes), не менее типичного обитателя тайги. Однако она, как и близкая к ней ронжа, или кукша (Cractesinfaustus), может питаться ягодами, насекомыми и даже амфибиями.
Обычна в тайге также свиристель (Bombycillagarrulus). Эта прожорливая птица зимой появляется в средних широтах. Питается она ягодами рябины, калины, боярышника и можжевельника, способствуя тем самым распространению данных растений, так как семена их часто не перевариваются в желудке птиц и сохраняют всхожесть. Распространены и белобровый дрозд (Turdusmusicus), вьюрок (Fringillamontifingilla), синицы и хищники — беркуты (Aquilachysaetus), мохноногий сыч (Aegoliusfunereus), длиннохвостая неясыть (Strixuralensis). Часто встречаются трехпалые дятлы (Picoidestridactylus), не имеющие первого пальца на ногах. Живут они в дуплах, которые выдалбливают в елях или лиственницах. Дятлы приносят огромную пользу лесу, истребляя насекомых-вредителей, обитающих под корой, особенно личинок короедов. С многочисленными водоемами таежной зоны связаны чирок-свистунок (Anascrecca), большой крохаль (Mergusmerganser) и чернозобая гагара (Gaviaarctica).
Рептилий и амфибий из-за холодного климата в тайге мало. Из них следует назвать живородящую ящерицу (Lacertavivipara), остромордую лягушку (Ranaterrestris) и реже встречающегося тритона Triturusvulgaris.
Пресноводные рыбы в тайге те же, что и в тундре, но ихтиофауна здесь богаче за счет многочисленных сигов (Coregonus).
Большинство растительноядных насекомых тайги приурочены к хвойным породам и в период массового размножения наносят им большой ущерб. Таковы шелкопряд-монашенка (Ocneriamonacha), сосновый шелкопряд (Dendrolimuspini), еловая огневка (Dioryctriaabietella), гусеницы которой поедают семена ели, пилильщики Cephaleia и рогохвосты Birex. Большое количество в тайге жуков-короедов из родов Ips, Pityogenes, Polygraphus. Очень характерен для европейской тайги жук-восковик (Trichiusfasciatus), родственник наших бронзовок.
Среди менее известных насекомых есть типичные «указатели тайги», т. е. встречающиеся только в таежных условиях. К их числу относятся жужелицы Pelophilaborealis и Trachypachiszettersted, усач Lepturarubra, клопы Araduspictus, живущие под корой хвойных, и тли хермесы (Adelginae), вызывающие образование на деревьях специфических галлов, и др.
В тайге немало и кровососущих — комаров, мошек, мокрецов. Выплаживаясь в массе, они делают жизнь животных и людей труднопереносимой.
3.Зона лиственных лесов
охватывает западный массив широколиственного леса, расположенный в виде полосы в Европе и частично в Западной Сибири между тайгой и лесостепью. Эта зона считается переходной между лесными и травянистыми зонами. Широколиственные леса в умеренных широтах относятся к числу наиболее многоярусных и богатых сообществ. Флористический состав здесь гораздо разнообразнее, чем в зоне тайги. Преобладают дуб, липа, ясень, клен, а в Западной Европе к ним присоединяется бук. Большинство древесных пород регулярно плодоносит. Поскольку желуди, орешки и семена содержат много питательных веществ, в лесах подобного типа широко распространены семеноядные животные. Богатый растительный опад дает обильную пищу педо-бионтам — дождевым червям, личинкам двукрылых и жуков, мелким нематодам, коллемболам и панцирным клещикам. Масса насекомых привлекает в леса насекомоядных птиц и зверьков.
Фауна широколиственных лесов очень богата. Из млекопитающих для зоны характерны благородный олень (Cervuselaphus), образующий в своем ареале много подвидов, косуля (Capreolus), дикий кабан (Susscrofa), в историческое время обитал зубр (Bisonbonasus), а еще раньше тур (Bosprimigenius). Небольшая популяция зубра обосновалась сегодня в заповедниках России, Белоруссии и Польши.
Наиболее типичным хищником зоны до недавнего времени был дикий лесной кот (Felissilvestris), которого сегодня становится все меньше, а в ряде районов ареала его совсем не стало. Обычны волк, лиса и барсук, а из куньих — черный хорек (Mustelaputorius) и европейская норка (Mustelalutreola), придерживающаяся берегов рек и других водоемов.
Огромное количество в данной зоне и грызунов. Заяц-русак (Lepuseuropaeus) в настоящее время расселяется через зону широколиственного леса на север, вытесняя беляка. Это связано с вырубкой лесов — русак придерживается открытых пространств. Самыми характерными жителями лиственного леса следует считать сонь — садовую (Eliomysquercinus), лесную (Dryomysnitedula) и полчка (Glisglis). Все они ведут ночной образ жизни, питаются орехами, желудями и плодами, а зимой впадают в спячку. В этой же зоне находится основная часть ареала бобра (Castorfiber), связанного с водой и питающегося побегами и корой лиственных пород. Из мышевидных грызунов наиболее обычна рыжая полевка (Clethrionomysglareolus), лесная (Apodemussylvaticus) и желто-горлая (A. flavicollis) мыши.
Из насекомоядных самым многочисленным видом становится обыкновенный еж (Erinaceuseuropaeus). Он широко распространен также в лесостепи и даже в степи. Сохранившаяся популяция русской выхухоли обитает главным образом в этой же зоне.
Можно перечислить и типичных для широколиственных лесов птиц, разнообразие которых постоянно растет. Это сугубо лесные птицы. Например, несколько видов дятлов, самым характерным из которых является зеленый дятел (Picusviridis). Из голубей встречаются вяхирь (Columbapalumbus), которого иногда называют витютнем, клинтух (Columbaoenas) и обыкновенная горлица (Strep-topeliaturtur). Последняя нередко встречается на полях, где собирает опавшие при уборке зерна. Совы представлены рядом видов и прежде всего неясытью (Strixaluco), сплюшкой (Otusscops) и сипухой (Tytoalba).
Из хищников следует назвать осоеда (Pernisapivorus), орла-карлика (Hieraaetuspennatus), змееяда (Circaetusgallicus) и мелкого сокола-пустельгу (Faicotinnunculus) — очень полезную птицу, основным кормом которой являются насекомые и грызуны.
Воробьиные настолько многочисленны, что можно отметить лишь наиболее привязанных к широколиственным лесам. Это лесной жаворонок, или юла (Lullulaarborea), красноголовый сорокопут (Laniussenator), певчий (Turdusphilomelos) и черный (Turdusmerula) дрозды. Последний охотно селится в парках и даже на улицах, что обычно для Западной Европы. Соловей (Luscinialuscinia) — лучший певец среди европейских птиц — также весьма обычен на опушках леса или в парках. К ним присоединяются зарянка (Erithacusrubecula), пеночка-трещотка (Phylloscopussibilator), синица-лазоревка (Paruscoeruleus). Из вьюрковых с широколиственными лесами связаны дубонос (Coccothraustescocco-thraustes), зеленушка (Chlorischloris) и прекрасная иволга (Oriolusoriolus) — украшение европейской орнитофауны. Иволги приносят огромную пользу, истребляя вредителей леса — волосатых гусениц, которых не едят большинство других птиц.
Преимущественно к зоне лиственных лесов приурочены и вороновые — грач, серая и черная вороны, галка. Эти птицы давно уже стали спутниками человека, и их чаще можно увидеть в городах и селах, чем в лесу.
Разнообразие и численность рептилий и амфибий в этой зоне постоянно растут. Болотная черепаха (Emysorbicularis) распространена всюду, хотя в густонаселенных областях Западной Европы она становится все более редкой. Из ящериц на западе зоны встречается зеленая (Lacertaviridis), прыткая же ящерица (L. agilis) доминирует всюду. Из безногих ящериц обычна веретеница (Anguisfragilis), которую благодаря змеевидным движениям нередко принимают за змею. Эта совершенно безобидная рептилия приносит пользу, уничтожая слизней и личинок насекомых. Змеи представлены обыкновенной гадюкой (Viperaberus), ужом (Natrix) и медянкой (Coronellaaustriaca). Медянку часто несправедливо считают ядовитой и уничтожают.
Среди амфибий очень распространены квакша (Hylaarborea) — единственный европейский вид этого семейства, краснобрюхая жерлянка (Bombinabombina), жаба-повитуха (Alytesobstericans), самцы которой носят выметанную самками икру на своих бедрах до вылупления головастиков, прудовая (Ranaesculenta) и озерная (Ranaridibunda) лягушки. Последняя поедает главным образом насекомых, но иногда нападает и на мелких птиц. В рыборазводных прудах озерная лягушка питается мальками рыб и таким образом наносит определенный вред.
Пресноводные рыбы представлены осетровыми (немецкий осетр, белуга, стерлядь и др.) и карповыми (густера, плотва, лещ, голавль, европейский горчак и др.).
Беспозвоночные, особенно насекомые, достигают в подобласти небольшого разнообразия. Перечислим только самых типичных. Из бабочек это переливницы Apaturairis и A. ilia, ленточница LimenitisCamilla, пестрокрыльница (Araschnialevana), известная своим сезонным диморфизмом, перламутровки Brenthisino, Clos-sianadia, сатиры Minoisdryas, Erebiamedusa, голубянка коридон (Lysandracoridon) и масса других. Из жуков характерны хищные красотелы (Calosoma), охотящиеся на гусениц бабочек, жужелицы рода Carabus, особенно луговая (С. arsensis) и жужелица-про-круст (С. coriaceus), жук-олень (Lucanuscervus) — украшение европейских дубрав, красивый усач Rosaliaalpina, многочисленные листоеды, долгоносики и др.
Следует остановиться и на особенностях фауны лесостепи — переходной полосы, или подзоны, отделяющей лесную зону от открытых пространств степей. Существующий в области контакта различных сред пограничный эффект приводит, как известно, к интенсификации формообразовательного процесса. Это проявляется и в лесостепи в виде особенного фаунистического богатства и высокой насыщенности ценозов. К-В.Арнольди (1965) изучал вопрос о специфичности лесостепной фауны и привел большой список видов насекомых, характерных именно для лесостепи (жужелицы-брызгуны — 7 видов, листоеды Chrysochusasclepiadeus, ряд многоножек и наземных моллюсков). Подобные факты отмечаются и для птиц. Проблема самобытности и автохтонности животного мира лесостепи требует дальнейшего анализа, но его богатство бесспорно.
Подразделение зоны широколиственных лесов по фаунисти-ческому принципу (в отличие от зоны тайги) сводится к выделению двух провинций — Западноевропейской и Восточноевропейской. Граница между ними проходит в западных частях Белоруссии, Украины и на востоке Молдавии. Она примерно совпадает с границей распространения бука и граба.
Разница между фаунами этих провинций выражается в следующем. В Западноевропейской провинции распространены такие виды, как косуля, лесная кошка, малый подорлик, западный сарыч, сипуха, канареечный вьюрок, красноголовый сорокопут, мухоловка-белошейка, пятнистая саламандра, жаба-повитуха, немецкий осетр и масса эндемичных для Западной Европы насекомых: сатиры альциона и статилинус, золотая жужелица и др. Восточноевропейская провинция имеет смешанную фауну. Некоторые западноевропейские виды проникают и сюда, но чаще они заменены викарными видами или даже сибирскими фаунистическими элементами. Для этой провинции характерны выхухоль, сибирская косуля, черный коршун, восточный соловей, овсянка-дубровник и специфичные виды насекомых.
Зона европейских широколиственных лесов относится к территориям, издавна освоенным и преобразованным человеком. Поэтому в большинстве районов аборигенная фауна сохранилась только частично (особенно в Западной Европе). Ее можно наблюдать в заповедниках и резерватах, имеющихся в большинстве европейских стран.
4.Зона степей
почти полностью видоизменена деятельностью человека. Еще в исторические времена степи тянулись относительно узкой полосой от Венгерской низменности до Барабинской низменности в Сибири. Южной границей степной зоны служит побережье Черного моря, горы Крыма и Кавказа, полупустыни и пустыни Казахстана и Средней Азии.
Степная фауна характеризуется прежде всего отсутствием настоящих лесных видов. Отдельные же представители мезофильных групп, связанные с древесными породами, размещаются в инт-розональных биотопах — лесных балках, пойменных лесах вдоль рек и т. д.
Среди млекопитающих копытные в наше время совершенно вытеснены из степей человеком, но ранее здесь были большие стада диких лошадей (тарпанов), сайгаков и диких быков, или туров. Из хищников в степях существуют только эвритопные волк и лиса, а также специфические для этой зоны степной хорек (Putoriuseversmanni), истребляющий сусликов, и перевязка (Vormelaperegusna).
Наиболее же характерными для степи зверьками должны считаться грызуны и прежде всего суслики — малый (Citelluspyg-maeus), крапчатый (С. suslicus) и на крайнем западе зоны серый (С. citellus). Кое-где сохранился и самый крупный грызун европейских степей — байбак, или степной сурок (Marmotabobak). Но на большей части наших степей сурков уже нет и остались только сурчины — холмы высотой до 1 м, а в поперечнике 8—15 м, возникшие в результате рытья животными сложных нор. Ночной образ жизни ведут тушканчики — прыгающие зверьки с мощными задними ногами и длинным хвостом с кисточкой на конце. В степной зоне подобласти их несколько видов: земляной заяц (Allactagamajor), малый тушканчик (A.elater), емуранчик (Stylodipustelum). Очень характерны ведущие подземный образ жизни слепыш обыкновенный (Spalaxmicrophthalmus) — любитель цветных целинных степей, и слепушонка (Ellobiustalpinus). Из мышевидных грызунов обычны полевки Microtussocialis.
Заяц-русак представлен особым западным подвидом. Из насекомоядных повсюду встречается обыкновенный еж, и лишь на востоке подобласти появляется ушастый еж (Erinaceusauritus), широко распространенный на равнинах Средней Азии.
Орнитофауна степей содержит кроме широко распространенных эвритопных видов или же спутников человека многие типичные именно для этой зоны виды. Из куриных в степи встречаются серая куропатка (Perdixperdix) и перепел (Coturnixcotumix), который активно расселяется и в другие зоны. Крупная дрофа Otistarda в прошлом была здесь одной из очень многочисленных птиц, ее стаи насчитывали до 200—300 особей; с распашкой целинных степей и интенсивной охотой, в том числе и браконьерской, от былого обилия ничего не осталось, и дрофа теперь стала очень редкой. Та же судьба постигла и стрепета (Otistetrax). Если дрофа еще кое-где гнездится среди посевов озимых, то стрепет на распаханных и освоенных землях не живет.
Журавль-красавка (Anthropoidesvirgo) — житель степного юга. Когда-то он был широко распространен от низовьев Дуная до Казахстана и Юго-Восточной Сибири. Сейчас это крайне редкая на западе степной зоны птица, а на Украине она практически исчезла.
Хищные птицы представлены степным орлом (Aquilarapax), гнездящимся на земле и питающимся грызунами, степным лунем (Circusmacrourus), также гнездящимся на земле, и степной пустельгой (Faiconaumanni), которая истребляет в большом количестве насекомых, особенно саранчовых.
Из птиц других семейств заслуживают упоминания золотистая щурка (Meropsapiaster), селящаяся колониями в глинистых обрывах, удод (Upupaepops) и, безусловно, жаворонки. В степях Евро-пейско-Обской подобласти их несколько видов: полевой (Alaudaarvensis), хохлатый (Galeridacristata), малый (Galandrellacineria), aна востоке — черный (Melanocoryphayeltoniensis). Песня жаворонка знакома всем, кто бывал в степи.
Богата и фауна водоплавающих птиц, населяющих водоемы степной зоны. На побережье Черного моря встречаются своеобразные утки-пеганка (Tadornatadorna) и огарь (Т. ferruginea).
Из рептилий распространены крупные полозы, особенно желтобрюх (Coluberjugularis), достигающий 2-метровой длины, степная гадюка (Viperaursini), ящурка разноцветная (Eremiasarguta), которая, как правило, приурочена к пескам.
Из амфибий в степи чаще всего встречаются озерная лягушка и зеленая жаба, отличающаяся большой эвритопностью. Оба вида истребляют много вредных насекомых.
Пресноводные рыбы бассейна Черного и Каспийского морей практически те же, что и в водоемах зоны широколиственных лесов. Особенно характерны осетровые, а среди них — стерлядь.
Беспозвоночные степной зоны включают уже виды средиземноморского и среднеазиатского распространения. Таковы паукообразные ~ сольпуга из рода Galeodes, юхинфусский тарантул (Lycosasingoriensis) — крупный паук, роющий вертикальные норки в земле, ядовитые каракурты. В степной зоне найдены пауки семейств Atypidae и Zoropsidae, обнаруженные в других зонах.
Из насекомых в степях обычны и многочисленны саранчовые. Ряд их видов серьезно вредит сельскохозяйственным растениям. Это итальянский прус, мароккская, а также перелетная саранча. Однако чаще встречаются одиночные саранчовые, или кобылки, коньки и травянки. Появляются и богомолы из родов Mantis, Bolivaria и Iris. Тараканы также имеют своего представителя — степного таракана (Ectobiusduskei) — единственного степняка в этой мезофиль-ной и лесной группе. Типично степные виды жуков есть среди чернотелок, златок, листоедов, усачей, жужелиц и других семейств. Многие из них являются «указателями степи», или индикаторными видами. Таковы крупные жужелицы из рода Carabus — С. bes-sarabicus, С. besseri, усачи рода Dorcadion, листоеды Cryptocephalusgamma и С. lateralis и др. Из бабочек отметим лугового мотылька — известного вредителя, степную зорьку (Zegriseupheme), сатира-бризеиду, совок (яровую, стеблевую и др.). Среди мух значительное место в степной фауне занимают хищные ктыри, особенно выделяется гигантский ктырь (Satanasgigas) — самая крупная муха в нашей фауне. В степной зоне много муравьев из родов Camponotus, Proformica, Cataglyphis (бегунки) и Polyergus (амазонки).
Наземные моллюски придерживаются в основном интрозональ-ных биотопов, но около 15 видов их (из 71) можно отнести к типичным обитателям степей европейской части подобласти. Это виды рода Helicella (ксерофилы по природе), несколько видов рода Helix (к ним относится и виноградная улитка) и др. Степные моллюски являются промежуточными хозяевами паразита скота — ланцевидной двуустки.
Большинство дождевых червей степной зоны относятся к широко распространенным видам, иногда с всесветным распространением. Но 2 вида — Eiseniagordejeffl и Dendrobaenaintermedius — характерны только для зоны степей. Полезная роль дождевых червей в почвообразовательном процессе общеизвестна. Но они могут приносить определенный вред, поскольку являются носителями и передатчиками паразитарных заболеваний домашних животных (свиней, птиц).
Итак, необходимо подчеркнуть, что степь как тип ландшафта формировалась при активном участии животного населения. Это в первую очередь относится к почвообразованию, в котором принимали участие дождевые черви, насекомые, крупные пауки, роющие грызуны. В заволжских степях выбросы сурков в прошлом столетии занимали 39,9 м3
на гектар. Если почву сурчин распределить равномерно на этой площади, то она образовала бы слой в 4 мм. Большой объем занимают выбросы сусликов, слепышей и других землероев. Все эти выбросы играют важную роль в создании микрорельефа степей, а также в смене растительного покрова. Существенным является воздействие на растительность ряда насекомых. К примеру, при массовых размножениях молдавская огневка объедает типчак и его место занимают вероника и другие двудольные. Весьма заметны изменения степной растительности, вызываемые пасущимися в степях копытными. У водопоев, вдоль троп и на местах стоянок в прошлом и в настоящее время происходит чрезмерный выпас и уничтожение растительности. Однако если в доисторические времена это делали стада тарпанов, сайгаков и, может быть, зубров, то сейчас пастбищная дигрессия — результат выпаса овец.
Длинная и узкая степная зона, естественно, подразделяется по фаунистическому принципу по меньшей мере на две провинции. Понтийская провинция охватывает степи Восточной Европы и юга европейской части России, степи Крыма, Предкавказья, Донецкие и правобережной Украины. В последних водятся многие виды (например, крот и малый слепыш, жук-красавчик, жужелица Бессера), которые не распространяются к востоку за Днепр. Это подчеркивает значение нижнего течения Днепра как фаунистической границы. Река здесь служит и западной границей ареалов малого суслика, степной пеструшки, тушканчика. В степном
Крыму отсутствует ряд видов, характерных для Донецких степей. Там, к примеру, нет слепыша, степной пеструшки, байбака, полевой мыши и т. д. В предкавказских степях снова появляются крот и особый вид хомяка. Восстановился здесь ареал сайгака.
Восточная часть степной зоны в пределах подобласти относится к Урало-Барабинской провинции, которая постепенно переходит в полупустыни и пустыни, относящиеся уже к другой области Палеарктики. Фауна ее обогащается за счет среднекав-казских элементов. Появляется лисица корсак, гигантский слепыш заменяет обыкновенного, суслики представлены особыми видами (или формами большого суслика) — рыжеватым и краснощеким. Очень характерна степная пищуха (Ochotonapusilla), aиз птиц — кречетка, или степная пигалица (Chettusiagregaria), и черный живоронок.
Ангарская подобласть.
Она охватывает Восточную Сибирь. Ее западная граница проходит по долине Енисея («линия Иогансено»). Южная — от Алтая до северной оконечности Большого Хингана и далее вплоть до Охотского моря. Остров Сахалин большинство зоо-географов тоже относят к Ангарской подобласти.
Отличия животного мира Восточной Сибири отмечались еще во времена А. Ф. Миддендорфа (1869). Очень четко показал разницу в составе орнитофауны таежной Восточной и Западной Сибири П.П.Сушкин (1925). Выделение же этого региона в самостоятельную подобласть и уточнение ее границ принадлежат И.И.Пузанову (1949). Фауна Ангарской подобласти аналогично Европейско-Обской подразделяется на ландшафтные зоны.
1.Зона тундр Восточной Сибири
содержит большое количество циркумполярных элементов — видов, распространение которых охватывает безлесные районы Крайнего Севера материков Евразии и Северной Америки. И все же, как ни похожи природные условия этой зоны в обеих подобластях, фаунистические различия их достаточно выразительны.
Млекопитающие представлены леммингами либо тех же видов, что и на западе (но другими подвидами), либо своим видом — желтобрюхим леммингом (Lemmuschrysogaster). В тундру заходят и виды, характерные больше для альпийской (гольцовой) зоны восточносибирских хребтов. Это северная пищуха (Ochotonaalpina), черношапочный сурок (Marmotacamschatica) и суслик длиннохвостый (Citellusundulatusparryi). Белый медведь и волк представлены самостоятельными подвидами.
Среди водоплавающих птиц, для которых тоже свойствен цир-кумполярный ареал, есть и типичные восточносибирские виды:
гусь-белошей (Ansercanagicus), белый гусь (A. coerulescens), черная казарка (Brantabemicia), сохранившаяся в немногочисленных районах тундры, кулик-дутыш (Calidrismelanotus) и др.
Необходимо отметить, что к фауне крайнего северо-востока Азии (Чукотка, бассейн р. Анадырь) примешивается много американских видов. Число их постепенно убывает к западу. Но уже в низовьях Лены вместо американских элементов появляются западносибирские.
2.Зона восточносибирской тайги
при всем сходстве с европейской и западносибирской имеет ряд специфических отличий. Огромную площадь здесь занимают леса из даурской лиственницы. На юге они сменяются лиственнично-сосновыми лесами. Ель встречается спорадически и только в условиях местного более влажного климата.
Распределение широтных зон в Восточной Сибири такое, что тайга, поднимаясь по хребтам, переходит в альпийскую зону, а на юге наблюдается чередование тайги с участием степей. Таким образом, широколиственные леса как зона отсутствуют.
Заенисейская тайга населена особыми формами копытных — оленем маралом (Cervuselaphussibiricus), близким к канадскому оленю, восточносибирскими подвидами лося и северного оленя. Самым характерным видом из копытных этой зоны следует считать кабаргу (Moschusmoschifenis) — безрогого оленя с сабле-образно выступающими верхними клыками. Кабарга населяет горную тайгу и питается хвоей пихты, сосны и кедра, а также травянистыми растениями и лишайниками. Кабарга — промысловый зверь. Его добывают ради мяса, шкуры и главным образом ради мускусной железы, секрет которой применяется в парфюмерии.
Хищники в зоне представлены несколькими местными подвидами соболя (Marteszibellina). Особенно ценится баргузинский соболь. Колонок (Mustelasibirica) и солонгой (Mustelaaltaica) широко распространены, но обычно не встречаются совместно. Рысь отличается крупными размерами и выделяется в особый забайкальский подвид. Лесной куницы, норки и черного хорька в Восточной Сибири нет.
Из грызунов здесь обитают бурундук (очень многочисленный в тайге Ангарской подобласти), лемминги Lemmusamurensis и Myopusschisticolor, белки и летяги. Насекомоядные отличаются большим своеобразием. В зоне водятся особый виц крота Talpaaltaicaи даурский еж, несколько эндемичных видов землероек-бурозубок (Sorex).
Специфична и орнитофауна заенисейской тайги. Вместо европейского глухаря здесь встречается каменный глухарь (Tetraoparvirostris), рябчика обыкновенного сменяет дикуша (Falcipennisfalcipennis). Обращает на себя внимание поведение этой птицы: будучи потревожена, она не пытается скрыться, а взлетает на нижние ветки и спокойно усаживается, глядя на человека. За такую доверчивость местные жители называют ее «смиренным рябчиком». Добывают дикушу без выстрела — снимают с ветки петлей, привязанной к палке. Воробьиные восточносибирской тайги включают в себя как широко распространенные таежные виды, так и характерные только для нее. Из последних назовем сибирского (Turdussibiricus) и темного (Т. obscurus) дроздов, серого снегиря (Pyrrhulacinerea), розовую чечевицу (Erythrmarosea), сибирскую мухоловку (Muscicapasibirica), толстоклювую пеночку (Herbivoculaschwarzi), синего соловья (Lusciniacyane). Ангарская подобласть — родина овсянки-дубровника (Emberizaaureola), расселившейся на запад.
Амфибии и рептилии из-за сурового климата в тайге Восточной Сибири встречаются крайне редко. Сибирский углозуб, или четырехпалый тритон (Hynobiuskeyserlingi) — единственный в Палеарктике представитель восточноазиатского рода Hynobius из подотряда скрытожаберных. Обыкновенная жаба представлена местным подвидом Bufobufoasiaticus. Из змей есть гадюка и щитомордник (Ancistrodonhalys), а из ящериц — только живородящая.
Фауне пресноводных рыб свойственно прежде всего наличие эндемичных байкальских семейств голомянок и широколобок, или байкальских бычков. Последних в водах Байкала 24 вида из 8 родов. Голомянок же всего 2 вида. Большая голомянка дает ценный жир. Чрезвычайно интересно также присутствие чукучанов из американского семейства чукучановых рыб (Catostomidae). В реках Восточной Сибири из этого семейства встречается всего 1 вид, а в Северной Америке оно представлено 18 родами и 84 видами. Тихоокеанские лососи рода Oncorhynchus, относящиеся к ценным промысловым рыбам — кета, горбуша, чавыча, красная (нерка), кижуч и семга — проходные рыбы, нерестящиеся в реках Восточной Сибири. Большинство других рыб входит в циркумполярную зону.
Беспозвоночные тайги Ангарской подобласти включают массу эндемичных видов. Среди насекомых можно отметить сибирского аполлона (Parnassiusstubbendorfl), желтушку Coliasviluensis, кедрового шелкопряда (Dendrolimussibiricus) — серьезного вредителя хвойных пород, несколько своеобразных жужелиц-брызгунов, в частности Carabusmeander и С. hummeli, усачей Pronocerabrevicollis и Xylotrechusaltaicus, рогохвостов Paururusermak и многих других. Большое количество видов не идет на восток дальше долины Енисея, например бабочка черный аполлон (Parnassiusmnemosyne).
3.Зона лесостепи и степи
выделена довольно условно. Под ней подразумеваются отдельные пятна степей среди тайги, с одной стороны, и более или менее длинные языки тайги либо сосново-березовых лесов в окружении полупустынь или сухих степей Северной Монголии — с другой. В Забайкалье встречаются разно-травные степи с холмистым рельефом. Степь богата водоемами — речками, озерами. Зима здесь малоснежная и суровая. Соответственно и животный мир весьма пестр и не составляет единого целого. Тем не менее ряд видов придерживается именно таких ландшафтов.
Млекопитающие представлены антилопой дзереном (Ргосарга gutturosa), многочисленными сурками тарбаганами (Marmotasibirica), даурской пищухой (Ochotonadaurica) и даурским сусликом (Citellusdauricus), нередок заяц-толай (Lepustolai), распространенный также в пустынях Средней Азии и даже Аравии и Северо-Восточной Африки. Цокор (Myospalaxmyospalax) сменяет слепыша и также ведет подземный образ жизни, питаясь сочными клубнями и корешками растений. Другие виды расселены значительно шире, и степи Ангарской подобласти — только часть их ареала (волк, лисица, корсак, ушастый еж, джунгурский хомячок и др.).
Орнитофауну заенисейских степей нельзя назвать слишком бедной, но типичных видов здесь немного. Если полевой жаворонок и журавль-красавка обитают по открытым пространствам умеренной Евразии, то саджа, или копытка (Syrrhaptesparadoxus), дрофа Дыбовского (Otisdybovskii), бородатая куропатка (Perdixdaurica) и некоторые другие птицы характерны в основном для этой зоны.
То же относится и к рептилиям. Среди них есть и широко распространенные виды, и степные монгольско-сибирские типа монгольской ящурки (Eremiasargus).
Большие горные хребты Ангарской подобласти нарушают зональность ландшафтов и приводят к фаунистическим инверсиям. Хребты Алтая, Саян, Хэнтей, Яблоновый и Становой пересекают лесостепи и степи или окружены ими. Альпийская зона этих горных хребтов похожа своей растительностью на тундру и на севере сливается с ней. Зато на юге в высокогорья длинными языками проникают степные формации вместе со свойственными им животными.
В высокогорьях юга Ангарской подобласти из копытных обитает баран аргали (Ovisammon), а в восточной ее части — снежный баран, или чубук (Oviscanadensis), точнее — азиатские его подвиды, так как ареал вида охватывает не только северо-восток Азии, но и северо-запад Америки. Сибирский козерог, или горный козел (Caprasibirica), живет не только в указанных районах подобласти, но и в горах Средней Азии, Афганистана и Северо-Западного Китая.
Весьма обычны и многочисленны в горах грызуны. Это камчатский сурок (Marmotakamtschatica) с подвидами, пищуха (Ochotonaalpina) и даже суслики, заходящие из степей. Только для альпийской зоны Ангарской подобласти характерны каменные полевки (род Alticola) — представители самых высокогорных млекопитающих. Один из их видов — лемминговидная полевка (A. macrotislemminus) — интересен тем, что на зиму белеет. Питаются полевки травами и лишайниками.
Среди птиц высокогорий распространены бекас-отшельник, или горный дупель (Gallinagosolitaria), алтайский улар (Tetraogalusaltaicus), белая и тундряная куропатка и другие птицы, усиливающие сходство альпийской тундры с зональной. Это сходство особенно подчеркивается насекомыми, имеющими типичные арктоальпийские ареалы (бабочки рода Erebia). В ряде случаев арктоальпийские виды на юге Ангариды выступают в качестве ледниковых реликтов.
Продвижение степных элементов фауны в высокогорья, а через них и в тундру можно рассматривать как последствия аридиза-ции климата в послеледниковую эпоху.
Область Древнего Средиземья лежит к северу от Северного тропика, охватывает аридные и субаридные территории, окружающие Средиземное море и входящие в состав Средней и Центральной Азии.
Природные условия ее при всем разнообразии характеризуются прежде всего аридностью климата. Осадки выпадают главным образом в холодное время года, лето сухое, что обусловило развитие пустынь, протянувшихся широкой полосой от Северной Африки до Центральной Азии.
На западе области (в странах, окружающих Средиземное море) произрастают вечнозеленые жестколиственные леса, а в районах, подвергающихся воздействию человека, — ксерофитные кустарники и полукустарники, образующие формации, называемые маквис и томилляры. Южнее и восточное уже встречаются полупустыни и настоящие пустыни, занятые ксерофильными полукустарниками (полыни, солянки, парнолистник) и древовидными растениями (саксаулы, песчаная акация), среди которых в зимне-весеннее время вегетируют эфемероиды (мятлики, луки, тюльпаны, ферулы) и эфемеры (костры, вероники, малькольмии).
Чередование в рельефе области понижений и гор обусловливает большое разнообразие растительности, что сказывается и на распределении животного мира. Здесь преобладают виды, связанные с открытыми ландшафтами. Из млекопитающих распространены семейства селевиниевых, тушканчиковых, многие песчанки, из грызунов — слепыши, обитают также виверровые, гиеновые, кошки (степная, барханная, манул, каракал), из копытных характерны ослы, горные бараны (муфлоны), антилопы, лань, из населяющих Палеарктику родов немало и эндемиков. Благодаря обилию пещер в горах многочисленны летучие мыши, в том числе подковоносы и длиннокрылы.
Орнитофауну составляют рябки, дрофы, жаворонки, пустынные сойки, из воробьиных — своеобразный канареечный вьюрок (родоначальник комнатной канарейки), испанский воробей, сорокопуты, славки Sylviaundata, S. hortensis, каменки Oenanthehispanicaи О. deserti. В условиях горного рельефа живут грифы из родов Gyps, Aegypius и Neophron, горные куропатки, или кеклики (Alectorisgraeca). Обитатели водоемов имеют явно тропический или субтропический облик, как, например, фламинго и пеликаны.
Рептилий здесь значительно больше, чем в других областях Па-леарктического полцарства. В основном это семейства гекконов из родов сцинковых гекконов (Teratoscincus) и геккончиков (Also-phylax), эндемичных для области, агамовых (среди них роды змееящериц (Ophiomorus) и круглоголовок (Phrynocephalus) распространены только в пределах области) и настоящих ящериц. Последние особенно разнообразны, а род Lacerta богат видами именно в Средиземноморье, что дает основание считать этот район местом происхождения представителей рода. Много в области также ящурок (Eremias). Семейство веретеницевых представлено крупным желтопузиком (Ophiosaurusapodus) с восточносредиземно-морским ареалом.
Среди змей довольно разнообразны ужи и полозы. Заслуживает упоминания эскулапов полоз, изображение которого (чаша с обвившейся вокруг змеей) служит эмблемой медицины. Из удав-чиков (Егух), распространенных в Африке и Азии, встречаются степной, или западный, восточный песчаный. Настоящие ужи (Colubrinae) представлены как широко распространенными, так и присущими только области родами. К последним относятся роды медянок (Coronella), литоринхов (Lytorhynchus), ринокаламусов (Rhynchocalamus), эйренисов (Eirenis). Лишь обыкновенная медянка выходит в Европе за пределы области на север. Среди ядовитых змей на первом месте стоят гадюковые и именно род Vipera (гадюка степная, носатая, кавказская, курносая, гюрза и др.), насчитывающий здесь не менее 10 видов. Из других гадюковых следует назвать виды родов Pseudocerastes и Echis (эфы). Есть и кобры (Naja) — очковая (некоторые подвиды не имеют очкового рисунка) и египетская, или гая. В области Древнего Средиземья встречается единственный палеарктический вид семейства слепозмеек (Typhlopidae).
В отличие от рептилий состав амфибий здесь беден. Из отряда хвостатых семейства протеев (Proteidae) в пещерных водоемах Югославии обитает европейский протей (Proteusanguinus). Этот вид живет в полной темноте (глаза скрыты под кожей). У ближайшего его родственника — американского протея (правда, из другого рода) глаза развиты и хорошо заметны. В области Древнего Средиземья распространены и несколько родов настоящих саламандр — Salamandra, Martensieles, Chioglossa, Salamandrina. Безлегочные саламандры (Plethodontidae), подавляющее большинство которых обитают в Северной Америке, представлены только одним родом, живущим в Южной Европе. Есть в области также эндемичные роды тритонов. Из бесхвостых амфибий встречаются чесночницы (сирийская и испанская), из жаб и настоящих лягушек — несколько эндемичных (закавказская, малоазиатская лягушки) и ряд широко распространенных видов, но они немногочисленны.
В реках и озерах области много пресноводных рыб. Однако следует сказать, что запад и восток ее значительно различаются по составу ихтиофауны. Средиземноморье характеризуется присутствием карпозубых — тропических рыб, таких, как афании (Aphanius) и цихлиды, что придает фауне несколько экзотический облик. В основном же здесь распространены карповые, лососевые, осетровые, в том числе севрюга, осетры, ложные лопатоносы (Pseudo-scaphirhynchus), живущие в бассейне ареала. Обыкновенные сомы (Siluridae) кроме европейского представлены эндемичным родом Parasilurus, один вид которого — сом Аристотеля — живет в реках Греции. В составе ихтиофауны области встречаются также горные (роды Glyptosternum и Glyptothorax) и клариевые (Clarias) сомы. Последние населяют водоемы африканской части Средиземья.
Беспозвоночные области Древнего Средиземья отличаются обилием теплолюбивых южных групп и вообще многочисленны и разнообразны. Здесь встречаются тропические группы, заходящие в область с юга: скорпионы, сольпуги, пауки-тарантулы и др. Множество родов насекомых имеют типичные древнесредиземноморские ареалы. Из бабочек к ним относится Zerynthia, из жуков — кравчики Broscus, Duvalius, Pimelia, Lethrus, (усачики) Dorcadion, (листоеды) Lachnaia, Stylosomus. Еще большее число родов энде-мичны для отдельных частей области. То же можно сказать о наземных моллюсках.
Область Древнего Средиземья расчленяется на две подобласти — Средиземноморскую и Сахаро-Гобийскую.
Средиземноморская подобласть.
К ней относятся горные районы Южной Европы (Пиренейский, Апеннинский и Балканский полуострова), север Африки (горные части Алжира, Туниса и Марокко), а также прилегающие к Средиземному морю части Малой Азии и Ближнего Востока.
Средиземноморская подобласть в основном покрыта лесами. Как уже говорилось, доминирующая растительная формация здесь — вечнозеленые жестколистые леса, а маквис и томилляры составляют вторичную формацию. В среднегорном поясе господствуют листопадные полуксерофитные кустарники, получившие название шибляк. В более высоких поясах развиты хвойные леса.
Из млекопитающих для фауны Средиземноморской подобласти в целом характерны некоторые копытные, например виды рода Ovis (муфлоны), лань (Cervusdama), которую завезли и в страны Западной Европы, Новую Зеландию, а также в Америку; виверровые хищники — генетта (Genettagenetta), ихневмон (Heprestesichneumon); мелкие кошки Felisocreata — на Сардинии. Медведи сохранились в горах Марокко, в Пиренеях и на Балканском полуострове. Из насекомоядных следует отметить пиренейскую выхухоль (Calemuspyrenaica).
Из многочисленных и разнообразных птиц в первую очередь необходимо назвать голубую сороку (Cyanopicacyanea), имеющую разорванный ареал, западная часть которого охватывает Пиренейский полуостров, а восточная — Дальний Восток. Обычны воробьи рода Petronia, канареечные вьюрки, славки Sylviaundata, S. hortensis и S. cantillans, пересмешка Hippolaispolyglotta, каменка Oenanthehispanica, тугайный соловей (Erythropygiagalactotes), средиземноморская славка-портной (Cisticolajuncidis). Из водоплавающих птиц типичен мраморный чирок (Anasangustirostris).
Неотъемлемым элементом средиземноморского горного ландшафта являются грифы и стервятники. Черный гриф (Aegypiusmonachus) и белоголовый сип (Gypsfulvus) наиболее свойственны этой подобласти, а обыкновенный стервятник (Neophronpercnopterus) распространен до Средней Азии и Индии. Фламинго, пеликаны и змеешейки составляют экзотический элемент средиземноморской орнитофауны.
Рептилии приспособлены к сухому и теплому климату. Они представлены большим количеством специфических видов, черепахи — родами Clemmys (водяные черепахи), Emys (болотная черепаха) и Testudo (наземные черепахи), в котором наиболее известна греческая черепаха (Testudograeca). Местами обычны также балканская (Т. hermanni) и окаймленная (Т. marginatta) черепахи.
Хамелеоны являются выходцами из Эфиопской области. Встречаются они только в Южном Средиземноморье (Chamaeleochamaeleon и другие виды). Гекконы же гораздо многочисленнее и нередко поселяются даже в домах. Они принадлежат к родам Eublepharis, Gymnodactylus, Tarentola. Есть в подобласти также агамы, но только на южном и восточном побережье Средиземного моря. Среди них выделяются шипохвосты (Uromastix), заходящие по окраинам песков из соседней Сахаро-Гобийской подобласти. Группа сцинков представлена на островах Средиземного моря красивой золотистой мабуей (Mabuyaaurata), а также веретенообраз-
ными (Chalcides) и длинноногими (Eumeces) сцинками. Многочисленны в Средиземноморье широко распространенные в Старом Свете настоящие ящерицы. Среди них эндемична жемчужная (Lacertalepida), до 75 см в длину, населяющая Пиренейский полуостров, Южную Францию, Италию и Северо-Западную Африку. Особенно много так называемых «степных ящериц», живущих на скалах и в населенных пунктах (L. muralis, L. saxicola и др.). Желтопузик, или блавор, как его называют на Адриатике, не менее обычен. Вараны же для Средиземноморья (в узком смысле) совершенно не характерны. Зато в Северо-Западной Африке встречается амфисбена Trogonophiswiegmanni из подотряда двуходок, широко распространенного в тропиках Старого и Нового Света. Ведет она наземный и роющий образ жизни и похожа на пестро окрашенного дождевого червя.
Среди змей масса ужей Coluber и полозов Elaphe. К последним относится эскулапова змея, а также красивый леопардовый полоз (Elapheleopardina), из ядовитых — виперы, или настоящие гадюки (аспид овая, курносая, носатая, гюрза).
Из малочисленных амфибий, безусловно, наибольшего внимания заслуживают европейский протей — эндемик западной части Югославии — и пещерные саламандры рода Hydromantes, обитающие в Южной Европе. Они относятся к семейству, распространенному в основном в Америке.
Богат и красочен мир насекомых Средиземноморья. Среди бабочек много эндемиков, как локальных, так и более широко распространенных. Это кавалеры Papiliohospiton, Zerynthiarumina и Archonapollinus, белянки Pierisatlantica, Euchloebelimnia, Anthocharisbelia и Gonepteruscleopatra, нимфалида Charaxesjasius, сатиры Melanargialarissa, M. occitanica, M. ines, Satyrusactaea и Pyroniacecilia, а также замечательный коконопряд Изабеллы (Attacusisabellae) — локальный эндемик Испании. Немало эндемиков среди жуков: крупные жужелицы рода Carabus — С. gigas, С. splendes, C.rutilans и др., светлянки рода Luciola, своеобразные чернотелки, навозники и т. д. Следует назвать и эфиопских по происхождению термитов. Из паукообразных характерны также скорпионы, которых особенно много на африканском побережье Средиземного моря.
Средиземноморская подобласть, исходя их фаунистического своеобразия составляющих ее территорий, может быть разделена на провинции: Макаронезийскую (острова Азорские, Канарские, Зеленого Мыса и Мадейра), Тирренскую, или Западно-Средиземноморскую, и Восточно-Средиземноморскую, куда относятся Балканский полуостров, часть Малой Азии, горные районы Сирии и Ливана, остров Кипр. Естественно, что соответствующие провинции содержат кроме эндемичного ядра и свойственных всей Средиземноморской подобласти видов примесь эфиопских (на юго-западе) и сахаро-гобийских видов.
Сахаро-Гобийская подобласть.
Она занимает громадную территорию от атлантического побережья Сахары на западе до пустыни Алашань на востоке. Таким образом, в ее пределы входят: Северная и Центральная Сахара от Марокко до Египта, Синайский полуостров, Северная Аравия, пустыни Сирии, Палестины, Месопотамии, Средняя, Передняя и большая часть Центральной Азии, внутренняя часть Малой Азии, Восточное и Юго-Восточное Закавказье, северный и частично западный Прикаспий.
Ландшафты равнин, низкогорий, плоскогорий и отдельных хребтов носят здесь характер пустынь. Впрочем, в горных странах нередки и экстразональные растительные формации (даже острова леса в горах). Разнообразие природных особенностей и фауни-стические отличия требуют разделения подобласти на три над-провинции (группы провинций согласно ботанико-географическим рангам) — Сахаро-Синдскую, Ирано-Туранскую и Центрально-Азиатскую.
Климат подобласти характеризуется очень большой солнечной радиацией, крайне незначительным количеством осадков и очень жарким летом. Среднемесячная температура июля приблизительно 30 °С (25—35°С). Почти вся Сахаро-Синдская территория подобласти лежит в пределах пояса тропического воздуха, или пояса пассатов. Условия существования здесь чрезвычайно суровы. На большей части пустынь Северной Африки выпадает меньше 50 мм осадков в год, а в ряде районов дождей вообще не бывает в течение нескольких лет. Зима теплая: средняя температура января 10—15 "С, суточные колебания ее достигают в среднем 15—20°С и больше.
Климат Центральной Азии резко континентальный: суточная амплитуда температур 35 °С, а годовая — до 90 "С. Осадков меньше 100 мм в год, и выпадают они в большинстве пустынь летом. Зима холодная, среднеянварская температура от —16 до —18°С, лето жаркое — в июле 20—32 °С. Сухость воздуха очень велика.
В Ирано-Туранской части более благоприятный климат, зима и весна сравнительно мягкие, суточные и годовые колебания температур меньше, осадков свыше 100 мм в год, выпадают они преимущественно зимой и весной. Лето жаркое, среднеиюльская температура 24—27 °С. Осень теплая и сухая. Таким образом, в Ирано-Туранской надпровинции в основном средиземноморский тип климата.
Эти особенности климата, в частности его аридность, определяют преобладание в подобласти ксерофильных форм или обусловливают выработку у животных таких приспособлений, которые помогают избежать отрицательного воздействия жары и сухости, а в некоторых районах — и низких температур. Об этом уже говорилось при характеристике ландшафтных зон и основных биотопов суши.
Рассмотрим главные группы животного мира подобласти (рис. 147, 148) и попытаемся показать смену видового состава при переходе от одной надпровинции к другой.
Млекопитающие представлены прежде всего грызунами и копытными, играющими важную роль в местных биоценозах. В фауне Сахаро-Синдской надпровинции присутствует оригинальный род африканских сусликобелок (Xerus), живущих в норах, но похожих на белок. В Ирано-Туранской надпровинции распространен родственный сусликобелкам тонкопалый суслик (Sper-mophilopsisleptodactylus) — обитатель песчаных пустынь. Очень типичны для подобласти тушканчики, хорошо приспособленные к специфическим условиям пустынь и полупустынь. Это ночные животные, выходящие из нор с наступлением темноты. Зиму тушканчики проводят в спячке. Роль их в пустынных биоценозах велика. Они оказывают большое влияние на почвенный покров и растительность, служат пищей многим хищникам. С запада на восток происходит смена систематического состава: в Сахаре обитают представители только одного рода Jaculus (песчаные тушканчики), в Средней Азии и Иране к ним добавляются гребнепалые (Paradipus), толстохвостые (Pygerethmus) и мохноногие (Dipus) тушканчики, а также земляные зайцы (Allactaga). В Центральной Азии преобладают трехпалые карликовые тушканчики (Salpingotus), длинноухий (Euchoreutesnaso) и карликовый пятипалый (Cardiocraniusparadoxus) тушканчики и весьма распространенный тушканчик-прыгун (Allactagasaltator), заходящий в горы до высоты 2000 м.
Не менее часто в подобласти встречается подсемейство хомяковых — песчанки. Они напоминают крыс, но имеют опушенный хвост. Род Gerbillus (карликовые песчанки) живет в пустынях Северной Африки, Аравии и Передней Азии. В Сахаре это самые массовые млекопитающие. В тех же местах обитают и татэры, или голоподошвенные песчанки, — основной в Африке природный резервуар чумы. Только для североамериканских пустынь характерен род жирнохвостых (Pachyuromys) и малых (Psammomys) песчанок. Однако малые песчанки рода Meriones доходят до Средней и Центральной Азии, в то время как род Brachiones живет лишь в пустынях Центральной Азии. Подобные примеры систематического викариата в пустынях подобласти весьма типичны.

Рис. 147. Характерные представители фауны области Древнего Средиземья:
1 —
муфлон; 2 —
генетта; 3 —
лань; 4 —
белоголовый сип; 5 — жемчужная ящерица; 6 —
греческая черепаха; 7 — голубая сорока; 8 —
протей; 9 —
пещерная саламандра; 10 —
бабочка архон; 11 —
шелкопряд изабелла; 12 —
фенек

Рис. 148. Характерные представители фауны области Древнего Средиземья
(Сахаро-Гобийская подобласть):
I —
тонкопалый суслик; 2 —
большой тушканчик; 3 —
большая песчанка; 4 —
хаус; 5 —
снежный барс; 6 —
архар; 7 — джейран; 8 —
як; 9 —
саджа; 10 —
улар;
II —
круглоголовка; 12 —
гюрза; 13 —
лопатонос; 14 —
чернотелка стернодес
Подсемейство хомяков на западе подобласти вообще отсутствует. Но в Передней, Средней и Центральной Азии встречаются виды родов Phodopus (мохноногие хомячки) и Mesocricetus (средние хомячки). Представитель последнего — сирийский, или золотистый, хомячок — разводится в лабораториях и в домашних условиях.
Немало в подобласти хищных зверей. В густых зарослях камыша кое-где сохранился тигр (в Средней Азии он исчез в 1940-е годы); в горах Тянь-Шаня и Памиро-Алтая изредка попадается снежный барс, или ирбис (Pantherauncia); леопард распространен в Африке и южной половине Азии, иногда встречается в Средней Азии. Мелкие кошки Felis характерны для всей подобласти, особенно степная (F. lybica); барханный кот (F. margarita) и каракал (F. caracal) не заходят в центральноазиатскую часть, а хаус (F. chaus) — в сахарскую. Резко отличается от кошек гепард, встречающийся иногда во всех пустынях подобласти.
Виверровые, столь типичные для Палеотропического царства, на западе Сахаро-Гобийской подобласти представлены генеттой (Genettagenetta). Чрезвычайно интересен медоед (Mellivoracapensis). Это сравнительно крупное животное с черным брюшком и седой спиной. Питается он рептилиями, песчанками, насекомыми. Ареал его — Южная и Центральная Африка, Передняя и Средняя Азия до Индостана включительно, встречается в Туркмении и находится там под государственной охраной.
Своеобразно распространение лисиц (Vulpes) и их родственников — фенеков. Fennicuszerda живет в пустынях Северной Африки, Синайского и Аравийского полуостровов, ведет ночной образ жизни, способен зарываться в песок. Эта миниатюрная лисичка (1,5 кг) имеет очень длинные уши (более 15 см при общей высоте животного 30 см). В Иране, Афганистане и на северо-западе Индостана живет небольшая афганская лисица (Vulpes сапа). Лисица корсак (V. corsac) обитает в полупустынях и сухих степях от Ирана до Монголии.
Выходцем из Эфиопской области следует считать полосатую гиену (Hyaenahyaena), которая отсутствует только в центрально-азиатской части подобласти.
Группа копытных в подобласти довольно многочисленна. Антилопы, по меньшей мере 6 родов, представлены в Сахаро-Синд-ской части белым ориксом (Oryxgazellaleucoryx), саблерогой антилопой (О. dammah), аддаксом (Addaxnasomaculatus) и 3 видами газелей (Gazella). В Ирано-Туранской части живет джейран (Gazellasubgutturosa), а в Центральной Азии кроме него — монгольский (Procapragutturosa) и тибетский (Р. picticaudata) дзерены, оронго (Pantholopushodgsoni), такин (Budorcastaxicolor) и сайгак (Saigatatarica).
Горные козлы и бараны распространены по горным системам от Северной Африки и Аравии до Тибета: на западе — нубийский козел (Capranubiana), в Ирано-Туранской надпровинции — безоаровый (С. aegagrus), винторогий (С. falconeri) и сибирский (С. sibirica) козел, или козерог (последний заходит в центрально-азиатскую часть). Гривистый баран (Ammotraguslervia) живет в Северной Африке, различные подвиды горного барана (Ovisammon) — так называемые муфлоны, архары и аргали — в Передней, Средней и Центральной Азии. Из быков встречается лишь тибетский як.
Орнитофауну равнин и гор Сахаро-Гобийской подобласти лучше охарактеризовать раздельно.
Типичными для пустынь птицами считаются саксаульные сойки, 2 вида которых населяют пустыни Средней Азии (Podocespanderi) и Монголии (Р. hendersoni). Будучи близкими родичами лесной птицы кедровки, саксаульные сойки приобрели ряд адаптации, позволивших им освоить новую экологическую нишу. К жизни в пустынях хорошо приспособлены также многие рябки (Pterocles), но особенно саджи, или копытки. В пустынях и полупустынях Средней и Центральной Азии живет обыкновенная саджа (Syrrhaptesparadoxus). Свое второе название — копытка — птица получила благодаря сращению оперенных пальцев ног, в результате чего образовалось подобие заячьей лапы. Толстые кожистые подошвы предохраняют ноги от ожогов при бегании по раскаленной почве в пустыне. В Тибете и на Восточном Памире, т. е. уже в Центральной Азии, водится горная саджа, придерживающаяся горных пустынь. Встречаются в пустынях и дрофы, особенно вихляй, или джек (Chlamydotisundulata), ареал которого охватывает все равнинные части подобласти, а также куропатки, из которых пустынная (Ammoperdixgriseogularis) обитает в Передней и на юге Средней Азии. Много на равнинах и жаворонков (малый, хохлатый и др.). Из других воробьиных «пустынниками» являются пустынный сорокопут, славка-скотоцерка, саксаульный воробей, пустынная каменка.
Особую группу биотопов на равнинах Сахаро-Гобийской подобласти представляют оазисы, тугаи и орошаемые участки. Наличие здесь древесно-кустарниковых насаждений и лучшие микроклиматические условия привлекают самых различных животных, и птицы среди них играют далеко не последнюю роль. Это прежде всего заметно в тугаях — ленточных лесах, располагающихся вдоль рек, текущих в пустынях. Тут встречаются фазаны (подвиды колхидского фазана), испанский воробей, тонкоклювая камышовка, большая бухарская синица, зимородок, изредка орел-змееяд. Большая часть птиц, населяющих тугаи, по крайней мере в среднеазиатской части подобласти, относится к широко распространенным. По ленточным лесам — своеобразным «экологическим желобам» — происходит расселение птиц с юга на север. Это касается великолепной райской мухоловки (Terpsiphoneparadisi) и синего дрозда, или синей птицы (Myophonuscoeruleus), из Индии. В более отдаленном прошлом такими путями пользовался розовый скворец (Pastorroseus). Многие из тугайных птиц переселяются в оазисы и становятся спутниками человека. Белый аист и египетская горлица (Steptopeliasenegalensis), например, обитают только в населенных пунктах африканской и среднеазиатской частей подобласти.
Очень большим своеобразием отличается горная орнитофауна Сахаро-Гобийской подобласти, особенно ее альпийские районы, лучше выраженные на востоке. Характерными высокогорными птицами из отряда куриных являются улары (Tetraogallus). В Передней Азии это каспийский улар (Т. caspicus). В том числе он встречается в горах Малого Кавказа и Копет-Дага. В горах Средней Азии живут еще 2 вида — гималайский (Т. himalayensis) и тибетский (Т. tibe-tanus) улары, распространенные и в хребтах Центральной Азии. Улары — крупные птицы, легко и ловко бегающие по крутым склонам. В полете они громко свистят. Улары не пьют воду, а ограничиваются влагой растений. На Памире оба вида уларов живут на высоте свыше 4000 м. В многоснежные зимы улары пасутся вместе со стадами козерогов и архаров, собирая семена на раскопанных ими участках. Птенцы уларов легко приручаются, и в кишлаках Бадахшана и Памира часто встречаются вполне крупные взрослые улары, выращенные в неволе.
Обильны в подобласти клушицы (Pyrrhocoraxpyrrhocorax), населяющие альпийскую зону, а также альпийские галки (Pyrrhocoraxgraculus), которые отсутствуют только в Северной Африке. Хищные птицы представлены черным грифом, типичным для высокогорий, белоголовым сипом, не заходящим в Центральную Азию, снежным грифом, или кумаем (Gypshimalayensis), обитателем Средней и Центральной Азии, кое-где к нему присоединяется бородач, или ягнятник (Gypaetusbarbatus), единственный гриф с оперенной головой и шеей. Среди воробьиных выделяются вьюрки — альпийский (Montifringillanivalis) и горный (Leucosticte), характерные для Средней и Центральной Азии.
Водоемы альпийской зоны привлекают на период гнездования оригинального индийского гуся (Anserindicus). Ареал его охватывает Среднюю и Центральную Азию, где высоко в горах гнездится также кулик-серпоклюв (Ibidorhynchastruthersi) — единственный представитель особого подсемейства куликов. Он хорошо плавает. Из-за хорошей способности птиц к расселению местный эндемизм их выражен гораздо слабее, чем в фауне зверей.
Рептилии в Сахаро-Гобийской подобласти представлены более богато, чем в других палеарктических подобластях. (Причины этого указывались при описании Средиземноморья.) Главным образом здесь распространены ящерицы и змеи. Отметим самых характерных из них.
Семейство гекконов, распространенное на всех материках, в Сахаро-Гобийской подобласти представлено большим количеством видов из разных родов. Из примитивного подсемейства зублефаров в Передней Азии обитает иранский зублефар (Eublepharisangamainyu). Сцинковые гекконы (Teratoscincus) живут в пустынях Средней и Центральной Азии, особенно в барханных песках. Для среднеазиатских песчаных пустынь типичен гребнепалый гек-кон (Crossobamon). В пустынях Сахаро-Синдской надпровинции распространены тонкопалые гекконы (Stenodactylus), а виды гек-кончиков (Alsophylax) и голопалых гекконов (Gymnodactylus) встречаются по всей подобласти. Широко распространены в подобласти и агамы. В отличие от гекконов их нет в Западном полушарии. К пустыням и полупустыням приурочены 2 рода — агамы и круглоголовки. Настоящих агам больше всего в Африке, но викарирующие виды сменяют друг друга на всем протяжении подобласти — от Сахары до Центральной Азии. Круглоголовки (Phrynocephalus), отличающиеся оригинальной способностью закручивать колечком кончик хвоста, встречаются от Восточного Закавказья до Северо-Западного Китая. В песчаных пустынях Средней Азии обитает крупная ушастая круглоголовка. Пожалуй, наиболее интересны из агамовых так называемые шипохвосты (Uromastix), населяющие сухие пустынные и скалистые местности Северной Африки, Юго-Западной и Центральной Азии. В Сахаре они обитают на окраинах оазисов и среди холмов на границе с песчаной пустыней. Хвост у этих ящериц короткий, плоский и покрыт сверху большими колючими чешуями. Крупные виды достигают 75 см в длину.
Есть в подобласти и представители семейства сцинков. Это ма-буи (золотистая мабуя распространена в Аравии, Передней и Малой Азии, а также на юге Средней Азии), гологлазы (Ablepharus) со сросшимися, как у змей, веками в виде прозрачного окошка, змееящерицы (Ophiomorus) с редуцированными конечностями, настоящие сцинки (Scincus) и др. Из настоящих ящериц в подобласти широко распространен род Eremias (ящурки), хотя есть и другие роды — змееголовки (Ophisops) из Северной Африки, Малой и Передней Азии, гребнепалые ящерицы (Acanthodactylus). Ящерицы, живущие в песках, имеют своеобразные приспособления для передвижения по рыхлому субстрату или же закапывания в песок — пальцы у них снабжены роговыми чешуйками и выростами, увеличивающими площадь. В Передней и Средней Азии по речным долинам и в предгорьях живет крупная безногая ящерица желтопузик. Есть она, как уже отмечалось, и в Средиземноморской подобласти. Один род семейства варанов представлен крупным серым вараном (Varanusgriseus). Ареал его охватывает всю подобласть, за исключением Центральной Азии.
Змеи подобласти не менее разнообразны. Это семейства слепунов (единственный вид подобласти — обыкновенная слепо-змейка — встречается и на юге Средней Азии), узкоротых змей (в Передней Азии), удавов (представители рода удавчиков — Егух), ужеобразных, аспидовых, гадюковых и ямкоголовых. Среди уже-образных выделяется род полозов Coluber. Одни виды его широко распространены в подобласти — разноцветный (С. ravergieri) и пятнистый (С. tyria), другие имеют малые ареалы, приуроченные в основном к Сахаро-Синдской (С. rhodorachis) или к Ирано-Ту-ранской (С. karelini) надпровинции. В пустынях Северной Африки и Передней Азии встречаются остромордые ужи, или литоринхи (Lythorhynchus), ведущие полуроющий образ жизни. Из них венценосный литоринх живет только в Сахаре и Ливийской пустыне, а афганский — в Передней Азии. Из других ужеобразных отметим еще род Psammophis, к которому относится знаменитая стрела-змея (Р. lineolatus) — житель песчаных пустынь Ирано-Туранской и Центрально-Азиатской надпровинции. Это стремительно передвигающееся животное питается только ящерицами, а молодые особи — насекомыми. Аспидовые представлены кобрами (Naja) — индийской (среднеазиатский подвид без очкового рисунка) и египетской, а гадюковые — рядом видов, среди которых наиболее известны степная гадюка, гюрза, рогатые гадюки и эфы. Рогатые гадюки (Cerastes) и эфы (Echis) передвигаются «боковым ходом»: задняя половина туловища забрасывается вперед и вбок, а затем к ней подтягивается передняя. При этом змея опирается только на концы туловища и на песке остается след в виде косых полосок. Гадюки и эфы способны зарываться в песок и издавать «шипение» трением боковых чешуек тела. Виды рода Cerastesживут лишь в Сахаре и на Аравийском полуострове, Echis — в Северной Африке, Передней Азии, Индии. Ямкоголовые змеи представлены щитомордником обыкновенным. Ареал его захватывает также часть Средней и Центральной Азии.
Черепах в подобласти мало. Из водяных черепах каспийская (Clemmyscaspica) обитает от Северо-Западной Африки до Туркмении, болотные черепахи (Emys) с единственным видом Е, orbi-cularis — в Северо-Западной Африке, Европе, Турции, Северном Иране, на Кавказе и западе Туркмении. Встречаются и сухопутные черепахи (Testudinidae), в пустынях Северной Африки живут 2 вида — египетская и средиземноморская. Последняя распространена до Ирана и Азербайджана. В Ирано-Туранской надпровинции обычна степная черепаха (Testudohorsfieldi).MHco ее вполне съедобно. Вместе с тем, поедая культурные растения, она иногда наносит заметный ущерб. Сухопутные черепахи подобласти рас-тительноядны. Весной они ведут активный образ жизни, а затем впадают в спячку.
Состав амфибий в Сахаро-Гобийской подобласти довольно беден. Из хвостатых в Центрально-Азиатской надпровинции живут только углозубы (Hynobius) — сибирский и семиреченский. Саламандра заходит лишь на запад Малой Азии. Там же попадается и малоазиатский тритон (Triturusvittatus). Бесхвостые представлены лягушками и жабами, из которых наиболее обычна серая жаба. В Северо-Восточной Африке живет нильская Лягушка, ведущая водный образ жизни.
Среди пресноводных рыб выделяются своеобразные осетровые — аральские лопатоносы (Pseudoscaphirhynchus), эндемики Амударьи и Сырдарьи. Всего их 3 вида. Из лососей есть аральский лосось, нерестящийся в Амударье, ручьевая форель (в верхнем течении Амударьи), хариусы (в Монголии). В нижнем течении Нила, т. е. в Северо-Западной Африке, обитает единственный представитель особого семейства — гимнарх (Gymnarchusniloticus), способный испускать электрические импульсы. Это явный экзот в палеарктической фауне рыб. Для рек и озер подобласти типичны карповые рыбы — плотва, кутум, ельцы (промысловое значение имеют чебаки), алтайские османы (в Центральной Азии), пескари (Gobio), усачи (Barbus) и храмули (Varicorhinus), средне- и центральноазиатские маринки (Schizothorax) и османы (Dipty-chus). В Центральной Азии, реже в других районах восточной и центральной частей подобласти многочисленны гольцы (Nema-chilus). Некоторые из них живут только в высокогорных водоемах, например тибетский голец (N. stoliczkai). Нередки в подобласти и сомы. Так, верховья Средней и Центральной Азии населяет тибетский сомик (Glyptosternumreticulatum), водоемы Сахары — сомы семейства Clariidae. По всей Сахаро-Синдской надпровинции распространены рыбки семейства карпозубых (Cyprinodon-tidae). Только там встречаются представители тропического семейства цихлид.
Многочисленны и разнообразны насекомые Сахаро-Гобийской подобласти. Вкратце их охарактеризовать нелегко, остановимся лишь на некоторых самых распространенных группах.
Для аридных территорий подобласти, включая и полупустыни, чрезвычайно типично обилие жуков-чернотелок (Tenebrionidae). По всей подобласти обитают крупные Pimelia, роды Pachyscelis, Catomus, в песках живут Sphenaria. На западе подобласти из чернотелок очень характерен род Mesostena, а также Asida и Erodius. Фауна Ирано-Туранской надпровинции отличается наличием эн-демичных родов и даже своеобразного эндемичного подсемейства Kuhitangiinae с единственным родом и видом Kuhitangiakryzhanovskii. В пустынях Турана большинство чернотелок составляют эндемичные виды, относящиеся к родам Stemodes, Pisterotarsa, Trigonoscelis и др. В Центральной Азии есть свои эндемики, в основном обитатели пустынных плоскогорий (Colposcelis, Anotalica, Platyope, Platyscelis и др.).
Подобная картина распространения свойственна большинству хрущей, листоедов, жужелиц и других семейств жуков. Из наиболее характерных представителей в песчаных пустынях живут жуки-хрущи из родов Pharaonus, Chioneosoma (белые хрущи), Achranoxia, очень своеобразны Dynamopus и Thynorycter — все с сильно развитыми морфологическими адаптациями к среде. Пустынные предгорья и низкогорья населены многочисленными жуками-красавчиками Lethrus, из которых только в Средней Азии 60 эндемичных видов. Чрезвычайно характерны также представители древнесре-диземноморского рода листоедов Thelyterotarsus, Oreomela, Xeno-mela и Stemoplatus.
В горах Сахаро-Гобийской подобласти мир насекомых очень богат и высокоэндемичен. Это бабочки аполлоны (Pamassius), сатиры Hyponophele и Chazara, саранчовые и кузнечики Gom-phomastax и Conophyma и др. Субтропический характер биоцено-зов подобласти подчеркивается наличием термитов, палочников, скорпионов, сольпуг и т. д.
Зоогеографическое деление Сахаро-Гобийской подобласти до настоящего времени не завершено и достаточно спорно — за основу берутся фаунистические центры, а количество их, согласно разным авторам, различно.
Область включает юго-восток в нетропической части азиатского материка: Приморье, Северный и Средний Китай, Корею, Японию (кроме северного острова Хоккайдо), Восточный Тибет и восточную часть южного склона Гималаев. Граница области не везде отчетлива, особенно на севере и юго-востоке, в то время как на западе она вполне четко идет по хребту Большой Хинган.
Природные условия области вкратце сводятся к следующему. Климат здесь теплый или умеренно теплый, выпадает много осадков, особенно летом. Смягчающее и увлажняющее влияние океана чувствуется почти везде. Основной растительной формацией является лиственный лес, снова появляющийся на Дальнем Востоке после перерыва в Сибири. Южные районы области расположены в зоне вечнозеленого леса. Большую территорию области занимают горы, причем ряд хребтов имеет хорошо выраженную альпийскую зону. Естественно, что в таких районах распределение климата и растительности подчиняется правилу вертикальной зональности.
Животный мир области очень разнообразен (рис. 149).

Рис. 149. Характерные представители фауны Восточно-Азиатской области:
1- пятнистый олень; 2-горал; 3-большая панда; 4-такин; 5-восточный широкорот; 6-гималайский медведь; 7-монал; 8-амурский полоз; 10-махаон маака; 11 — реликтовый усач; 12 — мандаринка ' Это объясняется географическим положением — область граничит с тропиками Юго-Восточной Азии. Примесь индо-малайских форм местами весьма значительна. Кроме того, здесь можно встретить и американские группы, расселившиеся в период материкового соединения на севере. Известную «экзотичность» придают фауне мар-тышкообразные обезьяны — макаки и тонкотелы. Первые представлены в горах Юго-Восточного Китая медвежьим макаком, а в Японии — японским (Macacusfuscata). Медвежий макак населяет не только равнинные леса, но и горные районы с суровыми зимами, где ему приходится в поисках пищи раскапывать снег. В хвойных и бамбуковых лесах Западного Китая на высоте до 3000 м можно встретить гималайских тонкотелов (Rhinopithecus). Только после установления в горах снежного покрова обезьяны спускаются в долины и кормятся в садах и на огородах.
Восточно-Азиатская область нередко рассматривается как возможный центр происхождения оленей. Действительно, в ней есть разные их группы — от мелких Elaphodus с торчащими клыками и водяных оленей эндемичного в Китае рода Hydropotes до крупных пятнистых (Cervusnippon) и беломордого оленя (С. albirostris), открытого Пржевальским в Тибете. Крупный олень Давида, или Милу (Elaphurusdavidianus), сохранился в настоящее время только в неволе, хотя в исторические времена был очень многочислен в Северо-Восточном Китае. В горах Западного Китая обитают оригинальные копытные из подсемейства козлов. Внешне они напоминают мускусного быка или некоторых антилоп. Речь идет о горалах (Nemorhaedusgoral), cepay (Capricomissumatraensis) и такинах (Budorcastaxicolor). Впрочем, особый подвид cepay живет в Японии. Обычна в области также косуля.
Насекомоядные включают большое число эндемиков. Это своеобразные виды семейства гимнуровых — китайский гимнур (Neo-tetracussinensis), живущий в горах Сычуани, кротовые из рода могера — большая (Mogerarobusta) и малая (М. wogura) могеры, распространенные в Китае, Японии и у нас в Приморье, а также оригинальный сычуаньский землеройкокрот (Uropsyllussoricipes).
Из грызунов распространены несколько видов пищух, энде-мичные южноазиатские полевки Eothenomys и крысовидные хомячки (Cricetulus). Индо-малайский элемент в фауне представляют белки рода Dremomys. Обитает в Японии и крайне примитивный древесный заяц (Pentolagusfurnessi).
Хищников в области много, и среди них кроме широко распространенных волков и лисиц немало оригинальных форм. В первую очередь это енотовидная собака (Nyctereutesprocyonoides), обитающая в лесах Юго-Восточной Азии. Она несколько напоминает американского енота-полоскуна (с «маской» на морде). Этот практически всеядный зверь неоднократно акклиматизировался в Европе, в том числе и в европейской части России. В горах живет красный волк (Cuonalpinus) из восточноазиатского рода. Медведи имеют несколько представителей. Из них наиболее типичен гималайский, или черный, медведь (Ursusthibetanus).
Оригинальны восточноазиатские панды, близкие к енотам, распространенным в Америке. Ареал малой панды (Ailurusfulgens) выходит за пределы области на юге, но родина этого горного вида — Юго-Западный Китай. Панда активна ночью, а днем спит на деревьях, кормится побегами бамбука, травой, фруктами, иногда поедает яйца и птенцов. Большая панда (Ailuropodame-lanoleuca) похожа на медвежонка, имеет массивное тело с короткими широкими ногами. Подушечки на пальцах и голая подошва помогают ей удерживаться на гладких стеблях бамбука. От медведей панду отличает довольно длинный хвост. Окраска большой панды белая с черными пятнами, вокруг глаз так называемые очки. Из-за скрытого образа жизни она изучена слабо, внесена в Красную книгу. По новым данным большую панду следует относить к отдельному семейству Ailurinidae, занимающему промежуточное положение между медведями и енотами. Таким образом, в фауне млекопитающих Восточно-Азиатской области есть эндемик высокого ранга.
Из других хищников назовем тигра, дымчатого леопарда, снежного барса, крупную куницу харзу. Индо-малайский элемент представляет здесь трехполосная циветта (Arctogalidiatrivirgata) из виверровых.
В богатую орнитофауну Восточно-Азиатской области входят разнообразные фазаны из родов Phasianus, Chrysolophus (золотой и алмазный), Syrmaticus (королевский), Tragopan (рогатый фазан) и Lophophoms (монал). Фазаны различных родов и видов в основном принадлежат китайской горной фауне. Своеобразны журавли, считающиеся в ряде районов священными птицами. Японский журавль (Grusjaponensis) еще держится в нашем Приморье и Приханкайской низменности, но в самой Японии он крайне редок. Этот вид занесен в Красную книгу. На грани исчезновения находится и красноногий ибис (Nipponianippon). Аисты представлены эндемичным Ciconiaboyciana, а утки — очень красивой мандаринкой (Aixgalericulata).
Много в области выходцев из соседней Индо-Малайской области. Это многочисленные зимородки из родов Halcuon, Ceryle и Alcedo, стрижи рода Chaetura (иглохвосты), широкороты (Eury-stomus) из сизоворонковых, белоглазки и даже птицы бюль-бюли и рыбный филин (Ketupa).
Типично палеарктические птицы включают самостоятельные виды. Перечислить их из-за большого разнообразия невозможно. Отметим только, что уже упоминавшаяся при описании фауны Средиземноморья голубая сорока после громадного перерыва снова появляется в Приморье, Китае и Японии.
Среди рептилий обращают на себя внимание трехкоготные черепахи из рода Trionyx (например, съедобный китайский трионикс).
Из змей обычны и разнообразны полозы рода Elaphe (амурский, красноспинный, тонкохвостый, японский и др.), почти эндемичны виды рода Dinodon, попадающиеся и в Приморье, и на Южном Сахалине, а в Японии есть ксенодерм. Аспидовые змеи отсутствуют. Гадюковые представлены крайне примитивной бирманской гадюкой (Azemiopsfeae) с короткими ядовитыми зубами и красивой, но смертельно ядовитой цепочной гадюкой (Viperarusseli), заходящей из Индии и Южного Китая. Из ямкоголовых змей обычен щитомордник восточный (Ancistrodonblomhoffi).
Среди ящериц замечательны лесные долгохвостики (Tachy-dromus), отличающиеся чрезвычайно длинным хвостом, которым они могут цепляться за ветки деревьев и в таком состоянии проводить целые ночи. Из гекконов и сцинков здесь встречаются несколько видов.
Среди амфибий наибольший интерес представляют хвостатые. Когтистые тритоны рода Onychodactylus, распространенные в Китае, Корее, Японии, и заходящие к нам в Приморье О. fischeri, снабжены роговыми когтями, что не свойственно амфибиям. У нашего вида нет легких, и дышит он через кожу. Исполинская саламандра (Megalobatrachusjaponicus), живущая в горных реках Японии и Восточного Китая, считается самым крупным современным земноводным (достигает 160 см). Мясо ее съедобно и употребляется в пищу, поэтому саламандр активно истребляют, и вид стал очень редким. Среди лягушек и жаб есть эндемичные виды.
Пресноводные рыбы на севере области представлены викарными видами палеарктических родов (горчак, калуга, китайский сом и др.), но уже в Китае ихтиофауна приобретает своеобразные черты. К ней относятся обитающие и в бассейне Амура касатко-вые сомы (Bagridae), опасные из-за выделяемой ими ядовитой слизи и острых колючек. Из карповых, достигающих в Юго-Восточной Азии особого разнообразия, в первую очередь следует упомянуть серебряного карася. Эта обычная в Китае рыба послужила материалом для селекции декоративных «золотых рыбок» (вуалехвостов, телескопов, комет и др.). Промысловое .значение имеют черный амур (Mylopharyngodonpiceus) и белый амур (Ctenopha-rynodonidella), близкие к усачам субэндемичные кони (Hemiba-rbus), желтоперы (Plagiognathops), белые и черные лещи, хищные желтощеки (Elophichthysbambusa) и целый ряд других. Среди мелких рыб многие обладают красивой расцветкой и интересным поведением, за что их охотно держат в аквариумах. Таковы лабиринтовые и некоторые карпозубые.
Насекомые Восточно-Азиатской области интересны не только из-за эндемизма многих из них, но и главным образом благодаря их фаунистическим связям. Здешняя форма насекомых включает много третичных реликтов. Только здесь встречается род Cupes из семейства жуков Cupedidae, представители которого живут и в тропиках, и в Северной Америке. Только здесь обитают семейства Cephaloidae (с североамериканскими связями) и Helotidae (с индо-малайскими связями). Представителей палеотропических или индо-малайских семейств, проникающих в Палеарктику через Восточно-Азиатскую подобласть, немало. Эндемичны для подобласти крупные и очень красивые металлически окрашенные жуки-брызгуны рода Carabus из подрода Coptolabus, к которому относятся С. ignimetalla и С. pustulifer из Китая, С. gehini из Японии, С. соnstricticollis из Кореи и Приморья. Самым интересным жуком считается громадный реликтовый усач (Callipogonrelictus), относящийся к роду, распространенному в Центральной и Южной Америке.
Из бабочек отметим кавалеров-хвостоносцев Papiliohelenus, Р. bianor явно тропического облика. Для многих шмелей (Bombussapporoensis, В. ussuriensis) характерны ареалы, уходящие в южные районы Восточной Азии.
Хотя энтомофауна Китая, Японии и нашего Приморья изучена далеко не полностью, можно полагать, что не менее 50 % ее видов эндемичны.
К этому региону относится та часть Северной Америки, которая лежит севернее тропика, острова Алеутские и Ванкувер (в Тихом океане), Ньюфаундленд, Гренландия и Бермудские (в Атлантике). Южная граница полцарства является одновременно северной границей Неогейского царства.
Рельеф Северной Америки определяется меридиональным хребтом Кордильер, тянущимся от Аляски до Панамского перешейка. На распределении животных сказывается дивергенция Кордильер, начинающаяся с широты 52°, в результате которой образуются два хребта: восточный, именуемый Скалистыми горами, и западный, названный Сьерра-Невада. Между ними располагается плоскогорье Большого бассейна, изрезанное каньонами. На востоке материка возвышаются горы Аппалачи. К северо-западу от них находится котловина Великих озер (Верхнее, Мичиган, Гурон, Эри и Онтарио).
В связи с громадной протяженностью материка с севера на юг в Северной Америке встречаются все типы климатов, кроме тропического. Климат тихоокеанского побережья мягче, чем атлантического.
По характеру растительности выделяются зоны тундры, тайги, широколиственных лесов, прерий. Субтропические хвойно-широколиственные леса простираются вдоль тихоокеанского побережья на западных склонах Сьерра-Невады и в Калифорнии. Субтропические пустыни занимают Мексиканское нагорье и прилежащие районы к северу от него. Это страна кактусов. Растительный покров здесь сильно нарушен деятельностью человека, в особенности в зоне широколиственных лесов на востоке и в прериях. Нетронутая природа в большинстве случаев сохранилась в национальных парках США и Канады.
Наиболее разнообразная фауна, в частности позвоночных, сосредоточена в южных районах, где самые благоприятные условия обитания. К северу это разнообразие постепенно, хотя и неравномерно, уменьшается до тех пор, пока не ограничится очень обедненной северной фауной, сходной с таковой в Северной Пале-арктике. На юге фауна Неарктики образует сложный переход к неотропической фауне (рис. 150).
Млекопитающие Северной Америки представлены 3 эндемичными и большим количеством широко распространенных семейств. Из сумчатых присутствует американский опоссум (Didelphis), распространенный от Южной Канады до севера Южной Америки. Насекомоядные очень своеобразны. Это многочисленные кротовые из родов, не встречающихся в Старом Свете: Scapanus, Scalopus, Neurotrichus и др. Причудливый звездонос (Condyluracristata) имеет на рыльце 22 мягких мясистых подвижных голых луча.

Рис. 150. Характерные представители фауны Неарктики
(млекопитающие и птицы):
1 —
овцебык; 2 —
снежная коза; 3 —
скунс; 4 —
поркупин; 5 — звездонос; 6
опоссум; 7 — вилорогая антилопа; 8 —
луговая собачка; 9 —
кардинал; 10
белоголовый орлан; 11 —
американский журавль; 12 —
луговой тетерев; 13
кондор
Среди грызунов выделяются эндемичные семейства гоферовых (Geomyidae) и аплод оптовых (Aplodontidae). Много зайцев, в том числе обычный беляк, несколько видов диких кроликов Sylvilagus, среди которых водяной и болотный хорошо плавают и даже ныряют. Один вид пищух населяет северо-запад страны. Мешотчатопры-гуновые (Heteromyidae) содержат ряд эндемичных родов из юго-западных штатов США. Это семейство распространено до северных частей Южной Америки. Беличьи представлены сурками, эндемич-ными луговыми собачками (Cynomys), сусликами и эндемичными родами антилоповых сусликов (Ammospermophilus) и бурундуковых белок (Tamiasciurus). Настоящих белок также много, а летяги представлены эндемичным родом Glaucomys. Немало эндемиков и среди мышовок, хомяковых, полевковых. Выделяется ряд белено-гих хомячков, или оленьих мышей, как их называют американцы.
Хищники, кроме волка и лисицы, населяющих и Евразию, имеют своих представителей из семейства собачьих. Это эндемич-ный рыжий волк, американские карликовые лисицы (из них Vulpesmacrotis отличается громадными ушами) и эндемичный род серых лисиц (Urocyon), ведущих полудревесный образ жизни. Медведи представлены крупным бурым медведем (гризли) и барибалом (Euarctosamericanus), близким к гималайскому. Из енотов типичен полоскун, акклиматизированный и в Старом Свете. Многочисленны куньи. Среди них выделяются голаркты — ласка, горностай, росомаха, эндемичный американский барсук (Taxidea) — и оригинальные скунсы, или вонючки (Mephitis). Из кошек можно назвать пуму (хищник панамериканского распространения), заходящую из Южной Америки ягуарунди и голарктическую рысь.
Копытные представлены эндемичным семейством вилорогов (Antilocapridae), населяющим прерии запада Северной Америки. Из оленей распространены уже известные нам северный олень, или карибу, благородный олень и ряд видов американских оленей Odocoileus. Лось представлен очень крупным подвидом, а быки — бизоном. Палеарктических оленей заменяют пекари.
Ежи, крысы, настоящие мыши отсутствуют. Непарнокопытных тоже нет, хотя первые этапы эволюции лошадей происходили в Новом Свете.
Трудно в краткой форме охарактеризовать орнитофауну Неарктики. Среди водоплавающих и околоводных птиц процент голарктов особенно велик. Таковы гагары и поганки, утки (в том числе кряква, широконоска, шилохвость и др.), гаги. Среди гусей встречаются эндемики: американский лебедь, канадская казарка, очень красивая каролинская утка, заменяющая здесь мандаринку и относящаяся к тому же роду Aix. Среди дневных хищников, с одной стороны, встречаются роды и виды, общие и для Палеарк-тики (беркут, ястреб-перепелятник, кречет), а с другой — эндемичные: белоголовый орлан (Haliaeetusleucocephalus), коршун -слизнеед (Rosthramussociabilis) и вилохвостый коршун (Elanoidesforflcatus). Вместо стервятников в Северной Америке распространены грифы-индейки (Cathartes), а на юго-западе даже калифорнийский кондор (Gymnogypscalifornianus).
Из тетеревиных здесь обитает обыкновенная индейка (Meleagrisgallopavo), родоначальник домашних индеек. Живут и американские кукушки (Coccyzus, Geococcys). Они интересны тем, что сами насиживают яйца, а не паразитируют, как европейская кукушка. Дятлы представлены яркими и красивыми видами (золотой, медный).
Воробьиные крайне разнообразны и красочны из-за обилия южно-американских семейств. Этотирановые (Tyrannidae), котинги, пересмешники (Mimidae), древесницы (Parulidae), трупиалы (Icteridae). На севере полцарства много палеарктических видов: свиристели, крапивники, корольки, пищухи, чечетки и др. Домового воробья завезли сюда из Европы. Обращает на себя внимание отсутствие скворцов.
На крайнем юге к неарктической орнитофауне присоединяются экзотические колибри и попугаи.
Фауна рептилий отличается полным отсутствием настоящих ящериц и гадюковых змей. Место агам занимают игуановые — анолисы (Anolis) и фринозомы (Phrynosoma). Амфисбены, которых много в Южной Америке, в Северной Америке также имеют своего представителя — пятнистую двуходку, обитающую в гнездах муравьев и термитов. Эндемичное семейство ядозубов (Нею-dermatidae) объединяет всего 2 вида североамериканских ядовитых ящериц, единственных ящериц в мире, обладающих настоящими ядовитыми железами. Укус ядозуба опасен и для человека. Обитающие здесь безногие ящерицы семейства Anniellidae также эндемичны (рис. 151).
Среди змей подавляющее большинство относится к американским семействам и родам. Наиболее характерны ямкоголовые — щитомордники (Ancistrodon) и гремучие змеи (Sistrurus и Crotalus) (последние отличаются наличием «трещотки» на конце хвоста). Все они очень ядовиты и опасны для домашних животных и человека. Аспидовых мало, причем кобры вообще отсутствуют. Удавы (калифорнийский, резиновая змея), обитающие в этом подцар-стве, небольших размеров.
Несколько неожиданным кажется разнообразие черепах. Встречаются представители семейств каймановых (Chelydridae), пресноводных, сухопутных и трехкоготных черепах. Из сухопутных наибольшее значение имеют так называемые гоферовые черепахи (Gopherus), живущие на юге США. Черепаха-гофер, населяющая сухие песчаные местности, роет длинные (до 12м) норы. В этих норах селятся также лягушки, змеи, кролики, опоссумы и даже еноты. Мясо гофера съедобно и вкусно.
В бассейне Миссисипи живет аллигатор — близкий родственник китайского аллигатора.

Рис. 151. Характерные представители фауны Неарктики (рептилии и амфибии):
1 —
гремучая змея; 2 —
ядозуб; 3 —
миссисипский аллигатор; 4 —
каймановая черепаха; 5 — лягушка-бык; 6 —
полосатый сирен
Амфибии замечательны большим, чем в других частях Северного полушария, обилием хвостатых. Это скрытожаберники (Сгурtobranchus), амбистомы, сиреневые (Sirenidae), настоящие и безлегочные саламандры и, наконец, эндемичные лягушки Ascaphustruei, относящиеся к семейству гладконогих, или лиопельмид. Кроме того, есть представители чесночниц, жаб, квакш и настоящих лягушек, среди которых лягушка-бык достигает больших размеров (до 20 см в длину).
Пресноводные рыбы Неарктики включают эндемичный род отряда амиеобразных, или ильных, рыб (Amiiformes) с одним семейством Amiidae и видом Amiacalva. Это одно их самых примитивных семейств среди ганоидных рыб. Осетровые, населяющие воды Северного полушария, представлены осетрами (атлантическими, тупорылыми, озерными, белыми) и американскими лопатоносами (Scaphirhynchus) — близкими родичами аральских лопатоносов Палеарктики. Семейство веслоносых (Polyodontidae) имеет одного представителя в бассейне Миссисипи (веслонос Polyodon), а другого в Китае (Psephurus). Разрывы ареалов подобного типа характерны и для других животных.
Неарктике свойственны рыбы монотипичного субэндемично-го отряда панцирникообразных (Lepidosteiformes) с одним семейством каймановых рыб, или панцирников, известных с мелового периода. Наиболее обычна панцирная щука (Lepidosteusosseus). Лососи и сиги распространены в основном на севере, как и в Палеарктике. Из настоящих костистых рыб отряда карпообразных есть чукучаны (Catostomidae), обитают они и на северо-востоке Сибири, и в Китае (по одному роду).
На крайнем севере Неарктики живет далия (Dallia), которую можно встретить и на северо-востоке Азии. В бассейнах Великих озер и Миссисипи обитает еще один замечательный эндемик — афредодер из эндемичного семейства Aphredoderidae. Наконец, совершенно необычно выглядят слепоглазковые (Amblyopsidae) — тоже эндемичное семейство с 3 родами и 5 видами. Живут они в подземных водах карстовых пещер средней части Северной Америки и лишены зрения.
Некоторые беспозвоночные Неарктики указывают на родство ее фауны с палеарктической, другие же весьма своеобразны или связаны с фауной Неотропики. Для пресноводных моллюсков характерно семейство Unionidae с несколькими сотнями видов. Два десятка из них обитают в водах Палеарктики. Оригинальная фауна насекомых проявляется главным образом среди жуков, в то время как бабочки, особенно дневные, маловыразительны. Есть в Северной Америке представители таких тропических групп, как термиты и привиденьевые.
Фаунистическое деление Нёарктики впервые было произведено Уоллесом, вычленившим в ней четыре области (или подобласти, если принимать Неарктику за область). Мы же выделяем только Канадскую и Сонорскую.
К этому царству относятся Центральная и Южная Америка, за исключением ее крайнего юга (Патагонии, Огненной Земли и Фолклендских (Мальвинских) островов). Северная граница проходит по южному краю Мексиканского плоскогорья. К Неогее также относятся острова: Большие Антильские, Багамские, Малые Антильские, Галапагос, Хуан-Фернандес и ряд мелких островков.
Фауна Южной Америки долгое время развивалась в изоляции от других фаунистических центров. Это и обусловило фаунисти-ческое своеобразие Неогеи. В данном регионе распространены примитивные группы млекопитающих, в том числе сумчатые, эндемичные отряды и семейства птиц, рептилий, амфибий, пресноводных рыб, насекомых и других беспозвоночных.
В пределах царства выделяются 2 фаунистические области — Неотропическая и Карибская.
Северная граница области совпадает с границей всего царства. Южная проходит по умеренным широтам юга материка, оставляя за пределами области (и царства) юг Чили и Аргентины (в том числе Огненную Землю, Фолклендские и другие группы островов), а также острова Карибского моря, образующие самостоятельную область.
Природные условия Неотропической области разнообразны. Через нее вдоль западного края материка тянется система гигантских хребтов — Кордильер, или Анд, отдельные вершины которых превышают 7000 м. На северо-востоке Южной Америки располагается сравнительно невысокое Гвианское нагорье, а на востоке — Бразильское нагорье, где берут начало правые притоки Амазонки. Остальные части материка представляют собой равнины. Климат области разнообразный: влажный тропический — в Центральной Америке в бассейне Амазонки; периодически засушливый, жаркий, субтропический и даже пустынный — в южной части материка.
Распределение растительности зависит от характера климата. Аллювиальные равнины в бассейне Амазонки, а также ряд других районов тропической зоны покрыты влажными экваториальными лесами (гилеями). Общая площадь южноамериканских гилей превышает 5 млн км2
. Годовое количество осадков здесь равно 2500— 3000 мм, среднемесячная температура колеблется в пределах 23—29 "С, а среднегодовая — 24—27 °С. Послеполуденные ливни в гилее — обычное явление. Характерная особенность амазонского леса — обилие заболоченных или периодически затопляемых участков. По облику экваториальные влажные леса Южной Америки сходны с африканскими лесами этого же типа, хотя по составу флоры богаче (более 15 тыс. видов цветковых растений!). Необычайно обильны семейства бромелиевых и кактусовых, почти не встречающихся в Старом Свете. Особенно пышно развиты эпифиты, орхидеи, папоротники. В заболоченных лесах.растут цекропии с ходульными корнями и полыми стволами. Везде масса пальм.
При продвижении с севера на юг и увеличении продолжительности сухого периода дождевые леса сменяются полувечнозелеными сезонными лесами, затем появляются листопадные, после них — колючее редколесье, далее — заросли кактусов и, наконец, саванны, или льяносы. Западные склоны Кордильер и прибрежная равнина покрыты редкой пустынной растительностью.
Фауна Неотропической области (рис. 152, 153) отличается высоким уровнем эндемизма, большой оригинальностью и наряду с этим дефектностью.

Рис. 152. Характерные представители фауны Неотропической
области (млекопитающие):
1 — муравьед тамандуа; 2 — броненосец; 3 —
ленивец; 4 —
цепкохвостая обезьяна; 5 — обезьяна игрунка; 6 —
древесный дикобраз; 7 — вампир; 8 —
капибара; 9 —
дикая морская свинка; 10 —
ягуар; // — соленодон; 12 —
лама

Рис. 153. Характерные представители фауны Неотропической
области (от птиц до насекомых):
I —
каменный петушок; 2 —
тукан; 3 —
королевский гриф; 4 —
колибри; 5 — червяга; 6—
солнечная цапля; 7— пипа; 8—
анаконда; 9—
нанду; 10—
гоацин;
II —
паламедея; 12 —
попугай ара; 13 —
пиранья; 14 —
черепаха матамата; 15 —
бабочка гелеконица
Среди млекопитающих обращает на себя внимание эндемич-ный отряд неполнозубых (Edentata), включающий семейства муравьедов (Myrmecophagidae), броненосцев (Dasypodidae) и ленивцев (Bradypodidae). Приматы все без исключения относятся к надсемейству широконосых обезьян Ceboidae, эндемичному для Центральной и Южной Америки. Оно состоит из 2 семейств — игрунковых (Callitrichidae) и цебид (Cebidae). Почти все их представители имеют цепкий хвост, типичный для лесных зверей области. Грызуны разнообразны, причем ядро их фауны составляет 10 эндемичных семейств подотряда дикобразных, не считая более широко распространенных беличьих и мышей. Это оригинальные древесные дикобразы (Erethizonthidae), свинковые (Caviidae), к которым относится известная «морская свинка», водосвинковые (Hydrochoeridae) с самым крупным в мире грызуном капибарой, агутиевые (Dasyproctidae), шиншилловые (Chinchillidae), восьми-зубые (Octodontidae) и др. Рукокрылые представлены 9 семействами; из них зайцеротые, дискокрылые, листоносые, дымчатые летучие мыши и кровососы вампиры эндемичны.
Прочие отряды и семейства млекопитающих являются относительно молодыми вселенцами и включают небольшое количество родов, где немало эндемичных видов.
Из непарнокопытных в Неотропической области встречаются только тапиры, состоящие из 3 видов. Парнокопытные включают следующие семейства: пекари (Tayassuidae), обитающие и в Северной Америке, верблюдовые (Camelidae), куда входят своеобразные ламы (род Lama с 2 дикими и 2 одомашненными видами), и олени (5 родов), относящиеся к подсемейству американских оленей Odocoileinae. Некоторые из оленей (пуду и мазама) живут в лесах и на высокогорьях.
Отряд хищных представлен несколькими семействами, среди которых наиболее многочисленны енотовые (Procyonidae) — носуха (Nasua), кинкажу (Potos) и др. Из кошек обычны оцелот (Felispardalis), мелкая онцилла, ягуарунди, панамериканская пума (Felis concolor) и крупный ягуар (Pantheraonca), житель густых лесов, нападающий даже на крупных тапиров. Из медведей распространен Tremarctosornatus, имеющий узкий ареал. Из псовых следует отметить гривистого волка рода Chrysocyon.
Птицы Неотропики необычайно разнообразны и оригинальны. Эндемичных семейств среди них не менее 25. Не зря Дарлингтон называет Южную Америку «континентом птиц».
В орнитофауне материка присутствуют цапли, ибисы, аисты, утки, хищные птицы, кукушки, совы, голуби, попугаи, дятлы, стрижи и некоторые другие тропикополитные и более широко распространенные семейства. В то же время здесь нет журавлей, дроф, птиц-носорогов, т. е. групп, типичных для Старого Света, незначительно и количество представителей подотряда певчих птиц (Oscines), что, вообще говоря, противоречит характеру материковой фауны.
Однако наибольший интерес, безусловно, представляют энде-мичные семейства, насчитывающие около 1500 видов (1/6 всей фауны птиц области). К их числу относятся 2 вида из эндемич-ного семейства и отряда нанду (Rheidae). Обитают нанду в открытых ландшафтах. Также эндемичен отряд Tinamifbrmes с семейством тинаму (Tinamidae), включающие плохо летающих лесных или степных птиц — отдаленных родственников американских страусов. Из куриных эндемичны семейства краксов, или гокко (Cracidae), гнездящихся на деревьях, и гоацинов (Opisthocomidae). Гоацины — странные птицы с сомнительными родственными связями. Некоторые выделяют их в особый отряд. Будучи близок к куриным, гоацин одновременно сходен с некоторыми кукушками. Летает он плохо. У птенцов на каждом крыле есть два развитых когтя, которыми они пользуются при лазании по веткам. В отличие от других крупные птенцы гоацина умеют плавать.
Журавлиные в области представлены эндемичными семействами кариам (Cariamidae), трубачей (Psophiidae) и солнечных цапель (Eurypygidae). С водоемами связаны своеобразные паламедеи (Anhimidae), образующие эндемичный подотряд гусеобразных. Это крупные грузные птицы величиной с лебедя, без плавательной перепонки между пальцами. На сгибе крыла имеются две мощные шпоры, из-за которых паламедеи называют шпорцевыми гусями. Эти шпоры птицы используют для защиты от хищников, при драках в брачный период.
Из козодоев эндемично семейство жиряков (Steatornithidae), относящихся к особому подотряду. Единственный вид жиряков, так называемый гуахаро, — крупная колониальная птица (до 1 м в размахе крыльев), устраивающая гнезда в глубоких пещерах. Она ведет ночной образ жизни и питается фруктами, обычно плодами пальм. Способность к эхолокации позволяет жирякам передвигаться в пещерах при полной темноте. Птенцы гуахаро, усиленно откармливаемые родителями, накапливают огромные жировые запасы и весят больше, чем взрослые. Местные жители используют жир гуахаро в пищу и для освещения.
Наиболее известными птицами Южной Америки являются колибри (Trochilidae). В этом семействе, насчитывающем массу родов и около 30 % видов, собраны чрезвычайно ярко окрашенные мелкие и мельчайшие птички Нового Света, причем подавляющее большинство их живет в Неотропической области. Многие виды обитают в горах, даже выше лесной зоны. Колибри — великолепные летуны, способные, подобно ночным бабочкам, «зависать» над цветками. Питаются они нектаром и мелкими насекомыми, добывая их из глубины цветка.
Попугаи (Psittacidae), будучи тропикополитными птицами, достигают в Неотропике особого разнообразия. Только здесь встречаются короткохвостые амазона Amazona и крупные яркие ара Ага с длинными хвостами. Эндемичны также момоты (Momotidae) из отряда ракшевых, роющие, подобно зимородкам, норы в береговых обрывах. Дятловые представлены не только настоящими дятлами, но и туканами (Ramphastidae), отличающимися непропорционально большим клювом яркой окраски, которые питаются плодами.
Воробьиные птицы Неотропики исключительно своеобразны. Они относятся к подотряду кричащих (Tyranni) и включают ряд эндемичных семейств: топоколов, гусеницеедов, муравьеловок, печников, древолазов, манакинов, котинг, траворезов и др.
У древолазов (Dendrocolaptidae) передние пальцы сращены у основания, а заостренный хвост при лазании по стволам деревьев создает хорошую опору. Клюв у них бывает самой различной формы — от короткого прямого до длинного серповидного. Печники (Fumariidae) получили название из-за способности отдельных видов строить из глины громадные гнезда, похожие на печи. Мана-кины (Pipridae) замечательны своим токованием, иногда коллективным, при котором они кричат и щелкают крыльями. Среди котинговых (Cotingidae) — лесных птиц, иногда называемых также звонарями, — выделяются формы с необычайным оперением (с хохлами на голове, перьевыми гребнями, кисточками и т. п.). Другие котинговые, как, например, скалистые, или каменные, петушки (род Rupicoia), привлекают внимание брачным поведением. У них бывает групповой ток, и тогда ярко-оранжевые самцы распускают хохолок, вытягивают крылья и хвосты и проделывают своеобразные движения — «танцы».
В целом орнитофауна Южной Америки резко отличается от фаун Старого Света, хотя в ее составе есть и представители широко распространенных групп.
Рептилии Неотропической области менее оригинальны, что объясняется большой древностью класса и в связи с этим давностью расселения на земном шаре. Из черепах распространены семейства каймановых, или мускусных, черепах (Chelydridae), встречающиеся и в Северной Америке; немногочисленные пресноводные черепахи (Emydidae), большинство которых живет в тропиках Восточной Азии, и сухопутные (Testudinidae), включающие здесь всего 2 вида. Из подотряда бокошейных черепах (Pleurodira) 2 семейства — пеломедузы (Pelomedusidae) и змеиношенные (Chelidae) — имеют представителей в Неотропике. Напомним, что другие виды семейств живут в Африке и на Мадагаскаре (пеломедузы) или же в Австралийской области (змеи-ношейные черепахи).
Из ящериц необыкновенного разнообразия достигают игуаны (Iguanidae). Их здесь сотни видов, большинство характерны для тропиков. О реликтовом распространении двух родов на Мадагаскаре и одного в Полинезии уже говорилось раньше. Среди американских игуан встречаются крупные древесные формы, мелкие горные, скальные, степные, полуводные и др. У рода василисков самцы имеют на голове и спине кожистые выросты в виде гребней, что придает им фантастический вид. Морские игуаны (Amblyrhynchus) живут только на Галапагосских островах и питаются водорослями. Есть в Южной Америке также сцинки и гекко-ны, но их меньше, чем в Старом Свете. Настоящих ящериц и варанов нет.
Из змей заслуживает упоминания семейство вальковатые змеи (Anilidae) с рудиментами задних ног. Они встречаются в бассейне Амазонки, а также в Инд о-Малайской области. Южноамериканский род (сверташки) представлен очень красивым видом — коралловой сверташкой. В окраске ее чередуются ярко-красные и черные цвета в виде колец. Удавовые относятся к подсемейству Boinae, характерному только для Нового Света. Среди них отметим настоящего удава (Constrictor), анаконду (Eunectesmuri-nus) — крупнейшего удава земного шара, связанного с водоемами бассейна Амазонки и Ориноко и нападающего на приходящих на водопой животных, и узкобрюхих удавов (Corallus). Семейство аспид овых (Elapidae), распространенное в тропиках и субтропиках земного шара, представлено в Неотропике немногими видами — стройным и коралловым аспидами (род Micrurus). Своей яркой окраской они очень похожи на сверташек.
Настоящих гадюк в Новом Свете нет, но их с успехом замещают ямкоголовые гадюки (Crotalinae), к которым относятся гремучие змеи. Последние являются строго американскими формами. У всех ямкоголовых имеется термолокаторный орган в виде двух ямок между ноздрями и глазами. Тепловое (инфракрасное) излучение действует на термолокатор и позволяет охотящейся ночью змее точно определять местоположение своей жертвы. В Центральной и Южной Америке насчитывается около 40 видов ямкоголовых, среди которых жарарака, бушмейстер и каскавела крайне ядовиты.
Крокодилы и кайманы весьма многочисленны в реках Неотропической области.
Из земноводных в Центральной и Южной Америке распространены отряды безногих, бесхвостых и хвостатых, причем последние представлены исключительно бедно. Безногие из семейства червяг (Caeciliidae) — обитающие в тропиках животные, в Южной Америке их 16 видов из 3 эндемичных родов. Живут червяги в почве, нередко в муравейниках и термитниках, иногда в воде, кормятся червями и насекомыми. Из хвостатых и в Северной, и в Южной Америке обитают безлегочные саламандры (Plethodontidae).
Очень многочисленны и разнообразны лягушки и жабы. В Неотропической области встречаются знаменитые пипы (Pipa), откладывающие яйца в ячейки на спине и вынашивающие их до выхода молодых животных, свистуны (Leptodactylidae), настоящие жабы (Bufonidae), секрет кожных желез которых ядовит. Среди последних выделяется громадная ага, не избегающая и солоноватых вод. Древесные лягушки, или квакши (Hylidae), разнообразны и красивы. Среди них следует отметить квакшу-кузнеца, отгораживающую глиняными валиками специальные участки в водоемах для икрометания; бразильскую квакшу, смазывающую дупла на деревьях особой смолой для их водонепроницаемости (дождевая вода попадает в дупло, и в нее самка откладывает яйца); квакш -филломедуз, поселяющихся в кронах высоких деревьев, где они склеивают листья в виде сумки и откладывают в нее яйца. Развившиеся из них головастики падают в воду, где метаморфоз завершается. Наконец, сумчатые квакши имеют на спине особую выводковую сумку, в которой вынашивают яйца. Настоящие же лягушки (Ranidae) отсутствуют в южной половине материка, а в северную проникли сравнительно недавно.
Пресноводные рыбы Неотропической области достигают огромного богатства при полном, однако, отсутствии карповых, столь характерных для тропиков Азии. Среди южноамериканских рыб выделяются двоякодышащие из семейства Lepidosirenidae. Близкие родственники их живут в Африке. Из тропикополитных костноязычных (Osteoglossidae) встречаются представители родов аравана и арапаима с большим ячеистым плавательным пузырем, стенки которого пронизаны густой сетью капилляров. Такой пузырь может действовать как легкое, усваивая кислород. Арапаима — одна из крупнейших пресноводных рыб (длина до 3 м, масса до 180 кг).
Эндемичное семейство электрических угрей (Electrophoridae) содержит единственный вид, населяющий небольшие реки северо-востока Южной Америки и притоков Амазонки и питающийся рыбой. Громадные электрические органы этой крупной рыбы служат для защиты и нападения — они способны давать разряд 650 В при силе тока всего 0,5—0,75 А.
Из карпообразных (не карповых) многочисленны харацино-видные (около 800 видов). Среди них широкую известность получила пиранья — представитель семейства пираньевых (Serrasalmidae). Острые зубы и сильные челюсти позволяют им выхватывать куски мяса из тела животных любой величины. Пираньи мгновенно собираются в стаи и нападают на плывущих через реку животных, запах крови чрезвычайно их возбуждает. Мелкие хара-циновые (Characidae) — красивые рыбки, которых охотно держат в аквариумах. Это так называемые тетры, неоны и пр. Представители харациновых живут в водоемах Африки.
Необыкновенно разнообразны сомовые — семейства Doradidae, Pimelodidae, Pygididae (ведут паразитический образ жизни), Aspre-dinidae и другие (всего 12 семейств). Они эндемичны. Сомиков каллихтов нередко держат в аквариумах. Из колючеперых для области типичны семейства нандовых (Nandidae) и цихловых (Cich-lidae), обитающих также в тропической Африке. К последним относится замечательная скалярия, которую часто разводят в аквариумах.
Наземных беспозвоночных Неотропики, особенно насекомых, невозможно охарактеризовать кратко. Здесь обитают роскошно окрашенные бабочки — кавалеры (Papilonidae) и морфо (Мог-phidae), знаменитые своей мимикрией геликониды, огромные бражники и урании. Еще Уоллес писал, что в окрестностях города Пара можно собрать не менее 700 видов дневных бабочек. Среди жуков встречаются настоящие гиганты — рогачи, дровосеки-титаны, многие златки. Щелкуны рода Pyrophoris способны излучать яркий свет — это самые знаменитые светляки в мире. Перечислить все группы насекомых Южной Америки здесь невозможно, для этого потребовалось бы издание отдельного тома. Отметим лишь немногих. Нежалящие пчелы — мелипоны устраивают гнезда открыто на ветках деревьев. Очень велико разнообразие муравьев, их тут больше, чем в тропиках других частей земного шара. Наиболее известны бродячие эцитоны — пугало сельвы. О них Г.Бэйтс (1859), живший в Бразилии долгое время, сказал: «В долине Амазонки господствует не человек, а муравей». Действительно, колонны этих хищников отпугивают все живое. Муравьи са-уба, относящиеся к листорезам, вырезают на листьях низкорослых деревьев круглые кусочки и затем переносят их в подземные гнезда. На пережеванной массе листьев сауба выращивают грибы, которыми и питаются. При основании новой колонии молодая самка вылетает из гнезда с кусочком грибницы в челюстях, чтобы на новом месте заложить плантацию грибов для будущего муравейника.
Местные сколопендры выделяются гигантскими размерами (до 25 см длиной). В Южной Америке многочисленны крупные пауки птицеяды. Их жертвами оказываются не только насекомые, но и ящерицы, лягушки и даже птенцы. В лесах масса наземных пиявок.
В целом необходимо сказать, что современная фауна Неотропической области, точнее, ее позвоночные животные — результат смешения древней эндемичной фауны третичного времени и новой фауны, проникшей из Азии через Северную Америку.
На основании особенностей фауны данной области в ней можно выделить 3 подобласти: Гвиано-Бразильскую, Центрально-Американскую и Чилийскую, или подобласть пампасов.
К данной области относятся Большие Антильские (Куба, Ямайка, Гаити и Пуэрто-Рико), Багамские, Малые Антильские острова, Тринидад. Это настоящая островная область, поскольку фауна ее обладает типичными для острова чертами.
Природные условия Карибской области вполне благоприятны. Большая часть островов покрыта (или была покрыта в недалеком прошлом) тропическими лесами, вместе с тем здесь есть и другие формации растительности.
Наземных млекопитающих в области очень мало. Насекомоядные представлены удивительными щелезубами, живущими на Кубе и Гаити: два вида их — кубинский (Atopogalecubana) и гаитянский (Solenodonparadoxus) — образуют эндемичное семейство Solenodontidae, близкое к землеройкам. Из грызунов есть хутие-вые — хутии (роды Capromys и Geocapromys), а также загути (Plagiodontia). Островной опоссум и один вид енота (на Багамских островах) дополняют короткий список наземных млекопитающих. Зато рукокрылых очень много: это плодоядные листоносые, вампиры, зайцегубые, рыбоядные, листобородые.
Орнитофауна области интересна благодаря присутствию на Больших Антильских островах эндемичного семейства тоди (Todi-dae) из отряда ракш. Клюв у тоди приплюснутый сверху вниз, за что их еще называют плоскоклювами. Ярко окрашенные птички живут большей частью в лесах, питаются насекомыми, схватывая их на лету. На каждом из островов имеется по эндемичному виду, на Гаити их 2. Много эндемиков среди колибри, трогонов, кукушек, тиранов и других семейств. Всего зарегистрировано около 140 видов, из которых 25 % эндемики. Больше всего эндемиков на Ямайке.
Рептилии представлены несколькими видами черепах, 2 видами крокодилов (1 из них эндемик Кубы), многочисленными ящерицами (в том числе роющими амфисбенами) и немногими змеями. Ядовитая копьеголовая змея (Bothropsatrox) встречается только на Малых Антильских островах.
Из амфибий нет ни безногих, ни хвостатых. Здесь обитает лишь 5 родов лягушек, среди которых — квакши, южноамериканские свистуны и короткоголовые лягушки (только на Кубе).
Пресноводные рыбы представлены североамериканской панцирной щукой (на Кубе), хромисами, а также живородящими и яйце-родящими карпозубыми (на Кубе и Гаити). Здесь обычна и знаменитая рыбка гамбузия, истребляющая личинок малярийных комаров.
Это царство включает крайний юг материка Южной Америки (Патагонию и Огненную Землю), Австралию, Тасманию, Новую Зеландию и субантарктические острова Мирового океана. Таким образом, Нотогея — настоящее островное царство, для которого невозможно дать единую фаунистическую характеристику. Единственное, что объединяет входящие в Нотогею земли (кроме сходства, обусловленного древностью их происхождения), — это чисто негативный признак: бедность и одностороннее развитие животного мира (рис. 154, 155).
В царстве Нотогея выделяются 3 области — Австралийская, Новозеландская и Патагонская (Голантарктическая).
Сюда относятся Австралия, крупный остров Тасмания и ряд мелких островов (Кенгуру, Кинг, Флиндерс и др.).
Характер природных условий Австралии определяется ее географическим положением, рельефом и климатом. Северная часть материка находится в тропическом поясе, центральная — в субтропическом, а южная — в умеренном. Горные хребты располагаются по периферии, перехватывая основную массу осадков, и центральные части материка испытывают недостаток увлажнения. Западное побережье окаймлено низкими горами Дарлинг и Виктории, по восточному побережью тянется Большой Водораздельный хребет, отдельные вершины которого достигают 2200 м. Наибольшее количество дождей выпадает на северо-востоке Австралии (в Квинсленде), достаточно их и на острове Тасмания. В связи с этим в указанных районах растительность представлена лесами: на северо-востоке тропическими, к югу от них субтропическими, еще южнее хвойно-широколиственными (вечнозелеными). Во влажных субтропиках Австралии растут крупные эвкалипты. На Тасмании распространены широколиственные леса из нотофагуса и древовидных папоротников.

Рис. 154. Характерные представители фауны Нотогеи (Австралийская область):
1 —
ехидна; 2
— утконос; 3 —
гигантский кенгуру; 4 —
сумчатый дьявол; 5 — вомбат; 6 —
коала; 7 — сумчатый волк; 8 —
кус-кус; 9 —
сумчатый крот; 10 —
черный лебедь; 11 —
лирохвост; 12 —
эму

Рис 155. Характерные представители фауны Нотогеи (Австралийская,
Новозеландская и Патагонская области):
1 -
мара; 2 -
вискача; 3 -
киви; 4 -
гаттерия; 5 -
молох; 6 -
какаду; 7 -кеа; 8 - рогозуб; 9 - плащеносная ящерица; 10 -
скалистый пингвин; 11 -
королевский пингвин
Юго-запад Австралии покрыт относительно сухими и редко-стойными лесами. В остальной же части материка — самой засушливой — встречается либо своеобразная растительность из низкорослых жестколиственных эвкалиптов или вечнозеленых акаций (так называемый скрэб), либо редкие кустарниковидные солянки. Большие пространства в Австралии занимают пустыни, где расте-
ния встречаются только вдоль лощин, по берегам соленых озер или временных водостоков. Австралийские пустыни довольно разнообразны: песчаные, глинистые, галечниковые, солончаковые и всевозможные их варианты. Для песчаных пустынь типичны заросли ксероморфных жестколиственных злаков из родов спинифекс и триодия.
Самой характерной особенностью австралийской фауны является присутствие однопроходных и сумчатых млекопитающих.
Однопроходные, или первозвери, представляют собой эндемичный подкласс с крайне примитивными признаками: у них имеется клоака, отсутствуют соски, размножение осуществляется путем откладывания яиц. В этом подклассе и единственном отряде Monotremata всего 2 семейства: утконосы (Ornithorhynchidae) и ехидны (Tachyglossidae), причем к последнему относится и новогвинейский род проехидна. Однопроходные — звери специализированные. Утконос благодаря непромокаемому меху, плавательным перепонкам на ногах, роговому чехлу на челюстях («клюв») ведет водный образ жизни. Ехидны приспособлены к рытью, питаются муравьями и термитами. У них крепкие ноги, мощные когти, морда вытянута в клюв, язык длинный и клейкий. Ехидны найдены в Австралии и в ископаемом состоянии.
Сумчатых (отряд Marsupialia) в Австралийской области насчитывается 8 семейств: хищные сумчатые (Dasyuridae); намбаты, или сумчатые муравьеды (Myrmecobiidae); сумчатые кроты (Notoryc-tidae); бандикуты (Peramelidae), относящиеся к подотряду многорезцовых; из подотряда двурезцовых — семейства поссумов, или фалангистов (Phalangeridae); сумчатые медведи, или коала (Phas-colarctidae); вомбаты (Vombatidae) и кенгуру (Macropodidae). Разнообразие и богатство сумчатых в области поразительны. Занимая те же экологические ниши, что и плацентарные звери в других областях земного шара, сумчатые приобрели с ними большое конвергентное сходство. На собачьих похожи сумчатый волк и сумчатый дьявол, на настоящих кротов — сумчатые кроты, на тушканчиков — сумчатый прыгун, на сурков — неуклюжие вомбаты, общеизвестный коала напоминает игрушечного медведя, хотя и занимает ту же экологическую нишу, что и южноамериканский ленивец. Тем не менее при всем разнообразии представителей отдельных семейств они обладают четко выраженными признаками отряда: у них нет плаценты, детеныши рождаются мелкими и недоразвитыми, имеются сумчатые кости, а у большинства и сумка, строение мозга простое, температура тела ниже, чем у плацентарных.
В Австралийской области обитают и высшие звери, но они представлены только грызунами и рукокрылыми. Собака же динго — недавний вселенец в фауну Австралии.
Грызуны в Австралии относятся к семейству мышиных. Кроме крыс, широко расселившихся по всему земному шару, на материке выделяется субэндемичное подсемейство полуводных бобровых крыс (Hydromyinae), отдельные представители которых доходят до Новой Гвинеи. Рукокрылые состоят как из плодоядных, так и из насекомоядных летучих мышей. Среди них много энде-мичных видов.
В настоящее время фауна млекопитающих Австралии сильно изменена человеком и в известной мере «европеизирована» благодаря завозу европейских видов.
Птицы Австралийской области разнообразны и распределяются среди 50 семейств. К бескилевым принадлежит семейство эму (Dromaedidae), эндемичное для материка, а также казуары. Впрочем, казуары населяют только северо-восточную часть Австралии, занятую тропическими дождевыми лесами. Основная же масса их видов живет на Новой Гвинее, т.е. за пределами Нотогеи. Из гусеобразных в Австралии водится красивый черный лебедь, которого разводят в декоративных целях в парках Европы. Из видов семейств, родина которых лежит за пределами Австралии, следует назвать белого ястреба, скопу, аистов и др. Семейство сорных кур (Mega-podiidae) представлено в Австралии многими видами, населяющими и засушливые местности центра материка, и леса его северо-восточной части. Как известно, сорные куры зарывают свои яйца в песок, почву, вулканический пепел или в кучи гниющего растительного мусора, и инкубация их происходит за счет тепла, поступающего извне. У австралийской сорной индейки и глазчатой курицы самцы создают из листьев с прослойками песка сложные «инкубаторы» и в зависимости от температуры, которую проверяют при помощи клюва, по мере надобности снимают или добавляют слои. Самка же только приходит время от времени откладывать яйца.
Очень своеобразны австралийские попугаи. Среди них выделяются какаду, некоторые лори (например, щеткоязычный горный попугай), плоскохвостые, к которым относится ночной попугай (Geopsittacusoccidentalis), строящий настоящее гнездо, что для попугаев не характерно. На юге Австралии живет элегантный попугайчик из рода Neophema, обитающего исключительно в Австралии и Новой Зеландии. Широко известен попугай розелла, ставший спутником человека. Местами он наносит ущерб урожаю пшеницы, клевера, а также фруктов. Но наиболее известен из плос-кохвостых все-таки волнистый попугайчик, которого часто содержат в клетках как комнатную птицу.
Среди многочисленных зимородков выделяется громадный смеющийся зимородок (Dacelogigas), буровато-серой окраски, питающийся змеями и ящерицами.
Эндемичны для Австралии семейства лирохвостов (Menuridae) и кустарниковых птиц (Atrichornithidae). Виды лирохвостов получили название из-за того, что крайняя пара рулевых перьев у них лентообразная, изогнутая, длиной до 60 см, средняя — длинная, узкая, напоминает струны. Благодаря этому хвост похож на старинный музыкальный инструмент — лиру. Населяют лирохвосты леса и кустарниковые заросли юго-востока материка.
Медососовые (Meliphagidae) — также весьма характерное семейство птиц, распространенное в Австралии и Полинезии. Виды его отличаются большим разнообразием, напоминающим адаптивную радиацию сумчатых. Медососы — потребители нектара, пыльцы и насекомых. Чаще всего они ведут древесный образ жизни и только изредка опускаются на землю в поисках корма. Многие из них играют важную роль в опылении эвкалиптов.
Преимущественно в Австралии живут виды семейств Gralli-nidae (сорочьи жаворонки), Artamidae (лесные ласточки) и Cracticidae (австралийские сороки).
В общем островные черты проявляются и на материке, что выражается в известной дефектности фауны при высоком уровне эндемизма.
Своеобразные черты присущи и фауне рептилий. На австралийском материке обитают змеиношейные черепахи (Chelidae), распространенные также в Южной Америке и на Новой Гвинее. Представители этого семейства не втягивают шею, а закладывают ее сбоку под панцирь. Питаются они пресноводной рыбой и беспозвоночными, активно преследуя их в реках и озерах. Гекконы разнообразны и зачастую внешне причудливы из-за расплющенного или сильно утолщенного хвоста. У австралийского голопало-го геккона в хвосте откладывается запас жира. Почти эндемично семейство чешуеногов (Pygopodidae), обладающих змееобразно вытянутым туловищем. Передние ноги у них отсутствуют, а задние часто редуцированы. В семействе 13 видов, 12 из них найдено в Австралии, 1 — на Новой Гвинее.
Из широко распространенного семейства агам несколько представителей имеют крайне своеобразный вид. На севере и северо-западе Австралии живет плащеносная ящерица (Chlamydosauruskingi), имеющая вокруг шеи кожную перепонку («воротник»), который ящерица поднимает и опускает при помощи особого мускула. У самцов развернутый воротник представляет собой круг диаметром до 15 см, а его яркая расцветка отпугивает врага или соперника. Замечательна способность ящерицы бегать на задних ногах, держа туловище почти вертикально над землей.
В Австралии водится еще одна интересная ящерица — молох (Molochhorridus) — единственный вид своего рода. Живет он в песчаных пустынях, питается муравьями. Все тело этой небольшой (около 20 см) ящерицы покрыто многочисленными шипами и колючками. Кожа очень гигроскопична и впитывает воду, как фильтровальная бумага. При этом масса ящерицы возрастает почти на 30 %. Благодаря такому приспособлению молох запасает влагу редко выпадающих дождей и выживает в крайне засушливых условиях.
Сцинки представлены в Австралии эндемичными родами (колючие сцинки, или эгернии, и др.). Короткохвостый сцинк (Trachy-saurusrugosa) — живородящий: развивающиеся в яйцеводах самки эмбрионы не покрыты яйцевыми оболочками и получают питание из крови матери через зачаточную плаценту. Настоящих ящериц в области нет, зато много варанов, и среди них самый мелкий в семействе короткохвостый варан не превышает 20 см в длину.
Среди австралийских змей часть относится к широко распространенным группам, часть — к эндемикам. Это тропикополит-ные слепуны, питоны (например, ромбический питон, аметистовый и др.), ужеобразные и главным образом аспидовые змеи. Причем по численности ядовитые формы явно преобладают над неядовитыми. В Австралии аспидовые змеи (других ядовитых змей здесь нет) представлены наиболее древними и примитивными видами, и следует отметить, что эволюция их привела к возникновению форм, конвергентно сходных с гадюками и ямкоголо-выми. Из 22 родов аспидовых, характерных для области, отметим денисоний, ехидн, крайне ядовитых и агрессивных тайпанов (длина до 3,5 м), тигровых змей, похожих на гадюк акантофисов, и др. Считается, что у тигровой змеи наиболее сильный яд среди всех наземных змей.
Крокодилы встречаются только в северной части Австралии.
Из амфибий распространены лишь бесхвостые — жабы, свистуны и квакши. Среди свистунов выделяется эндемичное подсемейство Criinidae, включающее 56 видов. Квакши, обычно живущие в лесах, имеют в Австралии и пустынных представителей, пользующихся временными водоемами (красная квакша). Пустынная австралийская жаба (Chiroleptesplaticephalus) из подсемейства австралийских жаб обитает в пустынях центральной части материка и отличается способностью запасать воду в полости тела и в подкожных полостях. Местные жители используют ее как источник питьевой воды. Единственный вид еще одного рода австралийских жаб — Myobatrachus — живет в термитниках.
Из пресноводных рыб Австралии особенно замечательны двоякодышащие и костноязычные, или аравановые. Среди первых известен рогозуб (Neoceratodusforsteri) — представитель эндемич-ного семейства. Эта крупная рыба (до 175 см и свыше 10 кг) населяет медленно текущие реки северо-востока. В засушливые сезо-
ны, когда реки пересыхают, рогозуб поселяется в углублениях дна, где сосредоточивается перегретая гниющая вода, и переходит там к дыханию атмосферным воздухом при помощи единственного легкого. К семейству костноязычных относится Sclero-pagesleichhardtii. Он также способен дышать атмосферным воздухом, но уже с помощью плавательного пузыря, стенки которого пронизаны кровеносными сосудами. Эти рыбы характерны для тропиков.
Исключительно интересны рыбы семейства галаксиевых (Gala-xiidae). Они распространены только в Южном полушарии: 24 вида в Австралии и на Тасмании, 20 — в Новой Зеландии, 7 — на юге Южной Америки, 2 вида в Южной Африке, 3 рода эндемичны для области. Причины такого распространения до сих пор не установлены. Некоторые считают это свидетельством прежнего соединения материков, другие — результатом расселения через соленые воды, поскольку многие галаксиевые солевыносливы и могут жить в устьях рек, впадающих в море. Карповых рыб в Нотогее нет.
Беспозвоночные Австралийской области менее роскошны, чем в Юго-Восточной Азии. Великолепные дневные бабочки — кавалеры и другие встречаются лишь на северо-востоке материка. В ряде районов преобладают разнообразнейшие моли, которых здесь больше, чем всех европейских бабочек, вместе взятых. Среди насекомых можно отметить и многочисленных муравьев из примитивного подсемейства понерин, обитающих и в Африке, но здесь они представлены наиболее древними видами. К ним относятся муравьи-бульдоги с громадными челюстями и жалом. Знамениты медовые муравьи (Melophorus), у которых рабочие особи с непомерно раздутыми от сладкой жидкости телами висят на потолке камеры муравейника, сохраняя для остальных запасы сиропа. Термиты сравнительно немногочисленны в данной области. Они делают главным образом подземные жилища. Среди фитофагов обильны прямокрылые, особенно саранчовые.
Вообще энтомофауне Австралийской области присущи наиболее примитивные виды из семейств, которые широко распространены по земному шару. Таковы, например, австралийские жуки-листоеды, жужелицы и др.
Среди дождевых червей особенно интересно семейство мега-сколецид (Megascolecidae). Это настоящие гиганты (до 2,5 м), характеризующиеся чаще всего антибореальным распространением.
Эта островная область объединяет оба крупных острова Новой Зеландии (Северный и Южный), а также ряд мелких островков — Окленд, Кермадек, Чатем и др.
Новая Зеландия — гористая страна, причем горы имеют вид расчлененных хребтов с альпийской зоной, ледниками и озерами. Следы вулканической деятельности проявляются здесь в обилии горячих источников. Климат страны мягкий, осадков много — от 815 до 1090 мм в год. Средняя годовая температура 12-13 °С, минимальная температура не ниже 5 "С. Основной ландшафт — густой влаголюбивый лес тропического типа, несмотря на то что климат умеренный. Для новозеландских лесов характерно обилие папоротников, в том числе древовидных, достигающих большой высоты. В лесах растет и нотофагус — типичная антибореальная порода. Вдоль восточного берега Южного острова тянется полоса степей.
Фауна области крайне дефектна, что выражается прежде всего в отсутствии наземных млекопитающих (о завезенных видах речь будет идти ниже). Единственный грызун — маорийская крыса, — очевидно, был завезен мореплавателями маорийцами еще до появления здесь европейцев. Млекопитающие представлены 2 видами летучих мышей: один — из древнего эндемичного семейства футлярокрылов (Mystacinidae), другой — из австралийского рода, недавно вселившийся в местную фауну. Новозеландский футля-рокрыл — единственный представитель семейства, ведущий и наземный образ жизни. Питается он насекомыми, а также падалью.
Для Новой Зеландии характерна неповторимая группировка нелетающих птиц. Еще в историческое время здесь жили представители 2 вымерших ныне семейств гигантских моа, достигавшие 3—5 м высоты. Многочисленные полуископаемые остатки позволили реконструировать внешний облик этих птиц. Оказалось, что они близки к казуарам. Из существующих в настоящее время бескрылых птиц назовем киви — представителей эндемичного отряда Apterygiformes семейства Apterygidae. В отряде 1 род с 3 видами. Киви — птица размером с курицу, у нее зачаточные крылья, крошечные глаза и длинный клюв, на конце которого помещаются ноздри (единственный случай в классе птиц). Эти лесные птицы ведут ночной образ жизни, добывая пищу (червей, насекомых) в сырой почве при помощи хорошо развитого обоняния. Кроме киви из нелетающих птиц в области живут пастушки рода Ocydromus. Они также ведут ночной образ жизни, но питаются плодами. Не летает и знаменитый пастушок такахе (Notornis-mantelli) — одна из самых редких птиц Новой Зеландии. Она долгое время считалась вымершей, но заново была «открыта» в 1948 г. Небольшая популяция ее (200—300 экз.) живет сейчас в резервате. Такахе занесена в Международную красную книгу. Есть в местной фауне 2 нелетающих вида попугаев (так называемые совиные попугаи, или какало), отличающиеся мягкими перьями. Эти представители эндемичного подсемейства, питающиеся ягодами, пока еще сохранились в густых буковых горных лесах, но уже стоят на грани исчезновения.
Из других птиц Новой Зеландии отметим эндемичные семейства новозеландских крапивников (Xenicidae) и гуйий (Callaeidae), причем первые — плохие летуны. Кроме того, есть эндемичные роды и виды из хищных и сов, кукушек, голубей, зимородков и различных воробьиных.
В фауне рептилий присутствует гаттерия, или туатара, — единственный на земном шаре представитель отряда клювоголовых (Rhynchocephalia) семейства клинозубых (Sphenodontidae). Гаттерия — наиболее древнее из современных пресмыкающихся животное, древний реликт Новой Зеландии. У нее есть настоящий теменной глаз, сошниковые зубы, остатки хорды. Живет гаттерия в норах на морском побережье и питается червями и насекомыми. Вид этот строго охраняется и занесен в Международную красную книгу.
Из других рептилий отметим гекконов (2 эндемичных рода, оба яйцеживородящие) и сцинков. Змей и черепах в Новой Зеландии нет. Амфибии представлены 2-3 видами лягушек, из рода Leiopelma семейства гладконогих (Leiopelmidae). Лиопельмы — реликтовые древние животные, эндемики Новой Зеландии. Они отличаются тем, что у них нет перепонок между пальцами, позвонки амфицельные, а ребра сохраняются в течение всей жизни.
Пресноводные рыбы немногочисленны — 1 вид угря, несколько видов семейств Retropinnidae и Aplochitonidae, близких к лососям, но встречающихся только в Южном полушарии, и уже известные нам галаксииды.
Фауна беспозвоночных довольно бедна. Дождевые черви (настоящие и гигантский) имеют родственников в Австралии и на Мадагаскаре. Скорпионов нет. Насекомых масса, но они маловыразительны. Среди бабочек отсутствуют кавалеры и белянки, но молеобразных много. Жуки представлены главным образом жужелицами и долгоносиками. Термитов всего 3—4 вида. Эндемизм в фауне насекомых развит весьма значительно.
В эту область входят территории юга Южной Америки с умеренным климатом (юг Чили и Аргентины, включая Огненную Землю), архипелаг Хуан-Фернандес, Фолклендские (Мальвин-ские) острова и ряд мелких островных групп, а также отдельные острова, разбросанные в южной части Мирового океана: Керге-лен, Крозе, Южная Георгия, Тристан-да-Кунья, остров Амстердам и др. Ботанико-географы выделяют эту область давно, зоо-географы же юг Южной Америки рассматривали как подобласть Неотропики. Между тем исследования последнего времени вскрывают глубокое своеобразие фауны данного региона, что объясняется его геологической историей.
Область очень фрагментарна. Для нее характерно преобладание степных пространств (пампасов), а южнее — полупустынь и даже пустынь. Растительный покров здесь разреженный и состоит из редких дерновин, злаков, кустиков и подушек болакса и азорел-лы. Холодные высокогорья Кордильер — «пуны» — близки по растительности к полупустыням на равнинах. На архипелаге Чилоэ и на Огненной Земле растут влажные леса, состоящие из нотофагуса.
В фауне Патагонской области встречаются виды, свойственные Неотропике (колибри, броненосцы), но число их невелико.
Типичными для области следует считать ценолестовых сумчатых млекопитающих. Южнее других распространен род Notodelphis(до 47° ю. ш.). В Чили и на островах Чилоэ живут представители рода Dromiciops. Только в Чили обитают Rhyncholestes и Marmosa. Эта группа млекопитающих объединяет в себе признаки хищных и растительноядных сумчатых и выделяется в особый подотряд Coenolestoidea, эндемичный для данной области. Ценолесты найдены в ископаемом состоянии на территории Патагонии в слоях раннетретичного времени. На крайней южной оконечности материка в настоящее время сумчатых, по-видимому, нет.
Из отряда грызунов в Патагонской области есть не менее своеобразные виды кавий (свинковых), например мара (Dolichotispatagonica), достигающая 1 м длины, вискачи Lagostomus и Lagi-dium из шиншилловых, а также туко-туко (Ctenomys) из семейства гребнемышиных. Последний населяет территорию от 15° ю. ш. до Огненной Земли. Родина нутрии также находится здесь.
Из птиц следует отметить нанду Дарвина (Rheapennata), черношейного лебедя, утку эндемичного рода Merganetta, которая близка к новозеландским уткам, ржанку эндемичного рода Chub-bia, зобатых бегунков (Thinocoridae).
Рептилии немногочисленны. Это некоторые игуаны (род Lio-laemus доходит до Огненной Земли), тейи, примитивные ужеобразные змеи. Из амфибий в Чили обитает ринодерма, а из рыб — галаксииды.
Гораздо богаче в области представлены беспозвоночные. Они включают большое количество своеобразных эндемиков, нередко высокого ранга. О.Л.Крыжановский (1977) приводит характерные примеры относительно распространения ряда жужелиц. Так, для Патагонии эндемичны трибы Cnemacanthini и Antarctiini. Чрезвычайно своеобразно распространение отдельных эндемичных родов: красивые крупные Ceroglossus имеют ближайших родственников среди голарктических красотелов (Calosoma), жужелицы трибы Migadopini обитают в Патагонии, Новой Зеландии и Австралии, жуки-рогачи из подсемейства Chiasognathinae населяют леса Чили, Австралии и юга Африки.
На островах Тристан-да-Кунья вулканического происхождения единственными наземными позвоночными являются птицы: 2 эндемичных рода нелетающих пастушков, 1 эндемичный род дрозда, 2 эндемичных рода вьюрков. По мнению специалистов, все эти птицы связаны родством с южноамериканскими (пата-гонскими) группами.
Фолклендские (Мальвинские) острова отличаются умеренно холодным климатом и полным отсутствием древесной растительности. Фауна позвоночных здесь очень бедная, амфибий и рептилий вообще нет. Единственный эндемик — громадная лисица — родственна южноамериканским видам рода Dusicyon. Несомненно, эти острова были некогда частью Патагонии. Впрочем, Дар-лингтон высказал предположение, что предки фолклендской лисицы достигли архипелага на дрейфующих льдах.
Несмотря на то, что острова Субантарктики расположены в приполярной зоне, климат здесь океанический, прохладный и сырой, близкий к климату высокогорий тропической зоны. Древесной растительности нет. Многие растения имеют вид подушек (азорелла на Кергелене и др.). В фауне преобладают морские звери и птицы — королевский и золотоволосый пингвин, буревестники и эндемичное семейство белых ржанок (Chionididae), морские слоны — крупные тюлени. Стада последних на Кергелене и Южной Георгии насчитывают до 250 тыс. голов. Сходство растительного и животного мира субантарктических островов достаточно велико для объединения их в одну область.
С конца ледникового периода к факторам, определяющим условия существования животных и формирования фауны, добавилась деятельность человека. Начиная с каменного века люди на всем земном шаре не только ограничивают численность популяций отдельных видов — объектов охоты, но и полностью их уничтожают. Д.Фишер (1976) в предисловии к Международной красной книге приводит такие примеры.
Плейстоценовая фауна, существовавшая 10 тыс. лет назад, характеризовалась крупными и узкоспециализированными животными, а поэтому была особенно уязвима. В Европе человек разумный появился около 250 тыс. лет назад. Люди развивали охотничье искусство, добывали огонь, овладевали орудиями труда, теснили своих главных соперников — пещерного льва, саблезубого тигра, пещерного медведя — и занимали их убежища. По мере приобретения опыта и совершенствования орудий охоты воздействие человека на животных усиливалось. Примерно 100 тыс. лет назад исчезли лесные слоны и бегемоты, несколько позже ~ лесные носороги. Гигантский олень сохранился вплоть до железного века. Предки индейцев Северной Америки достигли этого материка не позже 15 тыс. лет назад, когда плейстоценовая фауна уже была сформирована, и есть доказательства, что они охотились примитивным оружием — дротиками — на американского мастодонта, гигантскую ламу, колумбийского мамонта. То же происходило и на других материках. В результате плейстоценовая фауна сохранилась только в Африке.
Люди каменного века сталкивались с последствиями собственного «плейстоценового перепромысла» и вырабатывали различные охотничьи законы, правила раздела добычи, системы запретов, т. е. учились жить в гармонии с природой.
Переход к скотоводству, а затем и земледелию не исключал охоты, но она уже не была главным средством к существованию. Новая деятельность человека (раскорчевка лесов, распашка земель, выпас скота) вызывала изменение среды обитания. Воздействие на животный мир планеты становилось все более разрушительным. Если первоначально оно было локальным, то после великих географических открытий и путешествий европейских мореплавателей (XV—XVI вв.) постепенно превращалось в глобальное.
Развитие капитализма, захват колоний и беспощадная эксплуатация природных ресурсов распространили влияние оснащенного техникой человека на самые отдаленные уголки планеты. Можно сказать, что человек стал важнейшим фактором, действующим как прямо (истребляя одни виды и расселяя другие), так и опосредованно — через изменение условий их обитания. Косвенно влияние по масштабам и результатам стало доминирующим. В целом антропическое воздействие носит двоякий характер — положительный и отрицательный.
Прямое влияние человека на фауну земного шара привело к уничтожению ряда животных как видов. По данным Б. Гржимека (1977), за последние 400 лет уничтожено 130 видов млекопитающих и птиц, из них 76 — уже после Первой мировой войны. Еще 550 видов находятся на грани полного истребления.
По данным Н.А.Гладкова, А.В.Михеева и В.М.Галушина (1975), до 1800 г. вымерло 33 вида млекопитающих и 30 видов птиц. С 1801 по 1850 г. вымерло соответственно 2 и 20 видов, с 1851 по 1890 г. — 31 и 50, а с 1901 по 1950 г. — 40 видов млекопитающих и 150 видов и подвидов птиц. Помимо полного исчезнове-
ния видов широкий размах приняло частичное вымирание, иначе говоря, исчезновение их в отдельных странах и регионах. С точки зрения зоогеографии это не что иное, как сокращение и фрагментация ареалов. Те же авторы приводят и конкретные примеры. Например, в Австралии исчезло 7 видов кенгуру, а в одном только австралийском штате Новый Южный Уэльс из 52 видов сумчатых 11 вымерло под влиянием выпаса овец и прямого уничтожения. В штате Алабама (США) исчезло 3 вида змей, а в штате Луизиана — 4 вида лягушек вследствие применения ядохимикатов. На Кавказе под воздействием человека вымерло 9 видов зверей и среди них лев, дикий бык тур, гепард, бобр, лось, зубр и тигр.
Скорость вымирания видов животных пропорциональна скорости и глубине изменений, вносимых человеком в природу. При грубом вторжении людей природа претерпевает быстрые и глубокие изменения. Это видно на примере перемен в животном мире Австралии, Африки и Северной Америки. В то же время фауна Европы и Азии пострадала значительно меньше и именно из-за того, что процесс антропического воздействия начался давно, а вызванные им изменения природы происходили медленно. Животные, теснимые человеком, находили себе новые убежища и постепенно приспосабливались к новым условиям.
Что же касается островных фаун, то здесь дело обстояло совсем иначе. Н.А. Гладков (1959) приводит следующие данные. На Гавайских островах вымерло 26 форм птиц, или 60 % всей их фауны. На островах Лусон и Мидуэй (Филиппины) из местных видов исчезло 3, т. е. 60 %, на Маскаренских островах из 28 местных видов вымерло 34, или 86 %. Это наиболее высокий процент исчезнувших видов в мире.
В ряде случаев точная датировка исчезновения и его причины для многих видов устанавливаются с трудом. Но часто причины и время исчезновения известны. Приведем только наиболее характерные примеры.
Европейская степная дикая лошадь — тарпан — в прошлом была очень многочисленной в степях юго-восточной Европы. В определенной мере и сама степная растительность сформировалась под воздействием громадных стад диких копытных, в том числе и тарпана. Однако повсеместная охота на него из-за мяса, но главным образом распашка степей привели к тому, что популяции этого вида неуклонно уменьшались по численности и в конце концов исчезли. В восточной Европе тарпан окончательно исчез в первые десятилетия XIX в., а в южнорусских степях табуны встречались еще в 30-х годах, отдельные же группы можно было видеть в 60-е годы, но к концу столетия и они исчезли. Последний тарпан на территории нашей страны был убит в 1879 г.
Дикий бык тур, родоначальник европейского домашнего рогатого скота, в доисторические времена был обычным животным Европы, Сибири, Малой Азии и Северной Африки. В результате охоты на него, а также вырубки лесов и других перемен в ландшафтах тур постепенно вытеснялся из большей части своего ареала и к XV в. сохранился лишь в Польше и России. Но уже в начале XVI в. он стал редким видом и содержался только в загонах и лесных охраняемых участках в бассейнах Вислы и Буга. Точно известно, что последний экземпляр тура погиб в 1627 г.
Зебра квагга, южный подвид одного из африканских видов зебр, в начале XIX в. была обычным животным Южной Африки. Она отличалась от остальных зебр наличием полос только на морде и шее. Когда началось освоение территории под сельскохозяйственные культуры, местные жители — европейские поселенцы (буры) — принялись истреблять крупных животных. Жертвой стала и квагга. Ее в массе отстреливали, но использовали главным образом шкуры, которые шли на экспорт или на изготовление мешков под зерно. На большей части своего ареала квагга была уничтожена за какие-нибудь 50 лет, а последние экземпляры исчезли к 1880 г.
Не менее плачевна судьба и некоторых птиц, бывших ранее обычными, но в настоящее время от них остались только музейные чучела.
Бескрылая гагарка — крупная нелетающая птица, в прошлом населявшая скалистые побережья и острова Северной Атлантики. Как объект охоты гагарка была известна с доисторических времен. О численности популяций этой птицы можно судить по тому, что нередко целые суда загружались ее мясом и яйцами. В результате хищнической охоты численность бескрылой гагарки начала резко убывать к началу XVII в., а в 1844 г. у берегов Исландии была убита последняя пара.
Странствующий голубь еще в первой половине XIX в. был одной из самых массовых птиц Северной Америки. По данным американских орнитологов, в штате Висконсин на площади в 2200 км2
гнездилось около 136 млн птиц. В 1810 г. только одна из колоний насчитывала более 2 млн особей. Пока охотой на голубей ради пищи занимались индейцы, численность их не уменьшалась, так как изымалась только небольшая часть популяции. Положение резко изменилось после прибытия европейских переселенцев, имевших огнестрельное оружие. Голубей начали убивать не только ради мяса, но и как вредителей полей. Размах истребления виден из одного только факта, что в штате Мичиган (США) за один лишь сезон было убито 1,5 млн этих птиц. Таким образом, сокращение ареала странствующего голубя и уменьшение плотности его популяций — прямое следствие истребления. Уже в 80-х годах XIX в. отмечалось резкое уменьшение численности голубей, а к 90-м годам они встречались крайне редко. В настоящее время ни одного экземпляра странствующего голубя не осталось ни в природе, ни в зоопарках мира.
Вообще размах прямого уничтожения некоторых видов животных в Северной Америке в прошлом превосходит всякое воображение. Весьма показательна в этом отношении история бизонов. В Северной Америке ко времени появления европейцев насчитывалось не менее 60 млн бизонов — ближайших родственников европейского зубра. С ними было связано все существование индейцев: мясо бизонов служило пищей, шкуры шли на одежду и своеобразные палатки — вигвамы, бизоньи жилы служили нитками. У многих племен бизон был тотемом. В конце 30-х годов XIX в. началось истребление его переселенцами. Только в 1877 г. было убито 100 тыс. бизонов, а к 1889 г. южная популяция их полностью исчезла. В США специальные отряды стрелков ежегодно убивали примерно 2,5 млн бизонов, главным образом для того, чтобы обречь на голодную смерть индейские племена и заставить их покинуть родные земли. В результате от многомиллионных стад в США и Канаде осталось меньше тысячи голов. Лишь в 1905 г. было организовано общество по спасению бизонов. Сейчас они охраняются, и количество их увеличилось до нескольких десятков тысяч голов.
В Азии полностью истреблена дикая популяция оленя милу (сохранился пока только в зоопарках), сведены до минимального количества азиатский лев, островные виды носорогов, дикий двугорбый верблюд, иранская лань. Из птиц исчезли аравийский страус и некоторые виды фазанов, а многие другие представлены столь малым числом особей, что шансов на их выживание в природе крайне мало.
На Африканском материке особенно пострадали крупные представители копытных и хищных. Эти животные беспощадно уничтожались приезжими охотниками. Ежегодно за границу увозили десятки тысяч шкур жирафов, зебр, буйволов и львов, не говоря уже о бивнях слонов. В 30-х годах XX столетия были уничтожены сотни тысяч (по 20-30 тыс. ежегодно) крупных зверей с целью ликвидации сонной болезни в Южной Африке. Однако, как и следовало ожидать, переносчики возбудителя этой болезни — мухи цеце — стали питаться кровью других животных (в том числе птиц и мелких зверьков), и варварская операция закончилась крахом. Массовое истребление животных в Африке производилось также для того, чтобы на освободившихся местах заложить плантации. После Второй мировой войны по этой причине погибли тысячи жирафов, буйволов, носорогов и антилоп. Много животных гибнет в настоящее время от рук браконьеров.
Фауна Южной Америки — одна из самых оригинальных и богатых на земном шаре. Тем не менее и на этом материке хищническое истребление животных привело к сокращению ареалов многих видов. Стала очень редкой шиншилла, обладающая ценным мехом. Уменьшилась численность кошек — ягуара, оцелота, онциллы, добываемых из-за капризной моды на дамские манто. За один только 1967 г. через таможни США прошло 115 458 шкур оцелота. Истребляется гигантская выдра, за шкуру которой платят огромные суммы. Если в 1940-х годах из Перу в США вывозилось до 2 тыс. шкур ежегодно, то в 1960-е годы это число снизилось до 500. Резко сократилось количество лам — викуньи и гуанако. Истребление коснулось очкового медведя, гривистого волка и других зверей. Исчезают южноамериканские кайманы, из кожи которых делают обувь и сумки. Становятся редкими роскошные попугаи.
В Австралии обитают самые примитивные и архаичные млекопитающие земного шара. Фауна этого материка является, в сущности, островной и так же уязвима, как и любая островная фауна. Потери здесь невосполнимы. К началу колонизации Австралии целый ряд животных был уже истреблен. Наиболее известна история сумчатого волка. На материке он исчез до появления европейцев, но на острове Тасмания всего лишь столетие назад животных было еще много. Так как сумчатому волку приписывали истребление овец, его уничтожали любыми средствами. С 1888 по 1914 г. правительство выплачивало премии за каждого убитого зверя, что привело к гибели более 3 тыс. особей. Теперь же за его убийство взимается огромный штраф. Однако после 1961 г. сумчатый волк не регистрируется. Истреблено полностью 4 вида кенгуру; путем больших усилий спасли коалу, уничтожается также дикая собака динго, а из птиц — клинохвостый орел.
Крайне неблагоприятно сложилась судьба и многих животных на других островах. Полностью уничтожены громадные нелетающие птицы моа в Новой Зеландии; дронт и отшельник (солитер) — на Маскаренских островах. Безжалостно истреблялись слоновые черепахи Галапагосских островов. Съедобное мясо их вывозили десятками тонн. В настоящее время они сохранились в очень небольшом количестве и охраняются законом.
Значительно позже началась эксплуатация животных ресурсов Мирового океана. Китобойный промысел нанес океанской фауне трудновосполнимый ущерб. Некоторые виды китов практически уничтожены, популяции других стали настолько малочисленными, что охота на них оказалась уже нерентабельной. Первые китобои выходили в прибрежные части океана на небольших лодках и охотились с помощью ручных гарпунов. В XVII в. не осталось китов в гренландских водах, а в начале XVIII в. хищническая охота англичан, голландцев и датчан привела к исчезновению китов у Шпицбергена. После 1865 г. американцы на китобойных судах вели широкий промысел гладких китов и кашалотов ради китового жира и спермацета. Промысел велся круглый год без ограничения. В 60-е годы XIX в. пушечный гарпун стали начинять взрывчаткой, что ознаменовало начало последнего века китобойного промысла. Под угрозой оказались такие гиганты, как синий кит и финвал, которых до сих пор спасала их величина. С 1904 г. ведется промысел китов в Антарктике.
С 1923 г. норвежцы стали строить плавучие фабрики — суда, которые принимали на борт и полностью перерабатывали туши, доставленные легкими судами-китобойцами. Уже в 60-е годы XX в. в мире было более 250 таких судов, что обеспечивало китобойным флотилиям возможность длительного автономного плавания. В результате только за 30 лет мировая популяция синих китов уменьшилась в 100 раз — до 1 тыс. голов. С 60-х годов началась интенсивная добыча полосатиков. В результате всего этого в северных водах Мирового океана синий кит, горбач, финвал и кашалот стали вымирающими видами. В настоящее время промысел китов запрещен. Пример с китами показывает, что виды даже с космополитными ареалами и высокой численностью в прошлом при интенсивном уничтожении могут исчезать. Но если киты еще имеют шансы сохраниться в Мировом океане как зоологические виды, то другие морские млекопитающие могут вообще исчезнуть без следа, как это произошло всего за 27 лет с морской, или стелле-ровой, коровой у Командорских островов.
Прямое воздействие человека на животный мир выражается в образовании искусственных ареалов путем случайного, неумышленного или, наоборот, планомерного завоза животных в новые районы.
Случайный завоз возможен с морским и сухопутным транспортом. К днищам кораблей прикрепляется и разносится масса морских животных. Таким образом в Черном море появился новый вид усоногих раков — американский морской желудь. Из дальневосточных вод в Балтику попал китайский мохнатый краб. Используя транспортные средства, переселяются сухопутные животные — спутники человека — тараканы, комары, москиты и др. С посадочным материалом неумышленно были развезены по свету многие насекомые-вредители — филлоксера, щитовки, жуки и т. д. Общеизвестны примеры широкого «ассортимента» случайно завезенных животных в крупные морские порты. Неудивительно, что легко расселяющиеся с помощью человека животные нередко имеют космополитное распространение. Они составляют наиболее молодой слой местных фаун.
Все большее значение приобретает сознательный завоз животных в различные страны. Теперь уже трудно перечислить акклиматизированные виды зверей, птиц, рыб, моллюсков и насекомых. Из Южной Америки в Европу завезли нутрию. Сейчас она стала обычным видом в Средней Европе и у нас в России. Прекрасно прижился дикий европейский кролик в Австралии. В Новой Зеландии почти половину фауны составляют интродуцированные виды.
При описании островных фаун уже упоминалось о том, что они имеют дефектную структуру и во многих случаях открыты для вселения новых видов. Прекрасный пример роли человека как фактора, содействующего изменению фаун и расширению ареалов ряда видов, демонстрирует новозеландская биота. По данным Ч.Эл-тона (1960), на Новой Зеландии прижился 31 новый вид птиц и 34 вида млекопитающих, завезенных из Европы, Азии, Австралии, Америки и Полинезии. Остров, не имевший до колонизации сначала полинезийцами, а затем европейцами ни одного вида млекопитающих, получил их в большом количестве. Среди них еж, горностай, хорек, ласка, европейская собака, кошка, черная и серая крысы, мышь, кролик, заяц, серна — из Европы; тар, олени-аксис, замбар — из Азии; американский лось, вапити, вир-гинский олень и мазама — из Америки; опоссум и кенгуру — из Австралии. Кроме того, были завезены многие виды птиц. Так, из Европы завезли диких гусей, крякв, певчих дроздов, грача, зяблика, щегла и даже домового воробья; из Индии — павлина, из Австралии — белого какаду, черного лебедя, один вид попугаев, из Америки — виргинского перепела, калифорнийскую куропатку и канадскую казарку.
Согласно данным А-А.Насимовича (1961), всего было переселено 150 видов зверей из 9 отрядов, главным образом парнокопытных (48 видов), грызунов (36 видов) и хищных (34 вида).
Из беспозвоночных очень интенсивно завозили насекомых с целью борьбы с вредителями-сорняками. Так, в Канаду с 1910 по 1955 г. для борьбы с 68 видами вредных насекомых было переселено около 1 млрд особей хищных и паразитических насекомых, относящихся к 220 видам. В фауну видов европейской части России совсем недавно вошел амброзиевый листоед — жук, завезенный из Канады для уничтожения карантинного сорняка американского происхождения — амброзии.
Обогащается также морская и пресноводная фауна благодаря завозу ценных видов рыб, ракообразных и моллюсков. К примеру, в Европу завезли американскую радужную форель, а в реки Северной Америки — европейскую форель. Черноморская кефаль переселена в Каспийское море вместе со своими кормовыми объектами — червями и моллюсками. Дальневосточные лососи успешно прижились у берегов Европы, т. е. в Атлантике.
К сожалению, переселение животных не всегда проводилось с учетом их экологии. Это нередко оканчивалось вытеснением местных видов пришельцами, особенно на островах. Сильно пострадала оригинальная фауна Новой Зеландии, Австралии, островов Карибского моря, где более конкурентоспособные обитатели Европы и Северной Америки вытеснили примитивных аборигенов, имевших сходные экологические ниши.
Косвенное воздействие человека на фауну земного шара особенно проявляется в наше время. Сводится оно к нарушению привычных, сложившихся в течение истории вида, экологических связей со средой (пищевых, территориальных, биоценотических). Вырубка лесных массивов, распашка целинных земель, постройка населенных пунктов, введение в круговорот веществ на Земле отходов промышленного производства — основные факторы косвенного антропического влияния.
Сплошная вырубка лесов практиковалась еще в доисторические времена как в Европе, так и на Ближнем Востоке. В результате лесной фауне нанесен невосполнимый ущерб — многие виды исчезли навсегда, ареалы других фрагментировались, площадь, занимаемая животными, становилась меньше, что лишало их нормальных условий существования и приводило к вымиранию изолированных популяций. Из-за вырубки лесов в Европе и Северной Америке исчезли многие популяции медведей, оленей, белок, дятлов, огромного количества видов лесных беспозвоночных. Сокращение лесных массивов в тропической зоне земного шара угрожает гибелью тысячам видов, которые не могут жить вне леса. В первую очередь это лемуры, лесные виды обезьян, азиатские фазаны, вест-индские попугаи, африканские турако, американские туканы. По существу, речь идет о лесной фауне тропиков в целом. Распашка целинных степей в Европе и Азии, прерий в Северной Америке настолько изменила облик животного мира этих районов, что о существовании в наше время коренной степной фауны говорить уже не приходится. Уменьшается площадь болот, а вместе с ними исчезают болотные и околоводные птицы. Сокращение ареала белого и черного аиста в Европе вызвано именно этой причиной.
Вторая половина XX
в. принесла животному миру земного шара еще большую опасность. Имеется в виду загрязнение биосферы химическими веществами — отходами промышленного производства, пестицидами, радиоактивными соединениями и др. Эти вещества особенно опасны для животных, так как они являются конечными звеньями пищевых цепей.
Особенно страдают от загрязнения обитатели рек, озер и морей. К примеру, на американском побережье Атлантики в 1963 г. по этой причине в реке Потомак погибло 9 млн рыб. Участок океана в 50 км2
около Нью-Йорка называют мертвым морем, потому что все живое в нем погибло. Нефтяное загрязнение Мирового океана достигло огромных размеров. Особенно страдают от него уникальные биоценозы коралловых рифов.
Подобных примеров, к сожалению, немало, и перечислять их нет необходимости, однако следует сказать, что нарушение экологических законов приводит к кризисным ситуациям в биосфере, от которых в конечном счете страдает все человечество.
Число непрерывно исчезающих видов не уменьшается, о чем свидетельствует Международная красная книга. На 1 января 1979 г. в ней насчитывалось 321 вид зверей, 485 видов птиц, 181 вид амфибий и рептилий, многие виды рыб. Внесение вида в Красную книгу — сигнал опасности и одновременно призыв к спасению животных. В нашей стране также была создана Красная книга, где в 1984 г. числилось 94 вида и подвида млекопитающих, 80 видов птиц, 37 видов пресмыкающихся, 9 видов земноводных и 9 видов рыб.
Сохранение животного мира — неотложная проблема, которая должна решаться в глобальном масштабе. Пока она довольно успешно разрешена лишь в отдельных странах. Животный мир можно сохранить только в его естественном окружении. С этой целью создаются заповедники. Первые охраняемые «парки» известны еще из глубокой древности. Л. К. Шапошников (1969) приводит сведения о 720 заповедниках, национальных парках и других охраняемых участках, существовавших к началу 70-х годов XX в. Число их увеличивается.
Основная
Адольф Т.Д., Бутьев В. Т., Михеев А. В., Орлов В.И.
Руководство к лабораторным занятиям по зоологии позвоночных. — М., 1977.
Беклемишев К. В.
Зоология беспозвоночных: Курс лекций. — М., 1979.
Бобринский И. А.
География животных. — М., 1951.
Бобринский Н.А., Зенкевич Л. А., БирштейнЯ.А.
География животных. — М., 1946.
Воронов А. Г.
Биогеография. — М., 1963.
Гептнер В. Г.
Общая зоогеография. — М., 1936.
Дарлингтон Ф.
Зоогеография: Пер. с англ. / Под ред. Н.А. Гладкова. — М.: Прогресс, 1966.
Догель В. А.
Зоология беспозвоночных. — 7-е изд. — М., 1981.
Зеликман А. А.
Малый практикум по зоологии беспозвоночных. — М 1965.
Карташев Н.Н., Соколов В.Е., Шилов И. А.
Практикум по зоологии позвоночных. — М., 1981.
Лопатин И. К.
Зоогеография. — Минск, 1989.
Лопатин И. К.
Общая зоология. — Минск, 1983.
Лопатин И. К.
Основы зоогеографии. — Минск, 1980.
Натали В. Ф.
Зоология беспозвоночных. — М., 1975.
Наумов Н. П., Карташев Н. Н.
Зоология позвоночных: В 2 ч. — М., 1986.
Пузанов И. И.
Зоогеография. — М., 1938.
Фролова Е.Н., Щербина Т. В., Михина Т.Н.
Практикум по зоологии беспозвоночных — М., 1985.
Шарова И.Х., Абдурахманов Г.М., Матвеева И.Г.
Зоология беспозвоночных. — М., 1993.
Шарова И.Х.
Зоология беспозвоночных. — М., 1999.
Дополнительная
Абдурахманов Г. М., Исмаилов Ш. И., Лобанов А. Л.
Новый подход к проблеме объективного зоогеографического районирования. — Махачкала, 1995.
Бей-Биенко Г.Я.
Общая энтомология. — М., 1980.
Беклемишев В. Н.
Основы сравнительной анатомии беспозвоночных. —
М., 1964.-Т. 1,2.
Второе П.П., Дроздов Н.Н.
Биогеография материков. — М., 1974.
Иванов А. В.
Происхождение многоклеточных. — Л., 1968.
Крыжановский О. Л.
К вопросу о предмете зоогеографии и методах зоогео-
графических исследований // Журнал общей биологии. — 1976. — Т. 37. —
Вып. 4.
Крыжановский О. Л. О
принципах зоогеографического районирования суши // Зоологический журнал. — 1976. — Т. 55. — Вып. 7.
Леме Ж.
Основы биогеографии: Пер. с фр. / Под ред. А. Г. Воронова. — М., 1976.
Лобье Л.
Оазисы на дне океана / Пер. с фр. — Л., 1990.
Росс Г., Росс Ч., Росс Д.
Энтомология. — М., 1985.
Серавин Л.Н.
Простейшие... Что это такое? — Л., 1984.
Тарасов В.В.
Простейшие патогенные для человека. — М., 1987.
Хаусман К.
Протозоология. — М., 1988.
Чернов Ю.И.
Природная зональность и животный мир суши. — М., 1975.
СОДЕРЖАНИЕ
ЧАСТЬ I 3
ОСНОВЫ зоологии.. 3
Введение. 3
Общие сведения о зоологии.. 3
Роль животных в биогенном круговороте 7
и значение в жизни биосферы.. 7
Роль животных в жизни и хозяйстве человека.. 10
Из истории зоологии.. 15
Планы строения животных.. 19
Система животного мира.. 21
ПОДЦАРСТВО ПРОСТЕЙШИЕ,
ИЛИ ОДНОКЛЕТОЧНЫЕ (PROTOZOA)22
Тип саркомастигофоры
(SARCOMASTIGOPHORA)24
Подтип саркодовые (SARCODINA)24
Надкласс корненожки (RHIZOPODA)24
Надкласс актиноподы (ACTINOPODA)27
Подтип жгутиконосцы (MASTIGOPHORA)29
Класс растительные жгутиконочцы
(PHYTOMASTIGOPHOREA)30
Класс животные жгутиконосцы
(ZOOMASTIGOPHOREA)31
Отряд воротничковые
жгутиконосцы (CHOANOFLAGELLIDA)31
Отряд кинетопластиды (KINETOPLASTIDA)31
Отряд дипло (DIPLOMONADIDA)33
Отряд трихомонадидыTRICHOMONADIDA)33
Отряд гипермастигиды (HYPERMASTIGIDA)33
Подтип опалины (OPALINATA)33
Класс опалины (OPALINATEA)33
Тип апикомплексы (APICOMPLEXA)33
Класс споровики (SPOROZOEA)33
Подкласс грегарины (GREGARINIA)35
Подкласс кокцидии (COCCIDIA)36
Отряд кокцидии (COCCIIDA)37
Отряд кровяные споровики (HAEMOSPORIDIA)37
Тип инфузории, или ресничные
(CILIOPHORA, или INFUSORIA)40
ПОДЦАРСТВО МНОГОКЛЕТОЧНЫЕ
(METAZOA)44
Происхождение многоклеточных.. 44
Принципы классификации многоклеточных.. 51
Надраздел паразои (PARAZOA)53
Тип губки (PORIFERA, ИЛИ SPONGIA)53
Надраздел эуметазои (EUMETAZOA)57
Раздел лучистые (RADIALIA)58
Тип кишечнополостные (COELENTERATA)58
Класс гидрозои (HYDROZOA)58
Класс сцифоидные (SCYPHOZOA)62
Класс коралловые полипы (ANTHOZOA)63
Тип гребневики (CTENOPHORA)65
Класс гребневики (CTENOPHORA)66
Раздел билатеральные (BILATERIA)67
Подраздел первичноротые (PROTOSTOMIA)67
Тип плоские черви (PLATHELMINTHES)67
Класс моногенеи, или моногенетические
сосальщики (MONOGENEA)71
Класс трематоды,
или сосальщики (TREMATODA)71
Класс ленточные черви (CESTODA)73
Тип круглые червм (NEMATHELMINTHES)76
Класс собственно круглые черви,
или нематоды (NEMATODA)76
Тип кольчатые черви (ANNELIDA)79
Класс многощетинковые, или полихеты
(POLYCHAETA)82
Класс малощетинковые (OLIGOCHAETA)84
Класс пиявки (HIRUDINEA)85
Ттип членистоногие (ARTHROPODA)86
Подтип жабродышащие (BRANCHIATA)87
Класс ракообразные (CRUSTACEA)87
Подтип хелицеровые (CHELICERATA)92
Класс паукообразные (ARACHNIDA)93
Подтип трахейные (TRACHEATA)99
Надкласс многоножки (MYRIAPODA)100
Надкласс шестиногие (HEXAPODA)100
Класс скрыточелюстные насекомые. 101
(INSECTA-ENTOGNATHA)101
Класс открыточелюстные насекомые
(INSECTA-ECTOGNATHA)102
Отряд стрекозы (ODONATA)109
Отряд таракановые (BLATTOPTERA)109
Отряд термиты (ISOPTERA)109
Отряд прямокрылые (ORTHOPTERA)109
Отряд равнокрылые (HOMOPTERA)110
Отряд полужесткокрылые, или клопы (HETEROPTERA)110
Отряд жесткокрылые, или жуки (COLEOPTERA)110
Отряд сетчатокрылые (NEUROPTERA)111
Отряд чешуекрылые, или бабочки (LEPIDOPTERA)111
Отряд перепончатокрылые (HYMENOPTERA)112
Отряд двукрылые, или мухи (DIPTERA)113
Тип моллюски (MOLLUSCA)114
Подтип боконервные (AMPHINEURA)117
Класс панцирные (POLYPLACOPHORA)117
Подтип раковинные (CONCHIFERA)117
Класс моноплакофоры (MONOPLACOPHORA)118
Класс брюхоногие (GASTROPODA)118
Класс двустворчатые (BIVALVIA)120
Класс головоногие (CEPHALOPODA)123
Тип погонофоры (POGONOPHORA)126
Подраздел вторичноротые животные
(DEUTEROSTOMIA)132
Тип иглокожие (ECHINODERMATA)132
Подтип асторозои (ASTEROZOA)133
Класс морские звезды (ASTEROIDEA)133
Подтип эхинозои (ECHINOZOA)137
Класс морские ежи (ECHINOIDEA)137
Класс голотурии (HOLOTHUROIDEA)138
Подтип кринозои (CRINOZOA)139
Класс морские лилии (CRINOIDEA)139
Тип полухордовые (HEMICHORDATA)141
Класс кишечнодышащие (ENTEROPNEUSTA)142
Класс крыложаберные (PTEROBRANCHIA)142
Тип хордовые (CHORDATA)142
Подтип бесчерепные (ACRANIA)143
Класс головохордовые (CEPHALOCHORDATA)143
Подтип личиночнохордовые, или оболочники.. 147
(UROCHORDATA, или TUNICATA)147
Класс асцидий (ASCIDIAE)149
Класс сальпы (SALPAE, или THALIACEA)149
Класс аппендикулярии
(APPENDICULARIAE, или LARVACEA)150
Подтип позвоночные, или черепные. 151
(VERTEBRATA, или CRANIATA)151
Надкласс бесчелюстные (AGNATHA)154
Подтип челюстноротые (GNATHOSTOMATA)161
Надкласс рыбы (PISCES)161
Класс хрящевые рыбы (CHONDRICHTHYES)163
Класс костные рыбы (OSTEICHTHYES)169
Подкласс лучеперые рыбы (ACTINOPTERYGII)179
Надотряд ганоидные (GANOIDOMORPHA)179
Отряд осетрообразные (ACIPENSERIFORMES)180
Костистые рыбы (TELEOSTEI)180
Отряд сельдеобразные (CLUPEIFORMES)181
Отряд лососеобразные (SALMONIFORMES)181
Отряд углеобразные (AJNGUILLIFORMES)181
Отряд карпообразные (CYPRINIFORMES)181
Отряд сомообразные (SILURIFORMES)182
Отряд трескообразные (GADIFORMES)182
Отряд кефалеобразные (MUGILIFORMES)182
Отряд окунеобразные (PERCIFORMES)182
Отряд камбалообразные (PLEURONECTIFORMES)182
Подкласс лопастеперые рыбы
(SARCOPTERYGII)183
Надотряд кистеперые рыбы (CROSSOPTERYGIMORPHA)183
Надотряд двоякодышащие рыбы (D1PNEUSTOMORPHA)184
Надкласс четвероногие (TETRAPODA)184
Класс земноводные или амфибии (AMPHIBIA)185
Отряд хвостатые амфибии (URODELA, или CAUDATA)197
Отряд безногие амфибии (APODA)197
Отряд бесхвостые амфибии (ANURA)197
Класс пресмыкающиеся, или рептилии (REPTILIA)198
Подкласс черепахи CHELONIA, или ESTUDINES)208
Подкласс архозавры (ARCHOSAURIA)208
Отряд крокодил (CROCODILIA)208
Подкласс лепидозавры (LEPIDOSAURIA)208
Отряд клювоголовые (RHYNCHOCEPHALIA)208
Отряд чешуйчатые (SQUAMATA)209
Подотряд ящерицы (SAURIA)209
Подотряд змеи (OPHIDIA, или SERPENTES)209
Класс птицы (AVES)210
Надотряд плавающие (IMPENNES)225
Надотряд типичные, или новонёбные
птицы (NEOGNATHAE)225
Отряд страусообразные (STRUTHIONIFORMES)225
Отряд нандуобразные (RHEIFORMES)226
Отряд кивиобразные (APTERYGIFORMES)226
Отряд курообразные (GALLIFORMES)226
Отряд голубеобразные (COLUMBIFORMES)226
Отряд журавлеобразные (GRUIFORMES)227
Отряд ржанкообразные (CHARADRIIFORMES)227
Отряд гусеообразные (ANSERIFORMES)227
Отряд голенастные, или
листообразные (CICONIIFORMES)228
Отряд соколообразные, или хищные
птицы (FALCONIFORMES)228
Отряд совообразные (STRIGIFORMES)228
Отряд кукушкообразные (CUCULIFORMES)228
Отряд ракшеобразные(CORACIIFORMES)229
Отряд дятлообразные (PICIFORMES)229
Отряд воробьинообразные (PASSERIFORMES)229
Класс млекопитающие, или
звери (MAMMALIA, или THERIA)230
Подкласс клоачные,
или первозвери (PROTOTHERIA)247
Подкласс живородящие млекопитающие,
или настоящин вери (THERIA)248
Иинфракрасс сумчатые,
или низшие звери (METATHERIA)248
Инфракласс высшие звери,
или плацентарные (EUTHERIA, или PLACENTALIA)249
Отряд насекомоядные (INSECTIVORA)250
Отряд шерстокрылы (DERMOPTERA)250
Отряд рукокрылые (CHIROPTERA)250
Отряд зайцеобразные (LAGOMORPHA)250
Отряд грызуны (RODENTIA)251
Отряд хищные (CARNIVORA)251
Отряд ластоногие (PINNIPEDIA)252
Отряд китообразные (СЕТАСЕА)253
Отряд даманы (HYKACOIDEA)254
Отряд хоботные (PROBOSCIDEA)254
Отряд сирены (SIRENIA)255
Отряд непарнокопытные (PERISSODACTYLA)255
Отряд мозоленогие (TYLOPODA)256
Отряд парнокопытные (ARTIODACTYLA)257
Подотряд нежвачные
(NONRUMINANTIA, или SUIFORMES)257
Подотряд жвачные (RUMINANTIA)258
Отряд приматы (PRIMATES)260
Подотряд полуобезьяны (PROSIMII)260
Подотряд человекоподобные приматы,
или обезьяны (ANTHROPOIDEA, или SIMIA)261
ЧАСТЬ II
ОСНОВЫ ЗООГЕОГРАФИИ
ОБЩАЯ ХАРАКТЕРИСТИКА
ЗООГЕОГРАФИИ.. 263
Основные разделы зоогеографии.. 263
Связи зоогеографии с другими науками.. 264
Цели и задачи зоогеографии.. 265
ОЧЕРК РАЗВИТИЯ ЗООГЕОГРАФИИ.. 267
БИОСФЕРА.. 279
Факторы среды.. 280
Экологическая толерантность животных.. 280
Условия существования и распространения
животных в море. 281
Условия существования и распространения
животных в пресных водах.. 285
Условия существования и распространения
наземных животных.. 287
АРЕАЛ.. 299
Общие сведения об ареале. 299
Картирование ареалов. 299
Типология ареалов. 302
Расселение животных.. 316
Центры распространения и
происхождения видов. 323
ПОНЯТИЕ О ФАУНЕ.. 325
Структура фауны.. 326
Сравнительный анализ фаун.. 327
Эндемизм.. 327
Генезис фуны.. 329
Островные фауны.. 329
ПРИНЦИПЫ И МЕТОДЫ... 333
ЗООГЕОГРАФИЧЕСКОГО РАЙОНИРОВАНИЯ.. 333
Новый подход к проблеме объективного.. 334
зоогеографического районирования. 334
ЗООГЕОГРАФИЧЕСКОЕ ДЕЛЕНИЕ
МИРОВОГО ОКЕАНА.. 337
Роль океанов в биосфере и их значение
для человека.. 337
Фаунистическое расчленение литорали.. 339
Тропический регион. 341
Бореальный регион. 342
Антибореальный регион. 345
Фаунистическое расчленение пелагиали.. 347
Тропический регион. 347
Бореальный регион. 349
Антибореальный регион. 351
ЗООГЕОГРАФИЧЕСКОЕ РАЙОНИРОВАНИЕ
КОНТИНЕНТАЛЬНЫХ ВОДОЕМОВ.. 352
ЗООГЕОГРАФИЧЕСКОЕ ДЕЛЕНИЕ СУШИ.. 358
Краткая характеристика.. 358
Царство Палеогея. 359
Эфиопская область. 360
Мадагаскарская область. 367
Индо-Малайская область. 370
Полинезийская область. 376
Царство Арктогея. 378
Палеарктическое подцарство.. 379
Европейско-Сибирская область. 382
Область Древнего Средиземья. 398
Восточно-Азиатская, или. 411
Гималайско-Китайская, область. 411
Неарктическое подцарство.. 416
Царство Неогея. 421
Неотропическая область. 422
Карибская, или Антильская, область. 430
Царство Нотогея. 431
Австралийская область. 431
Новозеландская область. 438
Патагонская, или Голантарктическая, область. 440
АНТРОПИЧЕСКОЕ ВОЗДЕЙСТВИЕ
НА ФАУНУ ЗЕМНОГО ШАРА.. 442
ЛИТЕРАТУРА.. 450
[1]
Анаморфоз — развитие, при котором из яйца выходит личинка с неполным числом члеников, восполняемых в ходе нескольких линек.
[2]
Авторы намеренно используют термин «антропический фактор», считая его более подходящим, чем термин «антропогенный фактор».
[3]
Биом —
совокупность организмов и среды в определенной ландшафтно-географической зоне.
[4]
Биота
— совокупность организмов, объединенных общей областью распространения.
[5]
Согласно правилу Бергманна, у гомойотермных животных одного вида или группы близких видов размеры тела больше в холодных частях ареала, а в более теплых — меньше. Эти изменения связаны с широтой и с высотой местности.
[6]
В этом царстве нередко выделяют только одну область - Голарктическую, подразделяемую на подобласти.
|
