|
Реферат
«Модификационная и мутационная изменчивость»
1.
Модификационная изменчивость
Различные признаки организма неодинаково реагируют на изменение внешних условий. Одни из них очень пластичны и изменчивы, другие менее изменчивы, наконец, третьи лишь в очень малой степени могут быть изменены условиями среды. У рогатого скота удой во многом зависит от кормления и ухода. Хорошо известно, что удой можно значительно повысить подбором нужных количеств кормов и т. п. От условий кормления и содержания в меньшей степени, чем количество молока, зависит его жирномолочность. Процент жира в молоке — довольно постоянное свойство породы, хотя изменением пищевого рациона его тоже удается несколько изменить. Гораздо более постоянным признаком является масть. При самых различных условиях она почти не изменяется. Не следует, однако, думать, что окраска шерсти совсем не зависит от условий развития. У некоторых млекопитающих на окраску шерсти влияет температура окружающей среды. Например, порода горностаевых кроликов характеризуется тем, что при обычных условиях большая часть шерсти кролика белая, черная шерсть развивается лишь на ушах, лапах и хвосте. Если выбрить или выщипать шерсть на спине, то при положительных температурах опять вырастает белая шерсть. Но если кролика держать при низкой температуре (около 0°), то вместо белой вырастет черная шерсть.
Можно сказать, что наследственна в данном примере способность развивать белую шерсть на спине при высоких температурах и черную — при низких.
Норма реакции.
У организмов проявление действия генов и генотипа в целом зависит от условий среды. Эта форма изменчивости, не связанная с изменением генотипа, носит название модификационной.
Пределы модификационной изменчивости, для разных признаков и при разных условиях, как это показано на рассмотренных примерах, могут быть очень различными. Пределы модификационной изменчивости признака называют его нормой реакции.
Одни признаки (например, молочность) обладают очень широкой нормой реакции, другие (окраска шерсти) — гораздо более узкой.
На основе только что рассмотренных фактов мы можем углубить представление о сущности явления наследственности. Наследуется не признак, как таковой, а способность организма (его генотипа) в результате взаимодействия с условиями развития давать определенный фенотип.
Не существует, например, наследственного признака какой-либо породы рогатого скота давать 4000 л
молока в год. Этот признак выявляется лишь при определенном режиме кормления и содержания.
Реклама

Широкая норма реакции (широкая приспособляемость) в природных условиях может иметь важное значение для сохранения и процветания вида. Однако отклонения, вызванные внешними условиями, не изменяют генотипа, они лежат в пределах нормы его реакции.
Управление доминированием.
Доминантный и рецессивный признаки определяются, прежде всего, особенностями гена и его влиянием на развитие признака. Поскольку, однако, фенотип всегда зависит от двух начал: генотипа и условий развития, то можно ожидать, что, меняя условия развития, можно воздействовать на характер доминирования признака у гибридов. Наиболее полно вопрос о возможности управления доминированием и направлением индивидуального развития организма был разработан на плодово-ягодных растениях И. В. Мичуриным. Мичурин установил важную закономерность доминирования признаков у гибридов. На ряде примеров по гибридизации плодовых деревьев он показал, что у гибридов преимущественно доминируют те признаки, которые в окружающей среде встречают наиболее благоприятные условия для своего развития.
Проводя, например, многочисленные скрещивания западноевропейских и американских сортов плодовых деревьев (из стран с мягким климатом) с местными сортами из Тамбовской области (с суровым континентальным климатом), Мичурин выращивал гибриды не в оранжереях, а в условиях открытого грунта. В этих условиях проявлялось доминирование признаков высокой зимостойкости, свойственной местным сортам.
Статистические закономерности модификационной изменчивости.
В течение всей жизни, от момента оплодотворения и до самой смерти, организмы подвергаются действию самых разнообразных условий среды. Нельзя представить себе двух растений одного вида, произрастающих, например, на лугу или в лесу, условия жизни, которых были бы совершенно одинаковы. Поскольку эти условия никогда не бывают совершенно сходными, фенотипы разных особей тоже не вполне тождественны. Если мы измерим длину и ширину листьев, взятых с одного дерева, то увидим, что размеры их варьируют в довольно широких пределах. Эта изменчивость — результат разных условий развития листьев на ветвях дерева; генотип их одинаков. Если некоторое количество листьев расположить в порядке нарастания или убывания признака (например, длины), то получатся ряд изменчивости данного признака, который носит название вариационного ряда.
Этот ряд слагается, как мы видим, из отдельных вариант. Как часто встречаются отдельные члены вариационного ряда? Если мы подсчитаем число отдельных вариант в вариационном ряду, то увидим, что частота встречаемости их неодинакова. Чаще всего будут встречаться средние члены вариационного ряда, а к обоим концам ряда частота встречаемости будет снижаться.
Реклама

Рассмотрим это на примере изменчивости числа колосков в колосе пшеницы. Подсчитывая число колосков в разных колосьях, устанавливаем, что это число варьирует от 14 до 20. Возьмем, не выбирая подряд 100 колосьев и определим частоту встречаемости разных вариант. Мы увидим, что чаще всего встречаются колосья со средним числом колосков (16—18), реже — с большим или меньшим числом их. Вот результат одного из таких подсчетов:
число колосков в колосе 14 15 16 17 18 19 20
количество колосьев 2 7 22 32 24 8 5
Верхний ряд цифр — выписанные в ряд варианты от наименьшей к наибольшей. Нижний ряд — частота встречаемости каждой варианты. Если сложить ряд нижних цифр, мы получим 100, что и соответствует числу отдельных наблюдений. Распределение вариант в вариационном ряду можно выразить наглядно на графике, который построим следующим образом. По горизонтали (на оси абсцисс) отложим на одинаковых расстояниях отдельные варианты в нарастающем порядке. На оси ординат нанесем числа, соответствующие частоте встречаемости каждой варианты в произвольно выбранном масштабе. На горизонтальной оси восстановим перпендикуляры до уровня, соответствующего частоте встречаемости каждой варианты. Точки пересечения перпендикуляров с линиями, соответствующими частоте встречаемости вариант, соединим прямыми. Получим кривую, которая выражает изменчивость числа колосков в колосе пшеницы. Наиболее высокая точка кривой изменчивости соответствует варианте с наибольшей частотой встречаемости. Графическое выражение изменчивости признака, отражающее как размах вариаций, так и частоту встречаемости отдельных вариант, называют вариационной кривой.
Какими же биологическими причинами вызвано такое распределение вариант в вариационном ряду? Причина этого — внешняя среда и воздействие ее на организм. С самого начала жизни, в течение всего периода развития и до самой смерти каждый организм подвергается действию самых различных факторов среды. Среди семян пшеницы, высеянных на поле, нельзя найти два семени, развитие которых протекало бы в совершенно одинаковых условиях. Глубина заделки в почву, физические свойства почвы, взаимодействие и конкуренция с соседними растениями, увлажнение, освещенность и т. п. — все это варьирует в различных направлениях и отражается на развитии фенотипа. Для того чтобы получить крайнее (наиболее выраженное или наиболее слабое) развитие признака, нужно, чтобы все эти многочисленные факторы среды действовали примерно в одном направлении. Чтобы получить большой колос с многими колосками, нужно, чтобы вся сумма факторов оказалась наиболее благоприятной из всех возможных сочетаний условий среды. На самом деле большинство растений испытывает воздействия различного характера. Одни благоприятствуют развитию признака, другие задерживают его. При этом фенотип их оказывается где-то среди средних вариант вариационного ряда. Чем больше стандартизированы условия развития, тем меньше выражена модификационная изменчивость, тем короче будет вариационный ряд. Чем разнообразнее условия среды, тем шире модификационная изменчивость. Размах вариаций зависит и от генотипа. Признаки с широкой нормой реакции дают широкий размах изменчивости, а признаки с узкой нормой реакции
— незначительный размах.
Чтобы дать объективную характеристику изменчивого признака, недостаточно ограничиться изучением одной или немногих особей. Нужно установить размах изменчивости и построить вариационную кривую. Выражением развития признака является его средняя величина. Она вычисляется как средняя арифметическая всего вариационного ряда. Численное выражение признака для каждой варианты умножают на число вариант.
Все эти произведения складывают и затем делят на общее число вариант. Это может быть выражено следующей формулой:
М = 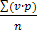 , ,
где М – средняя величина, v – вариант, p - частота встречаемости варианты, s — знак суммирования и n — общее число вариант вариационного ряда. Вычислим по этой формуле среднюю величину для приведенного выше вариационного ряда числа колосков пшеницы. Умножим каждую варианту на частоту ее встречаемости. Это составит:
14X2 = 28; 15X7=105; 16X22 = 352; 17X32 = 544; 18X24=432; 19X8 = 152; 20X5=100.
Далее суммируем все эти произведения, что дает в данном случае 1713. Разделив эту сумму на общее число вариант ряда, которое равняется 100, получим среднюю величину, равную 17,13.
2.
Мутационная изменчивость
Разнообразные формы и проявления модификационной изменчивости не затрагивают генотипа организма. Наряду с модификациями существует другая форма изменчивости, меняющая генотип.
Эту форму изменчивости называют генотипической или мутационной, а отдельные изменения — мутациями.
Существование наследственных изменений было известно Дарвину. Вся его теория эволюции вытекает из учения о естественном отборе наследственных изменений. Наследственная изменчивость — необходимая предпосылка естественного и искусственного отбора. Однако во времена Дарвина еще отсутствовали опытные данные по наследственности и законы наследования не были известны. Это не давало возможности строго различать разные формы изменчивости в зависимости от наследования.
Понятие мутаций было введено в науку голландским ботаником де Фризом. У растения ослинник (энотера) он наблюдал появление резких скачкообразных отклонений от типичной формы растения, причем эти отклонения оказались наследственными. Дальнейшие исследования на различных объектах— растениях, животных, микроорганизмах — показали, что явление наследственной (мутационной) изменчивости свойственно всем организмам. Мутации затрагивают разнообразные стороны строения и функции организма. Например, у дрозофилы известны мутационные изменения формы крыльев (вплоть до полного их исчезновения), окраски тела, развития щетинок на теле, формы глаз, их окраски (красные, желтые, белые, вишнёвого цвета и т. п.), а также многих физиологических признаков (продолжительность жизни, плодовитость, стойкость к разным повреждающим воздействиям и т. п.). Первоначальные представления де Фриза о том, что мутации всегда крупные наследственные изменения, дальнейшими исследованиями не подтвердились. Наряду с резкими отклонениями гораздо чаще встречаются небольшие мутации, лишь немногим отличающиеся от исходных форм. Тем не менее, указанный еще де Фризом признак мутаций — их скачкообразный характер и наследственность — остается в силе. Мутации совершаются в различных направлениях, и обычно они не являются приспособительными, полезными для организма изменениями.
Существуют и такие наследственные изменения, которые в гомозиготном состоянии вызывают гибель (такие мутации называются летальными).
Генные мутации.
Различают несколько типов мутаций по характеру изменений хромосомного аппарата. Наиболее распространенными являются мутации, не связанные с видимыми в микроскоп изменениями строения хромосом. Такие мутации представляют собой качественные изменения отдельных генов и носят название генных мутаций.
На основании исследований, проведенных главным образом на микроорганизмах за последнее время, установлено, что такие мутации связаны с преобразованием химической структуры ДНК, входящей в состав хромосом. Последовательность оснований определяет состав образующейся на ДНК
молекулы РНК, а она в свою очередь обусловливает последовательность аминокислот при синтезе белковой молекулы. Химическая основа генных мутаций заключается в изменении расположения нуклеотидов в цепочке ДНК.
Хромосомные мутации.
Наряду с генными известен ряд других мутаций, связанных с видимыми преобразованиями хромосом, которые доступны непосредственному микроскопическому изучению. К числу таких изменений относится, например, переход части одной хромосомы на другую, ей не гомологичную, поворот участка внутри хромосомы на 180° и ряд других структурных изменений отдельных хромосом.
Особую группу мутаций представляют собой изменения числа хромосом. Эти мутации сводятся к появлению лишних или утере некоторых хромосом. Такого рода изменения в хромосомном составе происходят при нарушении в силу каких-либо причин нормального хода мейоза, когда вместо нормального распределения хромосом между полюсами ахроматинового веретена и затем дочерними клетками обе гомологичные хромосомы отходят к одному полюсу. Обычно такого рода нарушения оказываются неблагоприятными, снижающими жизнеспособность.
Особый тип наследственных изменений представляет собой явления полиплоидии
, которое выражается в кратном увеличении числа хромосом. Возникновение полиплоидов обычно связано с нарушением процессов митоза или мейоза. Эти нарушения сводятся к тому, что хромосомы проделывают нормальный митотическйй цикл, а веретено деления, служащее для «растаскивания» хромосом к противоположным полюсам, не функционирует. В результате хромосомы не расходятся к полюсам и не образуют дочерних ядер, а остаются в том же ядре. Если этот процесс имеет место в соматической клетке с диплоидным набором хромосом, то сразу возникает клетка с удвоенным диплоидным набором (такие клетки называются тетраплридными). Они имеют, следовательно, вместо двух гаплоидных наборов четыре (4 n).
Если это наблюдается при мейозе, то конъюгирующие гомологичные хромосомы не расходятся к противоположным полюсам и возникают диплоидные гаметы. Если такая гамета при оплодотворении сольется с нормальной гаплоидной, то возникает триплоидная зигота (с тройным набором хромосом— 3 n).
Если же обе гаметы окажутся диплоидными, то возникает тетраплоидная зигота.
Полиплоидные виды довольно часто наблюдаются у растений и очень редко встречаются у животных. Полиплоидные мутации у растений по сравнению с диплоидами часто характеризуются более мощным ростом, большим размером и весом семян и плодов и т.п. В генетико-селекционной работе по созданию высокопродуктивных сортов растений явление полиплоидии широко используется на практике.
В настоящее время разработаны методы, позволяющие получать полиплоиды экспериментально, воздействуя на делящуюся клетку некоторыми ядами (например, колхицином), разрушающими веретено, но не препятствующими удвоению хромосом.
Мы видим, таким образом, что мутации всегда связаны с изменениями в хромосомах. Если эти изменения происходят в половых клетках, то они проявляются в том поколении, которое развивается из половых клеток. Но изменения могут иметь место и в соматических клетках. Тогда они получают название соматических мутаций. Такие мутации приводят к изменению признака только части организма, развивающегося из измененных клеток.
У животных соматические мутации не передаются последующими поколениям, поскольку из соматических клеток новый организм не возникает. Другое дело у растений. При помощи отводков и прививок иногда удается сохранить возникшее изменение, и оно оказывается стойким, наследственным.
Частота и причины мутаций.
Как часто происходят мутациии каковы причины их возникновения? Прежде чем ответить наэтот вопрос, нужно иметь в виду, что учет возникающих мутаций представляет очень большие трудности. Большинство мутаций рецессивны. Они возникают в генах, локализованных в хромосомах половых клеток. Гамета, несущая вновь возникшую рецессивную мутацию, при оплодотворении обычно соединяется с гаметой, которая такой же мутации не несет. Поэтому вновь возникшая рецессивная мутация фенотипические не проявится. Однако в последующих поколениях она будет размножаться вместе с несущей ее хромосомой и распространяться среди особей данного вида. Лишь когда соединятся две гаметы, несущие одну и ту же рецессивную мутацию, она проявится фенотипически.
Исследования показали, что в природных условиях мутация каждого отдельно взятого гена происходит очень редко. На первый взгляд может возникнуть представление, что такая малая изменчивость гена не может дать достаточного материала наследственной изменчивости для естественного отбора. На самом деле это не так. У организма имеется несколько тысяч генов, так что общее число мутаций оказывается значительным. Для той же дрозофилы, например, высчитано, что около 5% ее гамет несут какую-нибудь мутацию. Прямые исследования распространения мутаций в природных популяциях дрозофилы, проведенные в разных географических зонах, показали, что они «насыщены» разнообразными мутациями, большинство которых, однако, в силу рецессивности не проявляется видимо. Значительная стойкость гена имеет большое биологическое значение. Действительно, если бы гены легко и часто изменялись, то существование видов стало бы невозможным, ибо в каждом поколении организмы превращались бы в нечто совершенно новое, не похожее на родителей. Относительная стойкость видов — важное условие приспособления организма к окружающей среде.
Способность к мутированию — одно из основных свойств гена. Разумеется, каждая отдельная мутация вызывается какой-то причиной. Однако в большинстве случаев эти причины остаются нам неизвестными. Мутации связаны с изменениями во внешней среде.
Это убедительно доказывается тем, что различными внешними факторами удается резко повысить число возникающих мутаций. Особенно эффективно действующими факторами экспериментального получения мутаций оказываются такие, которые влияют на нуклеиновые кислоты. Это вполне понятно, так как материальной основой генов служит ДНК.
Впервые в опыте резкое повышение числа возникающих наследственных изменений было получено действием лучей Рентгена. Под влиянием рентгенизации число получаемых мутаций удалось повысить в 150 раз и даже более. С тех пор экспериментальное получение мутаций было осуществлено на самых различных организмах — от бактерий и вирусов до млекопитающих и цветковых растений. Кроме лучей Рентгена и других форм ионизирующей радиации, мутации могут быть вызваны самыми различными химическими и физическими воздействиями: температурой, изменением газового режима, влажности и т. п. Любые изменения, затрагивающие процессы обмена веществ, оказывают свое влияние и на мутационный процесс. Результаты исследований по экспериментальному получению мутаций показали, что в основном дело сводится к увеличению их частоты. Экспериментально вызываемые наследственные уклонения совершаются в различных направлениях, так же как и естественный процесс мутационной изменчивости. Лишь в самое последнее время намечаются некоторые пути воздействия на направление мутаций. Эти новые возможности базируются на глубоком проникновении в механизм процесса синтеза нуклеиновых кислот.
Экспериментальное получение мутаций имеет и большое практическое значение, так как резко повышает наследственную изменчивость, давая, таким образом, материал для отбора.
Важная закономерность была установлена Н. И. Вавиловым. Она известна под именем закона гомологических рядов наследственной изменчивости. Сущность этого закона сводится к тому, что виды и роды, генетически близкие
(т. е. связанные друг с другом единством происхождения), характеризуются сходными рядами наследственной изменчивости.
Зная наследственные изменения у одного вида, можно предвидеть нахождение сходных изменений у родственных видов и родов. Несколько примеров гомологических рядов изменчивости в семействе злаков даны в прилагаемой таблице, в которой показано наличие или отсутствие различных признаков в пределах разных родов этого семейства.
У животных мы также встречаемся с проявлением этой закономерности. Например, у грызунов существуют гомологические ряды по окраске шерсти.
Закон гомологических рядов имеет важное практическое значение, он облегчает поиски наследственных уклонений.
Список литературы
1. Азимов А. Краткая история биологии. М.,1997.
2. Кемп П., Армс К. Введение в биологию. М.,2000.
3. Либберт Э. Общая биология. М.,1978 Льоцци М. История физики. М.,2001.
4. Найдыш В.М. Концепции современного естествознания. Учебное пособие. М.,1999.
5. Небел Б. Наука об окружающей среде. Как устроен мир. М.,1993.
|
