| Контрольная работа
по физиологии человека и животных
Вариант № 7
«Механизмы регуляции газообмена»
Вариант-7. Механизмы регуляции газообмена
1. Газовый состав крови человека и высших животных. Процессы, определяющие содержание газов во внутренней среде организма в условиях покоя и при напряженной работе.
2. Внешнее дыхание, его механизм и значение в системе гомеостаза газового состава крови.
3. Условия, способствующие газообмену в легких. Потенциальные резервы легочного газообмена.
4. Жизненная емкость легких, дыхательный, дополнительный, резервный остаточный воздух. Минутный объем дыхания (МОД). Возрастные особенности внешнего дыхания.
5. Зависимость между интенсивностью внешнего дыхания и газовым составом крови. Описание опытов, иллюстрирующих эту зависимость.
6. Представление о дыхательном центре. Роль в регуляции дыхания СО2
и О2
.
7. Значение в дыхании импульсов с рецепторов легких, дыхательных мышц, дыхательных путей и скелетной мускулатуры.
8. Особенности процесса саморегуляции дыхания.
9. Роль в регуляции дыхания коры больших полушарий. Примеры изменения дыхания под влиянием корковой регуляции.
10. Регуляция дыхания при мышечной работе, при пониженном атмосферном давлении.
11. Кривая диссоциации-оксигемоглобина. Роль гемоглобина в поддержании газового гомеостаза.
1.
Газовый состав крови человека и высших животных. Процессы, определяющие содержание газов во внутренней среде организма в условиях покоя и при напряженной работе.
Дыхание (respiratio)— совокупность процессов, обеспечивающих поступление из атмосферного воздуха в организм кислорода, использование его в биологическом окислении органических веществ и удаление из организма углекислого газа. В результате биологического окисления в клетках освобождается энергия, идущая на обеспечение жизнедеятельности организма. Нормальная жизнедеятельность клеток возможна лишь при условии удаления конечных продуктов метаболизма, к числу которых относится углекислый газ.
Различают внешнее Д., транспорт газов кровью (Газообмен) и тканевое. Внешнее Д.— это совокупность процессов газообмена, осуществляемых с помощью дыхательных мышц, бронхолегочного аппарата и системной регуляции, обеспечивающей вентиляцию легочных альвеол и диффузию газов через альвеолярно-капиллярные мембраны. Во внешнем Д. выделяют две стадии: конвекционный транспорт газов в альвеолы (вентиляция); диффузия из альвеол в кровь легочных капилляров кислорода и в обратном направлении углекислого газа.
Реклама

Газообмен— совокупность процессов обмена газов между организмом и окружающей средой; состоит в потреблении кислорода и выделении углекислого газа с незначительными количествами газообразных продуктов и паров воды. Интенсивность Г. пропорциональна интенсивности окислительно-восстановительных процессов, происходящих во всех органах и тканях, и находится под регулирующим влиянием нервной и эндокринной систем.
Газообмен обеспечивается функциями нескольких систем организма. Наибольшее значение имеют внешнее, или легочное, дыхание, обеспечивающее направленную диффузию газов через альвеолокапиллярные перегородки в легких и обмен газов между наружным воздухом и кровью; дыхательная функция крови, зависимая от способности плазмы растворять и способности гемоглобина обратимо связывать кислород и углекислый газ; транспортная функция сердечно-сосудистой системы (кровотока), обеспечивающая перенос газов крови от легких к тканям и обратно; функция ферментных систем, обеспечивающая обмен газов между кровью и клетками тканей, т.е. тканевое дыхание.
Дыхательная функция крови определяется количеством связанных с гемоглобином и растворенных в плазме О2
и СО2
, а также условиями, обеспечивающими диссоциацию молекул HbO2 и HbCO2 необходимую для Г. между тканями и легкими. Кроме О2
и СО2
в крови в небольших количествах растворены азот, аргон, гелий, окись углерода, метан. Азот содержится в крови в количестве, которое соответствует растворимости его в воде при температуре тела и при парциальном давлении его в атмосфере. Содержание азота в крови составляет 1,2 объемных процента, при тех же условиях вода может растворить 0,9 объемных процента. Азот в артериальной и в венозной крови содержится в равных количествах. Участия в дыхании азот не принимает.
В артериальной крови содержится 18-20 объемных процентов О2 и 50-52 объемных процентов СО2. В венозной крови – 10-12 объемных процентов О2 и 55-57 объемных процентов СО2. Артериальная кровь насыщена О2 на 96%, а венозная – на 66%.
Содержание газов в жидкости в физически растворенном виде зависит от его напряжения и от коэффициента растворимости (закон Генри — Дальтона), соответствующего объему газа (в мл), физически растворяющегося в 1 мл жидкости при напряжении газа, равном 1 атм, или 760 мм рт. ст. Для цельной крови при t° 37° коэффициент растворимости кислорода равен 0,024, углекислоты — 0,49, азота — 0,012. Чем выше напряжение газа, тем больше, при прочих равных условиях, его объем, растворяемый в жидкости, в т.ч. в крови. При парциальном давлении кислорода в альвеолярном воздухе, равном 95 мм рт. ст.(при давлении 14231 Па), в 100 мл артериальной крови растворено около 0,30 мл О2
в смешанной венозной крови при снижении напряжения кислорода до 40 мм рт. ст. в 100 мл крови на долю физически растворенного кислорода приходится около 0,11 мл. Количество растворенного СО2
в 100 мл артериальной и венозной крови соответственно составляет 2,6 и 2,9 мл, значительно больше, чем растворимость О2. Почти весь О2 в крови соединен с гемоглобином. Содержание СО2 в крови в 17-18 раз больше, чем это соответствует его растворимости при температуре тела и при парциальном давлении, при котором он находится в крови. Большая часть СО2 находится в крови не в растворенном состоянии, а в виде углекислых соединений.
Реклама

Человек поглощает в покое в среднем за 1 час на каждый килограмм массы тела 300 см3 О2 (в среднем 420-500 дм3 в сутки) и выделяет в 1 час на каждый килограмм массы тела 250 см3 СО2 (в среднем 380-450 дм3 в сутки). Водяного пара выделяется в сутки 450 см3.
Вдыхаемый атмосферный воздух относительно постоянен по своему составу. В нем содержится 20,96% О2; 0,04% СО2 и 79,0% N. Выдыхаемый воздух содержит 16,4% О2; 4,1% СО2 и 79,5% N. Приведенные цифры относятся к сухому воздуху при температуре 0º и давлении 101080 Па.
Напряжение СО2 в альвеолярном воздухе незначительно колеблется в зависимости от пола, возраста, мышечной работы и колебаний барометрического давления.
Несмотря на колебания частоты дыхания, содержание СО2 в альвеолярном воздухе остается относительно постоянным, так как учащение или урежение дыхания возмещается уменьшением или увеличением глубины дыхания.
Газообмен у детей связан с различиями в регуляции щелочно-кислородного равновесия. Например, у ребенка 5 лет в выдыхаемом воздухе в 3 раза меньше СО2, чем у взрослых.
2.
Внешнее дыхание, его механизм и значение в системе гомеостаза газового состава крови.
Акт дыхания состоит из трех процессов:
внешнее, или легочное, дыхание - обмен газов между организмом и окружающей средой; внутреннее, или тканевое, дыхание, протекающее в клетках; транспорт газов кровью, т.е. перенос кислорода кровью от легких к тканям и углекислого газа от тканей к легким.
Легочный тип дыхания является наиболее совершенным, он характерен для высокоорганизованных животных - птиц и млекопитающих, а также для человека. У более низкоорганизованных животных дыхание жаберное, трахейное и кишечное.
Органы дыхания подразделяются на воздухоносные пути и дыхательную часть. К воздухоносным путям относятся носовая полость, гортань, трахея и бронхи; к дыхательной части относится паренхима легких - легочные альвеолы, в которых происходит газообмен. Дыхательная система развивается как вырост вентральной стенки передней кишки; эта связь сохраняется в окончательной стадии развития - верхнее отверстие гортани открывается в глотку. Воздух проходит в гортань через полость носа или рта и глотку (их объединяют под названием «верхние дыхательные пути»). Для дыхательных путей характерно наличие хрящевого состава в их стенках (в результате чего стенки дыхательных путей не спадаются) и мерцательного эпителия на слизистой оболочке дыхательных путей. Ворсинки слизистого эпителия колеблются против движения воздуха и гонят наружу вместе со слизью инородные частицы, загрязняющие воздух.
Воздух в полость носа поступает через ноздри, носовая полость делится перегородкой на две половины, а сзади с помощью хоан сообщается с носоглоткой. Стенки носовой полости образованы костями и хрящами, выстланы слизистой оболочкой. Проходя через полость носа, воздух согревается, увлажняется и очищается. В полости носа находятся обонятельные луковицы, благодаря которым человек воспринимает запах. С носовой полостью связаны воздухоносные пазухи соседних костей - околоносовые пазухи (придаточные пазухи носа). Из носовой полости воздух попадает в носоглотку, затем в ротовую и гортанную части глотки, куда открывается гортань. Воздух сюда может поступать также через рот. Гортань располагается в области шеи на уровне 4-6 шейных позвонков, по бокам ее располагаются доли щитовидной железы, а сзади - глотка. Гортань образована хрящами. Надгортанник прикрывает вход в гортань во время глотания. Изнутри гортань покрыта слизистой оболочкой с мерцательным эпителием. На боковой стороне гортани справа и слева имеется углубление - желудочек гортани. Гортань служит для проведения воздуха и одновременно является органом звукообразования. В образовании звуков участвуют две голосовые связки: правая и левая, состоящие из эластических соединительных волокон. Связки натянуты между щитовидным и черпаловидными хрящами и ограничивают голосовую щель. При напряжении голосовых связок выдыхаемый воздух приводит их в колебание, в результате чего возникают звуки. Напряжение или расслабление голосовых связок, а также расширение или сужение голосовой щели зависят от сокращения мышц гортани (все мышцы гортани поперечнополосатые). Далее гортань переходит в трахею - трубку длиной около 12 см, состоящую из хрящевых полуколец. Задняя стенка трахеи мягкая (состоит из соединительнотканной перепонки), прилегает к пищеводу. Изнутри она также выстлана слизистой оболочкой, содержащей железы, которые выделяют слизь. Из области шеи трахея переходит в грудную полость и делится на два бронха (бифуркация трахеи). Бронхи входят в легкие и там делятся на бронхи меньшего диаметра. В грудной полости расположено два легких. Они имеют форму конуса: верхняя - суженная часть - верхушка, а нижняя - более широкая - основание. На стороне каждого легкого, обращенной к сердцу, располагаются углубления (ворота легкого), через которые проходят бронх, нерв легкого, кровеносные и лимфатические сосуды. Бронх в каждом легком ветвится. Бронхи, как и трахея, в стенках содержат хрящи. Самые мелкие разветвления бронхов называются бронхиолами, они не имеют хрящей и желез, но снабжены мышечными волокнами и способны сужаться (спазмы бронхиол). Правое легкое состоит из трех, а левое из двух долей. Каждый отдел легкого состоит из сегментов: в правом легком 11 сегментов, в левом - 10. Каждый сегмент в свою очередь состоит из множества легочных долек. Бронхиолы переходят в расширения - альвеолярные ходы, на стенках которых находятся выпячивания, называемые легочными пузырьками, или альвеолами (диаметр их 0,2-0,3 мм). Стенки альвеол состоят из однослойного эпителия и к ним примыкают капилляры. Через стенки альвеол и осуществляется газообмен: в кровь из альвеол поступает кислород, а обратно - углекислый газ. В легких взрослого человека насчитывается около 300-400 млн альвеол, их общая поверхность составляет около 100 м2. Легкие покрыты серозной оболочкой - плеврой. Плевра состоит из двух слоев - пристеночного и внутренностного. Около каждого легкого плевра образует плевральный мешок. Пристеночный листок прилегает к грудной клетке, а внутренностный - сросся с легким. Между двумя листками плевры имеется щелевидное пространство - полость плевры, в которой находится серозная жидкость, увлажняющая листки плевры, благодаря чему уменьшается трение плевры во время дыхания. В полости плевры воздуха нет и давление там отрицательное. Плевральные полости между собой не сообщаются.
Вдох осуществляется следующим образом: под влиянием нервных импульсов сокращаются мышцы, принимающие участие в дыхании (диафрагма, межреберные мышцы и др.). Диафрагма опускается (уплощается), за счет чего увеличивается вертикальный объем грудной полости. В этом акте принимают участие и другие мышцы, увеличивая горизонтальный объем легких. При вдохе легкие растягиваются, давление в них падает и становится ниже атмосферного. Таким образом создается разность давления между атмосферным и легочным воздухом, и наружный воздух устремляется в легкие. При выдохе мышцы расслабляются (диафрагма при этом поднимается), ребра опускаются, объем грудной клетки уменьшается, легкие сжимаются, давление в них повышается (выше атмосферного) и воздух по воздухоносным путям устремляется наружу. В спокойном состоянии взрослый человек дышит 16-20 раз в минуту. У детей дыхание более частое - до 60 вдохов в минуту. У нетренированных людей при физической нагрузке ритм дыхания учащается. Учащается дыхание при многих заболеваниях, но глубина его часто снижается. Во время сна дыхание урежается. Различают брюшной тип дыхания (преобладает у мужчин), когда объем грудной клетки увеличивается преимущественно в результате сокращения диафрагмы, и грудной (у женщин) - в результате сокращения других дыхательных мышц, когда увеличивается поперечный размер грудной клетки.
3.
Условия, способствующие газообмену в легких. Потенциальные резервы легочного газообмена.
Обмен газов в легких происходит в альвеолах. Альвеолы лежат в сети эластичных соединительных волокон, выполняющих опорную функцию и придающих легким эластичность и упругость. Важное физиологическое значение в газообмене легких имеет сурфактант – нерастворимая в воде тонкая пленка фосфолипида, покрывающая внутреннюю поверхность альвеол и стабилизирующая силы поверхностного натяжения. Сурфактант, противодействуя влиянию сил поверхностного натяжения при изменении диаметра альвеол, препятствует развитию слипания альвеол – ателектазу. Общая поверхность всех альвеол, через которую диффундируют газы, составляет у взрослого человека примерно 100-120 м2
, что более чем в 50 раз превышает поверхность кожи. Этим определяется резкое снижение роли кожи как органа дыхания у высших позвоночных, в том числе и у человека.
Стенки альвеол очень тонки, и газы легко диффундируют через них из полостей альвеол в кровеносные капилляры. Через легочные капилляры протекает около 5 л крови/мин. Суммарная толщина мембраны, разделяющей альвеолярный воздух и кровь, составляет 4 мкм и образована двумя слоями плоских клеток: эндотелия кровеносных капилляров и эпителиальных клеток, выстилающих поверхность альвеол. Скорость газообмена через мембрану достаточна велика, чтобы установилось почти полное равновесие между кровью, протекающей через легочные капилляры, и воздухом, содержащимся в альвеолах, хотя кровь пребывает в кровеносных капиллярах альвеол не более 2 с.

Система газообмена в легочной альвеоле и тканях:
1-полость легких, 2-альвеола, 3-кровеносные капилляры легких, 4-кровеносные капилляры тканей, 5-клетки тканей, 6-система наружного дыхания, 7-система внутреннего дыхания
Для физиологических процессов газообмена основное значение имеет парциальное давление, которое развивают газы в альвеолах (пропорционально их процентному содержанию).
Таблица. Состав воздуха, %
| Воздух
|
О2
|
СО2
|
N
|
| Вдыхаемый
|
20,9-21,00
|
0,02-0,03
|
79,02-78,97
|
| Выдыхаемый
|
16,40
|
4,10
|
79,50
|
| Альвеолярный
|
13,70
|
5,6
|
80,70
|
Атмосферный воздух значительно меняет свой газовый состав в результате газообмена с кровью в легочных альвеолах.
| Газы воздуха
|
Парциальное давление, мм рт. ст. (кПа)
|
| Водяные пары
|
47 (6,3)
|
| Азот
|
571 (76,1)
|
| Кислород
|
102 (13,6)
|
| Диоксид углерода
|
40 (5,3)
|
Диффузия О2
и СО2
происходит вследствие разности между парциальным давлением этих газов в альвеолярном воздухе и напряжением их в крови. Напряжение СО2
и О2
есть концентрация этих газов в газовой смеси, с которой растворимые газы находятся в равновесии при атмосферном давлении. Каждый газ растворяется в жидкости в зависимости от своего парциального давления. Напряжение О2
в артериальной крови равно 100 мм рт. ст., а СО2
– 40 мм рт. ст., в венозной крови соответствующие значения этих величин – 40 и 46 мм рт. ст. В альвеолярном воздухе парциальное давление О2
составляет 102, а СО2
– 40 мм рт. ст. Разность между напряжением газов в венозной крови и их давлением в альвеолярном воздухе равна для О2
примерно 62, а для СО2
– 6 мм рт. ст. Вследствие более низкого парциального давления СО2
в альвеолярном воздухе (по сравнению с напряжением его в венозной крови) он переходит путем диффузии из крови легочных капилляров альвеолярный воздух. В результате более низкого парциального давления О2
в альвеолярном воздухе, по сравнению сего напряжением в венозной крови, О2
поступает из альвеолярного воздуха в кровь капилляров легких. Потребность человека в кислороде составляет примерно 350 мл/мин (при физической работе эта величина возрастает до 5000 мл/мин). Условия газообмена в альвеолах легко обеспечивают такой уровень потребления О2
организмом: разности в парциальных давлениях в 1 мм рт. ст. уже достаточно, чтобы в кровь из альвеол перешло 250 мл О2
(реально наблюдаемая разность напряжений О2
в альвеолярном воздухе и крови составляет 62 мм рт. ст.).
Обмен О2
и СО2
в легких включает диффузию газа через жидкость: О2
диффундирует вначале из альвеолярного воздуха в плазму крови, а затем из плазмы в эритроциты и соединяется с гемоглобином, образуя оксигемоглобин. В то время как диффузия газов через газовую среду есть простая функция молекулярной массы, диффузия газа через жидкость определяется расстоянием и концентрационным градиентом. Эта зависимость описывается законом Фика: Ф=∆dv/dx, где Ф – скорость диффузии на единицу поверхности, а dv/dx – математическое выражение, характеризующее интенсивность диффузии с расстоянием v. Диффузионная константа ∆ варьирует при изменении температуры и давления. При давлении примерно в 100 кПа (1 атм) ∆ увеличивается на 3% на каждый 1°С.

Рис. Обмен газов через стенку альвеолы
4.
Жизненная емкость легких, дыхательный, дополнительный, резервный остаточный воздух. Минутный объем дыхания (МОД). Возрастные особенности внешнего дыхания.
Легкие окружены герметически замкнутой плевральной полостью, в которой содержится 1—5 мл серозной жидкости, распределенной по легочной поверхности тонким слоем (толщина этого слоя 5—10 мкм). Благодаря этому висцеральный и париетальный листки плевры способны скользить относительно друг друга, что позволяет легким следовать за изменениями формы грудной клетки, не деформируясь.
В спокойном состоянии взрослый человек вдыхает и выдыхает в среднем 500 см3
воздуха (от 300 до 600). Этот объем воздуха называется дыхательным. Не весь дыхательный объем поступает в легочные альвеолы. Часть поглощенного воздуха остается в ротовой полости, носоглотке, гортани, трахее и бронхах. Эта часть дыхательных путей, в которых поглощенный воздух не приходит в соприкосновение с кровью, называется мертвым или вредным пространством. Его объем у взрослого человека равен 140-160 см3
и несколько увеличивается при расширении бронхов вследствие расслабления их мускулатуры и , наоборот, уменьшается при сужении бронхов, сокращении их мускулатуры.
Объем легких определяется силой PL
, которая равна разности между давлением в альвеолах РА
и внутриплевральным (внутригрудным) давлением Ppl
RL
= РА
— Ppl
. В нормальных условиях легкие всегда растянуты. На их поверхности создается некоторое напряжение, обусловленное растяжением эластических элементов легочной ткани (так называемая эластическая тяга легких) и силами поверхностного натяжения в стенках альвеол. Эти силы обеспечивают отрицательное давление в плевральной полости (по отношению к давлению в альвеолах). Поэтому Ppl
, численно равно эластической тяге легких EL
. В конце выдоха внутриплевральное давление ниже атмосферного на 3—5 см вод. ст. (0,3—0,5 кПа), а на высоте вдоха — на 6—8 см вод. ст. (0,6—0,8 кПа).
При спокойном Д. в течение одного вдоха в легкие поступает 400—500 мл воздуха (дыхательный объем — ДО), а при максимально глубоком вдохе — еще примерно 1500 мл воздуха (резервный, или дополнительный, объем вдоха — РОвд.
). После спокойного выдоха человек может дополнительно выдохнуть примерно 1500 мл воздуха (резервный объем выдоха — РОвыд.
), такое же количество воздуха остается в легких после максимального выдоха (остаточный объем легких — ООЛ). Объем воздуха в легких в конце спокойного выдоха называется функциональной остаточной емкостью (ФОЕ), которая представляет собой сумму показателей РОвыд.
и ООЛ. Наибольшее количество воздуха, которое можно выдохнуть после максимального вдоха, — это жизненная емкость легких, которая является суммой величин показателей ДО, РОвд.
, РОвыд.
и в среднем равна 3500 мл. Общая емкость легких (ОЕЛ), т.е. количество воздуха, содержащееся на высоте максимального вдоха, равна сумме показателей жизненной емкости легких (ЖЕЛ) и ООЛ.
Все три объема – дыхательный, резервный объем вдоха и резервный объем выдоха – составляют жизненную емкость легких, сокращенно ЖЕЛ.

Она представляет собой наибольший объем воздуха в легких, который может быть набран при однократном вдохе.
У человека жизненная емкость легких равна 3-4 дм3
. Жизненная емкость увеличивается с возрастом в связи с ростом грудной клетки и легких. С 18 до 35 лет она максимальна, а после 35-40 лет уменьшается. У женщин жизненная емкость меньше, чем у мужчин. Жизненная емкость возрастает с увеличением длины тела (на каждые 5 см роста она возрастает на 400 см3
) и объема туловища (который в среднем в 7 раз больше жизненной емкости).
Измерение жизненной емкости легких называется спирометрией. Даже после сильнейшего выдоха в легких человека остается около 1 дм3
воздуха, который не может быть удален. Этот объем воздуха, остающийся в легких и после смерти, называется остаточным. С возрастом у людей остаточный объем возрастает. Общая емкость легких состоит из жизненной емкости и остаточного объема.
Из числа указанных показателей наибольшее практическое значение имеют ДО, ЖЕЛ и ФОЕ. Величина ЖЕЛ зависит от возраста, пола, размеров и положения тела, степени тренированности. У женщин ЖЕЛ в среднем на 25% меньше, чем у мужчин. Благодаря функциональной остаточной емкости поддерживается постоянство содержания кислорода и углекислого газа в альвеолярном воздухе независимо от дыхательных движений. Поскольку ФОЕ в несколько раз больше ДО, изменение состава альвеолярного воздуха при вдохе относительно невелико. В среднем ФОЕ у людей молодого возраста составляет 2,4 л, а в пожилом возрасте — 3,4 л. У женщин ФОЕ примерно на 25% меньше, чем у мужчин.
Минутный объем дыхания (МОД) — объем вдыхаемого или выдыхаемого воздуха за 1 мин — рассчитывается путем умножения величины дыхательного объема на частоту дыхания. В покое МОД равен 6—9 л. Максимальная вентиляция легких колеблется в диапазоне 80—90 л/мин, достигая у тренированных лиц 170 л/мин.
При условии нормальной функции легких только около 2
/3
каждого дыхательного объема достигают альвеол, снабжаемых смешанной венозной кровью, и т.о. принимают участие в газообмене. Этот объем составляет так называемую эффективную, или альвеолярную, вентиляцию. Остающаяся треть, не участвующая в газообмене, — мертвое, или вредное, пространство. Оно включает анатомическое мертвое пространство (объем газа, который вентилирует воздухоносные пути) и альвеолярное мертвое пространство (объем газа, который можно рассматривать как не принимающий участие в газообмене на альвеолярном уровне).
Объем анатомического мертвого пространства зависит от размеров легких и составляет примерно 2,22 мл на 1 кг массы тела. В клинической практике объем анатомического мертвого пространства у взрослого человека принято считать равным 150 мл. Альвеолярное мертвое пространство не имеет четких анатомических границ. У здорового человека в положении лежа на спине анатомическое и альвеолярное мертвое пространство почти равны, при этом альвеолярное мертвое пространство минимально мало. В вертикальном положении, когда в высоко расположенные альвеолы не поступает воздух, оно может возрастать на 60—80 мл. При тяжелых заболеваниях легких с выраженным несоответствием между легочной вентиляцией и кровоснабжением альвеол альвеолярное мертвое пространство может увеличиваться на 100—120 мл.
Часть минутного объема дыхания, достигающая альвеол, называется альвеолярной вентиляцией. Именно от этой величины зависит поддержание газового состава альвеолярного воздуха. Поскольку объем мертвого пространства постоянен, альвеолярная вентиляция тем больше, чем глубже дыхание.
Состав альвеолярного воздуха на относительно постоянном уровне поддерживают механизмы регуляции внешнего Д. Недостаточная вентиляция легких (гиповентиляция) ведет к увеличению в крови напряжения углекислоты (гиперкапния) и уменьшению напряжения кислорода (гипоксемия). Следствием этих изменений является увеличение вентиляции легких (гипервентиляция) за счет как увеличения глубины Д. (гиперпноэ), так и возрастания частоты дыхательных движений (тахипноэ).
Избыток вентиляции легких ведет к снижению напряжения углекислоты в крови (гипокапния) и повышению напряжения кислорода (гипероксия). В результате дыхательные движения ослабевают (брадипноэ) или временно прекращаются (апноэ). Особыми типами вентиляции легких являются диспноэ — одышка, сопровождающаяся неприятным субъективным ощущением недостаточности или затруднения Д., и ортопноэ — выраженная одышка, связанная с застоем крови в легочных капиллярах в результате левожелудочковой сердечной недостаточности.
На вентиляцию легких влияют многие неспецифические факторы. К ним относятся прежде всего колебания температуры окружающей среды. Сильные холодовые или тепловые воздействия на кожу могут приводить к возбуждению дыхательного центра. Кроме того, Д. зависит от изменений температуры тела: как ее повышение (при лихорадочном состоянии), так и незначительное понижение (умеренная гипотермия) сопровождаются увеличением вентиляции легких. Резкое охлаждение (глубокая гипотермия) приводит к угнетению дыхательного центра. К неспецифическим факторам, влияющим на Д., относится также боль.
Величина минутного объема дыхания зависит от напряжения углекислого газа в артериальной крови и цереброспинальной жидкости. Напряжение углекислого газа воспринимается периферическими и центральными хеморецепторами. Периферические рецепторы локализуются в параганглиях каротидных синусов, расположенных у места деления общей сонной артерии на внутреннюю и наружную (каротидные тельца), и параганглиях дуги аорты (аортальные тельца). Хеморецепторы, находящиеся в этих образованиях, возбуждаются при повышении напряжения углекислого газа, уменьшении рН и снижении напряжения кислорода в артериальной крови.
Центральные хеморецепторы, настроенные на содержание в крови СО2
и ионов Н+
, расположены преимущественно на вентральной поверхности продолговатого мозга. Импульсация от хеморецепторов обеспечивает быстрые реакции дыхательного центра. При небольшом понижении напряжения кислорода в крови наблюдается рефлекторное учащение ритма дыхания, а при незначительном повышении напряжения углекислоты в крови происходит рефлекторное углубление дыхательных движений.
Дыхательный центр получает афферентные импульсы не только от хеморецепторов, но и от барорецепторов сосудистых рефлексогенных зон, а также от механорецепторов легких, дыхательных путей и дыхательных мышц. Именно импульсы от механорецепторов легких, поступающие к дыхательному центру по блуждающим нервам, определяют рефлекторную зависимость частоты и глубины Д. от объема легких (рефлексы Брейера и Геринга). У животных после перерезки блуждающих нервов Д. не прекращается, но становится реже и глубже. Жизненно важная функция Д. возможна лишь при ритмическом чередовании вдоха и выдоха и регулируется сложным нервным механизмом.
Газовую смесь в альвеолах, участвующую в газообмене, называют альвеолярным воздухом, или альвеолярной смесью газов. Содержание в ней различных газов относительно постоянно, что обеспечивает заключительный этап внешнего Д. — переход из альвеол в кровь легочных капилляров кислорода и в обратном направлении углекислого газа. Этот процесс представляет собой диффузию, движущей силой его является градиент парциальных давлений кислорода и углекислого газа.
У новорожденных и недоношенных детей отмечается выраженная дыхательная аритмия вследствие незрелости дыхательного центра. В периоде новорожденности частота дыхания оставляет 40—60 в минуту, она увеличивается даже при незначительном возбуждении. Вследствие горизонтального положения ребер и слабого развития дыхательных мышц Д. у новорожденных поверхностное, осуществляется во многом за счет диафрагмы. Метеоризм, аэрофагия, гепатолиенальный синдром способны уменьшать альвеолярную вентиляцию.
Чем меньше возраст ребенка, тем уже дыхательные пути: диаметр долевых бронхов новорожденного — 1,5, в 1 год — 3, в 4 года — 3,5 мм у взрослых — 5—6 мм). Давление в плевральной полости у новорожденных также отличается некоторыми особенностями. Через несколько минут после первого вдоха это давление на высоте вдоха составляет — 10 см вод. ст. — 1 кПа). Однако в конце выдоха разница между внутриплевральным и атмосферным воздухом у новорожденного равно 0. По мере роста организма эластическая тяга легких в конце выдоха постепенно увеличивается. Склонность к обструкции, образованию ателектазов, а также повышенная васкуляризация слизистых оболочек способствуют генерализации воспалительных процессов. Функционально дети первых дней жизни адаптированы к ацидозу и гипоксии. Наличие фетального гемоглобина в крови у новорожденных улучшает связывание кислорода в условиях низкого рН. Отклонения от индивидуальных физиологических норм дети переносят даже несколько лучше, чем взрослые. Однако чем меньше возраст ребенка, тем сильнее воздействие на Д. наркотиков и различных токсических веществ.
При старении организма вследствие изменений костно-мышечного аппарата грудной клетки, воздухоносных путей, легочной паренхимы, сосудов малого круга кровообращения условия легочной вентиляции ухудшаются. Снижается максимальная вентиляция легких — объем воздуха, проходящий через легкие за определенный промежуток времени при Д. с наиболее возможными частотой и глубиной. В результате увеличения физиологического мертвого пространства уменьшается доля альвеолярной вентиляции в минутном объеме Д. Снижение эффективности вентиляции связано с нарушением равномерности распределения вдыхаемого воздуха из-за потери легочной тканью эластичности, наличия ателектатических участков, а также нарушения бронхиальной проходимости. Отмечается несовершенство адаптивных механизмов внешнего Д. Так, у лиц пожилого и старческого возраста МОД увеличивается преимущественно за счет частоты, а не глубины Д., максимум респираторных реакций смещается к концу нагрузки, нарушается четкое соотношение между интенсивностью работы и изменением дыхательной функции, удлиняется период восстановления сдвигов, увеличивается кислородный долг.
5. Зависимость между интенсивностью внешнего дыхания и газовым составом крови. Описание опытов, иллюстрирующих эту зависимость
.
Главным фактором, определяющим уровень дыхательных движений в организме, является концентрация СО2
в крови. Повышение его содержания увеличивает возбудимость структур дыхательного и пневмотаксического центров, в результате чего усиливается и учащается дыхание. Первый вдох у новорожденного также связан с увеличением концентрации СО2
в крови ребенка после отделения его от плаценты. Эта концентрация, достигнув порогового значения, активизирует нервные структуры дыхательного центра. Экспериментальными исследованиями показано, что основным фактором, стимулирующим деятельность структур дыхательного центра, является не уменьшение количества О2
, а увеличение СО2
в крови (состояние гиперкапнии). С понижением концентрации СО2
в крови связана задержка вдыхании, наступающая после нескольких глубоких дыхательных движений, так как при этом из крови вымывается большое количество СО2
и концентрация последнего в ней падает ниже порогового (т.е стимулирующего вдох) значения (наступает гипокапния). Стимулирующий эффект повышенного содержания СО2
в крови связан как с прямым действием его на клетки дыхательного центра, так и с опосредованным рефлекторным влиянием на дыхательный ритм через хеморецепторы кровеносных сосудов. Прямые доказательства участия СО2
в генерации нервных импульсов получены на гигантских нервных клетках ганглия моллюска Aplysia. Увеличение СО2
в среде приводило к деполяризации клеточной мембраны и возникновению группы потенциалов действия. (Физиология человека и животных /под ред. Когана А.Б.)
Другим важным фактором регуляции постоянства газового состава крови является рефлекторный механизм регуляции глубины и частоты дыхательных движений путем активации хеморецепторов каротидных синусов. У основания каждой из внутренних сонных артерий расположены каротидные синусы, в которых находятся хеморецепторные клетки, чувствительные к изменениям химического состава крови. Аналогичные хеморецепторные образования расположены в дуге аорты. Понижение напряжения О2 в крови, стимулируя хеморецепторы сосудистых стенок, вызывает рефлекторное учащение дыхания.
В острых и хронических экспериментах показано, что рефлекторное изменение (уселение) дыхательных движений через хеморецепторы каротидных синусов является основным механизмом регуляции дыхания в зависимости от содержания О2 в артериальной крови. О решающей роли каротидных и аортальных рефлексогенных зон в поддержании постоянства напряжения О2 в крови свидетельствует тот факт, что после денервации этих зон реакция дыхания на гипоксию исчезает, в то время как реакция дыхания на гиперкапнию сохраняется. Следовательно, физиологическое значение СО2 в организме определяется не только его участием в механизме рефлекторной саморегуляции дыхания, но и важной ролью в прямой активации структур дыхательного центра. Перерезка обоих блуждающих нервов на шее приводит к резкому замедлению ритма дыхательных движений, движение при этом становится более глубоким. Экспериментально показано, что по блуждающим нервам распространяются центростремительные нервные импульсы, оказывающие постоянное влияние на деятельность нейронов дыхательных центров. Центростремительный характер этой импульсации доказывается тем, что раздражение центрального отрезка перерезанного нерва почти всегда оказывает четкий эффект на дыхательные движения. В составе блуждающего нерва выявлены две группы волокон: одни оказывают влияние, аналогичное раздражению любого сенсорного нерва, другие – являются специфическими афферентными путями дыхательной системы. Перерезка волокон второй группы приводит к длительным изменениям характера дыхательных движений.
Поддержание постоянства газового состава крови по отношению к СО2 обеспечивается в основном прямой реактивностью дыхательного центра на высокое содержание СО2 в крови. При низком напряжении О2 в крови прямая реакция структур дыхательного центра имеет гораздо меньшее значение, чем активация хеморецепторов каротидных и аортальной зон. Хеморецептивные реакции каротидных и аортальной зон сосудистой стенки усиливают прямые гуморальные эффекты на дыхательный центр, так как рефлекторная реакция на гиперкапнию и гипоксемию (уменьшение О2 в крови) качественно протекает так же, как и прямая реакция нейронов дыхательного центра на гуморальные стимулы. Обычно при гиперкапнии наступает углубление дыхания в то время, как гипоксемия вызывает значительное увеличение частоты дыхательных движений.
6.
Представление о дыхательном центре. Роль в регуляции дыхания СО2
и О2
.
Понятие о дыхательном центре появилось в 18 в. Современные электрофизиологические методы исследования позволили выявить определенные группы так называемых дыхательных нейронов.
К дыхательным нейронам относят нервные клетки, импульсная активность которых меняется в соответствии с фазами дыхательного цикла. Различают инспираторные нейроны, которые разряжаются в фазу вдоха, экспираторные, активные во время выдоха, и целый ряд нейронных популяций, активность которых или занимает часть определенной фазы дыхательного цикла (ранние, поздние), или включается в моменты перехода инспираторной фазы в экспираторную либо экспираторной в инспираторную.
Центральный дыхательный механизм входит в состав ретикулярной формации ствола мозга. Подавляющая масса дыхательных нейронов сосредоточена в двух группах ядер: дорсальной и вентральной.
Активность центрального дыхательного механизма управляется стимулами, исходящими от хеморецепторов и механорецепторов дыхательной системы. Главная особенность работы этого механизма – линейное нарастание активности инспираторных нейронов на протяжении вдоха и резкий обрыв инспираторной активности, знаменующий окончание вдоха и переход к выходу. Центральный дыхательный механизм продолговатого мозга обладает автоматией, т.е. постоянной ритмической активностью. Важнейшую роль играет импульсация, поступающая от хеморецепторов.
Регуляция Д. осуществляется ЦНС. Рефлекторные сокращения дыхат. мускулатуры обеспечиваются двигательными нервами, ядра к-рых расположены в передних рогах серого вещества спинного мозга. Ритмичную смену вдоха и выдоха, координацию деятельности спинномозговых нервов обеспечивает дыхательный центр (ДЦ), расположенный в продолговатом мозге. В варолиевом мосту находится пневматаксиче-ский центр, к-рый совместно с ДЦ служит регулятором ритма Д. В регуляции ритма Д., его частоты и глубины большое значение имеют лёгочные рецепторы, импульсация от которых по блуждающим нервам поступает в ДЦ. Главным фактором, регулирующим Д., является концентрация СО2 в крови (повышение его содержания ведёт к усиленным сокращениям дыхательной мускулатуры и увеличению МОД) и сопровождается удалением избыточного СО2 из организма. Гомеостатический механизм регуляции содержания О2 и СО2 в крови связан с наличием в сонных артериях рецепторов, чувствительных к изменениям химич. состава крови и обеспечивающих быстрые реакции ДЦ на изменения напряжения О2 и СО2 в крови. Центральные хеморецепторы, расположенные на поверхности продолговатого мозга, реагируют на изменения СО2 в ликворе. Регуляция Д. направлена не только на автоматич. поддержание гомеостатич. констант парциального давления О2 и СО2, но и на предупреждение возможных отклонений. При нарушениях Д. и механизмов его регуляций возникают изменения газового состава крови.
7.
Значение в дыхании импульсов с рецепторов легких, дыхательных мышц, дыхательных путей и скелетной мускулатуры.
Регуляция дыхательного ритма осуществляется нервной системой и гуморальным путем. Центр дыхания располагается в продолговатом мозгу. В дыхательном центре различают два отдела: отдел вдоха и отдел выдоха, функции которых взаимосвязаны. При возбуждении отдела вдоха происходит торможение отдела выдоха и наоборот. Участвуют в регуляции дыхания специальные скопления нервных клеток в мосту и в промежуточном мозге. Кроме того, в спинном мозге находятся группы клеток, отростки которых идут в составе спинномозговых нервов к дыхательным мышцам. В дыхательном центре попеременно возбуждение сменяется торможением. При вдохе легкие расширяются, их стенки растягиваются, что раздражает окончания блуждающего нерва. Возбуждение передается к дыхательному центру и тормозит его деятельность. Мышцы перестают получать возбуждение от дыхательного центра и расслабляются: грудная клетка опускается, ее объем уменьшается и происходит выдох. При расслаблении центростремительные волокна блуждающего нерва перестают возбуждаться (нет тормозящего действия на дыхательный центр), и дыхательный центр, не получая тормозящих импульсов, вновь возбуждается - наступает очередной вдох. Таким образом, происходит как бы саморегуляция: вдох вызывает выдох, а выдох - вдох. Другой причиной изменения деятельности дыхательного центра является концентрация углекислоты в крови. Она является специфическим возбудителем дыхания: повышение концентрации углекислоты в крови (особенно в крови, омывающей дыхательный центр) приводит к возбуждению дыхательного центра - дыхание становится частым и глубоким. Глубокое и частое дыхание продолжается до тех пор, пока концентрация углекислоты в крови не снизится до нормального уровня. На понижение концентрации углекислоты в крови дыхательный центр отвечает понижением возбудимости вплоть до полного прекращения своей деятельности на некоторое время - до установления нормального уровня углекислоты. Ведущим физиологическим механизмом, влияющим на дыхательный центр, является рефлекторный, за которым следует гуморальный. Дыхание подчинено коре головного мозга, о чем свидетельствует факт произвольной задержки дыхания (хотя и на очень короткое время) или изменения частоты и глубины дыхания. Это подтверждается учащением дыхания человека при эмоциональных состояниях. Возбуждение дыхательного центра также может вызвать пониженное содержание кислорода в крови и некоторые лекарственные препараты, поступающие в кровь. С дыханием связаны и такие защитные акты, как кашель и чиханье. Они осуществляются рефлекторно, а их центры находятся в продолговатом мозге. Кашель возникает в ответ на раздражение слизистой оболочки гортани, глотки или бронхов (при попадании пыли, частиц пищи и др.). При кашле после глубокого вдоха воздух с силой выталкивается из дыхательных путей, приводя в движение голосовые связки (возникает характерный звук); вместе с выдыхаемым воздухом удаляются и раздражители. Акт чиханья возникает в ответ на раздражение слизистой оболочки носа; причины его те же, что и при кашле.
Остановка дыхания, которая наблюдается в результате понижения концентрации углекислоты в крови, называется апноэ. Нарушение ритма дыхания - одышка и учащенное дыхание - происходит вследствие повышения концентрации углекислоты в крови и называется диспноэ (гиперпноэ). Возникновением апноэ объясняется способность некоторых людей нырять и оставаться несколько минут под водой. Ныряльщик перед погружением в воду производит несколько частых и глубоких дыхательных движений, вследствие чего в крови понижается концентрация углекислоты. Ныряльщикам за жемчугом удается пробыть под водой до 5 минут, но только после длительных тренировок. А при задержке дыхания на 1-1,5 минуты наступает диспноэ - усиленное дыхание. При задержке дыхания концентрация углекислоты в крови повышается и как следствие - увеличивается возбудимость дыхательного центра до тех пор, пока содержание углекислоты в крови не понизится. С повышенной концентрацией углекислоты в крови связан и первый вдох новорожденного.
8. Особенности процесса саморегуляции дыхания
.
Координированные сокращения дыхательных мыщц обеспечиваются ритмической активностью нейронов дыхательного центра. Такие нейроны сгруппированы в целом ряде структур мозгового ствола. Неотъемлемым звеном аппарата регуляции дыхания являются также хеморецепторные и механорецепторные системы, обеспечивающие нормальную работу центрального дыхательного механизма в соответствии с потребностями организма в обмене газов.
Центральный дыхательный механизм мозгового ствола и его афферентные системы обеспечивают поддержание легочной вентиляции, адекватной уровню газообмена в тканях организма, т.е. выполняют гомеостатическую функцию. Дыхательные мышцы участвуют в локомоторных актах, поддержании позы, фонаторной функции. Поэтому нейрональные структуры спинного и продолговатого мозга, управляющие респираторной мускулатурой, находятся под контролем вышележащих центров супрапонтинного уровня – мозжечка, среднего и промежуточного мозга, больших полушарий
Важное значение имеет влияние этих центров в регуляции дыхания, связанной с двигательной активностью организма. Особую роль играет кортикальная регуляция дыхания у человека. Это связано прежде всего с участием дыхательного аппарата в речевой функции. Человек может управлять дыханием произвольно. Это позволяет ему менять частоту и глубину дыхания и даже задерживать его. Но длительность произвольного апноэ ограничена: как только парциальное давление накапливающегося в крови СО2 достигает определенного уровня, сильное возбуждение хеморецепторов (императивнй стимул) выводит дыхание из-под произвольного контроля и дыхательные движения возобновляются.
Центральный дыхательный механизм жестко ограничивает вмешательство в регуляцию дыхания со стороны супрапонтинных структур, охраняя жизненно важную гомеостатическую, газообменную функцию дыхательной системы.
9.
Роль в регуляции дыхания коры больших полушарий. Примеры изменения дыхания под влиянием корковой регуляции.
Дыхательный центр в продолговатом мозге находится под влиянием нервных импульсов из опоясывающей извилины, полосатых тел, зрительных бугров, подбугровой области, среднего мозга, мозжечка и премоторных областей больших полушарий головного мозга.
Раздражение лобных долей больших полушарий головного мозга вызывает изменения дыхательных движений (В.Я.Данилевский, 1874). У животных были выработаны условные рефлексы на дыхание. После удаления участков коры больших полушарий, осуществляющих высшую регуляцию дыхания, безусловно-рефлекторные изменения дыхания сохранялись, а затем восстанавливались.
Влияние больших полушарий головного мозга на дыхание доказывается тем, что дыхательные движения могут быть по желанию изменены: дыхание можно замедлить или задержать на некоторое время или, наоборот, участить или углубить. Психические влияния и внушения вызывают изменение дыхательных движений. У людей вентиляция легких увеличивается при действии сигналов, предшествующих мышечной деятельности.
Высшая, наиболее тонкая форма регуляции дыхания – обеспечение соответствия дыхательных движений условиям жизни – осуществляется при участии больших полушарий головного мозга.
Предполагается, что нейроны больших полушарий более возбудимы к повышению напряжения углекислоты в крови, чем дыхательный центр в продолговатом мозге.
10.
Регуляция дыхания при мышечной работе, при пониженном атмосферном давлении.
Регуляция дыхания при мышечной работе
При усиленной мышечной работе в крови, протекающей через мышцы, увеличивается давление СО2
, повышается концентрация водородных ионов и температура крови, что благоприятствует отдаче О2
в мышцах.
Эти изменения осуществляются рефлекторно и нервно-гуморально. В результате значительного усиления обмена веществ, поступления в кровь углекислоты, молочной и других кислот и накопления в крови водородных ионов и возбуждения дыхательного центра легочная вентиляция при мышечной работе увеличивается как за счет учащения дыхания, так и за счет углубления.
Парциальное давление СО2
в артериальной крови во время мышечной работы не изменяется или незначительно увеличивается (не соответственно величине физической нагрузки). При тяжелой мышечной работе оно меньше, чем при легкой работе. Возбудимость дыхательного центра к действию СО2
во время мышечной работы не увеличивается. Эти факты доказывают, что ведущая роль в увеличении дыхания при мышечной работе принадлежит не гуморальному, а рефлекторному механизму (рефлексам из химических рецепторов кровеносных сосудов, из механических рецепторов дыхательных мышц, диафрагмы и двигательного аппарата). Особенно возрастает число афферентных импульсов из рецепторов дыхательных мышц и диафрагмы при дыхании в противогазе, что приводит к рефлекторному усилению вентиляции легких. Ведущая роль в изменениях дыхания при мышечной работе принадлежит корригирующим влияниям больших полушарий головного мозга и условным рефлексам, главным образом при действии словесных раздражителей. Условные рефлексы на дыхательный аппарат тем больше выражены, чем ближе начало выполнения трудовых или физических упражнений (предрабочее и предстартовое состояние). Изменения дыхания во время речи обусловлены только нервными влияниями.
Рефлекторная регуляция дыхания в соответствии с интенсивностью мышечной работы связана с изменениями физико-химических свойств крови, влияющих на возбудимость дыхательного центра. Максимальный уровень вентиляции легких ограничен пределом возбудимости дыхательного центра. Повышение этого предела при чрезмерном увеличении притока к нему афферентных импульсов уменьшает вентиляцию легких. При произвольном усилении дыхания обмен веществ в дыхательных мышцах больше, чем при произвольном.
При интенсивной мышечной работе вследствие увеличения вентиляции легких возрастает парциальное давление кислорода в альвеолярном воздухе и резко снижается содержание кислорода в венозной крови в результате увеличения его потребления в мышцах. Поэтому артериально-венозная разность кислорода в крови достигает 160 см3
и больше.
Легочная вентиляция обычно увеличивается параллельно интенсивности физической работы. Чем напряженнее работа, тем больше воздуха проходит через легкие и тем больше газообмен. При гимнастических упражнениях вентиляция легких увеличивается в 2-7 раз, а при беге на средние дистанции – даже до 20 раз.
При интенсивной длительной мышечной работе, беге, езде на велосипеде, плавании нередко «через» некоторое время наступает очень сильная отдышка, затруднение дыхания и ощущение стеснения в груди, учащаются сердцебиения, повышается кровяное давление («мертвая точка»). Это состояние преодолевается волевым усилием, следовательно, импульсами с больших полушарий, и тогда исчезает ощущение стеснения в груди и удушье, а самочувствие улучшается. Дыхание становится ровным и спокойным, значительно повышается работоспособность («второе дыхание»).
При «мертвой точке» в крови накапливаются углекислота и молочная кислота и повышается концентрация водородных ионов. Это вызывает дальнейшее углубление дыхания, резко усиленную вентиляцию легких, которая сопровождается увеличением выделения СО2
в выдыхаемый воздух и падением концентрации водородных ионов; то же наступает и благодаря увеличенному выделению молочной кислоты при потоотделении во время «второго дыхания».
Усиленная вентиляция легких перед интенсивной мышечной работой предупреждает наступление «мертвой точки».
При подъеме тяжестей, лазании, гребле и других физических упражнениях происходит натуживание – закрытие голосовой щели после вдоха и задержка дыхания при напряжении выдыхательной мускулатуры, что приводит к повышению давления в легких и сужению легочных сосудов.
Регуляция дыхания при пониженном атмосферном давлении.
 Парциальное давление СО2
в альвеолярном воздухе остается относительно постоянным и при повышении и при понижении атмосферного давления в окружающей среде. Это имеет большое физиологическое значение, так как создает необходимое условие для нормальной возбудимости дыхательного центра и делает возможным дыхание на больших высотах. Постоянство парциального давления СО2
в альвеолярном воздухе обусловлено регулированием дыхания. Это было доказано в опытах, проведенных на людях в стальной камере, сконструированной для изучения влияния высоких и низких атмосферных давлений (барокамера). Парциальное давление СО2
в альвеолярном воздухе остается относительно постоянным и при повышении и при понижении атмосферного давления в окружающей среде. Это имеет большое физиологическое значение, так как создает необходимое условие для нормальной возбудимости дыхательного центра и делает возможным дыхание на больших высотах. Постоянство парциального давления СО2
в альвеолярном воздухе обусловлено регулированием дыхания. Это было доказано в опытах, проведенных на людях в стальной камере, сконструированной для изучения влияния высоких и низких атмосферных давлений (барокамера).
В отличие от СО2
парциальное давление О2
в альвеолярном воздухе повышается при увеличении атмосферного давления и понижается при уменьшении этого давления.
При подъеме на высоту падает атмосферное давление и уменьшается парциальное давление О2
в альвеолярном воздухе. Это вызывает нарушение снабжения организма О2
. Корличество кислорода в крови становится недостаточным (гипоксемия). Гипоксемия сопровождается значительной потерей углекислоты за счет выдыхания СО2
при увеличении легочной вентиляции.
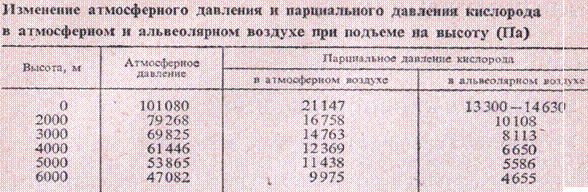
При быстром подъеме на 3-4 км у некоторых здоровых людей начинает развиваться гипоксемия, появляется высотная, или горная, болезнь, выражающаяся в ряде расстройств (головная боль, одышка, усталость, понижение зрения, слуха и др.). У большинства людей горная болезнь отмечается на высоте 4,5-5,5 км. Некоторые люди могут переносить без признаков горной болезни высоту 6 км.
При медленном подъеме на высоту или у живущих на высоте организм успевает приспособиться и не испытывает кислородного голодания (высотная акклиматизация). при этом усиливается легочная вентиляция, что увеличивает парциальное давление О2
в легких и насыщение им артериальной крови. Учащается пульс, увеличиваются минутный объем сердца и кровяное давление, количество эритроцитов и гемоглобина за счет депонированной крови, что повышает кислородную емкость крови на 20-25%. Увеличивается новообразование эритроцитов и содержание в них гемоглобина.
11.
Кривая диссоциации-оксигемоглобина. Роль гемоглобина в поддержании газового гомеостаза.
Большая часть О2
и СО2
переносится в форме связи их с гемоглобином в виде молекул HbO2 и HbCO2. Максимальное количество кислорода, связываемое кровью при полном насыщении гемоглобина кислородом, называется кислородной емкостью крови. В норме ее величина колеблется в пределах 16,0—24,0 об% и зависит от содержания в крови гемоглобина, 1 г которого может связать 1,34 мл кислорода (число Хюфнера). В клинике определяют степень насыщения артериальной крови кислородом, представляющую собой выраженное в % отношение содержания кислорода в крови к ее кислородной емкости.
Связывание кислорода гемоглобином является обратимым процессом, зависимым от напряжения кислорода в крови (при понижении напряжения кислорода оксигемоглобин отдает кислород), что отражается так называемой кислородно-диссоциационной кривой гемоглобина, а также от других факторов, в частности от рН крови. Как правило, все эти факторы смещают кривую диссоциации оксигемоглобина, увеличивая или уменьшая ее наклон, но не изменяя при этом ее S-образную форму.
Рис. Кислородно-диссоционная кривая гемоглобина.
  
Присоединение О2
к Hb, образование HbO2 и освобождение его (восстановление Hb) зависят от концентрации О2
и в меньшей степени – СО2
. Ни у одного животного напряжение О2
в крови не уравновешено непосредственно парциальным давлением этого газа в атмосфере (т.е. предельным значением О2
в газовой смеси окружающей среды). Дыхательные пигменты у большинства животных насыщаются при значительно более низком значении парциального давления. Диапозон значений парциального давления, в котором осуществляется либо присоединение, либо отщепление О2
является одной из наиболее важных характеристик дыхательного пигмента у различных животных. Эти значения парциального давления определяют условия, в которых функционирует данный дыхательный пигмент.
В области парциальных давлений О2
от 80 до 100 мм рт. ст. (т.е. колебаний, которые могут легко наступить в альвеолярном воздухе в связи с изменением внешней газовой среды организма) кривая диссоциации имеет пологий вид. Это означает, что насыщение гемоглобина кислородом при таких колебаниях парциального давления О2
меняется незначительно (от 93 до 96%). При обычных условиях дыхание Hb почти полностью насыщен О2
. Крутое падение кривой при парциальном давлении О2
, меньшем чем 60 мм рт. ст. (напряжение, характерное для тканей организма), свидетельствует о том, что при этих условиях кровь, протекающая через капилляры, интенсивно освобождает О2
, используемый функционирующими клетками и тканями организма, в которых напряжение О2
невелико. Кривая диссоциации HbO2 имеет S-образную форму. Нижний отрезок кривой определяет свойства Hb в зоне низкого парциального давления О2
, характерного для тканей организма; средняя часть кривой характеризует условия в венозной крови, верхний отрезок – в альвеолах легких.
Диссоциация HbO2 зависят от температуры: при одинаковом парциальном давлении О2
в окружающей среде HbO2, отдает больше О2
при температуре тела организма (37-38ºС), чем при более низкой температуре. Присоединение О2
к Hb в определенной мере регулируется концентрацией СО2
в крови.
СО2
реагируя с водой, образует угольную кислоту Н2СО3, поэтому при повышенном напряжении СО2
кислотность крови увеличивается. Увеличение кислотности крови приводит к уменьшению способности Hb связывать О2
. Такой механизм регуляции количества HbO2, обеспечивает эффективную систему переноса О2
в организме. В капиллярах функционирующих тканей концентрация СО2 достаточно велика, поэтому совместное действие низкого напряжения О2
и высокого напряжения СО2 ведет к освобождению гемоглобином большого количества кислорода. В капиллярах легких (или жабр) напряжение СО2 ниже, и под действием низкого напряжения СО2 и высокого напряжения О2
гемоглобин присоединяет к себе большее количество О2
.
Особенности транспортной функции переносчиков О2
, определяемые различиями в экологических условиях существования различных животных, проявляются в соответствующих изменениях кривой диссоциации HbO2. Кривые диссоциации у амфибий, обитающих главным образом на суше, сдвинуты вправо по сравнению с соответствующими кривыми для видов животных, обитающих в воде. Чем доступнее кислород для животного, тем выше давление, при котором происходит насыщение им гемоглобина крови. Так, у сухопутных черепах Hb имеет низкое сродство к О2
; напряжение полунасыщения Р50
(напряжение О2
, которое обеспечивает 50%-насыщение им гемоглобина) равно 28 мм рт.ст., в то время как у водных черепах сродство Hb к
О2
выше – Р50
равно 12 мм рт. ст.
Литература:
1. Воробьева Е.А., Губарь А.В., Сафьянникова Е.Б. Анатомия и физиология. М., 1975.
2. Гальперин С.И. Физиология человека и животных. М., 1977.
3. Общий курс физиологии человека и животных. В 2 кн./Под ред. А.Д.Ноздрачева. – М., 1991.
4. Физиология человека и животных. В 2 ч./Под ред. А.Б.Когана. – М., 1984.
|
