Наследственность и влияние на человека факторов среды: введение
Особенности наследственности при индивидуальном ответе на воздействие химических факторов внешней среды - это обычное для человеческих популяций явление. Генетические изменения
глутадион S-
трансферазы ,
CYP1A2 ,
N-ацетилтрансферазы и
параоксоназы являются примерами взаимосвязи изменений метаболизма с восприимчивостью индивидуума к канцерогенам и другим токсическим веществам окружающей среды.
Большую роль играют также наследуемые
изменения рецепторов , например рецепторов, связанных с устойчивостью к ретиноевой кислоте при острой лейкемии промиелоцитов, с парадоксальным ответом на антиандрогены при раке простаты и с пигментозом сетчатки.
Также большую роль играют наследуемые разновидности
белков, взаимодействующих с лекарствами , например вызывающие индуцированную антибиотиками глухоту, лечение гликокортикоидами альдостеронизма и гипертонию, QT-синдром и заболевание легких, вызываемое бериллием. Эти примеры показывают, как знание молекулярных механизмов ответа может повлиять как на профилактические меры для восприимчивых индивидуумов, так и на лечение генетических нарушений, возникающих при воздействии химических веществ окружающей среды.
Канцерогены и другие вредные вещества из окружающей среды обычно влияют лишь на часть популяции, подвергающейся их воздействию. Даже
курение , одна из основных причин сердечнососудистых заболеваний и рака, приводит к возникновению рака легких лишь у 5-10% курильщиков (
Taylor, 1990 ). На вопрос о том, чем же отличаются люди с меньшей или большей восприимчивостью, ответ такой же, как и в случае многих других биологических явлений - это обусловлено специфическими взаимодействиями наследственности и окружающей среды. Большинство воспринимает наследственность, как основной фактор, определяющий физические и умственные признаки организма, забывая о ее влиянии на различную реакцию индивидуумов на воздействие окружающей среды. А это воздействие может вызывать передающиеся по наследству изменения генетического материала.
Изучение влияния наследственности на восприимчивость человека к химическим веществам и есть сфера исследований
фармакогенетики .
Реклама

Практикующие фармакогенетики исследуют эту проблему на всех уровнях - от гена до популяции. Обобщая, можно сказать, что различия ответа на воздействие различных веществ можно характеризовать на двух уровнях: генетическом и биохимическом. Характерные особенности того и другого изучаются с целью определения взаимодействия экологии и наследственности. С этой целью подходы и методы фармакологии, генетики, биохимии, популяционной и молекулярной генетики и эпидемиологии используются при анализе фармакологических, токсикологических и эпидемиологических наблюдений за индивидуумами, близнецами, семьями и популяциями.
Многие генетические нарушения в ответ на внешние воздействия являются гетерогенными (т.е. у разных людей участвуют разные гены), хотя большинство считаются моногенными (т.е. в определенной семье дефектен только один чувствительный локус). Подобно другим наследственным особенностям, эти дефекты связаны с новыми мутациями, и так же как и генетические и демографические свойства различных этногеографических популяций могут варьировать, число и этиология факмакогенетических характеристик может также различаться. Они могут быть связаны с опустошающими болезнями и смертью, что делает их исследование клинически необходимым. Главная цель фармакогенетиков, это использование знаний о влиянии наследственности на чувствительность к воздействию экзогенных химических веществ в целях профилактики возможных нарушений. Так, раннее выявление раковых заболеваний, связанных с экологическими условиями, если и не может повлиять на их исход, то выявление вызвавших их причин дает возможность профилактики восприимчивых индивидуумов.
Аномальная реакция на внешние факторы характерна как для
полигенных , так и для
моногенных болезней ; мутации в одном гене часто вызывают сильные и нередко опасные для жизни
аллергические реакции на лекарственные средства (
табл. 65.6 ). Реакции на
изониазид , некоторые
бета-адреноблокаторы и
трициклические антидепрессанты существенно зависят от наследственных факторов. Кроме того, реакция на лекарственные средства изменена при
недостаточности Г-6-ФД ,
острой перемежающейся порфирии ,
гемохроматозе и
предрасположенности к злокачественной гипертермии .
Неправильная интерпретация побочного действия лекарственного средства может привести к катастрофическим последствиям для больного, поэтому любые необычные или аллергические реакции следует считать наследственно обусловленными, пока не доказано обратное. К счастью, при необычной реакции выбор лечения прост: больному и его родственникам, входящим в группу риска, не следует принимать опасный препарат.
Реклама

Развитию заболевания при наследственной предрасположенности способствуют не только лекарственные средства, но и другие факторы. При
недостаточности альфа1-антитрипсина курение особенно опасно для гомозигот (возможно, и для гетерозигот), предрасположенных к
эмфиземе . Больные с
пигментной ксеродермой чувствительны к
ультрафиолетовому излучению . Исключение
молока из рациона в раннем возрасте предотвращает опасные для жизни осложнения при
галактоземии .
Режим питания важен при большинстве форм
гиперлипопротеидемий .
К сожалению, современное общество подвержено влиянию множества новых внешних факторов. Например, широкое использование
аспартама в качестве
заменителя сахара представляет особый риск для больных
фенилкетонурией .
Мутагены
Мутагены — физические и химические факторы воздействие которых на живые организмы вызывает изменения наследственных свойств (генотипа).
Мутагены разделяются на: физические (рентгеновские и гамма-лучи.
радионуклиды, протоны, нейтроны и пр.), физико-химические (волокна, асбест), химические (пестициды, минеральные удобрения, тяжелые металлы и др.).
биологические (некоторые вирусы, бактерии). За всю историю своего развития человечество накопило (главным образом за счет естественного мутационного процесса) так называемый генетический груз, проявляющийся в наследственных, генетически обусловленных заболеваниях. Здоровье нынешних будущих поколений людей в значительной степени зависит от того, какой генетический груз получен в наследство от предыдущих, какое количество мутаций накоплено человечеством. На данный момент известно около 2 тысяч генетических дефектов, затрагивающих только часть общего числа локусов в геноме, а так как считается, что за поколение естественно возникает немногим более одной генной мутации (в геноме), частота их в среднем мала (на ло-кус за поколение) и не может угрожать существованию популяций, При этом примерно четверть общего объема мутаций обусловлена энергией естественного фона радиации.
Вместе с тем генные мутации, обусловливающие небольшие биохимические аномалии в организме, возможно, более часты. Проблема заключается в том, что ускорение частоты мутаций ведет к увеличению числа особей с врожденными дефектами и вредными отклонениями, передающимися по наследству, причем мутации в неполовых (соматических) клетках, как правило, могут вызывать рост злокачественных новообразований (спонтанный рак).
Расчеты показывают (Н.
Дубинин, 1958), что удвоение частоты мутаций настолько увеличивает объем генетического груза, что это может стать опасным для существования популяций. Существует выход из такого кризисного состояния — это путь эволюционных изменений, однако приспособление к мутагенам в процессе отбора требует от популяции огромного числа генетических жертв и времени.
В особенности видам, представленным сравнительно малым числом особей, с медленной сменяемостью поколений, труднее было бы приспособиться к высокому мутагенному фону среды, Больше шансов на выход из генетического кризиса, обусловленного ростом мутагенных загрязнений (повышением темпа мутаций), имеют биологические виды с высокой численностью особей, с быстрой сменяемостью поколений, например микроорганизмы, Хорошо известно явление резистентности их к широко распространенным антибиотикам, сульфаниламидным препаратам, так же как и появление устойчивых к пестицидам рас бактерий, грибов, насекомых. Главная опасность загрязнения окружающей среды мутагенами, как полагают генетики, заключается в том, что вновь возникающие мутации, не «переработанные» эволюционно, отрицательно повлияют на жизнеспособность любых организмов.
И если поражение зародышевых клеток может привести к росту числа носителей мутантных генов и хромосом, то при повреждении генов соматических клеток возможно возрастание числа раковых заболеваний.
Более того, существует глубокая связь различных на первый взгляд биологических эффектов. Например, мутагены окружающей среды влияют на величины рекомбинаций наследственных молекул, являющихся также источником наследственных изменений.
Возможно и влияние на функционирование генов, что может быть причиной, например, тератологических отклонений (уродств), наконец, вероятны поражения ферментных систем, что изменяет различные физиологические особенности организма, вплоть до деятельности нервной системы, а следовательно, сказывается и на психике.
Генетическая адаптация популяций человека к возрастающему загрязнению биосферы мутагенными факторами принципиально невозможна. В отличие от грубых хромосомных повреждений наследственного материала точковые генные мутации, обладающие способностью накапливаться в поколениях, представляют основную трудность для обнаружения в популяциях. Выявление их важно именно потому, что такие мутации будут в значительной мере ответственны за проявления генетического груза в ближайших поколениях. Определенные перспективы прямой регистрации генных точковых мутаций создает возможность слежения за изменениями в строении редких и мономорфных белков (Н.
Дубинин, Ю.
Алтухов, 1975).
Как показано Ю.
Алтуховым, спонтанные мутации приводят к изменению таких белков, и метод улавливает скрытые (в гетерозиготе) вновь возникшие мутации у особей.
А это необходимо как для выявления новых мутаций, вызванных загрязнением окружающей среды, так и для оценки изменений темпов мутирования, объема генетического груза, а тем самым и генетических последствий. Прежде всего необходимо оценить мутагенность различных загрязнений на высокочувствительных биологических тест-системах, в том числе и тех, которые могут поступить в биосферу, и если риск для человека доказан, то принимать меры для борьбы с ними. Формируется задача скрининга — просеивания загрязнений с целью выявления мутагенов и выработки специального законодательства для регулирования их поступления в окружающую среду.
И таким образом, контроль генетических последствий загрязнения в комплексе содержит в себе две задачи: испытание на мутагенность факторов среды различной природы и мониторинг популяций. На данный момент в мире уже имеется большое число квалифицированных лабораторий, в которых проводятся достаточно точные испытания.
Только за последнее десятилетие предложено свыше трех десятков тест-систем, часть которых предназначена для выявления точковых мутаций.
Задача состоит в разработке комплексных тест-систем, которые могли бы давать ответ на вопрос, в каких условиях потенциально мутагенные факторы могут стать действующими — в зависимости от каких путей попадания в организм и особенностей внутриклеточного обмена веществ, активирующего или, наоборот, подавляющего мутагенный эффект.
Комплексные наборы биологических тест- систем для массового скрининга предназначены для выявления всех типов мутационных повреждений хромосом и .генов и должны быть чувствительны к малым дозам мутагенов.
Ведь последствия суммарного и длительного воздействия низких доз мутагенов создают наибольший вклад в увеличение генетического груза: достаточные для индукции точковых мутаций, способных накапливаться в поколениях, они к тому же наиболее распространены в окружающей среде.
Генные мутации
1. Изменчивость, ее причины и методы изучения. Классификация форм изменчивости. Фенотипическая изменчивость и ее компоненты. Наследуемость признаков
2. Мутационная изменчивость. Основные положения мутационной теории. Общие свойства мутаций.
3. Генные мутации. Последствия мутаций. Методы выявления генных мутаций.
4. Общие закономерности мутационного процесса. Механизмы возникновения генных мутаций. Инсерционный мутагенез
1. Изменчивость, ее причины и методы изучения. Классификация форм изменчивости. Фенотипическая изменчивость и ее компоненты. Наследуемость признаков
Самовоспроизведение с изменением – это одно из основных свойств жизни. Термин «изменчивость» служит для обозначения различных понятий; как и большинство других терминов, он полисемантичен
(многозначен). Юрий Александрович Филипченко различал два основных подхода к определению изменчивости.
1. Изменчивость как состояние. В этом значении термин «изменчивость» служит для обозначения отличий биологических объектов друг от друга в данный момент времени. Всегда существуют различия между частями одного организма, между разными организмами в популяции, между разными внутрипопуляционными группировками, между популяциями.
2. Изменчивость как процесс. В этом значении термин «изменчивость» служит для обозначения изменения биологического объекта во времени. В этом случае изменчивость отражает развитие особи, отличие потомков от родителей.
Любая наблюдаемая изменчивость является фенотипической. В свою очередь, фенотипическая, или общая изменчивость включает три компонента:
ü Наследственная
(генетическая
, или генотипическая изменчивость
) – в значительной мере обусловлена влиянием генетических факторов. Например, в сходных условиях выращивается несколько сортов одного вида растений. Тогда различия между результатами эксперимента (например, урожайность) обусловлены генетическими особенностями каждого сорта. В основе генетической изменчивости лежит мутационная и комбинативная изменчивость.
ü Ненаследственная
(модификационная
) изменчивость
– в значительной мере обусловлена действием негенетических (экзогенных
) факторов. Например, один сорт растений выращивается в разных условиях. Тогда различия между результатами эксперимента (например, урожайность) обусловлены влиянием условий выращивания растений.
ü Неконтролируемая
(остаточная изменчивость
) – обусловлена неконтролируемыми (по крайней мере, в данном эксперименте) факторами.
Для разных признаков влияние генотипа и условий среды на общую фенотипическую изменчивость неодинаково. Например, окраска шерсти, жирномолочность у крупного рогатого скота, масса яиц у кур зависят, в основном, от особенностей породы (т.е. от генотипа) – эти признаки обладают высокой наследуемостью. Другие признаки: качество шерсти, общая удойность у КРС, яйценоскость у кур – зависят, в основном, от условий выращивания и содержания – эти признаки обладают низкой наследуемостью.
2. Мутационная изменчивость. Основные положения мутационной теории. Общие свойства мутаций
Термин «мутация» (от лат. mutatio
– изменение) долгое время использовался в биологии для обозначения любых скачкообразных изменений. Например, немецкий палеонтолог В.Вааген называл мутацией переход от одних ископаемых форм к другим. Мутацией называли также появление редких признаков, в частности, меланистических форм среди бабочек.
Современные представления о мутациях сложились к началу XX столетия. Например, российский ботаник Сергей Иванович Коржинский в 1899 г. разработал эволюционную теорию гетерогенезиса, основанную на представлениях о ведущей эволюционной роли дискретных (прерывистых) изменений.
Однако наиболее известной стала мутационная теория голландского ботаника Хьюго (Гуго) Де Фриза (1901 г.), который ввел современное, генетическое понятие мутации для обозначения редких вариантов признаков в потомстве родителей, которые не имели этого признака.
Де Фриз разработал мутационную теорию на основе наблюдений за широко распространенным сорным растением – ослинником двулетним, или энотерой (Oenothera
biennis
). У этого растения существует несколько форм: крупноцветковые и мелкоцветковые, карликовые и гигантские. Де Фриз собирал семена с растения определенной формы, высевал их и получал в потомстве 1…2% растений другой формы. В дальнейшем было установлено, что появление редких вариантов признака у энотеры не является мутацией; данный эффект обусловлен особенностями организацией хромосомного аппарата этого растения. Кроме того, редкие варианты признаков могут быть обусловлены редкими сочетаниями аллелей (например, белая окраска оперения у волнистых попугайчиков определяется редким сочетанием aabb
).
Основные положения мутационной теории Де Фриза остаются справедливыми и по сей день (разумеется, с некоторыми современными уточнениями):
Положения мутационной теории
Де Фриза
|
Современные уточнения
|
| 1 |
Мутации возникают внезапно, без всяких переходов. |
существует особый тип мутаций, накапливающихся в течение ряда поколений (прогрессирующая амплификация в интронах). |
| 2 |
Успех в выявлении мутаций зависит от числа проанализированных особей. |
без изменений |
| 3 |
Мутантные формы вполне устойчивы. |
при условии 100%-ной пенетрантности и 100%-ной экспрессивности |
| 4 |
Мутации характеризуются дискретностью (прерывистостью); это качественные изменения, которые не образуют непрерывных рядов, не группируются вокруг среднего типа (моды). |
существуют ликовые мутации, в результате которых происходит незначительное изменение характеристик конечного продукта |
| 5 |
Одни и те же мутации могут возникать повторно. |
это касается генных мутаций; хромосомные аберрации уникальны и неповторимы |
| 6 |
Мутации возникают в разных направлениях, они могут быть вредными и полезными. |
сами по себе мутации не носят адаптивный характер; только в ходе эволюции, в ходе отбора оценивается «полезность», «нейтральность» или «вредность» мутаций в определенных условиях; при этом «вредность» и «полезность» мутаций зависит от генотипической среды |
В настоящее время принято следующее определение мутаций:
Мутации – это качественные изменения генетического материала, приводящие к изменению тех или иных признаков организма
.
Организм, во всех клетках которого обнаруживается мутация, называется мутантом
. Это происходит в том случае, если данный организм развивается из мутантной клетки (гаметы, зиготы, споры). В ряде случаев мутация обнаруживается не во всех соматических клетках организма; такой организм называют генетической мозаикой
. Это происходит, если мутации появляются в ходе онтогенеза – индивидуального развития. И, наконец, мутации могут происходить только в генеративных клетках (в гаметах, спорах и в клетках зародышевого пути – клетках-предшественницах спор и гамет). В последнем случае организм не является мутантом, но часть его потомков будет мутантами.
Различают «новые» мутации (возникающие denovo) и «старые» мутации. Старые мутации – это мутации, появившиеся в популяции задолго до начала их изучения; обычно о старых мутациях едет речь в генетике популяций и в эволюционной теории. Новые мутации – это мутации, появляющиеся в потомстве немутантных организмов (♀ АА
× ♂ АА
→ Аа
); обычно именно о таких мутациях идет речь в генетике мутагенеза.
Мутация – это случайное явление, т.е. невозможно предсказать: где, когда и какое изменение произойдет. Можно только оценить вероятность мутации в популяциях, зная фактические частоты определенных мутаций. Например, вероятность появления у кишечной палочки устойчивости к тетрациклину равна 10–10
(одна десятимиллиардная), поскольку лишь одна из 10 миллиардов клеток обнаруживает устойчивость к этому антибиотику (зато все потомство этой бактерии будет устойчивым к тетрациклину).
Установлено, что мутабильность гена (т.е. частота появления определенной мутации) зависит от природы гена: существуют гены, склонные к мутированию, и относительно стабильные гены.
Вероятность события – это математическая абстракция, математическое ожидание того, или иного события. Вероятность случайного события лежит в пределах от 0 до 1. Математическое ожидание определяется вне опыта (априорно), на основании дедуктивных рассуждений. Например, при подбрасывании монеты вероятность выпадения «орла» равна вероятности выпадения «решки» и равна 50% или 0,5: Р
О
=Р
Р
=0,5.
Однако в биологии вероятность многих событий не может быть найдена вне опыта, например, вероятность рождения ребенка с синдромом Дауна. Тогда понятие математической вероятности подменяется понятием статистической вероятности
. Статистическая вероятность определяется опытным путем (апостериорно). Численно статистическая
, или апостериорная вероятность
события равна относительной частоте
этого события. Например, на 700 новорожденных приходится один ребенок с болезнью Дауна. Тогда статистическая вероятность рождения ребенка с этим заболеванием равна 1/700 ≈ 0,0014.
Относительная частота колеблется около некоторого постоянного числа, которое и является математическим ожиданием события. Чем больше проведено наблюдений, тем больше апостериорная вероятность приближается к математическому ожиданию данного события.
Множественные аллели и генокопии
В одном и том же гене могут возникать разные мутации: тогда возникают серии множественных аллелей
. Например, у мушки дрозофилы ген white
(«уайт» – белый), определяющий окраску глаз представлен последовательно доминирующими аллелями: w
+
(темно-красные глаза) > w
ch
(вишневые) > wa
(абрикосовые) > w
bf
(тускло-желтые) > w
(белые) и т.д. У кроликов ген, определяющий степень выраженности альбинизма, представлен последовательно доминирующими аллелями: C
(нормальная, неальбинистическая окраска) > c
ch
(шиншилловая) > c
h
(горностаевая) > с
(полный альбинизм). У мышей ген, определяющий общую окраску тела, также представлен последовательно доминирующими аллелями: AY
(желтая) > AL
(агути со светлым брюхом) > A
(агути, норма) > at
(черная с подпалинами) > a
(черная). Исходное, нормальное состояние аллеля традиционно называется дикими типом (часто обозначается символом +). Диким типом называют также нормальный генотип и нормальный фенотип. Сочетание двух мутантных аллелей называется компаундом
(например, w
ch
//
w
bf
или at
//
a
).
В то же время, в неаллельных, т.е. в разных генах (или в разных участках одного и того же гена) могут возникать мутации со сходным фенотипическим эффектом. Такие мутации называются генокопиями
. Генокопии необходимо учитывать в здравоохранении (сходные наследственные заболевания – например, гемоглобинопатии – могут быть обусловлены разными мутациями), в селекционном процессе (например, мутациями в разных генах может быть обусловлена устойчивость к фитопатогенным грибам у разных сортов растений).
Общие свойства мутаций
В настоящее время считается, что многие мутации не оказывают существенного влияния на жизнеспособность особей; такие мутации называются нейтральными
. Нейтральность мутаций часто обусловлена тем, что большинство мутантных аллелей рецессивно по отношению к исходному аллелю. Однако существуют мутации, приводящие к гибели организма (летальные) или заметно снижающие его жизнеспособность (полулетальные). В определенных условиях мутации могут повышать жизнеспособность организмов (как в примере с серповидноклеточной анемией).
По способности передаваться при половом размножении различают соматические
и генеративные мутации
. Соматические мутации не затрагивают половые клетки и не передаются потомкам. В результате соматических мутаций возникают генетические мозаики
. Генеративные мутации происходят в половых клетках и могут передаваться потомкам. При участии мутантных половых клеток образуются полностью мутантные организмы.
Мутации возникают как в аутосомах, так и в половых хромосомах; соответственно различают аутосомные мутации и мутации, сцепленные с полом. Кроме того, по возможности проявления в фенотипе различают доминантные, полудоминантные и рецессивные мутации (заметим, что подавляющее большинство мутаций является рецессивными).
Мутантный аллель может возвращаться в исходное состояние. Тогда первоначальная мутация называется прямой (например, переход А → а
), а другая – обратной мутацией, или реверсией (например, обратный переход а → А
).
Классификации мутаций
Мутации классифицируют на основании различных критериев. Например, по уровню фенотипического проявления различают следующие мутации: биохимические
(изменяется структура белков); физиолого-биохимические
(изменяется обмен веществ); онтогенетические
(изменяется характер онтогенеза); физиолого-репродуктивные
(изменяются плодовитость, границы репродуктивного периода); анатомо-морфологические
(изменяется внутреннее и внешнее строение организмов); этологические
(поведенческие).
По уровню организации генетического материала, затронутого изменением, все мутации делят на генные
, хромосомные
и геномные
.
3. Генные мутации. Последствия мутаций. Методы выявления генных мутаций
Генные мутации выражаются в изменении структуры отдельных участков ДНК. По своим последствиям генные мутации делятся на две группы: мутации без сдвига рамки считывания
и мутации со сдвигом рамки считывания
.
Мутации без сдвига рамки считывания
происходят в результате замены нуклеотидных пар, при этом общая длина ДНК не изменяется. В результате возможна замена аминокислот, однако из-за вырожденности генетического кода возможно и сохранение структуры белка.
Пример 1
. Замена аминокислотного остатка в составе полипептида
(миссенс–мутации
). В состав молекулы гемоглобина человека входят две a–цепи (a–цепь закодирована в 16-ой хромосоме) и две b–цепи (b–цепь закодирована в 11-ой хромосоме). В состав b–цепи входит 146 аминокислотных остатков, при этом в нормальной b–цепи шестым аминокислотным остатком является глутаминовая кислота. С участием нормальной b–цепи образуется нормальный гемоглобин – HbA. В нетранскрибируемой нити участка ДНК, кодирующего b–цепь, глутаминовая кислота закодирована триплетом ГАА. Если же в результате мутации в ДНК произойдет замена триплета ГАА на триплет ГТА, то на месте глутаминовой кислоты в молекуле гемоглобина в соответствии с генетическим кодом появится валин. В итоге вместо гемоглобина HbA появится новый гемоглобин – HbS. Такая замена всего лишь одного нуклеотида и одной аминокислоты приводит к развитию тяжелого заболевания – серповидноклеточной анемии
.
На клеточном уровне эта болезнь проявляется в том, что эритроциты приобретают форму серпа и теряют способность к нормальному транспорту кислорода. Гомозиготы HbS/HbS умирают в раннем детстве. Зато гетерозиготы HbA/HbS характеризуются слабо измененными эритроцитами. При этом изменение формы эритроцитов значительно повышает устойчивость гетерозигот к малярии. Поэтому в тех регионах Земли, где свирепствует малярия (например, в Африке), отбор действовал в пользу гетерозигот. Таким образом, серповидноклеточная анемия – это пример относительности «полезности» и «вредности» мутаций.
Пример 2
. Мутация без замены аминокислотного остатка в составе полипептида
(сеймсенс–мутации
). Если в нетранскрибируемой нити участка ДНК кодирующего b–цепь гемоглобина, произойдет замена триплета ГАА на триплет ГАГ, то из-за избыточности генетического кода замены глутаминовой кислоты не произойдет. В итоге структура b–цепи гемоглобина не изменится, и в эритроцитах будет обнаруживаться только нормальный гемоглобин HbA. Таким образом, вовсе не любая генная мутация проявляется в фенотипе.
Особую группу образуют ликовые мутации
, в результате которых происходит незначительное изменение характеристик конечного продукта. Это связано с заменой аминокислотных остатков в пассивной части белка: такие замены не оказывают существенного влияния на структуру и функции белка.
Мутации со сдвигом рамки считывания
(фреймшифты
) происходят в результате вставки или потери нуклеотидных пар, при этом общая длина ДНК изменяется. В результате происходит полное изменение структуры белка.
Однако если после вставки пары нуклеотидов происходит потеря пары нуклеотидов (или наоборот), то аминокислотный состав белков может восстановиться. Тогда две мутации хотя бы частично компенсируют друг друга. Это явление называется внутригенной супрессией.
| Структура ДНК и белка до мутаций
|
| ДНК |
А Т Г
|
Г Г Ц
|
А Т Ц
|
Г Г Ц
|
| Т А Ц
|
Ц Ц Г
|
Т А Г
|
Ц Ц Г
|
| ß |
ß |
ß |
ß |
| мРНК |
А У Г
|
Г Г Ц
|
А У Ц
|
Г Г Ц
|
| белок |
метионин |
глицин |
изолейцин |
глицин |
| Мутации и их последствия
|
| вставка пары А – Т
|
потеря пары Ц – Г
|
| ДНК |
А Т Г
|
А Г Г
|
Ц А Т
|
Г Г Ц
|
| Т А Ц
|
Т Ц Ц
|
Г Т А
|
Ц Ц Г
|
| ß |
ß |
ß |
ß |
| мРНК |
А У Г
|
А Г Г
|
Ц А У
|
Г Г Ц
|
| белок |
метионин |
аргинин |
гистидин |
глицин |
Мутации со сдвигом рамки считывания составляют ~ 80% от всех генных мутаций. Вставки иначе называются инсерциями
, а потери – эксцизиями
. Процесс образования вставок называется инсерционным мутагенезом
. Инсерционный мутагенез необходимо учитывать в генной инженерии.
Нонсенс–мутации
. Особую группу генных мутаций составляют нонсенс–мутации
с появлением стоп–кодонов. Нонсенс–мутации могут возникать как вследствие замен нуклеотидных пар, так и с потерями или вставками. С появление стоп–кодонов синтез полипептида вообще обрывается. В результате могут возникать нуль–аллели
, которым не соответствует ни один белок.
Кроме того, мутация в одном гене может подавлять мутации, происходящие в других (неаллельных) генах. Это явление называется межгенной супрессией
.
Дополнение 1
. Существуют особые мутации, влияющие на экспрессию генов у эукариот
1. Мутации, изменяющие степень компактизации ДНК. В гигантских политенных хромосомах и в хромосомах типа ламповых щеток описаны мутации, инактивирующие ген, расположенный в каком-либо одном участке ДНК, т.е. блокирующие декомпактизацию хроматина. Скрещивание гетерозигот по таким регуляторным мутациям в F
2
дает расщепление 3:1, указывая на то, что они затрагивают единичные менделирующие факторы.
2. Гомеозисные мутации. Изменяют порядок экспрессии генов. Фенотипический эффект гомеозисных мутаций заключается в превращении одних органов в другие. Например, у мушки дрозофилы мутация группы bithorax
, контролирующих развитие грудных и брюшных сегментов у дрозофилы, может приводить к появлению крылоподобных образований вместо галтеров; мутации группы antennapedia
выражаются в том, что у насекомых на месте антенн вырастают ножки; мутации ophthalmoptera
– развитие крыла из имагинального диска глаза; мутации proboscipedia
– развитие ноги или части антенны (в зависимости от температуры) вместо хоботка; у мутантов tumorous
head
ткани головы замещаются другими типами тканей, включая структуры, характерные для гениталий.
Дополнение 2
. Некоторые мутации обладают плейотропным действием, т.е. приводят к изменению сразу нескольких признаков.
Пример 1.
Ароматические аминокислоты – триптофан, фенилаланин, тирозин – образуются из хоризмовой кислоты. Если некоторая мутация заблокирует хотя бы один этап синтеза хоризмовой кислоты, то клетка (организм) утрачивает способность к синтезу сразу трех аминокислот.
Пример 2
. Один и тот же фермент (трансаминаза) контролирует синтез валина (из α–кетоизовалериановой кислоты) и изолейцина (из α–кето–β–метилвалериановой кислоты). Если некоторая мутация нарушит функции этого фермента, то клетка (организм) утрачивает способность к синтезу сразу двух аминокислот.
Методы выявления генных мутаций
Сложность выявления генных мутаций связана, во-первых, с рецессивностью большинства мутаций (вероятность их фенотипического проявления ничтожно мала), а во вторых с летальностью многих из них (мутанты не выживают).
Все множество методов выявления генных мутаций можно разделить на две группы: методы генетического анализа и биохимические методы.
1. Методы генетического анализа
основаны на скрещивании возможных носителей мутации с тестерными линиями (линиями-анализаторами). Самый простой метод – это скрещивание носителей предполагаемой мутации с соответствующей рецессивно-гомозиготной линией, т.е. обычное анализирующее скрещивание.
Однако этот метод не позволяет выявить неизвестные мутации, а также летальные мутации. Поэтому создаются специальные тестерные линии для учета летальных мутаций.
Например, у мушки дрозофилы синтезирована тестерная линия М
–5 (Мёллер–5), которая характеризуется особой структурой X
–хромосом у самок. В этих хромосомах имеются аллели с определенным фенотипическим проявлением (доминантный аллель B
– полосковидные глаза; рецессивный аллель wa
– абрикосовые глаза; кроме того, имеется еще один аллель – sc
, контролирующий отсутствие щетинок, но он в анализе обычно не учитывается). В хромосомах М
–5 изменен порядок генов: имеется одна большая инверсия и одна малая, расположенная внутри большой (инверсии будут рассмотрены ниже); такое строение хромосом исключает появление кроссоверных особей при скрещивании мушек М
–5 с другими линиями.
Для выявления мутаций используются самцы дикого типа – с нормальными X
–хромосомами (аллели В
+
и w
+
– нормальные красные глаза, sc
+
– нормальные щетинки; нормальный порядок генов). Эти самцы подвергаются обработке мутагенами (факторами, повышающими частоту мутаций). В результате в их половых клетках часть X
–хромосом мутирует, т.е. в них возникают мутации. Обработанные самцы скрещиваются с самками М
–5. В первом поколении (F1) все самки имеют полосковидные темно-красные глаза, а самцы – абрикосовые полосковидные глаза. Кроме того, часть самок получает от отцов по нормальный X
–хромосоме, а часть – по мутантной X
–хромосоме. Все самцы получают от матерей М
–5 только немутантные хромосомы с аллелями В
и wa
. В F1 рецессивные мутации у самок, даже если они есть, не дают летального эффекта, поскольку они находятся в гетерозиготном состоянии: мутантная X
–хромосома дикого типа от отца сочетается с немутантной М
–5–хромосомой от матери.
Затем гибриды первого поколения скрещиваются между собой, и потомство каждой самки выращивается отдельно. Часть самок несет немутантную X
–хромосому дикого типа, и в их потомстве обнаруживаются немутантные самцы дикого типа. Однако некоторая часть самок несет мутантную X
–хромосому дикого типа с летальной мутацией; соответственно их сыновья, получившие такие хромосомы, не выживают, и самцы дикого типа в потомстве самок–носительниц не обнаруживаются.
Ниже приведены схемы скрещивания, иллюстрирующие принцип использования метода Мёллер–5 (символом l
обозначены летальные мутации).
| Р
:
|
♀ wa
B // wa
B
|
×
|
♂ w+
B+
// Y
|
– обработка самцов |
| абрикосовые полосковидные |
красные нормальные |
– окраска и форма глаз |
| GP
:
|
wa
B
|
w+
B+
|
– немутантная X
–хромосома |
| w+
B+
l
|
– мутантная X
–хромосома |
| Y
|
– Y
–хромосома |
| F1
:
|
♀ w+
B+
// wa
B
|
♀ w+
B+
l // wa
B
|
♂ wa
B // Y
|
| красные полосковидные глаза; без летальных мутаций |
красные полосковидные глаза; носители летальных мутаций |
абрикосовые полосковидные глаза |
1 вариант скрещивания – без летальных мутаций
| F1
:
|
♀ w+
B+
// wa
B
|
×
|
♂ wa
B // Y
|
| красные полосковидные глаза |
абрикосовые полосковидные глаза |
| G
1
:
|
w+
B+
|
wa
B
|
| wa
B
|
Y
|
| F2
:
|
♀ w+
B+
// wa
B
|
♀ wa
B // wa
B
|
♂ w+
B+
// Y
|
♂ wa
B // Y
|
| красные полосковидные глаза |
абрикосовые полосковидные глаза |
красные нормальные глаза |
абрикосовые полосковидные глаза |
2 вариант скрещивания – при наличии летальных мутаций
| F1
:
|
♀ w+
B+
l // wa
B
|
×
|
♂ wa
B // Y
|
| красные полосковидные глаза |
абрикосовые полосковидные глаза |
| G1
:
|
w+
B+
l
|
wa
B
|
| wa
B
|
Y
|
| F2
:
|
♀ w+
B+
l // wa
B
|
♀ wa
B // wa
B
|
♂ w+
B+
l // Y
|
♂ wa
B // Y
|
| красные полосковидные глаза |
абрикосовые полосковидные глаза |
самцы не обнаруживаются (летали) |
абрикосовые полосковидные глаза |
В настоящее время, кроме тестерной линии М
–5 используются и другие тестерные лини мушек дрозофил и других модельных объектов. Например, существуют тест-системы, позволяющие выявлять мутации X
-хромосомах самцов в первом же поколении, а также мутации в аутосомах. Применение этих линий позволяет изучать закономерности мутационного процесса, однако классический генетический анализ далеко не всегда можно использовать для выявления мутаций в популяциях человека и многих других организмов.
2. Биохимические методы
выявления мутаций
исключительно разнообразны и основаны на применении различных методик.
а).
Методики, основанные на выявлении определенных биохимических продуктов мутантных генов. Легче всего выявлять мутации по изменению активности ферментов или по утрате какого-либо биохимического признака. Например, у микроорганизмов на селективных питательных средах выявляются ауксотрофные формы, не способные синтезировать определенные вещества (по сравнению с нормальными, прототрофными формами).
б)
. Методики, основанные на непосредственном выявлении измененных нуклеиновых кислот и белков с помощью гель-электрофореза в сочетании с другими методиками (блот-гибридизации, авторадиографии).
4. Общие закономерности мутационного процесса. Механизмы возникновения генных мутаций
Причины возникновения мутаций
По причинам возникновения различают спонтанные и индуцированные мутации.
Спонтанные (самопроизвольные) мутации
возникают без видимых причин. Эти мутации иногда рассматривают как ошибки трех Р
: процессов репликации, репарации и рекомбинации ДНК
. Это означает, что процесс возникновения новых мутаций находится под генетическим контролем организма. Например, известны мутации, которые повышают или понижают частоту других мутаций; следовательно, существуют гены-мутаторы и гены-антимутаторы.
Индуцированные мутации
возникают под действием мутагенов
. Мутагены – это разнообразные факторы, которые повышают частоту мутаций. Впервые индуцированные мутации были получены отечественными генетиками Г.А. Надсоном и Г.С. Филипповым в 1925 г. при облучении дрожжей излучением радия.
Различают несколько классов мутагенов:
ü Физические мутагены: ионизирующие излучения, тепловое излучение, ультрафиолетовое излучение.
ü Химические мутагены: аналоги азотистых оснований (например, 5-бромурацил), альдегиды, нитриты, метилирующие агенты, гидроксиламин, ионы тяжелых металлов, некоторые лекарственные препараты и средства защиты растений.
ü Биологические мутагены: чистая ДНК, вирусы, антивирусные вакцины.
ü Аутомутагены – промежуточные продукты обмена веществ. Например, этиловый спирт сам по себе мутагеном не является. Однако в организме человека он окисляется до ацетальдегида, а это вещество уже является мутагеном.
Общие закономерности мутагенеза
Мутации возникают не мгновенно. Вначале под воздействием мутагенов возникает предмутационное состояние клетки. Различные репарационные системы стремятся устранить это состояние, и тогда мутация не реализуется. Основу репарационных систем составляют различные ферменты, закодированные в генотипе клетки (организма). Таким образом, мутагенез находится под генетическим контролем клетки; это – не физико-химический, а биологический процесс.
Например, ферментные системы репарации вырезают поврежденный участок ДНК, если повреждена только одна нить (эту операцию выполняют ферменты эндонуклеазы), затем вновь достраивается участок ДНК, комплементарный по отношению к сохранившейся нити (эту операцию выполняют ДНК-полимеразы), затем восстановленный участок сшивается с концами нити, оставшимися после вырезания поврежденного участка (эту операцию выполняют лигазы).
Существуют и более тонкие механизмы репарации. Например, при утрате азотистого основания в нуклеотиде происходит его прямое встраивание (это касается аденина и гуанина); метильная группа может просто отщепляться; однонитевые разрывы сшиваются. В некоторых случаях действуют более сложные, малоизученные системы репарации, например, при повреждении обеих нитей ДНК.
Однако при большом числе повреждений ДНК они могут стать необратимыми. Это связано с тем, что: во-первых, репарационные системы могут просто не успевать исправлять повреждения, а во-вторых, могут повреждаться сами ферменты систем репарации, необратимые повреждения ДНК приводят к появлению мутаций – стойких изменений наследственной информации.
Физические мутагены
К физическим мутагенам относятся: ионизирующее излучение (альфа–, бета–, гамма–, нейтронное и рентгеновское излучение), коротковолновое ультрафиолетовое излучение, СВЧ–излучение, действие экстремальных температур.
Действие ионизирующего излучения основано на ионизации компонентов цитоплазмы и ядерного матрикса. При ионизации возникают высокоактивные химические вещества (например, свободные радикалы), которые различным образом действуют на клеточные структуры. Рассмотрим наиболее изученные механизмы мутагенного воздействия ионизирующего излучения.
1.
Непосредственное воздействие
частиц с высокой энергией на ДНК, которое приводит к ее разрывам: одиночным (под воздействием гамма-квантов, рентгеновских лучей) или множественных (под воздействием альфа-частиц, нейтронного излучения). Это универсальный механизм возникновения хромосомных перестроек на всех стадиях клеточного цикла, но он действует очень грубо – обычно клетки теряют способность к нормальному делению и погибают. К разрывам ДНК приводит и ультрафиолетовое облучение.
2
. Опосредованное воздействие
ионизирующих факторов связано с нарушением структуры ферментов, контролирующих репликацию, репарацию и рекомбинацию ДНК. Этот механизм наиболее эффективно действует на синтетической стадии интерфазы. При больших дозах мутагенов клетки погибают. (Поскольку раковые клетки делятся непрерывно, то облучение является универсальным средством подавления развития метастазов при онкологических заболеваниях – непрерывно делящиеся раковые клетки более уязвимы, чем медленно пролиферирующие или непролиферирующие нормальные клетки.)
Опосредованное воздействие ионизирующих факторов индуцирует самые разнообразные генные и хромосомные мутации. При опосредованном действии ионизирующих факторов их мутагенный эффект может быть снижен с помощью специальных веществ – радиопротекторов. К радиопротекторам относятся различные антиоксиданты, взаимодействующие с продуктами ионизации. В то же время, мутагенный эффект может быть усилен, например, высокая температура повышает мутагенный эффект радиации.
3. Особенности мутагенного действия ультрафиолетовых лучей
. ДНК интенсивно поглощает жесткий ультрафиолет с длиной волны ≈ 254 нм. Основным продуктом является образование нуклеотидных димеров: два нуклеотида, расположенных рядом в одной цепи ДНК, «замыкаются» сами на себя, образуя пары «тимин–тимин» или «тимин–цитозин». При репликации ДНК напротив такой пары в достраивающейся цепи могут стать два любых нуклеотида, т.е. принцип комплементарности не выполняется. Ультрафиолетовый свет – это сравнительно мягкий мутаген, поэтому его широко используют в селекции растений, облучая проростки.
4. Особенности мутагенного действия экстремальных температур
. Собственный мутагенный эффект экстремальных температур не доказан. Однако очень низкие или очень высокие температуры нарушают деление клетки (возникают геномные мутации). Экстремальные температуры усиливают действие других мутагенов, поскольку снижают ферментативную активность репарационных систем.
Химические мутагены
К химическим мутагенам относятся самые разнообразные вещества. Рассмотрим мутагенное действие некоторых из них.
Алкилирующие агенты
. Вызывают алкилирование ДНК (например, метилирование, этилирование и т.д.). В результате при репликации ДНК нарушается принцип комплементарности, и происходит замена нуклеотидных пар: ГЦ → АТ; ГЦ → ЦГ; ГЦ → ТА
Некоторые из алкилирующих агентов в природе не встречаются, их не распознают ферменты защитных систем. Такие вещества называются супермутагенами (например, N-метил-N-нитрозомочевина). Супермутагены применяются в селекции растений для получения индуцированных мутаций; их используют также как стимуляторы роста (в сверхмалых концентрациях).
Гидроксиламин
. Избирательно аминирует цитозин, что также нарушает принцип комплементарности при репликации ДНК. В результате происходит замена ГЦ → АТ.
Нитриты
. Осуществляют окислительное дезаминирование гуанина, аденина, цитозина. Также нарушается принцип комплементарности при репликации ДНК. В результате происходит замена АТ → ГЦ.
Аналоги оснований
. Это вещества, сходные с «обычными» азотистыми основаниями. Однако они способны образовывать комплементарные пары с разными «нормальными» основаниями. Например, при репликации ДНК напротив гуанина вместо цитозина достраивается 5-бромурацил (аналог тимина). В дальнейшем напротив 5-бромурацила достраивается аденин, а напротив аденина – обычный тимин. Этот же процесс может идти и в противоположную сторону. В результате происходят замены: ГЦ → АТ или АТ → ГЦ.
Существует множество иных химических факторов, обладающих мутагенным, канцерогенным и тератогенным действием. Например, ионы тяжелых металлов, связываясь с ферментами репликации, репарации и рекомбинации, снижают их ферментативную активность. Таким образом, не являясь собственно мутагенами, ионы тяжелых металлов способствуют появлению мутаций. Кроме того, нужно учесть, что воздействие совершенно разных мутагенов может приводить к сходным результатам.
Опасность загрязнения окружающей среды мутагенами
Во второй половине ХХ века над биосферой нависла угроза загрязнения мутагенами. Любая популяция способна выдержать лишь определенный груз мутаций. Увеличение частоты мутаций может привести к снижению устойчивости популяций из-за нарушения генетического гомеостаза. Необходимо дальнейшее усиление биомониторинга – контроля за состоянием окружающей среды с помощью биологических систем (от англ. monitor
– предостерегающий). В качестве профилактических мер следует использовать развитие «безотходных» технологий, ограничение производства веществ с мутагенным действием, усиление всех видов контроля за состоянием потенциально опасных предприятий: АЭС, химические и микробиологические производства, научно-промышленные установки биотехнологического характера.
Установлено, что мутагены при определенных условиях оказывают канцерогенное и тератогенное действие
. Канцерогены – это факторы, провоцирующие развитие онкологических заболеваний; тератогены – это факторы, провоцирующие развитие различных аномалий, уродств. Тератогенный эффект дают многие лекарственные препараты. Например, в 1960-е гг. на Западе широко использовалось снотворное талидомид
, применение которого привело к рождению большого числа детей с недоразвитыми конечностями.
Наряду с тератами
– уродствами – часто встречаются морфозы
– изменения, которые не ведут к утрате органом его функций. Отличить мутагенное действие от тератогенного сравнительно легко: тераты (уродства) являются ненаследственными модификациями, они предсказуемы (направлены) и не сохраняются в последующих поколениях. Например, серая окраска тела у дрозофилы – это нормальный признак. В то же время известна мутация yellow
– желтое тело (эту мутацию легко получить искусственно, обрабатывая родительских особей различными мутагенами; при этом разные мутагены могут давать одинаковый фенотипический эффект). Если же личинкам дрозофилы добавлять в корм азотнокислое серебро, то все эти личинки разовьются в мух с желтым телом. Но, если от этих желтых мух получить потомство и выращивать его на обычной питательной среде, то все потомки вновь станут серыми. Таким образом, в данном случае «пожелтение» тела мух – это не мутация, а модификация, или фенокопия (модификация, по фенотипу копирующая мутацию); азотнокислое серебро в данном случае является не мутагеном, а тератогеном.
Мутации – источник генетической изменчивости популяций
Частота возникновения мутаций.
Эволюция была бы невозможной, если бы генетические программы воспроизводились абсолютно точно. Как вы знаете, копирование генетических программ – репликация ДНК – происходит с высочайшей, но не абсолютной точностью. Изредка возникают ошибки – мутации. Частота мутаций не одинакова для разных генов, для разных организмов. Она возрастает, иногда очень резко, в ответ на воздействие внешних факторов, таких как ионизирующая радиация, некоторые химические соединения, вирусы и при изменениях внутреннего состояния организма (старение, стресс и т.п.).
Средняя частота мутаций у бактерий оценивается как 10-9
на ген на клетку за поколение. У человека и других многоклеточных она выше и составляет 10-5
на ген на гамету за поколение. Иными словами только в одной из 100 тысяч гамет ген оказывается измененным. Казалось бы, это ничтожно малая величина. Следует помнить, однако, что генов в каждой гамете очень много. По современным оценкам геном человека содержит около 30 тысяч генов. Следовательно, в каждом поколении около трети человеческих гамет несут новые мутации по какому-нибудь гену.
Таким образом, несмотря на чрезвычайную редкость каждой отдельной мутации, в каждом поколении появляется огромное количество носителей мутантных генов. Благодаря мутационному процессу генотипы всех организмов, населяющих Землю, постоянно меняются; появляются все новые и новые варианты генов (аллели), создается огромное генетическое разнообразие, которое служит материалом для эволюции.
Вредные, полезные и нейтральные мутации.
Мутации различаются по своим фенотипическим эффектам. Большинство мутаций, по-видимому, вовсе никак не сказываются на фенотипе. Их называет нейтральными мутациями. Большой класс нейтральных мутаций обусловлен заменами нуклеотидов, которые не меняют смысла кодонов.
Такие замены называют синонимическими.
Например, аминокислота аланин кодируются триплетами ГЦУ, ГЦЦ, ГЦА и ГЦГ. Если в результате мутации ГЦУ превращается в ГЦЦ, то белок, синтезированный по измененной программе, остается тем же самым. Если мутация изменяет смысл кодона (несинонимическая мутация) и одна аминокислота заменяется другой, это может привести к изменению свойств белка.
Большинство несинонимических мутаций оказывается вредными. Они нарушают скоординированное в ходе предшествующей эволюции взаимодействие генетических программ в развивающимся организме, и приводят либо к его гибели, либо к тем или иным отклонениям в развитии. Только очень малая доля вновь возникающих мутаций может оказаться полезной.
Следует помнить, однако, насколько условна эта классификация. Полезность, вредность, или нейтральность мутации зависит от условий, в которых живет организм. Мутация нейтральная или даже вредная для данного организма и данных условиях, может оказаться полезной для другого организма и в других условиях, и наоборот. Жуки и комары не могли знать заранее, что люди изобретут ДДТ и другие инсектициды и подготовить мутации защиты. Тем не менее, эти мутации возникали – они обнаруживаются даже в тех популяциях насекомых, которые с инсектицидами не встречались. В то время, когда насекомые не сталкивались с инсектицидами, эти мутации были нейтральными. Но как только люди стали применять инсектициды – эти мутации стали не просто полезными, они стали ключевыми для выживания. Те особи, которым по наследству досталась такая мутация, вовсе не нужная их родителям, жившим в доинсектицидную эру, приобрели колоссальное преимущество перед теми, кто такой мутации не имел.
Естественный отбор «оценивает» вредность и полезность мутаций по их эффектам на выживание и размножение мутантных организмов в конкретных экологических условиях. При этом вредность мутации, как правило, обнаруживается немедленно, а ее полезность часто определяется задним числом: мы называем полезными те мутации, которые позволяют популяциям адаптироваться к изменяющимся условиям среды.
Чем сильнее фенотипический эффект мутации, тем вреднее такая мутация, тем выше вероятность того, что такая мутация будет отбракована отбором. Как правильно отметил Ч.Дарвин, природа не делает скачков. Ни одна сложная структура не может возникнуть в результате мутации с сильным фенотипическим эффектом. Новые признаки не возникают мгновенно, они формируется медленно и постепенно путем естественного отбора случайных мутаций со слабыми фенотипическими эффектами, которые чуть-чуть изменяют старые признаки.
Мутации случайны и не направлены.
Принципиальным положением мутационной теории является утверждение, что мутации случайны и не направлены. Под этим подразумевается, что мутации изначально не адаптивны. Применение инсектицидов не ведет к направленному возникновению мутаций устойчивости к ним у насекомых. Инсектициды могут приводить к общему повышению частоты мутаций, в том числе и мутаций в генах устойчивости к ним, в том числе и таких мутаций, которые эту устойчивость повышают. Но на одну такую «адаптивную» мутацию в «нужном» гене возникают десятки тысяч любых других – нейтральных и вредных - мутаций в генах, которые не имеют никакого отношения к устойчивости к инсектицидам.
Организм не может знать, какие мутации будут полезны в следующем поколении. Нет и не может быть механизма, который бы обеспечивал направленное появление полезных для организма мутаций. Это утверждение следует из всего того, что мы знаем о принципах кодирования, реализации и передачи генетической информации. Мы уже говорили о том, что ДНК – это не чертеж, а рецепт создания организма. Говорят, что генотип определяет фенотип. Не следует понимать эту фразу буквально. Генотип определяет не сам фенотип, а последовательности биохимических и морфогенетических реакций, которые, взаимодействуя друг с другом, определяют развитие фенотипических признаков. Изменения генотипа влекут за собой изменения фенотипа, но не наоборот. Как бы не менялся фенотип организма в ответ на воздействия внешней среды – его изменения не могут привести к изменению генов, которые этот организм передаст следующему поколению.
Роль хромосомных и геномных мутаций в эволюции.
Все перечисленные выше характеристики верны для всех типов мутаций – генных, хромосомных и геномных. Однако, такие геномные и хромосомные мутации как полиплоидия
(кратное увеличение количества хромосом) и дупликации
(удвоения определенных участков хромосом) играют особую роль в эволюции. Это связано с тем, что они увеличивают количество генетического материала и тем самым открывают возможность возникновения новых генов с новыми свойствами.
Расшифровка генома человека и других организмов показала, что многие гены и участки хромосом представлены в нескольких копиях. К ним относятся множество генов, отвечающих за синтез рибосомной РНК, гистонов (белков, участвующих в упаковке ДНК в хромосомах) и многих других. Таких генов нужно много для того, чтобы обеспечить высокий уровень синтеза, контролируемых ими продуктов. Следует ли из этого, что множественные копии этих генов возникли для этого
? Конечно же, нет. Удвоение всего генома или его отдельных участков происходило случайно. При этом удваивались не только эти гены, но и многие другие. Естественный отбор, однако, «поступал» с этим лишними копиями по-разному. Некоторые копии оказались полезными, и естественный отбор поддерживал их в популяциях. Другие оказались вредными, поскольку «больше - не всегда лучше». В этом случае отбор или отбраковывал носителей таких копий, или способствовал размножению таких особой, у которых излишние копии генов терялись в результате других хромосомных мутаций – делеций
. Были, наконец, и нейтральные копии, присутствие которых никак не сказывалось на приспособленности их носителей.
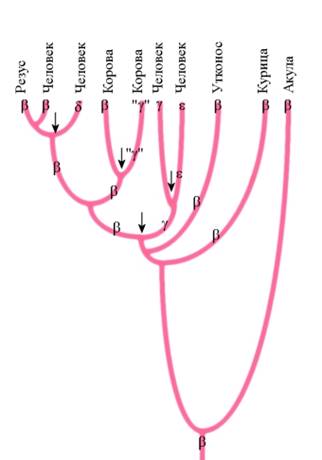 |
| Филогенетическое древо глобиновых генов. Ген глобина в ходе эволюции несколько раз дуплицировался (отмечено стрелками). Его добавочные копии затем приобретали новые свойства и функции. Из гена бета-глобина общего предка возникли гены гамма-, дельта-, эпсилон-глобинов – белков, которые выполняют иные функции, чем бета-глобин. |
Эти лишние копии становились резервом эволюции. Мутации в таких «резервных генах» не так строго отбрасывались отбором, как мутации в основных, уникальных генах. Резервным генам было «позволено» меняться в более широких пределах. Со временем они могли приобретать новые функции и становиться все более и более уникальными. Ярким примером последствий такого процесса является многочисленное и разнообразное семейство генов глобинов млекопитающих. Анализ последовательности нуклеотидов в этих генах показывается, что все они произошли в результате серии последовательных удвоений одного-единственного гена. За каждым удвоением следовало накопление случайных мутаций и постепенное изменение их функций, синтезируемых ими белков.
Когда мы сравниваем кариотипы разных видов млекопитающих, мы обнаруживаем, что в ходе эволюции этих видов происходили и закреплялись и другие хромосомные мутации, такие как транслокации и инверсии. Кариотип человека отличается от шимпанзе и других антропоидов одной транслокацией и несколькими инверсиями. За десятки миллионов лет независимой эволюции в кариотипах человека и землеройки возникли и закрепились десятки различных транслокаций и инверсий. Эти хромосомные перестройки не могли бы закрепиться, если бы они резко нарушали жизнеспособность или плодовитость их носителей.
В результате транслокаций и инверсий меняется взаимное расположение генов и, следовательно, характер их взаимодействия. В настоящее время хорошо известно, какую важную роль в проявлении генов играют их регуляторные элементы. Эти элементы, как правило, находятся в тех же хромосомах, что и контролируемые гены, но часто на большом расстоянии от них. Отрыв гена от его регуляторного элемента, обусловленный инверсией или транслокаций, или соединение этого гена с чужим регуляторным элементом может приводить к значительным изменениям в функции гена – времени его проявления в развитии, типе клеток, в которых этот ген активен, в количестве синтезируемого белка. К таким же последствиям может приводить и перемещение мобильных генетических элементов, которые могут захватывать и переносить с места на место регуляторные элементы.
В геноме обнаружены участки, где довольно часто происходят разрывы хромосом, ведущие к образованию хромосомных перестроек. Найдены и участки преимущественной локализации мобильных генетических элементов. Интересно, что во многих случаях это одни и те же участки. Таким образом, мы можем говорить о неслучайном распределении этих участков по геному. Однако, и как все остальные мутации, хромосомные перестройки и перемещения мобильных элементов случайны. Они случайно меняют функции генов, находящихся вблизи точек разрывов, они случайно распределяют гены по геному. Они приводят к тому, что возникает множество новых «коалиций» генов, а приспособительная ценность этих «коалиций» оценивается отбором.
Мутационный процесс является важнейшим фактором эволюции.
Мутационный процессизменяет гены и порядок их расположения в хромосомах и тем самым увеличивает генетическое разнообразие популяций. Он создает избыточные копии генов и тем самым открывает возможность усложнения организмов. Мутации возникают случайно и не направленно. Адаптивная ценность каждой мутации не постоянна. Она определяется взаимодействием мутантного аллеля с другими генами организма и с условиями среды, в которой развивается и живет мутантный организм.
Мутационный процесс, создавая новые аллели генов, изменяет частоты аллелей в популяциях. Пусть частота спонтанного мутирования аллеля B
(например, черной окраски лисиц) в аллель b
( красной окраски) равна 10-5
на гамету за поколение.Тогда частота аллеля b
в популяции будет медленно, но неуклонно возрастать в каждом поколении на одну десятитысячную, если этому возрастанию не будут препятствовать или способствовать другие факторы эволюции. В принципе, благодаря только мутационному процессу новый аллель может практически полностью вытеснить старый аллель из популяции. Однако, это будет происходить крайне медленно. Для того чтобы снизить частоту аллеля B
от 0,50 до 0,49 нужно 2000 поколений. Но дальше процесс пойдет еще медленнее. На снижение от 0,10 до 0,09 – уйдет 10000 поколений. Чем ниже частота аллеля, тем медленнее она будет изменяться за счет мутаций. Если бы мутационный процесс был единственным фактором эволюции, то сама эволюция происходила гораздо медленнее, чем на самом деле. Частоты генов в популяциях меняются не только и не столько за счет мутационного процесса, но благодаря действию других факторов эволюции.
Мута́ция
— стойкое (то есть такое, которое может быть унаследовано потомками данной клетки или организма) изменение генотипа, происходящие под влиянием внешней или внутренней среды. Процесс возникновения мутаций получил название мутагенеза
.
 Причины мутаций Причины мутаций
Мутации делятся на спонтанные
и индуцированные
. Спонтанные мутации возникают самопроизвольно на протяжении всей жизни организма в нормальных для него условиях окружающей среды с частотой около 10 в −9 степени — 10 в −12 на нуклеотид за клеточную генерацию. Индуцированными мутациями называют наследуемые изменения генома, возникающие в результате тех или иных мутагенных воздействий в искусственных (экспериментальных) условиях или при неблагоприятных воздействиях окружающей среды.
Мутации появляются постоянно в ходе процессов, происходящих в живой клетке. Основные процессы, приводящие к возникновению мутаций - репликация ДНК, нарушения репарации ДНК и генетическая рекомбинация.
Связь мутаций с репликацией ДНК
Многие спонтанные химические изменения нуклеотидов приводят к мутациям, которые возникают при репликации. Например, из-за деаминирования цитозина напротив него в цепь ДНК может включаться урацил (образуется пара У-Г вместо канонической пары Ц-Г). При репликации ДНК напротив урацила в новую цепь включается аденин, образуется пара У-А, а при следующей репликации она заменяется на пару Т-А, то есть происходит транзиция.
Связь мутаций с рекомбинацией ДНК
Из процессов, связанных с рекомбинацией, наиболее часто приводит к мутациям неравный кроссинговер. Он происходит обычно в тех случаях, когда в хромосоме имеется несколько дуплицированных копий исходного гена, сохранивших похожую последовательность нуклеотидов. В результате неравного кроссинговера в одной из рекомбинантных хромосом происходит дупликация, а в другой - делеция.
Связь мутаций с репарацией ДНК
Спонтанные повреждения ДНК встречаются довольно часто, такие события имеют место в каждой клетке. Для устранения последствий подобных повреждений имеется специальные репарационные механизмы (например, ошибочный участок ДНК вырезается и на этом месте восстанавливается исходный). Мутации возникают лишь тогда, когда репарационный механизм по каким-то причинам не работает или не справляется с устранением повреждений. Мутации, возникающие в генах белков, ответственных за репарацию, могут приводить к многократному повышению (мутаторный эффект) или понижению (антимутаторный эффект) частоты мутирования других генов. Так, мутации генов многих ферментов системы эксцизионной репарации приводят к резкому повышению частоты соматических мутаций у человека, а это, в свою очередь, приводит к развитию пигментной ксеродермы и злокачественных опухолей покровов.
Мутагены
Основная статья
: Мутагены
Существуют факторы, способные заметно увеличить частоту мутаций — мутагенные факторы. К ним относятся:
- химические мутагены — вещества, вызывающие мутации,
- физические мутагены — ионизирующие излучения, в том числе естественного радиационного фона
, ультрафиолетовое излучение, высокая температура и др.,
- биологические мутагены — например, ретровирусы, ретротранспозоны.
Классификации мутаций
Существует несколько классификаций мутаций по различным критериям. Мёллер предложил делить мутации по характеру изменения функционирования гена на гипоморфные
(измененные аллели действуют в том же направлени, что и аллели дикого типа; синтезируется лишь меньше белкового продукта), аморфные
(мутация выглядит, как полная потеря функции гена, например, мутация white
у Drosophila), антиморфные
(мутантный признак изменяется, например, окраска зерна кукурузы меняется с пурпурного на бурый) и неоморфные
.
В современной учебной литературе используется и более формальная классификация, основанная на характере изменения структуры отдельных генов, хромосом и генома в целом. В рамках этой классификации различают следующие виды мутаций:
- генные
- хромосомные
- геномные
.
Геномные
: — полиплоидизация
(образование организмов или клеток, геном которых представлен более чем двумя (3n, 4n, 6n и т. д.) наборами хромосомами) и анеуплоидия (гетероплоидия) - изменение числа хромосом, не кратное гаплоидному набору (см. Инге-Вечтомов, 1989). В зависимости от происхождения хромосомных наборов среди полиплоидов различают аллополиплоидов, у которых имеются наборы хромосом, полученные при гибридизации от разных видов, и аутополиплоидов, у которых происходит увеличение числа наборов хромосом собственного генома, кратное n.
При хромосомных мутациях
происходят крупные перестройки структуры отдельных хромосом. В этом случае наблюдаются потеря (делеция) или удвоение части (дупликация) генетического материала одной или нескольких хромосом, изменение ориентации сегментов хромосом в отдельных хромосомах (инверсия), а также перенос части генетического материала с одной хромосомы на другую (транслокация) (крайний случай — объединение целых хромосом, т.н. Робертсоновская транслокация, которая является переходным вариантом от хромосомной мутации к геномной).
На генном
уровне изменения первичной структуры ДНК генов под действием мутаций менее значительны, чем при хромосомных мутациях, однако генные мутации встречаются более часто. В результате генных мутаций происходят замены, делеции и вставки одного или нескольких нуклеотидов, транслокации, дупликации и инверсии различных частей гена. В том случае, когда под действием мутации изменяется лишь один нуклеотид, говорят о точковых мутациях. Поскольку в состав ДНК входят азотистые основания только двух типов — пурины и пиримидины, все точковые мутации с заменой оснований разделяют на два класса: транзиции (замена пурина на пурин или пиримидина на пиримидин) и трансверсии (замена пурина на пиримидин или наоборот). Возможны четыре генетических последствия точковых мутаций: 1) сохранение смысла кодона из-за вырожденности генетического кода (синонимическая замена нуклеотида), 2) изменение смысла кодона, приводящее к замене аминокислоты в соответствующем месте полипептидной цепи (миссенс-мутация), 3) образование бессмысленного кодона с преждевременной терминацией (нонсенс-мутация). В генетическом коде имеются три бессмысленных кодона: амбер — UAG, охр — UAA и опал — UGA (в соответствии с этим получают название и мутации, приводящие к образованию бессмысленных триплетов - например амбер-мутация), 4)обратная замена (стоп-кодона на смысловой кодон).
По влиянию на экспрессию генов
мутации разделяют на две категории: мутации типа замен пар оснований
и типа сдвига рамки считывания (frameshift)
. Последние представляют собой делеции или вставки нуклеотидов, число которых не кратно трем, что связано с триплетностью генетического кода.
Первичную мутацию иногда называют прямой мутацией
, а мутацию, восстанавливающую исходную структуру гена, — обратной мутацией,
или реверсией. Возврат к исходному фенотипу у мутантного организма вследствие восстановления функции мутантного гена нередко происходит не за счет истинной реверсии, а вследствие мутации в другой части того же самого гена или даже другого неаллельного гена. В этом случае возвратную мутацию называют супрессорной. Генетические механизмы, благодаря которым происходит супрессия мутантного фенотипа, весьма разнообразны.
Последствия мутаций для клетки и организма
Мутации, которые ухудшают деятельность клетки в многоклеточном организме, часто приводят к уничтожению клетки (в частности, к программируемой смерти клетки, — апоптозу). Если внутри- и внеклеточные защитные механизмы не распознали мутацию и клетка прошла деление, то мутантный ген передастся всем потомкам клетки и, чаще всего, приводит к тому, что все эти клетки начинают функционировать иначе.
Мутация в соматической клетке сложного многоклеточного организма может привести к злокачественным или доброкачественным новообразованиям, мутация в половой клетке
— к изменению свойств всего организма-потомка.
В стабильных (неизменных или слабо изменяющихся) условиях существования большинство особей имеют близкий к оптимальному генотип, а мутации вызывают нарушение функций организма, снижают его приспособленность и могут привести к смерти особи. Однако в очень редких случаях мутация может привести к появлению у организма новых полезных признаков, и тогда последствия мутации оказываются положительными; в этом случае они являются средством адаптации организма к окружающей среде и, соответственно, называются адаптационными
.
Роль мутаций в эволюции
При существенном изменении условий существования те мутации, которые раньше были вредными, могут оказаться полезными. Таким образом, мутации являются материалом для естественного отбора. Так, мутанты-меланисты
(темноокрашенные особи) в популяциях березовой пяденицы (Biston betularia) в Англии впервые были обнаружены учеными среди типичных светлых особей в середине XIX века. Темная окраска возникает в результате мутации одного гена. Бабочки проводят день на стволах и ветвях деревьев, обычно покрытых лишайниками, на фоне которых светлая окраска является маскирующей. В результате промышленной революции, сопровождающейся загрязнением атмосферы, лишайники погибли, а светлые стволы берез покрылись копотью. В результате к середине XX века (за 50-100 поколений) в промышленных районах темная морфа почти полностью вытеснила светлую. Было показано, что главная причина преимущественного выживания черной формы - хищничество птиц, которые избирательно выедали светлых бабочек в загрязненных районах.
Если мутация затрагивает «молчащие» участки ДНК, либо приводит к замене одного элемента генетического кода на синонимичный, то она обычно никак не проявляется в фенотипе (проявление такой синонимичной замены может быть связано с разной частотой употребления кодонов). Однако методами генного анализа такие мутации можно обнаружить. Поскольку чаще всего мутации происходят в результате естественных причин, то в предположении, что основные свойства внешней среды не менялись, получается, что частота мутаций должна быть примерно постоянной. Этот факт можно использовать для исследования филогении
- изучения происхождения и родственных связей различных таксонов, в том числе и человека
. Таким образом, мутации в молчащих генах служат для исследователей своеобразными «молекулярными часами». Теория "молекулярных часов" исходит также из того, что большинство мутаций нейтральны, и скорость их накопления в данном гене не зависит или слабо зависит от действия естественного отбора и потому остается постоянной в течение длительного времени. Для разных генов эта скорость, тем не менее, будет различаться.
Исследование мутаций в митохондриальной ДНК (наследуется по материнской линии) и в Y-хромосомах (наследуется по отцовской линии) широко используется в эволюционной биологии для изучения происхождения рас и народностей, реконструкции биологического развития человечества.
Проблема случайности мутаций
В 40-е годы среди микробиологов была популярна точка зрения, согласно которой мутации вызываются воздействием фактора среды (например. антибиотика), к которому они позволяют адаптироваться. Для проверки этой гипотезы был разработан флуктуационный тест и метод реплик.
Флуктуационный тест Лурия-Дельбрюка заключается в том, что небольшие порции исходной культуры бактерий рассеивают в пробирки с жидкой средой, а после нескольких циклов делений добавляют в пробирки антибиотик. Затем (без последующих делений) на чашки Петри с твердой средой высевают выживших устойчивых к антибиотику бактерий. Тест показал. что число устойчивых колоний из разных пробирок очень изменчиво - в большинстве случаев оно небольшое (или нулевое), а в некоторых случаях очень высокое. Это означает, что мутации, вызвавшие устойчивость к антибиотику, возникали в случайные моменты времени как до, так и после его воздействия.
Метод реплик (в микробиологии) заключается в том, что с исходной чашки Петри, где на твердой среде растут колонии бактерий, делается отпечаток на ворсистую ткань, а затем с ткани бактерии переносятся на несколько других чашек, где рисунок их расположения оказывается тем же, что на исходной чашке. После воздействия антибиотиком на всех чашках выживают колонии, расположенные в одних и тех же точках. Высевая такие колонии на новые чашки, можно показать, что все бактерии внутри колонии обладают устойчивостью.
Таким образом, обоими методами было доказано, что «адаптивные» мутации возникают независимо от воздействия того фактора, к которому они позволяют приспособиться, и в этом смысле мутации случайны. Однако несомненно, что возможность тех или иных мутаций зависит от генотипа и канализована предшествующим ходом эволюции (см. Закон гомологических рядов в наследственной изменчивости
). Кроме того, закономерно различается частота мутирования разных генов и разных участков внутри одного гена. Также известно, что высшие организмы используют «целенаправленные» (то есть происходящие в определенных участках ДНК) мутации в механизмах иммунитета
. С их помощью создаётся разнообразие клонов лимфоцитов
, среди которых в результате всегда находятся клетки, способные дать иммунный ответ на новую, неизвестную для организма болезнь. Подходящие лимфоциты подвергаются положительной селекции, в результате возникает иммунологическая память.
МУТАГEНЫ (от мутации и греч. -genes-рождающий, рожденный), хим. и физ. факторы, вызывающие наследств. изменения - мутации. Впервые искусств. мутации получены в 1925 Г. А. Надсе-ном и Г. С. Филипповым у дрожжей действием радиоактивного излучения радия; в 1927 Г. Мёллер получил мутации у дрозофилы действием рентгеновских лучей. Способность хим. в-в вызывать мутации (действием иода на дрозофилы) открыта в 1932 В. В. Сахаровым.
М. химические.
Различают М. прямого действия -соед., реакц. способность к-рых достаточна для хим. модификации ДНК, РНК и нек-рых белков, и промутагены - в-ва, к-рые сами по себе инертны, но превращ. в организме в М. (в осн. в результате ферментативного окисления системой микро-сомных многоцелевых оксидаз). Последние часто называют "конечными" М. Так, немутагeнный 1,2-бензопкрен (бен-зо[а]пирен) в организме окисляется до 7,8-дигидрокси-9,10-эпокси-7,8,9,10-тетрагидро-1,2-бензопирена, один из стерео-изомеров к-рого обладает мутагенной, а также канцерогенной активностью:
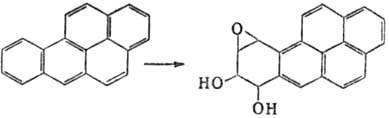
Др. пример - немутагенный N-нитрозодиметиламин, к-рый в печени подвергается ферментативному окислит. деметили-рованию и превращ. в высокомутагенный и канцерогенный метилдиазогидроксид (см. Канцерогенные вещества).
Мишенью действия М. в клетке являются гл. обр. ДНК и, возможно, нек-рые белки. К последним относят в осн. белки, играющие структурную роль в организации генома или принимающие участие в репликации (самовоспроизведении молекулы нуклеиновых к-т), рекомбинации (перераспределении генетич. материала родителей в потомстве) или репарации (восстановлении поврежденной структуры ДНК).
Среди М. наиб. обширен класс электроф. алкилирующих М., к к-рым относят не только типичные алкилирующие агенты (диазоалканы, эфиры серной к-ты и алкансульфо-кислот), но и эфиры фосфорной и азотной к-т, аминоэтили-рующие реагенты (2-хлорэтиламин, этиленимин и их производные), оксиэтилирующие агенты (этиленоксид и его производные) и альдегиды. К этому же классу М. относят N-нитрозо-N-алкиламиды карбоновых к-т, N-нитрозо-N-алкилуретаны, N-нитрозо-N-алкилмочевины, N-алкил-N-нитрозо-N'-нитрогуанидины, являющиеся, по-видимому, наиб. активными из известных М. Эти соед. сами по себе лишены алкилирующих св-в, но при их гидролитич. распаде образуются активные алкилдиазогидрохсиды (иногда считают, что образуются своб. алкилкарбкатионы, что менее вероятно).
Электроф. реагентами являются также подавляющее большинство конечных М., образующихся из синтетич. и прир. в-в. Пример последних - глюкозид циказин (b-D-глю-козид метилазоксиметанола), к-рый под действием ферментов трансформируется в метилдиазогидроксид.
Механизм мутагенного действия простейших алкилирующих агентов довольно хорошо изучен. Характер повреждений ДНК при воздействии этих агентов м.б. предсказан с помощью ф-лы Свена-Скотта:
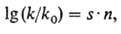
где k- константа скорости бимолекулярной р-ции алкили-рующего агента с нуклеофилом; k0
- константа скорости бимолекулярной р-ции алкилирующего агента с водой, выбранной в качестве стандарта; n-константа, характеризующая нуклеофильность субстрата; .s-мера чувствительности скорости р-ции к изменению п.
Из ф-лы следует, что при алкилировании биополимеров, имеющих неск. нуклеоф. центров, доля продуктов алкили-рования центра с низкой нуклеофильностью должна быть выше при действии агента с низкой константой s.
Наибю нуклеоф. центр в молекуле ДНК-положение 7 в гуанине. Поэтому диметилсульфат и метилметансульфоиат (s соотв. 0,9 и 0,86) алкилируют ДНК в осн. по этому положению (см. ф-лу I; везде R-цепь ДНК), а выход продуктов алкилирования центров с меньшей нуклеофильностыо мал. При действии на ДНК этилметансульфоната и N-нитрозо-N-метилмочевины (s соотв. 0,67 и 0,42) доля 7-алкилгуанина в общем выходе продуктов алкилирования снижается и возрастает вклад продуктов алкилирования центров с низкой нуклеофильностью - межнуклеотидных фосфатных групп и атомовкислорода в основаниях. При алкилировании ДНК N-нитрозо-N-этилмочевиной (s ок. 0,26) осн. продукт р-ции -алкилир. остатки фосфорной к-ты и О-алкилир. основания: О6
-этилгуанин (II), О2
-этилцитозин (III), О2
- и О4
-этилтимины (соотв. IV и V).
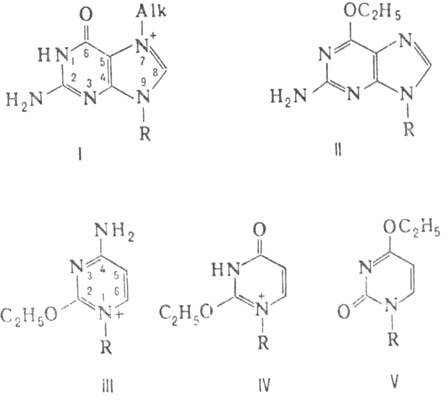
Синтез новой нити ДНК на ДНК-матрице, содержащей 7-алкилгуаниновые звенья, проходит без ошибок и мутаций не возникает. Напротив, полинуклеотид, синтезированный на матрице, содержащей О-алкилир. звенья, содержит ошибочно включенные пуриновые и пиримидиновые основания. С возрастанием способности к О-алкилированию у простейших алкилирующих агентов возрастает канцерогенная активность.
Мутации, возникающие при действии метилметансульфо-ната, не алкилирующего основания ДНК по атому О, являются следствием ошибок работы ферментовклетки, восстанавливающих исходное (неизмененное) состояние ДНК.
Спектр повреждений ДНК при действии нуклеоф. мутаге-нов (гидроксиламин, О-метилгидроксиламин, гидразины, бисульфит Na) значительно уже. В осн. это модификация цитозина, направление и механизм к-рой иллюстрирует след. схема:
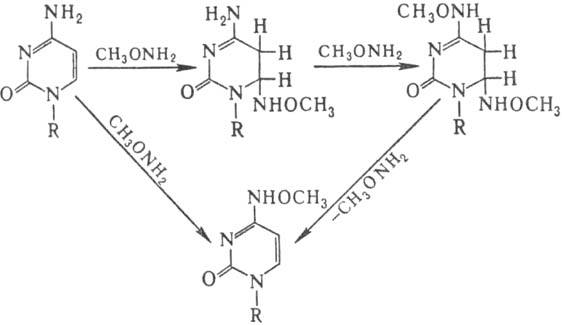
Др. нуклеоф. агенты реагируют аналогично, но константы скорости отдельных стадий могут существенно меняться. Особенность действия бисульфита на ДНК-замена цитози-новых звеньев на урацилъные в результате дезаиминирова-ния цитозина по схеме:
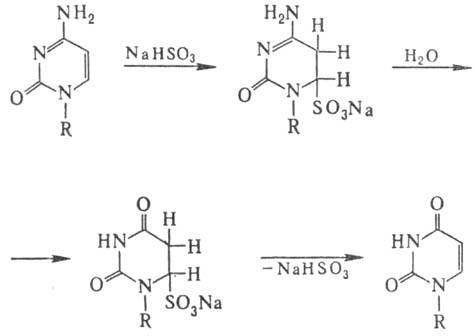
Ряд М. вызывают мутации, не связываясь ковалентно с ДНК. Так, нек-рые гетероциклич. соед. (напр., акридин и его производные), обладающие жесткой плоской структурой молекулы, встраиваются (интеркалируют) между смежными, расположенными стопкой, парами оснований двойной спирали ДНК. В этом случае матричный синтез на ДНК протекает с ошибками. В синтезируемой нити ДНК оказывается на один нуклеотид больше или меньше обычного и возникают мутации.
Особый класс М. составляют соед., представляющие со-бой аналоги оснований ДНК-5-галогенурацилы, 2-амино-и 6-метиламинопурины и др. Галогенурацилы включаются в ДНК при матричном синтезе вместо тимина, 2-амино-пурин-вместо аденина. Вследствие различий в положении кетоенольного равновесия у тимина и галогенурацилов (при включении последних в ДНК) увеличивается частота ошибочных спариваний оснований и возникают ошибки при репликации.
Существуют также М., ингибирующие синтез предшественниковДНК. Мишень таких М.-ферменты, синтезирующие компоненты ДНК. Считается, что в результате подавления синтеза предшественников происходит замедление или даже остановка синтеза ДНК. В этих условиях повышается вероятность того, что решшкац. система клетки может пропустить отсутствующий нуклеотид, либо включить вместо отсутствующего нуклеотида другой, ошибочный; следствие обоих событий-мутация.
Мутагенные и канцерогенные св-ва хим. в-в тесно связаны между собой. Поэтому выявление возможных М. в окружающей среде, испытание на мутагенность продуктов пром. синтеза (красители, лек. ср-ва, пестициды и др.)-важная задача совр. генетики. Разработаны тест-системы для экспресс-обнаружения М. Наиб. часто используют тест Б. Эймса и его модификации. Для их осуществления используют специально полученные штаммы бактерий Salmonella typhimurium, к-рые не способны синтезировать гистидин из-за генетич. нарушений. Этот штамм поэтому не может расти в среде, в к-рой отсутствует эта аминокислота. Др. его особенность-способность в результате обратной мутации приобретать исходную способность синтезировать гистидин из обычных предшественников (NH3
и др.). Частота обратных мутаций заметно увеличивается под действием М. и может служить критерием их активности. В тестах используют лишенную гистидина питат. среду, в к-рую добавляют экстракт из печени крыс, содержащий ферменты эндоплаз-матич. ретикулума, способные превращать в-во в мутаген-ную (канцерогенную) форму. В случаях, когда в-во обладает мутагенной активностью, наблюдается активный рост колоний бактерий. Время тестирования-ок. 24 ч (на тестирование с использованием эксперим. животных затрачивается 2-3 года).
М. физические.
Мутации при действии физических М. возникают так же, как и при действии М. химических. Вначале возникает первичное повреждение ДНК. Если оно не будет полностью исправлено в результате репарации, то при послед. репликативном синтезе ДНК будут возникать мутации. Специфика мутагенеза (процесса возникновения мутаций) при действии физ. факторов связана с характером первичных повреждений генома, вызываемых ими.
Подробно изучены повреждения ДНК, возникающие в результате действия электромагн. излучения разной длины волны. Электромагн. излучение с длиной волны больше 300 нм не поглощается ДНК, однако в нек-рых случаях может оказывать мутагенное действие, механизм к-рого заключается в поглощении кванта света молекулой сенсибилизатора и передачей энергии возбуждения на ДНК. При действии света с длиной волны 200-300 нм (lмакс
поглощения ДНК 260 нм) происходит поглощение квантов света хромофорными группами ДНК (пуриновые и пиримидиновые основания) и переход последних в возбужденное состояние. В обоих случаях б.ч. поглощенной энергии рассеивается и основания ДНК возвращаются в исходное, невозбужденное, состояние, но часть возбужденных оснований подвергается фотохим. трансформации.
В наиб. степени это относится к тимину, остальные основания более устойчивы. Энергия излучения, поглощенная тимином, локализуется в осн. на двойной связи цикла; следствием этого является образование в составе ДНК димеров:
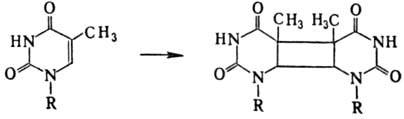
Возбужденная двойная связь способна также присоединять нуклеофилы с образованием, напр., нестойкого гидрататимина:
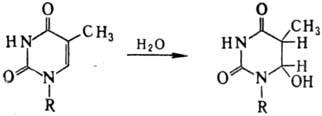
При облучении ДНК дальним УФ светом (185 нм) возможно и фотоокислениетимина с образованием его гидроперок-сида (ф-ла VI).
Предполагают, что осн. предмутац. повреждение ДНК, возникающее при действии УФ света, связано с образованием димеров тимина.
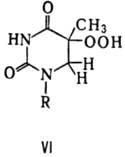
Электромагн. излучения еще более высокой энергии (рентгеновское и у-излучение) способны ионизовать в-во. Ионизация происходит случайным образом, поэтому молекулы, являющиеся наиб. распространенными в объекте, больше других под вергаются ионизации. При облучении живой материи, на 70-90% состоящей из воды, б. ч. энергии будет поглощена молекуламиводы и поэтому мутагенный эффект при действии этих агентов возникает гл. обр. вследствие модификации ДНК продуктами радиолизаводы. Наиб. вклад в развитие радиац. поражения ДНК вносит радикал ОН.
. При взаимод. с ДНК 80% всех радикалов ОН.
атакуют основания ДНК, остальные-дезоксирибозную часть молекулы. Возникающие первичные продукты затем вступают в разнообразные вторичные р-ции как с теми же продуктами радиолизаводы, так и с кислородом, белками, низкомол. компонентами клетки, а также подвергаются диспропорционированию, изомеризации, гидролизу. Возникает широкий спектр разнообразных изменении первичной и вторичной структуры ДНК: измененные основания, апури-новые и апиримидиновые сайты (участки с удаленными основаниями), разрывы связей в дезоксирибозе, одно- и двунитевые разрывы цепей ДНК. Точная роль каждого из возникающих повреждений структуры ДНК в формировании мутагенного эффекта все еще остается невыясненной. Предполагают, что ключевую роль в этом процессе играют продукты радиолизатимина.
В тимине первичной атаке радикалом ОН.
подвергается двойная связь цикла с образованием двух радикалов с радикальными центрами в положениях 5 и 6, с преобладанием первого. В присут. кислорода из радикала образуется гидро-пероксид, к-рый уже при комнатной т-ре разлагается с раскрытием цикла:

Радиационно-хим. изменения цитозина также протекают через стадию образования аналогичного, но еще более нестабильного гидропероксида. В случае цитозина и аденина возможно также дезаминирование оснований. Пуриновые основания (аденин, гуанин) реагируют с радикалом ОН.
с меньшей скоростью. Идентифицированы, напр., продукт гидроксилирования аденина (8-гидроксиаденин), а также продукты раскрытия имидазольного кольца этих оснований.
В дезоксирибозном остатке молекулыДНК радикал ОН.
способен атаковать и отрывать атом Н из любого положения цикла. Конечный результат радиационно-хим. модификации этой части молекулыДНК - образование малонового диальдегида, сопровождающееся разрывом N-гликозидной связи с отщеплением неизмененного основания, и, в итоге, однонитевых разрывов ДНК. Затем уже возникают и двунитевые разрывы ДНК в результате статистич. накопления однонитевых разрывов в противоположных участках двух цепей ДНК.
Известно прямое действие радиации на молекулы-мишени, в результате к-рого макромолекулы превращ. в своб. радикалы по схеме:
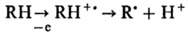
Судьба образовавшихся радикалов R.
, вероятно, схожа с судьбой радикалов, возникающих при непрямом действии радиации.
Механизм действия др. видов ионизирующего излучения (a-частицы, протоны, нейтроны, электроны, ускоренные ядра более тяжелых элементов) близок к механизму действия ионизирующего электромагн. излучения. Отличия обусловлены гл. обр. разницей в массе, заряде, энергии и глубине проникновения излучения в объект, способом ионизации макромолекул и др. Имеются сведения, что воздействие нек-рых др. физ. факторов, напр. звуковых колебаний, вибрации, могут также привести к мутации.
Изменчивость и мутации
Изменчивость – это способность организмов приобретать новые признаки. Изменчивость организмов связана как с изменчивостью генотипа, так и с влиянием окружающей среды.
Признаки в популяции (совокупности свободно скрещивающихся особей, длительное время существующих на более-менее обособленной территории) могут изменяться как непрерывно, так и дискретно. К дискретно изменчивым признакам относится, например, группа крови у человека. В этом случае признак может принимать лишь некоторые формы; промежуточные формы отсутствуют. Дискретная изменчивость возникает по признакам, контролируемым одним или двумя генами. При этом внешние условия редко влияют на фенотип.
1
|
| Непрерывная изменчивость и закон нормального распределения |
Признаки, контролируемые множеством генов, обычно меняются непрерывно, причём их крайние проявления (большие отклонения от среднестатистического значения признака) наблюдаются у относительно небольшого количества особей. У большинства же особей рассматриваемый признак не очень сильно отличается от его среднего значения. Примерами непрерывной изменчивости являются изменения массы или длины, а также формы тела. Непрерывная изменчивость в значительной мере обусловлена влиянием на фенотип внешней среды.
Наследственную изменчивость (т.е. изменчивость, передающуюся по наследству от организма к организму, от клетки к клетке) можно разделить на комбинативную и мутационную. Источником комбинативной изменчивости является мейоз. Обмен генами между гомологичными хромосомами в профазе I мейоза, случайная ориентация и последующее независимое расхождение хромосом, слияние мужской и женской гамет в зиготу являются причиной практически неограниченного различий между особями в популяции. Такая изменчивость получила название комбинативной; при этой форме наследственной изменчивости происходит перетасовка имеющихся в популяции генов (аллелей), но не образование новых.
Случайное соединение генов в новых комбинациях является источником генетического разнообразия, но всё же не может породить крупных изменений в генотипе, которые приведут к образованию нового вида. Причиной таких изменений могут быть
мутации – изменения структуры (генные мутации) или количества ДНК (хромосомные мутации) в организме. Мутации возникают случайным образом; на их частоту влияют продолжительность жизни организма и внешние условия (например, коротковолновое излучение). Усиливают мутации и некоторые вещества: кофеин, формальдегид, никотин, некоторые лекарственные препараты и пищевые добавки. Изменчивость, обусловленная мутациями, получила название мутационной.
Мутации, возникающие в половых клетках, передаются последующим поколениям, в то время как мутации, возникающие в прочих (соматических) клетках, наследуются только дочерними клетками.
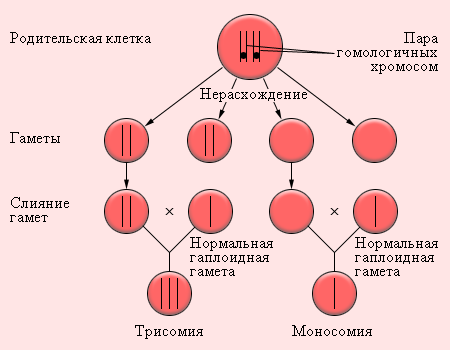
2
|
| Нерасхождение хромосом |
Геномные мутации возникают, как правило, в ходе мейоза и приводят к приобретению или утрате отдельных хромосом (
анэуплоидии) или гаплоидных наборов хромосом (
полиплоидии). Анэуплоидия чаще всего выражается в наличии добавочной хромосомы или отсутствии одной из хромосом. Её причиной является нерасхождение хромосом при мейозе. Зиготы, в которых количество хромосом меньше, чем обычно, как правило, не развиваются, но зиготы с увеличенным набором хромосом могут развиваться во взрослые организмы. Обычно это сопровождается значительными аномалиями в строении и функционировании организма. Типичным примером анэуплоидии является
синдром Дауна у человека (трисомия по 21-ой хромосоме)..
В результате полиплоидии количество хромосом в гамете увеличивается в целое число раз. Это происходит из-за того, что хромосомы делятся и расходятся, но цитоплазма и клеточная мембрана остаются общими, что приводит к возникновению дочерней клетки с большим ядром. Эта клетка впоследствии делится, давая начало новым особям. Полиплоидные растения обычно более крупные и устойчивые к заболеваниям. Полиплоидия может вызываться как случайным образом, так и искусственным путём (например, воздействием на половые клетки некоторыми химическими веществами). Одной из форм этой мутации является
аллополиплоидия – удвоение числа хромосом у стерильной особи, в результате чего она приобретает способность к размножению. Полиплоидия особенно распространена у растений, размножающихся вегетативным путём; более половины цветковых растений (и большинство культурных растений) – полиплоиды.
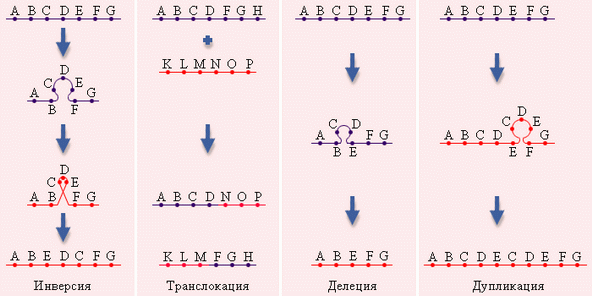
3
|
| Хромосомные перестройки |
В результате другой разновидности хромосомных мутаций изменяется взаимное расположение генных локусов. Подобные хромосомные перестройки могут выражаться в
инверсии (повороте на 180° части генного кода по отношению к окружающим генам),
транслокации (перемещению одних локусов относительно других), которые не изменяют генотипа, но могут привести к изменению фенотипа,
делеции (утрате хромосомой каких-либо участков) и
дупликации (повторению части генов в хромосоме), приводящих к изменению генотипа.
Мутировать могут не только хромосомы, но и отдельные гены. При этом изменяется последовательность нуклеотидов в определённом участке хромосомы, что влечёт за собой изменение последовательности аминокислот в белковой цепи. Отдельные нуклеотиды могут просто добавляться, дублироваться, заменяться на другие, удаляться из цепи, перемещаться по отношению к другим нуклеотидам. Генные мутации в соматических клетках могут порождать клетки с повышенной скоростью деления, что приводит к образованию опухолей. Опухоли, оказывающие негативное влияние на организм (злокачественные опухоли), являются причиной раковых заболеваний.
Многие мутации заканчиваются летальным исходом, другие не оказывают никакого влияния на организм. Некоторые мутации приводят к усилению благоприятных признаков, повышению генетического разнообразия организма, что способствует изменчивости популяции и может привести к образованию новых видов.
Загрязнение окружающей среды может негативно влиять на плод
Пыль и другие виды загрязнения окружающей среды могут влиять не только на животных и людей, но также и на развивающиеся плоды, о чем сообщают исследователи. Они обнаружили, что генетические мутации, которые, как известно, вызываются некоторыми загрязняющими агентами, могут передаваться через сперму мышам-зародышам. Вероятно, то же самое может произойти и с людьми.
У мышей, вдыхавших загрязненный воздух от сталелитейного завода, гораздо чаще рождались дети с явными генетическими мутациями по сравнению с мышами, вдыхавшими отфильтрованный воздух, что и обнаружила группа ученых из Макмастерского университета в г. Торонто.
"Наше исследование определяет переносимые воздухом частицы, которые вносят свой вклад в стимулирование наследственных мутаций у мышей, однако прямая связь между мутациями и влиянием на здоровье пока не была установлена", - пишут они.
"Несмотря на это, структурные изменения в ДНК были выявлены и в сперме людей после контакта с загрязненным воздухом", - пишут исследователи.
Загрязнение воздуха также было связано с сердечно-сосудистыми заболеваниями, раком легких и врожденными пороками развития, говорят ученые, ссылаясь на многочисленные исследования.
Кристофер Сомерс (Christopher Somers) и коллеги поместили мышей в клетках в точках, в которые ветер доносил загрязнение со сталелитейного завода в г. Гамильтон (штат Онтарио). Одни мыши получали отфильтрованный воздух, а другие получали то, что приносит ветер.
"Мыши, вдыхавшие отфильтрованный воздух в индустриальной области города, имели на 52% меньше мутаций со стороны отца, чем мыши, вдыхавшие неотфильтрованный воздух", - пишут они.
Пока неясно, каким образом вдыхаемые загрязняющие вещества вызывают мутации.
|
