Контрольная работа №1
2. Сущность жизни и характерные свойства живого организма. Клетка как носитель жизни.
2.1 Жизнь – это особая, высшая по сравнению с физической и химической, форма движения материи, которая возникла на определенном этапе ее исторического развития и представлена на нашей планете большим количеством индивидуальных систем.
«Жизнь, - по определению Ф. Энгельса, - есть способ существования белковых тел, и этот способ существования состоит по своему существу в постоянном самообновлении химических основных частей этих тел». Таким образом, Ф. Энгельс охарактеризовал «белковые тела» как материальный субстрат жизни, а обмен веществ – как основной фактор его существования.
Понятие о белковых телах близко к современным представлениям о протопласте, состоящем из белков, нуклеиновых кислот, липидов, сложных углеводов и других органических веществ. Оно не совпадает с понятием о химически индивидуальных белках, которые можно выделить и изолировать из живых организмов. Всюду, где возникла жизнь, были найдены белки. Они составляют структурную основу протопласта клетки, биокатализаторов (ферментов), запасных веществ, играют решающую роль во всех жизненных процессах и выполняют самые разнообразные функции. Протопласту как полимолекулярной системе свойственны форма движения, характерная для жизни, биологический обмен веществ, который представляет собой основу жизненных процессов, происходящих в растительных организмах, является совокупностью большого количества биохимических и биофизических реакций в клетках, связанных в единое целое.
Современное естествознание расширило и конкретизировало определение сущности жизни, данное Ф. Энгельсом. Было выяснено, что развитие любых организмов тесно связано не только с белками, но и с нуклеиновыми кислотами ДНК и РНК – носителями наследственной информации об организме. Основными молекулами живых систем (организмов) являются биополимеры: белки (полипептиды) и ДНК и РНК (полинуклеотиды), а основной признак жизни – самовоспроизведение самообновление белковых тел, в основе которого лежит саморепликация, т.е. удвоение молекулы ДНК с передачей рождающейся клетке генетической информации. В соответствии с этим академик Г. И. Гольданский дает следующее определение сущности жизни: жизнь есть форма существования биополимерных тел (систем), способных к саморепликации в условиях постоянного обмена веществом и энергией с окружающей средой.
Реклама

Обмен веществ состоит из ассимиляции и диссимиляции, т.е. включает процессы синтеза и распада. С помощью обмена веществ осуществляется взаимодействие организма с окружающей средой. Каждому организму свойственна наследственность, которая обеспечивает определенный порядок обмена веществ, исторически сложившийся под непосредственным влиянием внешних условий. Кроме того, растительный организм обладает свойством избирательности, которая выражается потребностью в определенных условиях, необходимых для его роста и развития.
2.2 Одним из величайших достижений XIX в. является учение о клеточном строении организмов. Ф. Энгельс считал создание клеточной теории не менее великим завоеванием человеческих знаний, чем открытие закона превращения энергии и эволюционной теории. Клеточная теория служит доказательством единства строения и развития всех живых существ.
Открытие клеток и разработка клеточной теории строения животных и растительных организмов оказали огромное влияние на развитие биологии и медицины. С применением электронного микроскопа успешно проводятся исследования субмикроскопической организации клетки, открыты неизвестные ранее структуры, о существовании которых даже не догадывались. Это прежде всего различные полимембранные системы, на которых осуществляются процессы обмена веществ. Познание ультраструктуры клетки и интегрирование явлений обмена веществ в ней дали возможность проникнуть в сущность биохимических процессов и превращение энергии в клетке на молекулярном уровне. Молекулярная организация тесно связана со структурой и функцией, структурой и обменом, что дает определенные представления о живой клетке как единой морфологической физиологической диалектической системе.
Таким образом клетка является элементарной единицей полноценной живой системы. Клетку следует считать важнейшим этапом в развитии жизни на Земле. Она является морфологической и физиологической структурой, элементарной единицей растительных и животных организмов. Возникновение многоклеточности в процессе филогенеза и онтогенеза сопровождается постепенным ограничением и даже потерей клетками их физиологической активности и генетической потенции, т.е. оставаясь гомологическими структурами, клетки перестают быть аналогичными. На современном этапе развитии науки можно считать, что комплексы клеток – это своеобразные, в известной мере индивидуализированные системы (гистосистемы).
Реклама

В онтогенезе постепенно создаются системы клеток определенной структуры. Клетки в этих системах утрачивают свою индивидуальность, т.е. теряют способность к независимой жизнедеятельности. Это явление называют клеточной интеграцией.
32. растительная клетка как осмотическая система. Связь между осмотическим давлением и концентрацией клеточного сока.
Клетка представляет собой осмотическую систему. Растительная клетка окружена клеточной стенкой, которая обладает эластичностью и может растягиваться. Плазмалемма и тонопласт обладают избирательной проницаемостью, легко пропускают воду и менее проницаемы для растворенных веществ. В этом можно убедиться рассмотрев явления плазмолиза и тургора.
Если поместить клетку в раствор более высокой концентрации, чем в клетке, то протоплазма (цитоплазма) отстает от клеточной стенки – это явление получило название плазмолиз. Плазмолиз происходит в результате того, что под воздействием более концентрированного внешнего раствора вода выходит из клетки.
При помещении клетки в чистую воду или слабоконцентрированный раствор вода поступает в клетку. Объем вакуоли возрастает, клеточный сок давит на цитоплазму и прижимает ее к клеточной оболочке. Под влиянием внутреннего давления клеточная стенка растягивается, а клетка переходит в напряженное состояние (тургор).
Явление плазмолиза показывает, что клетка жива и цитоплазма сохранила полупроницаемость. По форме и скорости плазмолиза можно судить о вязкости цитоплазмы. Плазмолиз позволяет определить величину осмотического потенциала (плазмолитический метод).
Силу, с которой вода входит в клетку, называют сосущей силой S. Она тождественна водному потенциалу клетки. Величина сосущей силы определяется осмотическим давлением клеточного сока (Р) и тургорным гидростатическим давлением (Т) в клетке. Тургорное давление равно противодавлению клеточной стенки, возникающему при ее эластичном растяжении.
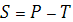
В условиях разной оводненности соотношение компонентов этого уравнения меняется.
Осмотическое давление всегда можно рассчитать по формуле:
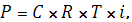
Где С – концентрация раствора в молях; Т – абсолютная температура; i– изотонический коэффициент; R – газовая постоянная.
Определение величины осмотического потенциала (сосущей силы) имеет большое значение. Величина сосущей силы позволяет судить о способности растений поглощать воду из почвы и удерживать ее. Осмотический потенциал колеблется в пределах от 5 до 200 бар. У большинства растений средней полосы данный показатель колеблется от 5 до 30 бар. Установлены определенные градиенты осмотического потенциала в пределах растения. Так, в тканях стебля отрицательный осмотический потенциал возрастает от периферии к центру и от основания к верхушке.
В проводящих элементах стебля и корня, как правило, отрицательная величина осмотического потенциала очень низка. В листьях осмотический потенциал колеблется от 10 до 18 бар.
Осмотический потенциал различен у разных жизненных форм. У древесных пород он более отрицателен, чем у кустарников, а у кустарников более отрицателен, чем у травянистой растительности. У светолюбивых растений осмотический потенциал более отрицателен, чем у тенелюбивых.
Говоря о поступлении воды в клетку, надо учитывать, что наряду с осмотическими силами в клетках существуют силы набухания. Набухание связано со способностью гидрофильных каллоидов притягивать к себе молекулы воды. Набухание может рассматриваться как особый вид диффузии, так как движение воды также идет по градиенту концентрации. Водный потенциал клеток становится более отрицательным, благодаря присутствию органических веществ, связывающих воду.
Возможным механизмом поступления воды является электроосмос, т.е. из-за разности потенциалов мембраны тонопласта, возникшего на наружной и внутренней сторонах. Движение воды может быть вызвано накоплением катионов, которое, в свою очередь, происходит под влиянием электрических потенциалов.
15. Нуклеиновые кислоты, их структура. Функциональные группы нуклеиновых кислот.
Нуклеиновые кислоты – это высокомолекулярные полимерные органические соединения, в состав которых входят основания пуриновой (аденин, гуанин) и пиримидиновой группы (цитозин, урацил, тимин), сахар (рибоза или дизоксирибоза) и фосфорная кислота. К нуклеиновым кислотам относятся рибонуклеиновая кислота (РНК), которая состоит из рибозы, оснований (аденина, гуанина, цитозина, урацила) и фосфорной кислоты (молекулярная масса 1-2 млн), и дезоксирибонуклеиновая кислота (ДНК), состоящая из дезоксирибозы, оснований (тимина, аденина, гуанина, цитозина) и фосфорной кислоты (молекулярная масса 4 – 8 млн). ДНК сосредоточена главным образом в ядре клетки, а РНК – в цитоплазме преимущественно в рибосомах.
Много нуклеиновых кислот содержится в молодых, интенсивно растущих тканях и органах (зародыши семян, пыльца, глазки клубней картофеля, кончики корней). В молодых листьях и конусах нарастания побегов нуклеиновых кислот значительно больше, чем в старых листьях и стеблях. Обычно в листьях и стеблях большинства растений нуклеиновые кислоты составляют 0,1-1% массы сухого вещества.
ДНК – высокомолекулярный полимер, состоящий из четырех нуклеотидов. Нуклеотиды ДНК включают пуриновые (аденин, гуанин) и пиримидиновые (цитозин, тимин) азотистые основания, сахар – дезоксирибозу и остатки фосфорной кислоты. Молекула ДНК представляет собой двойную спираль закрученных полимерных цепей, в каждой из которых нуклеотиды ковалентно связаны через остаток фосфорной кислоты с сахаром (десоксирибоза). Между собой две полинуклеотидные цепи взаимодействуют водородными связями, возникающими междуповернытыми внутрь спирали пуриновыми (аденин, гуанин) и пиримидиновыми (цитозин, тимин) основаниями.
При взаимодействии азотистых оснований аденин всегда взаимодействует с тимином, а гуанин с цитозином. Таким образом, возможны четыре варианта пар оснований: А-Т, Т-А, Г-У, У-Г. это явление называется комплементарностью и лежит в основе воспроизводства самой ДНК, синтеза, РНК и синтеза белков в клетке.
Молекула ДНК удваивается путем репликации каждой из двух ее цепей. Поэтому каждая двойная спираль молекулы ДНК состоит из «старой» и «новой» полинуклеотидных цепей.
Все РНК также построены из нуклеотидов четырех оснований: аденина и гуанина, цитозина и урацила.
Урацил заменяет в РНК тимин ДНК. В качестве пентозы используется рибоза. Размеры РНК разнообразны. Матричная РНК имеет от нескольких тысяч до десятков тысяч нуклеотидов и составляет до 3% суммарной РНК клетки.
42. Транспирация как физиологический процесс. Факторы, определяющие величину транспирации.
Транспирация – это физиологический процесс испарения воды растением. Количество воды, которое испаряет растение, во многом превосходит объем содержания в нем воды. Экономный расход воды составляет одну из важнейших проблем для сельскохозяйственного производства. К.А. Тимирязев называл транспирацию необходимым физиологическим злом.
Если мы будем выращивать растения в условиях высокой влажности воздуха, но транспирация будет идти со значительно меньшей интенсивностью. Однако рост растений будет одинаков или даже лучше там, где влажность воздуха выше, транспирация меньше. Вместе с тем транспирация необходима организму:
1. транспирация спасает растение от перегрева, который ему грозит на прямом солнечном свете. Температура транспирирующего листа на 5-7 градусов ниже температуры окружающего воздуха;
2. при высокой температуре разрушаются хлоропласты и угнетается процесс фотосинтеза (оптимальная температура для фотосинтеза 30-35ºС);
3. транспирация создает непрерывный ток воды из корневой системы к листьям и связывает все органы растения в единое целое;
4. с транспирационным током передвигаются растворимые минеральные и частично органические питательные вещества, при этом интенсивнее транспирация, тем быстрее идет процесс.
Транспирацию можно охарактеризовать следующими показателями: интенсивность транспирации, транспирационный коэффициент, продуктивность транспирации, относительная транспирация.
Интенсивность транспирации – это количество воды, испаряемой растением с единицы площади листьев в единицу времени. Эта величина колеблется от 0,1 до 1,5 г/дм /ч.
Транспирационный эффект – показывает, сколько единиц воды необходимо транспирировать растению, чтобы создать единицу сухого вещества. Транспирационный коэффициент зависит от вида растений, ярусности листьев, условий внешней среды. На величину транспирационного коэффициента оказывает влияние минеральное питание.
Продуктивность транспирации – это количество сухого вещества, накопленного растением при транспирации 1 литра воды. Этот показатель колеблется от 1 до 8 г.
Относительная транспирация – отношение воды, транспирируемое листом, к воде, испаряемой со свободной водной поверхности.
Регулирование устьичной транспирации – осуществляется открытием или закрытием устьиц. Их движение обусловлено различными факторами. Как мы уже отмечали, основным, обусловливающим движением устьиц является содержание воды в замыкающих клетках (изменение тургора). Различают гидропассивное и гидроактивное открытие и закрытие устьиц.
Гидропассивная реакция – это закрытие устьичных щелей, вызванное тем, что окружающие паренхимные клетки, переполненные водой, механически сдавливают замыкающие клетки. В результате сдавливания устьица не могут открыться. Гидропассивное движение обычно наблюдается после сильных поливов и может служить причиной торможения процесса фотосинтеза, а также скажется на тех процессах, которые связаны с током воды по растению. Гидроактивная реакция открывания и закрывания – это движение замыкающих клеток, вызванное применением содержания воды. Это связано с изменением концентрации осмотически активных веществ в процессе фотосинтеза, в замыкающих клетках.
В последнее время появились сведения о регулировании движения устьиц АБК (абсцизовой кислотой). АБК тормозит образование ферментов, участвующих в гидролизе крахмала, поступление калия.
Транспирация растения зависит не только от степени открытия устьиц, но и от внешних условий. Внешние условия влияют и на степень открытия устьиц, и на процесс транспирации. Зависимость транспирации от условий среды подчиняется уравнению Дальтона:
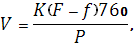
Где V – интенсивность транспирации количества воды, испарившейся с единицы поверхности;
К – коэффициент диффузии;
F – упругость паров воды, насыщающих данное пространство;
f – упругость паров воды в окружающем пространстве при температуре испаряющей поверхности;
Р – атмосферное давление в момент опыта.
Из приведенного уравнения видно, что прежде всего испарение пропорционально разности (F-f), т.е. ненасыщенности атмосферы парами воды, или дефициту влажности. Чем больше дефицит влажности воздуха, тем ниже его водный потенциал, тем быстрее идет испарение. Однако следует учесть, что данное явление смягчается в силу устьичного и внеустьичного регулирования транспирации, чем это следовало ожидать согласно формуле Дельтона.
Следующим фактором, влияющим на транспирацию, является температура. С повышением температуры возрастает.
Особое влияние на транспирацию оказывает свет. Его влияние проявляется в следующем: на свету зеленые листья поглощают определенные участки спектра, повышается температура листа и, следовательно, усиливается процесс транспирации. Действие света на транспирацию усиливается тем больше, чем выше содержание хлорофилла. На свету увеличивается проницаемость цитоплазмы.
Почва и растение образуют единую водную систему, поэтому уменьшение содержание воды в почве снижает содержание воды в растении и, как следствие, транспирацию.
Из внешних факторов существенное влияние на процесс транспирации оказывает ветер – движение атмосферы. Ветер перемещает более насыщенный парами воды слой воздуха с поверхности листа в более дальние слои. В силу этого усиливается прежде всего кутикулярная транспирация. Более сильное влияние на транспирацию оказывает ветер у тех растений, которые имеют тонкую или поврежденную кутикулу.
Интенсивность транспирации зависит и от ряда внутренних факторов, и прежде всего от содержания воды в листьях. Всякое уменьшение содержания воды в листьях уменьшает транспирацию. Транспирация зависит и от концентрации клеточного сока. Чем концентрированнее клеточный сок, тем слабее транспирация. Интенсивность транспирации зависит от эластичности клеточных стенок.
С увеличением возраста растений интенсивность транспирации снижается. На процесс транспирации влияет смена дня и ночи. В ночной период суток транспирация резко сокращается из-за снижения температуры, повышения влажности воздуха, отсутствия света. Исследования показывают, что ночная транспирация составляет 3-5% от дневной.
Основной ход транспирации зависит от соотношения метеорологических факторов. Максимум транспирации наблюдается в середине дня.
Транспирация зависит от величины листовой поверхности, чем она (листовая поверхность) больше, тем сильнее процесс транспирации.
Контрольная работа №2
10. Физиологические нарушения при недостатке отдельных элементов питания.
Корневое питание – это процесс поглощения и усвоения ими химических элементов из окружающей среды. Чтобы выяснить какие элементы питания необходимы растениям, их выращивают в условиях вегетационного опыта, на специально составленных питательных смесях и наблюдают за ростом и развитием.
Различают следующие методы искусственных культур: водных, песчаных и почвенных. Метод водных культур впервые был разработан немецким физиологом Н. Кнопом и Ю. Саксом в 70-х годах 19 столетия.
В состав питательных смесей входят такие необходимые для растений макроэлементы, как азот, фосфор, калий, кальций, магний и сера. Исключая из нормальной смеси некоторые элементы, установили, что нормальное развитие растений возможно только при наличии в питательном растворе следующих элементов: неметаллов –азота, фосфора, серы и бора; металлов – калия, магния, кальция, железа, меди, цинка, марганца, кобальта, молибдена.
Для нормального развития растений необходимы также микро- и ультрамикроэлемнты и нельзя заменить один другим.
Буферные свойства питательных смесей определяют по наличию основных и кислых групп. Чем выше буферность смеси, тем меньше изменяется ее рН при неэквивалентном поглощении ионов корнями растений. В растительном организме важную роль в качестве буферов играют органические кислоты (винная, яблочная, лимонная, щавелевая) и их соли.
Анализ почвенного раствора или водной вытяжки дает определенное представление о наличии доступных элементов питания для растений на данной почве.
Однако они являются приближенными, поскольку корни различных растений обладают растворяющей способностью.
Индикатором плодородия почвы служит само растение. Во многих случаях при недостатке элементов минерального питания на растениях появляются характерные симптомы. В ряде случаев эти признаки голодания могут помочь установить функции данного элемента, а также необходимость его дополнительного внесения в почву. Симптомы голодания зависят также от способности передвижения данного элемента питания по растению. Соединение таких элементов, как N, P, S, K, Mgи некоторые другие, легко передвигаются, и растение способно к их вторичному использованию. В случае недостатка какого-либо из этих элементов они передвигаются из более старых к более молодым органам. В силу этого симптома голодания проявляются в первую очередь на старых листьях. Вместе с тем такие элементы и их соединения, как Са, Fe, B, Zn плохо передвигаются по растению. Эти элементы не способны ко вторичному использованию (реутолизации). Симптомы голодания проявляются в отношении этих элементов в первую очередь на самых молодых листьях и органах.
Внешние признаки голодания в отношении отдельных элементов проявляются следующим образом.
Азот.
Ярким признаком недостатка азота является пожелтение листьев, связанное с недостатком хлорофилла. У некоторых растений наблюдается усиленный синтез антоциана и в связи с этим появление красноватого оттенка черешков и жилок листьев. Листья преждевременно опадают.
Фосфор.
При фосфорном голодании на листьях, незрелых плодах появляются мертвые некротические пятна. Окраска листьев становится голубовато-зеленая или темно-зеленая. Стебли имеют слаборазвитую проводящую систему.
Калий.
При недостатке калия на листьях появляются желтые пятна. Некротические участки, края и кончики листьев часто скручиваются.
Магний.
Поскольку магний входит в состав хлорофилла, то первым признаком голодания является интенсивное пожелтение паренхимы листа (особенно между жилками).
Сера.
Признаки серного голодания очень близки к тем, которые наблюдаются при недостатке азота. Листья желтеют, появляется антоциановая окраска. Однако эти признаки прежде всего на молодых листьях.
Кальций.
При недостатке кальция повреждаются и отмирают меристематические зоны стебля, корня и листьев. В свою очередь это тормозит процессы роста.
Железо.
Недостаток железа вызывает хлороз листьев, в первую очередь молодых, хлороз проявляется между жилками листа.
Марганец.
При недостатке марганца на листьях появляются желтые и некротические пятна. Особенно чувствительны к недостатку марганца хлоропласты.
Медь.
При недостатке меди белеют и отмирают кончики листьев, затем хлорофилл разрушается по краям листовой пластинки. Листья теряют тургор, растение завядает, листья и плоды плодовых деревьев покрываются бурыми пятнами.
Цинк.
Недостаток цинка приводит к уменьшению размера листьев и к изменению их формы, междоузлия укорачиваются, на листьях проявляется хлороз.
Бор.
Первым симптомом при недостатке этого элемента является остановка роста побегов и корней, листья становятся толще, скручиваются, цветки не образуются, клетки плохо дифференцируются.
Молибден.
При дефиците молибдена листья по краям приобретают серую, а затем коричневую окраску, теряют тургор, а затем отмирают (остаются живыми только жилки).
69. Условия и причины вымерзания растений. Морозоустойчивость.
Морозоустойчивость – способность растений переносить температуру ниже 0ºС. Разные растения переносят зимние условия, находясь в различном состоянии. У однолетних растений зимуют семена, нечувствительные к морозам, у многолетних – защищенные слоем земли и снега клубни, луковицы и корневища. У озимых растений и древесных пород ткани под действием отрицательных температур могут замерзнуть и даже промерзнуть насквозь, однако растения не погибают. Способность этих растений перезимовывать обусловливается их достаточно высокой морозоустойчивостью.
Поврежденные морозом растения имеют вид как бы обваренных, они утрачивают тургор, листья их буреют и засыхают. При оттаивании клубней картофеля или корнеплодов сахарной свеклы вода легко вытекает из тканей. Такое явление длительное время объясняли разрывом клеточных стенок под влиянием льда, образующегося в тканях растений. Однако установлено, что лед образуется главным образом в межклетниках и клеточные стенки остаются неповрежденными. Гибель растений под влиянием морозов обусловливается изменениями, происходящими в протопласте, его коагуляцией. Физико-химические преобразования в протопласте происходят вследствие оттягивания воды образующимися в межклетниках в межклетниках кристаллами. Кроме того, протопласт подвергается сжатию со стороны растущих в межклетниках кристаллов. В результате наступает необратимая денатурация коллоидов протопласта клеток и отмирание тканей. Если льда образуется немного, то после оттаивания растение сможет остаться живым. Так, в листьях капусты, выдержанных при температуре минус 5-6ºС, образуется некоторое количество льда, воздух из межклетниках вытесняется, и листья становятся прозрачными. Однако образование льда в межклетниках неопасно, и после оттаивания листья возвращаются в нормальное состояние.
Каждая клетка имеет свою границу обезвоживания и сжатия. Переход этих границ, а не только снижение температуры – причина гибели клеток. Такое явление следует рассматривать не как непосредственное влияние холода на протопласт, а как действие, вызывающие обезвоживание протопласта вследствие вымораживания воды. Убедительным доказательством этого служит состояние переохлаждения (без образования льда), которое растения переносят без вреда; при тех же температурах, но с образованием льда в тканях растения гибнут. Не все растения одинаково реагируют на образование льда в тканях. Например, клубни картофеля, георгина погибают сразу; капуста, лук переносят умеренное промораживание; растения северных широт, озимые злаки (рожь, пшеница) выдерживают понижение температур до – 15 - 20ºС. Еще более выносливы зимующие почки лиственных и иглы хвойных деревьев. Нечувствительность к морозам достигается физико-химическими изменениями в клетках. В зимующих листьях и других частях растений накапливается много сахара, а крахмала в них почти нет. Сахар защищает белковые соединения от коагуляции при вымораживании, и поэтому его можно назвать защитным веществом. При достаточном количестве сахара в клетках повышаются водоудерживающие силы коллоидов протопласта, увеличивается количество прочносвязанной и уменьшается содержание свободной воды. Связанная с коллоидами вода при действии низких температур не превращается в лед. У ряда древесных пород в результате преобразования углеводов в коровой паренхиме накапливаются жиры и липоиды, которые не замерзают и проявляют защитное действие в зимний период. Кроме того, белковые вещества, частично гидролизуясь, переходят из менее устойчивой в более устойчивую форму азотосодержащих веществ – аминокислоты, которые при замерзании в меньшей мере подвергаются денатурации.
Современное учение о морозоустойчивости растений основано на том, что это свойство формируется в процессе онтогенеза под влиянием условий внешней среды в соответствии с генотипом, связано с явлением покоя и не является постоянным. У вегетирующих растений легко вымерзают растущие и закончившие рост растения. Выносливость растений к низким температурам в этот период незначительная. Степень морозоустойчивости клеток во много зависит от состояния плазмолеммы, обмена веществ, образования сложных органических соединений и массы цитоплазмы. В клетках внутренние слои цитоплазмы повреждаются раньше, чем плазмалемма, которая способна к быстрому новообразованию за счет молекул остальной массы цитоплазмы. Недостаточно закаленные клетки не могут быстро восстанавливать плазмалемму. На степень морозоустойчивости растений большое влияние оказывают сахара, регуляторы роста и другие вещества, образующиеся в клетках.
И. И. Туманов разработал теорию закаливания растений, повышающего их устойчивость к действию низких температур. Сущность ее заключается в том, что у растений под влиянием низких положительных температур накапливаются сахара и другие соединения – первая фаза закаливания. Дальнейшее повышение морозоустойчивости происходит уже при отрицательных температурах, но не повреждающих клетки, - вторая фаза закаливания. Она идет сразу же после первой при температуре немного ниже 0ºС. В этой фазе наблюдается частичная потеря воды клетками. Под действием сахаров, накопившихся в клетках, изменяются биоколлоиды и возрастает относительное количество коллоидно-связанной воды. Такие изменения придают биоколлоидам устойчивость к низким температурам.
У озимых злаков, не имеющих периода покоя в первой фазе закаливания при относительно низкой температуре (до 10º) и солнечной погоде накапливаются углеводы. Если осенью погода ясная и прохладная, озимые хлеба хорошо перезимовывают, так как первая фаза закаливания у них происходит в благоприятных условиях. При закаливании озимых растений свет необходим не только для накопления в клетках защитных веществ в процессе фотосинтеза, но и для поддержания ультраструктуры протопласта и ростовых процессов. Растения озимой пшеницы можно закаливать и в темноте при 2ºС, если их корни или узлы кущения погрузить в раствор сахарозы. Такие растения выдерживают морозы до 20ºС.
Обнаружено, что в период закаливания растений высокоморозоустойчивого сорта озимой пшеницы при температуре, близкой к 0ºС, количество сахаров в хлоропластах листьев увеличивалось в 2,5 раза. Вместе с тем в пересчете на белок в хлоропластах их накапливается меньше, чем в целом листе, так как значительная часть сахаров локализуется в клеточном соке, что, по-видимому, предотвращает образование льда в вакуолях. Однако наблюдения за динамикой накопления сахаров показали, что во время закаливания содержание их в хлоропластах в пересчете на белок возрастает в 3 раза, а в листьях – лишь в 1,5 раза. В хлоропластах содержатся те же формы сахаров, что и в листьях. Повышение содержания сахаров в хлоропластах в значительной мере зависит от соотношения ряда физиологических процессов, протекающих при температурах, близких к 0ºС. Так, с понижением температуры при закаливании растений интенсивность дыхания снижается сильнее, чем фотосинтез, в результате чего наблюдается задержка ростовых процессов. Повышение содержания сахаров в хлоропластах коррелирует с морозоустойчивостью растений. Следовательно, сахара оказывают стабилизирующие действие на клеточные структуры.
Установлено, что содержание не замерзающей (связанной) воды в тканях зимостойкой пшеницы почти в 3 раза выше по сравнению с незимостойкой. В районах с неустойчивой весной и частым возвращением весенних холодов новые побеги у пшеницы образуются медленно и неодновременно из почек, находящихся в состоянии покоя в узлах кущения. Наоборот, у злаковых, возделываемых в районах с устойчивым и большим снеговым покровом, наблюдается дружное прорастание всех спящих почек узла кущения, что свидетельствует о приспособленности к прохождению озимыми растениями так называемого зимнего покоя. При этом глубина залегания узла кущения и мощности его развития зависят от качества семян, способа посева, обработки почвы и оказывают большое влияние на перезимовку озимых. Обычно, когда узел кущения находится близко от поверхности (1,5 см), такие растения менее зимостойки, чем растения с более глубоким залеганием узла кущения (3-4 см).
Зная природу морозоустойчивости, можно значительно повысить устойчивость растений к низким температурам.
82. диагностика засухоустойчивости. Физиологическое обоснование селекции на засухоустойчивости
.
Способами диагностики засухоустойчивости могут быть прямые методы оценки и косвенные. Прямые методы: полевой метод, метод завядания в сосуде, метод засушника, суховейная камера, лабораторный метод крахмальной пробы.
Повышение засухоустойчивости растений. Внесение удобрений, особенно фосфорных, улучшает рост и развитие и способствует экономному расходованию воды. П А. Генкель предлагал проводить предпосевное закаливание семян. Адаптация к обезвоживанию происходит в семенах, которые перед посевом после однократного намачивания вновь подсушиваются. Для растений, выращенных из таких семян, характерны морфологические признаки ксероморфности. Одним из самых эффективных мероприятий по борьбе с засухой является орошение. Основная задача при орошении – это добиться наибольшего эффекта с наименьшим количеством воды.
Влияние засухи на растение. Если в почве имеется недостаточное количество доступной для растений воды, то отрицательное влияние перегрева становится особенно сильным. Перегрев вызывает поражение растения, называемое запалом. Запал обнаруживается через некоторое время в виде различных окрашенных некротических пятен на листьях. На пшенице появляются желтые пятна, на овсе – красные, у большинства растений – коричневые.
Встречается и другой вид повреждений от атмосферной засухи – захват. Он наблюдается реже, чем запал, и появляется в том случае, когда при сравнительно не очень высоких температурах наблюдается сильный ветер и большая сухость воздуха. При этом листья просто высыхают, сохраняя зеленую окраску. Недостаток воды в ткани растений может возникнуть в жарку солнечную погоду к середине дня, при этом увеличивается сосущая сила листьев, что активирует поступление воды из почвы. Растения регулируют уровень водного дефицита открытием или закрытием устьиц. В этот период происходит временное завядание листьев. Обычно в вечерние и утренние часы это явление устраняется.
Отсутствие в почве доступной для растений воды приводит к глубокому завяданию. Это завядание чаще всего приводит к гибели растений. Характерным признаком устойчивости водного дефицита является сохранение его в тканях утром, прекращение выделения пасаки из срезанного стебля. Действие засухи приводит в первую очередь к уменьшению в клетках свободной воды, что нарушает гидратные оболочки белков цитоплазмы и сказываются на функциях белков – ферментов.
При длительном завядании снижается активность ферментов синтеза и активируются гидролитические процессы, что приводит к возрастанию содержания в клетках низко молекулярных белков. В результате гидролиза полисахаридов в тканях накапливаются растворимые углеводы. В листьях снижается количество РНК, наблюдается распад полирибосомных комплексов. Возрастает концентрация вакуолярного сока, облегчается выход ионов из клеток.
Происходит снижение скорости фотосинтеза из-за недостатка углекислого газа, нарушение синтеза хлорофилла и АТФ, изменение в течение фотохимических реакций и задержки оттока ассимилянтов из листьев. При обезвоживании растений не приспособленных к засухе, значительно усиливается интенсивность дыхания, а у засухоустойчивых растений такое явление не наблюдается.
В условиях водного дефицита тормозятся клеточное деление и особенно растяжение, что приводит к формированию мелких клеток. В следствии этого задерживается рост листьев, стебля растения в целом. При водном дефиците происходит гидролиз полимеров, в том числе и белка. Распад белков идет с образованием аммиака, который может оказывать отравляющее действие на клетки.
Приспособления растений к засухе. Известно, что растения неодинаково реагируют на перегрев и обезвоживание в разный период онтогенеза. У каждого вида в онтогенезе имеется такой период, когда недостаток воды резко сказывается на течении всех физиологических процессов, этот период называется критическим. Из этого, однако, не следует, что остальные периоды своего развития растение не нуждается в воде и не страдает от ее недостатка.
Установлено, что вязкость цитоплазмы, ее эластичность в критический период резко падает, что и является одним из условий высокой чувствительности растений к перегреву и обезвоживанию в этот период развития. В критический период происходят интенсивные ростовые процессы и образование новых органов – цветков.
53. Физиология формирования плодов.
Созревание клубнеплодов и корнеплодов имеет особенность. Клубнеобразование у картофеля начинается в фазу бутонизации. Во время созревания клубней в основном наблюдается синтез крахмала и белка. В начальный период клубнеобразования интенсивность синтеза крахмала незначительна, много свободных сахаров, в соотношении крахмал/сахар доходит до 10, тогда как во время интенсивного клубнеобразования в начале отмирания листьев это отношение достигает 60 и более. Установлено, что максимальное содержание крахмала наступает раньше, чем заканчивается период вегетации. Дальнейшее нахождение клубней на растении не желательно, так как приводит к снижению содержания крахмала из-за расходования его на дыхание. У поздних сортов картофеля процессы накопления сухого вещества и крахмала в клубнях продолжается до конца вегетации. Уборку таких сортов необходимо проводить в наиболее поздние сроки. Максимальное содержание крахмала в клубнях отмечено, когда примерно 80% листьев засыхает. Поэтому сроки уборки картофеля должны определяться не полным засыханием ботвы, а максимальным содержанием сухого вещества и крахмала. В клубнях картофеля при их созревании меняется соотношение активности a- и b-амилазы и фосфорилазы. По мере созревания клубней происходит снижение активности амилазы, одновременно с синтезом крахмала в клубнях идет накопление белков, но их соотношение зависят от сорта. Клубнекартофель имеет кислую реакцию (рН сока 5,6-6,2), что связано содержанием органических кислот, среди которых преобладает лимонная. В клубнях картофеля содержится и много витаминов, каротин.
В корнеплодах сахарной свеклы основной составной частью является сахароза – до 20%, на воду приходится до 75%, в сухом веществе содержится так же мальтоза, рафиноза, глюкоза и фруктоза.
Общее количество азотистых веществ в корнеплодах составляет 0,15-0,25%, это в основном свободные аминокислоты и белки.
Изучение ферментативной активности сахарной свеклы показало, что на ранних этапах развития активна инвертаза, но по мере роста свеклы содержание этого фермента падает к 45 дню уже не обнаруживается, а становится активным ферментом сахорозосинтеза.
Процесс созревания сочных плодов, которые имеют мясистую ткань, сопровождается изменением физических и химических свойств ткани, конституции мякоти, вкуса, аромата и т.д.
К состоянию полной спелости в яблоках, плодах цитрусовых, томата возрастает количество сахаров и значительно уменьшается количество органических кислот. Состав сахаров не одинаков. Так, у томатов содержание сахара идет за счет моноз, а содержание сахарозы постоянно. Твердость плода зависит от количества пектиновых и дубильных веществ в нем. В зеленых плодах томата на нерастворимую фракцию пектиновых веществ приходится 2/3 общего его количества, в зрелых же лишь 4%, а остальная часть пектиновых веществ превращается в другие соединения или переходят в растворимые фракции. Можно искусственно ускорять дозревание плодов. Для этого плоды можно обрабатывать этиленом в течении 2-3 суток в плотно закрытых камерах из расчета 1 объем газа на 1000 объема воздуха при температуре 18-20ºС и относительной влажности воздуха 70-85%.
При вызревании плодов газообмен смещается к анаэробному дыханию – в них возрастает количество спирта, этилена.
|
