Концепция материальности сознания.
1. Нейрон – структурно – функциональная единица нервной системы..2
2. Механизм проведения нервного импульса…………………………... 3
3. Материальная основа высшей нервной деятельности………………. 8
4. Физиология восприятия……………………………………………….12
5. Физиологические механизмы памяти. ……………………………….14
6. Литература……………………………………………………………...15
1. Нейрон – структурно – функциональная единица нервной системы
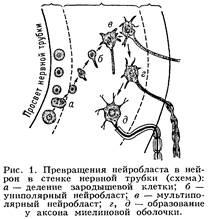 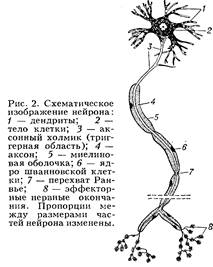 Нейрон - (от греч. neuron — нерв), неврон, нервная клетка, основная функциональная и структурная единица нервной системы, принимает сигналы, поступающие от рецепторов и др. Нейрон перерабатывает их и в форме нервных импульсов передаёт к эффекторным нервным окончаниям, контролирующим деятельность исполнительных органов (мышцы, клетки железы или др.). Образование нейрона происходит при эмбриональном развитии нервной системы: на стадии нервной трубки развиваются нейробласты, которые затем дифференцируются в нейрон (рис. 1). В процессе дифференцировки формируются специализированные части нейрона (рис. 2), которые обеспечивают выполнение его функций. Для восприятия информации развились ветвящиеся отростки — дендриты, обладающие избирательной чувствительностью к определённым сигналам и имеющие на поверхности т. н. рецепторную мембрану. Процессы местного возбуждения и торможения с рецепторной мембраны, суммируясь, воздействуют на триггерную (пусковую) область — наиболее возбудимый участок поверхностной мембраны нейрона, служащий местом возникновения (генерации) распространяющихся биоэлектрических потенциалов. Для их передачи служит длинный отросток — аксон, или осевой цилиндр, покрытый электровозбудимой проводящей мембраной. Достигнув концевых участков аксона, импульс нервный возбуждает секреторную мембрану, вследствие чего из нервных окончаний секретируется физиологически активное вещество — медиатор или нейрогормон. Кроме структур, связанных с выполнением специфических функций, каждый нейрон, подобно др. живым клеткам, имеет ядро, которое вместе с околоядерной цитоплазмой образует тело клетки, или перикарион. Здесь происходит синтез макромолекул, часть которых транспортируется по аксоплазме (цитоплазме аксона) к нервным окончаниям. Структура, размеры и форма нейрона сильно варьируют. Сложное строение имеют нейроны коры больших полушарий головного мозга, мозжечка, некоторых др. отделов центр, нервной системы. Для мозга позвоночных характерны мультиполярные нейроны. Нейрон - (от греч. neuron — нерв), неврон, нервная клетка, основная функциональная и структурная единица нервной системы, принимает сигналы, поступающие от рецепторов и др. Нейрон перерабатывает их и в форме нервных импульсов передаёт к эффекторным нервным окончаниям, контролирующим деятельность исполнительных органов (мышцы, клетки железы или др.). Образование нейрона происходит при эмбриональном развитии нервной системы: на стадии нервной трубки развиваются нейробласты, которые затем дифференцируются в нейрон (рис. 1). В процессе дифференцировки формируются специализированные части нейрона (рис. 2), которые обеспечивают выполнение его функций. Для восприятия информации развились ветвящиеся отростки — дендриты, обладающие избирательной чувствительностью к определённым сигналам и имеющие на поверхности т. н. рецепторную мембрану. Процессы местного возбуждения и торможения с рецепторной мембраны, суммируясь, воздействуют на триггерную (пусковую) область — наиболее возбудимый участок поверхностной мембраны нейрона, служащий местом возникновения (генерации) распространяющихся биоэлектрических потенциалов. Для их передачи служит длинный отросток — аксон, или осевой цилиндр, покрытый электровозбудимой проводящей мембраной. Достигнув концевых участков аксона, импульс нервный возбуждает секреторную мембрану, вследствие чего из нервных окончаний секретируется физиологически активное вещество — медиатор или нейрогормон. Кроме структур, связанных с выполнением специфических функций, каждый нейрон, подобно др. живым клеткам, имеет ядро, которое вместе с околоядерной цитоплазмой образует тело клетки, или перикарион. Здесь происходит синтез макромолекул, часть которых транспортируется по аксоплазме (цитоплазме аксона) к нервным окончаниям. Структура, размеры и форма нейрона сильно варьируют. Сложное строение имеют нейроны коры больших полушарий головного мозга, мозжечка, некоторых др. отделов центр, нервной системы. Для мозга позвоночных характерны мультиполярные нейроны.
Реклама

В таком нейроне от клеточного тела отходят несколько дендритов и аксон, начальный участок которого служит триггерной областью. На клеточном теле мультиполярного нейрона и его дендритах имеются многочисленые нервные окончания, образованные отростками др. В ганглиях беспозвоночных чаще встречаются униполярные нейроны в которых клеточное тело несёт лишь торфич. функцию и имеет единственный, т. н. вставочный, отросток, соединяющий его с аксоном. У такого нейрона, по-видимому, может не быть настоящих дендритов и рецепцию синаптических сигналов осуществляют специализированные участки на поверхности аксона. Нейрон с двумя отростками назаваемые биполярными; такими чаще всего бывают периферично чувствительные нейроны имеющие один направленный наружу дендрит и один аксон. В зависимости от места, которое нейрон занимает в рефлекторной дуге, различают чувствительные (афферентные, сенсорные, или рецепторные) нейроны получающие информацию из внешней среды или от рецепторных клеток; вставочные нейроны (или интернейроны), которые связывают один нейрон с другим; эффекторные (или эфферентные) нейроны посылающие свои импульсы к исполнительным органам (напр., мотонеироны, иннервирующие мышцы). Нейрон классифицируют также в зависимости от их химической специфичности, т. е. от природы физиологически активного вещества, которое выделяется нервными окончаниями данного нейрона (например, холинергический нейрон секретирует ацетилхолин, пептидер-гический — то или иное вещество пептидной природы и т. д.). Разнообразие и сложность функций нервной системы зависят от числа составляющих её нейронов (около 102
у коловратки и более чем 1010
у человека).
2
. Механизм проведения нервного импульса.
Для перехода сигнала из одной клетки в другую существуют специальные межклеточные соединения - синапсы. Хотя контактирующие участки нейронов тесно прилегают друг к другу, между ними, как правило, остается разделяющая их синаптическая щель, ширина которой - всего несколько десятков нанометров. Непременное условие успешной деятельности нейронов - их "территориальный суверенитет", а содружественную работу обеспечивают синапсы. Электрический импульс не может преодолеть без существенных потерь энергии даже такую короткую межклеточную дистанцию, поэтому в большинстве случаев требуется преобразование информации из одной формы в другую, например из электрической в химическую, а затем - вновь в электрическую. Как же это происходит?
Реклама

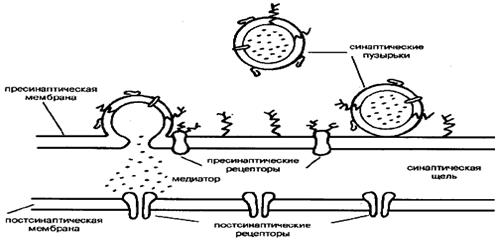 Рис.3. Рис.3.
Схема синапса. Вверху - участок нервного окончания, ограниченный пресинаптической мембраной, в которую встроены пресинаптические рецепторы; синаптические пузырьки внутри нервного окончания наполнены медиатором и находятся в разной степени готовности к его освобождению; мембраны пузырьков и пресинаптическая мембрана содержат пресинаптические белки. Внизу - участок управляемой клетки, в постсинаптическую мембрану которой встроены постсинаптические рецепторы
Синаптическая передача осуществляется последовательностью двух пространственно разобщенных процессов: пресинаптического по одну сторону синаптической щели и постсинаптического по другую (рис.3). Окончания отростков управляющего нейрона, повинуясь пришедшим в них электрическим сигналам, высвобождают в пространство синаптической щели специальное вещество-посредник (медиатор). Молекулы медиатора достаточно быстро диффундируют через синаптическую щель и возбуждают в управляемой клетке (другом нейроне, мышечном волокне, некоторых клетках внутренних органов) ответный электрический сигнал. В роли медиатора выступает около десятка различных низкомолекулярных веществ: ацетилхолин (эфир аминоспирта холина и уксусной кислоты), глутамат (анион глутаминовой кислоты), ГАМК (гамма-аминомасляная кислота), серотонин (производное аминокислоты триптофана), аденозин и др. Они предварительно синтезируются пресинаптическим нейроном из доступного и относительно дешевого сырья и хранятся вплоть до использования в синаптических пузырьках, где, словно в контейнерах, заключены одинаковые порции медиатора (по несколько тысяч молекул в одном пузырьке).
В настоящее время хорошо известны основные этапы процесса освобождения медиатора. Нервный импульс, т. е. электрический сигнал, возникает в нейроне, распространяется по его отросткам и достигает нервных окончаний. Его преобразование в химическую форму начинается с открывания в пресинаптической мембране кальциевых ионных каналов, состояние которых управляется электрическим полем мембраны. Теперь роль носителей сигнала берут на себя ионы кальция. Они входят через открывшиеся каналы внутрь нервного окончания. Резко возросшая на короткое время примембранная концентрация ионов кальция активизирует молекулярную машину освобождения медиатора: синаптические пузырьки направляются к местам их последующего слияния с наружной мембраной и, наконец, выбрасывают свое содержимое в пространство синаптической щели.
Синапсы выполняют функцию усилителей нервных сигналов на пути их следования. Эффект достигается тем, что один относительно маломощный электрический импульс освобождает сотни тысяч молекул медиатора, заключенных до того во многих синаптических пузырьках. Залп молекул медиатора синхронно действует на небольшой участок управляемого нейрона, где сосредоточены постсинаптические рецепторы - специализированные белки, которые преобразуют сигнал теперь уже из химической формы в электрическую.
Синапсы - удобный объект регулирования потоков информации. Уровень усиления сигнала при его передаче через синапс можно легко увеличить или уменьшить, изменяя количество освобождаемого медиатора, вплоть до полного запрета на передачу информации. Теоретически это можно осуществить путем направленного воздействия на любой из этапов высвобождения медиатора.
Именно на выяснение механизмов управления синаптической передачей и были устремлены усилия исследователей в лаборатории биофизики Института эволюционной физиологии и биохимии им. И. М. Сеченова РАН (Санкт-Петербург). Их эффективному проведению способствовала поддержка Международного научного фонда, позволив, в частности, оперативно приобрести современные приборы и разнообразные химические реактивы.
В качестве подходящего объекта исследования выбрали мышцу лягушки, изолированную вместе с управляющим нервом, такой препарат обладает главными достоинствами: однотипностью синапсов, удобством проведения разнообразных и достаточно тонких экспериментов, способностью изолированных тканей длительное время сохранять свою жизнеспособность. Поскольку основные свойства синапсов в разных тканях весьма сходны, полученные сведения можно использовать для анализа синаптической передачи в мозгу.
Свойства нервно-мышечных синапсов диктовали и методы регистрации. Одиночный акт передачи сигнала через синапс длится всего несколько тысячных долей секунды (миллисекунд), поэтому для его регистрации возможны только электрические измерения. С этой целью изолированный препарат помещают в ванночку с солевым раствором, с той же концентрацией ионов, как в плазме крови лягушки. Нерв укладывают на раздражающие электроды, а в мышечное волокно, в непосредственной близости с синапсом, вводят внутриклеточные микроэлектроды. Последние представляют собой тонкие стеклянные трубки, диаметр вытянутого кончика которых не превышает одного-двух микрометров. Микроэлектрод, заполненный концентрированным раствором хлористого калия, может служить для измерения разности потенциалов между внутренностью мышечного волокна и его поверхностью. Специальная система усилителей биоэлектрических сигналов позволяет одновременно регистрировать и быстрые изменения электрического тока, вызванные действием медиатора, и удерживать потенциал на мембране мышечного волокна на заданном уровне. Электрические синаптические ответы затем можно вывести на экран осциллографа (рис.4), записать на магнитную ленту либо после преобразования в цифровую форму проанализировать на компьютере. Такая система позволяет экспериментатору оценивать изменения синаптической передачи в ходе опыта.
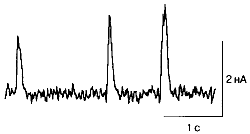 Спонтанные постсинаптические токи, генерируемые мышечным волокном. Каждый сигнал - ответ на действие порции медиатора, освободившегося из синаптических пузырьков Спонтанные постсинаптические токи, генерируемые мышечным волокном. Каждый сигнал - ответ на действие порции медиатора, освободившегося из синаптических пузырьков
Из работ наших предшественников уже было известно, что окончания двигательного нерва, управляющего сокращениями мышцы, чувствительны к некоторым химическим агентам, способным изменять количество освобождаемого медиатора. На поверхности пресинаптической мембраны нервных окончаний тоже имеются рецепторы, белковые "датчики", которые опознают действующий на них агент и запускают процесс регулирования. Нам предстояло выяснить, как это происходит, каким образом химический сигнал, воспринятый нервным окончанием, влияет на освобождения медиатора. Для этого надо было проследить всю цепочку молекулярных процессов - от воздействия химического агента на рецептор до конечной мишени его влияния. Прежде всего, какие же химические агенты модулируют освобождение медиатора? Наибольший интерес для нас представляло пресинаптическое действие самих медиаторов. Биологический смысл использования одних медиаторов для регуляции освобождения других очевиден. Такой механизм обеспечивает вмешательство третьего нейрона в "разговор" первых двух: выделяемый им медиатор действует на нервное окончание управляющего нейрона и регулирует освобождение медиатора, предназначенного для передачи сигнала управляемому нейрону. Здесь, как правило, используются медиаторы разной химической природы: один для передачи по основному пути, а другой - для ее модуляции. Но медиатор может и сам управлять своим освобождением. Если из нервного окончания освобождается избыточное количество медиатора, это в конце концов приводит к нерациональным энергетическим затратам на синтез необходимых веществ и на восстановление режима готовности к новому рабочему акту. Наконец, перевозбуждение может стать губительным для управляемой клетки. Значит, необходима саморегулирующаяся система, основанная на принципе отрицательной обратной связи. Действительно, медиатор в нервно-мышечных синапсах, ацетилхолин, будучи выброшен в синаптическую щель, не только выполняет свою прямую обязанность - передать информацию (например, двигательный приказ мышечному волокну, действуя на его постсинаптические рецепторы), но и способен регулировать собственное освобождение, действуя через пресинаптические рецепторы нервных окончаний.
Принципиальная схема регуляции должна выглядеть следующим образом: медиатор действует на пресинаптический рецептор, изменения состояния рецептора передаются через некие промежуточные стадии на системы, способные влиять на выброс медиатора, - и в результате снижается вероятность этого события.
Нужно было ответить на целый ряд вопросов, логически связанных друг с другом. Прежде всего определить, какой тип пресинаптического рецептора вовлечен в процесс саморегулирования. Оказывается, природа и здесь подошла к решению проблемы рационально. Для передачи двигательных сигналов используется один тип рецептора, рассчитанный на быстрое их преобразование, а для регулирования - рецепторы другого типа, работающие гораздо медленнее, но зато обеспечивающие высокий коэффициент усиления сигнала. Несмотря на то, что оба типа рецептора реагируют на один и тот же медиатор (ацетилхолин), в эксперименте, используя различия в их чувствительности к синтезированным химическим аналогам ацетилхолина, можно избирательно действовать на постсинаптические (пусковые) или на пресинаптические (регулирующие) рецепторы (рис.5).
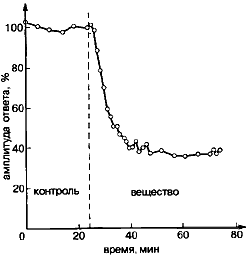
Эксперимент на мышце лягушки, демонстрирующей угнетения освобождения ацетилхолина из двигательных нервных окончаний в результате действия одного из его аналогов на пресинаптические рецепторы. Видно, что постсинаптические токи, вызванные периодическим раздражением нерва, при действии вещества значительно уменьшают свою амплитуду по сравнению с контрольным периодом
Следующая проблема: где расположена клеточная система, способная непосредственно модулировать освобождение медиатора, в пресинаптической мембране или внутри нервного окончания? Пресинаптическая мембрана, действительно, содержит элементы, от деятельности которых зависит реализация сигнала. Это пути, используемые ионами калия и кальция для прохождения через клеточную мембрану, - соответственно калиевые и кальциевые ионные каналы. Может быть, активация пресинаптических рецепторов изменяет потоки этих ионов и тем самым модулирует освобождение медиатора?
Проведенные нами эксперименты отвергли это предположение. Оказалось, что активация пресинаптических рецепторов в равной мере подавляет освобождение медиатора, как вызванное раздражением двигательного нерва (оно, действительно, зависит от состояния кальциевых и калиевых каналов), так и спонтанное освобождение, на которое эти факторы существенно не влияют. Если в эксперименте кальциевые и калиевые каналы заблокированы, то регулирующее действие медиатора сохраняется. Более того, возможность регулирования не зависит от концентрации кальция внутри нервного окончания.
Эти результаты привели нас к важному выводу: конечную мишень регулирования надо искать внутри нервного окончания, в структурах, непосредственно ответственных за освобождение медиатора. Но прежде необходимо понять: как активированный пресинаптический рецептор передает эстафету дальше, внутрь клетки, каков механизм усиления этого сигнала? Типичный для большинства живых клеток способ решения подобной задачи состоит в сопряжении рецептора, расположенного на поверхности мембраны клетки, с универсальным передаточным устройством, внутриклеточным специализированным белком, который назвали Г-белком. Этот белок составлен, как правило, из нескольких субъединиц, образующих единый комплекс. Активация рецептора приводит к его обратимому разделению, и одна из составных частей обретает относительную свободу передвижения. Это позволяет ей взаимодействовать с целым рядом клеточных устройств: активировать или подавлять работу транспортных систем клетки, регулировать деятельность внутриклеточных ферментов и т. п. Таким образом, регулирующий сигнал в результате многократного усиления в конце концов изменяет определенную клеточную функцию.
Используется ли этот типичный механизм в исследуемом нами случае? Известно, что Г-белки описанного выше типа чувствительны к некоторым токсинам, продуцируемым микробами, например коклюшной палочкой или холерным вибрионом. Наши эксперименты дали отрицательный ответ: ни коклюшный, ни холерный токсины не влияли на регуляцию освобождения медиатора. В то же время пресинаптическое действие медиатора сильно зависело от температуры: при снижении температуры раствора на 10 градусов оно исчезало, хотя сам медиатор высвобождался. Это означает, что в процесс передачи регулирующего сигнала вовлечены реакции, требующие поступления энергии извне, например ферментативные. Был получен также ряд косвенных свидетельств в пользу того, что природа нашла здесь не совсем обычный способ регуляции. В частности, можно предполагать, что белки, подобные Г-белкам, располагаются непосредственно на поверхности мембраны синаптических пузырьков.
Дальнейшее выяснение конкретного объекта регулирования предусматривает анализ возможного участия белковых компонентов, из которых собрана молекулярная машина освобождения медиатора. И тут мы подходим к проблеме наиболее трудной как для исследования, так и для непротиворечивого изложения. Дело в том, что существование такой машины и ее многокомпонентность очевидны. И хотя эту задачу пытаются решить представители самых разных направлений современной нейробиологии, пока видны лишь только контуры общей схемы. Основы подхода предложили нейрохимики. Они детально исследовали белки, которые можно обнаружить только в нервных окончаниях, справедливо полагая, что молекулярная машина освобождения медиаторов построена из них. Таких белков набралось более десятка. Затем к делу подключились иммунохимики. Вводя выделенные индивидуальные белки в кровь животных (кроликов, мышей), вызывали выработку у них защитных антител, способных избирательно взаимодействовать с определенным белком или только с его участком. Такие антитела могут служить точным и высокочувствительным инструментом исследования.
Специалисты по электронной микроскопии использовали антитела для картирования пресинаптических белков, т. е. выявления их расположения в нервном окончании в разных стадиях функционирования. Сейчас большие надежды возлагаются на искусство генетиков. Обнаружив участки генома, ответственные за воспроизведение того или иного белка, можно вывести линию мышей-мутантов, лишенных этого белка. А далее дело физиологов определить, в чем и насколько изменилась работа молекулярной машины, какую роль играла в ней удаленная деталь. Это долгая и кропотливая работа, дающая подчас самые неожиданные результаты. Нобелевский лауреат, биохимик А. Сент-Дьерди когда-то сравнивал исследователя с ребенком, который стремится разобрать подаренную игрушку на составные части, чтобы понять, как она устроена, хотя лучше бы научиться ее собирать. Пожалуй, сегодня исследователи механизмов освобождения только приступают к этому этапу познания. Полученные нами данные о том, что влияние пресинаптических рецепторов может быть адресовано непосредственно внутриклеточным компонентам, открывают новую перспективу в изучении механизмов как освобождения медиатора, так и регулирования функциональных связей между нейронами. Действительно, если, лишив животное определенного пресинаптического белка, удастся устранить регулирования, то будет найдена искомая конечная мишень. У нас уже накопились косвенные улики, позволяющие подозревать один из таких белков. Проверка этой гипотезы - предмет наших будущих исследований.
Ионные каналы
- пути прохождения ионов через мембрану клетки. Изменение их состояния (открыто-закрыто) используется для регулирования работы нейронов.
Медиатор
- низкомолекулярное вещество, освобождаемое пресинаптическим нервным окончанием и обеспечивающее перенос сигнала в синапсе.
Постсинаптическая мембрана
- участок мембраны управляемой клетки, входящий в состав синапса.
Пресинаптическая мембрана
- участок мембраны нервного окончания, входящий в состав синапса.
Рецепторы
- белковые молекулы, содержащиеся в пост- и пресинаптических мембранах и воспринимающие химические сигналы медиаторов.
Синапс
- структура, обеспечивающая функциональный контакт между нейронами, а также между нейронами и управляемыми клетками.
Синаптическая щель
- пространство разделяющее пре- и постсинаптическую мембраны.
Синаптические пузырьки
- структурные образования, накапливающие и хранящие медиатор вплоть до момента его освобождения в синаптическую щель. Мембрана синаптических пузырьков содержит разнообразные белки, предположительно участвующие в освобождении медиатора.
3. Материальная основа высшей нервной деятельности.
Нервная система - совокупность структур в организме животных и человека, объединяющая деятельность всех органов и систем и обеспечивающая функционирование организма как единого целого в его постоянном взаимодействии с внешней средой. Нервная система воспринимает внешние и внутренние раздражения, анализирует эту информацию, отбирает и перерабатывает её и в соответствии с этим регулирует и координирует функции организма. Нервная система образована главным образом нервной тканью, основной элемент которой — нервная клетка с отростками, обладающая высокой возбудимостью и способностью к быстрому проведению возбуждения. Эволюция нервной системы претерпеваемая ею в ходе филогенеза, отличается большой сложностью. У простейших — одноклеточных организмов — нервная система отсутствует, но у некоторых инфузорий имеется внутриклеточная сеточка, выполняющая функцию проведения возбуждения к др. элементам клетки. В процессе дальнейшей эволюции строение нервная система усложняется. У свободно живущих кшечнополостных происходит образование и погружение в глубь тела скоплений нервных клеток — нервных узлов (ганглиев), — связи между которыми устанавливаются преим. при помощи длинных отростков (нервных волокон, нервов). Появление такого диффузно-уз-лового типа строения сопровождается развитием специализированных воспринимающих нервных структур (рецепторов), дифференцирующихся в соответствии с воспринимаемым ими видом энергии. Проведение возбуждения становится направленным. У позвоночных животных тип строения нервной системы резко отличается от узлового типа, обычно присущего беспозвоночным. Центральная нервная система (ЦНС) представлена нервной трубкой, расположенной на спинной стороне тела, и состоит из спинного и головного мозга. В эмбриональном развитии позвоночных нервная система образуется из наружного зародышевого листка — эктодермы (сперва в виде нервной пластинки, сворачивающейся в желобок, а затем превращающейся в нервную трубку). Зачаточные экто-дермальные клетки дифференцируются на нейробласты (клетки, дающие начало нейронам) и спонгиобласты (образующие клетки нейроглии). Из эктодермальных клеток, путём их миграции, формируются и периферические узлы, а совокупность отростков некоторых нейробластов образует черепномозговые и спинномозговые нервы, относимые к периферич.
Как спинной, так и головной мозг позвоночных покрыт рядом оболочек и заключён в костные покровы — череп и позвоночник. В процессе эволюции происходит дальнейшее усложнение структуры нервной системы и усовершенствование всех форм её взаимодействия с внешней средой; при этом всё большее значение приобретают прогрессирующие в своём развитии передние отделы головного мозга. У рыб передний мозг почти не дифференцирован, но у них хорошо развиты задний, а также средний мозг; наибольшего развития у рыб достигает мозжечок. У земноводных и пресмыкающихся задний мозг занимает относительно меньший объём, чем у рыб, мозжечок же уступает в развитии среднему мозгу, который делится на 2 части, образуя двухолмие. Усложняется структурно и функционально передний мозговой пузырь, он дифференцируется на промежуточный мозг и 2 полушария с развитой нервной тканью, образующей т. н. первичную кору мозга. Передний мозг, первоначально связанный лишь с обонянием, затем приобретает и более сложные функции. Несколько обособленное место в эволюционном ряду занимают птицы, у которых доминируют структуры т. н. мозгового ствола, т. е. средний мозг и только те части переднего мозга, которые расположены в глубине полушарий (базальные ганглии, промежуточный мозг); сильно развит у птиц и мозжечок; кора головного мозга дифференцирована слабо. Высшего развития нервная система достигает у млекопитающих. Головной конец нервной трубки в эмбриогенезе делится у них на 5 пузырей: передний — даёт начало большим полушариям и промежуточному мозгу, средний — среднему мозгу, задний — делится на собственно задний (варолиев мост и мозжечок) и продолговатый мозг. Кора больших полушарий головного мозга образует многочисленные борозды и извилины. Первичная полость нервной трубки превращается в желудочки мозга и спинномозговой канал. Нейронная организация мозга крайне усложняется. Развитие и дифференциация структур нервной системы у высокоорганизованных животных обусловили её разделение на соматическую и вегетативную нервную систему. Особенность строения вегетативной нервной системы та, что её волокна, отходящие от ЦНС, не доходят непосредственно до эф-фектора, а сначала вступают в периферич. ганглии, где оканчиваются на клетках, отдающих аксоны уже непосредственно на иннервируемый орган. В зависимости от того, где расположены ганглии вегетативной нервной системы и некоторых её функциональных особенностей вегетативную нервную систему делят на 2 части: парасимпатическую и симпатическую. Кроме нервных клеток, в структуру нервной системы входят глиальные клетки. Нейроны являются в известной мере самостоятельными, единицами — их протоплазма не переходит из одного нейрона в другой. Взаимодействие между нейронами осуществляется благодаря контактам между ними. В области контакта между окончанием одного нейрона и поверхностью другого в большинстве случаев сохраняется особое пространство — синап-тич. щель — шириной в несколько сот аксон. Основные функции нейронов: восприятие раздражении, их переработка, передача этой информации и формирование ответной реакции. В зависимости от типа и хода нервных отростков (волокон), а также их функций нейроны подразделяют на: а) рецепторные (афферентные), волокна которых проводят нервные импульсы от рецепторов в ЦНС; тела их находятся в спинальных ганглиях или ганглиях черепно-мозговых нервов; б) двигательные (эфферентные), связывающие ЦНС с эффекторами; тела и дендриты их находятся в ЦНС, а аксоны выходят за её пределы (за исключением эфферентных нейронов вегетативной нервной системы, тела которых расположены в периферич. ганглиях); в) вставочные (ассоциативные) нейроны, служащие связующими звеньями между афферентными и эфферентными нейронами; тыла и отростки их расположены в ЦНС. Деятельность нервной систем основывается на двух процессах: возбуждении и торможении, Возбуждение может быть распространяющимся или местным — не распространяющимся, стационарным (последнее открыто Н. Е. Введенским в 1901) Торможение — процесс, тесно связанный с возбуждением и внешне выражающийся в снижении возбудимости клеток. Одна из характерных черт тормозного процесса — отсутствие способности к активному распространению по нервным структурам (явление торможения в нервных центрах впервые было установлено И. М. Сеченовым в 1863). Клеточные механизмы возбуждения и торможения подробно изучены. Тело и отростки нервной клетки покрыты мембраной, постоянно несущей на себе разность потенциалов (т. н. мембранный потенциал). Раздражение расположенных на периферии чувствит. окончаний афферентного нейрона преобразуется в изменение этой разности потенциалов. Возникающий вследствие этого нервный импульс распространяется по нервному волокну и достигает его пресинаптического окончания, где вызывает выделение в синаптическую щель высокоактивного химического вещества — медиатора. Под влиянием последнего в постсинаптической мембране, чувствительной к действию медиатора, происходит молекулярная реорганизация поверхности. В результате постсинап-тическая мембрана начинает пропускать ионы и деполяризуется, вследствие чего на ней возникает электрич. реакция в виде местного возбуждающего постсинап-тич. потенциала (ВПСП), вновь генерирующего распространяющийся импульс. Нервные импульсы, возникающие при возбуждении особых тормозящих нейронов, вызывают гиперполяризацию пост-синаптич. мембраны и, соответственно, тормозящий постсинаптич. потенциал (ТПСГТ). Помимо этого, установлен и др. вид торможения, формирующийся в пре-синаптич. структуре,— пресинаптич. торможение, обусловливающее длительное снижение эффективности синаптич. передачи . В основе деятельности нервной системы лежит рефлекс, т. е. реакция организма на раздражения рецепторов, осуществляемая при посредстве нервной систем . Термин «рефлекс» был впервые введён в зарождавшуюся физиологию Р. Декартом в 1649, хотя конкретных представлений о том, как осуществляется рефлекторная деятельность, в то время ещё не было. Все рефлекторные процессы связаны с распространением возбуждения по определённым нервным структурам — рефлекторным дугам. Осн. элементы рефлекторной дуги: рецепторы, центростремительный (афферентный) нервный путь, внутрицентральные структуры различной сложности, центробежный (эфферентный) нервный путь и исполнительный орган (эффектор). Различные группы рецепторов возбуждаются раздражителями разной модальности (т. е. качественной специфичности) и воспринимают раздражения, исходящие как из внешней среды (экстерорецепторы — органы зрения, слуха, обоняния и др.), так и из внутренней среды организма (инте-рорецепторы, возбуждающиеся при механических,. химических, температурных и др. раздражениях внутри органов, мышц и др.). Нервные сигналы, несущие в ЦНС информацию от рецепторов по нервным волокнам, лишены модальности, и обычно передаются в виде серии однородных импульсов. Информация о различных характеристиках раздражении кодируется изменениями частоты импульсов, а также приуроченностью нервной импульсации к определённым волокнам (т. н. пространственно-временное кодирование). Совокупность рецепторов данной области тела животного или человека, раздражение которых вызывает определённый тип рефлекторной реакции, называют рецептивным полем рефлекса. Такие поля могут накладываться друг на друга. Совокупность нервных образований, сосредоточенных в ЦНС и ответственных за осуществление данного рефлекторного акта, обозначают термином нервный центр. На отдельном нейроне в нервной системе может сходиться огромное число окончаний волокон, несущих импульсы от др. нервных клеток. В каждый данный момент в результате сложной синаптич. переработки этого потока импульсов обеспечивается дальнейшее проведение лишь одного, определённого сигнала — принцип конвергенции, лежащий в основе деятельности всех уровней нервной системы («принцип конечного общего пути» Шеррингтона, получивший развитие в трудах Ухтомского и др.). Пространственно-временная суммация синаптич. процессов служит основой для различных форм избирательного функционального объединения нервных клеток, лежащего в основе анализа поступающей в нервной системы информации и выработки затем команд для выполнения различных ответных реакций организма. Такие команды, как и афферентные сигналы, передаются от одной клетки к другой и от ЦНС к исполнит, органам в виде последовательностей нервных импульсов, возникающих в клетке в том случае, когда суммирующиеся возбуждающие и тормозящие синаптич. процессы достигают определённого (критического для данной клетки) уровня — порога возбуждения.
Несмотря на наследственно закреплённый характер связей в основных рефлекторных дугах, характер рефлекторной реакции может в значит, степени изменяться в зависимости от состояния центр, образований, через которые они осуществляются. Так, резкое повышение или понижение возбудимости центр, структур рефлекторной дуги может не только количественно изменить реакцию, но и привести к определённым качественным изменениям в характере рефлекса. Примером такого изменения может служить явление доминанты. Важное значение для нормального протекания рефлекторной деятельности имеет механизм т. н. обратной афферента-ции — информации о результате выполнения данной рефлекторной реакции, поступающей по афферентным путям от исполнит, органов. На основании этих сведений в случае, если результат неудовлетворителен, в сформировавшейся функциональной системе могут происходить перестройки деятельности отд. элементов до тех пор, пока результат не станет соответствовать уровню, необходимому для организма (П. К. Анохин, 1935). Всю совокупность рефлекторных реакций организма делят на две основные группы: безусловные рефлексы — врождённые, осуществляемые по наследственно закреплённым нервным путям, и условные рефлексы, приобретённые в течение индивидуальной жизни организма путём образования в ЦНС временных связей. Способность образования таких связей присуща лишь высшему для данного вида животных отделу нервной системы (для млекопитающих и человека — это кора головного мозга). Образование условнореф-лекторных связей позволяет организму наиболее совершенно и тонко приспосабливаться к постоянно изменяющимся условиям существования. Условные рефлексы были открыты и изучены И. П. Павловым в кон. 19 — нач. 20 вв. Исследование условно-рефлекторной деятельности животных и человека привело его к созданию учения о высшей нервной деятельности (ВНД) и анализаторах. Каждый анализатор состоит из воспринимающей части — рецептора, проводящих путей и анализирующих структур ЦНС, обязательно включающих её высший отдел. Кора головного мозга у высших животных — совокупность корковых концов анализаторов; она осуществляет высшие формы анализаторной и интегра-тивной деятельности, обеспечивая совершеннейшие и тончайшие формы взаимодействия организма с внешней средой. Нервная система обладает способностью не только немедленно перерабатывать поступающую в неё информацию при помощи механизма взаимодействующих синаптич. процессов, но и хранить следы прошлой активности (механизмы памяти). Клеточные механизмы сохранения в высших отделах нервной системы длительных следов нервных процессов, лежащие в основе памяти, интенсивно изучаются. Наряду с перечисленными выше функциями нервной системы осуществляет также регулирующие влияния на обменные процессы в тканях — адаптационно-трофическую функцию (И. П. Павлов, Л. А. Орбели, А. В. Тонких и др.). При перерезке или повреждении нервных волокон свойства иннервируемых ими клеток изменяются (это касается как физико-химич. свойств поверхностной мембраны, так и биохимических процессов в протоплазме), что, в свою очередь, сопровождается глубокими нарушениями в состоянии органов и тканей (напр., трофич. язвами). Если иннервация восстанавливается (в связи с регенерацией нервных волокон), то указанные нарушения могут исчезнуть. Изучением строения, функций и развития нервной системы у человека занимается неврология.
4.Физиология восприятия.
Восприятие сложная система процессов приёма и преобразования информации, обеспечивающая организму отражение объективной реальности и ориентировку в окружающем мире. Восприятия вместе с ощущением выступает как отправной пункт процесса познания, доставляющий ему исходный чувственный материал. Будучи необходимым условием процесса познания, восприятия в этом процессе всегда так или иначе опосредуется деятельностью мышления и проверяется практикой. Вне такого опосредования и проверки восприятия может выступать источником как истинного знания, так и заблуждения, иллюзии. К числу процессов восприятия относятся: обнаружение объекта в воспринимаемом поле; различение отд. признаков в объекте; выделение в объекте информативного содержания, адекватного цели действия; ознакомление с выделенным содержанием и формирование образа (или «оперативной единицы» восприятия). Большой вклад в развитие науки о восприятия внесли философы, астрономы, физики, художники — Аристотель, Демокрит, И. Кеплер, Леонардо да Винчи, М. В. Ломоносов, и мн. др. Внимание психологов и физиологов долгое время было сосредоточено на изучении сенсорных (чувственных) эффектов, возникающих под влиянием тех или иных объективных воздействий, в то время как самый процесс восприятия оставался за пределами исследования. Методология такого подхода опиралась на сенсуализм в теории познания, особенно развитый Дж. Локком и франц. материалистами (П. Кабанис и Э. Кондильяк). В психологии этот подход получил наиболее ясное выражение в концепции восприятия согласно которой сенсорный образ возникает в результате воздействия внешних агентов на воспринимающие органы чувств пассивно созерцающего субъекта. Ограниченность такого подхода — игнорирование деятельности субъекта, исследование лишь результатов процесса восприятия представление о корковом звене анализаторов как о субстрате сенсорных процессов, месте, где якобы происходит преобразование нервных процессов в идеальные психические образы,— практически затрудняла разработку способов управления процессами восприятия в целях его совершенствования и развития, а теоретически вела либо к различным субъективно-идеалистическим теориям, либо к отказу от естеств.-науч. объяснения восприятия. Решающий шаг в преодолении пассивной «рецепторной» концепции был сделан советскими психологами, которые, исходя из методологии диалектического материализма и сеченовского понимания рефлекторной природы сенсорных процессов, рассматривают восприятия как своеобразное действие, направленное на обследование воспринимаемого объекта и на создание его копии, его подобия. В 60х гг. 20 в. исследования восприятия ведутся представителями различных специальностей на различных уровнях процессов приёма и переработки информации. На уровне входа воспринимающих систем (сетчатка глаза, кортиев орган уха и т. д.) исследуются анатомо-морфологич., биофиз., электро-физиол. особенности деятельности рецепторов. Восприятие изучается также на нейронном, психофизиол., психологич., социаль-но-психологич. уровнях. В кибернетике и бионике ведутся многочисленые исследования по созданию технических устройств, имитирующих работу органов чувств. Результаты разнообразных исследований восприятия публикуются в десятках журналов, посвящённых преимущественно проблемам восприятия. Следует, однако, подчеркнуть, что до наст. времени не удалось построить единую теорию восприятия, которая интегрировала бы результаты многочисленых исследований. С большими трудностями сталкиваются попытки моделирования таких свойств восприятия, как осмысленность, предметность, константность и др. Согласно современным представлениям, совокупность процессов восприятия обеспечивает субъективное, пристрастное и вместе с тем адекватное отражение объективной реальности. Адекватность образа восприятия (его соответствие действительности) достигается благодаря тому, что при его формировании происходит уподобление (А. Н. Леонтьев), т. е. подстраивание воспринимающих систем к свойствам воздействия: в движении руки, ощупывающей предмет, в движении глаза, прослеживающего видимый контур, в движениях гортани, воспроизводящих слышимый звук, и т. д.— во всех этих случаях создаётся копия, сопоставимая с оригиналом; сигналы рассогласования, поступая в нервную систему, выполняют корректирующую функцию по отношению к формирующемуся образу и соответственно к практическим действиям, реализующимся на основе этого образа. Следовательно, восприятия представляет собой своеобразный саморегулирующийся процесс, обладающий механизмом обратной связи и подчиняющийся особенностям отражаемого объекта. Важное свойство восприятия — возможность перестройки чувств, моделей воздействующего на субъект внешнего мира, смены способов их построения и опознавания. Один и тот же объект может служить прототипом многих перцептивных (от лат. perceptio — восприятие) моделей. В процессе их формирования они уточняются, из объекта извлекаются инвариантные свойства и признаки, что приводит в итоге к тому, что мир воспринимается таким, каким он существует на самом деле. Целенаправленные процессы восприятия (перцептивные действия) выступают в своей развёрнутой, внешней форме лишь на ранних ступенях онтогенеза, где наиболее отчётливо обнаруживаются их структура и их роль в формировании образов восприятия. В дальнейшем они претерпевают ряд последоват. изменений и сокращений, пока не облекаются в форму мгновенного акта «усмотрения» объекта, который был описан представителями гештальт-психологии и ошибочно принимался ими за исходную генетически первичную форму восприятия.
Любая живая система обладает выработанным алфавитом, т. е. определённой совокупностью образов или перцептивных моделей. Если на фазе построения образа объекта происходит уподобление воспринимающих систем свойствам воздействия, то на фазе опознания или оперирования сложившимися образами характеристики и направленность процесса уподобления существенно изменяются (А. В. Запорожец): с одной стороны, субъект воссоздаёт с помощью собств. движений и действий некоторое подобие, образ воспринимаемого объекта; с др. стороны, происходит перекодирование, перевод получаемой информации на «язык» оперативных единиц восприятия или перцептивных моделей, уже освоенных субъектом. Эта вторая сторона выражает тот факт, что одновременно с уподоблением воспринимающей системы субъекта объекту происходит уподобление объекта субъекту, и только это двустороннее преобразование приводит к формированию полноценного, адекватного и вместе с тем субъективного образа объективной реальности. В развитых процессах восприятия имеются спец. перцептивные действия; на их основе производится выделение информативного содержания, по которому субъект может сличать предъявленный объект с накопленными им перцептивными моделями, осуществлять собственно процесс сличения и, наконец, опознание и отнесение объектов к тому или иному классу, т. е. их категоризацию. Процессы опознания требуют значительно меньше времени, чем процессы формирования образа; для сличения и идентификации нужно только извлечь из предъявленного объекта некоторые инвариантные свойства и признаки. Воспринимающие системы (особенно ярко это проявляется в зрении) обладают некоторой «манипулятивной» способностью: субъект в короткое время имитирует процессы формирования образа, как бы с разных сторон рассматривая объект и находя такую позицию, при которой максимально облегчаются процессы сличения и идентификации. В целостном акте поведения существует ещё одна своеобразная форма уподобления: процессы переструктурирования и трансформации образа с целью привести информацию к виду, пригодному для принятия решения. В таком процессе уподобления решается задача изменения реальности, адекватного планам и задачам поведения. Такому изменению реальности предшествует преобразование образа ситуации, как правило, не осознаваемое субъектом, но тем не менее вносящее существ, вклад в решение стоящих перед ним жизненных задач. Восприятия — не пассивное копирование действительности, а активный творческий процесс познания. Изучение восприятия имеет важное значение и свои специфич. стороны в области эстетики, педагогики, спорта и т. д.
5. Физиологические механизмы памяти.
Память способность к воспроизведению прошлого опыта, одно из основных свойств нервной системы, выражающееся в способности длительно хранить информацию о событиях внешнего мира и реакциях организма и многократно вводить её в сферу сознания и поведения. Память в нейрофизиологии. Память свойственна человеку и животным, имеющим достаточно развитую центральную нервную систему (ЦНС). Объём памяти длительность и надёжность хранения информации, как и способность к восприятию сложных сигналов среды и выработке адекватных реакций, возрастают в ходе эволюции по мере увеличения числа нервных клеток (нейронов) мозга и усложнения его структуры. Физиологические исследования памяти обнаруживают 2 основных этапа её формирования, которым соответствуют два вида памяти: кратковременная и долговременная. Кратковременная память характеризуется временем хранения информации от секунд до десятков минут и разрушается воздействиями, влияющими на согласованную работу нейронов (электроток, наркоз, гипотермия и др.). Долговременная память, время хранения информации в которой сравнимо с продолжительностью жизни организма, устойчива к воздействиям, нарушающим кратковременную память. Переход от первого вида памяти ко второму постепенен. Нейрофизиологи полагают, что кратковременная память основана на активных механизмах, поддерживающих возбуждение определённых нейронных систем. При переходе к долговременной памяти связи между нейронами, входящими в состав таких систем, фиксируются структурными изменениями в отд. клетках. Опыты с иссечением участков коры больших полушарий головного мозга и электрофизиологические исследования показывают, что -«запись»- каждого события распределена по б. или м. обширным зонам мозга. Это позволяет думать, что информация о разных событиях отражается не в возбуждении разных нейронов, а в различных комбинациях со возбужденных участков и клеток мозга. Нервные клетки не делятся в течение жизни, и новые реакции могут вырабатываться и запоминаться нервной системой только на основе создания новых связей между имеющимися в мозге нейронами. Новые нейронные системы фиксируются за счёт изменений в межнейронных кон тактах — синапсах, в которых нервный импульс вызывает выделение спец. химические вещества — медиатора, способного облегчить или затормозить генерацию импульса следующим нейроном. Долго временные изменения эффективности синапсов могут быть обусловлены изменениями в биосинтезе белков, от которых зависит чувствительность синоптической мембраны к медиатору. Установлено, что биосинтез белков активируется при возбуждении нейронов на разных уровнях организации ЦНС, а блокада синтеза нуклеиновых кислот или белков затрудняет или исключает формирование долговременной памятью. Очевидно, что одна из функций активации синтеза при возбуждении — структурная фиксация нейронных систем, что и лежит в основе долго временной памяти. Имеющиеся экспериментальные данные не позволяют пока решить, происходит ли проторение путей распространения возбуждения за счёт увеличения проводимости имеющихся синапсов или в результате возникновения дополнит, межнейронных связей. Оба возможных механизма нуждаются в интенсификации белкового синтеза. Первый — сводится к частично изученным явлениям клеточной адаптации и хорошо согласуется с представлением об универсальности основная биохимическая систем клетки. Второй — требует направленного роста отростков нейронов и, в конце концов, кодирования поведенческой информации в структуре химических агентов, управляющих таким ростом и заложенных в генетическом аппарате клетки. Для исследования памяти применяют методы клинич. и экспериментальной психофизиологии, физиологии поведения, морфологии и гистохимии, электрофизиологии мозга и отдельных нейронов, фармакологические методы, а также методы аналитической биохимии. В зависимости от задач, подлежащих решению, исследование механизмов памяти осуществляется на разных объектах — от человека до культуры нервных клеток.
Литература:
1. Анохин П. К., Биология и нейрофизиология условного рефлекса, М., 1968;
2. 2Бериташвили И. С., Память позвоночных животных, ее характеристика и происхождение, 2 изд., М., 1974;
3. Агранов Б., Память и синтез белка, в кн.: Молекулы и клетки, пер. с англ., в. 4, М., 1969;
4. Волков Н.Н., Восприятие предмета и рисунка, М., 1950;
5. Соколов Е. Н., Восприятие и условный рефлекс, М., 1958;
6. Ананьев Б. Г., Психология чувственного познания, М., 1960;
7. Леонтьев А. Н., Проблемы развития психики, 2 изд., М., 1965;
8. Розенблатт Ф., Принципы нейродинамики, пер. с англ., М., 1965;
9. Ярбус А. Л., Роль движений глаз в процессе зрения, М., 1965;
10. Шехтер М. С., Психологические проблемы узнавания, М., 1967;
11. Восприятие и действие, М., 1967;
12. Большая советская энциклопедия.
|
