Я - тоже муха:
Мой краток век.
А чем ты, муха,
Не человек?
ВИЛЬЯМ БЛЕЙК.
"Муха"
Если почистить фрукты и то, что считается несъедобным, день-другой не выносить на помойку, можно заметить, как вокруг этого при родного растительного материала разовьется свояособая жизнь. Вокруг отвергнутых человекомостатков пищи начнут роиться маленькиемушки размером по крайней мере на порядок, как говорят математики (т.е. примерно в 10 раз), меньше, чем обычные комнатные мухи. Они настолько малы, что, если заведется всего несколько штук, их можно и не заметить. Размеры мушек составляют только 2-3,5 мм. Однако это крохотное создание вошло в историю науки, а следовательно, и в историю человечества как бесценный объект генетических исследований.
В русский язык уже прочновошло их на звание - дрозофила,
в точности повторяющее латинское наименование рода мух семейства плодовых мушек Drosophilia. На земном шаре существует свыше 1000 видов этих прелестных существ, и наибольшее распространение они получили в субтропиках и тропиках - на одних только Гавайских островах обитает более 300 видов дрозофилы. На территории же нашей страны их на порядок меньше. Наиболее используемым в науке видом является Drosophilia melanogaster.Сами плодовые мушки питаются соком растений, гниющими растительными остатками, а личинки - микроорганизмами. Их жизненный цикл очень короток, и развитие от яйца до мухи занимает в среднем 10 суток. Можно получить массу удовольствий, часами разглядывая эти крошечные создания под микроскопом с небольшим увеличением, лучше под бинокулярным микроскопом, позволяющим получать объемное изображение. В нем легко разглядеть детали строения переливающихся крылышек, удивительно красиво посаженные глазки, прямые или вилочкообразные щетинки и многое, многое другое. Малые размеры, плодовитость и ряд других преимуществ перед большинством живых организмов на долгое время сделали дрозофилу главным объектом генетики, и не один нобелевский лауреат кроме своего могучего интеллекта обязан и ей своими высшими научными достижениями.
Одним из нобелевских лауреатов, которому дрозофилы оказали неоценимую услугу, был замечательный американский зоолог и генетик Томас Хант Морган. Именно его имя использовалось для шельмования отечественных биологов в период "лысенковщины", когда были введены ругательные тогда слова вейсманисты-морганисты, менделисты-морганисты или просто морганисты.
Морган родился в 1866 г. в семье дипломата. Мать его была внучкой композитора Фрэнсиса Скопа Ли сочинившего американский национальный гимн. Не испытывая влияния биологов, Томас с детства интересовался биологическими объектами - он приносил в дом окаменелости, собрал коллекцию различных птиц, а первые свои научные исследования он выполнил, используя морских пауков.
Реклама

В 1902 г. американский биолог Уильям С. Саттон высказал предположение, что единицы наследственности (гены) размещаются в хромосомах. Морган решил его опровергнуть, считая, что хромосомы не являются носителями наследственности, а возникают на ранних стадиях развития. Теперь мы уже знаем, что в конце концов он изменил свое мнение на противоположное, доказав прямую роль хромосом в процессах наследования макроскопических признаков живых организмов. Для выполнения этой работы в 1908 г. лучшего объекта, чем дрозофила, Морган найти не смог. Ему понадобилось вырастить и изучить несколько миллионов мушек, чтобы прийти к твердому убеждению, что хромосомы напрямую связаны с наследственностью. И это только один ученый. Дрозофилу использовали наверняка не менее нескольких сотен генетиков, и, следовательно, число мушек, понадобившихся науке, сравнимо с численностью людей на земном шаре.
Результаты некоторых экспериментов Моргана с дрозофилой, казалось, противоречили менделевскому закону независимого наследования, согласно которому каждый организм обладает генами, контролирующими тот или иной признак, и наследование одного признака, например пола, не зависит от наследования другого - например цвета глаз. Оказалось, что некоторые признаки все же связаны между собой, т.е. их сочетание встречается у потомков чем следует из законов Менделя.Так, например, белоглазость - мутантный признак - почти всегда встречается только у самцов. Это явление Морган назвал сцеплением с полом. Тенденция к сцеплению подсказало ученому, что гены, по-видимому, располагаются на одной и той же хромосомев тесной близости друг к другу. Было обнаружено что таких сцепленных групп у дроздофилы - четыре, и эта величина в точности совпала числом пар хромосом
Свои эксперименты Морган проводил в помещении, которое он называл мушиной комнатой. В 1914 г. в эту комнату явился студент-выпускник Герман Джозеф Меллер, которому впоследствии на той же дрозофиле суждено было открыть мутации под действием рентгеновского излучения. А пока Морган, Меллер и другие сотрудники старались ответить на вопрос, почему гены, расположенные на одной и той же хромосоме, наследовались реже, чем этого можно ожидать.Они предположили, что хромосомы, собранные в пары,могут расщепляться и обмениваться своими участками, генами, и назвали этот процесс кроссинговером. Предположение основывалось на обнаруженном бельгийским ученым Ф.А.Янсеном в 1909 г. (с помощью светового микроскопа) тесном переплетении хромосом.
Реклама

Морган рассуждал так: чем больше расстояние между двумя генами в одной хромосоме, тем больше вероятность разрыва. Если это верно, то гены не будут наследоваться вместе, и наоборот - гены, расположенные в хромосоме близко друг от друга, имеют меньше шансов быть разделенными, т.е. верна гипотеза американского генетика Альфреда Генри Стертеванта о том, что сцепление двух генов в хромосоме определяется величиной линейного расстояния между ними. Иными словами, была высказана замечательная мысль, что, гены расположены вдоль хромосомы линейно, т.е. представляют собой линейную матрицу. Используя данные о частотах кроссинговера, Морган первым начал составлять хромосомные (или генетические) карты, где в линейной последовательности указывались гены, ответственные за тот или иной макроскопический признак (цвет глаз или брюшка, формой щетинок или крыльев и т.д.). Так, если частота обмена между двумя генами равна 5, то это означает, что они расположены в одной и той же хромосоме на расстоянии 5 условных линейных единиц. В дальнейшем имя Моргана, как ранее имена ряда других выдающихся ученых, например Дальтона, Ньютона, Джоуля, Ангстрема и других, стали использовать для наименования этих единиц, и в настоящее время они называются морганидами. В 1933 г. Т. X. Моргану была присуждена Нобелевская премия по физиологии и медицине за открытия, связанные с ролью хромосом в наследственности.
Морган изучал в основном самопроизвольно возникающие мутации. Но они могут быть вызваны и искусственно, например путем физических или химических воздействий. Первым физическим мутагенным фактором было рентгеновское излучение. Его использовал ученик Моргана Герман Меллер. Химический мутагенез еще в 30-е гг. открыл наш соотечественник Иосиф Абрамович Рапопорт, герой Великой Отечественной войны (он дважды был представлен к званию Героя Советского Союза) и настоящий герой науки во времена борьбы с "лысенковщиной". Однако опубликовать свои результаты о мутагенном действии формальдегида и других карбонильных соединений он смог только в 1946 г. одновременно с шотландкой Шарлоттой Ауэрбах, сообщившей в научной печати об аналогичных свойствах иприта. И опять объектом исследования было неприметная мушка.
Ричард Эксел (RichardAxel) и его коллеги изучили функции двух десятков генов, ответственных за восприятие вкуса у дрозофилы. Об этом - статья в Cell
от 9 марта.
Группа Эксела продолжила исследования молекулярных биологов из Йельского университета Питера Клайна и Джона Карлсона, идентифицировавших комплекс генов под общим названием GR. Эти гены отвечают за формирование вкусовых рецепторов дрозофил: кодируемые ими белки присутствуют преимущественно в хоботках, ножках и усиках насекомых.
Эксел и его коллеги обнаружили большое сходство между GR и обонятельными генами дрозофилы. Это позволяет предположить, что у отдаленных предков мух за распознавание вкуса и запаха отвечали одни и те же гены, и лишь в процессе эволюции постепенно произошло их разделение на "вкусовые" и "обонятельные".
Исследователи считают, что дальнейшее изучение белков, отвечающих за распознавание вкуса и запаха, может привести к революции в сельском хозяйстве. Разобравшись в структуре молекул; можно будет создавать экологически чистые препараты, способные сделать сельскохозяйственные культуры невкусными или дурно пахнущими для насекомых-вредителей.
Различия в организации эу- и гетерохроматина дрозофилы.
Наиболее существенные свойства эу- и гетерохромати на, характеризующие различия в их организации, при ведены в табл. 1.
Даже первого взгляда на эту таблицу достаточно чтобы увидеть, насколько эу- и гетерохроматин различны по строению и функционированию. Некоторые из свойств очень интересны, и мы рассмотрим их боле детально.
Эффект положения мозаичного типа
Одним из удивительных свойств гетерохроматина является его способность передавать компактизованное состояние на эухроматиновые фрагменты хромосом, перенесенные в его соседство с помощью хромосомных перестроек (рис. 2). Гены в перенесенном фрагменте инактивируются, хотя и не во всех клетках одного и того же органа. Например, если ген w+
у дрозофилы, обеспечивающий нормальный красный цвет глаз у мухи, переносится с помощью инверсии In
(
l
)
wm
4
в новое положение - в окружение прицентромерного гетерохроматина, в части клеток он инактивируется. В результате на фоне нормально окрашенных участков глаза будут появляться пятна из неокрашенных белых клеток, в которых ген w+
инактивирован - образуется как бы мозаика из окрашенных и неокрашенных клеток (см. рис. 2). Это явление; называемое эффектом положения мозаичного типа, в настоящее время изучают весьма интенсивно, поскольку исследователи полагают, что оно является удобной моделью для понимания генетического контроля механизмов компактизации - декомпактизации хроматина.
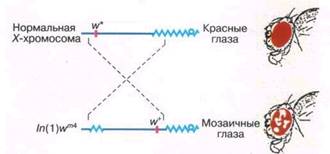
Рис. 2. Схема, иллюстрирующая эффект положения мозаичного типа - генетическую инактивацию эухроматинового фрагмента хромосомы, содержащего ген
w
+
,
перенесенного в соседство гетерохроматина
Во всех этих работах было сделано сенсационное открытие: наследственность можно преднамеренно изменять в лабораторных условиях. Последнюю точку в исследовании хромосом и генов как линейных матриц, по-видимому, поставил еще один наш соотечественник, лауреат Кимберовской премии (премии по генетике,дополняющей Нобелевские по физиологии и медицине) Николай Владимирович Тимофеев-Ресовский, который совместно с немецкими учеными Клаусом Циммерманом и Максом Дельбрюком (еще одним нобелевским лауреатом) в 30-х гг. определил размер гена. И уже не в условных единицах, а в обычных единицах длины (например, в нанометрах). Полученные величины великолепно совпали с более поздними данными о размерах ДНК.
Гены Y-хромосомы.
Еще на заре рождения генетики, в 1916 году, американский ученый К. Бриджес установил, что экспериментально полученные самцы дрозофилы без Y-хромосомы (то есть ХО в отличие от нормальных самцов XY) имеют нормальную жизнеспособность и строение всех органов, но они полностью стерильны. В последующих экспериментах было показано, что Y-хромосома дрозофилы содержит только девять генов, из которых шесть влияют на способность самцов оставлять потомство (фертильность). Оставшиеся три гена - это bobbed
(
bb
),
серия или кластер генов, кодирующих рибосомную РНК и активность которых приводит к образованию ядрышка (нужно упомянуть, что второй ядрышкообразующий ген bb
у дрозофилы находится также в гетерохроматиновом районе, но Х-хромосомы). Ген bb,
состоящий из повторенных фрагментов, занимает около 5% всей ДН К Y-хромосомы.
В пределах гена bb
находятся участки, контролирующие процесс коньюгации хромосом в мейозе. Дело в том, что в мейозе спариваются гомологичные хромосомы за счет конъюгации гомологичных последовательностей нуклеотидов ДНК. Поскольку половые Х- и Y-хромосомы морфологически и функционально совершенно различны, вопрос о механизмах спаривания этих элементов в мейотической профазе I достаточно актуален. Начиная с 1930-х годов накапливались данные о наличии участков спаривания в гетерохроматине Х-хромосомы, в районе локализации гена bobbed
.
Их назвали сайтами collohores
(
col
).
В 1990 году удалось показать, что ответственными за опознание Х- и Y-хромосом и их последующую конъюгацию и расхождение в мейозе являются короткие последовательности нуклеотидов длиной в 240 п.н., расположенные в промежутках между генами рибосомной РНК, как в Х-, так и Y-хромосоме. Участок локализации локуса со/ занимает в Y-хромосоме около 7% ее длины. Удаление bb
с
помощью хромосомных нехваток (делений) полностью нарушает правильную конъюгацию половых хромосом.
Еще один ген - crystal
(
cry
) влияет на поведение хромосом в мейозе и правильное формирование гамет. Разрывы участка хромосом, занимаемого этим геном, не приводят к развитию каких-либо фенотипических изменений у самцов дрозофил. Однако при полном или частичном удалении этого участка с помощью делений в первичных сперматоцитах, в клетках, из которых образуются сперматозоиды, появляются белковые кристаллы, а во время мейоза нарушается расщепление хромосом. Интересно отметить, что есть еще один ген, расположенный в эухроматине Х-хромосомы, - Stellate
(
Ste
),
который взаимодействует с геном crystal
.
При этом, если в Х-хромосоме присутствует нормальный аллель гена Stellate
(Ste+
),
кристаллы имеют игловидную форму, если мутантный Ste
-
-
они приобретают вид звезды. Ген Ste
+
был клонирован, и в результате анализа ДНК было показано, что он содержит тандемно повторенную (до 200 раз) последовательность длиной 1250 п.н. Нужная степень повторенности этого фрагмента соответствует аллелю Ste+
(игловидные кристаллы у Ste+
/0
самцов, то есть тех, которые не имеют Y-хромосомы). Высокая степень повторенности приводит к образованию звездовидных кристаллов у Ste-
/О. Транскрипты гена Ste
-
находят в семенниках. Ген Ste+
кодирует бета-субъединицу фермента казеин-киназы-2. Этот белок, по-видимому, вовлечен в процессы конденсации хромосом и их последующего расхождения по гаметам.
Присутствие нормального аллеля гена crystal
ингибирует накоплениеРНК гена Ste
+
.
По существующим представлениям сгу+
контролирует активность гена Ste+
: удаление Y-хромосомы приводит к сверхпродукции Ste+
-PHK, в результате чего избыток белка этого гена кристаллизуется в сперматоцитах и нарушает их функциональные возможности, что и приводит к стерильности.
У D
.
melanogaster
найдено шесть факторов фертильности самцов (
kl
-5,
kl
-3,
kl
-2,
kl
-1,
ks
-
I
и
ks
-2
нарис. З) Из них три очень больших: kl-5, kl-3
и ks
-1 —
занимают по 10% Y-хромосом каждый, то есть примерно по 4000 т.п.н.
Интересно проявляется активность факторов фертильности у дрозофилы. В 1961 году три немецких ученых (G.F. Меуег, О. Hess, W. Beermann) описали особые нитевидные структуры в ядрах развивающихся сперма тоцитов D
.
melanogaster
,
которые впоследствии стал называть петлями (рис. 5). Такие структуры нашли фактически у всех 50 изучаемых видов дрозофилы. Показано, что петли - это декомпактизованные, а следовательно, активные участки Y-хромосом. В них синтезируется РНК и накапливаются белки. Каждая петля ядре данного вида дрозофилы имеет характерные размеры, ультраструктуру и внешний вид (см. рис. 5). У других видов морфология набора петель другая.
О том, что петли формируются из материала Y-xpомосомы, свидетельствуют следующие факты.
1. У самцов, не имеющих Y-хромосомы (ХО), нет ипетель, а у особей с двумя Y-хромосомами (XYY)ониприсутствуют в двойном наборе. Если происходит делеция части Y-хромосомы, обнаруживаются не все петли. В линиях с дупликациями частей Y-хромосом число петель соответственно увеличивается.
2. У межвидовых гибридов морфология петель такая же, как и у вида - донора Y-хромосомы.
Более детальный анализ показал, что гены ферментильности самцов локализованы в петлях.
1.Сначала были установлены корреляциимежду числом генов и петель. Затем, используя хромосомные – перестройки, установили прямое соответствие в их локализации. Так, фактор kl
-5
соответствует петле А, поскольку и петля, и фактор располагаются между точками разрывов одних и тех же перестроек (см. рис. 3). Фактор kl
- 3
расположен в петлеB,
ks
-1 -
в петле С.
2. При удалении делециями хотя бы одной петли самец становится стерильным.
После получения клонов ДНК из Y-хромосом дрозофил появилась возможность анализа молекулярной организации этой хромосомы. Общая длина петель составляет около 1000 мкм, или 1/12 всей длины ДНК в Y-хромосоме. Функции остальных 11/12 пока неизвестны. В состав ДНК Y-хромосомы входят два типа повторенных последовательностей.
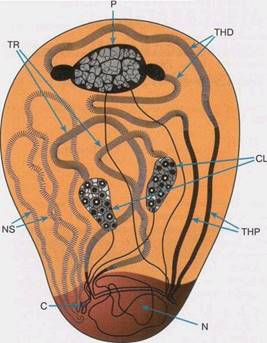
Рис. 3. Общий вид ядра спермотоцита у самца
Drosofilia
hydei
(из [1], с.62).
TR
,
P
,
THD
,
CL
,
THP
,
NS
– названия петель, С – центромера,
N
- ядрышко
Вывод:
Таким образом, муха Дрозофила играет большую роль, как объект генетических исследований. Исследование ее генов принесло известность многим генетикам. Изучая гены Дрозофилы, ученые открыли много законов, таких как: закон Моргана - сцепленного наследования генов и закон Менделя. К тому же у Дрозофилы нашли много различных генов, отвечающих за наследование некоторых признаков. В последствии многие из этих открытий применялись к людям. И если бы не было этих мух, ученые еще долго бы не открыли законы наследования у людей.
Таблица 1.
| Свойства |
Эухроматин |
Гетерохроматин |
| Доля генома |
67% |
33% |
| Расположение в хромосомах |
Плечи хромосом |
В прицентромерных областях, вся Y-хромосом |
| Состояние компактности в клеточном |
В ходе митотического и мейоти- |
На протяжении всего клеточного цикла |
| Цикле |
ческого делений |
| Компактизующее влияние на приближен |
Не оказывает |
Участки эухроматина, приближенные к гетеро |
| ные участки хромосом (эффект положе |
хроматину, также становятся компактными, |
| ния мозаичного типа) |
гены в них инактивируются |
| Способность объединяться с другими |
Не отмечена |
Гетерохроматиновые участки объединяйте; |
| районами хромосом |
образуя хромоцентры |
| Образование хромосомных перестроек |
Обычная частота обнаружения |
Повышенная частота обнаружения |
| Расположение в клеточном ядре |
По всему объему ядра |
Главным образом на ядерной оболочке |
| Время синтеза ДНК в клеточном цикле |
Первые 3/4 периода синтеза ДНК |
Последняя половина S-периода. Завершени |
| в интерфазе (S-периода) |
процесса репликации ДНК сильно задержано |
| Дифференциальная окраска специфичес |
Отсутствие окраски |
Интенсивная окраска |
| кими красителями (С-окраска) |
| Фракции ДНК по степени повторенности |
-90% уникальных последователь |
Основная масса ДНК представлена высокопое |
| ностей и -10% умеренно повто |
торенными фракциями, в меньшей степен |
| ренных |
умеренными повторами и совсем мало уникал! |
| ных последовательностей |
| Наличие особых компактизующих бел |
Почти отсутствует |
Обильно присутствует по всему гетерохромг |
| ков, например белка НР1 |
тину |
| Варьирование количества материала в |
Заметное варьирование не обна |
Варьирование количества гетерохроматина un |
| хромосомах |
ружено |
роко представлено в каждой хромосоме |
| Генетическое содержание |
Основная часть всех генов генома |
Гены почти отсутствуют |
| локализована в эухроматине |
Литература:
«Соросовский Образовательный Журнал» том 6 №2 2000
И. Ф. Жимулев «Молекулярная и генетическая организация гетерохромотина в хромосомах дрозофилы».
Биология №14 1996.
А.А. Замятнин «Хромосомные матрицы, илиОда в честь плодовой мушки-дрозофилы»
Газета «Поиск» №11 2001
|