| Содержание
Введение 2
1. Основные факты о строении клеточной мембраны 3
2. Общие представления о проницаемости 4
3. Перенос молекул через мембрану
4
3.1. Диффузия 5
3.2 Уравнение Фика 6
3.3 Пассивный транспорт 7
3.3.1 Отличия облегченной диффузии от простой 8
4. Закон Дарси 8
5. Активный транспорт 9
6. Строение и функции ионных каналов 11
Заключение
15
Список литературы 17
ВВЕДЕНИЕ
Мембранный транспорт – транспорт веществ сквозь клеточную мембрану в клетку или из клетки, осуществляемый с помощью различных механизмов – простой диффузии, облегченной диффузии и активного транспорта.
Важнейшее свойство биологической мембраны состоит в ее способности пропускать в клетку и из нее различные вещества. Это имеет большое значение для саморегуляции и поддержания постоянного состава клетки. Такая функция клеточной мембраны выполняется благодаря избирательной проницаемости, т.е. способностью пропускать одни вещества и не пропускать другие. Легче всего проходят через липидный бислой неполярные молекулы с малой молекулярной массой (кислород, азот, бензол). Достаточно быстро проникают сквозь липидный бислой такие мелкие полярные молекулы, как углекислый газ, оксид азота, вода, мочевина. С заметной скоростью проходят через липидный бислой этанол и глицерин, а также стероиды и тиреоидные гормоны. Для более крупных полярных молекул (глюкоза, аминокислоты), а также для ионов липидный бислой практически непроницаем, так как его внутрення часть гидрофобна. Так, для воды коэффициент проницаемости (см/с) составляет около 10-2, для глицерина – 10-5, для глюкозы – 10-7, а для одновалентных ионов – меньше 10-10.
Перенос крупных полярных молекул и ионов происходит благодаря белкам-каналам или белкам-переносчикам. Так, в мембранах клеток существуют каналы для ионов натрия, калия и хлора, в мембранах многих клеток – водные каналы аквапорины, а также белки-переносчики для глюкозы, разных групп аминокислот и многих ионов. Активный и пассивный транспорт.
Мембраны формируют структуру клетки и осуществляют ее функции. Нарушение функций клеточной и внутриклеточной мембран лежит в основе необратимого повреждения клеток и, как следствие, развитие тяжелых заболеваний сердечно-сосудистой, нервной, эндокринной системы.
Реклама

1. Основные факты о строении клеточной мембраны.
К клеточным мембранам относятся плазмолемма, кариолемма, мембраны митохондрий, ЭПС, аппарата Гольджи, лизосом, пероксисом. Общей чертой всех мембран клетки является то, что они представляют собой тонкие (6-10 нм) пласты липопротеиновой природы, (липиды в комплексе с белками). Основными химическими компонентами клеточных мембран являются липиды (40%) и белки (60%); кроме того, во многих мембранах обнаружены углеводы (5-10%).
Плазматическая мембранна окружает каждую клетку, определяет ее размер и обеспечивает сохранение различий между содержимым клетки и внешней средой. Мембрана служит высокоизбирательным фильтром и отвечает за активный транспорт веществ, то есть, поступление в клетку питательных веществ и вывод наружу вредных продуктов жизнедеятельности. Наконец, мембрана ответственна за восприятие внешних сигналов, позволяет клетке реагировать на внешние изменения. Все биологические мембраны представляют собой ансамбли липидных и белковых молекул, удерживаемых вместе с помощью нековалентных взаимодействий.
Основу любой молекулярной мембраны составляют молекулы липидов, образующих бислой. К липидам относится большая группа органических веществ, обладающих плохой растворимостью в воде (гидрофобность) и хорошей растворимостью в органических растворителях и жирах (липофильность). Состав липидов в разных мембранах неодинаков. Например, плазматическая мембрана, в отличие от мембран эндоплазматической сети и митохондрий обогощена холестерином. Характерными представителями липидов, встречающихся в клеточных мембранах, являются фосфолипиды (глицерофосфатиды), сфингомиелины и из стероидных липидов – холестерин.
Особенностью липидов является разделение их молекул на две функционально различные части: гидрофобные неполярные, не несущие зарядов («хвосты»), состоящие из жирных кислот, и гидрофильные, заряженные полярные «головки». Это определяет способность липидов самопроизвольно образовывать двухслойные (билипидные) мембранные структуры толщиной 5-7 нм.
Первые опыты, подтверждающие это, были проведены в 1925 году.
Формирование бислоя является особым свойством молекул липидов и реализуется даже вне клетки. Важнейшие свойства бислоя: способность к самосборке – текучесть – ассиметричность.
2. Общие представления о проницаемости.
Xарактеристика мембран, стенок сосудов и эпителиальных клеток, отражающая способность проводить химические вещества; различают активную (активный транспорт веществ) и пассивную П. (фагоцитоз <phagocytosis> и пиноцитоз <pinocytosis>); пассивная и (в ряде случаев) активная П. (крупных молекул) обеспечиваются мембранными порами, П. для низкомолекулярных веществ (например, ионов) обеспечивается специфическими мембранными структурами с участием молекул-переносчиков.
Реклама

3. Перенос молекул через мембрану.
Так как внутренняя часть липидного слоя гидрофобна, он представляет собой практически непроницаемый барьер для большинства полярных молекул. Вследствие наличия этого барьера, предотвращается утечка содержимого клеток, однако из-за этого клетка была вынуждена создать специальные механизмы для транспорта растворимых в воде веществ через мембрану. Перенос малых водорастворимых молекул осуществляется при помощи специальных транспортных белков. Это особые трансмембранные белки, каждый из которых отвечает за транспорт определенных молекул или групп родственных молекул.
В клетках существуют также механизмы переноса через мембрану макромолекул (белков) и даже крупных частиц. Процесс поглощения макромолекул клеткой называется эндоцитозом. В общих чертах механизм его протекания таков: локальные участки плазматической мембраны впячиваются и замыкаются, образуя эндоцитозный пузырек, затем поглощенная частица обычно попадает в лизосомы и подвергается деградации.
3.1 Диффузия (лат. diffusio — распространение, растекание, рассеивание) — процесс переноса материи или энергии из области с высокой концентрацией в область с низкой концентрацией (против градиента концентрации). Самым известным примером диффузии является перемешивание газов или жидкостей (если в воду капнуть чернил, то жидкость через некоторое время станет равномерно окрашенной). Другой пример связан с твёрдым телом: если один конец стержня нагреть или электрически зарядить, распространяется тепло (или соответственно электрический ток) от горячей (заряженной) части к холодной (незаряженной) части. В случае металлического стержня тепловая диффузия развивается быстро, а ток протекает почти мгновенно. Если стержень изготовлен из синтетического материала, тепловая диффузия протекает медленно, а диффузия электрически заряженных частиц — очень медленно. Диффузия молекул протекает в общем ещё медленнее. Например, если кусочек сахара опустить на дно стакана с водой и воду не перемешивать, то пройдёт несколько недель, прежде чем раствор станет однородным. Ещё медленнее происходит диффузия одного твёрдого вещества в другое. Например, если медь покрыть золотом, то будет происходить диффузия золота в медь, но при нормальных условиях (комнатная температура и атмосферное давление) золотосодержащий слой достигнет толщины в несколько микрометров только через несколько тысяч лет.
Все виды диффузии подчиняются одинаковым законам. Скорость диффузии пропорциональна площади поперечного сечения образца, а также разности концентраций, температур или зарядов (в случае относительно небольших величин этих параметров). Так, тепло будет в четыре раза быстрее распространяться через стержень диаметром в два сантиметра, чем через стержень диаметром в один сантиметр. Это тепло будет распространяться быстрее, если перепад температур на одном сантиметре будет 10 °C вместо 5 °C. Скорость диффузии пропорциональна также параметру, характеризующему конкретный материал. В случае тепловой диффузии этот параметр называется теплопроводность, в случае потока электрических зарядов — электропроводность. Количество вещества, которое диффундирует в течение определённого времени, и расстояние, проходимое диффундирующим веществом, пропорциональны квадратному корню времени диффузии.
Диффузия представляет собой процесс на молекулярном уровне и определяется случайным характером движения отдельных молекул. Скорость диффузии в связи с этим пропорциональна средней скорости молекул. В случае газов средняя скорость малых молекул больше, а именно она обратно пропорциональна квадратному корню из массы молекулы и растёт с повышением температуры. Диффузионные процессы в твёрдых телах при высоких температурах часто находят практическое применение. Например, в определённых типах электронно-лучевых трубок (ЭЛТ) применяется металлический торий, продиффундировавший через металлический вольфрам при 2000 °C.
3.2 Уравнение Фика
В большинстве практических случаев вместо химического потенциала применяется концентрация C. Прямая замена µ на C становится некорректной в случае больших концентраций, так как химический потенциал связан с концентрацией по логарифмическому закону. Если не рассматривать такие случаи, то выше приведённую формулу можно заменить на следующую:
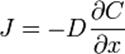
которая показывает, что плотность потока вещества J пропорциональна коэффициенту диффузии D и градиенту концентрации. Это уравнение выражает первый закон Фика (Адольф Фик — немецкий физиолог, установивший законы диффузии в 1855 г.). Второй закон Фика связывает пространственное и временное изменения концентрации (уравнение диффузии):
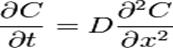
Коэффициент диффузии D зависит от температуры. В ряде случаев в широком интервале температур эта зависимость представляет собой уравнение Аррениуса.
Процессы диффузии имеют большое значение в природе:
Питание, дыхание животных и растений;
Проникновение кислорода из крови в ткани человека.
3.3 Пассивный транспорт
Пассивный транспорт – это перенос веществ из мест с большим значением электрохимического потенциала к местам с его меньшим значением.
При опытах с искусственными липидными бислоями было установлено, что чем меньше молекула и чем меньше она образует водородных связей, тем быстрее она дифундирует через мембрану. Итак, чем меньше молекула и чем более она жирорастворима (гидрофобна или неполярна), тем быстрее она будет проникать через мембрану. Диффузия веществ через липидный бислой вызывается градиентом концентрации в мембране. Через липидные и белковые поры сквозь мембрану проникают молекулы нерастворимых в липидах веществ и водорастворимые гидратированные ионы (окруженные молекулами воды). Малые неполярные молекулы легко растворимы и быстро диффундируют. Незаряженные полярные молекулы при небольших размерах также растворимы и диффундируют.
Важно, что вода очень быстро проникает через липидный бислой несмотря на то, что она относительно нерастворима в жирах. Это происходит из-за того, что ее молекула мала и электрически нейтральна.
Осмос – преимущественное движение молекул воды через полупроницаемые мембраны (непроницаемые для растворенного вещества и проницаемые для воды) из мест с меньшей концентрацией растворенного вещества в места с большей концентрацией. Осмос – по сути дела, простая диффузия воды из мест с ее большей концентрацией, в места с меньшей концентрацией воды. Осмос играет большую роль во многих биологических явлениях. Явление осмоса обусловливает гемолиз эритроцитов в гипотонических растворах.
Итак, мембраны могут пропускать воду и неполярные молекулы за счет простой диффузии.
3.3.1 Отличия облегченной диффузии от простой:
1) перенос вещества с участием переносчика происходит значительно быстрее;
2) облегченная диффузия обладает свойством насыщения: при увеличении концентрации с одной стороны мембраны плотность потока вещества возрастает лишь до некоторого предела, когда все молекулы переносчика уже заняты;
3) при облегченной диффузии наблюдается конкуренция переносимых веществ в тех случаях, когда переносчиком переносятся разные вещества; при этом одни вещества переносятся лучше, чем другие, и добавление одних веществ затрудняет транспорт других; так из сахаров глюкоза переносится лучше, чем фруктоза, фруктоза лучше, чем ксилоза, а ксилоза лучше, чем арабиноза и. т. д.;
4) есть вещества, блокирующие облегченную диффузию – они образуют прочный комплекс с молекулами переносчика, например, флоридзин подавляет транспорт сахаров через биологическую мембрану.
4.Закон Дарси
Закон Дарси (Анри Дарси, 1856) — закон фильтрации жидкостей и газов в пористой среде. Получен экспериментально. Выражает зависимость скорости фильтрации флюида от градиента напора:
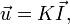
где: — скорость фильтрации, K — коэффициент фильтрации, — градиент напора. Закон Дарси связан с несколькими системами измерений. Среда с проницаемостью 1 Дарси (Д) позволяет протекать 1 см³/с жидкости или газа с вязкостью 1 сп (мПа·с) под градиентом давления 1 атм/см, действующего на площадь 1 см². 1 миллидарси (мД) равен 0,001 Дарси.
В системе измерения СИ, 1 Дарси эквивалентен 9,869233×10−13м² или 0,9869233 мкм². Такое преобразование обычно аппроксимируется как 1 мкм². Следует заметить, что это число, обратное к 1,013250 — коэффициент преобразования из атмосфер в бары.
Транспорт сквозь липидный бислой (простая диффузия) и транспорт при участии мембранных белков
5. Активный транспорт
Другие белки-переносчики (их иногда называют белки-насосы) переносят через мембрану вещества с затратами энергии, которая обычно поставляется при гидролизе АТФ. Этот вид транспорта осуществляется против градиента концентрации переносимого вещества и называется активным транспортом.
Симпорт, антипорт и унипорт
Мембранный транспорт веществ различается также по направлению их перемещения и количеству переносимых данным переносчиком веществ:
1) Унипорт - транспорт одного вещества в одном направлении в зависимости от градиента
2) Симпорт - транспорт двух веществ в одном направлении через один переносчик.
3) Антипорт - перемещение двух веществ в разных направлениях через один переносчик.
Унипорт осуществляет, например, потенциал-зависимый натриевый канал, через который в клетку во время генерации потенциала действия перемещаются ионы натрия.
Симпорт осуществляет переносчик глюкозы, расположенный на внешней (обращенной в просвет кишечника) стороне клеток кишечного эпителия. Этот белок захватывает одновременно молекулу глюкозы и ион натрия и, меняя конформацию, переносит оба вещества внутрь клетки. При этом используется энергия электрохимического градиента, который, в свою очередью создается за счет гидролиза АТФ натрий-калиевой АТФ-азой.
Антипорт осуществляет, например, натрий–калиевая АТФаза (или натрий–зависимая АТФаза). Она переносит в клетку ионы калия. а из клетки - ионы натрия.
Работа натрий-калиевой АТФазы как пример антипорта и активного транспорта
Первоначально этот переносчик присоединяет с внутренней стороны мембраны три иона Na + . Эти ионы изменяют конформацию активного центра АТФазы. После такой активации АТФаза способна гидролизовать одну молекулу АТФ, причем фосфат-ион фиксируется на поверхности переносчика с внутренней стороны мембраны.
Выделившаяся энергия расходуется на изменение конформации АТФазы, после чего три иона Na + и ион (фосфат) оказываются на внешней стороне мембраны. Здесь ионы Na + отщепляются, а замещается на два иона K + . Затем конформация переносчика изменяется на первоначальную, и ионы K + оказываются на внутренней стороне мембраны. Здесь ионы K + отщепляются, и переносчик вновь готов к работе.
Более кратко действия АТФазы можно описать так:
1) Она изнутри клетки "забирает" три иона Na + ,затем расщепляет молекулу АТФ и присоединяет к себе фосфат
2) "Выбрасывает" ионы Na + и присоединяет два иона K + из внешней среды.
3) Отсоединяет фосфат, два иона K + выбрасывает внутрь клетки
В итоге во внеклеточной среде создается высокая концентрация ионов Na + , а внутри клетки - высокая концентрация K + . Работа Na + , K + - АТФаза создает не только разность концентраций, но и разность зарядов (она работает как электрогенный насос). На внешней стороне мембраны создается положительный заряд, на внутренней - отрицательный.
6. Строение и функции ионных каналов.
Модель возбудимой мембраны предполагает регулируемый перенос ионов калия и натрия через мембрану. Однако, непосредственный переход иона через липидный бислой весьма затруднен, поэтому плотность потока ионов была бы очень мала, если бы ион проходил непосредственно через липидную фазу мембраны. Это и ряд других соображений дали основание считать, что в мембране должны быть некоторые специальные структуры – проводящие ионы.
Такие структуры были найдены и названы ионными каналами. Подобные каналы выделены из различных объектов: плазматической мембраны клеток, постсинаптической мембраны мышечных клеток и других объектов. Известны также ионные каналы, образованные антибиотиками.
Основные свойства ионных каналов:
1) селективность;
2) независимость работы отдельных каналов;
3) дискретный характер проводимости;
4) зависимость параметров каналов от мембранного потенциала.
Рассмотрим их по порядку.
1. Селективностью называют способность ионных каналов избирательно пропускать ионы какого-либо одного типа.
Еще в первых опытах на аксоне кальмара было обнаружено, что ионы натрия и калия по-разному влияют на мембранный потенциал. Ионы калия меняют потенциал покоя, а ионы натрия - потенциал действия.
Измерения показали, что ионные каналы обладают абсолютной селективностью по отношению к катионам (катион-селективные каналы), либо к анионам (анион-селективные каналы). В то же время через катион-селективные каналы способны проходить различные катионы различных химических элементов, но проводимость мембраны для неосновного иона, а значит и ток через нее, будет существенно ниже, например, для натриевого канала калиевый ток через него будет в 20 раз меньше. Способность ионного канала пропускать различные ионы называется относительной селективностью и характеризуется рядом селективности - соотношением проводимостей канала для разных ионов, взятых при одной концентрации.
2. Независимость работы отдельных каналов. Прохождение тока через отдельный ионный канал не зависит от того, идет ли ток через другие каналы. Например, калиевые каналы могут быть включены или выключены, но ток через натриевые каналы не меняется. Влияние каналов друг на друга происходит опосредованно: изменение проницаемостей каких-либо каналов (например натриевых) меняет мембранный потенциал, а уже он влияет на проводимости прочих ионных каналов.
3. Дискретный характер проводимости ионных каналов. Ионные каналы представляют собой субъединичный комплекс белков, пронизывающий мембрану. В центре его существует трубка, сквозь которую могут проходить ионы.
Количество ионных каналов на 1 мкм поверхности мембраны определяли с помощью радиоактивно-меченного блокатора натриевых каналов - тетродотоксина. Известно, что одна молекула ТТХ связывается только с одним каналом. Тогда измерение радиоактивности образца с известной площадью позволило показать, что на 1 мкм аксона кальмара находится около 500 натриевых каналов. Впервые это было обнаружено в 1962 г. в исследованиях проводимости бислойных липидных мембран (БЛМ) при добавлении в раствор, омывающий мембрану, микроколичеств некоторого вещества, индуцировавшего возбуждение. На БЛМ подавали постоянное напряжение и регистрировали ток. Запись тока во времени имела вид скачков между двумя проводящими состояниями.
Результаты экспериментов выполненных на различных ионных каналах показали, что проводимость ионного канала дискретна и он может находиться в двух состояниях: открытом или закрытом. Выбросы тока обусловлены одновременным открытием 2-х или 3-х каналов. Переходы между состояниями ионного канала происходят в случайные моменты времени и подчиняются статистическим закономерностям. Нельзя сказать, что данный ионный канал откроется именно в этот момент времени. Можно лишь сделать утверждение о вероятности открывания канала в определенном интервале времени.
Ионные каналы описывают характерными временами жизни открытого и закрытого состояний.
4. Зависимость параметров канала от мембранного потенциала. Ионные каналы нервных волокон чувствительны к мембранному потенциалу, например натриевый и калиевый каналы аксона кальмара. Это проявляется в том, что после начала деполяризации мембраны соответствующие токи начинают изменяться с той или иной кинетикой. На языке "ионных каналов" этот процесс происходит следующим образом. Ион-селективный канал имеет так называемый
"сенсор" -некоторый элемент своей конструкции, чувствительный к действию электрического поля (см. рисунок). При изменении мембранного потенциала меняется величина действующей на него силы, в результате эта часть ионного канала перемещается и меняет вероятность открывания или закрывания "ворот" – своеобразных заслонок, действующих по закону "все или ничего".
Структура ионного канала
Ион-селективный канал состоит из следующих частей погруженной в бислой белковой части, имеющей субъединичное строение; селективного фильтра, образованного отрицательно заряженными атомами кислорода, которые жестко расположены на определенном расстоянии друг от друга и пропускают ионы только определенного диаметра; воротной части.
"Ворота" ионного канала управляются мембранным потенциалом и могут находиться как в закрытом состоянии (штриховая линия), так и в открытом состоянии (сплошная линия). Нормальное положение ворот натриевого канала – закрытое. Под действием электрического поля увеличивается вероятность открытого состояния, ворота открываются и поток гидратированных ионов получает возможность проходить сквозь селективный фильтр.
Если ион "подходит" по диаметру, то он сбрасывает гидратную оболочку и проскакивает на другую сторону ионного канала. Если же ион слишком велик по диаметру, как например, тетраэтиламмоний, он не в состоянии пролезть сквозь фильтр и не может пересечь мембрану. Если же, напротив, ион слишком мал, то у него возникают сложности в селективном фильтре, на сей раз связанные с трудностью сбросить его гидратную оболочку. У "подходящего" иона сброшенная вода замещается на связи с атомами кислорода, расположенными в фильтре, у "неподходящего" иона стерическое соответствие хуже. Поэтому ему труднее пройти через фильтр и проводимость канала для него ниже.
Блокаторы ионных каналов либо не могут пройти сквозь него, застревая в фильтре, либо, если это большие молекулы как ТТХ, они стерически соответствуют какому-либо входу в канал. Так как блокаторы несут положительный заряд, их заряженная часть втягивается в канал к селективному фильтру как обычный катион, а макромолекула закупоривает его.
Таким образом, изменения электрических свойств возбудимых биомембран осуществляется с помощью ионных каналов. Это белковые макромолекулы, пронизывающие липидный бислой, которые могут находиться в нескольких дискретных состояниях. Свойства каналов, селективных для ионов калия, натрия и кальция могут по-разному зависеть от мембранного потенциала, что и определяет динамику потенциала действия в мембране, а также отличия таких потенциалов в мембранах разных клеток.
Заключение
Любая молекула может пройти через липидный бислой, однако скорость пассивной диффузии веществ, т.е. перехода вещества из области с большей концентрацией в область с меньшей, может сильно отличаться. Для некоторых молекул это занимает столь длительное время, что можно говорить об их практической непроницаемости для липидного бислоя мембраны. Скорость диффузии веществ через мембрану зависит главным образом от размера молекул и их относительной растворимости в жирах.
Легче всего проходят простой диффузией через липидную мембрану малые неполярные молекулы, такие как О2, стероиды, тиреоидные гормоны, а также жирные кислоты. Малые полярные незаряженные молекулы - СО2, NH3, Н2О, этанол, мочевина - также диффундируют с достаточно большой скоростью. Диффузия глицерола идёт значительно медленнее, а глюкоза практически не способна самостоятельно пройти через мембрану. Для всех заряженных молекул, независимо от размера, липидная мембрана непроницаема.
Транспорт таких молекул возможен благодаря наличию в мембранах либо белков, формирующих в липидном слое каналы (поры), заполненные водой, через которые могут проходить вещества определённого размера простой диффузией, либо специфических белков-переносчиков, которые избирательно взаимодействуя с определёнными лигандами, облегчают их перенос через мембрану (облегчённая диффузия).
Кроме пассивного транспорта веществ, в клетках есть белки, активно перекачивающие определённые растворённые в воде вещества против их градиента, т.е. из меньшей концентрации в область большей. Этот процесс, называемый активным транспортом, осуществляется всегда с помощью белков-переносчиков и происходит с затратой энергии.
Наружная часть канала сравнительно доступна для изучения, исследование внутренней части представляет значительные трудности. П. Г. Костюком был разработан метод внутриклеточного диализа, который позволяет изучать функцию входных и выходных структур ионных каналов без применения микроэлектродов. Оказалось, что часть ионного канала, открытая во внеклеточное пространство, по своим функциональным свойствам отличается от части канала, обращенной во внутриклеточную среду.
Именно ионные каналы обеспечивают два важных свойства мембраны: селективность и проводимость.
Селективность, или избирательность, канала обеспечивается его особой белковой структурой. Большинство каналов являются электроуправляемыми, т. е. их способность проводить ионы зависит от величины мембранного потенциала. Канал неоднороден по своим функциональным характеристикам, особенно это касается белковых структур, находящихся у входа в канал и у его выхода (так называемые воротные механизмы).
Уравнение Фика
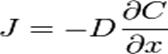
Знак «–» показывает, что суммарная плотность потока вещества при диффузии направлена в сторону уменьшения плотности, D –коэффициент диффузии. Формула показывает, что плотность потока вещества J пропорциональна коэффициенту диффузии D и градиенту концентрации. Это уравнение выражает первый закон Фика (Адольф Фик — немецкий физиолог, установивший законы диффузии в 1855 г.).
Ион-селективный канал состоит из следующих частей погруженной в бислой белковой части, имеющей субъединичное строение; селективного фильтра, образованного отрицательно заряженными атомами кислорода, которые жестко расположены на определенном расстоянии друг от друга и пропускают ионы только определенного диаметра; воротной части. Именно ионные каналы обеспечивают два важных свойства мембраны: селективность и проводимость. Кальциевые каналы играют существенную роль в клетках сердца.
Список литературы
1. Сайт http://www.biochemistry.ru/biohimija_severina/.html
2. Ю. И. Афанасьев, Н. А. Юрина, Е. Ф. Котовский и др. Гистология. М.
Медицина, 1999.
3. Сайт http://www.bibliotekar.ru/447/7.htm
4. Филлиппович Ю.Б. Основы биохимии. М., Высшая школа, 1985.Диффузия
5. Басниев К. С., Кочина Н. И., Максимов М. В. Подземная гидромеханика. // М.: Недра, 1993, с. 41—43
6. Геннис Р. Биомембраны. Молекулярная структура и функции. М., Мир, 1997
|
