КОНТРОЛЬНАЯ РАБОТА
«
Концепции современного естествознания
»
Тема: Гравитация и электродинамика. Организация живой материи. Каталитические реакции
Оглавление
1 Гравитационное и электромагнитное взаимодействия
2 Уровни организации живой материи
1.1 Молекулярный
1.2 Субклеточный
1.3 Клеточный
1.4 Органотканевый
1.5 Организменный
1.6 Популяционно-видовой
1.7 Биоценотический, биогеоценотический
1.8 Биосферный
3 Пример нескольких каталитических реакций. Принцип действия катализатора
Используемая литература
Фундаментальные взаимодействия — различные, не сводящиеся друг к другу типы взаимодействия элементарных частиц и составленных из них тел. На сегодня достоверно известно существование четырех фундаментальных взаимодействий: гравитационного, электромагнитного, сильного и слабого взаимодействий, причём электромагнитное и слабое взаимодействия, вообще говоря, являются проявлениями единого электрослабого взаимодействия. Ведутся поиски других типов взаимодействий, как в явлениях микромира, так и в космических масштабах, однако пока существование какого-либо другого типа взаимодействия не обнаружено.
Электромагнитное взаимодействие — одно из четырёх фундаментальных взаимодействий. Электромагнитное взаимодействие существует между частицами, обладающими электрическим зарядом. С современной точки зрения электромагнитное взаимодействие между заряженными частицами осуществляется не прямо, а только посредством электромагнитного поля.
С точки зрения квантовой теории поля электромагнитное взаимодействие переносится безмассовым бозоном — фотоном (частицей, которую можно представить как квантовое возбуждение электромагнитного поля). Сам фотон электрическим зарядом не обладает, а значит не может непосредственно взаимодействовать с другими фотонами.
Из фундаментальных частиц в электромагнитном взаимодействии участвуют также имеющие электрический заряд частицы: кварки, электрон, мюон и тау-частица (из фермионов), а также заряженые калибровочные бозоны.
Электромагнитное взаимодействие отличается от слабого и сильного взаимодействия своим дальнодействующим характером — сила взаимодействия между двумя зарядами спадает только как вторая степень расстояния (см.: закон Кулона). По такому же закону спадает с расстоянием гравитационное взаимодействие. Электромагнитное взаимодействие заряженных частиц намного сильнее гравитационного, и единственная причина, по которой электромагнитное взаимодействие не проявляется с большой силой на космических масштабах — электрическая нейтральность материи, то есть наличие в каждой области Вселенной с высокой степенью точности равных количеств положительных и отрицательных зарядов.
Реклама

В классических (неквантовых) рамках электромагнитное взаимодействие описывается классической электродинамикой.
Краткая сводка основных формул классической электродинамики
На проводник с током, помещенный в магнитное поле, действует сила Ампера:
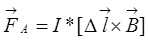
На заряженную частицу, движущуюся в магнитном поле, действует сила Лоренца:
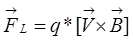
Гравита́ция (всеми́рное тяготе́ние, тяготе́ние) (от лат. gravitas — «тяжесть») — дальнодействующее фундаментальное взаимодействие, которому подвержены все материальные тела. По современным представлениям, является универсальным взаимодействием материи с пространственно-временным континуумом, и, в отличие от других фундаментальных взаимодействий, всем без исключения телам, независимо от их массы и внутренней структуры, в одной и той же точке пространства и времени придаёт одинаковое ускорение относительно локально-инерциальной системы отсчёта — принцип эквивалентности Эйнштейна. Главным образом, определяющее влияние гравитация оказывает на материю в космических масштабах. Термин гравитация используется также как название раздела физики, изучающего гравитационное взаимодействие. Наиболее успешной современной физической теорией в классической физике, описывающей гравитацию, является общая теория относительности, квантовая теория гравитационного взаимодействия пока не построена.
Гравитационное взаимодействие — одно из четырёх фундаментальных взаимодействий в нашем мире. В рамках классической механики, гравитационное взаимодействие описывается законом всемирного тяготения Ньютона, который гласит, что сила гравитационного притяжения между двумя материальными точками массы m1 и m2, разделёнными расстоянием R, пропорциональна обеим массам и обратно пропорциональна квадрату расстояния — то есть

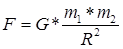
Здесь G — гравитационная постоянная, равная примерно 6,6725 *10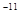 м³/(кг•с²). м³/(кг•с²).
Закон всемирного тяготения — одно из приложений закона обратных квадратов, встречающегося так же и при изучении излучений, и являющимся прямым следствием квадратичного увеличения площади сферы при увеличении радиуса, что приводит к квадратичному же уменьшению вклада любой единичной площади в площадь всей сферы.
Реклама

Поле тяжести потенциально. Это значит, что можно ввести потенциальную энергию гравитационного притяжения пары тел, и эта энергия не изменится после перемещения тел по замкнутому контуру. Потенциальность поля тяжести влечёт за собой закон сохранения суммы кинетической и потенциальной энергии и при изучении движения тел в поле тяжести часто существенно упрощает решение. В рамках ньютоновской механики гравитационное взаимодействие является дальнодействующим. Это означает, что как бы массивное тело ни двигалось, в любой точке пространства гравитационный потенциал зависит только от положения тела в данный момент времени.
Большие космические объекты — планеты, звезды и галактики имеют огромную массу и, следовательно, создают значительные гравитационные поля.
Гравитация — слабейшее взаимодействие. Однако, поскольку оно действует на любых расстояниях и все массы положительны, это тем не менее очень важная сила во Вселенной. Для сравнения: полный электрический заряд этих тел ноль, так как вещество в целом электрически нейтрально.
Также гравитация, в отличие от других взаимодействий, универсальна в действии на всю материю и энергию. Не обнаружены объекты, у которых вообще отсутствовало бы гравитационное взаимодействие.
Из-за глобального характера гравитация ответственна и за такие крупномасштабные эффекты, как структура галактик, черные дыры и расширение Вселенной, и за элементарные астрономические явления — орбиты планет, и за простое притяжение к поверхности Земли и падения тел.
Гравитация была первым взаимодействием, описанным математической теорией. Аристотель считал, что объекты с разной массой падают с разной скоростью. Только много позже Галилео Галилей экспериментально определил, что это не так — если сопротивление воздуха устраняется, все тела ускоряются одинаково. Закон всеобщего тяготения Исаака Ньютона (1687) хорошо описывал общее поведение гравитации. В 1915 году Альберт Эйнштейн создал Общую теорию относительности, более точно описывающую гравитацию в терминах геометрии пространства-времени.
Cложившееся к 60-м гг. 20 в. представление о структурности живого. Жизнь на Земле представлена индивидуумами определённого строения, принадлежащими к определённым систематическим группам, а также сообществами разной сложности. Индивидуумы обладают молекулярной, клеточной, тканевой, органной структурностью; сообщества бывают одновидовые и многовидовые. Индивидуумы и сообщества организованы в пространстве и во времени. По подходу к их изучению можно выделить неск. основных У. о. ж. м. на базе разных способов структурно-функционального объединения составляющих элементов: молекулярный, субклеточный, клеточный, органотканевый, организменный, популяционно-видовой, биоценотический, биогеоценотический, биосферный.
Любая живая система, как бы сложно она ни была организована, состоит из биологических макромолекул: нуклеиновых кислот, белков, полисахаридов, а также других важных органических веществ. С этого уровня начинаются разнообразные процессы жизнедеятельности организма: обмен веществ и превращение энергии, передача наследственной информации и др.
Молекулярный уровень составляет предмет молекулярной биологии, изучающей строение белков, их функции как ферментов или элементов цитоскелета, роль нуклеиновых кислот в хранении, репликации и реализации генетической информации, т. е. процессы синтеза ДНК, РНК и белков. На этом уровне достигнуты большие практические успехи в области биотехнологии и генной инженерии.
На уровне субклеточных, или надмолекулярных, структур изучают строение и функции органоидов (хромосом, митохондрий, рибосом и др.), а также др. включений клетки.
2.3 Клеточный
Клетка - структурная и функциональная единица, а также единица развития всех живых организмов, обитающих на Земле. На клеточном уровне сопрягаются передача информации и превращение веществ и энергии
Особый У. о. ж. м.— клеточный; биология клетки (цитология) — один из основных разделов современной биологии, включает проблемы морфологической организаций клетки, специализации клеток в ходе развития, функций клеточной мембраны, механизмов и регуляции деления клетки. Эти проблемы имеют особенно важное значение для медицины, в частности, составляя основу проблемы рака.
Изучение клеток, выступающих в роли самостоятельных организмов (бактерии, простейшие и некоторые другие организмы) и клеток, составляющих многоклеточные организмы.
Клетки, имеющие общее происхождение и выполняющие сходные функции, образуют ткани. Выделяют несколько типов животных и растительных тканей, обладающих различными свойствами.
У организмов, начиная с кишечнополостных, формируются органы (системы органов), часто из тканей различных типов.
Клетки многоклеточных организмов образуют ткани - системы сходных по строению и функциям клеток и связанных с ними межклеточных веществ. Ткани интегрируются в более крупные функциональные единицы, называемые органами. Внутренние органы характерны для животных; здесь они входят в состав систем органов (дыхательной, нервной и пр.). Например, система органов пищеварения - полость рта, глотка, пищевод, желудок, двенадцатиперстная кишка, тонкая кишка, толстая кишка, заднепроходное отверстие. Подобная специализация, с одной стороны, улучшает работу организма в целом, а с другой - требует повышения степени координации и интеграции различных тканей и органов.
На органотканевом уровне основные проблемы заключаются в изучении особенностей строения и функций отдельных органов и составляющих их тканей.
Этот уровень представлен одноклеточными и многоклеточными организмами.
Элементарной единицей организменного уровня служит особь, которая рассматривается в развитии - от момента зарождения до прекращения существования - как живая система. Возникают системы органов, специализированных для выполнения различных функций.
Совокупность организмов одного и того же вида, объединенная общим местом обитания, в которой создается популяция - надорганизменная система. В этой системе осуществляются элементарные эволюционные преобразования.
На организменном уровне изучают особь и свойственные ей как целому черты строения, физиол. процессы, в т. ч. дифференцировку, механизмы адаптации (акклимации) и поведения, в частности — нейрогумоарльные механизмы регуляции, функции ЦНС.
2.6 Популяционно-видовой
Организмы одного и того же вида, совместно обитающие в определенных ареалах, составляют популяцию. Сейчас на Земле насчитывают около 500 тыс. видов растений и около 1,5 млн. видов животных.
На популяционно-видовом уровне изучают факторы, влияющие на численность популяций, проблемы сохранения исчезающих видов, динамики генетического состава популяций, действие факторов микроэволюции и т. д. Для хозяйственной деятельности человека важны такие проблемы популяционной биологии, как контроль численности видов, наносящих ущерб хозяйству, поддержание оптимальной численности эксплуатируемых и охраняемых популяций.
Представлен совокупностью организмов разных видов, в той или иной степени зависящих друг от друга.
Биогеоценоз - совокупность организмов разных видов и различной сложности организации с факторами среды их обитания. В процессе совместного исторического развития организмов разных систематических групп образуются динамичные, устойчивые сообщества.
На биогеоценотическом и биоценотическом уровнях ведущими являются проблемы взаимоотношений организмов в биоценозах, условия, определяющие их численность и продуктивность биоценозов, устойчивость последних и роль влияний человека на сохранение биоценозов и их комплексов.
Высшая форма организации живого. Включает все биогеоценозы, связанные общим обменом веществ и превращением энергии.
Биосфера - совокупность всех биогеоценозов, система, охватывающая все явления жизни на нашей планете. На этом уровне происходит круговорот веществ и превращение энергии, связанные с жизнедеятельностью всех живых организмов.
На биосферном уровне современная биология решает глобальные проблемы, например, определение интенсивности образования свободного кислорода растительным покровом Земли или изменения концентрации углекислого газа в атмосфере, связанного с деятельностью человека.
Разделение живой материи и проблем биологии по уровням организации хотя и отражает объективную реальность, но в то же время является условным, т. к. почти все конкретные задачи биологии касаются одновременно нескольких уровней, а нередко и всех сразу. Например, проблемы эволюции или онтогенеза не могут рассматриваться только на уровне организма, т. е. без молекулярного, субклеточного, клеточного, органотканевого, а также популяционно-видового и биоценотического уровней; проблема регуляции численности опирается на молекулярный уровень, но касается также всех вышестоящих, включая такие аспекты, как, например, загрязнение всей биосферы. По наличию специфических элементарных единиц и явлений считается достаточным выделение 4 основных У. о. ж. м. (табл.). Представление об У. о. ж. м. наглядно отражает системный подход в изучении живой природы
3 Пример нескольких каталитических реакций. Принцип действия катализатора
Для того чтобы прошла химическая реакция, нужно, чтобы молекулы столкнулись. Из кинетической энергии таких столкновений можно почерпнуть энергию, необходимую для того, чтобы разорвать или ослабить химические связи в молекулах реагентов. В результате теплового движения за 1 с. происходят триллионы столкновений молекул, лишь редкие из которых приводят к химическому превращению. То есть реагируют только те молекулы, которые в момент столкновения обладают достаточной суммарной энергией. Эта энергия, называемая энергией активации (Еа), характеризует ту минимальную энергию, которой должна обладать молекула (или молекулы), чтобы вступить в химическую реакцию. Графически эта величина соответствует величине барьера (образование переходного состояния), который необходимо преодолеть для осуществления химической реакции. В результате многие даже термодинамически разрешенные (выгодные) реакции практически не идут из-за слишком высокой энергии активации.
Вспомним реакцию окисления аммиака, который способен гореть в чистом кислороде (с огромным трудом на воздухе) с образованием азота и паров воды. Реакция идет только при высокой температуре. Добавление же порошка оксида хрома (Cr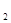 O O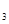 ) приводит к "огненному дождю" из этих частиц. Идет реакция (экзотермическая, то есть с выделением теплоты), продукты которой содержат NO и воду. Оксид хрома, являющийся в реакции катализатором, не изменяется, но направляет реакцию по энергетически более выгодному пути с меньшей энергией активации, при более низкой температуре в данном случае. ) приводит к "огненному дождю" из этих частиц. Идет реакция (экзотермическая, то есть с выделением теплоты), продукты которой содержат NO и воду. Оксид хрома, являющийся в реакции катализатором, не изменяется, но направляет реакцию по энергетически более выгодному пути с меньшей энергией активации, при более низкой температуре в данном случае.
Важно отметить, что катализатор не расходуется и не изменяет разности свободных энергий начального и конечного состояний реакции, то есть не влияет на общие термодинамические характеристики реакции, в частности не смещает ее равновесие (изменяя лишь время выхода в равновесное состояние). Действие катализатора заключается в понижении энергии активации и означает, что большая доля сталкивающихся молекул будет обладать достаточной энергией для преодоления барьера свободной энергии переходного состояния и протекания реакции.
За счет чего катализатор может понижать барьер свободной энергии (или свободную энергию переходного состояния)? Он может взаимодействовать с реагентами, давая принципиально иное переходное состояние или просто более стабильное (и, следовательно, с более низкой свободной энергией), чем образуемое в некаталитической реакции.
Рассмотрим некоторые основные механизмы катализа. Все они могут быть обнаружены при изучении действия ферментов как катализаторов. Пример - реакция гидролиза сложного эфира:
R'COOR" + HO R'COOH + R"OH.
Эта реакция включает нуклеофильную атаку свободной парой электронов кислорода воды по углеродному остатку карбонила (имеет частичный положительный заряд d+ в результате оттягивания электронной плотности на кислородный атом), образование переходного состояния и далее продуктов реакции.
В образующемся переходном состоянии атакующая молекула воды приобретает положительный заряд, а кислород карбонильной группы - отрицательный. Такое переходное состояние крайне невыгодно, то есть его образование требует высокой энергии активации. Это, в свою очередь, означает, что скорость реакции будет очень мала.
Использование катализатора в данном случае может существенно помочь процессу. Каким образом?
1. Кислотно-основный катализ (под действием Н+ или ОН-);
а) кислоты могут временно давать протон молекуле эфира:
Протонированная форма эфира (более реакционноспособная) затем атакуется молекулой воды аналогично схеме без катализатора с той лишь разницей, что переходное состояние недвухзарядное (более стабильное, чем предыдущее), следовательно, для такой реакции требуется меньшая энергия активации.
б) основания (ХО-) могут временно акцептировать протон (Н+), помогая стабилизировать двухзарядное переходное состояние, например таким образом:
2. Электростатический катализ. Переходное состояние можно стабилизировать электрическим полем иона, например положительно заряженный карбоний-ион (переходное состояние в случае катализа ферментом - лизоцимом) стабилизируется электрическим полем отрицательно заряженной карбоксильной группы остатка аспарагиновой кислоты. Конечно, энергия электростатического взаимодействия двух точечных зарядов зависит от свойств среды (диэлектрической постоянной), в которой это взаимодействие происходит. В водной среде это взаимодействие слабое, однако в органических растворителях, а также в активных центрах ферментов оно может вносить существенный вклад.
3. Ковалентный катализ (электрофильный или нуклеофильный). Роль ионов меди в предыдущем примере, помогающих оттянуть электроны из реакционного центра, можно обсуждать как форму электрофильного катализа.
Катализатор нуклеофильной природы, например третичный амин, имеющий неподеленную пару электронов на атоме азота, казалось бы, ничем не отличается от исходного реагента некаталитической реакции, в данном случае молекулы воды.
Более того, в этом случае мы видим и двухзарядное переходное состояние. Тем не менее реакция в присутствии катализатора идет, а при отсутствии его практически нет. Причина в том, что катализатор имеет более сильно выраженный нуклеофильный характер, чем атакующая группа (а следовательно, действует быстрее), а промежуточное соединение более реакционноспособное, чем исходное. Эффект повышения нуклеофильной способности тех или иных групп весьма ярко проявляется в ферментативном катализе. Более того, фермент может одновременно использовать несколько различных механизмов катализа, повышая эффективность химической реакции.
Действие ферментов, как и других катализаторов, как уже сказано выше, заключается в понижении свободной энергии активации реакции. Однако картина протекания реакции в присутствии фермента (профиль изменения свободной энергии) выглядит сложнее, чем для обычных катализаторов.
Огромную роль в катализе ферментами играет то, что, собственно, происходит до начала химической реакции, а именно образование так называемого фермент-субстратного комплекса Почему? Дело в том, что в этом случае реагирующие частицы оказываются сближенными и сориентированными до начала собственно химической реакции. Высокий барьер свободной энергии разбивается на несколько меньших, первый из которых характеризует неизбежные энергетические потери при сближении и ориентации молекул, связанные с затормаживанием их поступательного и вращательного движений. То есть в ферментативном катализе осуществляется перевод реакции во внутримолекулярный режим. Что дает такой перевод, отражает приведенный в качестве примера гидролиз аспирина.
4. Внутримолекулярный катализ. Гидролиз эфирной связи в случае аспирина ускоряется за счет внутримолекулярного общеосновного катализа.
Оказывается, что простой перевод реакции гидролиза эфирной связи во внутримолекулярный режим приводит к увеличению ее скорости в 100 раз. Как и за счет чего осуществляется такой перевод в ферментах?
Ферменты, как и все белки, строятся из аминокислот, в пространстве они организованы (свернуты) особым образом (третичная и четвертичная структуры белка). Высокая каталитическая активность ферментов обеспечивается функционированием специального участка - сложноорганизованного активного центра, в состав которого входят аминокислотные остатки, часто весьма удаленные друг от друга в первичной полипептидной последовательности.
Выше мы выяснили, что для успешного нуклеофильного катализа необходимо, чтобы катализатор был сильным нуклеофилом, по крайней мере более сильным, чем исходный реагент. В то же время химические группы, формирующие активный центр фермента, сами по себе, как правило, являются слабыми катализаторами соответствующих реакций. И тем не менее ферментативный катализ по своей эффективности превосходит все известные катализаторы. Таким образом, действие фермента представляет собой хорошо отлаженную в пространстве и времени систему.
Как уже отмечалось, перевод реакции во внутримолекулярный режим приводит к ее ускорению.
Следует отметить также, что фермент, и в частности его активный центр, - это не некое застывшее образование. В ходе взаимодействия с субстратом и химической реакции в молекуле фермента могут происходить структурные (конформационные) изменения, которые удается в некоторых случаях зафиксировать и которые свидетельствуют о неком "настраивании" активного центра.
Таким образом, общие принципы катализа применимы и к действию ферментов. Однако различные факторы, способствующие протеканию реакции, в ферментативном катализе действуют согласованно, а во многих случаях являются суммарным результатом ряда причин. Важнейшие из них: 1) многоточечное взаимодействие с субстратом, приводящее к сближению и правильной ориентации реагирующих групп; 2) перевод реакции во внутримолекулярный режим; 3) стабилизация переходного состояния реакции.
1. Горелов А.А. Концепции современного естествознания. М.: Центр, 2003.
2. Диксон М., Уэбб Э. Ферменты. М.: Мир, 1982. Т. 1-3.
3. Лавриненко В.Н. Концепции современного естествознания. М.: ЮНИТИ-ДАНА, 2006.
4. Рузавин Г.И. Концепции современного естествознания. М.: ЮНИТИ, 2001.
5. Фершт Э. Структура и механизм действия ферментов. М.: Мир, 1980. 432 с.
6. http://ru.wikipedia.org/wiki/Фундаментальные_взаимодействия
7. http://www.pereplet.ru/obrazovanie/stsoros/280.html
|
