АВТОРЕФЕРАТ
диссертации на соискание ученой степени доктора биологических наук
ОЦЕНКА СТРАТЕГИЙ РАЗМНОЖЕНИЯ И ПОДДЕРЖАНИЯ ЖИЗНЕСПОСОБНОСТИ ООМИЦЕТА
PHYTOPHTHORA
INFESTANS
В СВЯЗИ С СОВРЕМЕННЫМИ МЕТОДАМИ ЗАЩИТЫ КАРТОФЕЛЯ И ТОМАТА ОТ ФИТОФТОРОЗА
1. Общая характеристика работы
Назначение и структура работы
Актуальность работы.
Фитофтороз остается одной из самых опасных болезней картофеля и томата, не смотря на многолетние разноплановые мероприятия по их защите. Прямые потери урожая картофеля и томата от фитофтороза в полевых условиях достигают до 70 и 100% соответственно. По данным Института Картофелеводства НАН Беларуси и ВНИИКХ имени А.Г. Лорха за период с 1924 по 2009 г. 41 год из 85 лет характеризовались высоким развитием фитофтороза. Прямые потери картофеля от фитофтороза достигают 4 млн. тонн (около 15% по урожаю, до 40 млн. рублей в стоимостном отношении). Потери томата от фитофтороза в Средней полосе России достигли критических величин. И ЛПХ, и хозяйства за последние годы отказываются выращивать томат из-за «побурения» плодов.
С середины 1990‑ых годов наблюдается последующее усиление вредоносности фитофтороза. Это часто связывают с появлением половой стадии возбудителя. На фоне увеличивающейся встречаемости ооспор патогена наблюдают существенное увеличение агрессивности патогена и его устойчивости к применяемым системным, трансламинарным, и контактным фунгицидам. В этой связи, определение стратегий размножения и поддержания жизнеспособности популяций патогена, чему и посвящена настоящая работа, сейчас приобретает принципиальное значение.
Цель работы –
оценить жизнеспособность полевых популяций возбудителя фитофтороза, направленная на получение оперативных характеристик патосистемы картофель – томат – P
.
infestans
для оптимизации защитных мероприятий на этих культурах против фитофтороза.
Задачи работы
для разработки системы оценки жизнеспособности и агрессивности полевых популяций
P
.
infestans
1. Изучить потенциал бесполого размножения и агрессивность изолятов P
.
infestans
из России и одного из основных центров происхождения заболевания – Мексики.
Реклама

2. Изучить встречаемость структур полового размножения – ооспор в полевых популяциях P
.
infestans
из различных регионов России на картофеле и томате.
3. Оценить морфологические особенности, жизнеспособность, способность к прорастанию, и инфекционное значение ооспор P
.
infestans
с учетом их различного происхождения – гибридного, партеногенетического и от самооплодотворения.
4. На основе традиционных подходов к учетам возбудителя фитофтороза и полученных нами данных разработать новую комплексную методологию, позволяющую оценивать репродуктивные характеристики и агрессивность полевых популяций патогена.
по использованию разработанной системы оценки для решения
научно-теоретических проблем, связанных с изучением био-экологических особенностей современных популяций
P
.
infestans
5. Использовать разработанную систему оценки для получения первичных характеристик полевых популяций P
.
infestans
в пространстве и времени.
6. Оценить роль томата в связи с возможными сценариями развития эпифитотии фитофтороза на картофеле.
по использованию разработанной системы оценки для решения
практических проблем, связанных с совершенствованием подходов
защиты картофеля и томата от фитофтороза
7. Определить уровень устойчивости и наличие адаптивной устойчивости в современных полевых популяциях P
.
infestans
к фунгицидам.
8. Использовать разработанную систему оценки для определения сравнительной эффективности химического и селекционно-семеноводческого методов защиты картофеля от фитофтороза.
Научная новизна работы.
Проведена оценка различных характеристик ооспор P
.
infestans
– их встречаемость в природных популяциях патогена, морфология, образование в результате перекрестного оплодотворения, самооплодотворения и партеногенеза, сохранение жизнеспособности, способность к прорастанию и эпидемиологическое значение.
Разработана система оценки, позволяющая определять стратегии размножения и поддержания жизнеспособности полевых популяций P
.
infestans
. Она может быть очень полезна как при проведении популяционных исследований патогена, так и при полевых обследованиях посадок картофеля и томата, пораженных фитофторозом. Полученные данные характеризуют накопление структур первичной и вторичной инфекции патогена и, таким образом, могут быть использованы для прогнозирования развития заболевания.
На основе изучения патосистемы картофель – томат – P
.
infestans
pазработана модель развития эпифитотии на местном успешно адаптированном виде при условии последовательного внедрения родственного восприимчивого неприспособленного вида-интродуцента.
Реклама

Практическая значимость работы.
Разработана новая система оценки, позволяющая определять полевую устойчивость сортов картофеля и томата к фитофторозу, а также эффективность защитных мероприятий в условиях конкретного агроценоза.
Основные положения, выносимые на защиту
1. Новая научно-обоснованная система оценки развития фитофтороза в полевых условиях, сочетающая в себе подходы, применяемые в полевых учетах и лабораторных исследованиях.
2. Мультивариантность образования и дальнейшего развития ооспор P
.
infestans
при различных природных и производственных условиях.
3. Наличие на томате штаммов P
.
infestans
, высокоагрессивных по отношению к сортам картофеля со значительным уровнем полевой устойчивости.
4. Отсутствие ожидаемой отрицательной взаимосвязи между устойчивостью как к системным, так и к контактным фунгицидам, и агрессивностью по отношению к растениям-хозяевам у отдельных штаммов P
.
infestans
.
Апробация работы.
Материалы диссертации представлены на 19 конференциях: конференции «Современные проблемы микологии, альгологии и фитопатологии», Москва, МГУ имени М.В. Ломоносова (1998); Х Московском совещании по филогении растений. Москва, МГУ имени М.В. Ломоносова (1999); молодежной конференции ботаников в Ботаническом институте, Санкт-Петербург (2000); научных конференциях в РГАУ-МСХА имени К.А. Тимирязева (2000, 2002, 2007, 2009 г.); научной конференции «Памяти Грегора Менделя», Москва, ИОГен (2001); международном симпозиуме «Проблемы изучения и сохранения биоразнообразия европейских природных сообществ, Москва, МГУ имени М.В. Ломоносова (2001); 7‑м международном микологическом конгрессе, Осло (2002); первой и второй всероссийских конференциях «Современные проблемы иммунитета растений к вредным организмам», Санкт-Петербург (2002, 2008); всероссийской научно-практической конференции «Ставропольский государственный аграрный университет – 75 лет» (2004); юбилейной конференции, посвящённой 85‑летию кафедры микологии и альгологии МГУ имени М.В. Ломоносова (2004); втором всероссийском съезде по защите растений в Санкт-Петербурге (2005), конференции «Грибы и водоросли в биоценозах», Москва, МГУ имени М.В. Ломоносова (2006); международной конференции «Научное наследие Н.И. Вавилова – фундамент отечественного и мирового сельского хозяйства», РГАУ-МСХА имени К.А. Тимирязева (2007); 60‑м международном симпозиуме по защите растений, Бельгия, Гентский университет (2008); третьей научной конференции «Микология, микотоксикология и микозы», Сербия, Нови Сад (2009).
Публикации.
Основные результаты исследований представлены в 49 публикациях, 16 из них – в журналах, рекомендованных ВАК РФ.
Структура и объем работы.
Диссертация изложена на 427 страницах. Содержит вводную (3 главы), информационно-аналитическую (1 глава), экспериментальную (6 глав), и заключительную (3 главы) части; а также 2 приложения (4 страницы). Содержит 90 таблиц и 65 рисунков. Список литературы включает 1324 наименования, в том числе 956 иностранных.
Участие в работе
. Работа выполнена непосредственно соискателем и представляет собой завершенный научный труд. В выполнении экспериментальной части работы принимали участие аспиранты и студенты кафедры фитопатологии РГАУ-МСХА. Большую помощь оказал С.А. Кузнецов. Автор признателен зав. кафедрой фитопатологии РГАУ-МСХА профессору В.А. Шкаликову за создание условий для проведения работы, своему учителю заведующему кафедрой микологии и альгологии МГУ имени М.В. Ломоносова профессору Ю.Т. Дьякову, профессору ВНИИ карантина растений С.С. Ижевскому и профессору ВНИИ овощеводства К.Л. Алексеевой за консультации.
Введение
В этой главе детально разбирается актуальность тематики работы. Отмечается, что P
.
infestans
привлекает свое внимание как модельный объект. Полученные данные применимы и для других паразитов растений. В работе исследуется комплекс взаимосвязанных вопросов, имеющих как научно-теоретическое, так и практическое значение. В связи с преодолением полевой устойчивости сортов картофеля и томата это изучение агрессивности патогена, в связи с массовым применением против фитофтороза фунгицидов – уточнение уровня устойчивости к ним P
.
infestans
. Усиление агрессивности и устойчивости к фунгицидам может в ряде случаев определяться влиянием половой стадии возбудителя фитофтороза. На основе ее изучения создана и применена на практике комплексная методика, позволяющая оценивать стратегии размножения и поддержания жизнеспособности патогена. Наконец, детально проанализированы причины острой фитосанитарной ситуации, сложившейся в ряде регионов России на томате из-за поражения фитофторозом.
Характеристики патосистемы картофель-томат –
P
.
infestans
Рассмотрены ботанические характеристики растений-хозяев. Охарактеризованы основные направления селекции картофеля (Симаков и др., 2005) и томата (Гавриш, 2005). Приводятся основные этапы изучения P
.
infestans
и фитофтороза. Анализируются достижения мировых школ по изучению P
.
infestans
. Далее в работе рассматриваются бесполая фаза и cистема скрещивания P
.
infestans
. Анализируются работы о регуляции полового процесса, осуществляемой посредством «веществ совместимости» и половых гормонов (Lealetal., 1964; Ко, 1978; Elliott, 1983; Brasier, 1992; Kemmit, 1992). В настоящее время жизнеспособные ооспоры P
.
infestans
встречаются во многих странах.
Разбираются вопросы, связанные с популяционной биологией P
.
infestans
и миграционной теорией. Анализируются достоинства и недостатки последней. Также приводится анализ различий междупопуляциями P
.
infestans
на картофеле и томате.
Проанализированы методы защиты картофеля и томата от фитофтороза.
Исследования прорастания зооспорангиев и агрессивности
P
.
infestans
из России и Мексики
Бесполое спороношение P
.
infestans
играет основную роль во вторичном инфицировании картофеля и томата (Попкова, 1972). Поэтому изучение прорастания зооспорангиев и агрессивности популяций имеет большое значение в научном и в практическом аспектах.
Материалы и методы.
В исследовании использовали 13 изолятов P
.
infestans
с разными типами спаривания, выделенных с картофеля и томата в 2004 на Валдае и в Лаборатории защиты растений РГАУ-МСХА. Также в исследовании использовали 25 штаммов P
.
infestans
из коллекции 1997 г. исследовательского центра Pictipapaпри Минсельхозе Мексики (долина Толука, г. Метепек). Они охарактеризованы по типам спаривания, а также по изоферментным локусам Gpi
и Pep
‑1.
Для определения агрессивности мексиканских изолятов использовали сегменты листьев восприимчивого сорта картофеля Альфа, российских изолятов – диски клубней восприимчивого сорта Сантэ и устойчивого сорта Луговской (Филиппов, 2005). Подробнее методика описана в работах А.Н. Смирнова (Смирнов, 2008). Статистический анализ проводили посредством сравнения распределения рангов в изучаемых выборках на основе определения χ2
(Глотов и др., 1982).
Результаты.
Российские изоляты.
При 7о
С прорастание зооспорангиев было непрямым (зооспорами), при 20 и 25о
С – прямым (ростковыми трубками).
При 20о
С зооспорангии изолятов P
.
infestans
(особенно А2) прорастали в среднем на 28% эффективнее, чем при 25о
С. Различий по прорастанию зооспорангиев изолятов, выделенных с картофеля и томата, не выявили.
Изоляты P
.
infestans
продемонстрировали довольно разный диапазон показателей агрессивности на клубневых дисках сорта Сантэ. По всем показателям выявили высоко агрессивные, умеренно агрессивные и мало агрессивные изоляты. Клубневые диски сорта Луговской поразились довольно слабо всеми исследуемыми изолятами P
.
infestans
. Исключением были изоляты Д2 (с томата), а также К2.8 и С5.1 (с картофеля). Изолят Д2 поразил клубневые диски сорта Луговской даже лучше, чем Сантэ.
Достоверных различий по агрессивности между изолятами с типами спаривания А1 и А2 не было обнаружено. В целом, изоляты, выделенные с картофеля, были несколько агрессивнее к клубневым дискам сорта Сантэ, чем изоляты, выделенные с томата. На клубневых дисках сорта Луговской показатели агрессивности были на низком уровне для обеих групп изолятов, и различий между ними не обнаружили.
Определяющий вклад в итоговый индекс агрессивности (ИИА) вносят такие компоненты как интенсивность спороношения, размер некрозов и латентный период (табл. 1). Напротив, такие компоненты как частота инфекции и инкубационный период не влияли на ИИА.
Таблица 1. Коэффициенты корреляции между компонентами агрессивности на клубневых дисках сортов Сантэ (слева от диагонали) и Луговской (справа от диагонали)
| ЧИ |
РН |
ИС |
ИП |
ЛП |
ИИА |
| ЧИ |
- |
- |
- |
- |
- |
- |
| РН |
0,81±0,10
|
- |
0,87±0,07
|
0,03±0,29 |
-0,78±0,11
|
0,94±0,03
|
| ИС |
0,67±0,16
|
0,79±0,11
|
- |
0,10±0,29 |
-0,79±0,11
|
0,96±0,02
|
| ИП |
0,11±29 |
-0,13±0,28 |
-0,29±0,26 |
- |
0,08±0,29 |
-0,04±0,29 |
| ЛП |
-0,75±0,13
|
-0,80±0,10
|
-0,87±0,07
|
0,37±0,25 |
- |
-0,80±0,10
|
| ИИА |
0,57±0,20 |
0,81±0,10
|
0,95±0,03
|
-0,27±0,27 |
-0,81±0,10
|
- |
Примечание: ЧИ – частота инфекции; РН – размер некрозов; ИС – интенсивность спороношения; ИП – инкубационный период; ЛП – латентный период;
ИИА – итоговый индекс агрессивности.
Корреляционный анализ не выявил достоверных взаимосвязей между прорастанием зооспорангиев при разных температурах и компонентами агрессивности. Исключением была лишь довольно хорошая корреляция между прорастанием зооспорангиев при 25о
С и интенсивностью спороношения на клубневых дисках сорта Луговской.
Мексиканские штаммы
.
При 7о
С у большинства штаммов было зарегистрировано только непрямое прорастание зооспорангиев с образованием зооспор, с разной интенсивностью. При 20о
С наблюдалось прямое прорастание 5 – 10% зооспорангиев штаммов. При 25о
С у всех штаммов отмечалось только прямое прорастание до 10% зооспорангиев.
Прорастание зооспорангиев у большинства изученных штаммов при 25о
С было в среднем на 19% эффективнее, чем при 20о
С.
У штаммов с типами спаривания А1 и А2 не выявлено различий в прорастании зооспорангиев при 7о
С. Однако при 20о
и 25о
С штаммы с наиболее высокой интенсивностью прорастания зооспорангиев относились к типу спаривания А1. Прорастание зооспорангиев у штаммов, собранных в начале и в конце вегетации, было на одинаковом уровне. В прорастании зооспорангиев при исследуемых температурах существенных различий между штаммами с аллелями Gpi
86, штаммами с аллелем Pep
‑1
92 и штаммами без этих аллелей выявлено не было. Лишь штаммы с аллелем Pep
‑1
92 (генотип 92/100) уступали штаммам без этого аллеля (генотип 100/100) при 7о
С.
Из 25 проверенных штаммов 5 были высоко агрессивны, 11 умеренно агрессивны, 9 мало агрессивны, 2 неагрессивны. Штаммы P
.
infestans
А1 и А2 типов спаривания были сходными по большинству компонентов. Лишь по ИС штаммы А1 превосходили штаммы А2 (табл. 2).
Таблица 2. Сравнение средних значений и распределений (рангов с низкими, средними и высокими значениями) по компонентам агрессивности для штаммов P
.
infestans
типов спаривания А1 и А2, собранных в начале и конце вегетации, по критерию χ2
| Компо-нент агрессив-ности |
Средние для штаммов |
χ2
|
Достоверность различия между группами штаммов |
А1,
в начале вегетации
|
А2,
в начале
вегетации
|
А1,
в конце
вегетации
|
А2,
в конце вегетации
|
| ЧИ |
4,1 |
3,7 |
4,7 |
4,8 |
7,5 |
- |
| РН |
3,6 |
3,1 |
4,3 |
4,4 |
10,0 |
- |
| ИС |
2,6 |
1,7 |
3,5 |
2,3 |
15,8 |
+ |
| ИП |
3,1 |
3,3 |
3,3 |
3,3 |
10,9 |
- |
| ЛП |
3,4 |
3,6 |
3,5 |
3,5 |
8,7 |
- |
| ИИА |
4,9 |
3,5 |
6,5 |
4,2 |
7,6 |
- |
Примечания: см. табл. 1. При P=0,05 и 6 степенях свободы предельное значение χ2
12,6.
Существенных различий по компонентам агрессивности между штаммами с аллелем Gpi
86, и штаммами без этого аллеля выявлено не было. Напротив, по большинству компонентов агрессивности штаммы, имеющие аллель Pep
‑1
92 достоверно уступали штаммам, не имеющим этого аллеля.
Достоверная корреляция (r=0,62±0,13)была выявлена только между прорастанием зооспорангиев при температурах 20о
и 25о
С. Между прорастанием зооспорангиев при 7о
С и других температурах корреляции выявлено не было. Проведенный корреляционный анализ показал, что компоненты РН, ИС и ИИА хорошо коррелировали между собой.
Корреляции выявили только между прорастанием зооспорангиев при 7о
С и компонентами агрессивности, исключая инкубационный и латентный периоды. Достоверную корреляцию установили, однако, только для размера некрозов (r=0,64±0,12). Между прорастанием зооспорангиев при температурах 20о
С или 25о
С и компонентами агрессивности корреляции выявлено не было.
По всем компонентам агрессивность штаммов P
.
infestans
была значительно (в 1,5–17 раз) выше при 20о
С, чем при 12о
С. При 12о
С штамм «старого типа» значительно уступал почти по всем компонентам агрессивности самому себе при 20о
С. Если при понижении температуры штаммы с эндемичными и рекомбинантными генотипами теряли агрессивность в 6–17 раз, то штамм «старого типа» – в 9 раз.
Обсуждение результатов.
У многих мексиканских штаммов P
.
infestans
прорастание зооспорангиев было выше при 25о
С по сравнению с 20о
С. Напротив, у большинства российских изолятов P
.
infestans
прорастание зооспорангиев было выше при 20о
С. Это можно объяснить климатическими различиями подмосковного региона и долины Толука.
В целом, не выявлено существенной связи между компонентами агрессивности, измеренной при температуре 20–21о
С и прорастанием зооспорангиев. Показано, что агрессивность P
.
infestans
связана с интенсивностью образования зооспор лишь в некоторой степени.
Ни предшествующие, ни наши результаты не свидетельствуют о полном вытеснении штаммов «старого типа» (с типом спаривания А1 и генотипом 86/100 по изоферментному локусу глюкозо‑6 фосфат изомераза или Gpi
‑
1
) другими штаммами (с типом спаривания А1 или А2 без аллеля 86 по Gpi
‑1
) за счет различий по компонентам агрессивности. Есть основания полагать, что на замещение генотипов по Gpi
повлиял внешний фактор – применение против фитофтороза металаксил-содержащих фунгицидов (Gisi, Cohen, 1996; Иванюк и др., 2005; Смирнов, 2007b; 2007c).
Чувствительность российских изолятов
P
.
infestans
к фунгицидам, часто применяемым против фитофтороза картофеля и томата
Для оомицетов первые свидетельства адаптивной устойчивости к фунгицидам из фениламидной группы были получены в начале 1980‑ых годов (Bruin, Edgington, 1981). Также с середины 1990‑ых годов против фитофтороза начали использовать Ширлан, Амистар (Квадрис), Акробат и Браво (Cooketal., 1995; 1999; www.syngenta.com; Kuhnetal., 1991; Albertetal., 1991; 2002). Эти препараты важны для совершенствования стратегий антирезистентности, активно применяемых на практике.
Диффузное развитие мицелия P
.
infestans
в растении (в стебле) может продлиться до 1–2 месяцев (Keay, 1953; 1954a; Kable, MacKenzie, 1980; Harrison, 1992; Lapwood, 1977). Замедление развития мицелия может быть связано с периодами засушливых погодных условий. В случае применения фунгицидов, это может быть связано и с тем, что концентрация современных фунгицидов после обработок может быть существенно ниже в стебле, чем в ботве (Hammetal., 2006) и мицелий в стебле будет сохранять свою жизнеспособность (Kapsa, 2000). В этой связи представляет интерес, изменится ли чувствительность мицелия к фунгицидам в зависимости от его возраста.
Материалы и методы.
Для анализа чувствительности к Ридомилу (д. в. металаксил), Ширлану (д. в. флуазинам), Амистару (д. в. азоксистробин), Акробату (д. в. диметоморф), и Браво (д. в. хлороталонил) использовали 182 изолята P
.
infestans
из различных регионов России (Москва, Московская область, Новгородская область, регионы Кавказа), выделенных с 1998 по 2004 г. Часть изолятов была из коллекций С.Н. Еланского (МГУ имени М.В. Ломоносова), другую часть – из коллекции РГАУ-МСХА. В экспериментах по изучению изменения чувствительности к фунгицидам использовали 5 изолятов P
.
infestans
из ЛЗР РГАУ-МСХА (Д3.2, Д13, и ДТП10.1) и Новгородской области (Валдай; ВалС13, ВалС15). Эти изоляты представляют полевые популяции патогена, против которых на протяжении многих лет использовали различные фунгициды. Более подробно методы рассмотрены в работе А.Н. Смирнова и Ю.С. Троицкой (Смирнов, Троицкая, 2008). Для каждого изолята по отношению к каждому из исследуемых фунгицидов определяли: 1) минимальную [порядковую] подавляющую концентрацию, 2) минимальную [порядковую] сдерживающую концентрацию.
Чувствительность к фунгициду определяли как (1 – (дф
/дк
))·100%, где дф
и дк
– диаметры колоний на среде с фунгицидом и в контроле соответственно. Отрицательные показатели означали, что изолят на среде с фунгицидом рос лучше, чем в контроле. Формула взята из работы В. Умэруса (Umaerus, 1984) и адаптирована для определения чувствительности к фунгицидам изолятов P
.
infestans
Результаты.
Наименьшей минимальной порядковой концентрацией (10-4%
), подавляющей большое количество (не менее 25%) изолятов P
.
infestans
, характеризовался фунгицид Акробат. Далее располагались фунгициды Амистар (10-4
, 10-3%
), Ридомил (10-3%
), Браво (10-2
, 10-1%
) и Ширлан (100%
).
Кластерный анализ порядковых концентраций 5 фунгицидов, подавляющих рост различных изолятов P
.
infestans
, выявил умеренное сходство между эффективностью системного фунгицида Ридомил и ограниченно системного трансламинарного фунгицида Амистар. Остальные фунгициды кластеризовались независимо (рис. 1).
Наименьшей минимальной порядковой концентрацией (10-5%
), сдерживающей большое количество (не менее 25%) изолятов P
.
infestans
, характеризовался фунгицид Ридомил. Далее в порядке возрастания располагались фунгициды Акробат (10-5
, 10-4%
), Амистар (10-4%
), Браво (10-3
, 10-2%
) и Ширлан (10-2%
).
Кластерный анализ порядковых концентраций 5 фунгицидов, сдерживающих рост различных изолятов P
.
infestans
выявил довольно высокое сходство по эффективности ограниченно системных фунгицидов Акробат и Амистар, а также контактных фунгицидов Браво и Ширлан (рис. 2).
Изменения чувствительности к сублетальным концентрациям фунгицидов Ширлан и Амистар у исследуемых изолятов была различной. У изолятов Д2 и Д13.2 наблюдали как уменьшение, так и увеличение чувствительности. У изолята ДТП10.1 чувствительность в основном возрастала, у изолятов ВалС13 и ВалС15 – не изменялась (табл. 3).
Рисунок 1. Дендрограмма сравнения фунгицидов по концентрациям, подавляющим изоляты P
.
infestans
Рисунок 2. Дендрограмма сравнения фунгицидов по концентрациям, сдерживающим изоляты P
.
infestans
Таблица 3. Возраст исходных культур (в сутках) изолятов P
.
infestans
, для которых отмечена наименьшая чувствительность к фунгицидам
| Фунгицид |
Возраст
изолята
|
Изоляты |
| Д3.2 |
Д13 |
ДТП10.1 |
ВалС13 |
ВалС15 |
| Ширлан |
Значения |
30 |
15, 30 |
30 |
не выявлен |
не выявлен |
| P |
>0,991
|
>0,652
|
>0,65 |
<0,653
|
<0,65 |
| Амистар |
Значения |
60 |
60, 30 |
30, 15 |
не выявлен |
15, 60 |
| P |
>0,99 |
>0,99 |
>0,65 |
<0,65 |
>0,65 |
| Ридомил |
Значения |
30, 15 |
15 |
не выявлен |
30 |
60, 15 |
| P |
>0,65 |
>0,65 |
<0,65 |
>0,65 |
>0,65 |
1
возраст культур с наименьшей чувствительностью установлен достоверно, P>0,99;
2
возраст культур с наименьшей чувствительностью установлен на уровне тенденции, P>0,65;
3
возраст, при котором отмечена наименьшая чувствительностью культур, не установлен, P<0,65.
При последовательном пассировании
на средах с Ширланом при втором пассаже рост был крайне ограничен, с развитием прижатого мицелия, на третьем пассаже способность к росту терялась у всех изолятов P
.
infestans
. На средах с Амистаром такой результат наблюдали для изолятов ДТП10.1 и ВалС13. У изолята Д13 происходило существенное повышение чувствительности. Напротив, у изолята ВалС15 чувствительность к Амистару значительно не изменялась. На средах с Ридомилом выявили разные направления изменения чувствительности.
При перекрестном пассировании
наблюдали резкое повышение чувствительности, если использовали среды с Ширланом (для всех изолятов) или Амистаром (для всех изолятов, кроме ВалС15). Если при первом и / или втором пассаже использовали среды с Ридомилом, то во многих случаях наблюдалось понижение чувствительности. Наименее эффективными последовательностями фунгицидов оказались: Ридомил®Ридомил®Амистар, Ридомил®Амистар®Ридомил. Однако было выявлено несколько исключений. Так, изолят ВалС15 резко понизил чувствительность к Ширлану после двух пассажей на среде с Амистаром. Изоляты ВалС13 и ВалС15 понизили чувствительность к Амистару после двух пассажей на среде с Ридомилом. Изоляты Д13 и ВалС15 понизили чувствительность к Амистару после пассажей на средах с Ридомилом и Ширланом.
Обсуждение результатов.
Наше исследование четко показало, что популяции P
.
infestans
включают группы штаммов с разной устойчивостью к фунгицидам. Наличие штаммов с повышенной устойчивостью к одному или двум фунгицидам (в том чиле и контактным) – это потенциальная опасность для стратегий антирезистентности. Также важно отсутствие обратных корреляционных взаимосвязей между агрессивностью и устойчивостью к различным фунгицидам.
По мере инкубации изолятов P
.
infestans
определенной тенденции изменения чувствительности к фунгицидам выявлено не было.В полевых условиях преимущество могут получить изоляты, способные не уменьшать определенный уровень толерантности к фунгицидам до начала обработок.
Ооспоры
P
.
infestans
Изучая ооспоры P
.
infestans
, мы стремились изучать их в природе, в растениях-хозяевах, инфицированных естественным путем, и учитывали, что ооспоры могут иметь гибридное и негибридное происхождение.
Материалы и методы.
90 популяций P
.
infestans
(групп природных образцов растений-хозяев, пораженных фитофторозом, собранных в одном месте в течение одного вегетационного сезона) исследовали на встречаемость в них ооспор. Популяции образцов собирали в 14 регионах России: Москве, Московской, Ленинградской, Новгородской, Ярославской, Тульской, Брянской, Калужской, Тамбовской, Рязанской и Томской областях, Мордовии, Северном Кавказе, Ставропольском крае в 1997–2008 г. Количество собираемых образцов (листьев картофеля, листьев томата или плодов томата) зависело от размера изучаемой популяции: от 4 (из несколько растений-хозяев) до 80–100 (из довольно крупных массивов. Методы работы с ооспорами рассмотрены в работах А.Н. Смирнова и С.А. Кузнецова (Смирнов, Кузнецов, 2006).
Результаты.
Ооспоры обнаружили как в листьях картофеля, так в листьях и в плодах томата. Их встречаемость была неодинаковой (χ2
= 398,0 при 6 степенях свободы; критическое значение χ2
=12,6). Количество образцов листьев картофеля с ооспорами было низким, листьев томата – выше. Самым большим было количество образцов плодов томата с ооспорами (табл. 4).
Таблица 4. Встречаемость образцов с ооспорами P
.
infestans
| Происхождение образцов (растение-хозяин) |
Общее коли-чество образ-цов |
Число и процент образцов |
| в которых ооспоры не обнаруже-ны |
с редкой встречае-мостью ооспор |
с умеренной встречае-мостью ооспор |
с частой встречае-мостью ооспор |
| Картофель листья |
2888 |
1405 (95,6%) |
86 (3,0%) |
25 (0,9%) |
13 (0,5%) |
| Томат листья |
344 |
165 (88,1%) |
19 (5,5%) |
13 (3,8%) |
9 (2,6%) |
| Томат плоды |
445 |
290 (71,2%) |
48 (10,8%) |
28 (6,3%) |
52 (11,7%) |
Редкая встречаемость – не более 25 ооспор, умеренная – 26–250 ооспор, частая – более 250 ооспор в 50 полях зрения микроскопа (1 мм2
).
По четырем исследуемым признакам (диаметр, толщина стенки, наличие антеридия и форма оогония) в образцах 1999 г. выявили 40 морфологических типов ооспор. Среди них выявлено две большие группы. Первая представлена ооспорами типичной морфологии. Их диаметр (21 – 40 мкм) и форма (тяготеющая к шаровидной) были близки к типовым показателям (рис. 3). Вторая представлена ооспорами нетипичной морфологии (рис. 4). Их размер (15 – 20 мкм или 41 – 55 мкм) или форма (уклоняющаяся от шаровидной) отличались от характеристик ооспор типичной морфологии.
Анализы морфологии обнаруженных ооспор и соотношения типов спаривания мицелиев, их образующих, показали, что около 61% (от 31% до 83%) всех обнаруженных ооспор в 1999 г. имели негибридное происхождение. Доля гибридных ооспор составила лишь 2% (от 0% до 7%). Происхождение остальных ооспор было спорным. Совместное присутствие А1 и А2 в образцах не всегда приводило к образованию в них ооспор.
Добавление фунгицида Ридомил в среду оказывало сильный угнетающий эффект на образование гибридных ооспор.
Определение жизнеспособности ооспор P
.
infestans
до и после перезимовки показало, что после перезимовки большинство ооспор (от 50% и выше) сохранили свою жизнеспособность (табл. 5).
Таблица 5. Количество жизнеспособных ооспор от скрещивания изолятов А1 и А2 на ржано-томатном агаре и в плодах томата, определенная на основе плазмолизного теста, до и после перезимовки
| Годы |
Количество жизнеспособных ооспор, % |
| от скрещивания изолятов А1 и А2 на ржано-томатном агаре |
в плодах томата |
| до перезимовки |
после перезимовки |
до перезимовки |
после перезимовки |
| 2004–2005 |
60–70 |
60–80 |
80–90 |
60–85 |
| 2005–2006 |
57–70 |
32–44 |
90–100 |
28–50 |
| 2006–2007 |
90–95 |
69–90 |
52–94 |
69–90 |
Изучение прорастания ооспор дало следующие результаты.
За месяц инкубации скрещенных изолятов с примесью триходермы в районе скрещивания образовалось много нетипичных ооспор, до 80% которых прорастало короткой ростковой трубкой. Типичные ооспоры почти не прорастали.
Ооспоры от скрещиваний А1 и А2 на ржано-томатной среде и после перезимовки прорастали ростковой трубкой с частотой от 20 до 35%. Природные ооспоры из плодов томата прорастали с частотой, не превышающей 3%.
Дополнительный тест показал, что наибольшее влияние на прорастание типичных ооспор оказала обработка суспензией из мицелия и спор триходермы. Действие отрицательной температуры также усиливало прорастание ооспор.
Инокуляция суспензией ооспор ломтиков картофеля во влажных камерах не привело к их заражению. Нанесение ооспор на прорастающие клубни не привело к заражению выросших растений.
При заражении проростков томата установили, что они могут поражаться фитофторозом через почву, в которой содержатся ооспоры (или зооспорангии) P
.
infestans
в условиях, ослабляющих иммунитет растения. Однако более вероятно, что при этом проростки поражаются комплексом болезней по типу корневых гнилей, поражающих зародышевый корешок и гипокотиль. Фитофтороз входит в этот комплекс.
В полевом эксперименте, при нанесении суспензии ооспор на семенные клубни, массового поражения растений не обнаруживали. Исключением был очаг на сорте Дельфин (Лаборатория защиты растений РГАУ-МСХА, 2006 г.).
Обсуждение результатов.
Максимальное количество ооспор мы наблюдали в плодах томата. Справедливо утверждение, впервые сформулированное С.А. Кузнецовым, что в условиях Московской области плоды томата – это своеобразная «маленькая Мексика» (Смирнов, Кузнецов, 2005; Smirnovetal., 2008; Smirnov, Kuznetsov, 2009).
Мы полагаем, что в большинстве современных полевых популяций P
.
infestans
могут образовываться ооспоры всех трех типов по происхождению. Тип образуемых ооспор может в значительной степени регулироваться внешними факторами и условиями существования полевых популяций патогена (Turkensteenetal., 2000). Инфекционный цикл P. infestans
включает в себя несколько стратегий (Дьяков, 1992; 1998). Зооспоры и зооспорангии соответствуют r‑стратегии (размножение, низкая сопротивляемость), гибридные ооспоры – К-стратегии (жизнеспособность, высокая сопротивляемость) (Дьяков, 1992). Ооспоры от самооплодотворения и партеногенетические ооспоры соответствуют промежуточным стратегиям. Первые ближе к К-стратегии, вторые находятся между r- и К-стратегиями (Смирнов, 2003а).
Проведенные нами лабораторные и полевые эксперименты стали еще одним косвенным подтверждением того, что гибридные ооспоры могут иметь значительное инфекционное значение, которое, однако, может реализовываться не сразу, а в последующих вегетационных сезонах. Прямое исчерпывающее доказательство этого с использованием молекулярно-генетических маркеров RFLP получено в Голландии в начале 1990‑ых годов (Drenthetal., 1994).
Образование гибридных ооспор возможно только у жизнеспособных полевых популяций патогена. Самооплодотворение в бесполых полевых популяциях может быть этапом к гибридизации и со временем привести к кардинальным изменениям в генетической структуре полевых популяциях патогена. Вклад партеногенетических ооспор, на наш взгляд, в последние годы недооценивается. В определенных условиях (нежестких) именно партеногенетические ооспоры могут иметь существенное эпидемиологическое значение – вызывать ранние эпифитотии. Особенно это может быть актуально для томата, если такие ооспоры попадут как примесь на поверхность семенной кожуры семян.
Стратегии размножения и поддержания жизнеспособности полевых популяций
P
.
infestans
Соотношение бесполого и полового размножения имеет принципиальное значение в эволюции жизненных стратегий у грибов и псевдогрибов. Функцию размножения и распространения они осуществляют в бесполой фазе.
В этой связи, у P
.
infestans
целесообразно определять и исследовать компоненты долгосрочной и краткосрочной жизнеспособности (рис. 5). Долгосрочная жизнеспособность в природных условиях напрямую определяется половой стадией патогена, краткосрочная – в основном его агрессивностью. Агрессивность увеличивает общий потенциал патогена, но ослабляют его K‑потенции (Дьяков, 1992). При проявлении агрессивности на определенных сортах в последние годы выявлена некоторая специфичность (Маторина, личное сообщение; Flieretal., 2002; Макаров и др., 2003; Филиппов и др., 2004).
В основе нашего метода лежит единовременное определение встречаемости ооспор и зооспорангиев в тканях картофеля и томата.
Каждая популяция, каждый штамм P
.
infestans
может характеризоваться определенной агрессивностью. Однако в природе проявление агрессивности специфично корректируется рядом факторов – устойчивостью сортов и климатическими условиями (Youngetal., 2004). На восприимчивых сортах при благоприятных для фитофтороза условиях проявление агрессивности P
.
infestans
будет максимальным. На сортах с полевой устойчивостью проявление агрессивности будет сдерживаться (Carlisleetal., 2002; Pillet, 2003; Montarryetal., 2006; Flieretal., 2007).
Наконец, мы исходили из того, что все показатели должны выражаться в баллах, соответствующих определенному рангу. Согласно закону Вебера-Фехнера, ранговая оценка показателей болезни позволит избежать субъективных ошибок (Forbes, Korva, 1994).
Материалы и методы.
Использованы те же материалы, как в гл. 6. Обесцвечивание листьев и плодов томата проводили по методике А.Н. Смирнова и С.А. Кузнецова (Смирнов, Кузнецов, 2006). Обесцвеченные в 96% кипящем растворе этанола и 10% водного растворе хлорсодержащего средства образцы микроскопируют. При малом увеличении (около х150) проверяют 50 случайных полей зрения, каждое – примерно 1 мм2
. При поиске ооспор просматривают 50 полей зрения в тканях образца. При поиске зооспорангиев просматривают 25 полей зрения в тканях образца и 25 полей зрения – в свободном растворе. Если число обнаруженных ооспор или зооспорангиев не превышает 25, находится в интервале от 26 до 250, или превышает 250, то их встречаемость оценивают как редкую, умеренную или частую соответственно.
Далее подсчитывают индексы встречаемости зооспорангиев (ИЗ
) и ооспор (ИО
) в полевой популяции по формулам: ИЗ = 0,05·РС + 0,5·УС + ЧС, ИО = 0,05·РО + 0,5·УО + ЧO, где: РС – частота встречаемости образцов с редкими зооспорангиями, УС – частота встречаемости образцов с умеренными зооспорангиями, ЧС – частота встречаемости образцов с частыми зооспорангиями, РО – частота встречаемости образцов с редкими ооспорами, УО – частота встречаемости образцов с умеренными ооспорами, ЧО – частота встречаемости образцов с частыми ооспорами, %.
На основе предшествующего кластерного анализа (Смирнов, Кузнецов, 2001) распределения индексов можно выделить 4 основных стратегии размножения (рис. 6, 7): С
– слабая
, Б –
бесполая
, П –
половая
, и Д –
двойная
.
Распространенность болезни (Р
) и индекс развития болезни (И
) определяются по стандартным формулам (Попкова, Быченкова, 1969, с изменениями) (Практикум…, 1976): Р = n·100% / N; И = å(ai
·bi
) 100% / 5N, где n – число больных растений, ai
bi
– сумма произведений числа больных растений (ai
) на соответствующий им балл интенсивности поражения (bi
): [наименьший балл – 0
(отсутствие поражения), 1
– 0,1–10% растения поражено, 2
– 11–25% поражено, 3
– 26–50% поражено, 4
– 51–89% поражено, 5
(наибольший) – 90–100% растения поражено], N – общее число больных и здоровых растений.
Индекс агрессивности (ИА
) полевой популяции P
.
infestans
можно подсчитать так: ИА = (Р · И · ИЗ) / 10 000. ИАаналогиченпоказателюCFI
(Composite Fitness Index) (Tooley et al., 1986; Kadish, Cohen, 1988).
Индекс ИО характеризует способность полевой популяции формировать потенциал первичной инфекции непосредственно in
situ
, который следует отслеживать для предотвращения раннего развития фитофтороза (Evenhuisetal., 2007). Наибольшую интенсивность образования ооспор следует ожидать при промежуточном развитии заболевания, когда достаточно живой питающей ткани, достигаются высокие жизнеспособность мицелиев и вероятность контакта между А1 и А2. Наименьшую интенсивность образования
| ИО |
100
10,1
|
П
В популяции
регулярно образуются структуры полового размножения, образование
структур бесполого размножения ограничено
|
Д
В популяции регулярно образуются как структуры бесполого так и полового размножения
|
10,0
0
|
С
Популяция со слабым потенциалом размножения
|
Б
В популяции образуются преимущественно структуры бесполого размножения
|
| 0,1 20,0 |
20,1 100 |
ИЗ |
Рисунок 6. Стратегии размножения полевых популяций P
.
infestans
.
| ИО |
100
25,1
|
П2
|
Д2
|
Д4
|
Д6
|
25,0
10,1
|
П1
|
Д1
|
Д3
|
Д5
|
10,0
3,1
|
С3
|
Б3
|
Б6
|
Б9
|
3,0
0,1
|
С2
|
Б2
|
Б5
|
Б8
|
| 0 |
С1
|
Б1
|
Б4
|
Б7
|
| 0,1 20,0 |
20,1 40,0 |
40,1 60,0 |
60,1 100 |
ИЗ |
Рисунок 7. Подстратегии размножения полевых популяций P
.
infestans
.
ИО
|
100
10,1
|
Ж
Популяция слабо агрессивна, имеется регулярное образование структур выживания (ооспор)
|
О
Популяция агрессивна, имеется регулярное образование структур выживания (ооспор)
|
10,0
0
|
Н
Популяция слабо агрессивна, образование структур выживания (ооспор) ограничено
|
А
Популяция агрессивна, образование структур выживания (ооспор) ограничено
|
| 0,1 10,0 |
10,1 100 |
ИА |
Рисунок 8. Стратегии поддержания жизнеспособности полевых популяций P
.
infestans
| ИО |
100
25,1
|
Ж2
|
О2
|
О4
|
О6
|
25,0
10,1
|
Ж1
|
О1
|
О3
|
О5
|
10,0
3,1
|
Н3
|
А3
|
А6
|
А9
|
3,0
0,1
|
Н2
|
А2
|
А5
|
А8
|
| 0 |
Н1
|
А1
|
А4
|
А7
|
| 0 10,0 |
10,1 25,0 |
25,1 50,0 |
50,1 100 |
ИА |
Рисунок 9. Подстратегии поддержания жизнеспособности полевых популяций P
.
infestans
Ооспор следует ожидать при ограниченном развитии заболевания или наоборот при жестком развитии болезни (Romero-Montesetal., 2008).
На основе распределения этих индексов можно выделить 4 стратегии поддержания жизнеспособности: (рис. 8, 9): Н –
нет
агрессивности и образования ооспор; популяция слабо агрессивна, образование ооспор ограничено; А
– агрессивная
; популяция агрессивна, образование ооспор ограничено; Ж –
жизнеспособная
долгосрочно, популяция слабо агрессивна, имеется регулярное образование ооспор; и О
– есть оба
свойства, популяция агрессивна, имеется регулярное образование ооспор.
Мелкоделяночные эксперименты с проведением обработок фунгицидами Хлорокись меди (Гринбелт, норма расхода 6,5 кг/га), Ридомил голд (Сингента, норма расхода 2,5 кг/га), Ширлан (Сингента, норма расхода 0,4 л/га), Амистар [Квадрис] (Сингента, норма расхода 0,6 л/га) и Бактерицид (НПО «Каустик», норма расхода 10 л/га) 21.08 и 29.08. проводили в 2000, 2003 и 2004 годах на территории Лаборатории защиты растений РГАУ-МСХА.
При исследовании устойчивости сортов к фитофторозу для массива растений каждого сорта определяли индекс полевой устойчивости сорта – ИПУС = (1/ИА) ∙ 100. ИПУС
<10,0%
– сорт в основном восприимчив; 10,1–25,0%
– сорт малоустойчив; 25,1–50,0%
– сорт среднеустойчив; 50,1–100,0%
– сорт устойчив; > 100,0%
– сорт гиперустойчив, возможен существенный вклад и вертикальной устойчивости. 5‑бальная шкала полевой устойчивости сортов картофеля обоснована в предшествующих исследованиях (Попкова, Быченкова, 1969; Макаров и др., 2003). В соответствии с латвийской методикой В.А. Гауера (Гауер, 1965), биологическую эффективность применения сорта с полевой устойчивостью по показателю ИР (БИИр
) определяли по формуле: БИир
=((ИРs
-ИРпу
)/ ИРs
)ּ100, где ИРs
иИРпу
– индексы развития болезни на восприимчивом и устойчивом сортах. Определение БИИЗ
и БИИА
аналогично.
Результаты.
И на картофеле, и на томате преобладали бесполые популяции P
.
infestans
со стратегией размножения Б (рис. 10).
Популяции с регулярным бесполым и половым размножением встречались только на томате. Распределения стратегий размножения в популяциях с листьев картофеля, листьев томата и плодов томата различались друг от друга (χ2
= 18,4 при 6 степенях свободы; критическое значение χ2
12,6).
В листьях картофеля и томата преобладали популяции с подстратегиями Б4, Б1, Б2, Б7 и С2, в плодах томата – Б3 (табл. 6).
Для 89 популяций P
.
infestans
провели корреляционный анализ между распределениями индексов встречаемости зооспорангиев (ИЗ) и ооспор (ИО). В целом, между образованием зооспорангиев существует очень слабо выраженная тенденция к обратной взаимосвязи. Более выраженная тенденция к обратной взаимосвязи (r=-0,41) прослеживается только для плодов томата.
Далее приводятся флуктуации значении ИЗ и ИО для вегетационных периодов различных лет, отличающихся по погодным условиям (табл. 7).
 Рисунок 10. Число популяций P
.
infestans
с определенной стратегией размножения (С, Б, П и Д) на картофеле и томате в 1997–2006 г. Рисунок 10. Число популяций P
.
infestans
с определенной стратегией размножения (С, Б, П и Д) на картофеле и томате в 1997–2006 г.
Таблица 6. Число популяций P
.
infestans
с определенной подстратегией размножения в листьях картофеля, а также листьях и плодах томата
| Подстрате-гия |
Местонахождение популяций P
.
infestans
|
Подстрате-гия |
Местонахождение популяций P
.
infestans
|
| листья картофеля |
листья томата |
плоды томата |
листья картофеля |
листья томата |
плоды томата |
| С1 |
2 |
0 |
0 |
Б1 |
8 |
4 |
1 |
| С2 |
5 |
2 |
1 |
Б2 |
8 |
3 |
0 |
| С3 |
2 |
0 |
1 |
Б3 |
0 |
0 |
6 |
| П2 |
0 |
0 |
2 |
Б4 |
9 |
4 |
1 |
| Д1 |
0 |
1 |
2 |
Б5 |
8 |
0 |
1 |
| Д3 |
0 |
0 |
1 |
Б6 |
3 |
0 |
0 |
| Д5 |
0 |
1 |
0 |
Б7 |
5 |
1 |
1 |
| Б8 |
4 |
0 |
0 |
| Б9 |
1 |
0 |
0 |
| Общее число подстратегий |
11 |
7 |
10 |
| Индекс Шеннона |
3,23 |
1,85 |
2,94 |
Таблица 7. Характеристика развития P
.
infestans
на картофеле в Московской области в различные годы
Год
|
Характеристика года |
Среднее значение ИЗ |
Среднее значение ИО |
Подстратегии популяций
P
.
infestans
|
| 1997 |
Умеренный (с засухами) |
13,7±6,2 |
3,0±3,5 |
С2, С3 |
| 1999 |
Депрессивно-благоприятный |
63,7±15,6 |
1,4±2,2 |
Б8, Б7, Б9, Б4, Б2 |
| 2000 |
Умеренный |
31,5±8,7 |
0,4±0,2 |
Б2, Б5 |
| 2001 |
Депрессивно-умеренный |
37,5±12,5 |
0,1±0,1 |
Б4, Б1, Б2 |
| НСР095
|
12,5 |
2,3 |
ИЗ имел высокие значения в первой половине сезона, далее нередко наблюдали его уменьшение. ИО изменялся по-разному (рис. 11).
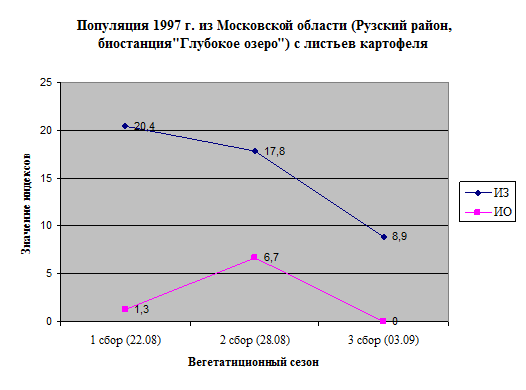
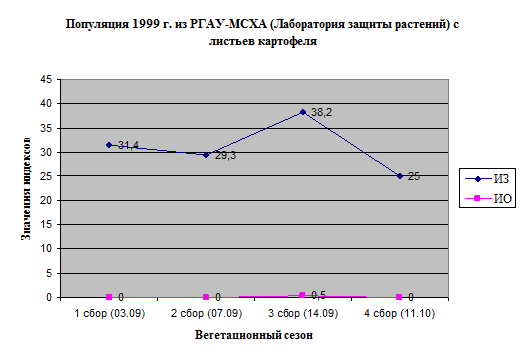 Рисунок 11. Изменение индексов ИЗ, ИО и стратегий размножения полевых популяций P
.
infestans
на картофеле в течение вегетационного сезона.
Значения ИЗ были выше в 1999 г., значения ИО – несколько выше в 1999 г. и 1997 г. Однако различия в ИО определялись вкладом отдельных популяций, в целом, различий по ИО мы не выявили.
Изучение влияния применения фунгицидов
Ридомил голд, Ширлан, Амистар, Оксихом и Бактерицид на изменение стратегий размножения и поддержания жизнеспособности полевых популяций P
.
infestans
в тесте 2003 г. показало следующие результаты.
Изменение стратегий размножения
(табл. 8).В контроле на восприимчивом сорте Сантэ наблюдалось увеличение индекса ИЗ (лишь к концу сезона он несколько уменьшался) и изменение подстратегии поддержания жизнеспособности от С1 к Б4. Напротив, на устойчивом сорте Луговской ИА оставался на одинаковом умеренном уровне, но потом снижался. В течение всего сезона подстратегия размножения изменялась от Б1 к С1.
Во всех вариантах на сорте Сантэ, кроме варианта с Ширланом, индекс ИЗ в общих чертах изменялся как в контроле. После обработки Ридомилом голд ИЗ достиг максимального значения – 100.
На сорте Луговской после обработки фунгицидами в ряде вариантов (Бактерицид, Хлорокись меди, Ридомил голд) значения ИЗ были высокими (особенно в варианте с Бактерицидом – близко к максимальному) и превышающими показатели в контроле. Стратегии размножения после обработки у этих вариантов были Б7 (Бактерицид) или Б4. Напротив, в вариантах с Ширланом и Амистаром наблюдали кардинальное уменьшение ИЗ, стратегия размножения после обработки была С1.
Изменение стратегий поддержания жизнеспособности
(табл. 8).В контроле на сорте Сантэ наблюдалось увеличение индекса ИА и изменение подстратегии поддержания жизнеспособности от Н1 к А1. Напротив, на сорте Луговской в течение всего сезона подстратегия поддержания жизнеспособности оставалась Н1.
В вариантах с обработкой Бактерицидом, Хлорокисью меди и Амистаром на сорте Сантэ наблюдали повышение ИА и измнение подстратегии поддрежания жизнеспособности как в контроле. Но после обработки Ширланом и Ридомилом голд наблюдали снижение ИА. В отличие от контроля, стратегия была Н1 в течение всего сезона.
Во всех вариантах на сорте Луговской наблюдали снижение ИА. Стратегия была Н1 на протяжение всего сезона, за исключением варианта с Ридомилом голд (до обработки – А1, после – Н1).
Полученное распределение свидетельствует о том, что с одной стороны – стандартный общепринятый показатель индекс развития болезни (ИР) и с другой стороны – предложенные нами показатели индекс образования зооспорангиев (ИЗ) и общий показатель индекс агрессивности (ИА) – самодостаточные и независимые друг от друга показатели, способные дополнять друг-друга и характеризующие полевые популяции P
.
infestans
в разных ситуациях.
Таблица 8. Изменение индексов и стратегий полевой популяции P
.
infestans
на сортах картофеля Сантэ и Луговской после обработки фунгицидом; тест 2003 г.
| Варианты |
Учитыва-емые показа-тели |
Учеты |
| 1 учет 19.08 |
2 учет 27.08 |
3 учет 09.09 |
4 учет 17.09 |
| Сорта |
| Сантэ* |
Луг.* |
Сантэ |
Луг. |
Сантэ |
Луг. |
Сантэ |
Луг. |
| Без обработки |
Контроль
(без обработки)
|
ИО |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
| ИЗ |
15,6 |
35,0 |
62,5 |
33,8 |
70,0 |
33,8 |
24,3 |
5,0 |
| ИР |
4 |
2,7 |
10,7 |
10,7 |
29,3 |
17,3 |
24,3 |
21,3 |
| ИА |
0,1 |
0,1 |
3,6 |
1,9 |
15,1 |
3,9 |
15,6 |
0,4 |
| СР |
С1 |
Б1 |
Б7 |
Б1 |
Б7 |
Б1 |
Б4 |
С1 |
| СПЖ |
Н1 |
Н1 |
Н1 |
Н1 |
А1 |
Н1 |
А1 |
Н1 |
| До обработки |
После обработки |
| Обработка Ридомилом голд |
ИО |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
| ИЗ |
5,0 |
5,0 |
33,8 |
57,1 |
100 |
57,5 |
35,7 |
5,0 |
| ИР |
2,7 |
2,7 |
22,7 |
22,7 |
4,0 |
4,0 |
16,0 |
2,7 |
| ИА |
0,1 |
0,1 |
7,7 |
13,0 |
0,8 |
0,3 |
3,8 |
0,1 |
| СР |
С1 |
С1 |
Б1 |
Б4 |
Б7 |
Б4 |
Б1 |
С1 |
| СПЖ |
Н1 |
Н1 |
Н1 |
А1 |
Н1 |
Н1 |
Н1 |
Н1 |
| Обработка Ширланом |
ИО |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
| ИЗ |
50,8 |
31,4 |
28,3 |
25,0 |
5,0 |
12,5 |
5,0 |
28,3 |
| ИР |
4,0 |
3,3 |
33,3 |
34,7 |
4,0 |
5,3 |
5,3 |
2,9 |
| ИА |
0,4 |
0,2 |
9,4 |
8,7 |
0,1 |
0,1 |
0,1 |
0,1 |
| СР |
Б4 |
Б1 |
Б1 |
Б1 |
С1 |
С1 |
С1 |
Б1 |
| СПЖ |
Н1 |
Н1 |
Н1 |
Н1 |
Н1 |
Н1 |
Н1 |
Н1 |
| Обработка Амистаром |
ИО |
0 |
1,3 |
0 |
0 |
0 |
0 |
0 |
0 |
| ИЗ |
38,6 |
16,3 |
40,0 |
56,9 |
81,3 |
16,3 |
12,5 |
16,3 |
| ИР |
6,0 |
5,3 |
2,7 |
28,0 |
22,7 |
9,3 |
36,7 |
13,3 |
| ИА |
0,7 |
0,2 |
0,1 |
15,9 |
17,2 |
0,7 |
4,1 |
1,2 |
| СР |
Б1 |
С2 |
Б1 |
Б4 |
Б7 |
С1 |
С1 |
С1 |
| СПЖ |
Н1 |
Н2 |
Н1 |
А1 |
А1 |
А1 |
Н1 |
Н1 |
| Обработка Оксихомом |
ИО |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
1,9 |
| ИЗ |
16,3 |
35,0 |
10,6 |
66,7 |
87,5 |
51,3 |
38,8 |
16,9 |
| ИР |
5,3 |
2,7 |
2,7 |
10,7 |
20,0 |
13,3 |
30,7 |
33,3 |
| ИА |
0,1 |
0,2 |
0,1 |
3,8 |
8,2 |
2,7 |
7,9 |
3,8 |
| СР |
С1 |
Б1 |
С1 |
Б7 |
Б7 |
Б4 |
Б1 |
С2 |
| СПЖ |
Н1 |
Н1 |
Н1 |
Н1 |
Н1 |
Н1 |
Н1 |
Н2 |
| Обработка Бактерици-дом |
ИО |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
| ИЗ |
20,6 |
45,6 |
40,0 |
33,0 |
75,6 |
93,8 |
22,5 |
5,0 |
| ИР |
1,3 |
2,7 |
4,0 |
22,7 |
44,0 |
10,7 |
36,0 |
22,7 |
| ИА |
0,1 |
0,1 |
0,2 |
7,5 |
24,4 |
4 |
5,4 |
0,7 |
| СР |
Б1 |
Б4 |
Б4 |
Б1 |
Б7 |
Б7 |
Б4 |
С1 |
| СПЖ |
Н1 |
Н1 |
Н1 |
Н1 |
А1 |
Н1 |
Н1 |
Н1 |
* Сантэ – восприимчивый в условиях Московской области сорт; Луг. (Луговской) – устойчивый сорт.
Сравнение мицелиев, чувствительных и устойчивых к металаксилу, из природных пораженных фитофторозом образцов по интенсивности образования зооспорангиев дало следующие результаты.
Из образцов картофеля и томата 2000, 2003 и 2004 г. выделяли изоляты, для которых определяли устойчивость к металаксилу и типы спаривания. Также в образцах определяли встречаемость зооспорангиев. Результаты анализа на основе критерия χ2
свидетельствуют о том, что, в целом, чувствительные и устойчивые к металаксилу изоляты P
.
infestans
(без значительной зависимости от их типов спаривания) в естественных условиях на листьях картофеля отличаются по интенсивности образования зооспорангиев. Среди устойчивых изолятов мы почти не выявили высокой интенсивности образования зооспорангиев. Устойчивые изоляты А1 и А2 по интенсивности образования зооспорангиев между собой не отличались.
Исследования полевой устойчивости сортов картофеля к фитофторозу проводили в 2000 и 2003–2008 г. Данные в довольно засушливые 2005 и 2007 г. не включены, так как в силу депрессивных условий они не могут показывать реальные показатели агрессивности P
.
infestans
, а следовательно и полевой устойчивости изучаемых сортов картофеля.
Протестированные сорта картофеля можно поделить на три группы.
Первая группа сортов
(табл. 9) характеризуется восприимчивостью или практически полным отсутствием полевой устойчивости к фитофторозу. По нашим данным, в нее входит большинство голландских сортов, а также многие российские сорта: Лина, Ресурс, Утенок, Русский сувенир, Чародей.
Трудно объективно охарактеризовать устойчивость голландского сорта Сатурна (2007 г.), показавшего значительную устойчивость на фоне засушливого вегетационного сезона и жестких обработок фунгицидами. Он все же поразился фитофторозом, когда другие обследованные сорта совсем не поразились фитофторозом и при отсутствии обработок. Исследование этого сорта целесообразно повторить.
Другой особый случай – шотландский сорт Джура (IsleofJura). Данные получили в середине вегетационного сезона 2006 г. после нескольких обработок фунгицидами. Поэтому говорить об устойчивости (хотя бы незначительной; не смотря на значение индекса ИПУС) этого сорта не приходится. Напротив, можно полагать что развитие фитофтороза на этом сорте было частично и временно сдержано фунгицидными обработками.
Вторая группа сортов
(табл. 9) характеризуется наличием полевой устойчивости к фитофторозу. Однако эта полевая устойчивость либо не работает к концу вегетационного сезона, либо не воспроизводится во всех проведенных испытаниях. По нашим данным, в эту группу входят российские сорта Невский, Жуковский ранний, Никулинский, Удача.
Третья группа сортов
(табл. 9) характеризуется наличием стабильной полевой устойчивости к фитофторозу. По нашим данным, в нее входят украинский сорт Луговской, российские сорта Водолей и Герань, белорусский сорт Ветразь, и голландский сорт Гранола.
Таблица 9. Характеристики полевой устойчивости к фитофторозу сортов картофеля, выведенных в четырех странах, с учетом подстратегий размножения и поддержания жизнеспособности полевых популяций P
.
infestans
в местах проведения испытаний
| № |
Сорт |
Страна, где выведен
сорт
|
Место тестиро-
вания
|
Год тести-рования |
ПР |
ПЖ |
ИПУС |
Ранг полевой устойчивости сорта |
| 1 |
Гранола |
Голландия |
ЛЗР |
2000 |
Б3 |
Н3 |
93,5 |
устойчив |
| 2 |
Сантэ |
Голландия |
ЛЗР |
2003 |
Б7 |
А1 |
6,6 |
восприимчив |
| ЛЦ |
2004 |
Б7 |
А1 |
4,9 |
восприимчив |
| ВНИИКХ |
2006 |
С1 |
А1 |
8,1 |
восприимчив |
| ЛЗР |
2006 |
Б4 |
А1 |
4,5 |
восприимчив |
| 3 |
Луговской |
Украина |
ЛЗР |
2003 |
Б1 |
Н1 |
25,6 |
среднеустойчив |
| ЛЦ |
2004 |
Б4 |
А1 |
4,2 |
восприимчив |
| ВНИИКХ |
2006 |
Б2 |
Н2 |
48,3 |
среднеустойчив |
| ВНИИКХ |
2006 |
С1 |
Н1 |
47,6 |
среднеустойчив |
| 4 |
Невский |
Россия |
ЛЗР |
2004 |
Б7 |
Н1 |
23,6 |
слабоустойчив |
| ЛЦ |
2004 |
Б4 |
А4 |
3,7 |
восприимчив |
| ВНИИКХ |
2006 |
Б1 |
А4 |
3,4 |
восприимчив |
| 5 |
Жуковский ранний |
Россия |
ЛЗР |
2004 |
Б6 |
Н3 |
1344,5 |
устойчив |
| ВНИИКХ |
2006 |
С1 |
А1 |
6,7 |
восприимчив |
| 6 |
Латона |
Голландия |
Валдай |
2004 |
Б7 |
А4 |
2,5 |
восприимчив |
| ЛЦ |
2004 |
Б4 |
А4 |
2,6 |
восприимчив |
| 7 |
Скарлет |
Голландия |
Валдай |
2004 |
С1 |
А1 |
7,9 |
восприимчив |
| 8 |
Ветразь |
Беларусь |
ЛЦ |
2004 |
С1 |
Н1 |
102,0 |
устойчив |
| 9 |
Никулинский |
Россия |
ЛЦ |
2004 |
Б1 |
А4 |
3,8 |
восприимчив |
| ВНИИКХ |
2006 |
Б1 |
Н1 |
62,5 |
устойчив |
| ВНИИКХ |
2006 |
Б1 |
А1 |
8,1 |
восприимчив |
| 10 |
Лина |
Россия |
ВНИИКХ |
2006 |
Б7 |
А4 |
2,5 |
восприимчив |
| 11 |
Удача |
Россия |
ВНИИКХ |
2006 |
Б4 |
Н1 |
32,7 |
среднеустойчив |
| ВНИИКХ |
2006 |
Б4 |
А4 |
2,8 |
восприимчив |
| ПС |
2006 |
Б3 |
Н3 |
12,3 |
слабоустойчив |
| ЛЗР |
2008 |
Б4 |
Н1 |
2,5 |
восприимчив |
| 12 |
Леди Розетта |
Голландия |
ВНИИКХ |
2006 |
Б5 |
А2 |
5,5 |
восприимчив |
| 13 |
Аусония |
Голландия |
ВНИИКХ |
2006 |
Б4 |
А4 |
3,0 |
восприимчив |
| 14 |
Водолей |
Россия |
ВНИИКХ |
2006 |
С1 |
Н1 |
40,0 |
среднеустойчив |
| 15 |
Герань |
Россия |
ВНИИКХ |
2006 |
Б4 |
Н1 |
35,7 |
среднеустойчив |
| 16 |
Ресурс |
Россия |
ВНИИКХ |
2006 |
Б4 |
А1 |
4,1 |
восприимчив |
| 17 |
Утенок |
Россия |
ВНИИКХ |
2006 |
Б7 |
А7 |
2,0 |
восприимчив |
| 18 |
Русский сувенир |
Россия |
ВНИИКХ |
2006 |
Б1 |
А1 |
4,6 |
восприимчив |
| 19 |
Чародей |
Россия |
ПС |
2006 |
Д3 |
О3 |
3,7 |
восприимчив |
| 20 |
Джура |
Шотландия |
Рогачево |
2006 |
Б8 |
Н1 |
10,5 |
слабоустойчив* |
| 21 |
Сатурна |
Голландия |
Рогачево |
2007 |
Б1 |
Н1 |
500 |
устойчив* |
| 22 |
Ильинский |
Россия |
ЛЗР |
2008 |
Б7 |
Н1 |
1,4 |
восприимчив |
Аббревиатуры: ПР – подстратегия размножения; ПЖ – подстратегия жизнеспособности; ИПУС – индекс полевой устойчивости сорта; ЛЗР – Лаборатория защиты растений РГАУ-МСХА; ПС – полевая станция РГАУ-МСХА; ЛЦ – лаборатория цветоводства РГАУ-МСХА; ВНИИКХ – Всероссийский НИИ картофельного хозяйства; сорта Джура и Сатурна помечены звездочками, их устойчивость к фитофторозу обсуждается в тексте.
Нам представляется, что без использования показателя ИЗ, то есть на основе исключительно визуальной оценки проводить испытания полевой устойчивости сортов картофеля к фитофторозу нецелесообразно. Это может исказить результаты учета.
Обсуждение результатов.
Применение шкалы встречаемости зооспорангиев и ооспор (на основе индексов ИЗ и ИО) позволяет отказаться от применения многочисленных статистических данных, нередко несущих небольшую смысловую нагрузку. Отличие индексов ИЗ и ИО от обычных показателей (процент образцов с ооспорами) заключается в том, что обычный показатель не учитывает встречаемость зооспорангиев и ооспор в образце.
В некоторых случаях значения встречаемости зооспорангиев или ооспор могут быть близкими к граничным значениям, разделяющим интервалы встречаемости. В таких случаях количество образцов в исследуемой выборке (характеризующей полевую популяцию) должно быть достаточно велико. Это позволит уменьшить риск случайных отклонений. Предложенную шкалу можно совмещать со всеми известными селекционными и фитопатологическими шкалами.
С биологической точки зрения очень важно, что реализация стратегий размножения и поддержания жизнеспоспособности популяций P
.
infestans
определяется как генетическими характеристиками штаммов, составляющих популяцию, так и экологическими факторами, оказывающими на нее влияние. Генетические характеристики определяют потенциал проявления репродуктивных свойств и агрессивности. Однако именно экологические факторы корректируют их непосредсвтенное фенотипическое проявление.
Полученные в 2003 г. данные свидетельствуют о том, что фунгициды Ширлан и Ридомил голд МЦ эффективны при защите картофеля от фитофтороза. Обработка ими, как правило, вела к снижению показателей ИЗ (только Ширлан), ИР и ИА, что адекватно отражалось на стратегиях размножения и поддержания жизнеспособности возбудителя (табл. 10).
Таблица 10. Эффект сорта и биологическая эффективность применения фунгицида против фитофтороза картофеля на делянках ЛЗР РГАУ-МСХА
| Сорт Сантэ |
Сорт Луговской |
| ИР |
ИЗ |
ИА |
ИР |
ИЗ |
ИА |
| Сорт |
-* |
-* |
-* |
40,9 |
51,8 |
74,1 |
| Бактерицид |
-50,0 |
-8,0 |
-62,0 |
-2,5 |
-229,5 |
-76,7 |
| Ширлан |
86,4 |
92,9 |
99,4 |
28,3 |
11,3 |
21,3 |
| Хлорокись меди |
31,8 |
-25,0 |
45,8 |
-17,8 |
-103,6 |
-44,1 |
| Ридомил голдМЦ |
86,4 |
-42,9 |
94,7 |
36,0 |
-122,1 |
18,0 |
| Амистар |
22,7 |
-16,1 |
-14,1 |
5,3 |
0,2 |
7,7 |
* сорт Сантэ взят как эталон восприимчивого сорта, сорт Луговской – устойчивого.
Лишь по ИЗ наблюдали отрицательную эффективность в случае обработки Ридомилом голд. Это связано с увеличением количества устойчивых штаммов (Троицкая, 2007), у которых обработки фениламидами не снижают, а иногда и провоцируют образование зооспорангиев. Напротив, Ширлан показал практически лечебный эффект (табл. 10).
И эффективные фунгициды, и устойчивость сорта в значительной степени сдерживали развитие фитофтороза. Если возделывание восприимчивого сорта будет зависеть от обработок, то для устойчивого сорта зависимость будет минимальной (Michelanteetal., 2000; табл. 10).
Индексы ИЗ и ИА в конце вегетационного сезона незадолго до уборки должны быть использованы как один из параметров прогнозирования лежкости клубней картофеля при хранении (Lapwood, 1977; Sato, 1980; Rotemetal., 1983b; Пшеченков и др., 2007).
Фитофтороз томата: современное положение, проблемы и возможные решения
В связи со сложившейся фитосанитарной ситуацией, необходимо охарактеризовать изменения, обнаруженные в последние годы для фитофтороза томата.
Материалы и методы.
Вредоносность фитофтороза на томате оценивали путем анализа специальной литературы (отчетов профильных институтов и региональных станций защиты растений), а также опроса агрономов, фермеров и опытных дачников из различных областей России. Анализ фитосанитарного риска (АФР) возникновения эпифитотии на картофеле, вызванной штаммами P. infestans
с томата, в условиях Московской области проводился по методу С.С. Ижевского (Ижевский, 2003) с изменениями, позволяющими использовать эту энтомологическую карантинную методику в фитопатологии.
Результаты.
Распространенность болезни в частных огородах, плантациях открытого грунта и небольших пленочных теплицах достигает 70–100% уже через 2–3 недели после появления первых признаков фитофтороза. Во многом этому способствуют грубейшие нарушения агротехники – такие как складывание пораженных растительных остатков в кучи и многолетнее выращивание томата на одном месте.
В случае, когда к пораженным фитофторозом посадкам томата непосредственно примыкали участки с картофелем, именно здесь на последних появлялись очаги фитофтороза. В крупных теплицах на территории Москвы и Московской области также наблюдались значительные отдельные вспышки фитофтороза. В некоторых хозяйствах, производящих семена и рассаду томата (в Ярославской области), были отмечены случаи поражения рассады, рекомендованной к реализации и высадке.
Более раннее проявление фитофтороза на томате и его разрушительный характер зарегистрировали в Тверской, Ярославской, Московской, Калужской, Рязанской и Пензенской областях. Но в более южных регионах фитофтороз не приводит к серьезным последствиям. Для Астраханской области фитофтороз томата не характерен.
Изоляты из пяти популяций, отличающихся по обработкам фунгицидами и наличием ооспор, проверили на устойчивость к фунгицидам. Для этого в лабораторных условиях на овсяном агаре определили концентрации фунгицидов, сдерживающие рост изолятов (табл. 11).
Таблица 11. Отношение штаммов из различных полевых популяций к фунгицидам Ридомил, Ширлан и Амистар в зависимости от обработки фунгицидом и образования ооспор
| Полевая популяция |
Растение-хозяин |
Ооспоры
в полевой популяции
|
Обработка фунгицидом |
Наиболее частые сдерживающие концентрации фунгицида (по д. в.), выявленные у штаммов полевой популяции после обработки этим фунгицидом |
| Ридомил, мкг/мл |
Ширлан, % |
Амистар,
%
|
| ЛЗР РГАУ-МСХА |
Картофель листья |
- |
+ |
10-1
, 10-2
, R1
|
10-2
, 10-3
|
10-4
|
| ЛЗР РГАУ-МСХА |
Картофель листья |
- |
-
(контроль)
|
10-1
, 1 |
10-2
|
10-5
, 10-6
,10-4
|
Рузский
район
|
Картофель листья |
+ |
- |
10-1
, 1, R |
10-1
|
10-4
|
Рузский
район
|
Томат листья |
+ |
- |
1 |
10-2
, 10-1
|
10-4
,10-3
,10-2
|
Рузский
район
|
Томат плоды |
+ |
- |
R, 1 |
10-2
|
10-4
|
1
R – устойчивые к ридомилу штаммы, для которых сдерживающей концентрации в
исследуемом диапазоне выявлено не было.
Оказалось, что изоляты, собранные после обработки Ридомилом-МЦ и Амистаром, были более устойчивы к фунгицидам, чем собранные с необработанного контроля. В варианте с Ширланом проявилась обратная закономерность. В вариантах из Рузского района устойчивость ко всем фунгицидам была еще выше. Так, наибольшая сдерживающая концентрация по Ширлану была выявлена для изолятов с листьев картофеля, по Амистару – с листьев томата, по Ридомилу – с плодов томата.
Есть все основания полагать, что в Рузском районе популяция P
.
infestans
с томата мигрировала на картофель, сохранив способность к образованию ооспор. В результате в этой агросистеме сформировалась очень жизнеспособная популяция возбудителя, толерантная к различным внешним воздействиям.
Обсуждение результатов.
Отмеченные факты свидетельствуют о том, что в ряде областей России на томате сложилось крайне неблагополучное фитосанитарное положение. Существующие рекомендации и меры защиты против фитофтороза томата (Филиппова, 1969; Гуркина, 2005) недостаточно эффективны против этих изменений.
В томатных популяциях P
.
infestans
, против которых не применяли фунгицидных обработок, но с образованием ооспор уровень устойчивости к новым фунгицидам оказался сравнимым и даже несколько превышающим аналогичный уровень в бесполых популяциях P
.
infestans
, против которых проводили обработки (табл 11). Столь контрастных различий между географически различными преимущественно бесполыми популяциями P
.
infestans
, против которых не проводили обработки, в предшествующем исследовании не обнаружили (Троицкая, 2007). Однако доказано, что при скрещиваниях между изолятами P
.
infestans
с разной степенью устойчивости к металаксилу часть потомков повышает уровень устойчивости по сравнению с родительскими изолятами (Knapovaetal., 2002).
Перенос дрозофилами зооспорангиев P
.
infestans
может значительно ускорять ход фитофтороза на томате (глава 9). Кроме того, он повышает вероятность контакта между мицелиями разных типов спаривания.
Небольшие по численности, томатные популяции патогена представляют собой мощный источник изменчивости и могут оказывать влияние на популяционную структуру вида P
.
infestans
в целом.
Интенсивность образования ооспор в плодах томата можно сравнивать с ситуацией, сложившейся в долине Толука на картофеле и диких пасленах (Flieretal., 2001). Плоды томата, богатые фосфолипидными соединениями, представляют собой среду, подходящую для полового процесса P
.
infestans
. Это объясняет ранние поражения томата фитофторозом, а также довольно тревожные сообщения из Бельгии о корневой гнили томата, вызываемой P
.
infestans
(Lievensetal., 2004).
Проведенные нами эксперименты по агрессивности логически указывают на две возможности преодоления полевой устойчивости картофеля (рис. 12).
Первая возможность
связана с попаданием томатных штаммов на небольшие делянки картофеля с восприимчивыми сортами, увеличением в количестве и усилением агрессивности. То, что изоляты с томата могут быть эффективны при заражении картофеля, было доказано в лабораторных опытах (Müller, 1936; Mills, Peterson, 1952; Лаврова, 2004).
Вторая возможность
связана с томатными штаммами, исходно агрессивными по отношению к устойчивым сортам картофеля. Возможность избирательного заражения устойчивых сортов высоко агрессивными изолятами P
.
infestans
доказана в Шотландии посредством использования маркеров SSR (Leesetal., 2007).
Представляет интерес выявленная деталь, что многие из исследованных нами агрессивных к сорту Луговской изолятов P
.
infestans
характеризовались способностью к прямому прорастанию зооспорангиев при температуре 25о
С. Эта температура оптимальна для прорастания зооспорангиев (Harrison, 1992), хотя этот признак может быть штаммоспецифичным (глава 4). Данное обстоятельство свидетельствует о том, что значительная способность зооспорангиев прорастать при 25о
С может повышать вероятность поражения фитофторозом устойчивых сортов картофеля.
Возможность заражения картофеля непосредственно томатными штаммами зарегистрирована посредством молекулярно-генетического анализа в предшествующем американском исследовании (Legard, Fry, 1995). В нашем исследовании (в рамках данной главы) молекулярно-генетический анализ не проводили. Но с учетом предшествующих данных наши полевые наблюдения и сравнительные результаты по агрессивности к картофелю и томату исследованных картофельных и томатных изолятов P
.
infestans
(глава 4) – это сильное косвенное свидетельсво способности к преодолению полевой устойчивости сортов картофеля томатными штаммами P
.
infestans
в условиях Московской области.
Высокий уровень опасности ситуации на томате в связи с его поражением фитофторозом связан еще и с частым образованием ооспор. И предшествующие, и наши исследования показали, что томат – серьезный источник сохранения и возобновления популяций возбудителя, в случае непосредственной территориальной близости посадок томата и картофеля представляющий угрозу и для последнего.
Анализ фитосанитарного риска показал опасность ситуации как для томата, так и для картофеля в условиях Московской области. Основной причиной сложившейся в связи с пораженаемостью фитофторозом ситуации на томате стало его масштабное выращивание в холодном климате. В таких условиях все сорта томата (в том числе и скороспелки) демонстрировали низкий уровень устойчивости к фитофторозу (Дементьева, 1970).
Помимо рассмотренной нами ситуации, есть много и других примеров, когда продвижение культуры в регион с климатическими условиями, отличающимися от основного ареала, приводило к аналогичным последствиям (Горленко, 1975). Все это позволяет сформулировать общую концепцию. Она заключается в следующем. При длительном сосуществовании неприспособленного интродуцированного восприимчивого вида растения-хозяина (в нашем случае – томат) и местного (приспособленного к своему ареалу, как в нашем случае, успешно адаптированного интродуцента картофеля) относительно устойчивого к опасному заболеванию вида растения-хозяина может произойти эрозия устойчивости местного приспособленного вида к данному заболеванию. Перечислим необходимые условия эрозии устойчивости местного сорта или вида растения-хозяина.
1. Высокий уровень родства (сходства) между неприспособленным интродуцированным восприимчивым видом и местным приспособленным (в том числе и успешно адаптированным интродуцентом) видом растений-хозяев.
2. Снижение или сохранение на низком уровне иммунитета вида или сорта неприспособленного интродуцированного восприимчивого вида в новых нетипичных для него условиях произрастания, его существование – только благодаря поддержке человека.
3. Произрастание неприспособленного интродуцированного восприимчивого вида и местного (в том числе и успешно адаптированного интродуцента) вида растений-хозяев близко друг от друга.
Высокий уровень родства (сходства) между интродуцированным неприспособленным восприимчивым и местным приспособленным видами растений-хозяев имеет принципиальное значение в случае общей болезни, вызываемой облигатным паразитом или факультативным сапротрофом с узкой или промежуточной специализацией. Эпифитотийная ситуация на местном растении-хозяине может реализоваться в четыре этапа (рис. 13).
На первом этапе
паразит переходит с местного приспособленного растения на неприспособленное восприимчивое растение-интродуцент.
В рассматриваемой патосистеме этот процесс шел в течение с 1840‑ых годов по 1940‑ые годы на томате, сначала свободном от P
.
infestans
, а потом начавшем поражаться ею.
На втором этапе
происходит мощное развитие патогена на неприспособленном восприимчивом интродуценте. В отдельные годы на последнем развиваются эпифитотии.
В рассматриваемой патосистеме этот процесс шел на томате в течение с 1950‑ых годов по 1990‑ые годы.
На третьем этапе
эпифитотии на неприспособленном восприимчивом растении-интродуценте становятся реальными. Этому могут способствовать нарушения агротехники, климатические факторы. Штаммы патогена с неприспособленного восприимчивого растения-интродуцента могут довольно активно заражать местное приспособленное растение. На местном приспособленном растении нарастает эрозия устойчивости и возникает предэфитотийная ситуация.
В рассматриваемой патосистеме этот процесс шел с конца 1990‑ых годов по настоящее время. В средней полосе России ему способствовало последовательное выращивание томата в условиях холодного климата и перенос зооспорангиев с ветром, потоками воды, а также плодовыми мушками.
На четвертом этапе
на местном приспособленном растении наступает жесткая эпифитотия. Его устойчивость преодолена. Потери достигают 80–100%, что может иметь большое экономическое значение. Защита возможна только при жестком использовании химического метода.
В рассматриваемой патосистеме без кардинальных изменений в подходе к защите томата и картофеля от фитофтороза этот сценарий может быть реализован с вероятностью 40–60% через 5–20 лет.
Стойкость неприспособленного восприимчивого растения-интродуцента к местным патогенам будет зависеть от эффективности и быстроты акклиматизации, а также вирулентности и агрессивности местных патогенов. Для родственных видов растений-хозяев имеет смысл составлять ряды патогенов, в том числе и общих для них, наподобие гомологических рядов Н.И. Вавилова. Все это актуально для России при направлении акклиматизации культур (возможно, даже на сортовом уровне) в северном направлении.
Искусственная поддержка неприспособленных восприимчивых растений-интродуцентов может усугубить положение в агроценозах. В естественных сообществах эта проблема бы не возникла, так как неприспособленные восприимчивые растения-интродуценты не перенесли эпифитотию. Рассмотренный принцип развития эпифитотий может представлять интерес для карантина растений при проведении АФР.
Возможная роль дрозофил в патосистеме «томат –
P
.
infestans
»
Дрозофил очень часто можно видеть на томатах, пораженных фитофторозом. Мы предположили, что дрозофилы могут переносить инфекционные структуры P
.
infestans
(Кузнецов, 2005; 2006; Кузнецов и др., 2006; Смирнов, Кузнецов, 2006). Эксперименты организовали таким образом, чтобы проверить возможность контактного и алиментарного способов переноса бесполых и половых структур P
.
infestans
дрозофилами.
Материалы и методы.
Дрозофил линии дикого типа Canton-S выращивали на агаризованной среде. Для получения зооспорангиев использовали 3‑недельные колонии изолятов P
.
infestans
ВалС13 и ВалС15 (Новгородская область, из коллекции РГАУ-МСХА) на ржано-овощном агаре (Caten, Jinks, 1968). Для получения ооспор на ржано-овощном агаре скрещивали изоляты из коллекции РГАУ-МСХАP
.
infestans
T25 (А1, Тульская область) и 2/20 (А2, Белоруссия). Для проверки возможности заражения зооспорангиями и / или ооспорами P
.
infestans
использовали ломтики клубней картофеля сорта Сантэ. Методы рассмотрены в работе А.Н. Смирнова и С.А. Кузнецова (Смирнов, Кузнецов, 2006).
Результаты
. В результате лабораторных модельных опытов доказали контактный способ переноса зооспорангиев.
Алиментарный способ переноса зооспорангиев дрозофилами подтвердить не удалось. Возможность переноса ооспор (за исключением единичных ооспор самой маленькой их фракции – до 20 µм) также не подтвердилась.
Обсуждение результатов
. Возможная роль дрозофилы в патосистеме «томат –
P
.
infestans
»в открытом грунте.
Часто встречаясь на томатных посадках, дрозофилы способны находить плоды томата с микротрещинами. Если хотя бы часть мушек ранее уже контактировала со спороношением на пораженных плодах, то создается возможность эффективного переноса вторичной инфекции от больных плодов к еще непораженным. Такой способ распространения фитофтороза на томате может быть сравнимым с традиционными способами распространения – воздушными токами и капельно-жидкой влагой (Новотельнова, 1974).
Дрозофилы могут преодолевать довольно большие расстояния, перенося при этом зооспорангии. Этому способствует тот факт, что температурные оптимумы развития этих двух организмов перекрываются.
Мы не исключаем того, что и некоторые другие насекомые, встречающиеся на томате, могут сходным образом переносить инфекционные структуры P
.
infestans
. Возможно, в определенных ситуациях их ограничение поможет снизить опасность фитофтороза.
Заключение
Посредством разработанной нами методологии можно анализировать соотношение бесполой и половой стадий, а также жизнеспособность и агрессивность конкретных полевых популяций P
.
infestans
. Это представляет интерес как для последующих научных исследований, так и для решения текущих практических задач.
К примеру, наша работа, в целом, не выявила существенных преимуществ А2 или А1 по агрессивности и вирулентности (последнюю изучали преимущественно в предшествующих исследованиях). Мы полагаем, что А2 может быть своеобразным биологическим резервом P
.
infestans
. В стрессовых ситуациях его частота возрастает, причем многие изоляты А2 характеризуются сбалансированностью по показателям жизнеспособности и устойчивостью к стрессовому фактору. Далее через скрещивания эти соответствующие гены в виде эффективных аллельных комбинаций передаются к А1.
Кроме того, наше исследование еще раз показало всю остроту ситуации, складывающейся с применением фунгицидов против фитофтороза картофеля и томата. Популяции P
.
infestans
способны активно приспосабливаться не только к системным, но и к контактным препаратам. Мы полагаем, что на фоне применения фениламидов оказались эффективными и миграционные, и внутрипопуляционные процессы, ведущие к замене генотипов в 1990‑ые годы. В этой связи все более очевидной становится необходимость использовать сорта картофеля с горизонтальной устойчивостью против фитофтороза.
Выводы
1. Сравнительное изучение прорастания зооспорангиев изолятов P
.
infestans
российского и мексиканского происхождления выявило следующее. Большинство исследованных изолятов P
.
infestans
характеризовались довольно высокими показателями непрямого прорастания зооспорангиев при температуре 7о
С. У большинства мексиканских штаммов P
.
infestans
прорастание зооспорангиев было выше при 25о
, чем при 20о
С. Напротив, у большинства исследованных российских изолятов P
.
infestans
оно было выше при 20о
по сравнению с 25о
С. Показано, что температура, оптимальная для прорастания зооспорангиев – штаммоспецифичный признак.
2. И в российских, и в мексиканских популяциях P
.
infestans
был выявлен широкий спектр изолятов по всем компонентам агрессивности. Агрессивность на восприимчивом сорте Сантэ в значительной степени определялась способностью к образованию зооспор при 5о
С. Напротив, многие из штаммов, агрессивных к среднеустойчивому сорту Луговской, характеризовались высокой способностью к прямому прорастанию зооспорангиев при 25о
С. Это свидетельствует о том, что способность зооспорангиев к прямому прорастанию может повышать вероятность поражения фитофторозом устойчивых сортов картофеля.
3. В Мексике, между штаммами P
.
infestans
с характеристиками «старого» и «нового» типов (А1 Gpi
86/100 и A1 Gpi
100/100 соответственно) существенных различий по агрессивности выявлено не было. Соответственно, в условиях Мексики мы не получили свидетельств того, что штаммы с «новыми» генотипами могли вытеснить штаммы со «старыми» генотипами благодаря преимуществам в агрессивности.
4. Обследование 90 полевых популяций P
.
infestans
в листьях картофеля (около 4000 образцов), а также листьях и плодах томата в 1997–2007 г. показало, что максимальное количество ооспор образовывалось в плодах томата. Часть популяций P
.
infestans
в плодах томата сочетали бесполое размножение и половой процесс, другие были бесполыми. В листьях томата и картофеля ооспоры также встречались, но, за отдельными исключениями, значительно реже. Популяции P
.
infestans
в ботве растений-хозяев были преимущественно бесполыми. В клубнях картофеля выявили лишь единичные ооспоры.
5. Ооспоры P
.
infestans
характеризовались значительной морфологической изменчивостью. По таким критериям как размер, толщина стенки, наличие антеридия и форма оогония выявили около 50 их морфологических типов. Некоторые обнаруженные морфологические типы ооспор напоминали ооспоры других фитофтор, а также пероноспоровых псевдогрибов. Были выявлены и различные стадии развития ооспор.
6. В ботве картофеля и томата образовывались в основном ооспоры негибридного происхождения, а в плодах томата – как гибридного, так и негибридного. Инфекционное значение ооспор определяются их происхождением и влиянием внешних факторов.
- Гибридные ооспоры типичной морфологии обладают высокой жизнеспособностью. Однако они почти не способны к непосредственному прорастанию. Для эффективного инфицирования растений-хозяев им нужен период покоя и воздействие внешних факторов.
- Ооспоры от самооплодотворения в природе образуются благодаря влиянию внешних факторов. Жизнеспособность ооспор от самооплодотворения – на довольно высоком уровне, но ниже гибридных ооспор.
- Партеногенетические (нетипичные) ооспоры образуются в стрессовых условиях и / или при нарушении хода полового процесса. При благоприятных условиях они легко прорастают почти без периода покоя, действие почвенных грибов может усилить прорастание. Их жизнеспособность и инфекционная спосбность довольно низка, но в определенных условиях они могут представлять существенную опасность.
7. Применение разработанного метода определения стратегий размножения и агрессивности полевых популяций P
.
infestans
после обработок фунгицидами на сортах картофеля Сантэ (восприимчивый) и Луговской (устойчивый) показало следующее.
- Ширлан и Ридомил голд были высоко эффективны при защите картофеля от фитофтороза. Применение этих фунгицидов существенно снижало репродуктивные показатели и агрессивность полевых популяций P
.
infestans
.
- Амистар показал довольно низкую эффективность против фитофтороза на картофеле. Он снижал показатели индексы развития болезни и образования зооспорангиев только на Луговском, на Сантэ после обработок индекс развития болезни даже возрастал.
- Хлорокись меди, как правило, была не эффективна при защите картофеля от фитофтороза.
- На сорте Луговской репродуктивные показатели и агрессивность P
.
infestans
были ниже, чем на сорте Сантэ.
8. Большинство сортов (как голландских, так и российских), тестируемых нашим методом, имели низкий уровень полевой устойчивости к фитофторозу. Использование этих сортов, характеризующихся высокой урожайностью и товарными качествами, возможно только в случае обработок фунгицидами. Наличие R
‑генов не предотвращало значительное развитие фитофтороза на этих сортах. Лишь несколько сортов продемонстрировали довольно высокий и стабильный уровень полевой устойчивости к фитофторорзу. Это сорта Луговской, Водолей, Герань и Ветразь.
9. Наименьшей минимальной порядковой концентрацией (10-4
%), подавляющей in
vitro
большое количество изолятов P
.
infestans
, характеризовался фунгицид Акробат. Далее в порядке возрастания концентраций располагались Амистар (10-4
, 10-3
%), Ридомил (10-3
%), Браво (10-2
, 10-1
%) и Ширлан (100
%).
10. Без предварительного контакта с фунгицидом изоляты P
.
infestans
in
vitro
демонстрировали различия в чувствительности к низким (сублетальным) концентрациям фунгицида. По мере инкубации изолятов P
.
infestans
на средах с фунгицидами определенной тенденции изменения чувствительности к фунгицидам выявлено не было.Подтверждено наличие адаптивной устойчивости P
.
infestans
как к системным (в большей степени) так и контактным фунгицидам с уточнением, что адаптивная устойчивость к тем или иным фунгицидам или их комбинациям – штаммоспецифический признак.
11. По сравнению с картофелем, на томате в средней полосе России сложилась очень тяжелая фитосанитарная обстановка в связи с поражением фитофторозом: очень жесткие эпифитотии, регулярное образование гибридных ооспор. Часть популяции P
.
infestans
на томате характеризуется высокой вредоносностью не только к томату, но также и к картофелю. Томат представляет собой опаснейший источник заражения для картофеля, повышает вероятность эрозии полевой устойчивости эффективных сортов картофеля.
Практические рекомендации
Разработанную нами методологию по определению стратегий размножения и поддержания жизнеспособности полевых популяций возбудителя фитофтороза картофеля и томата целесообразно использовать специалистам-фитопатологам в как федеральных, так и региональных сельскохозяйственных центрах различного уровня и статуса, связанных с защитой растений.
В научных учреждениях результаты, получаемые посредством нашей методологии, могут быть очень существенным дополнением к результатам современных популяционных исследований P
.
infestans
на основе определения типов спаривания, устойчивости к фунгицидам и патогенности с использованием молекулярно-генетических маркеров (PCR, RAPD, фингерпринты ДНК и др.)
Нашу методику можно использовать при определении полевой устойчивости сортов картофеля и биологической эффективности новых фунгицидов, поступающих в производство.
По нашей методике мы рекомендуем проведение учетов развития фитофтороза на картофеле в хозяйствах в течение вегетационного сезона для определения поражения ботвы, а следовательно получения важных первичных данных для прогнозирования поражения клубней фитофторозом как одного из факторов их лежкости.
Кроме того, наша методология позволит оценить запасы и инфекционную роль ооспор в растительных остатках. В этой связи, результаты текущих учетов, сделанных по нашему методу во второй половине августа – начале сентября, могут быть полезны при уточнении планирования посадок картофеля в последующие годы из-за возможности их первичного инфицирования возбудителем фитофтороза.
Список опубликованных работ по теме диссертации в журналах, рекомендованных вак рф (по профилю «Биологические науки»)
1. Осман А.Д. Сравнительная оценка действия фунгицида амистар на агрономически важные микроорганизмы и некоторые штаммы Phytophthora
infestans
/ А.Д. Осман, В.Т. Емцев, В.А. Калинин, А.Н. Смирнов, К.В. Быков // Известия ТСХА. – 1999. – Вып. 1. – С. 139–145.
2. Смирнов А.Н. Образование ооспор в полевых популяциях гриба Phytophthora
infestans
в Московской области / А.Н. Смирнов, С.Н. Еланский // Микология и фитопатология. – 1999. – Т. 33. – С. 421–425.
3. Еланский С.Н. Популяции Phytophthora
infestans
в Московской области I. Системы размножения / С.Н. Еланский, А.Н. Смирнов, А.В. Долгова, Ю.Т. Дьяков // Микология и фитопатология. – 1999. – Т. 33. – Вып. 5. – С. 346–352.
4. Еланский С.Н. Популяции Phytophthora
infestans
в Московской области II. Сравнительная структура популяций, паразитирующих на картофеле и томатах / С.Н. Еланский, А.Н. Смирнов, С.Ф. Багирова, Ю.Т. Дьяков // Микология и фитопатология. – 1999. – Т. 33. – Вып. 5. – С. 353–360.
5. Смирнов А.Н. Встречаемость и морфология ооспор, обнаруженных в природных популяциях Phytophthora
infestans
в Московской области в 1999 году / А.Н. Смирнов, С.А. Кузнецов // Известия ТСХА. – 2001. – Вып 4. – С. 116–133.
6. Смирнов А.Н. Определение происхождения ооспор в полевых популяциях гриба Phytophthora
infestans
/ А.Н. Смирнов // Известия ТСХА. – 2001. – Вып. 1. – С. 141–153.
7. Смирнов А.Н. Ооспоры Phytophthora
infestans
/ А.Н. Смирнов // Микология и фитопатология. – 2003а. – Т. 37. – С. 3–21.
8. Смирнов А.Н. Оценка происхождения ооспор в природных популяциях Phytophthora
infestans
Московской области / А.Н. Смирнов // Известия ТСХА. – 2003b. – Вып. 4. – С. 87–95.
9. Филиппов А.В. Горизонтальная устойчивость листьев картофеля к Phytophthora
infestans
и агрессивность изолятов патогена из разных географических регионов / А.В. Филиппов, Б.И. Гуревич, М.А. Кузнецова, А.Н. Рогожин, С.Ю. Спиглазова, А.С. Кравцов, Т.И. Сметанина, А.Н. Смирнов // Микология и фитопатология – 2004. – Т. 38. – С. 74–88.
10.Смирнов А.Н. Характеристики мексиканских штаммов P
.
infestans
из долины Толука при различных температурах / А.Н. Смирнов // Известия ТСХА. – 2006. – №2. – С. 66–73.
11.Смирнов А.Н. Определение стратегий размножения и жизнеспособности полевых популяций Phytophthora
infestans
/ А.Н. Смирнов, С.А. Кузнецов // Известия ТСХА. – 2006b. – Вып. 4. – С. 28–41.
12.Смирнов А.Н. Характеристика мексиканских штаммов Phytophthora
infestans
из долины Толука. I. Агрессивность / А.Н. Смирнов // Микология и фитопатология. – 2007b. – Т. 41. – С. 89–103.
13.Смирнов А.Н. Характеристика мексиканских штаммов Phytophthora
infestans
из долины Толука. II. Прорастание зооспорангиев / А.Н. Смирнов // Микология и фитопатология. – 2007c. – Т. 41. – С. 383–391.
14.Троицкая Ю.С. Отношение к фунгицидам и агрессивность изолятов фитофторы / Ю.С. Троицкая, А.С. Черняховская, А.Н. Смирнов // Агро XXI. – 2007. – №10–12. – С. 32–33.
15.Смирнов А.Н. Изменение чувствительности изолятов Phytophthora
infestans
к фунгицидам в зависимости от возраста мицелия и контакта с фунгицидами / А.Н. Смирнов, Ю.С. Троицкая // Микология и фитопатология. – 2008. – Т. 42. – Вып. 3. – С. 287–299.
16.Аль-Саади М.Н.С. Влияние экстрактов растительного происхождения на возбудителя фитофтороза картофеля / М.Н.С. Аль-Саади, А.Н. Смирнов, Ю.И. Помазков // Известия ТСХА. – 2009. – Вып 1. – С. 136–144.
17.Ощепкова Ю.И. Выделение липидпереносящего белка Ns-LTP1 из семян чернушки посевной (Nigella
sativa
) / Ю.И. Ощепкова, О.Н. Вешкурова, Е.А. Рогожин, А.Х. Мусолямов, А.Н. Смирнов, Т.И. Одинцова, Ц.А. Егоров, Е.В. Гришин, Ш.И. Салихов // Биоорганическая химия. – 2009. – Т. 35.
Список остальных работ, опубликованных по теме диссертации
18.Смирнов А.Н. Роль ооспорообразования в изменчивости подмосковных популяций Phytophthora
infestans
(Mont.) deBary. / А.Н. Смирнов, С.Н. Еланский // Сборник конференции «Современные проблемы микологии, альгологии и фитопатологии». – М.: МГУ, 1998. – С. 109–111.
19.Смирнов А.Н. Ооспоры Phytophthora
infestans
в плодах томатов в Московской области / А.Н. Смирнов, С.Н. Еланский, А.В. Долгова // Защита и карантин растений. – 1998. – №5. – С. 41.
20.Smirnov A.N. Report on work conducted in the Toluca Valley during May 2 – October 24 of 1998. / A.N. Smirnov // Cornell-Eastern Europe-Mexico (CEEM) International Collaborative Project in Potato Late Blight Control. Progress Report, July 1999. – Appendix 12.
21.Дьяков Ю.Т. Мезоэволюция Phytophthora
infestans
в агроценозах / Ю.Т. Дьяков, С.Ф. Багирова, А.В. Долгова, С.Н. Еланский, А.Н. Смирнов // Х Московское совещание по филогении растений. Материалы. – М., 1999. –C. 71–73.
22.Смирнов А.Н. Ооспоры гриба Phytophthora
infestans
(Mont.) deBary в Московской области / А.Н. Смирнов // Тезисы VII молодежной конференции ботаников в Санкт-Петербурге, 15–19 мая, 2000 г. – 2000. – С. 83.
23.Кузнецов С.А. Морфология и встречаемость зигот гриба Phytophthora
infestans
(Mont.) deBary, обнаруженных на территории Московской области в 1999 году / С.А. Кузнецов, А.Н. Смирнов // Сборник студенческих научных работ. М.: Издательство МСХА. – 2000. – Вып. 6. – С. 14–21.
24.Смирнов А.Н. Встречаемость и возможное происхождение ооспор Phytophthora
infestans
в Московской области в 1999 г. / А.Н. Смирнов, А.С. Кравцов, С.А. Кузнецов, В.П. Апрышко, С.Н. Еланский // Материалы научной конференции «Памяти Грегора Менделя». – М: Издательство МСХА. – 2001. – С. 122–123.
25.Смирнов А.Н. Происхождение ооспор Phytophthora
infestans
в пораженных фитофторозом образцах в Московской области / А.Н. Смирнов, С.А. Кузнецов, А.С. Кравцов, В.П. Апрышко, М.А. Побединская, С.Н. Еланский // Материалы международного симпозиума «Проблемы изучения и сохранения биоразнообразия европейских природных сообществ. – 2001. – С. 135.
26.Смирнов А.Н. Изучение биологии возбудителя фитофтороза картофеля / А.Н. Смирнов, С.А. Кузнецов, С.Н. Еланский // Доклады ТСХА. – 2001. – Вып. 273. – Часть 1 – С. 226–231.
27.Elansky S. Genotypic analysis of Russian isolates of Phytophthora infestans
from the Moscow region, Siberia and Far East / S. Elansky, A. Smirnov, Y. Dyakov, A. Dolgova, A. Filippov, B. Kozlovsky, I. Kozlovskaya, P. Russo, C. Smart, W. Fry // J. Phytopathol. – 2001. – V. 149. – P 605–611.
28.Еланский С. Н, Популяции Phytophthora
infestans
в Европейской части России / С.Н. Еланский, А.Н. Смирнов, А.С. Кравцов, Ю.Т. Дьяков, И.Н. Козловская, Б.Е. Козловский, Е.В. Морозова, Ф.Х. Аматханова // Материалы Первой Всероссийская конференции по иммунитету растений к болезням и вредителям. – Санкт-Петербург, 2002. – С. 81–82.
29.Филиппов А.В. Сравнение изолятов Phytophthora
infestans
из различных регионов по агрессивности к картофелю / А.В. Филиппов, М.А. Кузнецова, С.Ю. Спиглазова, Т.И. Сметанина, А.Н. Смирнов, А.С. Кравцов // Материалы Первая Всероссийская конференция по иммунитету растений к болезням и вредителям. – Санкт-Петербург, 2002. – С. 125–126.
30.Smirnov A.N. Investigations of oospores of Phytophthora infestans
in Moscow region / A.N. Smirnov, S.A. Kuznetsov, S.N. Elansky // The 7th
International Mycological Congress, Oslo, 11–17 August, 2002. – №892. – P. 268–269.
31.Смирнов А.Н. Способность экологического фактора влажность ограничивать образование ооспор P
.
infestans
/ А.Н. Смирнов, О.Н. Рудакова // Доклады ТСХА. – 2003. – Вып. 275. – С. 136–137.
32.Уланова Т.И. Устойчивость к фитофторозу некоторых перспективных линий диких Lycopersicon
hirsutum
/ Т.И. Уланова, С.Н. Еланский, А.В. Филиппов, Ю.Т. Дьяков, В.П. Апрышко, Б.Е. Козловский, А.Н. Смирнов, М.Д. Кофей // J. RussianPhytopathol. Soc. – 2003. – V. 4. P. 9–15.
33.Elansky S.N. Second locus of Peptidase as a marker for genetic investigations of Phytophthora infestans
/ S.N. Elansky, A.N. Smirnov // Botanica Lithuanica. – 2003. – V. 9. – P. 275–283.
34.Смирнов А.Н. К вопросу о фитофторозе на томате в некоторых областях Российской Федерации / А.Н. Смирнов // Интегрированная защита с.х.культур и фитосанитарный мониторинг в современном земледелии. Материалы всероссийской научно-практической конференции, СГАУ – 75 лет. – 2004. – С. 119–121.
35.Троицкая Ю.С. Отношение изолятов Phytophthora
infestans
из географически различных популяций к фунгицидам ридомил, ширлан и амистар / Ю.С. Троицкая, Е.Н. Федулова, А.Н. Смирнов // Сборник материалов юбилейной конференции, посвящённый 85‑летию кафедры микологии и альгологии Московского Государственного Университета им. М.В. Ломоносова. – 2004. – С. 137.
36.Троицкая Ю.С. Изменение чувствительности изолятов Phytophthora
infestans
к фунгицидам в результате последовательного использования и чередования фунгицидов / Ю.С. Троицкая, А.Н. Смирнов // Второй всероссийский съезд по защите растений. Санкт-Петербург. Материалы съезда, 2005, 5–10 декабря. – Т. 1. – С. 227–229.
37.Шкаликов В.А. – Иммунитет растений. / В.А. Шкаликов, Ю.Т. Дьяков, А.Н. Смирнов и др // – М.: КолосС, 2005. – 188 с.
38.Кузнецов С.А. Дрозофила как переносчик инфекционных структур Phytophthora
infestans
/ С.А. Кузнецов, О.В. Кузнецова, А.Н. Смирнов // Грибы и водоросли в биоценозах. Москва, МГУ. Материалы конференции, 2006, 31 января – 3 февраля. – С. 146–149.
39.Смирнов А.Н. Адаптивная устойчивость изолятов Phytophthora
infestans
к некоторым современным фунгицидам / А.Н. Смирнов, Ю.С. Троицкая // Грибы и водоросли в биоценозах. Москва, МГУ. Материалы конференции, 2006, 31 января – 3 февраля. – С. 146–149.
40.Кваснюк Н.Я. Интегрированная система защиты картофеля от грибных, вирусных и бактериальных болезней. / Н.Я. Кваснюк, Б.И. Гуревич, К.А. Можаева, Н.В. Гирсова, Е.В. Матвеева, А.Н. Игнатов, Л.Н. Жеребцова, Е.И. Филиппова, Б.В. Анисимов, Е.А. Симаков, А.Н. Смирнов, В.И. Черкашин // Практическое руководство. – М.: ФГНУ «Росинфорагротех», 2006. – 71 с.
41.Смирнов А.Н. Определение стратегий размножения и жизнеспособности полевых популяций Phytophthora
infestans
/ А.Н. Смирнов, С.А. Кузнецов // Защита и карантин растений. – 2006. – №9. – С. 30–31.
42.Смирнов А.Н. Фитофтороз томата / А.Н. Смирнов, С.А. Кузнецов // Защита и карантин растений. – 2006. – №3. – С. 20–23.
43.Смирнов А.Н. Взаимосвязь способов размножения видов рода Solanum
и Phytophthora
infestans
/ А.Н. Смирнов // Материалы международной конференции «Научное наследие Н.И. Вавилова – фундамент отечественного и мирового сельского хозяйства» – М.: ФГОУ ВПО РГАУ-МСХА имени К.А. Тимирязева. – 2007. – С. 182–183.
44.Антоненко В.В. Влияние регуляторов роста Новосил, Лариксин и Терпенол на заболеваемость картофеля фитофторозом и альтернариозом / В.В. Антоненко, А.Н. Смирнов // Сборник студенческих научных работ. – М.: Издательство РГАУ-МСХА. – 2008. – Вып. 14. – С. 52–56.
45.Мамонов А.Г. Изучение природных популяций Phytophthora
infestans
на устойчивость к металаксилу / А.Г. Мамонов, Н.Я. Кваснюк, А.Н. Смирнов // Сборник студенческих научных работ. – М.: Издательство РГАУ-МСХА. – 2008. – Вып. 14. – С. 56–60.
46.Рогожин Е.А. Антимикробные пептиды ежовника (Echinochloa
crusgalli
) / Е.А. Рогожин, Т.И. Одинцова, А.Н. Смирнов, Ц.А. Егоров, Е.В. Гришин // Современные проблемы иммунитета растений к вредным организмам. Вторая Всероссийская конференция. – Санкт-Петербург. – 2008. – С. 32–34.
47.Смирнов А.Н. Возможная взаимосвязь способов размножения растений-хозяев и их основных патогенов (грибов и псевдогрибов) / А.Н. Смирнов // Доклады ТСХА. – 2008. – Вып. 280. – С. 127–132.
48.Smirnov A.N. Formation and germination of Phytophthora infestans
oospores in different regions of Russia / A.N. Smirnov, S.A. Kuznetsov, N. Ya. Kvasnyuk, K.L. Deahl // Comm. Appl. Biol. Sci, Ghent University. – 2008. – V. 73/2. – P. 109–118.
49.Smirnov A.N. Strategies of reproduction of field Phytophthora infestans
populations on potato and tomato in some regions of Russia / A.N. Smirnov, S.A. Kuznetsov // / Proceedings Nat. Sci., Matica Srpska Novy Sad – Serbia. – 2009. – №116. – P. 149–157.
|
