| ДАЛЬНЕВОСТОЧНЫЙ ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ
ТИХООКЕАНСКИЙ ИНСТИТУТ
ДИСТАНЦИОННОГО ОБРАЗОВАНИЯ И ТЕХНОЛОГИЙ

А.П. Анисимов
КОНЦЕПЦИИ СОВРЕМЕННОГО ЕСТЕСТВОЗНАНИЯ. БИОЛОГИЯ
ВЛАДИВОСТОК
2000 г.
ВВЕДЕНИЕ.............................................................................................................................................................. 3
СЕГМЕНТ 1. ПРЕДМЕТ И ЗАДАЧИ БИОЛОГИИ.................................................................................... 3
СЕГМЕНТ 2. ЗНАЧЕНИЕ ОБЩЕЙ БИОЛОГИИ...................................................................................... 3
СЕГМЕНТ 3. МЕТОДЫ БИОЛОГИИ........................................................................................................... 5
СЕГМЕНТ 4. ОСНОВНЫЕ КОНЦЕПЦИИ СОВРЕМЕННОЙ БИОЛОГИИ....................................... 7
ТЕМА 1. КОНЦЕПЦИЯ СИСТЕМНОЙ МНОГОУРОВНЕВОЙ ОРГАНИЗАЦИИ ЖИЗНИ............... 8
СЕГМЕНТ 5. СИСТЕМНАЯ ОРГАНИЗАЦИЯ.......................................................................................... 8
СЕГМЕНТ 6. УРОВНИ ОРГАНИЗАЦИИ ЖИВОЙ МАТЕРИИ............................................................. 9
СЕГМЕНТ 7. МОЛЕКУЛЯРНО-ГЕНЕТИЧЕСКИЙ УРОВЕНЬ.......................................................... 10
СЕГМЕНТ 8. ОНТОГЕНЕТИЧЕСКИЙ УРОВЕНЬ................................................................................. 12
СЕГМЕНТ 9. ПОПУЛЯЦИОННО-ВИДОВОЙ УРОВЕНЬ.................................................................... 14
СЕГМЕНТ 10. БИОГЕОЦЕНОТИЧЕСКИЙ УРОВЕНЬ......................................................................... 15
ЗАКЛЮЧЕНИЕ ПО ТЕМЕ 1......................................................................................................................... 16
ТЕМА 2. КОНЦЕПЦИЯ МАТЕРИАЛЬНОЙ СУЩНОСТИ ЖИЗНИ...................................................... 17
СЕГМЕНТ 11. МЕХАНИЦИЗМ И ВИТАЛИЗМ В ИСТОРИИ БИОЛОГИИ.................................... 17
СЕГМЕНТ 12 ЖИВАЯ МАТЕРИЯ И ЕЕ ОСНОВНАЯ ФОРМА ДВИЖЕНИЯ. ОБМЕН ВЕЩЕСТВ И ЭНЕРГИИ В ЖИВОЙ СИСТЕМЕ......................................................................................................................................................... 19
СЕГМЕНТ 13. ТРАНСФОРМАЦИЯ И ИСПОЛЬЗОВАНИЕ ЭНЕРГИИ........................................... 22
Реклама

СЕГМЕНТ 14. БЕЛКИ – СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ОСНОВА ЖИЗНИ.................. 26
СЕГМЕНТ 15. ОПОРА И ДВИЖЕНИЕ...................................................................................................... 27
СЕГМЕНТ 16. ТРАНСПОРТ ВЕЩЕСТВ.................................................................................................. 28
СЕГМЕНТ 17. ФЕРМЕНТАТИВНЫЙ КАТАЛИЗ (БИОКАТАЛИЗ).................................................. 29
СЕГМЕНТ 18. ЗАЩИТНЫЕ РЕАКЦИИ. ИММУНИТЕТ.................................................................... 31
СЕГМЕНТ 19. СИГНАЛИЗАЦИЯ. ГОРМОНАЛЬНАЯ И НЕРВНАЯ РЕГУЛЯЦИЯ................... 33
ЗАКЛЮЧЕНИЕ ПО ТЕМЕ 2......................................................................................................................... 37
ТЕМА 3. КОНЦЕПЦИЯ БИОЛОГИЧЕСКОЙ ИНФОРМАЦИИ И САМОВОСПРОИЗВЕДЕНИЯ ЖИЗНИ. ОНТОГЕНЕЗ 38
СЕГМЕНТ 20. САМОВОСПРОИЗВЕДЕНИЕ - ВАЖНЕЙШЕЕ СВОЙСТВО ЖИЗНИ. ОБЩАЯ СХЕМА ОНТОГЕНЕЗА.............................................................................................................................................................................. 38
СЕГМЕНТ 21. ПРЕФОРМИЗМ И ЭПИГЕНЕЗ В ИСТОРИИ ЭМБРИОЛОГИИ............................. 39
СЕГМЕНТ 22. ГЕНОТИП И ФЕНОТИП ОРГАНИЗМА. ЦЕНТРАЛЬНАЯ ДОГМА МОЛЕКУЛЯРНОЙ БИОЛОГИИ 41
СЕГМЕНТ 23. РЕПЛИКАЦИЯ ДНК И РАЗМНОЖЕНИЕ КЛЕТОК................................................. 43
СЕГМЕНТ 24. ФОРМЫ РАЗМНОЖЕНИЯ ОРГАНИЗМОВ. КЛОНИРОВАНИЕ.......................... 46
СЕГМЕНТ 25. РАЗВИТИЕ ОРГАНИЗМА................................................................................................ 50
ЗАКЛЮЧЕНИЕ ПО ТЕМЕ 3......................................................................................................................... 55
ТЕМА 4. КОНЦЕПЦИЯ САМОРЕГУЛЯЦИИ ЖИВЫХ СИСТЕМ......................................................... 57
СЕГМЕНТ 26. САМОРЕГУЛЯЦИЯ И ГОМЕОСТАЗ............................................................................ 57
СЕГМЕНТ 27. ВНУТРИКЛЕТОЧНАЯ САМОРЕГУЛЯЦИЯ.............................................................. 58
СЕГМЕНТ 28. САМОРЕГУЛЯЦИЯ МНОГОКЛЕТОЧНОГО ОРГАНИЗМА................................. 61
СЕГМЕНТ 29. САМОРЕГУЛЯЦИЯ В ЭКОСИСТЕМАХ..................................................................... 65
ЗАКЛЮЧЕНИЕ ПО ТЕМЕ 4......................................................................................................................... 73
Реклама

ТЕМА 5. КОНЦЕПЦИЯ САМООРГАНИЗАЦИИ И БИОЛОГИЧЕСКОЙ ЭВОЛЮЦИИ. ФИЛОГЕНЕЗ 75
СЕГМЕНТ 30. ПРОБЛЕМА САМООРГАНИЗАЦИИ И НАУКА СИНЕРГЕТИКА........................ 75
СЕГМЕНТ 31. ВОЗНИКНОВЕНИЕ ЖИЗНИ НА ЗЕМЛЕ................................................................... 77
СЕГМЕНТ 32. ЭТАПЫ РАЗВИТИЯ ЖИЗНИ НА ЗЕМЛЕ И СОВРЕМЕННОЕ БИОРАЗНООБРАЗИЕ 82
СЕГМЕНТ 33. ФАКТОРЫ БИОЛОГИЧЕСКОЙ ЭВОЛЮЦИИ............................................................ 91
СЕГМЕНТ 34. ПРОИСХОЖДЕНИЕ И ЭВОЛЮЦИЯ ЧЕЛОВЕКА.................................................... 97
ЗАКЛЮЧЕНИЕ К ТЕМЕ 5.......................................................................................................................... 102
РЕКОМЕНДУЕМАЯ ЛИТЕРАТУРА............................................................................................................ 103
ВВЕДЕНИЕ
Сегмент 1. Предмет и задачи биологии
Сегмент 2. Значение общей биологии
Сегмент 3. Методы биологии
Сегмент 4. Основные концепции современной биологии
СЕГМЕНТ 1.
ПРЕДМЕТ И ЗАДАЧИ БИОЛОГИИ
Биология
- совокупность наук о жизни, о живой природе (греч. bios - жизнь, logos - учение). Современная биология - очень разнообразная и развитая область естествознания. Различают ряд частных биологических наук
по объектам исследования, такие как зоология (о животных), ботаника (о растениях), микробиология (о бактериях), вирусология (о вирусах), и другие, еще более мелкие подразделения (орнитология - о птицах, ихтиология - о рыбах, альгология - о водорослях и т.д.). Другое подразделение биологических наук - по уровням организации и свойствам живой материи: молекулярная биология и биохимия (химические основы жизни), генетика (наследственность), цитология (клеточный уровень), эмбриология, биология развития (индивидуальное развитие организмов), анатомия и физиология (строение и принципы функционирования организмов), экология (взаимоотношения организмов с окружающей средой), теория эволюции (историческое развитие живой природы).
Живой мир очень многообразен. Существует около 2 млн видов животных, около 500 тыс. видов растений, сотни тысяч грибов, тысячи видов и еще больше штаммов (разновидностей) бактерий. Многие виды еще не описаны. Структурная сложность, типы питания, жизненные циклы, исторический возраст этих групп организмов очень сильно различаются (сравните хотя бы организацию и образ жизни человека и его домашних спутников - таракана, комнатного растения, микробов и вирусов). Но все организмы должны иметь нечто общее, что отличало бы их от неживой природы. Это - обмен веществ и энергии, способность к размножению и развитию, изменчивость и адаптивная эволюция. Выявлением и характеристикой этих общих свойств живых организмов и их системных комплексов с неживой природой занимается так называемая общая биология
. По сути перед общей биологией стоит задача познать сущность жизни, ответить на вопрос - что есть жизнь. Именно эта общая концептуальная часть биологии предлагается в современной модели гуманитарного образования. Для чего это нужно?
СЕГМЕНТ 2. ЗНАЧЕНИЕ ОБЩЕЙ БИОЛОГИИ
Теоретическое и гуманитарное значение
общей биологии состоит прежде всего в формировании материалистического мировоззрения
. Основной вопрос философии - о соотношении материи (бытия) и сознания - по сути вопрос биологический. От выбора позиции (что первично - материя или сознание) складывается либо материалистическое, либо идеалистическое понимание природы и общества, формируются принципиально разные подходы в пользовании объектами природы, в оценке социальных явлений, в выработке политических стратегий. К сожалению, многие политики и даже философы с необыкновенной легкостью отдают свои предпочтения различным (часто просто модным) идеалистическим построениям, порой даже не задаваясь вопросом о том, что такое материя. Развитие реальной демократии и свободы совести в нашей стране породили волну совершенно неосмысленного обращения людей к мистике, астрологии и прочим маргинальным проявлениям культуры. В то время как огромный массив накопленных реальных научных знаний остается для большинства населения неизвестным и невостребованным. Задачи средней школы в этом плане выполняются с низкой эффективностью. Поэтому общее естественнонаучное просвещение студентов гуманитарных специальностей стало актуальной задачей современного образования именно в плане становления научного мировоззрения.
Другая гуманитарная задача биологии состоит в формировании у современного человека экологического мышления
, суть которого заключается в осознании себя частью природы и понимании необходимости охранять и рационально использовать природные ресурсы. Актуальность задачи несомненна, если учесть, что по некоторым прогнозам нынешние темпы и технологии промышленного освоения Земли уже через 50-100 лет приведут к необратимым изменениям среды обитания человечества. Это означало бы постепенное вымирание человека и большинства других объектов живой природы как биологических видов (что случилось, например, с динозаврами) и, в лучшем случае, замещение современных экологических сообществ новыми, более приспособленными к измененной среде обитания. Таким образом, понимание основ биологии и экологии необходимо каждому человеку и в особенности его технократической, гуманитарной и политической элите с целью сохранения и устойчивого развития биосферы Земли.
Практическое значение
биологии состоит в том, что она является научной основой всех технологий производства продовольствия
. Возможности экстенсивного воспроизводства продуктов питания на Земле практически исчерпаны. Целинные земли России и Казахстана, освоенные в 50-е и 60-е годы нашего столетия, явились чуть ли не последними резервами пахотных земель. Огромные площади ежегодно выводятся из сельскохозяйственного использования в результате их засоления, опустынивания, превращения в дно искусственных водоемов при строительстве гидроэлектростанций. По этим причинам современное сельское хозяйство обречено развиваться на основе интенсивных технологий. Простое возделывание овощей или пшеницы, выращивание скота, птицы и т.п. требует знания условий и динамики их размножения и роста, особенностей минерального и органического питания, совместимости с другими культурами, отношения к сорнякам, паразитам, бактериям и вирусам, которыми буквально кишит наша общая среда обитания. Особое значение в 20 веке приобрели методы генетических модификаций и селекции объектов сельскохозяйственного производства. Выведение новых пород животных и сортов растений, приспособленных к конкретным местным условиям - давняя практика. Но современная селекция не может базироваться на основе проб и ошибок, она использует точные, математизированные законы генетики. В процветающих фермерских хозяйствах США и других развитых стран селекционно-генетическая работа столь же обычна и обязательна, как и ежедневная уборка коровника или прополка грядок. Генетик здесь одна из самых востребованных профессий. В последние годы быстрыми темпами развиваются и новые биотехнологии, основанные на генной и клеточной инженерии, клонировании, получении трансгенных (с пересаженными генами), или генетически модифицированных (GM), организмов. Освоенные вначале на бактериях, эти методы уже используются для получения химерных животных и растений с заранее спланированными свойствами. И хотя GM-технологии в растениеводстве и животноводстве встречают у потребителей настороженный прием, по сути речь идет о биотехнологической революции, о формировании новой культуры и практики природопользования. И все эти вопросы находятся в поле исследования современной биологии.
Совершенно особое гуманитарно-практическое значение
имеет биология как теоретическая основа медицины
. Причины и механизмы большинства патологий (болезней) кроются в нарушениях работы генов и их продуктов - клеточных белков. Понять эти причины и механизмы - значит наполовину решить и проблему их устранения или лечения больного человека. Взаимодействие клеток с вирусами, сожительство с бактериями, формирование иммунитета к новым и новым антигенам, возникновение неконтролируемого ракового роста клеток, молекулярная природа памяти, развитие наркозависимости, причины старения ... - это огромный и нескончаемый перечень проблем, решаемых сегодня медико-биологической наукой. Отдельной главой стоит производство современных лекарств, в котором химики-фармацевты все более уступают место молекулярно-клеточным биологам. Геннно-клеточные инженерные технологии способны дать экологически и генетически чистые лекарства, а пересаженные гены могут вообще устранить хроническую болезнь, например, сахарный диабет.
В последние годы впрямую встала и проблема искусственного производства человека. Искусственное оплодотворение (при необходимости преодолеть мужское бесплодие) - давно и успешно решаемая задача. Но появилась принципиально новая технология зачатия и размножения путем клонирования потомства вообще без мужских половых клеток. Пока это сделано на животных (в Японии с 1990 г. выводят клонированных коров, в Великобритании получена знаменитая овечка по кличке Долли), но и в отношении человека методических препятствий для клонирования уже нет. Зато возникает масса чисто гуманитарных, этических и даже юридических проблем, решать которые можно имея хотя бы общее понимание биологического существа дела.
СЕГМЕНТ 3. МЕТОДЫ БИОЛОГИИ
Говоря о методах науки в широком смысле, имеют в виду не конкретные технологические приемы (методики), а методологические
принципы
, подходы к изучению объектов, явлений, их связей. В общем методы биологии те же, что и в других естественных науках.
Процесс научного познания принято разделять на две стадии: эмпирическую и теоретическую. Это разделение не абсолютно, так как эмпирическая стадия всегда развивается на основе предсуществующих теорий или гипотез, а на теоретической стадии обычно возникает необходимость в эмпирической проверке выдвигаемых новых гипотез.
На эмпирической стадии
используются следующие методы
.
Наблюдение
- изучение объектов живой природы в естественных условиях существования
. Это - непосредственное наблюдение (в буквальном смысле) за поведением, расселением, размножением животных и растений в природе, визуальное или инструментальное определение характеристик организмов, их органов, клеток, химический анализ состава и обмена веществ. Для этих целей в современной биологии применяют как традиционные средства полевых исследований - от бинокля до глубоководных аппаратов с видеокамерами ночного видения, так и сложное лабораторное оборудование - микроскопы, в том числе спектральные и электронные, биохимические анализаторы, радиоактивные метки, ультрацентрифуги, разнообразную измерительную аппаратуру.
Экспериментальный метод
(опыт) предполагает исследования живых объектов в условиях экстремального действия факторов среды
- измененной температуры, освещенности или влажности, повышенной нагрузки, токсичности или радиоактивности, измененного режима или места развития (удаление или пересадка генов, клеток, органов, интродукция животных и растений, космические полеты и т.п.). Экспериментальный метод позволяет выявить скрытые свойства, потенции, пределы адаптивных (приспособительных) возможностей живых систем, степень их гибкости, надежности, изменчивости.
Сравнительный (исторический) метод
выявляет эволюционные преобразования биологических видов и их сообществ
. Сопоставляют анатомическое строение, химический состав, структуру генов и другие признаки у организмов разного уровня сложности. При этом исследуются не только ныне живущие организмы, но и давно вымершие, сохранившиеся в виде окаменелых останков в палеонтологической летописи.
Любой из названных подходов требует количественного учета и математического описания структур и явлений. Биология все более становится точной наукой, хотя выявляемые в ней закономерности носят обычно вероятностный характер и описываются методами вариационной статистики. Это означает, что то или иное событие не строго детерминировано (предопределено), а ожидается с той или иной степенью вероятности. На основе выявляемых статистических закономерностей можно осуществлять математическое моделирование
биологических процессов и прогноз их развития
. Например, можно построить модель состояния жизни в водоеме через энное время при изменении одного, двух или более параметров (температуры, концентрации солей, наличия хищников и др.). Такие приемы стали возможны благодаря проникновению в биологию идей и принципов кибернетики - науки об управлении.
Системный метод
, как и кибернетический подход, относится к категории новых междисциплинарных методов исследования
. Живые объекты рассматриваются как
системы
, то есть совокупности элементов с определенными отношениями.
С учетом иерархичности живых систем каждый объект может рассматриваться одновременно как система и как элемент системы более высокого порядка. Поэтому принципы системной организации справедливы для всех уровней - от макромолекул до биосферы Земли
.
Широкое развитие системного движения в современной науке, в том числе и в биологии, означает постепенный переход от анализа к синтезу
.
Анализ
- это дискретный подход, углубление в структуру и функции отдельных элементов системы - внутри клетки, внутри организма, внутри экологического сообщества. Синтез
означает интегративный подход, изучение целостных характеристик системы - клетки, организма, биоценоза. Исследование всегда совершается сначала от общего к частному - анализ, а потом от частного к общему, но на новом уровне познания этого общего - синтез. С аналитическим подходом в биологии связаны открытия химической и микроструктурной организации живых объектов, выяснение видового разнообразия среди животных, растений, микроорганизмов, выявление генетической неоднородности организмов внутри популяций и другие внутренние характеристики систем. Постепенно объем накопленных аналитических данных становился достаточным для перехода к их синтезу. Так возникли синтетическая теория эволюции, нейро-гуморальная физиология, современная иммунология, молекулярно-клеточная биология, новая мегасистематика организмов, основанная на их комплексной характеристике - от экологии и анатомии до молекулярной генетики. Решается актуальная задача современного естествознания - создание целостной биологической картины мира.
Повышение интереса к синтезу в науке свидетельствует о переходе от эмпирической к теоретической стадии познания
. От получения фактов
, через их обобщение начинается выдвижение новых гипотез
, далее обычно следует их повторная эмпирическая проверка
(новые наблюдения, эксперименты, сравнения, моделирования). Эмпирическая проверка ведет либо к опровержению гипотезы, либо к ее подтверждению с той или иной степенью вероятности. Высоко достоверные гипотезы становятся законами
, из них слагаются теории
. Но и эти законы, теории носят относительный характер, так как рано или поздно могут быть пересмотрены.
Материал нашего пособия как раз и содержит такие общие теоретические, концептуальные положения современной биологии, хотя в определенной мере мы будем приводить и их эмпирические обоснования. Для начала сформулируем эти положение в общем виде, чтобы яснее были видны конечные цели и пути нашего экскурса в общую биологию.
СЕГМЕНТ 4. ОСНОВНЫЕ КОНЦЕПЦИИ СОВРЕМЕННОЙ БИОЛОГИИ
Та или иная естественнонаучная концепция
представляет некую взаимосвязанную группу понятий, гипотез, теорий, объясняющих какое-нибудь фундаментальное явление или свойство природы. Основные биологические концепции
объясняют феномен и свойства жизни
. К настоящему времени еще не сложилась какая-либо определенная система биологических концепций. Разные авторы формулируют и комбинируют их по-разному, хотя суть от этого обычно не страдает. Мы предлагаем свой вариант из пяти концептуальных обобщений современной биологии.
1. Концепция системной многоуровневой организации жизни
: все живые объекты являются системами разного уровня сложности. Биологические системы образуют непрерывную иерархию уровней структурно-функциональной организации.
2. Концепция материальной сущности жизни
: жизнь материальна, ее физико-химическую основу составляет обмен веществ и энергии. В философском смысле это означает первичность материи и вторичность сознания (материализм).
3. Концепция биологической информации и самовоспроизведения жизни
: живые организмы воспроизводятся на основе собственной (генетической) информации при взаимодействии с внешней (эпигенетической) информацией. Результатом этого взаимодействия является индивидуальное развитие организмов (онтогенез).
4. Концепция саморегуляции живых систем
: живые системы поддерживают относительное постоянство своих внутренних связей и условий функционирования (гомеостаз) на основе сочетания прямых положительных и обратных отрицательных связей.
5. Концепция самоорганизации и биологической эволюции
: живой мир возник в результате самоорганизации из неживых химических систем и претерпевает необратимое историческое развитие (филогенез) на основе наследственной изменчивости и естественного отбора популяций организмов, наиболее приспособленных к меняющимся условиям среды.
Предложенная схема не является перечнем биологических дисциплин и в этом смысле не соответствует традиционному построению учебников по обшей биологии (наиболее типичный план включает последовательно главы по биохимии и молекулярной биологии, цитологии, биологии индивидуального развития, генетике, теории эволюции, экологии). Формулируя концепции, мы использовали синтетический подход, хотя некоторые из них базируются преимущественно на одной-двух ключевых дисциплинах. Учащимся предлагается самим, используя общедоступную учебную литературу и комбинируя информацию из разных глав, найти дополнительный материал по предложенным концепциям современной биологии.
ТЕМА 1. КОНЦЕПЦИЯ СИСТЕМНОЙ МНОГОУРОВНЕВОЙ ОРГАНИЗАЦИИ ЖИЗНИ
Сегмент 5. Системная организация.
Сегмент 6. Уровни организации живой материи.
Сегмент 7. Молекулярно-генетический уровень.
Сегмент 8. Онтогенетический уровень.
Сегмент 9. Популяционно-видовой уровень.
Сегмент 10. Биогеоценотический уровень.
Заключение по теме 1.
СЕГМЕНТ 5. СИСТЕМНАЯ ОРГАНИЗАЦИЯ
В первой половине 20 века была создана общая теория систем
. Основным ее автором является австрийский биолог и философ Людвиг фон Берталанфи (1901-1972; основные труды - 1931-1968).
Согласно общей теории систем все объекты природы и общества являются системами
. Системы бывают космические, физические, технические, биологические, социальные, экономические и др.
Все многообразие природных объектов принято делить на микромир
- атомы и их элементарные частицы, макромир
- от молекул до материков и океанов и мегамир
- космические объекты и их системные объединения. Живые системы относятся к макромиру
. Что же такое система?
Система
- это совокупность элементов
, связанных определенными отношениями
и взаимодействующих по определенным законам композиции.
Вся совокупность структурных и функциональных отношений и взаимодействий составляет организацию
системы
. Организация системы как правило иерархична
, то есть имеет несколько соподчиненных уровней
. Живые объекты - типичные системы, имеющие структурную и функциональную упорядоченность, то есть определенную организацию и иерархию.
В категориях философии понятия «элемент» и «система» соотносятся как часть и целое
. Общефилософский закон определяет, что целое больше суммы его частей
. Это означает, что совокупность и взаимодействие частей (элементов) создает у целого(системы) некоторые новые качества
, отсутствующие у исходных частей (элементов). Такие новые качества, свойства определяются как эмерджентные свойства системы
(в буквальном смысле - вновь появляющиеся, непредвиденные свойства). В случае с биологическими системами эти новые качества представляют собой разные проявления жизни
. И как мы увидим далее, сама жизнь - это эмерджентное, качественно новое свойство, появляющееся на определенном уровне системной организации
.
Теоретически системы могут быть открытыми или закрытыми
- в зависимости от того, открыты или закрыты они для обмена с внешней средой веществом, энергией и информацией. Современное естествознание не находит абсолютно закрытых систем, хотя степень открытости безусловно варьирует. Таким образом, системный подход предполагает единство открытой системы
с окружающей средой
через посредство полупроницаемой пограничной поверхности
(рис.1). Понятие «пограничной поверхности» очень условно, так как может выражать совершенно различную по природе форму изоляции. Например, оболочка клетки или обшивка корабля выполнены из смеси или сплава химических веществ (белково-липидная пленка, стальные листы), птичья стая или студенческая группа объединены неуловимыми отношениями взаимопомощи или письменным распоряжением декана, солнечная система никаких оболочек не имеет, но удерживает свои границы за счет гравитационного поля.

Как уже отмечено, биологические системы отвечают всем общесистемным характеристикам - структурным, функциональным, топологическим и др. Они как правило хорошо структурированы, функционально специализированы, географически или экологически изолированы и представляют сложные многоуровневые иерархии.
СЕГМЕНТ 6. УРОВНИ ОРГАНИЗАЦИИ ЖИВОЙ МАТЕРИИ
Живая материя представляет иерархию взаимосвязанных и взаимоподчиненных уровней организации.
Иначе говоря - жизнь имеет многоуровневую организацию.
Между прочим, это означает, что любая система может рассматриваться как элемент более высокого уровня организации и, наоборот, элемент представляет систему для более низких уровней организации. То есть каждый уровень является одновременно и системой и элементом. Например, человек как организм является системой, состоящей из элементов-органов, и в то же время он сам является элементом - членом определенной популяции людей. Такой подход справедлив к любому живому объекту.
В целом же принято рассматривать четыре уровня организации живых систем
, что в значительной степени условно, так как в них можно выделить множество подуровней (табл. 1).
Таблица 1 (к сегменту 6).
Уровни и подуровни организации живых систем
| Уровни
|
Подуровни
|
| Молекулярно-генетический
|
Органическая молекула
Макромолекула, в том числе ген
Макромолекулярный комплекс,
в том числе вирус
Органоид клетки
|
| Онтогенетический
|
Клетка
Ткань
Орган
Организм
|
| Популяционно-видовой
|
Популяция
Вид
|
| Биогеоценотический
|
Сообщество, биоценоз
Биогеоценоз
Биосфера
|
Обозначенные в таблице уровни и подуровни представляют так называемые логические системы
, они отражают сложность и иерархию структурно-функциональной организации биосистем в настоящее время. Кроме того, можно выделить исторические системы
, условные объединения организмов, начиная с популяций, отражающие историю их происхождения и развития в ходе эволюции. Это - вид, род, семейство, отряд (порядок), класс, тип (отдел), царство, империя.
Дадим краткую характеристику структурно-функциональных (логических) уровней организации живых систем.
СЕГМЕНТ 7. МОЛЕКУЛЯРНО-ГЕНЕТИЧЕСКИЙ УРОВЕНЬ
На уровне макромолекул
степень сложности систем, по сравнению с обычными молекулами, растет. Однако этот уровень еще не достаточен для возникновения полноценной жизни.
Макромолекулами принято называть очень крупные, обычно полимерные (многозвенные) молекулы. В живых организмах различают четыре типа макромолекул: углеводы, липиды, белки и нуклеиновые кислоты (рис. 2). Они образуют химическую основу клеток, хотя некоторые углеводы и белки входят также в состав межклеточного вещества, обычно вместе с солями (основное вещество хряща, кости).
Рис. 2
. Структура основных макромолекул.
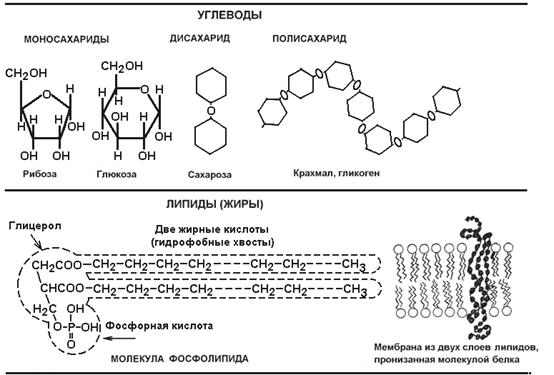
Углеводы
бывают простые - моносахариды (такие как глюкоза, лактоза) и сложные - полисахариды, образованные сотнями и тысячами соединенных моносахаридов. Некоторые полисахариды выполняют опорную функцию - целлюлоза (клетчатка) у растений, хитин у раков, насекомых, грибов. Но в основном углеводы используются как топливо для получения энергии (см. Тему 2).
Липиды,
или жироподобные вещества, имеют длинные «хвосты» из углеродно-водородных единиц, прикрепленные к «головке» - видоизмененной молекуле глицерина. Хвосты отталкивают воду (гидрофобны), поэтому два слоя липидных молекул, обращенные друг к другу хвостами, образуют водо- и иононепроницаемую пленку - мембрану. Из мембран построены оболочки клеток и некоторых внутриклеточных органоидов. Кроме того, липиды, как и углеводы, заключают в себе много энергии и используются в качестве топлива.
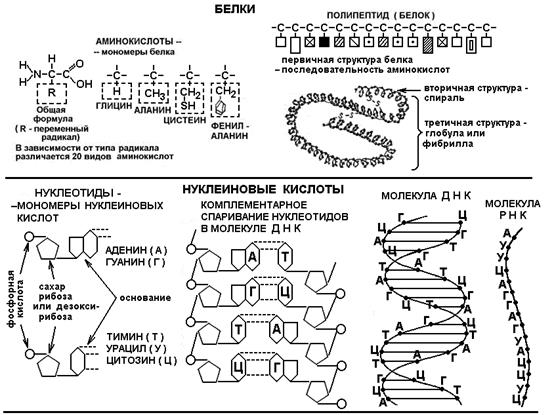
Белки
- основные биополимеры, так как выполняют большинство жизненных функций (см. Тему 2). Белковая цепь - полипептид - сложена из большого числа (50-100-500 и более) мономеров - аминокислот (включают аминогруппу -NH2
и кислотную группу -COOH). Имеется 20 разновидностей аминокислот, и чередование их беспорядочно (но строго определенно для каждого вида белка), так что возможное разнообразие белковых цепей бесконечно велико, что и дает возможность белкам выполнять очень разные функции. Наибольшим разнообразием отличаются белки-ферменты
- катализаторы биохимических реакций.
Нуклеиновые кислоты
(от латинского nuсleus - ядро) впервые были выделены из клеточных ядер и представляют самые сложные макромолекулы. Различают дезоксирибонуклеиновую кислоту - ДНК и рибонуклеиновую кислоту - РНК. ДНК - двухцепочечный полимер, РНК - одноцепочечный. Мономерами в обоих случаях являются довольно крупные и сложные молекулы - нуклеотиды. ДНК хранит информацию о структуре всех клеточных белков, РНК способствует ее реализации в момент синтеза новых белков (подробнее об этом см. Тему 3). Фрагмент ДНК, кодирующий структуру одной молекулы белка, называется геном
.
Макромолекулы обычно объединяются в макромолекулярные комплексы
, или даже в особые структуры, называемые органоидами
клетки (по аналогии с органами сложного организма). Типичными органоидами являются рибосомы - элементарные структуры, ведущие синтез белка, миофибриллы - сократимые нити в мышечных клетках, митохондрии - производители клеточной энергии, хромосомы - хранители ДНК, то есть генов.
Макромолекулы и их комплексы, гены, клеточные органоиды отвечают за отдельные свойства жизни - наследственность, синтезы, движение, энергетический обмен и др., но и эти свойства могут проявляться только в системе целостной клетки. Даже вирусы
, которые считаются внеклеточными формами жизни, вне клетки представляют фактически макромолекулярные кристаллы, не способные размножаться, синтезировать белки, усваивать энергию. Поэтому некоторые ученые вообще не считают вирусы живыми образованиями.
Таким образом, отдельные молекулярно-генетические структуры не обеспечивают того критического уровня сложности, который можно было бы назвать полноценной жизнью.
СЕГМЕНТ 8. ОНТОГЕНЕТИЧЕСКИЙ УРОВЕНЬ
Онтогенез
- это индивидуальное развитие организма, начиная от одной клетки (зиготы, образующейся при слиянии яйцеклетки и сперматозоида) до взрослого многоклеточного существа со множеством специализированных тканей и органов. Необходимость объединения этих подуровней в один онтогенетический уровень вызвана двумя причинами. Во-первых, зигота - по сути обычная клетка - уже представляет организм, хотя и на одноклеточной стадии развития. Во-вторых, в природе существуют не только многоклеточные, но и одноклеточные организмы как животного, так и растительного свойства - амеба, инфузория, эвглена, хлорелла и др. Бактерии - особо мелкие и безъядерные (прокариотные) клетки - тоже самостоятельные организмы, хотя живут обычно колониями. Так что понятия «клетка» и «организм» в определенных случаях совпадают.
Из сказанного следует очень важный вывод: клетка
является наименьшей, то есть элементарной живой системой
, так как ей присущи все свойства живого организма, свойства жизни как явления
. Клетка, как и многоклеточный организм способна питаться, поглощать энергию, синтезировать вещества, двигаться, реагировать на раздражители, размножаться, приспосабливаться и д.т
. Этому способствует достаточно высокая степень структурной дискретности - внутреннее расчленение клетки на органоиды, изолированные отсеки - особенно выраженная у высших, эукариотных клеток (рис. 3).
Рис. 3
. Схема организации про- и эукариотной клеток.
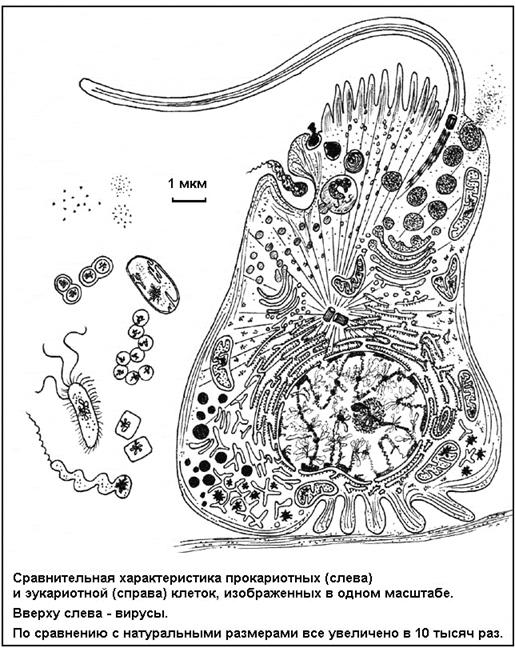
Существует нерешенная проблема клеточного уровня (подуровня), связанная с наличием в природе двух типов клеточной организации - прокариот и эукариот. Прокариоты
(доядерные)
- это мелкие (около 1 мкм) клетки, не имеющие ядра и других органоидов, типичных для эукариот. Наследственное вещество - ДНК - лежит свободно в цитоплазме, а прочие функциональные блоки тоже представлены небольшими макромолекулярными комплексами без оболочек. К прокариотам относятся все бактерии и так называемые сине-зеленые водоросли. Эукариоты
(с настоящим ядром)
- крупные (10-50 и более мкм) клетки, в которых ДНК в форме хромосом заключена в ядре и большинство рабочих структур, ферментов организовано в изолированных органоидах. Изолирующую роль для ядра и органоидов выполняют такие же липидно-белковые мембраны, как и мембрана клеточной поверхности. Эукариотную организацию имеют одноклеточные простейшие (амеба, инфузория и другие) и клетки многоклеточных организмов: грибов, растений, животных, включая человека. Суть проблемы не в размерных и даже не в структурных различиях двух типов клеток, а в том, что некоторые органоиды эукариотных клеток, такие как митохондрии и хлоропласты, похожи на прокариот - бактерий и сине-зеленых водорослей. Они имеют собственную ДНК, аппарат синтеза белка (рибосомы), систему энергообеспечения и, таким образом, мало зависят от других структур клетки, в частности от ядерной ДНК. На этом основании разработана симбиотическая гипотеза
о происхождении эукариотной клетки на основе симбиоза (взаимовыгодного объединения) некогда самостоятельных прокариотных клеток. В таком случае про- и эукариотные клетки не только по уровню сложности, но и по происхождению должны представлять разные - низший и высший - подуровни клеточного уровня организации. Этот пример показывает, что приведенная и общепринятая система уровней организации жизни не отражает всей сложности отношений между уровнями и подуровнями. Да и число подуровней можно увеличить, поскольку иерархическая сложность систем на самом деле значительно богаче.
Ткани
и органы
представляют основные промежуточные подуровни между клеткой и организмом
. Естественно, что эти подуровни можно выделить только у многоклеточных животных, растений, грибов.
Например, у человека различают эпителиальную (покровную) ткань, мышечную, нервную и соединительную (рыхлую, плотную, хрящевую, костную, кровь и лимфу). Ткани состоят из клеток и межклеточного связующего вещества. Органы состоят из разных тканей. Так, сердце кроме основной мышечной ткани включает рыхлую соединительную, кровь, нервные элементы и эпителиальные оболочки. Головной мозг наряду с нервными клетками содержит питающие их кровеносные сосуды, желудочки, выстланные специальным эпителием. Многие органы объединены в системы органов (пищеварительную, кровеносную и др.).
Наконец, многоклеточный организм
, как и отдельная клетка, представляет законченный и устойчивый уровень биологической организации
. Организм, или особь, способен к самостоятельному существованию, размножению и развитию
.
СЕГМЕНТ 9. ПОПУЛЯЦИОННО-ВИДОВОЙ УРОВЕНЬ
Вид
- важнейшая биологическая категория, которая определяется как совокупность особей (организмов), обладающих наследственным сходством по морфологическим, физиологическим, генетическим, эколого-географическим признакам, способных свободно скрещиваться и давать плодовитое потомство. Со времен Карла Линнея (выдающийся шведский натуралист 18 века) биологические виды обозначаются двойным наименованием на латинском языке - первое слово обозначает род, второе - вид. Например, Phaseolus vulgaris - фасоль обыкновенная, Passer domesticus - воробей домовый, Homo sapiens - человек разумный.
Главное в определении вида (его главный критерий) - способность особей скрещиваться и, более того, оставлять плодовитое потомство. В диких условиях особи разных видов не скрещиваются. Искусственно можно скрестить лошадь и осла, но их потомство - мул - бесплодно. Так что лошадь и осел - разные виды.
Каждый вид занимает на Земле определенный ареал
- территорию или акваторию (эколого-географический критерий вида). Иногда это - небольшой, изолированный участок, например, Манчжурская тайга для амурского тигра. Такие виды называют эндемичными, или эндемиками. В других случаях вид распространен по всему земному шару - виды-космополиты. Чаще ареал вида бывает разорван, вид существует отдельными группировками - популяциями.
Популяция
- некоторая изолированная совокупность особей одного вида, длительное время населяющая определенный ареал и способная к свободному скрещиванию. Кроме ареала популяция имеет и определенную экологическую нишу. Если ареал - это адрес популяции, то экологическая ниша - ее образ жизни: состав пищи, враги, водный режим, ярус леса и т.п. Но главное качество популяции как единицы воспроизведения и эволюции биологических видов - доступность ее особей к свободному скрещиванию, то есть свободная комбинаторика родительских генов. Постепенное расхождение генетической структуры популяций рождает новые виды. Поэтому иногда трудно провести грань между популяцией и видом, поэтому эти категории и рассматриваются в рамках одного уровня организации (подробнее см. тему 5).
СЕГМЕНТ 10. БИОГЕОЦЕНОТИЧЕСКИЙ УРОВЕНЬ
На этом уровне рассматриваются экологические системы: сообщество, биогеоценоз, биосфера.
Сообщество -
совокупность популяций разных видов на определенной территории
. Обычно специалисты (ботаники, зоологи, микробиологи) выделяют в сообщества объекты определенной категории: растительное сообщество - фитоценоз, сообщество животных - зооценоз, микроорганизмов - микробоценоз. Тогда совокупность всех совместно обитающих сообществ разных видов, представленных на ареале отдельными популяциями, образует высшее сообщество
- биоценоз
. Популяции разных видов в сообществе или биоценозе тесно взаимодействуют на основе разделения пищи и ярусов, взаимного использования продуктов обмена, отношений хищник-жертва, паразит-хозяин и т. д.
Любое живое сообщество, весь биоценоз способны существовать в определенных условиях внешней среды
. Для наземных сообществ это - почва определенного типа, температура, влажность, освещенность; для водных - минеральный состав, соленость и аэрация воды, те же температура и освещенность, глубина, течения и др. Совокупность этих неживых (абиотических) факторов среды обитания сообществ обозначается как биотоп
(дословно - место жизни)
.
Важнейшее обобщение современной экологии состоит в том, что
неживая среда и населяющий ее биоценоз обмениваются веществом и энергией, находятся в тесном взаимодействии, поэтому биотоп и биоценоз складываются в единую систему
- биогеоценоз
. Биогеоценозы - это естественные (природные) экосистемы: лесные, степные, болотные, озерные, речные, морские и др. Но человек создает и искусственные экосистемы - в частности, агроценозы (сельскохозяйственные плантации, птицефабрики, животноводческие фермы и т.п.), аквариумы и рыборазводные пруды, очистные сооружения со специально подобранными сообществами микробов, водорослей, моллюсков-фильтраторов, наконец, космические станции с уникальным внутренним климатом и биологическим равновесием.
Высшим экосистемным объединением на Земле является
биосфера -
земная оболочка, населенная живыми существами
. Основоположником учения о биосфере Земли является выдающийся российский натуралист и философ Владимир Иванович Вернадский
(1863-1945)
. Основная мысль этого учения и созданной Вернадским науки биогеохимии состоит в том, что живой и неживой мир нашей планеты един, организмы и компоненты среды связаны обменом (круговоротом) веществ и энергии
. Вершиной творческого наследия Вернадского является его представление о ноосфере
- биосфере, обогащенной разумом человека
. Разумная деятельность людей активно преобразует состав биосферы и становится все более важным фактором ее необратимой эволюции. Только к концу 20 века человечество начало понимать эту простую истину и задумалось над тем, как сохранить существующее равновесие.
Существуют ли живые системы более высоких уровней организации, чем биосфера Земли? Другими словами - существует ли жизнь вне Земли, в каких-нибудь дальних или ближних космических системах? И совсем тривиально - есть ли жизнь на Марсе? Наука пока не знает ответа на эти вопросы. Ученые предполагают, что по крайней мере на Марсе - ближайшей к нам планете - есть условия если не для жизни, то для переживания простых организмов типа бактерий в состоянии спор. При похожих условиях в ледяных толщах Антарктиды обнаружены микроорганизмы. Но Антарктида когда-то была ближе к экватору Земли, в составе единого материка Гондваны, и жизнь сохранилась здесь от давних времен. Существует ли жизнь на Марсе - должны показать ближайшие исследования этой планеты, в частности, планируемая на начало нового века экспедиция американских астронавтов.
ЗАКЛЮЧЕНИЕ ПО ТЕМЕ 1
Все объекты природы являются системами. Живые системы имеют разную степень сложности - от молекул до биосферы - и представляют в совокупности многоступенчатую иерархию уровней организации. Каждый уровень организации жизни имеет свои специфические свойства, закономерности структуры, функции, развития, приобретает новые качественные характеристики. Принципиальный качественный скачок наблюдается при переходе от макромолекулярных комплексов к клеткам - появляется качество жизни как свойство определенного уровня сложности материи
.
Наиболее устойчивыми живыми системами являются клетка, организм, биогеоценоз.
Что же это за новое качество - жизнь? В чем ее сущность, отличие от «нежизни»? Этот принципиальный общенаучный вопрос мы рассмотрим в следующей теме.
ТЕМА 2. КОНЦЕПЦИЯ МАТЕРИАЛЬНОЙ СУЩНОСТИ ЖИЗНИ
Сегмент 11. Механицизм и витализм в истории биологии.
Сегмент 12. Живая материя и ее основная форма движения. Обмен веществ и энергии в живой системе.
Сегмент 13. Трансформация и использование энергии.
Сегмент 14. Белки - структурно-функциональная основа жизни.
Сегмент 15. Опора и движение.
Сегмент 16. Транспорт веществ.
Сегмент 17. Ферментативный катализ.
Сегмент 18. Защитные реакции. Иммунитет.
Сегмент 19. Сигнализация. Гормональная и нервная регуляция.
Заключение по теме 2.
Концепция сущности жизни имеет прямое отношение к основному вопросу философии, суть которого в соотношении бытия и сознания, материи и духа. Диалектический материализм решает этот вопрос так: бытие, материя - первичны; сознание, дух - вторичны. То есть сознание как высшее отличие живого от неживого является свойством материи. В таком понимании жизнь есть форма существования особо сложной материи.
Современное понимание материи позволяет полнее охарактеризовать сущность жизни, что мы и сделаем в сегменте 12. В последующих разделах мы объясним с материальных позиций основные функции жизни - использование энергии, движение, транспорт веществ, ферментативный катализ, иммунитет, механизмы сигнализации и нервной деятельности, размножение и другие. Однако прежде полезно дать историческую справку о развитии взглядов на жизнь в среде философов и ученых.
СЕГМЕНТ 11. МЕХАНИЦИЗМ И ВИТАЛИЗМ В ИСТОРИИ БИОЛОГИИ
Исторически существовало две противоположные точки зрения
на этот вопрос - материалистическая и идеалистическая
. Первая получила название механицизма, вторая - витализма.
Механицизм
(от греческого mechane - орудие, сооружение) объяснял жизнь исходя из обычных механических или физических форм движения и превращения материи. Механицизм - односторонний метод познания, так как основан на признании механической формы движения материи единственно объективной. Было несколько механистических трактовок сущности жизни.
Собственно механицизм
- жизнь объяснялась на основе принципов классической ньютоновской механики. Ее каждый прошлый и будущий шаг может быть просчитан. Рождение, жизнь и смерть также циклично закономерны, как восход и заход солнца. Эта трактовка имеет сейчас лишь исторический интерес.
Машинная теория
была популярна в 17-18 веках (Декарт и др.). Жизнь представлялась как сумма физических и химических процессов, которые подобно машинным процессам протекают на статичных, неизменных структурах. В этой трактовке отсутствовала идея развития, эволюции. Живой мир рассматривался как сложный физико-химический механизм, работающий в заданном режиме.
Механицизм как физикализм
возник в конце 19 - начале 20 веков и получил законченные формы к середине 20 века. Согласно представлениям физикалистов жизнь развивается, но по сути она представляет простые физико-химические процессы
. Причем сложнейшие биологические процессы сначала сводятся к более простым химическим, а химические в свою очередь сводятся к еще более простым - физическим. Такие отношения между уровнями разной сложности обозначаются как принцип сводимости
(сложные уровни и процессы сводятся
к простым). Метод познания, основанный на принципе сводимости, или редукции, называется редукционизмом
(от латинского reductio - отодвигание назад, возвращение к прежнему состоянию), так что физикализм в своем методологическом применении выступает как крайняя форма редукционизма
. Для физикализма, как и для механицизма в целом, характерно отрицание качественной специфики более сложных материальных образований, поскольку сложное сводится к более простым элементам, целое - к сумме его частей.
Развитию физикализма как естественнонаучного мировоззрения способствовали успехи физики и химии второй половины 19 века и первой половины 20 века, открытие в живых телах свойств и законов движения (в физико-химическом смысле) неорганических тел. Жизнь сводится к процессам обмена веществ и энергии по химическим и физическим законам. Приведем некоторые характерные высказывания видных ученых в духе физикализма.
Бертран Рассел - английский философ, математик, логик; написано в 1951 году: «Нет причины считать, что живая материя подчиняется иным законам, чем те, которые управляют живой материей, и есть достаточные основания полагать, что все поведение живой материи удастся теоретически объяснить средствами физики и химии». Таким образом, надо полагать, биология станет частью физики и химии.
Эрвин Шредингер - австрийский физик, квантовый механик заявил в 1946 году, что «живая материя, хотя она и не отклоняется от установленных к настоящему времени физических законов, вероятно подчиняется и другим, еще не открытым физическим законам, которые, когда они будут ясно показаны, составят такую же неотъемлемую часть физики, как и первые».
То есть любые, пока что неразгаданные формы проявления жизни рано или поздно будут объяснены как физические процессы. Бурное развитие физики второй половины 20 века, открытие новых элементарных частиц и физических полей, успехи кибернетики и теории информации все более полно объясняют сложные материальные взаимодействия в природе, в том числе и в живых организмах, и все меньше тайн остается в понимании сложных биологических процессов. Но сама по себе физико-химическая интерпретация жизненных реакций не давала в прошлом и не дает сейчас ответа но вопрос: где кончается неживая природа и начинается живая? А что предлагали по этому поводу идейные противники механицистов - виталисты?
Витализм
(от латинского vitalis - жизненный, живой) утверждает, что живое не сводится только к физико-химическим явлениям, в нем действуют еще и особые «жизненные силы».
Витализм - давняя концепция, его корни, как и корни механицизма, уходят в классическую древность. Великий античный философ Аристотель (IV век до н. э.) ввел понятие «энтелехия», которое противопоставляется «материи» и означает конечную причину, цель, идею о совершенстве формы организма, которая и управляет развитием. По определению Аристотеля живой природе присуща «цель в самой себе».
В начале 18 века немецкий врач и химик Шталь - автор известной в химии теории флогистона, опровергнутой позже Лавуазье - развивал в медицине виталистическую теорию, известную под названием анимизма (от латинского anima -душа, дух). По Шталю главное для живого организма - его душа, она управляет телом и не допускает его распада.
В 19 веке состоялись выдающиеся открытия химии и физики, виталисты быстро теряли своих сторонников. Виталисты утверждали, что органические вещества могут возникать только с помощью «жизненной силы», но уже в 1828 г. Вёлер из неорганических веществ синтезировал мочевину - азотсодержащее органическое вещество животного происхождения. Знаменитый французский микробиолог Луи Пастер считал, что разложение сахара (брожение, дыхание) - особое свойство живых клеток, но в 1897 г. Бухнер получил из дрожжей ферментный экстракт и провел брожение сахаров в бесклеточной системе, то есть без всякой «жизненной силы». Сильный удар по витализму нанесло открытие Рубнера: в начале 20 века он установил, что закон сохранения энергии действует и в органическом, живом мире.
Однако идея энтелехии не была преодолена окончательно. В начале 20 века система витализма наиболее полно была изложена Хансом Дришем - видным немецким биологом и философом. Опираясь на открытые им эмбриональные регуляции, Дриш утверждал, что развитие организма не сводится к реализации предустановленного, заранее спланированного экстенсивного (пространственного) разнообразия, как утверждали механицисты, но происходит переход интенсивного (непространственного) разнообразия в экстенсивное. Этот переход свойствен только живым системам и осуществляется под действием специфически витального фактора - энтелехии.
Заметим, что признание энтелехии, жизненной силы часто ведет к антропоморфическим образам: учение о субстанциональной душе, психической силе. На этих понятиях основываются так называемый психовитализм (психизм), мистицизм. Поддержанию таких понятий способствуют очень большие и пока не разрешенные трудности в понимании принципов работы мозга, векторов эмбрионального развития, направленного и «целесообразного» характера биологической эволюции. Положительное значение витализма состояло в критике механистических взглядов на биологическую причинность, в стимулировании работ по биологической информации.
С развитием системного подхода и современного учения о самоорганизации (синергетики) причины специфической живой
организации стали искать не во внешних силах, а в самопроизвольно и эмерджентно (см. сегмент 5) возникающих новых свойствах достаточно сложных
систем. Специфика живого не отрицается, но она выводится как естественное свойство наиболее сложно организованной материи. Некий пороговый уровень сложности органических макромолекул - прежде всего белков и нуклеиновых кислот - и является той гранью, за которой (той «причиной», по которой) возникает качество жизни.
В общефилософском смысле можно говорить о переходе количественных изменений в качественные. Понять эту качественную специфику - наша дальнейшая задача. Однако одного философского рационализма
(от латинского ratio - разум), мудрствования здесь мало - нужны достоверные эмпирические
(опытные) знания. Их добывают естественные науки, в том числе биология.
СЕГМЕНТ 12 ЖИВАЯ МАТЕРИЯ И ЕЕ ОСНОВНАЯ ФОРМА ДВИЖЕНИЯ. ОБМЕН ВЕЩЕСТВ И ЭНЕРГИИ В ЖИВОЙ СИСТЕМЕ
Живая материя имеет в основе те же физические свойства, что и неживая. Понятие «материя» многогранно. В целом материя
представляет совокупность вещества и поля
, которые выступают как разные виды материи
. Веществом называют объекты и системы, обладающие массой покоя. Поле - это виды материи, не имеющие массы покоя. Например, электромагнитное поле представляет излучение в форме квантов (порций) энергии. Существует также гравитационное поле, нейтринное излучение. Между веществом и полем нет строгой границы, так как элементарные частицы вещества, например электроны, обладают одновременно корпускулярными и волновыми свойствами (дуализм волны и частицы). Эти базовые положения квантовой (волновой) механики были сформулированы в 20-30-е годы 20-го века. Выдающимся представителем этой новой науки был датский физик Нильс Бор.
Живая материя представляет особо сложное вещество и, соответственно, сложное многофакторное поле. Именно уровень сложности
делает материю живой, хотя внутри нее действуют простые физические и химические законы
. По уровню сложности материи разграничиваются и сферы внимания естественных наук. Атомы - поле деятельности физики, молекулы - объект химии, с уровня макромолекул начинается биология, так как с этого уровня сложности появляются качественно новые свойства, характеризующие живую материю.
Биологические макромолекулы
- это белки, липиды, углеводы и нуклеиновые кислоты. Их краткая характеристика уже дана в сегменте 7 и на рис. 2. Подчеркнем еще раз, что белки и нуклеиновые кислоты представляют апериодические полимеры, так как их мономеры - 20 видов аминокислот в белках и 4 вида нуклеотидов в ДНК и РНК - чередуются беспорядочно. Это и является источником огромного структурного разнообразия живой материи, какого нет в неживой природе.
Любая материя существует в
движении
. В широком смысле под движением материи понимают ее постоянное развитие, изменение, преобразование вещества в поле и обратно
. Для понимания основной формы движения живой материи сначала необходимо усвоить важнейшее исходное понятие - 2-ой закон термодинамики
. Суть его в том, что в природе изначально существует фундаментальная асимметрия, неравновесие
в распределении вещества и поля (энергии), поэтому самопроизвольно все физические процессы (движение материи) направлены к достижению равновесного
состояния
. Это означает переход материи из упорядоченного, структурированного состояния, когда есть сгустки и разреженные участки вещества и поля, к диффузному, гомогенному распределению вещества и поля в пространстве. В таком диффузном состоянии материя имеет минимальную свободную энергию
- энергию, способную совершить работу, и, напротив, максимальную энтропию
-рассеянную долю энергии, не способную к совершению работы. Это правило касается всех самопроизвольных процессов, в том числе колебательных: горячее тело рано или поздно остывает (энергия рассеивается); прыгающий мяч снижает амплитуду и в итоге останавливается и т. д. Эти процессы дезорганизации материи самопроизвольно необратимы.
Сформулированное 2-ое правило термодинамики полностью справедливо и для живой материи, которая в основе подчиняется законам физики и самопроизвольно стремится к распаду, к равновесному состоянию с минимальной свободной энергией и максимальной энтропией.
На рис. 4 эти процессы показаны в левой части схемы и означают смерть
живой материи
, ее превращение в неживую.
Рис. 4
. Термодинамические процессы в живой материи.

Однако живая материя, пока она действительно живая, остается неравновесной, структурированной, высоко упорядоченной
. В ней имеется свободная, готовая совершить работу энергия, а энтропия минимальна. Такое состояние поддерживается
за счет притока внешней энергии
и ее трансформации в энергию химических связей макромолекул.
Концентрация вещества и поля, то есть повышение внутренней свободной энергии материи происходит в процессе разнообразных биосинтезов
(образования сложных веществ из простых), сопряженных с поглощением внешней энергии.
Это и есть жизнь -
противоположность смерти
(правая часть схемы на рис. 4). Поскольку основной формой внешней энергии для поддержания жизни является солнечный свет, формулу жизни можно конкретизировать:

Таким образом, взаимодействие потоков простого вещества и энергии в ходе биосинтеза живого вещества и обратный процесс распада, то есть обмен веществ и энергии
, составляет фундаментальное свойство жизни, основную форму движения живой материи.
Этому определению вторят разнообразные характеристики жизни, как-то:
Жизнь
- это специфическая форма движения материи (кругооборот материи, обмен веществ и энергии) с поддержанием упорядоченного неравновесного состояния (с высокой свободной энергией и низкой энтропией) за счет поглощения и трансформации внешней энергии.
Жизнь
- это поддержание высокой упорядоченности (низкой энтропии) в среде с меньшей упорядоченностью (высокой энтропией).
Жизнь
препятствует росту энтропии.
Жизнь
- это синтез вещества и поля.
Далее мы подробнее остановимся на вопросах биосинтеза веществ и использования энергии, но не вдаваясь в глубокие детали во избежание потери главной мысли - о материальной сущности жизни.
СЕГМЕНТ 13. ТРАНСФОРМАЦИЯ И ИСПОЛЬЗОВАНИЕ ЭНЕРГИИ
Итак, материальная сущность жизни проявляется, прежде всего, в непрерывном обмене веществ и энергии, который происходит между живой системой (клеткой, организмом, биоценозом) и окружающей его внешней средой. В этом смысле биологические системы являются открытыми
.
Разные организмы потребляют разные виды энергии
, в связи с чем их делят на аутотрофные и гетеротрофные. Аутотрофные организмы
(дословно - самопитающиеся) способны поглощать энергию неживой природы.
Прежде всего это зеленые растения, а также бурые, красные и сине-зеленые водоросли, использующие солнечный свет для процесса фотосинтеза
- образования органического вещества глюкозы из неорганических воды и углекислого газа. К аутотрофам относятся также некоторые бактерии, способные к реакциям хемосинтеза
- синтеза органических веществ за счет энергии простых химических реакций. При этом первичная энергия (солнечная или химическая) преобразуется в энергию химических связей сложных органических молекул
, так что аутотрофы как бы сами создают себе пищу. Гетеротрофные организмы
(питающиеся за счет других) - человек, все животные, грибы, а также многие бактерии - получают пищу в виде готовых органических веществ, произведенных аутотрофами, в основном растениями.
В составе этой пищи они получают и энергию, заключенную в химических связях. Если органическое вещество пищи расщепить на более простые вещества, освобождается энергия. По сути гетеротрофы получают ту же солнечную энергию, но преобразованную зелеными растениями в химическую. Отсюда ясна огромная роль растительных организмов как посредника в энергетическом обеспечении животных и человека
. Избавиться от этой зависимости, получать какую-либо энергию прямо из неживой природы человечество еще не научилось. И хотя академик В. И. Вернадский выдвигал такую научную задачу, дальше фантастических произведений дело не продвинулось и вряд ли продвинется в обозримом будущем. Поэтому для биологов всего мира одной из приоритетных задач остается понять во всех деталях механизм фотосинтеза, с тем чтобы максимально интенсифицировать его в растениях и по возможности воспроизвести в искусственных условиях.
Рис. 5

Рассмотрим несколько подробнее реакции энергетического обмена
.
Независимо от исходного источника энергии все организмы - как аутотрофы, так и гетеротрофы - сначала переводят энергию в удобное для дальнейшего использования состояние. Это - так называемые макроэргические (богатые энергией) связи в молекулах аденозинтрифосфорной кислоты - АТФ
(рис. 5). Образуются молекулы АТФ из аденозинди
фосфорной (АДФ) или аденозинмоно
фосфорной (АМФ) кислоты и свободных молекул фосфорной кислоты, но при непременном поглощении внешней энергии - солнечной или химической (эндотермическая реакция). Количество энергии, запасенное в макроэргической связи, на порядок больше, чем в обычных связях, например, внутри молекулы глюкозы, поэтому в составе АТФ энергию удобно хранить и транспортировать в пределах клетки. В местах потребления этой энергии АТФ распадается на АДФ и фосфат (при крайней необходимости даже на АМФ и два фосфата), а освобожденная энергия расходуется на ту или иную работу - синтез глюкозы в хлоропластах растительных клеток, синтез белков и других макромолекул, транспорт веществ в клетку и из клетки, движение и др. (см. рис. 5 и 6). АДФ (АМФ) и фосфат могут снова соединиться, захватив очередную порцию внешней энергии, а потом разрушиться и отдать энергию в работу. Циклические преобразования АТФ многократно повторяются. Таким образом, АТФ выступает в качестве универсального переносчика энергии внутри клетки, своеобразной разменной монетой в энергетических платежах за внутриклеточные процессы
.
Рис. 6
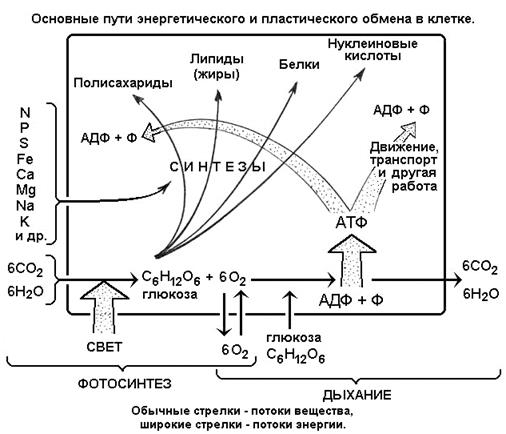
После того, как нам стала ясна роль АТФ и ее цикл, вся проблема клеточной энергетики сводится к пониманию первичных источников энергии
и механизмов ее перевода в АТФ
. В общем виде ситуация такова: у фотосинтетических аутотрофных организмов синтез АТФ из АДФ и фосфата генерируется солнечной энергией, у гетеротрофов - энергией от окисления пищевых продуктов (см. рис. 5). Таким образом,
растениям для синтеза АТФ нужен свет
, животным и человеку нужна органическая пища
.
Свет
является первичным
источником энергии
, он используется в реакциях фотосинтеза
у растений
. По конечной сути реакция фотосинтеза довольно проста:
6СО2
+ 6H2
O + энергия света = С6
Н12
О6
+ 6О2
(рис. 6): с помощью энергии света из углекислого газа и воды синтезируется 6-углеродное органическое вещество - глюкоза (моносахарид), и в качестве «лишнего» продукта образуется кислород, который уходит в атмосферу. На самом деле реакция более сложная, она состоит из двух стадий: световой и темновой. Сначала на свету с помощью особого Mg-содержащего белка хлорофилла вода расщепляется на кислород и водород, а энергия водорода передается на синтез АТФ. Только потом, в темновой стадии, водород соединяется с углекислым газом и образуется глюкоза. При этом часть АТФ расщепляется, отдавая энергию глюкозе.
Глюкоза вместе с минеральными веществами, поступающими в растение из почвы (азот, сера, фосфор, железо, магний, кальций, калий, натрий и др.), становится основой для более сложных синтезов - образуются полисахариды, липиды, белки, нуклеиновые кислоты, из которых строятся рабочие структуры клеток. Но и эти синтезы, как и синтез глюкозы, требуют энергетических затрат. Прямое использование света здесь невозможно (эволюция не создала таких энергетических переходов), поэтому некоторая часть глюкозы тратится как энергетический субстрат, то есть глюкоза
становится вторичным
источником энергии
. Глюкоза расщепляется и отдает энергию - сначала на синтез АТФ, а после расщепления АТФ - на биосинтезы макромолекул (рис. 6). Значительная часть АТФ, как уже сказано выше, расходуется на другую работу - транспорт веществ, движение клетки и др.
Наиболее эффективно глюкоза расщепляется с участием кислорода: C6
H12
O6
+ 6O2
= 6CO2
+ 6H2
O + энергия. По химической сути это - полное окисление (горение!) глюкозы. В живой клетке это «горение» происходит замедленно, поэтапно, так что энергия выделяется малыми порциями, и большая ее часть (около 55 %) используется на синтез АТФ, остальная рассеивается в виде тепла. Полное окисление одной молекулы глюкозы обеспечивает синтез 38 молекул АТФ. Поскольку кислород для окисления мы вдыхаем с атмосферным воздухом, то и на химическом уровне окисление глюкозы кислородом называют дыханием
.
На рис. 6 все описанные процессы схематично показаны для аутотрофной
растительной клетки
. Ее главная черта - способность к фотосинтезу, который обеспечивает первый этап построения органического вещества, в форме глюкозы. Но и дыхание в полной мере присуще растениям, так как именно этот процесс извлекает энергию из глюкозы (а также из жиров и лишних белков), переводит ее временно в АТФ и далее в сложные макромолекулы. Эта же схема, но с изъятием реакции фотосинтеза, соответствует и гетеротрофному
метаболизму животных клеток
. В этом случае глюкоза (а также другие углеводы, жиры, трофические белки и др.) поступают в клетку извне в готовом виде. Часть этих материалов идет на дыхание (в топку, для извлечения энергии через синтез АТФ), а часть - после некоторой переделки - на синтез новых макромолекул как строительный материал . Таким образом, пища у гетеротрофов (то есть и у нас с вами) имеет двойное назначение - энергетическое и пластическое (строительное)
.
На рис. 6 следует обратить внимание на единство процессов энергетического и пластического (строительного) метаболизма клетки
.
Энергия поглощается из внешней среды, преобразуется в АТФ прежде всего для осуществления строительных процессов, для построения живой материи. Или обратно: построение живой материи, то есть синтез макромолекул из простых неорганических веществ, возможен только с поглощением внешней энергии. Точки пересечения стрелок энергетического и пластического обмена означают места сопряжения энергетического и пластического метаболизма
. В живой клетке таких мест очень много. Сопряжение светового потока и синтеза глюкозы (реакция фотосинтеза) происходит в хлоропластах
- специальных органоидах растительных клеток, сопряжение дыхания (окисления глюкозы) и синтеза АТФ - в митохондриях
, сопряжение распада АТФ и синтеза клеточных белков - в рибосомах
и т. д. (см. сегменты 7 и 8 - о строении клетки, а также рис. 3).
С точки зрения обсуждаемой проблемы - материальной сущности жизни - все рассмотренные процессы обмена веществ и энергии в клетке представляют простые физико-химические реакции
. Мы не углублялись в механизмы преобразования энергии в хлоропластах и митохондриях, но если бы сделали это, то убедились бы в их изначально физической сущности. Основные события здесь связаны с распадом атомов водорода на элементарные заряженные частицы - протоны (Н+) и электроны (е-) и их индуцированным переносом по разные стороны мембраны внутри хлоропластов и митохондрий. В этих органоидах, как в электрических батарейках, создается разность электрических потенциалов порядка 0,2 вольта, которая при необходимости реализуется в электрический (протонный) ток, а этот ток, в свою очередь, вращает и активирует ферменты синтеза АТФ. Аналогичные физико-химические преобразования мы увидим и в других клеточных структурах.
СЕГМЕНТ 14. БЕЛКИ – СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ОСНОВА ЖИЗНИ
Как уже отмечено в сегменте 7, белки представляют полимерные молекулы длиной в 50-100-500 и более мономеров - аминокислот
. Имеется 20 разновидностей аминокислот, различных по химическим свойствам их свободных концов - радикалов.
Особо выделим аминокислоты со свободным электрическим зарядом. Так, аспарагиновая и глутаминовая кислоты имеют отрицательный заряд (лишняя –СОО- группа), аргинин, лизин, аспарагин и глутамин - положительный (лишняя –NH+ группа). Некоторые аминокислоты имеют свободную гидроксильную группу (–OH) . В совокупности активные группы придают молекуле белка электрический заряд - положительный или отрицательный в зависимости от соотношения аминокислот с разными зарядами. Кроме того, такие аминокислоты как цистеин и метионин имеют на свободном конце серу (группа –SH или –SCH3
) и могут формировать между собой -S-S- мостики, стягивая участок полипептидной цепи в петлю.
Эти и другие особенности белковой молекулы, которые целиком зависят от набора и порядка чередования аминокислот (то есть от ее первичной структуры
), придают каждой молекуле в водном окружении неповторимую вторичную и третичную (трехмерную) структуру
. В каждой белковой молекуле в строго определенных местах есть активные участки, нередко содержащие какой-нибудь металл (железо - Fe, магний - Mg, медь -Cu и др.) или другие специфические соединения. Эти участки обычно и отвечают за особые функции белков.
Важнейшее свойство белковой молекулы, объясняющее механизм ее функционирования, это -
способность обратимо изменять свою третичную структуру (трехмерную форму) в ответ на какое-либо раздражение
. Раздражителем чаще всего выступает энергетический разряд от расщепления молекулы АТФ.
Такая обратимая денатурация, или конформационная перестройка, и есть совершаемая молекулой работа
. Конформационная перестройка молекулы подобна циклу сжатия и расслабления пружины: при внешнем давлении пружина сжимается и приобретает внутренний запас энергии, а на обратном ходу совершает работу. В зависимости от структуры белковой молекулы и ее местонахождения совершаемая работа, то есть функция, будет различна. Ниже будет показана роль белков в реализации
основных жизненных функции: опорно-двигательной, транспортной, каталитической, защитной, сигнальной
.
Обычно называют еще энергетическую
функцию белков. Действительно, белки в своей химической структуре несут большой запас энергии. Они могут распадаться на отдельные аминокислоты, которые, в свою очередь, подобно глюкозе окисляются в митохондриях до углекислого газа и воды и отдают энергию на синтез АТФ. Однако этот путь в энергетическом обмене используется как резервный, в тех случаях, когда исчерпаны запасы углеводов и жиров - основных энергоемких субстратов. Использование белков в энергообмене расточительно для клетки, так как аминокислоты представляют дефицитный продукт, многие из них вообще не могут синтезироваться у животных, а получаются от растительных продуктов (незаменимые аминокислоты). Белки имеют уникальную структуру, которой дано более высокое предназначение. В этом смысле расщеплять белки для извлечения энергии - все равно, что топить печь ассигнациями. Кроме того, аминокислоты предварительно необходимо освободить от азота (дезаминировать), чтобы уровнять их с продуктами полураспада глюкозы, а это и дополнительная работа и потеря важнейшего элемента из организма (азот в составе простых соединений выводится с мочой). Таким образом, белки в энергетическом обмене используются в крайних случаях - при голодании, болезни, возрастных метаморфозах. Основные же функции белков, перечисленные выше и рассматриваемые ниже, значительно более важны и изящны, поскольку в их реализации используется уникальность структуры белка, его неповторимые индивидуальные формы и активные центры.
СЕГМЕНТ 15. ОПОРА И ДВИЖЕНИЕ
Опорно-двигательная функция
- одна из важных в обеспечении жизнедеятельности клеток и всего организма
. Движутся органоиды внутри клетки, сами клетки в воде или по твердой поверхности, организмы в их среде обитания. Движение невозможно без опоры, поэтому и в клетке и в организме формируются комплексные опорно-двигательные
структуры. Например, у человека мышцы связаны со скелетом, и только в такой комбинации работа мышц становится эффективной.
У позвоночных животных и человека опорные соединительные ткани
имеют в межклеточной основе специальный волокнистый белок коллаген (при вываривании костей, сухожилий или связок он набухает и дает клей, отсюда и название белка - клей дающий). В костях межклеточная коллагеновая основа для большей твердости минерализована солями кальция. Имеются опорные белковые микронити и микротрубочки и в самих клетках, они придают клеткам определенную форму, поддерживают отростки (например, в нервных волокнах).
В клетках мышечной ткани
имеются специальные сократительные нити - миофибриллы, которые обеспечивают сокращение клеток и всей мышцы. Каждая миофибрилла сложена из молекул белков двух основных видов - актина и миозина (рис. 7). Актин образует прочные микронити, по которым буквально шагают ножки миозина. Каждый шаг - это изменение третичной структуры молекулы миозина, его обратимая денатурация под действием энергии АТФ
. Когда одна ножка, зацепившись за микронить, изгибается, другая ножка распрямляется и цепляется за новое место на микронити. Потом они меняются ролями - первая ножка отцепляется от микронити и распрямляется, а вторая изгибается и подтягивает нить. Каждый шаг требует энергии АТФ. Таким образом, при наличии АТФ ножки быстро бегут вдоль микронити, а точнее - тянут нить к себе, так как обращенные в разные стороны пучки миозина скреплены хвостиками, и ножки разных пучков тянут свои микронити в противоположных направлениях (см. рис. 7).
Рис. 7
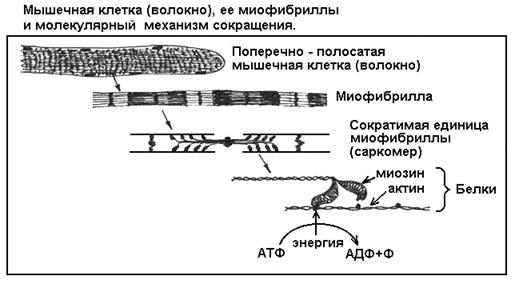
Главное, что следует вынести из описания мышечной функции, это понимание простой физико-химической сущности движения
, которая сводится к изменению объемной формы молекулы белка миозина
. Эта форма, или третичная структура, в свою очередь зависит от силы химических связей внутри молекулы. При энергетическом разряде от распада молекулы АТФ сила связей резко, но кратковременно изменяется - молекула «вздрагивает» (сгибается и тут же разгибается), совершая работу. Суть реакции миозина настолько проста, что воспроизводится in vitro («в стекле», в пробирке, то есть в искусственных условиях) на чистых белках, выделенных из клеток, или даже на мертвых клетках при добавлении АТФ. Добавим, что аналогичный механизм движения, но с другими белками, известен для ресничек и жгутиков, которыми снабжены многие одноклеточные организмы (например, инфузория), сперматозоиды, некоторые покровные (эпителиальные) ткани. Итак, движение - важное проявление жизни - имеет вполне материальную сущность
.
СЕГМЕНТ 16. ТРАНСПОРТ ВЕЩЕСТВ
Перенос веществ между организмом и средой - необходимый этап в процессах обмена веществ
. Это - поглощение пищевых материалов и вынос отходов, поглощение кислорода и удаление углекислого газа, перенос неорганических ионов, воды и других веществ. Способы и механизмы поглощения и выделения веществ различаются у организмов разного уровня сложности - у одноклеточных и многоклеточных, животных и растений
. На высших уровнях организации - у многоклеточных животных - в эти процессы включаются мышечные и ресничные механизмы
: преследование и захват добычи, глотание и перистальтика по кишечнику у позвоночных, нагнетание водных потоков и фильтрация планктона у сидячих беспозвоночных - например, у двустворчатых моллюсков мидии, гребешка. У высших растений для транспорта воды и солей от корней к стеблю и листьям большое значение имеет осмотическая диффузия
- движение растворов по градиенту концентрации, то есть из среды с высокой концентрацией вещества в среду с низкой концентрацией. Этот транспорт имеет простую физико-химическую основу и не требует энергетических затрат. Движение синтезированных веществ (сахаров) в обратную сторону, напротив, требует энергии.
Переносу малых молекул часто способствует их связывание с более крупными молекулами-переносчиками
. Типичный пример - перенос кислорода
из атмосферного воздуха, вдыхаемого через легкие, в кровь и далее во все клетки организма (для окисления пищевых субстратов и извлечения энергии - см. сегмент 13). В этом случае кислород из полости легких под осмотическим давлением поступает в кровеносные капилляры, далее в специальные клетки - эритроциты
(красные кровяные тельца), где химически связывается с особым белком гемоглобином
. Принцип связывания кислорода с гемоглобином очень прост. В составе этого белка имеются атомы железа
, которые и окисляются кислородом
- железо временно превращается в окисел (как обычное железо на воздухе превращается в ржавчину). Однако связь железа с кислородом непрочная, так что по мере углубления эритроцитов в ткани организма, где собственная концентрация кислорода ниже, гемоглобин отдает связанный с ним кислород, который теперь диффундирует в клетки, а далее в митохондрии. Решающее значение в транспорте кислорода имеет конфигурация (третичная структура) молекулы гемоглобина, от которой зависит окислительно-восстановительная способность связанного железа
. При заболевании, которое называется серповидно-клеточной анемией (эритроциты имеют нетипичную форму серпа), замена всего одной аминокислоты в составе гемоглобина так изменяет его форму, что делает железо не эффективным в отношении кислорода. Дыхание нарушается.
Критическим моментом в транспорте веществ между организмом и средой - будь то одно- или многоклеточный, животный или растительный организм - является преодоление клеточных оболочек
. Оболочка клетки включает липидно-белковую мембрану (плазмалемму) и углеводную стенку (см. сегмент 8 и рис. 3).
Клеточная стенка, даже очень толстая, как у растительных клеток, не представляет серьезного препятствия для воды и растворенных в ней веществ, но плазмалемма имеет настоящие барьерные свойства, так как ее основу составляют два слоя липидов (жиров), практически не проницаемых для водных растворов (см. рис. 2, 3 - строение липидов и мембран). Через билипидный слой легко проникают лишь газы (мелкие электрически нейтральные молекулы) и жирорастворимые вещества (спирты, ацетон и др.). Но клетке нужны прежде всего питательные органические и минеральные вещества. Для переноса заряженных частиц - ионов, а также небольших органических молекул - аминокислот, сахаров и т.п. в липидной мембране вставлены многочисленные белковые поры
. Мембрана представляет как бы белковую мозаику по липидному фону. Белки мембранных пор, как и большинство других белков, взаимодействуют с водой, так что пора представляет фактически водный канал. Однако благодаря белковой компоненте каждый такой канал имеет избирательную проницаемость
- для ионов K+, Na+, Ca2
+, Cl- и др. Вместе с ионами в клетку проникают и связанные с ними органические молекулы - сахара и аминокислоты, так осуществляется питание клеток. Рассмотренный случай представляет пассивный транспорт веществ
, он не требует затрат энергии. Совершенно особый тип каналов представляют ионные насосы, способные перекачивать ионы против градиента их концентрации. Как следует из самого термина, насосы могут работать только с затратой энергии, и такой способ перемещения веществ называют активным транспортом
. Белковая пора насоса способна расщеплять молекулу АТФ и за счет извлеченной энергии проталкивать ионы против их диффузионной силы. Созданный таким образом наружный избыток ионов возвращается обратно пассивным путем, но несет с собой нужные органические вещества. Так сочетание активного и пассивного транспорта обеспечивает питание клеток. Избыток ионов натрия снаружи клетки используется также для возбуждения электрического тока и проведения нервного возбуждения (см. сегмент 19).
Даже поверхностное рассмотрение механизмов мембранного транспорта веществ показывает, что,
как и в случае мышечного сокращения, работа по активному транспорту сводится к обратимой конформационной перестройке молекул белковых пор с преобразованием энергии АТФ
. Направленный в обратную сторону пассивный транспорт ионов и питательных веществ представляет простую диффузию, но является следствием предшествующего активного транспорта. Все эти процессы подчиняются в основе известным физико-химическим закономерностям.
СЕГМЕНТ 17. ФЕРМЕНТАТИВНЫЙ КАТАЛИЗ (БИОКАТАЛИЗ)
Катализ - это возбуждение или изменение скорости химических реакций с помощью внешних добавок - катализаторов
. Катализ используется в химической промышленности при получении аммиака, серной и азотной кислот, моторного топлива, разнообразных полимеров. Широко распространен катализ и в живых организмах. С участием катализаторов реализуется генетическая информация и осуществляются все процессы обмена веществ и энергии
. Катализ происходит в клетках, в межклеточных жидкостях и полостях, в пищеварительном тракте.
В роли катализаторов биохимических реакций выступают специальные белки, называемые ферментами
. При сравнительно низких концентрациях реагирующих веществ и при нормальных температурах (не выше 40 градусов) клеточные реакции ускоряются с помощью ферментов в миллионы и миллиарды раз! Различают анаболитические
, направленные на синтез, и катаболитические
, направленные на распад, ферментативные реакции
. Несколько разных ферментов могут образовывать цепочки (метаболические пути), по которым совершаются цепные реакции: вещество (субстрат) передается от фермента к ферменту и подвергается поэтапной перестройке - усложнению или, наоборот, деградации. Чаше всего такие полиферментные комплексы выстраиваются на мембранах клетки или ее органоидов.
Как действуют ферменты?
Ферменты очень специфичны по типу катализируемой реакции. Известно несколько тысяч ферментов разного действия, и каждый из них имеет специфическую, отличную от других пространственную структуру. Молекула фермента представляет свернутую полипептидную нить, в определенном месте которой включен небелковый компонент - кофермент (часто это бывают витамины, ионы металлов). Именно этот специфичный участок отвечает за функцию фермента, он называется активным центром. Благодаря молекулярной специфичности своих активных центров ферменты узнают и захватывают нужные вещества-субстраты, сближают их для соединения (синтеза) или, напротив, разделяют в реакциях распада (лизиса). При этом сам фермент не разрушается, а лишь изменяет свою третичную белковую структуру - происходит кратковременная и обратимая денатурация молекулы, ее конформационная перестройка
(рис. 8).
Рис. 8
Хотя ферменты и понижают многократно энергию активации соответствующих реакций, для их осуществления все же требуется определенная энергия
. Энергия может генерироваться окислительно-восстановительными кофакторами - такими, как НАД, ФАД или гем. Ферменты класса синтетаз (лигазы) способны расщеплять молекулу АТФ до АДФ и фосфата и используют выделенную энергию на свою конформационную перестройку
(см. сегмент 13 и рис. 5). После произведенной работы по синтезу сложного вещества большая часть энергии не исчезает и не уходит в виде тепла - она лишь переходит в новые химические связи синтезируемого вещества. Впоследствии - в случае расщепления этого вещества - энергия может быть снова извлечена для производства другой работы.
Даже не вдаваясь в детали ферментативного катализа, видно, что эта важнейшая биологическая функция реализуется как сумма простых физико-химических реакций. Важнейшие из них - стереохимическое молекулярное узнавание субстрата ферментом и конформационная перестройка молекулы белка-фермента, суть которой, как и в реакциях мышечного сокращения или активного транспорта, состоит в обратимом изменении силы химических связей внутри молекулы
. Изучение зависимости скорости ферментативной реакции от концентрации фермента и субстрата показало, что в простейших случаях она описывается уравнением Михаэлиса-Ментен. Из этого следует, что в основе ферментативных реакций лежат те же законы и принципы, на которых основаны обычные химические реакции. Однако единая теория ферментативного катализа до сих пор не разработана, так как механизмы протекания ферментативных реакций очень сложны, многофакторны и разнообразны, зависят от большого числа переменных величин, от взаиморасположения ферментов на мембранах и в ряде случае пока не поддаются математическому описанию. В этом и проявляется эмерджентная специфика биологических реакций как специфика нового уровня сложности
(см. сегмент 5).
СЕГМЕНТ 18. ЗАЩИТНЫЕ РЕАКЦИИ. ИММУНИТЕТ
Живые системы - организмы, клетки, клеточные органоиды - должны сохранять биологическую (биохимическую) индивидуальность
. Присоединение к ним чужеродных молекул, контакт с вирусами, бактериями, другими паразитами приводит к искажению соответствующих структур и их функций, например, ферментов или клеточных мембран, не говоря уже о прямом токсическом вреде в случае попадания паразитов в организм, в его клетки. Совокупность чужеродных агентов, противоположных своей генетической сущностью данному организму (чужие или искаженные молекулы, вирусы, бактерии, клетки) называют антигенами
. В виду чрезвычайной важности поддержания биологической индивидуальности, в эволюции организмов возникает и постепенно совершенствуется система реагирования и защиты от антигенов - иммунитет
. Частное проявление иммунитета - невосприимчивость к инфекционным заболеваниям. Изучением защитных реакций организма, направленных на сохранение его структурной и функциональной целостности и биологической индивидуальности, занимается наука иммунология.
Иммунные отношения организма с его антигенами и соответствующие защитные механизмы очень сложны, но хотя бы общее представление о них должен иметь каждый культурный человек. Уже у растений имеются такие защитные факторы, как неспецифические фитонциды - летучие вещества, убивающие бактерий (их много, например, в чесноке и луке), восковый налет на покровных тканях, замуровывание фитопатогенов в клеточных стенках, накопление токсических продуктов в погибших клетках и другие. У животных обособляются специальные клетки - фагоциты, способные пожирать «пришельцев», и другие более специализированные клетки. Причем уровень сложности иммунной системы возрастает в эволюции от простых - беспозвоночных к более организованным позвоночным животным, и наиболее развиты защитные механизмы у млекопитающих, в том числе у человека.
Наш организм прежде всего проявляет различные формы врожденного неспецифического иммунитета
- эволюционно древние, присущие низшим животным. Это - кожные и слизистые барьеры, бактерицидное действие ряда кислот в выделениях потовых и сальных желез, стенок желудка и кишечника, разрушение бактериальных стенок особым ферментом лизоцимом, присутствующим в слезной жидкости, и другие. Проникшие в организм бактерии устраняются фагоцитами и специальными антибактериальными белками, против вирусов действует особый белок интерферон. Кроме того, у высших животных и человека развивается сложная иммунная система, включающая красный костный мозг, вилочковую железу - тимус, селезенку, лимфоузлы, лимфоидную ткань пищеварительных и дыхательных путей (например, в миндалинах - гландах, в аппендиксе). Иммунная система формирует и поддерживает так называемый приобретенный специфический иммунитет.
Центральное место среди клеток иммунной системы занимают лимфоциты
. При контакте с чужеродными антигенами, в зависимости от их природы и структуры, иммунная система дает различные формы иммунного ответа: образование В-лимфоцитами и выделение в кровь специфических белков - антител
(гуморальный иммунитет); размножение Т-лимфоцитов, избирательно реагирующих на чужеродные или собственные мутантные клетки (клеточный иммунитет); появление долгоживущих Т- и В-лимфоцитов «иммунологической памяти», которые при повторной встрече с антигенами способны к быстрому и усиленному ответу; формирование иммунологической толерантности (дословно - терпимости), которая выражается в избирательном отсутствии ответа на данный антиген при повторной встрече; возникновение аллергии - повышенной чувствительности к специфическому антигену. Между прочим, иммунологический конфликт возникает у организма не только с вирусами, бактериями и чужеродными клетками, но также с паразитическими червями (глистами, или гельминтами), пересаженными органами, злокачественными опухолями и даже у беременной матери с ее собственным плодом.
Можно ли чрезвычайно сложные и многообразные защитные механизмы иммунитета свести к простым физико-химическим реакциям, как мы это сделали в отношении биологического движения, транспорта и катализа? Наиболее тонкое распознавание антигенов и максимальную иммунную специфичность обеспечивают антитела - специальные иммунные белки, вырабатываемые лимфоцитами и называемые иммуноглобулинами
. В течение эмбрионального развития и после рождения появляется множество лимфоцитов, активированных на выработку определенного вида антител - против соответствующего вида антигенов. В результате еще до встречи с антигеном в организме предсуществуют группы лимфоцитов, запрограммированных синтезировать антитела ко множеству (не менее 10 тысяч !) различных антигенов. Молекулярная структура антител-иммуноглобулинов хорошо изучена. Это гликопротеидные, то есть белковые в основе, но содержащие углеводную надстройку, молекулы. У них имеются вариабельные по аминокислотному составу концевые участки, которые, как и у ферментов, образуют активный центр. Активный центр антитела
представляет своеобразную молекулярную полость особой конфигурации, которая по размерам и структуре соответствует детерминантным (распознаваемым) участкам молекулы антигена
.
Таким образом, активный центр определяет способность антитела специфически связываться с определенным антигеном. Выполняется принцип молекулярной комплементарности - дополнительности
, подобно тому, как ключ комплементарно соответствует своему замку. Множественные аминокислотные замены в вариабельных частях иммуноглобулинов создают неисчерпаемый набор активных центров, способных связывать любой природный или искусственный антиген. Таким образом, в основе иммунной специфичности, в том числе при распознавании вирусов, бактерий и чужих клеток, лежит простое соответствие стереохимической структуры молекул. Как и в предыдущих примерах с другими функциями белков, в данном случае проявляется вполне материальная сущность сложнейшей реакции, свойственной только живым организмам.
Остается добавить, что препараты специфических антител (так называемые иммунные сыворотки), получаемые от искусственно иммунизированных животных, широко используются для диагностики, предупреждения и лечения инфекционных заболеваний и в некоторых других случаях в медицине и экспериментальной биологии. Например, при укусах ядовитых змей применяют специфическую к данному яду антитоксическую (противоядную) сыворотку. При заражении вирусом клещевого энцефалита собственный иммунитет организма усиливают инъекциями гамма-глобулина. При подозрении на раковую опухоль точный диагноз может дать реакция крови больного на раковые антигены; даже раннюю беременность сегодня легко идентифицируют по обнаружению в крови или моче женщины специфических антигенов зародыша.
Наиболее сложная задача современной иммунологии - найти способы защиты от вируса иммунодефицита человека (ВИЧ)
, вызывающего страшное заболевание - синдром приобретенного иммунодефицита, или СПИД
. Этот вирус избирательно поражает лимфоидные клетки - те самые лимфоциты, которые и должны обеспечивать иммунитет, но, будучи пораженными, не справляются со своими обязанностями. Больной СПИДом начинает серьезно страдать от обычных инфекций, которые здоровый человек переносит как несложную простуду. Чаще всего роковым событием становится появление мутантных злокачественных клеток, которые в отсутствие иммунного надзора разрастаются в раковую опухоль. ВИЧ передается с кровью или половым путем. Эффективные средства лечения пока не разработаны. Единственным способом защиты остается
профилактика от контактов с чужой кровью (для этого и применяют одноразовые шприцы), а также здоровые и осмотрительные отношения в половой жизни
.
СЕГМЕНТ 19. СИГНАЛИЗАЦИЯ. ГОРМОНАЛЬНАЯ И НЕРВНАЯ РЕГУЛЯЦИЯ
Для координации поведения и взаимодействия организмов, а также органов и клеток внутри организма необходима сигнализация
.
Животные используют прежде всего запахи
, то есть химическую сигнализацию. Пахучие вещества - разнообразные молекулы - выделяются у животных кожными железами, с выдыхаемым воздухом, с мочой и другими жидкостями, а у растений листьями, корой, цветками. Воспринимаются запахи у различных животных обонятельными органами или диффузно рассеянными в покровах хеморецепторными клетками. Различают, с одной стороны, аттрактанты - привлекающие вещества, в том числе феромоны - привлекающие полового партнера, с другой - репелленты - отпугивающие вещества. Аттрактанты и репелленты очень эффективны в борьбе с вредными насекомыми, улитками, грызунами и другими животными. Диапазон их действия может достигать нескольких километров. Пахучие травы, масла применяли с древних времен. В настоящее время в качестве аттрактантов и репеллентов все чаще используют синтетические препараты, которые своей молекулярной структурой как бы подражают природным пахучим веществам.
Многие организмы используют для общения свет
. Свечением обладают некоторые бактерии, грибы, простейшие, медузы, ракообразные, насекомые, рыбы. Специальные вещества - люминофоры выделяют порции световой энергии под действием коротковолнового излучения, электрического разряда или химической реакции. Это явление, называемое люминесценцией, известно также в неживой природе - например, свечение белого фосфора, сернистных соединений кальция, бария, стронция. Биолюминесценция используется разными организмами для освещения и приманки добычи, отпугивания хищников, привлечения полового партнера.
Важное значение в сигнализации у животных имеют также звуки, позы, жесты
. В эволюции человека эти формы сигнализации становились ведущими. Придавая особое значение речи
как форме общения людей, академик И.П. Павлов назвал ее второй сигнальной системой
, противопоставив всем остальным сигналам, объединенным в понятие первой сигнальной системы
. Особенность речи как способа сигнализации состоит в том, что в словах содержится обобщение бесчисленных сигналов первой сигнальной системы, и, таким образом, слова становятся «сигналами сигналов». Речь является одним из проявлений и в то же время инструментом высшей нервной деятельности человека (см. ниже).
Внутри организма для взаимодействия между органами, тканями и отдельными клетками и для восприятия сигналов из внешнего мира используются гормональные и нервные механизмы регуляции
.
Гормональная регуляция
осуществляется с помощью биологически активных веществ, среди которых главную роль играют гормоны
(от греческого hormao - побуждаю, привожу в движение). Гормоны выделяются в кровь особыми эндокринными железами, или железами внутренней секреции (гипофиз, надпочечник, щитовидная, поджелудочная, половые и другие железы). Доставка гормонов к органам-мишеням также осуществляется через жидкие среды организма - кровь, лимфу и межклеточные жидкости, поэтому данный механизм передачи сигналов называется гуморальным (от латинского humor - жидкость). Эндокринная система находится под контролем центральной нервной системы. Поэтому нервное возбуждение всегда оборачивается волной гормональных воздействий, которые мобилизуют организм на адекватную реакцию. Например, при стрессовых ситуациях (страх, физическая перегрузка) надпочечники выбрасывают в кровь гормон адреналин, который резко повышает потребление кислорода и концентрацию глюкозы в крови, что, в свою очередь, приводит к увеличению выработки энергии. Таким образом, фактически реализуется единая нейрогормональная регуляция (подробнее о саморегуляции см. тему 4, сегмент 28).
Большинство гормонов по химической природе представляют пептиды (малые белки), бывают также стероиды (из класса липидов) и моноамины (переделанные аминокислоты). Каждый гормон воздействует на определенные клетки-мишени или несколько типов клеток. Восприятие гормонального сигнала осуществляется специальными молекулами-рецепторами (гликопротеидами или гликолипидами), расположенными в оболочке либо в цитоплазме клетки (рис. 9). Возбужденный рецептор, катализируя цепь химических реакций, передает сигнал дальше - на рабочие структуры клетки. В результате идет ответная реакция в форме секреции (выброса активных веществ), специфических синтезов, размножения и роста клеток. Таким образом, гормоны участвуют в регуляции функционирования, роста и развития организма. На всех этапах от узнавания гормона клеточным рецептором до ответной реакции осуществляются элементарные физико-химические процессы: молекулярное узнавание на основе стереохимического соответствия (ключ-замок), обратимое конформационное изменение («вздрагивание») молекул, переход энергии из одной реакции в другую и т. п.
Рис. 9

Нервная регуляция
происходит с помощью особых нервных клеток (нейронов), имеющих длинные отростки и связанные в нервные цепи или сети разной сложности. Нервная регуляция имеется уже у гидр и медуз - наиболее простых многоклеточных животных, а наивысшего развития достигает у позвоночных, особенно у человека с его развитым головным и спинным мозгом, вегетативной системой ганглиев и локальными скоплениями нейронов во внутренних органах. Буквально каждый участок тела пронизан нервными отростками и их разветвленными окончаниями, что позволяет организму иметь информацию о состоянии условий среды во всех его точках и управлять этими состояниями - как правило с участием гормональной регуляции. На основе нервных связей формируются сложные программы внутренней регуляции органов, поведения и высшей нервной деятельности.
Наиболее сложным проявлением высшей нервной деятельности у человека выступает сознание
как высший уровень психической активности. Важнейшей функцией сознания является мышление
с его основными операциями абстрагирования, обобщения, опосредствования и др. Мышление направлено на осознание существа предметов и явлений, созидание новых идей, мысленное построение действий и предвидение их последствий. Мышление представляет высшую форму активного отражения объективной реальности.
Способна ли современная наука объяснить природу нервной деятельности, понять тонкие механизмы работы мозга? В нейробиологии остается много вопросов и белых пятен, поскольку речь идет о самой сложной форме проявления жизни, но элементарные процессы изучены достаточно хорошо. Как уже сказано выше, структурной единицей нервной ткани является нервная клетка - нейрон. Нейроны имеют многочисленные разветвленные связи, особенно сложные в коре головного мозга. Связи распространяются, с одной стороны, на чувствительные рецепторы (кожные, зрительные, слуховые, обонятельные, рецепторы внутренних органов), а с другой - на все регулируемые исполнительные органы (мышцы, пищеварительный тракт, железы и др.). Элементарным явлением в нервной регуляции выступает рефлекс
- ответная реакция органа (организма) на внешнее или внутреннее раздражение, осуществляемая через нервную систему (рис. 10). Представление о рефлексах было выдвинуто еще в 17 веке французским натуралистом и философом Р. Декартом, относившим их к автоматическим непроизвольным действиям. Российский физиолог И.М. Сеченов в 1863 г. утверждал, что «все акты сознательной и бессознательной жизни по способу происхождения суть рефлексы». В 20 веке эта концепция была развита И. П. Павловым в учении о безусловных и условных рефлексах. Многочисленные и разнообразные рефлексы слагаются в сложные поведенческие акты, инстинкты, на их основе и развивается вся высшая нервная деятельность. У низших животных преобладают наследственно закрепленные безусловные рефлексы. а у человека доминируют приобретенные условные рефлексы, закрепляемые в процессах обучения, воспитания, трудовой деятельности.
Рис. 10
Известны и биофизические принципы работы нейронов. По отросткам нейронов сигналы могут передаваться на большие расстояния за сотые доли секунды. Коснитесь рукой горячего предмета - тут же последует рефлекторный ответ. А, между прочим, сигнал успевает пробежать по чувствительным нервным волокнам от пальцев в спинальные ганглии и далее в спинной мозг, переключиться на другие нервные клетки и вернуться к мышцам, отдергивающим руку от горячего предмета (см. рис. 10). Установлено, что передача сигнала по нервному волокну осуществляется с помощью электрических токов и электромагнитных полей
, генерируемых в поверхностной мембране нейрона.
Рассмотрим схему генерации и проведения нервного импульса (рис. 11).
Рис. 11
Изначально благодаря работе ионных насосов (см. сегмент 16, активный транспорт ионов) на мембране нервной клетки накапливается разность потенциалов (плюс снаружи, минус изнутри), достигающая 80 милливольт. Основным носителем внешнего положительного заряда являются ионы натрия. При раздражении участка мембраны раскрываются белковые поры, по которым ионы натрия устремляются в клетку (по закону простой диффузии). Поток заряженных частиц, в данном случае - поток ионов натрия по водно-белковому каналу, представляет электрический ток. Как хорошо известно, электрический ток порождает вокруг проводника электромагнитное поле; то же самое происходит в электромоторе на обмотках ротора. Возникшее электромагнитное поле тут же перебрасывается на соседние белковые поры, раскрывая их для ионов натрия. Порождается цепная реакция от одной поры к другой, которая распространяется вдоль всего нервного волокна. Завершается передача нервного импульса раздражением мембраны на кончике волокна и выбросом порции медиатора - вещества, возбуждающего следующую клетку. Если это будет мышечная клетка, последует сократительная реакция с участием микронитей и миозина (см. сегмент 15 и рис. 7). В соматической нервной системе сигналы проходят особенно быстро, так как большие отрезки волокон покрыты так называемой миэлиновой оболочкой, и электромагнитное поле «перескакивает» через эти участки, а не «ползет» по всем порам мембраны. Ситуация сравнима с той, когда лошадь под всадником или хищник, преследующий добычу, переходят с бега рысью в галоп.
Заметим, что электрической возбудимостью и проводимостью обладают и другие ткани, в частности мышечные пучки сердца. Это позволяет организовать его ритмичную, бесперебойную и в известной мере автономную работу. В случае остановки сердца, если в нем не произошли сильные структурные нарушения, восстановить работу можно разрядами электрического тока, что и делается в экстренной медицине.
Электрическая активность проводников мозга, сердца и других органов слагается в некоторое суммарное биополе каждого отдельного органа и всего организма. Это электромагнитное поле легко регистрируется и служит ценным диагностическим признаком при выявлении заболеваний сердца и мозга (электрокардиография, электроэнцефалография). Особо чувствительные люди - экстрасенсы - способны улавливать своими рецепторами колебания чужого биополя и даже ставить некоторые диагнозы.
Из рассмотрения механизмов биологической сигнализации видно, что в их основе лежат элементарные физико-химические процессы. Белково-углеводные рецепторы клеточных мембран стереохимически распознают различные сигнальные молекулы - аттрактанты и репелленты, гормоны и медиаторы. Восприятие света, а значит и зрительных образов, основано на электромагнитном возбуждении белковых рецепторов в мембранах светочувствительных клеток - колбочек и палочек - в сетчатке глаза. Во всех случаях реагирующие молекулы претерпевают обратимую конформационную денатурацию. В передачу сигналов часто вовлекаются ферментативные реакции, где рабочим моментом также является конформационная перестройка. Практически все сигнальные и регуляторные процессы происходят с затратой энергии. Солнечная энергия в клетках растений генерирует синтез глюкозы, окисление глюкозы у животных дает энергию для синтеза молекул АТФ. Из макроэргических связей АТФ энергия переходит в работу - превращается в другие химические связи, в электрическую, световую, механическую энергию и рассеивается в космос в виде остаточного тепла. Как и другие виды жизнедеятельности, сигнализация, гормональная и нервная деятельность представляют варианты существования и преобразования материи - вещества и поля.
ЗАКЛЮЧЕНИЕ ПО ТЕМЕ 2
Жизнь материальна
, ее физико-химическую основу составляет обмен веществ и энергии. Материя, в том числе живая - это объективная реальность, она существует вне сознания, вне духа, независимо от какой-либо «жизненной силы». Материя
первична, тогда как сознание
вторично, производно от материи, то есть представляет свойство живой материи, одну из форм ее движения
. Движение - всеобщее свойство материи. Это даже больше, чем свойство - это ее способ существования. В этом смысле невозможно разделить саму живую материю и ее функциональные проявления, в том числе невозможно разделить живой мозг и его продукт - сознание. Жизнь - это особенная форма движения особенно сложно
устроенной материи, качественно (эмерджентно) отличная от форм движения неорганической материи.
Это новое качество - жизнь - порождается как сумма свойств составляющих элементов. Количество переходит в качество, целое оказывается больше суммы его частей. Аналогично - формы организации и движения социальных систем отличны от форм организации и движения отдельных живых индивидуумов. Современная биология имеет достаточно фактических оснований для материалистического объяснения сущности жизни.
ТЕМА 3. КОНЦЕПЦИЯ БИОЛОГИЧЕСКОЙ ИНФОРМАЦИИ И САМОВОСПРОИЗВЕДЕНИЯ ЖИЗНИ. ОНТОГЕНЕЗ
Сегмент 20. Самовоспроизведение - важнейшее свойство жизни. Общая схема онтогенеза.
Сегмент 21. Преформизм и эпигенез в истории эмбриологии.
Сегмент 22. Генотип и фенотип организма. Центральная догма молекулярной биологии.
Сегмент 23. Репликация ДНК и размножение клеток.
Сегмент 24. Формы размножения организмов. Клонирование.
Сегмент 25. Развитие организма.
Заключение по теме 3.
СЕГМЕНТ 20. САМОВОСПРОИЗВЕДЕНИЕ - ВАЖНЕЙШЕЕ СВОЙСТВО ЖИЗНИ. ОБЩАЯ СХЕМА ОНТОГЕНЕЗА
Из материала предыдущей темы следует, что основные проявления жизни - обмен веществ и энергии, движение, биокатализ, иммунитет, сигнализация - обеспечиваются разнообразными белками
. Каждый вид белка имеет строго определенную первичную структуру - набор и последовательность аминокислот в полипептидной цепи. Первичная структура предопределяет вторичную и третичную структуры, а третичная структура принципиально важна для функционирования молекулы - от нее зависит стереохимическое узнавание других молекул, дозированные конформационные перестройки, проведение ионов, выстраивание скелетных нитей и т. п.
Однако зададимся вопросом: как долго может работать белковая молекула? Характеризуя биокатализ, мы отмечали, что белки-ферменты в реакциях субстратов сами не разрушаются, они лишь многократно и обратимо денатурируют (см. сегмент 17, рис. 8). Значит ли это, что ферменты работают бесконечно долго и не претерпевают никаких нарушений структуры? Будем исходить из того, что белок - обычное материальное тело, испытывающее регулярные колебательные нагрузки (вспомните образ сжимающейся и разжимающейся стальной пружины). По всем законам сопротивления материалов даже в покоящихся телах, а тем более при регулярной, да еще пульсирующей нагрузке, в них возникают механические напряжения, которые рано или поздно приводят к искажениям структуры - разрывам или смещениям химических связей, нарушениям кристаллической решетки и т. п. Следует вывод: белки со временем теряют нативную (природную, естественную) структуру, они стареют, становятся непригодными к выполнению своих функций. Вместе с белками стареют, изнашиваются клетки, ткани, органы и в целом организмы. Следовательно, в живой природе должны быть механизмы замещения, или воспроизведения, стареющих белков, клеток и самих организмов. Такие механизмы действительно существуют.
В долгоживущих клетках (нервных, мышечных) происходит постоянный синтез новых белков
, идущих на замену изношенным и разрушенным. Такие клетки живут годами и даже десятилетиями, но они обновляются изнутри - подобно тому, как мы ремонтируем дом, не разрушая его как целое строение. В других тканях реализуется стратегия обновления клеточного состава путем регулярного деления молодых клеток
. Так, клетки крови обновляются за несколько недель, кожный эпителий - за неделю, кишечный эпителий - за 2 суток. Но и эти процессы не обеспечивают бессмертия организму. В нейронах накапливаются продукты распада и они погибают, репродуктивный потенциал соматических клеток истощается, наступает старение и смерть организма. Гибель организмов происходит также при их взаимодействии с внешней средой - одним просто не хватает пищи, другие становятся жертвами хищников, третьи погибают от природных катаклизмов (пожары, наводнения, холод и т. п.). Нужен радикальный механизм защиты жизни, ее дублирования, самовоспроизведения
. Для этих целей существуют особые, максимально защищенные от повреждений клетки, которые вовремя «покидают» организм и дают начало новому поколению. Это половые клетки
- предшественники бесконечной череды актов размножения организмов. Мужская и женская половые клетки - сперматозоид и яйцеклетка
- сливаясь, образуют одноклеточную зиготу
, которая в эмбриональном периоде многократно делится и превращается в зародыш
, а далее в постэмбриональном развитии формируется полноценный организм
. Организмы производят новые половые клетки, оставляют потомство и умирают (рис. 12). Весь цикл индивидуального развития
организма от образования зиготы до его смерти называется онтогенезом
.
Рис. 12
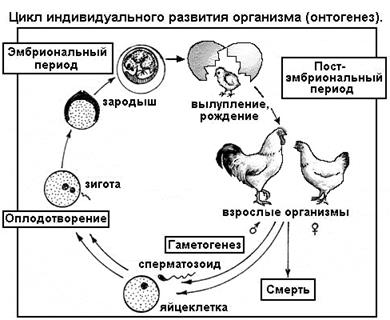
Важно то обстоятельство, что как отдельные клетки, так и целые организмы воспроизводят в поколениях прежний план организации и «образ жизни». Долгоживущие нервные клетки восстанавливают одни и те же органоиды, эритроциты во всех поколениях остаются эритроцитами, лейкоциты - лейкоцитами, а дети в целом похожи на родителей. В чем причина этого сходства? Где и в каком виде содержится и как реализуется информация о строении и свойствах организма
? Успехи современной генетики, цитологии, эмбриологии дают на эти вопросы достаточно ясные ответы. Для контроля развития используется два источника информации: внутренний - генетический
и внешний - эпигенетический
. Однако прежде, чем углубиться в эти понятия, посмотрим на проблему исторически и увидим, что путь к пониманию «золотой середины» лежал, как это обычно бывает, через непримиримый антагонизм альтернативных суждений.
СЕГМЕНТ 21. ПРЕФОРМИЗМ И ЭПИГЕНЕЗ В ИСТОРИИ ЭМБРИОЛОГИИ
В истории человечества существует давний интерес к природе размножения и развития. Эмбриология - наука о зародышевом развитии - одна из древнейших научных дисциплин. От античных времен ведут начало две противоположные точки зрения на причины и движущие силы индивидуального развития организмов - преформизм и эпигенез.
Сторонники преформизма
(от латинского praeformo - заранее образую, предобразую) исходили из того, что все формы, структуры и свойства будущего организма заложены в нем еще до рождения, даже в половых клетках
. Более того, уже в этом еще не родившемся организме содержатся невидимые (очень маленькие) зачатки будущих поколений, как вложенные одна в другую многочисленные матрешки убывающих размеров. Так, выдающийся врачеватель Древней Греции, основоположник медицины Гиппократ (IV в. до н. э.) обращал внимание на то, что цыпленок содержится в яйце в готовом виде со всеми необходимыми членами тела; в ходе его насиживания курицей происходит только рост, увеличение размеров уже предсуществующей формы (отсюда и «пре-формизм»). Наблюдения Гиппократа относятся к периоду зарождения эмбриологии, это ранний преформизм - как одно из ранних проявлений механицизма в биологии
(см. сегмент 11).
Расцвет преформизма приходится на XVII-XVIII в.в., когда эта точка зрения и оформляется в теорию преформации
. Излюбленными аргументами преформистов, кроме цыпленка в яйце, становятся личинка бабочки - куколка, которая содержит все части взрослого организма, многоклеточная шаровидная водоросль вольвокс с вложенными один в другой шарами 2-й и 3-й генерации (это вариант бесполого, или вегетативного, размножения - вроде внутреннего почкования). Уже известен в общих чертах онтогенетический цикл. Благодаря созданию микроскопа (Роберт Гук, 1665 г.) стали известны соматические и половые клетки. Когда стало ясно, что новый организм происходит от слияния яйца и сперматозоида, мнения преформистов о первоисточнике развития резко разделились. Большинство считало, что организм заложен в яйце (оно гораздо крупнее и содержит питательные вещества), тогда как сперматозоид лишь активирует яйцо к развитию. Сторонников этой теории называли овистами (от латинского ovum - яйцо). Другие - их называли анималькулистами (от латинского animalculum зверек, что означало сперматозоид, то есть микроскопический зверек) - видели предсуществующую форму организма именно в сперматозоиде, даже рисовали его в виде скрюченного человечка с хвостом. Яйцо по мнению анималькулистов является лишь питательной средой для развития сперматозоида, подобно тому, как плодородная почва служит кормилицей для прорастающего семени.
Как уже отмечено, преформизм можно отнести к разряду механистических представлений в биологии. По философской сути он является примитивным материализмом
, поскольку возникновение нового организма объяснялось существованием вполне реальных, материальных, хоть и микроскопических, зачатков. Однако в своей крайней форме и в завершенном логическом построении преформизм приходил к догме о заложенных в организме зачатках зародышей всех будущих поколений («вложение зародышей») и, таким образом, смыкался с идеей изначального сотворения живых существ богом, то есть становился «аргументом» в пользу креационизма
(от латинского creatio - создание, сотворение).
В противоположность преформизму сторонники эпигенеза
(от греческих epi - над, сверх, после и genesis - происхождение, возникновение) представляли зародышевое развитие как процесс, осуществляемый путем последовательных новообразований структур из недифференцированной массы оплодотворенного яйца
. Принципиально важным было решение вопроса о движущих силах развития. Эпигенетики невольно приходили к признанию неких внешних нематериальных факторов, управляющих морфогенезом. Так, уже Аристотель в противоречие Гиппократу утверждал, что ни в яйце, ни в семени нет готовых структур взрослого организма; развитием управляет некая высшая цель, жизненная сила - энтелехия
(см. сегмент 11 о витализме). В XVIII веке член Петербургской Академии наук К.Ф.Вольф провел детальное изучение развития яйца курицы и показал, что появление зачатков органов идет постепенно, на месте неоформленной массы желтка. Развитием, по мнению Вольфа, управляет «существенная (эссенциальная) сила» - по сути та же энтелехия.
Таким образом, эпигенез в законченном виде предлагает виталистическое
решение проблемы индивидуального развития. Энтелехия, существенная сила и т. п. нематериальные факторы синонимичны божественной воле, так что все учение становится явно идеалистическим
.
Как ни странно, но и преформисты, и эпигенетики в самой основе своих воззрений были правы. Если от преформизма взять наличие уже в половых клетках некоторого материального образа или программы строения будущего организма, а от эпигенеза поэтапность развития и необходимость его внешнего контроля (хотя бы со стороны материнского организма), то в итоге мы получим современную трактовку биологии индивидуального развития, которая признает наличие как генетических, так и эпигенетических начал развития. Суть этих начал мы и рассмотрим в следующих сегментах.
СЕГМЕНТ 22. ГЕНОТИП И ФЕНОТИП ОРГАНИЗМА. ЦЕНТРАЛЬНАЯ ДОГМА МОЛЕКУЛЯРНОЙ БИОЛОГИИ
Сегодня каждый школьник знает, что организм имеет наследственную (генетическую) информацию
о своих внешних признаках (строении, свойствах, поведении), которая передается от родителей к детям. Большинство сумеет объяснить, что эта информация в виде генов
хранится и передается в хромосомах
. Многие скажут, что ген - это участок молекулы ДНК, отвечающий за отдельный признак
. Кое-кто вспомнит, что ДНК обеспечивает синтез белков, которые, в основном, и формируют внешние признаки
. К этому следует добавить, что совокупность генов данного организма, то есть его генетических задатков, обозначается понятием генотип
(от греческого genos - род, происхождение)
, а то, что получается в итоге - совокупность внешних признаков организма - называется фенотипом
(от греческого phaino - являю, обнаруживаю)
. Наиболее думающие заметят, что под «внешними признаками» подразумеваются не только структурные свойства организма - форма частей тела, окраска и т. п., но также и функциональные характеристики - скорость роста, мышечная сила, характер питания, устойчивость к болезням и многое другое, что определяется функциями белков
. Наконец, самые памятливые должны подсказать нам, что фенотип зависит не только от генотипа, но и от условий среды
, в которых развивается организм, и что пределы варьирования фенотипа при неизменном генотипе обозначаются как норма реакции
. Вот в сущности и вся квинтэссенция молекулярной биологии развития. Но почему белкам нужна внешняя информация? Почему информация хранится в ДНК? Как реализуется и чем контролируется эта информация?
В сегменте 20 мы выяснили, что белки, как и целые клетки, организмы, стареют и разрушаются, поэтому их надо создавать заново. Синтез новых белков необходим в каждом новом поколении клеток и организмов, а в долгоживущих клетках он происходит ежедневно и ежечасно. Но каждый тип белка имеет уникальную, строго обязательную последовательность из аминокислот 20 разновидностей (первичная структура), которая должна быть воспроизведена в точности. В противном случае это будет искаженный белок или бессмысленный, не функциональный полипептид. Вот почему в каждой клетке должна быть информация
о первичной структуре белков
, причем эта информация должна копироваться и наследоваться, чтобы воспроизводить те же белки в поколениях. Проблемами наследования биологической информации занимается наука генетика
, а также возникшая на стыке химии, биологии и генетики молекулярная биология
. В понимание теоретических основ биологической информации внесла свой вклад и кибернетика
.
Заметим, что представление о генах как носителях наследственных признаков и сама наука генетика возникли еще в конце XIX века. В 1865 г. чешский монах Грегор Мендель при скрещивании разных сортов гороха открыл первые законы наследования отдельных признаков, доказал дискретность признаков, то есть их раздельное, независимое друг от друга существование и наследование. Представления о том, что такое гены, Мендель не имел и не мог иметь, так как ни ДНК, ни хромосомы тогда не были известны. Поскольку результаты наблюдений Менделя были опубликованы в мало доступном издании, о них узнали уже задним числом, когда в 1900 г. те же законы были переоткрыты другими учеными. К этому же времени стало ясно, что носителем генов являются микроскопические тельца - хромосомы, содержащиеся в клеточных ядрах, хорошо окрашиваемые (отсюда и название: от греческих chroma - окраска, цвет и soma - тело) и видимые во время деления клетки. Гораздо позднее - в 40-е годы XX века состоялось открытие ДНК и ее связи с хромосомами, а в 50-60-е годы установлена структура ДНК, выяснены причинные связи ДНК и белков, то есть генотипа и фенотипа.
Итак, в середине XX века состоялись великие научные открытия, обозначившие начало молекулярно-биологической революции в естествознании. Молодые исследователи Джеймс Уотсон из США и Френсис Крик из Великобритании в 1953 году расшифровали строение молекулы ДНК - материального носителя генетической информации (см. сегмент 7 и рис. 2). Следом был раскрыт и механизм передачи информации с ДНК на синтезируемый белок. Абстрактное до сих пор понятие гена приобрело материальный смысл и получило функциональное объяснение. Принцип функционирования ДНК (гена)при реализации наследственной информации был обозначен как центральная догма молекулярной биологии
. Вот ее краткая формула: ДНК
- РНК
- белок
.
Оставляя пока в стороне РНК, выделим самую суть догмы: информация о первичной структуре белка
(например, какого-либо фермента) заключена в молекуле ДНК
, то есть в молекулярной структуре гена
. Процесс передачи информации однонаправленный: по ДНК можно построить белок, но обратный процесс - синтез ДНК по белку - не возможен.
Вспомним, что белок и ДНК являются полимерами, но у них разные мономеры: аминокислоты в белке и нуклеотиды в ДНК (см. сегмент 7 и рис. 2). Таким образом, принцип кодирования
белка с помощью ДНК состоит в том, что последовательность
(порядок чередования) мономеров-аминокислот в белке зависит от последовательности мономеров-нуклеотидов в молекуле ДНК
. Важно понимать, что процесс «передачи информации» от ДНК на белок означает реакцию синтеза белка
, то есть процесс сборки новой полимерной белковой цепи из отдельных аминокислот, которые поступают в клетку в качестве пищи.
Как видно из формулы центральной догмы, передача информации, то есть синтез белка, осуществляется с помощью особого посредника - молекулы информационной РНК
(рибонуклеиновая кислота)
. РНК, как и ДНК, состоит из 4 типов нуклеотидов (только тимин - Т заменен на близкий ему урацил - У), но в отличие от ДНК представляет одноцепочечный полимер (см. сегмент 7 и рис. 2). РНК синтезируется из свободных нуклеотидов на одной из цепей молекулы ДНК по принципу комплементарности (дополнительности) и фактически является рабочей копией гена
, так как ее нуклеотиды повторяют нуклеотидный набор одной из цепей ДНК (рис. 13). Этот этап называется транскрипцией
(от латинского transcriptio - переписывание) и осуществляется в ядре клетки, где хранится ДНК. Теперь РНК переходит в цитоплазму, и по ней идет синтез белка с помощью так называемых транспортных РНК и специальных органоидов рибосом. Транспортные РНК выступают в роли «переводчика» с 4-буквенного языка нуклеотидов на 20-буквенный язык аминокислот, что следует из их структуры. Один конец тРНК имеет вывернутый наружу триплет (тройку) нуклеотидов - антикодон, который присоединяется к комплементарному триплету иРНК - кодону, а другой конец держит аминокислоту, строго соответствующую данному антикодону. Когда две молекулы тРНК, нагруженные аминокислотами, сойдутся на соседних кодонах иРНК, рибосома обеспечивает соединение висящих аминокислот (см. рис. 13). Так, шаг за шагом осуществляется синтез полимерной цепи белка. Этот второй этап передачи информации от РНК к белку называется трансляцией
(от латинского translatio - передача). Как видно, генетический код является триплетным
- он устроен так, что трем нуклеотидам цепи ДНК (РНК) соответствует одна аминокислота белковой цепи.
Рис. 13

Таким образом ДНК (ген) выполняет роль матрицы
(шаблона, образца) для синтеза белка
. Информационная РНК служит промежуточной матрицей, то есть рабочей копией гена. В целом в молекулярной биологии выполняется принцип: один ген - один белок
.
Почему именно ДНК выбрана Природой для кодирования структуры белков, не известно. В 20-х годах Российский биолог Н.Кольцов высказал идею о самовоспроизведении белков по матричному принципу. В отношении матричного принципа догадка была гениальной, но роль матрицы, как выяснилось позднее, выполняют не сами белки, а молекулы ДНК - полимера совершенно не похожего на полипептидные цепи белков. Согласно одной из новых гипотез (Костецкий, 1999) уже в момент зарождения жизни на Земле на основе перестройки кристаллов природного минерала апатита сразу возник комплекс ДНК и простого белка. Кристаллы апатита изначально имели многочисленные искажения кристаллической решетки («мутации»), что и послужило первичным источником разнообразия генов и, соответственно, белков. В последствии искажения нарастали, возрастало разнообразие структур и функциональных свойств белков. Таким образом, ДНК-белковое соответствие является изначальным свойством жизни.
СЕГМЕНТ 23. РЕПЛИКАЦИЯ ДНК И РАЗМНОЖЕНИЕ КЛЕТОК
Процесс биосинтеза белка - транскрипция и трансляция - это по сути реализация
имеющейся в клетках генетической информации, направленная на рост клеток и межклеточных структур, в итоге - на построение тела организма. Но, кроме синтеза белков, индивидуальное развитие включает размножение клеток
, а через половые клетки - и размножение самих организмов. Для этого
необходим механизм воспроизведения генетической информации
. Копирование генетической программы осуществляется посредством репликации ДНК
в цикле деления клетки
(рис. 14).
Рис. 14
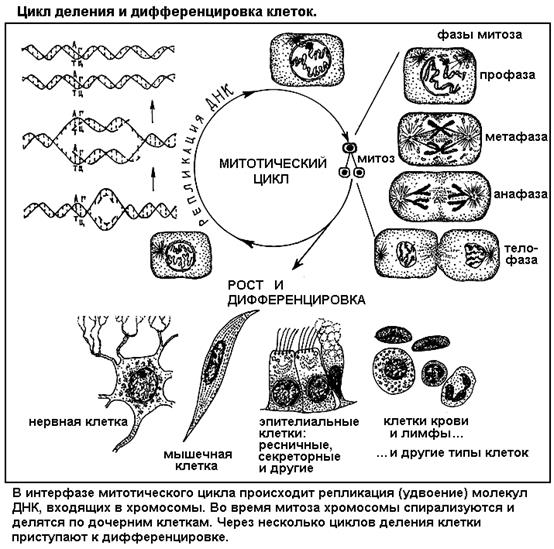
Репликация
(от латинского replicatio - повторение) означает процесс матричного синтеза молекулы ДНК, ведущий к ее самовоспроизведению (удвоению)
. Материнская молекула ДНК раскручивается и с помощью фермента ДНК-полимеразы достраивается по обнаженным цепям свободными нуклеотидами. Как и при синтезе РНК, подбор парных нуклеотидов идет по
принципу комплементарности
: аденин - тимин, гуанин - цитозин.
В результате происходит образование двух молекул ДНК, которые идентичны друг другу и, в то же время, повторяют материнскую молекулу. При этом каждая вновь образованная молекула содержит одну старую и одну новую цепочки нуклеотидов (полуконсервативный механизм).
Когда в 1953 г. Уотсон и Крик показали возможность копирования ДНК по принципу комплементарности, научная общественность была в шоке - настолько неожиданно прост и надежен оказался механизм наследственности! Не столько сама структура ДНК, сколько вытекающий из нее механизм самокопирования представлял наибольший интерес в их открытии. В 1962 г. Джеймс Уотсон и Френсис Крик вместе с Морисом Уилкинсом (именно его рентгенограммы ДНК использовались для обоснования биспиральной модели) были удостоены Нобелевской премии.
Ядерная ДНК находится в составе хромосом
(от греческих chroma - цвет, окраска и soma - тельце). Эти структуры хорошо окрашиваются специальными красителями и отчетливо видны во время деления клетки.
Каждая хромосома представляет одну гигантскую (до нескольких сантиметров длиной!), плотно свернутую молекулу ДНК, подразделенную на информационные участки - гены. Обычно полный комплект генов распределен в несколько хромосом разной величины, число которых строго специфично для каждого вида животных или растений (хромосомный набор, или кариотип вида). У человека в разных хромосомах содержится от сотен до нескольких тысяч генов, а всего в хромосомном наборе человека имеется порядка 60 тысяч разных генов (соответственно, столько же видов белков функционируют в нашем организме).
Начиная от зиготы, во всех поколениях клеток имеется один и тот же набор хромосом, причем этот набор двойной, или диплоидный
. Диплоидность возникает в момент оплодотворения - один хромосомный набор дает яйцеклетка, а второй - сперматозоид. Так, у человека в каждой соматической клетке содержится 23 хромосомы от матери и 23 такие же (за исключением одной - половой хромосомы) от отца; всего диплоидный набор человека составляет 46 хромосом, или 23 пары хромосом. Таким образом, каждый признак, то есть каждый белок, закодирован в клетке не одним, а двумя генами, от двух гомологичных (одноименных) хромосом. О преимуществах такого дублирования генов мы еще поговорим при рассмотрении полового размножения.
В цикле деления клетки в результате репликации ДНК в каждой хромосоме становится по 2 молекулы ДНК, а сами хромосомы приобретают вид Х-образных телец, так как состоят из 2 равных половинок - хроматид. Таким образом, число одинаковых генов в клетке временно становится равным четырем. Далее проходит митоз
- так называется процесс разделение хромосом на отдельные хроматиды (теперь они становятся самостоятельными хромосомами) и, далее, деление самой клетки надвое (см. рис. 14). В результате митоза в дочерних клетках воспроизводится исходный диплоидный набор хромосом,
поэтому каждая дочерняя клетка обладает той же информацией о белках, что и материнская клетка.
Деление клеток приводит к увеличению их числа. В крупных организмах насчитываются миллионы, миллиарды и триллионы клеток. На этой основе возможна дифференциация клеток
- появление структурно-функциональных различий между ними. В дифференцированных клетках синтезируются разные РНК и белки, строятся разные рабочие структуры, в итоге клетки имеют разный вид и выполняют разные функции. У человека в различных органах и тканях насчитывают более 200 типов клеток: разнообразные нервные, мышечные, эпителиальные, кровяные, опорные и другие клетки.
Таким образом, индивидуальное развитие
отдельной клетки или целого организма
слагается из 2 процессов: размножения и роста.
Соответственно этому на молекулярном уровне
процесс развитие включает 2 операции:
репликация ДНК - копирование генотипа;
синтез белков тела клетки или организма - построение фенотипа.
Так что все живые организмы представляют неразрывное единство фенотипа и генотипа
, то есть сомы (тела) и программы ее развития (ДНК), передающейся по наследству (соматическая и наследственная плазма Вейсмана).
Теперь схему центральной догмы молекулярной биологии
можно дополнить механизмом репликации ДНК, после чего она будет иллюстрировать всю молекулярно-биологическую сущность индивидуального развития (рис. 15)
.
Рис. 15

Внимательный читатель заметит, что рассмотренная схема индивидуального развития отражает только его генетическую (внутреннюю) сущность. Но еще в начале мы сказали (устами самого памятливого школьника), что внешний облик организма - его фенотип - зависит не только от генотипа, но и от условий среды
. При этом «условия среды» понимаются в широком смысле: это все, что так или иначе влияет на работу генов. Запомним это важное положение и вернемся к нему при рассмотрении механизмов эмбрионального развития, а пока продолжим разговор о размножении, но от клеток перейдем к целым организмам.
СЕГМЕНТ 24. ФОРМЫ РАЗМНОЖЕНИЯ ОРГАНИЗМОВ. КЛОНИРОВАНИЕ
Размножение организмов
является их важнейшим свойством и служит безграничному продолжению жизни в новых поколениях, при том, что отдельные особи смертны. Существует два способа размножения организмов - бесполое и половое
.
Бесполое размножение
наиболее распространено у растений
. Это известно любому дачнику. В практике широко используется размножение черенками (смородина), усами (земляника), клубнями (картофель) и другими частями тела. Поскольку новые растения вырастают из вегетативных (не половых) органов, этот механизм называют также вегетативным размножением. Реже встречается бесполое размножение
у животных
, причем с повышением уровня организации эта способность снижается, а у высших животных и человека она совсем утрачивается. Легко почкуются гидры и медузы. Плоские черви планарии восстанавливаются из оторванных частей тела. Морские звезды вырастают со всеми органами из отдельных оторванных лучей. Но если разрезать пополам насекомое, например, жука или муху, или позвоночное животное - рыбу, лягушку, не говоря уже о птицах и млекопитающих, то из этого ничего хорошего не выйдет.
Общим принципом бесполого размножения является участие одного «родителя»
. Важно и то, что дочерние особи образуются из обычных соматических (не половых) клеток, которые делятся митозом
, как это описано выше. Это означает, что все поколения имеют в клетках один и тот же набор хромосом, один и тот же генотип
- от одного родителя. Поэтому земляника, разводимая усами, или картофель при посадке клубнями будут сохранять из поколения в поколение одни и те же сортовые качества. С годами они заражаются вирусами, приобретают случайные нежелательные мутации и стареют, как стареет отдельный организм. Время от времени посадочный материал приходится менять на молодой, полученный при половом размножении. Но об этом чуть позже.
Одной из форм бесполого размножения является клонирование
растений и животных
. Теория и практика клонирования уже давно не новые, хотя заговорили об этом в последние годы, когда в 1997 году в Шотландии удалось вывести знаменитую овечку Долли. Но практичные японцы без шума в прессе выращивают клонированных коров уже с начала 90-х годов. Что же такое клонирование?
Клон - это совокупность генетически однородных особей, полученных путем простого митотического размножения клеток без полового процесса, без хромосомного комбинирования
. Клонированные организмы генетически идентичны, как однояйцовые братья или сестры. Этим клон отличается от потомков, полученных половым путем, у которых имеются хромосомы двух родителей. По сути потомство одного растения или животного, полученное обычным черенкованием или почкованием, это тоже клон. Но в последнее время клонирование особей ведут из отдельных клеток. Это повышает генетическую чистоту клона и имеет ряд других преимуществ. Схемы клонирования растений и животных приведены на рис. 16а и 16б
Рис. 16а
У растения берут какую-нибудь ткань, например, кусочек корнеплода моркови, помещают в колбу или пробирку с плотной питательной средой, добавляют гормоны роста. Через некоторое время клетки теряют признаки прежней дифференцировки и приступают к размножению. В это время клетки можно рассадить по одной штуке в множество колб или пробирок, и процесс пойдет с прежним темпом в каждой из них. Образуется клеточная масса (каллус), в которой далее идет дифференциация по разным тканям, формируются органы: корень, стебель, листья и в конце концов цветки. Растение в пробирке готово. Ценность метода клонирования растений заключается в том, что таким образом удается вырастить стерильный, не пораженный вирусами или бактериями, посадочный материал. Если это к тому же элитный сорт, урожай после пересаживания в открытый грунт будет превосходный. Естественно, делается это в специальных лабораториях, стерильно, с применением особых и дорогих сред и стимуляторов роста, поэтому широкого применения в практике клонирование растений пока не получило.
Рис. 16б
 Клонирование животных в хозяйственных масштабах также делает первые шаги. Главный резон этой технологии - рациональное использование генотипов элитных, особо ценных и редких производителей. Для клонирования берут соматические клетки эмбриона или даже взрослого животного. Так для выращивания овечки Долли взяли клетки молочной железы взрослой овцы. Клетки размножают в культуре, получается «посадочный» материал из генетически однородных клеток, которые представляют клеточный клон. Каждая клетка содержит ядро, а в ядре диплоидный (элитный!) набор хромосом. Теперь выращенные элитные ядра можно пересаживать в яйцеклетки других особей того же вида, предварительно удалив из них собственные ядра. Яйцеклетка с встроенным полноценным (то есть диплоидным) ядром представляет аналог зиготы, образуемой при обычном оплодотворении. Далее такую комбинированную зиготу имплантируют(пересаживают, вживляют) в матку взрослой самки для эмбрионального развития. Это может быть та же особь, от которой брали соматические клетки с ядрами, или особь, давшая яйцеклетку, или вообще другая самка. Так, злополучная Долли, зачатая без участия отца, имеет трех матерей, поскольку ядро и яйцеклетка были получены от двух разных овец-доноров, а реципиентом, принявшим зиготу для дальнейшего развития, была третья овца. В 1999 году было сообщено о клонировании обезьяны от разделенного на отдельные клетки 4-клеточного зародыша (после того, как зигота проделала два цикла деления). Эта методика значительно проще и дешевле, чем клонирование от более поздних стадий развития (как было в случае с Долли), и она по сути воспроизводит лишь то, что иногда происходит в природе. Именно так, но без участия экспериментатора, получаются обычные однояйцовые близнецы: начавшая делиться зигота по разным причинам не удерживает дочерние клетки, они расходятся и развиваются далее самостоятельно. Получаются абсолютно идентичные дети. Клонирование животных в хозяйственных масштабах также делает первые шаги. Главный резон этой технологии - рациональное использование генотипов элитных, особо ценных и редких производителей. Для клонирования берут соматические клетки эмбриона или даже взрослого животного. Так для выращивания овечки Долли взяли клетки молочной железы взрослой овцы. Клетки размножают в культуре, получается «посадочный» материал из генетически однородных клеток, которые представляют клеточный клон. Каждая клетка содержит ядро, а в ядре диплоидный (элитный!) набор хромосом. Теперь выращенные элитные ядра можно пересаживать в яйцеклетки других особей того же вида, предварительно удалив из них собственные ядра. Яйцеклетка с встроенным полноценным (то есть диплоидным) ядром представляет аналог зиготы, образуемой при обычном оплодотворении. Далее такую комбинированную зиготу имплантируют(пересаживают, вживляют) в матку взрослой самки для эмбрионального развития. Это может быть та же особь, от которой брали соматические клетки с ядрами, или особь, давшая яйцеклетку, или вообще другая самка. Так, злополучная Долли, зачатая без участия отца, имеет трех матерей, поскольку ядро и яйцеклетка были получены от двух разных овец-доноров, а реципиентом, принявшим зиготу для дальнейшего развития, была третья овца. В 1999 году было сообщено о клонировании обезьяны от разделенного на отдельные клетки 4-клеточного зародыша (после того, как зигота проделала два цикла деления). Эта методика значительно проще и дешевле, чем клонирование от более поздних стадий развития (как было в случае с Долли), и она по сути воспроизводит лишь то, что иногда происходит в природе. Именно так, но без участия экспериментатора, получаются обычные однояйцовые близнецы: начавшая делиться зигота по разным причинам не удерживает дочерние клетки, они расходятся и развиваются далее самостоятельно. Получаются абсолютно идентичные дети.
Проблема клонирования решена в биологическом плане, но встали чисто гуманитарные вопросы о допустимости клонирования человека. Шотландский ученый Ян Вильмут, который со своими сотрудниками и проделал опыты по выращиванию Долли (и наверное может считаться ее «крестным отцом»), высказался против клонирования людей. В ряде стран такие эксперименты запрещены законодательно. Но история науки и техники знает, как трудно удержать в стенах лаборатории выдающиеся открытия. Так было с расщеплением радиоактивных материалов, которое вылилось в гонку ядерных вооружений, так же, вероятно, будет и с клонированием человека. Последствия пока трудно предвидеть, но это вопрос не биологический. Это поле деятельности политиков, юристов, правозащитников, церкви - всей мировой общественности.
Половое размножение
принципиально отличается от бесполого и различных форм клонирования тем, что в образовании нового организма участвуют два родителя разного пола
. Как уже показано в сегменте 20 и на рис. 12, первая клетка нового организма - зигота - образуется от слияния яйцеклетки и сперматозоида. Их ядра несут по гаплоидному (одинарному) набору хромосом, так что ядро зиготы получает после слияния нормальный диплоидный набор. Но если при бесполом размножении во всех поколениях копируется одна и та же комбинация парных (аллельных) генов, то в зиготе всякий раз образуются новые комбинации из отцовских и материнских генов.
Надо понимать важную тонкость природы генов. Как мы уже знаем, ген - это участок молекулы ДНК, отвечающий за синтез одного вида белка. Но дело в том, что под действием различных факторов среды гены постоянно изменяются, то есть мутируют
, и, если эти мутации не приводят к полной потере структуры и функции кодируемого белка, такие организмы выживают, и мутация закрепляется в потомках. Если родители не являются близкими родственниками, то как правило, у них один и тот же ген чем-нибудь да различается, а если у них появляются дети, то эти различия комбинируются. Так, если одна половая клетка несет полноценный ген (исходный, или дикий, тип), а в другой этот же ген мутантный, то после слияния хромосом в зиготе полноценный ген доминирует над мутантным (доминантный и рецессивный гены, аллели), но если оба гена мутантные (рецессивные), причем каждый в своей степени, то в фенотипе проявится один из них. Это будет новый фенотипический признак, отсутствовавший у дикого предка. Кроме того, если углубиться в процесс формирования самих половых клеток, то можно увидеть, что в процессе мейоза (особое деление, которое и приводит от диплоидного набора хромосом к гаплоидному) идет так называемый кроссинговер
- схождение и коньюгация одноименных (гомологичных) хромосом, разрезание нитей ДНК, их переворот крест-накрест и сшивка в новом сочетании генов. Так что даже у одного родителя в разных половых клетках (допустим - в двух сперматозоидах) комбинации разнокачественных генов получаются разные. Поэтому у одной родительской пары дети различаются по многим признакам (кроме уже упомянутых случаев однояйцовых близнецов). Кроме того, яйцеклетка имеет большой объем цитоплазмы с питательными веществами и некоторыми дополнительными цитоплазматическими генами (нехромосомная ДНК ряда органоидов), которые тоже вносят свой вклад в генетическое разнообразие потомков.
Таким образом, в результате случайного сочетания отцовских и материнских хромосомных наборов, в которых одноименные гены могут заметно различаться, потомство получает самые разнообразные комбинации признаков родителей, так что у детей общий баланс генов - генотип - будет отличаться и от отцовского, и от материнского
. Поэтому у людей разный цвет кожи и глаз, разная форма носа, разные группы крови, разные предпочтения в пище (разные ферменты и рецепторы), разные интеллектуальные способности и т. д. и т. п. На Земле нет двух одинаковых людей, если они не однояйцовые близнецы.
Заметим, что половое размножение свойственно всем эукариотным организмам, начиная от одноклеточных водорослей и простейших и кончая человеком. (Даже у прокариот, у бактерий, где нет настоящей диплоидности и полового размножения, внехромосомные гены - плазмиды - могут мигрировать от одной клетки-особи к другой, внося генетическое разнообразие в популяцию). Те виды, которые хорошо размножаются бесполым путем (растения, низшие беспозвоночные - см. выше), через несколько вегетативных поколений обязательно вступают в половой процесс. Комбинативная изменчивость
особей одного вида, возникающая только при половом размножении, - очень важный фактор генетического разнообразия, которое служит материалом для отбора на лучшие комбинации признаков. В этом - движущий фактор эволюции.
Исключение полового размножения, переход на варианты клонирования обедняет генетическое разнообразие и становится тормозом эволюции. С другой стороны, бесполое размножение и клонирование позволяют тиражировать однажды полученную генетическую комбинацию во многих поколениях, что и используется в сельскохозяйственной практике.
СЕГМЕНТ 25. РАЗВИТИЕ ОРГАНИЗМА
Развитие нового организма, полученного половым путем, начинается от оплодотворенной яйцеклетки - зиготы и заканчивается смертью. В сегменте 20 мы уже рассмотрели общую схему онтогенеза, идущего от слияния половых клеток (см. рис. 12). Важно помнить, что в ядре зиготы содержатся два набора хромосом от двух родителей (гибридный генотип). При клонировании в целом выдерживается та же схема онтогенеза, с той лишь разницей, что зигота имеет подсаженное диплоидное ядро от одного родителя.
Биологическое развитие происходит по общим диалектическим
принципам развития
, которое можно наблюдать в неживой природе или в обществе. Чтобы убедиться в этом, сопоставим стадии «развития вообще», заимствованные нами из философского словаря, и стадии нормального онтогенеза многоклеточного организма, например, человека.
| Стадии «развития вообще»
|
Стадии онтогенеза человека
|
| Подготовка предпосылок развития -
внешнее движение, совершаемое пока что за пределами данной системы.
|
Предзародышевое развитие -
образование половых клеток (гаметогенез), формирование окружающей среды будущего организма.
|
| Возникновение -
переход к внутреннему движению и возникновение системы.
|
Оплодотворение -
слияние половых клеток, возникновение новой клетки - зиготы.
|
| Формирование -
преобразование новым процессом развития тех условий, из которых он возник.
|
Зародышевое развитие -
эмбриогенез, построение принципиально новой многоклеточной системы.
|
| Собственно развитие -
зрелость процесса развития, его существование на своей основе.
|
Послезародышевое развитие -
постэмбриогенез. У человека выделяют: период роста (0-20 лет),
репродуктивный период (20-50 лет),
период старения (после 50 лет).
|
| Умирание -
разрушение процесса развития.
|
Смерть - конец индивидуального развития, распад структуры.
|
Индивидуальное развитие системы, в том числе организма, происходит циклично
, так что восходящее развитие
всякий раз сменяется нисходящим
. Восходящее развитие идет от простого, низшего (предзиготическая стадия) к сложному, высшему (многоклеточный организм). Нисходящее - от сложного, высшего (многоклеточный организм) к простому, низшему (бесклеточная мертвая материя). Законы диалектики утверждают, что развитие как конечный процесс с самого начала в скрытом виде содержит тенденции, ведущие от низшего к высшему и обратно. То есть развитие имеет векторный, направленный характер
.
Чем же задается этот вектор? Чем детерминировано (предопределено) индивидуальное развитие?
Где программа развития и как она реализуется? Как из оплодотворенного яйца - из одной клетки - получается сложный многоклеточный организм, в котором тканевые клетки с одинаковым набором генов (одним генотипом) имеют разную структуру и функции (разный фенотип)? Поставленные вопросы составляют основной научный смысл науки эмбриологии или, говоря шире, биологии индивидуального развития. Они имеют и практическое медико-биологическое значение, так как нарушение процессов развития приводит к болезни и сокращает сроки полноценной жизни человека.
Чтобы сократить путь к пониманию программы развития, вспомним главную идею о том, что развитие находится под контролем двух начал - генетического (внутреннего) и эпигенетического (внешнего)
. Найдем эти начала в развивающемся организме.
Внутренняя, генетическая программа развития
заложена в ДНК зиготы. Это генотип организма.
Помним, что при размножении клеток - от зиготы до самой последней клетки тела - ДНК каждый раз удваивается и делится поровну, так что все клетки получают полный набор генов. В ДНК записана информация о всех белках организма.
При этом надо иметь в виду, что существуют гены и белки двух классов: структурные и регуляторные
. Первые обеспечивают построение рабочих структур клеток и межклеточного вещества, ферментативный катализ, транспорт и прочие жизненно важны функции. Вторые регулируют активность первых, то есть гены-регуляторы производят соответствующие регуляторные белки, которые управляют активностью структурных генов. Сейчас установлено, что и среди регуляторных генов есть взаимозависимость - одни гены активируются другими. Таким образом, гены образуют функциональные цепи с заранее предопределенной последовательностью включения
. Работает принцип домино: продукт первого гена активирует второй ген, продукт второго - третий и т. д. Благодаря слаженной работе таких конвейеров контролируются тесно увязанные шаги морфогенеза, развитие приобретает динамичный и направленный (векторный) характер
.
Однако организм - очень сложная система, чтобы ее развитие было выстроено по простому алгоритму домино. Отдельные морфогенетические процессы часто идут независимо и параллельно. В разных зачатках эмбриона, а потом в клетках разных тканей эти процессы расходятся, идет дифференциация клеток по функциям. Но при этом все клетки имеют один и тот же набор генов (!). Возникает ключевой вопрос проблемы клеточной дифференциации -
почему при одинаковом наборе генов синтезируются разные белки и получаются разные клетки?
Современная биология развития дает ответ и на этот сложный вопрос.
Внешняя, эпигенетическая программа развития
контролирует и направляет реализацию генетической программы.
Под действием внешних сигналов, биологически активных веществ, через посредство клеточных рецепторов
и внутриклеточных мессенджеров (молекул-посланников) происходит избирательная активация одних генов и подавление других
. В итоге в дифференцированных клетках разных органов и тканей работают не все гены, а только та их часть, которая ответственна за данную тканевую функцию. Генетики называют этот механизм дифференциальной экспрессией генов. Но встает новый вопрос: что является самой первой командой к дифференциации клеток? Ведь развитие начинается с одной клетки - зиготы.
Установлено, что в онтогенезе работают, сменяя друг друга, три системы регуляции.
1. Эмбриональная детерминация
развития на основе позиционной информации, заложенной в яйце.
В процессе роста и созревания яйцеклетки, когда она еще находится в материнском организме, в ее цитоплазме неравномерно откладываются различные РНК и белки-регуляторы, которые предопределяют будущий план раннего развития и ранней дифференциации клеток. Начало этой топологической неоднородности цитоплазмы яйца задает его полярное положение в яичнике: одним полюсом яйцо контактирует со стенкой (отсюда идет питание), другим обращено в просвет (здесь сосредотачиваются продукты сложных синтезов) (рис. 17а). Таким образом, еще до начала развития генетический материал (хромосомы в ядре) лежит в неоднородной, анизотропной среде, насыщенной биологически активными регуляторами. Уже в яйце мы имеем сочетание генетического (хромосомы) и эпигенетического (цитоплазма с регуляторами) факторов развития.
После оплодотворения зигота многократно делится, и дочерние ядра попадают в различно детерминированные участки цитоплазмы, содержащие разные регуляторы (рис. 17б). Эти регуляторы и становятся первыми внутренними индукторами дифференцировки эмбриональных клеток.
Рис. 17а,б
2. Эмбриональная индукция
- влияние одних зачатков на другие с помощью выделяемых клетками веществ-регуляторов.
Этот механизм включается в ранних зародышах и представляет по сути эмбриональную гуморальную регуляцию; внешние регуляторы - индукторы выступают в роли первых гормонов. Так, например на стадии гаструлы (двухслойный зародыш) под действием выделений внутреннего слоя клеток впячиваются и дифференцируются клетки будущей нервной системы (рис. 17 в).
Рис. 17в
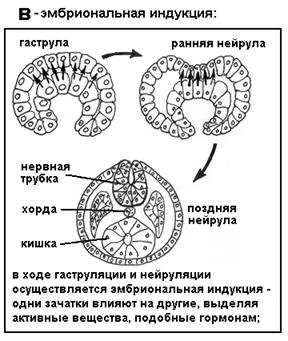
3. Нейрогормональная регуляция
дефинитивного (окончательного) типа, осуществляемая сложной системой желез внутренней секреции и нервной системой
(рис. 17 г).
Заметим, что гормоны вырабатываются железами под контролем нервной деятельности, а нервная система в свою очередь находится под воздействием внешней среды.
Рис. 17г
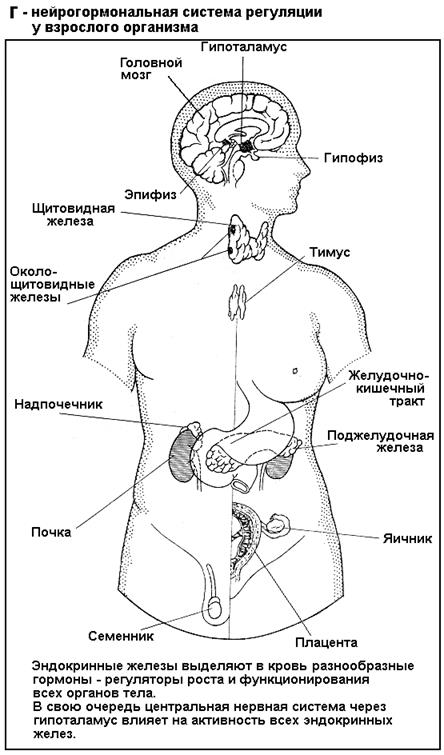
Таким образом, окружающая среда генотипа сформированного организма, например, человека, включает цепь управляющих факторов
: факторы собственно внешней среды (свет, тепло, другие природные явления, социальное окружение, трудовая деятельность, образование, воспитательные меры и т. п.), общий ритм и режим жизни, качество питания, активность нервной системы, гормональные регуляторы клеток, внутриклеточные мессенджеры и модуляторы активности генов. В этом же ряду стоят все окружающие нас живые организмы, в том числе паразиты, микробы, вирусы. В результате клетки и сами организмы могут иметь разную скорость размножения и роста, разную продолжительность жизни, разную интенсивность синтеза белков и, следовательно, разное проявление всех жизненных функций. Например, даже у однояйцовых близнецов, с абсолютно одинаковым генотипом, со временем выявляются фенотипические различия (рост, мышечная сила, структура кожи, трудовые навыки и др.), если они выросли в разных условиях, допустим - в городе и деревне. Вся совокупность внешних относительно генотипа условий и регуляторов - от природно-климатических до нейрогормональных и внутриклеточных - и составляет сложную эпигенетическую программу развития
, поскольку все эти факторы существенно влияют на генную активность, стимулируя одни гены и подавляя другие.
Из сказанного следуют важные определения:
фенотип - это совокупность всех признаков и свойств организма, формирующихся в процессе взаимодействия его генотипа (генетической структуры) и внешней среды;
в фенотипе никогда не реализуются все генетические возможности;
в конкретных условиях развивается конкретный фенотип
.
Таким образом, в развитии фенотипа, то есть конкретного организма со всеми его индивидуальными свойствами, имеет место единство генетического и эпигенетического начал,
проявляющих себя на разных уровнях организации жизни -
молекулярно-генетическом, клеточном, организменном.
На вопрос: что первично, курица или яйцо? - должен последовать ответ: первично и конечно единство яйца (генотипа, программы) и курицы (фенотипа, сомы). Результатом этого единства является развивающийся организм.
Думается, что каждый сумеет сделать и некоторые практические выводы
в отношении своего собственного организма и развития (это никогда не поздно), но в особенности в отношении развития своих будущих детей, которое начинается задолго до их появления на свет и даже до их зачатия. В первую очередь это касается будущих матерей, так как внешнее управление развитием закладывается уже в цитоплазме яйцеклетки, еще даже не оплодотворенной. Нарушения этой закладки в результате болезней, неправильного питания, употребления алкоголя, токсических веществ, лекарств и т. п. чревато развитием у зародыша уродств, врожденных заболеваний, а то и полным бесплодием. Не исключение и представители сильного пола. Названные внешние факторы приводят к потере подвижности сперматозоидов (одна из наиболее распространенных причин мужского бесплодия), нарушению их производства или повреждениям ДНК (мутациям), которые неминуемо передадутся клеткам ребенка. Важное полезное заключение для будущих родителей касается также роли воспитания и обучения в развитии личности. Врожденные (генотипические) интеллектуальные, художественные наклонности и даже физические задатки не проявятся в полной мере, если соответствующие гены не будут востребованы. А востребованы они будут при постоянной нагрузке, которая и создается в процессе воспитания, обучения, трудовой деятельности.
ЗАКЛЮЧЕНИЕ ПО ТЕМЕ 3
Самовоспроизведение и индивидуальное развитие - неотъемлемое свойство и способ существования организмов. Это свойство составляет основную суть онтогенетического уровня организации живой материи.
Возвращаясь к истории вопроса о движущих силах развития (см. сегмент 21), можно видеть, что противоборствовавшие сторонники преформизма и эпигенеза отражали «две стороны одной медали». Индивидуальное развитие организма детерминировано («преформировано») на уровне генетической информации, в молекулах ДНК, в генах. Но развитие находится под многосторонним эпигенетическим контролем и управлением. Единство генетических и эпигенетических факторов развития можно трактовать как единство преформизма и эпигенеза.
Философ сказал бы и был бы прав, что развитием движет единство и борьба противоположностей
. Другой добавил бы, что развитие есть переход количества в качество
, имея в виду увеличение числа клеток, их дифференциацию и последующую интеграцию в единый организм с помощью нейрогуморальной системы. Третий заметил бы, что онтогенез совершается циклично, а точнее - по спирали
, так как в каждом новом витке индивидуального развития есть нечто новое по сравнению с предыдущим витком родителей. Это и комбинативные генетические изменения, связанные с половым процессом, и возможные новые мутации, полученные в прежней жизни. Об этих свойствах процесса индивидуального развития мы вспомним при рассмотрении механизмов биологической эволюции.
ТЕМА 4. КОНЦЕПЦИЯ САМОРЕГУЛЯЦИИ ЖИВЫХ СИСТЕМ
Сегмент 26. Саморегуляция и гомеостаз.
Сегмент 27. Внутриклеточная саморегуляция
Сегмент 28. Саморегуляция многоклеточного организма
Сегмент 29. Саморегуляция в экосистемах
Заключение по теме 4
СЕГМЕНТ 26. САМОРЕГУЛЯЦИЯ И ГОМЕОСТАЗ
Саморегуляция
в системе - это внутреннее регулирование процессов с подчинением их единому стабильному порядку. При этом даже в меняющихся условиях среды живая система сохраняет относительное внутреннее постоянство своего состава и свойств - гомеостаз
(от греческих homoios - подобный, одинаковый и stasis - состояние).
Действительно, окружающая среда очень переменчива. Изменяются температура, освещенность, влажность. Для животных, да и для растений не регулярна доступность пищи. Донимают паразиты, хищники и просто конкуренты за среду обитания. Тем не менее, животные и растения выносят эти колебания среды, живут, растут, размножаются. Экологические сообщества долгое время сохраняют некий средний состав.
Человек как высший представитель животного царства также поддерживает свой внутренний гомеостаз - благодаря работе многочисленных управляющих механизмов. Так, несмотря на смену дня и ночи, зимы и лета, температура нашего тела поддерживается на одном и том же уровне - около 37 градусов (под мышкой 36,6 градуса). Кровяное давление варьирует в ограниченных пределах, так как регулируется благодаря иннервации стенок сосудов. Солевой состав крови и межклеточных жидкостей, содержание сахаров и других осмотически активных веществ (способных вызвать нежелательное перераспределение воды между структурами организма) также поддерживаются на оптимальных уровнях. Даже простое и, казалось бы, самопроизвольное стояние на двух ногах требует ежесекундной согласованной работы вестибулярного аппарата и многих мышц тела.
Основоположник идеи о физиологическом гомеостазе Клод Бернар (вторая половина XIX века) рассматривал стабильность физико-химических условий во внутренней среде как основу свободы и независимости живых организмов в непрерывно меняющейся внешней среде.
Саморегуляция происходит на всех уровнях организации биологических систем - от молекулярно-генетического до биосферного (об уровнях организации см. тему 1). Поэтому проблема гомеостаза в биологии носит междисциплинарный характер. Внутриклеточный гомеостаз изучают цитология и молекулярная биология, организменный - физиология животных и физиология растений, экосистемный - экология. Конкретные проявления этих механизмов мы рассмотрим ниже. Здесь же отметим, что для поддержания гомеостаза во всех системах используются кибернетические принципы саморегулирующихся систем
. Кибернетика - наука об управлении - объясняет принцип саморегуляции системы на основе прямых и обратных связей между ее элементами. Вспомним, что система - это совокупность взаимодействующих элементов. Прямая связь между двумя элементами означает передачу информации от первого ко второму в одну сторону, обратная связь - передача ответной информации от второго элемента к первому. Суть в том, что информационный сигнал - прямой или обратный - изменяет состояние системы, принимающей сигнал. И тут принципиально важно, какой по знаку будет ответный сигнал - положительный или отрицательный. Соответственно и обратная связь будет положительной или отрицательной.
В случае обратной положительной
связи первый элемент сигнализирует второму о некоторых изменениях своего состояния, а в ответ получает команду на закрепление этого нового состояния и даже его дальнейшее изменение. Цикл за циклом первый элемент с помощью второго (контрольного) элемента накапливает одни и те же изменения, его состояние стабильно изменяется в одну сторону (рис. 18 а). Эта ситуация характеризуется как самоорганизация, развитие, эволюция, и ни о какой стабильности системы говорить не приходится. Это может быть любой рост (клетки, организма, популяции), изменение видового состава в сообществе организмов, изменение концентрации мутаций в генофонде популяции, ведущее через отбор к эволюции видов. Естественно, что обратные положительные связи не только не поддерживают, но, напротив, разрушают гомеостаз.
Рис. 18
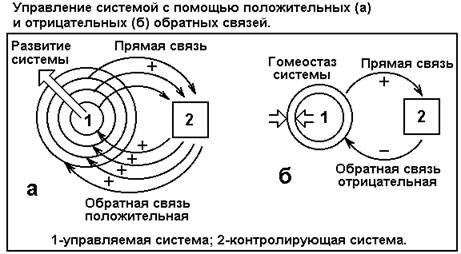
Обратная отрицательная
связь стимулирует изменения в регулируемой системе с противоположным знаком относительно тех первичных изменений, которые породили прямую связь. Первоначальные сдвиги параметров системы устраняются, и она приходит в исходное состояние. Цикличное сочетание прямых положительных и обратных отрицательных связей может быть, теоретически, бесконечно долгим, так как система колеблется около некоторого равновесного состояния (рис. 18б). Таким образом, для поддержания гомеостаза системы используется принцип отрицательной обратной связи
. Этот принцип широко применяется в автоматике. Так регулируется температура в утюге или холодильнике - с помощью термореле, уровень давления пара в автоклаве - с помощью выпускного клапана, положение судна, самолета, космического корабля в пространстве - с помощью гироскопов. В живых системах универсальный принцип обратной отрицательной связи работает во всех случаях, когда сохраняется гомеостаз.
Далее на конкретных примерах покажем саморегуляцию биологических систем разного уровня сложности.
СЕГМЕНТ 27. ВНУТРИКЛЕТОЧНАЯ САМОРЕГУЛЯЦИЯ
В клетке для поддержания гомеостаза используются в основном химические (молекулярные) механизмы регуляции. Наиболее важна регуляция генов
, от которых зависит производство белков, в том числе многочисленных и разнообразных ферментов.
Самая простая модель для демонстрации генного гомеостаза - регуляция выработки фермента для расщепления пищевого сахара у кишечной палочки. Эта бактерия является типичным гетеротрофом и поглощает из внешней среды несложные органические вещества, в том числе молочный сахар лактозу. Для расщепления и усвоения лактозы с определенного структурного гена, входящего в состав лактозного оперона (ген вместе с регуляторной областью) синтезируется информационная РНК и, далее, фермент (см. центральную догму молекулярной биологии - сегмент 22, рис. 13). Если сахар в среде отсутствует, фермент не вырабатывается, а при добавлении сахара активируется ген и идет синтез фермента. Но как только весь сахар будет клеткой использован, ген перестает работать. Как клетка узнает о присутствии сахара и его расходовании? Как оберегает свои гены от бесполезной работы и траты энергии? Регуляция генов у бактерий, как у всех прокариот, в целом организована гораздо проще, чем в эукариотных клетках. Оказывается, лактозный оперон у кишечной палочки работает по принципу отрицательной обратной связи, где в роли регуляторного «клапана» выступает особый участок оперона - оператор, а в роли регулятора сам пищевой субстрат - лактоза (рис. 19). Лактоза, поступившая в клетку, сама раскрывает структурный ген, используя для этого в качестве ключика операторный участок. Исчезновение лактозы автоматически приводит к закрытию гена.
Рис. 19
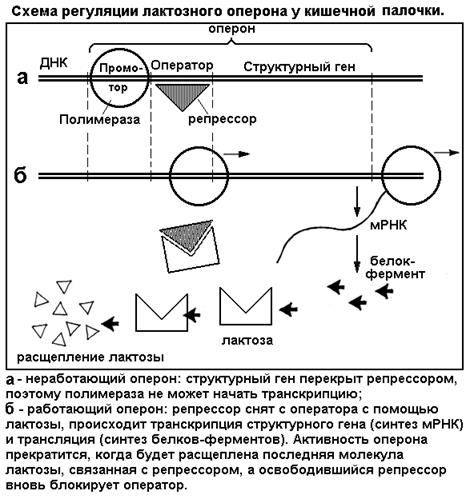
Лактозный оперон
- участок молекулы ДНК - состоит из трех частей: промотора, оператора и структурного гена. Промотор
- стартовый участок гена, сюда садится фермент РНК-полимераза, ведущий транскрипцию. Оператор -
пусковой барьер, в отсутствие лактозы закрытый специальным белком-репрессором. Структурный ген
(точнее - здесь находится цепочка, семейство генов) - основной участок ДНК, кодирующий и производящий через иРНК нужный белок-фермент. Пока оператор связан с белком-репрессором, полимераза не может стартовать и структурный ген не работает, синтез фермента отсутствует (см. рис. 19 а). Когда в клетку попадает лактоза, одна ее молекула связывается с репрессором и отнимает его от оператора. Теперь путь полимеразе открыт, идет синтез иРНК (транскрипция) и, далее, синтез соответствующего белка-фермента (трансляция) (рис. 19 б). Ферменты расщепляют поступивший в клетку сахар и в последнюю очередь ту его молекулу, которая связана с репрессором. Но когда будет переварена и эта последняя молекула, белок-репрессор освобождается и вновь блокирует оператор. Производство иРНК и фермента прекращается до поступления новой порции сахара. По своей простоте система регуляции гена концентрацией субстрата похожа на простые технические регуляторы. Напомним, однако, что у эукариот регуляция генной активности более сложная. Она включает возбуждение клеточных рецепторов гормонами или другими биологически активными веществами, запуск каскада реакций вторичных мессенджеров, которые поступают в ядро и избирательно активируют гены.
Другой пример простых саморегулирующихся систем, использующих обратную отрицательную связь, представляют ферментативные цепи, ингибируемые конечным продуктом
(рис. 20). Такие цепи обычно локализуются на поверхности внутриклеточных или наружных мембран и проводят комплексную переработку сложного субстрата в простой продукт. Суть регуляции состоит в том, что конечный продукт имеет стереохимическое сродство с первым ферментом. Связываясь с ферментом, продукт ингибирует (подавляет) его активность, так как полностью искажает его третичную структуру. Работает следующий регуляторный цикл. При повышении концентрации конечного продукта выше необходимого уровня его избыток ингибирует ферментную цепь (для этого достаточно остановить самый первый фермент). Ферментация прекращается, а свободный продукт расходуется на нужды клетки. Через некоторое время возникает дефицит продукта, блок с ферментов снимается, цепь активируется, и производство продукта снова растет.
Рис. 20
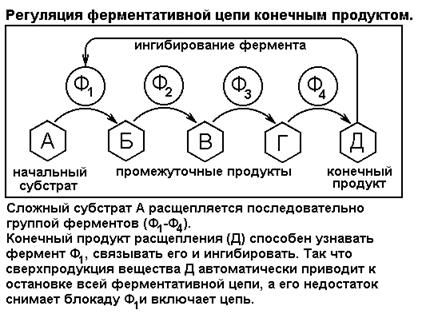
Третий пример- поддержание внутриклеточного осмотического гомеостаза
. В сегменте 19 мы говорили о механизме возникновения нервных импульсов и отмечали важную роль ионов натрия, концентрация которых снаружи клетки должна поддерживаться на более высоком уровне, чем внутри. Благодаря натриевым насосам, встроенным в мембрану клетки, удерживается нужный градиент ионов. Как только клетка получает избыток натрия, активируется натриевый насос (его фермент, расщепляющий АТФ и дающий энергию). Натрий выкачивается, его концентрация в клетке падает, что служит сигналом для отключения насоса.
Аналогично в клетках растений с помощью плазмалеммы (наружной мембраны) и вакуолей регулируется состав солей и питательных веществ. Плазмалемма обеспечивает приток в клетку необходимых ионов и воды из внешней среды и выделение балластных и избыточных ионов водорода, натрия, кальция. Мембрана вакуоли регулирует поступление в протоплазму запасных субстратов из вакуоли при их недостатке и удаление в вакуоль - при избытке.
Во всех рассмотренных случаях действует один и тот же принцип - саморегуляция системы на основе обратной отрицательной связи. Заметим, однако, что регулируемые параметры - концентрация солей, питательных веществ, конечного продукта ферментации или продукта генной активности - не бывают абсолютно постоянными, они поддерживаются в допустимых границах
. В каждом случае это свои физиологические границы, позволяющие нормально осуществлять клеточные функции. Аналогичный принцип мы увидим и на организменном уровне.
СЕГМЕНТ 28. САМОРЕГУЛЯЦИЯ МНОГОКЛЕТОЧНОГО ОРГАНИЗМА
Как мы только что видели, уже на клеточном уровне возникает необходимость поддержания специфических физико-химических условий, отличающихся от условий окружающей среды. У многоклеточных организмов появляется внутренняя среда, в которой находятся клетки различных органов и тканей, происходит усложнение и совершенствование механизмов гомеостаза
. В ходе эволюции формируются специализированные органы кровообращения, дыхания, пищеварения, выделения и др., участвующие в поддержании гомеостаза.
У морских беспозвоночных имеются механизмы стабилизации объема, ионного состава и рН жидкостей внутренней среды. Для животных, перешедших к жизни в пресных водах и на суше, а также у позвоночных, мигрировавших из пресных вод в море, сформированы механизмы осморегуляции, обеспечивающие постоянство концентрации солей внутри организма.
Наиболее совершенен гомеостаз у млекопитающих, что способствует расширению возможностей их приспособления к окружающей среде. В частности, обеспечивается постоянство объема крови и других внеклеточных жидкостей, концентрации в них ионов, осмотически активных веществ, постоянство рН крови, состава в ней белков, липидов и углеводов. У млекопитающих, а также у птиц, в узких пределах регулируется температура тела - их называют теплокровными животными.
Основную роль в поддержании гомеостаза организма играют нервная и гормональная системы регуляции
(см. рис. 17 г).
Наиболее важную интегрирующую функцию выполняет центральная нервная система
, особенно кора головного мозга. Большое значение имеет и вегетативная нервная система
, в частности ее симпатический отдел - система ганглиев (скоплений нервных клеток), расположенных по бокам позвоночника, в брыжейке и других частях тела (например, солнечное сплетение). Чувствительные нервные волокна охватывают сетью все внутренние органы, кровеносные сосуды, обеспечивая рефлекторную взаимосвязь между ними.
Гормональная регуляция
обеспечивается системой эндокринных желез
(от греческих endon - внутрь и krino - выделяю) - желез внутренней секреции. Центральная эндокринная железа - гипофиз - находится в голове и имеет прямую связь с головным мозгом (через посредство гипоталамуса), а ее гормоны через кровь воздействуют на все местные эндокринные железы- такие как щитовидная, паращитовидная, надпочечники, а также скопления эндокринных клеток в поджелудочной и слюнной железах, семенниках, яичниках, тимусе, плаценте и даже в сердце, желудке, кишечнике, почках. Выделяемые эндокринными железами гормоны с током крови (гуморально) распространяются ко всем органам-мишеням и участвуют в регуляции их роста (см. сегмент 25) и функционирования. Таким образом, фактически благодаря связи нервной и эндокринной систем осуществляется единая нейрогормональная саморегуляция организма.
В рамках данного пособия придется ограничиться лишь некоторыми примерами, демонстрирующими нейрогормональную регуляцию, без какой-либо претензии на комплексное освещение вопроса.
Примером сложной гомеостатической системы является система обеспечения оптимального артериального давления
. Изменение давления крови воспринимается барорецепторами сосудов - чувствительными нервными окончаниями, реагирующими на растяжение стенок сосудов при изменении внутреннего давления. Сигнал передается в сосудистые нервные центры, которые обратным сигналом изменяют тонус сосудов и сердечную деятельность. Одновременно включается система нейрогормональной регуляции и кровяное давление возвращается к норме.
Интересна и показательна регуляция пищевого поведения
у позвоночных животных и человека (рис. 21). В гипоталамусе - отделе головного мозга, ответственном за регуляцию вегетативных функций и связующем нервную и эндокринную системы (см. выше) - находятся центры голода и насыщения. В крови голодного животного (или человека) возникает недостаток глюкозы - простейшего сахара (углевода), который всасывается всеми клетками и расходуется для получения энергии (см. сегмент 13). Низкая концентрация глюкозы в крови приводит к раздражению центра голода. По нервным связям отдаются команды в мозг, на мышцы, и организуется поиск пищи. Когда пища найдена, включаются механизмы питания, пищеварения и всасывания продуктов в кровь. При этом белки перевариваются (расщепляются) до аминокислот, липиды до жирных кислот, а сложные углеводы до простых сахаров, в том числе глюкозы. Концентрация глюкозы в крови растет, что приводит к раздражению центра насыщения, далее к подавлению аппетита и прекращению питания. Когда глюкоза расходуется, ее концентрация в крови вновь понижается, отчего раздражается центр голода. Цикл повторяется. Поскольку гипоталамус связан и с нервными центрами, и со всей эндокринной системой, цикл пищевого поведения синхронизирован также с нервно-рефлекторной и гуморальной регуляцией желез пищеварительного тракта: выделяется слюна, желудочный сок, ферменты поджелудочной железы и кишечника, мобилизуется перистальтика.
Рис. 21
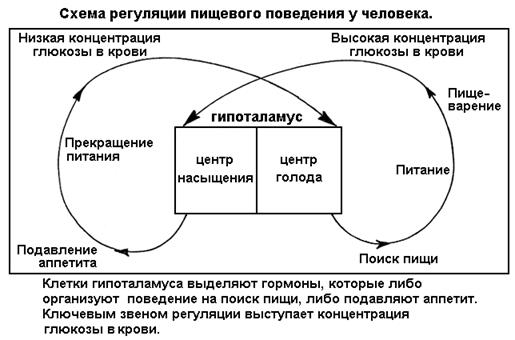
На основе процессов саморегуляции происходит морфологическая и функциональная гипертрофия органов в ответ на усиление нагрузки, на инфекцию, стрессовое воздействие
. В результате постоянных тренировок увеличиваются мышцы спортсмена, легкие ныряльщика. Увеличение нагрузки по прокачиванию крови ведет к гипертрофии сердца у тучного человека. Увеличивается и печень у больного человека. Характерная функциональная реакция развивается в ответ на гипоксию (недостаток кислорода): учащение пульса и увеличение числа эритроцитов, приводящие вместе к более быстрому обороту газов через организм. Или - реакция испуга, страха: выброс в кровь стрессового гормона адреналина ведет к повышению потребления кислорода, повышению концентрации глюкозы в крови, учащению пульса и мобилизации мышечной системы - все для мобилизации организма на оборону или избежание опасности. Другие системы при этом угнетаются - пропадают пищевые реакции, половые рефлексы и др. После исчезновения опасности все системы возвращаются в норму.
Механизм обратной отрицательной связи вовлечен в поддержание постоянства числа клеток в обновляющихся тканях
, таких как кровь, кишечный или кожный эпителий (рис. 22). В этих тканях имеется резерв недифференцированных клеток (например, красный костный мозг для крови), которые многократно делятся, дифференцируются, работают, стареют и отмирают. Считают, что зрелые клетки выделяют вещества, ингибирующие молодые делящиеся клетки. Выстраивается цепь взаимозависимых реакций: при избытке зрелых клеток продукция ингибитора высока и размножение клеток подавляется; уменьшение числа зрелых клеток в результате их естественной гибели сопровождается снижением концентрации ингибитора в среде; блок клеточных делений снимается; размножение молодых клеток усиливается; число зрелых клеток восстанавливается. Далее вновь возрастает продукция ингибитора и цикл повторяется. Общее число зрелых клеток в ткани колеблется около некоторого среднего уровня, резко не снижается и не повышается. По механизму передачи сигнала здесь мы имеем гуморальную систему, ингибитор работает как внутритканевой «гормон».
Рис. 22

К числу регуляторных систем, обеспечивающих внутреннее постоянство организма, кроме нервной и эндокринной, следует отнести иммунную систему
, которая отслеживает и поддерживает генетическую чистоту внутренней среды и тканей организма, устраняя проникшие вирусы, микробы или собственные мутантные клетки. Состав и принципы функционирования иммунной защиты были рассмотрены в сегменте 18. Теперь можно добавить, что сложный цикл выработки неспецифических и специфических защитных факторов (различных белков, в том числе антител), их взаимодействие с разнообразными чужеродными агентами (антигенами) и восстановление нормальной внутренней среды организма представляют звенья саморегулирующегося механизма. Это очень сложный, многокомпонентный механизм, в котором не сразу видны отдельные узлы саморегуляции, так как над ними или параллельно с ними работают другие управляющие механизмы.
Как и в случае с внутриклеточной регуляцией, мы должны заметить, что гомеостаз организма не бывает абсолютным.
Любые параметры: температура тела, артериальное давление, пищевое поведение, частота сердечных сокращений, число клеток в ткани и многие другие - находятся в колебательном режиме
. Это вытекает из самой природы механизма регуляции - прямая и обратная связи замкнуты в цикл, на оборот которого требуется определенное время. За это время регулируемая система успевает измениться в ту или иную сторону, что и выражается в колебании ее параметров. Но средний уровень параметра должен соответствовать норме, а коридор его колебаний не должен выходить за физиологические пределы. Если это все же происходит, говорят о патологических (болезненных) отклонениях в состоянии организма
.
Нормальные колебания функциональных характеристик организма происходят постоянно и называются
биоритмами
. Скорость синтеза белков в клетке колеблется в околочасовом (1,5 - 2 часа) ритме, большинство организменных ритмов имеют околосуточную периодичность, есть месячные, годичные и даже многолетние ритмы. Внутренний механизм, управляющий ритмами, принято называть биологическими часами, что подчеркивает связь биоритмов с астрономическим временем. Но заметим, что подавляющее большинство биоритмов являются наведенными, они сформированы под действием абиотических (небиологических) ритмов внешней среды. Это очевидно связанные с вращением Земли околосуточные ритмы, связанные с лунным циклом месячные ритмы и т. д. Поэтому биоритмы могут перестраиваться, и это происходит, например, когда мы перелетаем с востока на запад и наоборот. Но для этого требуется время, так как в один и тот же цикл (особенно суточный) бывают включены и жестко связаны друг с другом многие частные ритмы. И вообще колебательное состояние системы является наиболее устойчивым. Именно поэтому колебательное состояние внутренней среды организма выступает как важный фактор поддержания гомеостаза
.
СЕГМЕНТ 29. САМОРЕГУЛЯЦИЯ В ЭКОСИСТЕМАХ
Общее представление о структуре экологической системы было изложено при характеристике уровней организации жизни (тема 1). Напомним, что полноценная экосистема
представляет из себя биогеоценоз
- неразрывное единство биоценоза и биотопа. Биоценоз
- это сложное сообщество из популяций организмов разных видов - животных, растений, грибов, микроорганизмов, населяющих определенный ареал. При этом популяцией
обозначают совокупность особей одного вида, обитающих на данном ареале. Биотопом
называют всю совокупность факторов неживой среды, ареала, на котором обитает данный биоценоз.
Итак: биоценоз + биотоп = биогеоценоз (экосистема).
Прежде чем рассматривать механизмы саморегуляции в экосистемах, надо дать характеристику экологических факторов, без чего не возможно понять сути внутрисистемных экологических отношений.
Все условия среды, включая живые и неживые объекты, от которых зависит жизнь отдельного организма или популяции, обозначаются понятием экологические факторы
. Для конкретных популяций разные факторы могут быть необходимыми, вредными, безразличными (нейтральными). Экологические факторы делят на абиотические
и биотические
. Кроме того, в особую группу выделяют антропогенные
факторы, порожденные производственной деятельностью человека.
Абиотические факторы
- факторы неживой природы, в основном климатические. Сюда относятся свет, тепло («температура»), влажность, содержание в почве химических элементов, соленость морской воды, уровень радиации и др.
Биотические факторы
- это влияние одних живых организмов на другие. В зависимости от характера отношений различают несколько типов биотических факторов: конкуренцию, хищничество, паразитизм, симбиоз.
1. Конкуренция
- за среду обитания, пищу, свет, половых партнеров и другие условия. Конкуренция может быть внутривидовой - между особями одного вида и межвидовой - между особями разных видов животных или растений, обитающих на одном ареале и требующих одинаковых условий жизни. Конкуренция - обязательная форма отношений у рядом проживающих организмов и составляет одну из форм борьбы за существование.
2. Хищничество
- это способ добывания пищи и питания животных (редко - растений), при котором они ловят и поедают других животных. Жертва-хищник - одна из самых распространенных связей внутри сообщества. Внутривидовое хищничество известно как канибализм, распространено у хищных насекомых, пауков, рыб. Среди растений известны водная пузырчатка, болотные росянка, жирянка и другие, питающиеся насекомыми. Жертва захватывается быстрым смыканием листьев или лепестков, переваривается выделяемыми наружу ферментами и кислотами, и потом простые органические вещества всасываются клетками эпидермиса растения. Так пополняется недостаток азота в тканях растения.
3. Паразитизм
(от греческого parasitos - нахлебник) -
форма отношений, когда один организм (паразит) использует другого (хозяина) в качестве среды обитания и (или) источника пищи. Система паразит-хозяин представляет результат совместной эволюции двух видов (коэволюция) с сохранением равновесия, при выживании обоих. Паразиту не выгодно, чтобы хозяин погиб. Паразитизм распространен во всех царствах, типах, классах - как внутри этих таксонов, так и между ними. Вирусы, не имея своих метаболических систем, паразитируют в клетках бактерий, грибов, растений, животных. Паразитические бактерии поселяются в тканях грибов, растений, в кишечнике и других полостях животных (однако большинство бактерий ведет свободный образ жизни - в почве, воде, гниющих остатках). Грибы могут паразитировать на растениях и животных. Растения бывают паразитами растений же (повилика), а животные - животных. Среди животных наиболее известны паразитические черви (у человека - аскарида, острицы, цепень и др.) и насекомые (вши, клопы и др.).
4. Симбиоз
(от греческого symbiosis - совместная жизнь) - взаимовыгодное сожительство. Примерами симбиоза являются клубеньковые бактерии и бобовые растения (бактерии получают от растения пищу, растение от бактерии - усвоенный азот), грибы и корни растений, термиты и живущие в их кишечнике жгутиковые простейшие, переваривающие клетчатку. Аналогичные отношения имеются у человека с кишечной микрофлорой - разнообразными бактериями, переваривающими определенные компоненты пищи. Нарушение этого симбиоза - дисбактериоз - ведет к расстройствам кишечника, нарушениям пищеварения.
Любой экологический фактор имеет меру интенсивности
: тепловой фактор выражают температурой среды и измеряют в градусах Цельсия или Кельвина; освещенность измеряют в люксах или количестве световых часов в сутках, световых дней в году; интенсивность инвазии паразитами выражают в количестве паразитов на одну зараженную особь и т. д.
Для каждого организма, популяции существуют пределы выносливости того или иного экологического фактора
- от минимальной до максимальной интенсивности, в пределах которых организмы сохраняют жизнеспособность. Более узкую зону, в которой условия среды наиболее благоприятны, составляет оптимум экологического фактора
. В зависимости от природы фактора кривая степени его благоприятности имеет разный вид (рис. 23). Для таких факторов, как радиация или наличие паразитов зона оптимума начинается от нуля (то есть лучше, когда их нет совсем). Для влажности воздуха, освещенности, температуры и т. п. степень благоприятности фактора описывается кривой нормального распределения (колоколообразная кривая), отставленной от абсолютного нулевого значения. То есть влага, свет, тепло необходимы организмам в определенном диапазоне интенсивностей.
Рис. 23
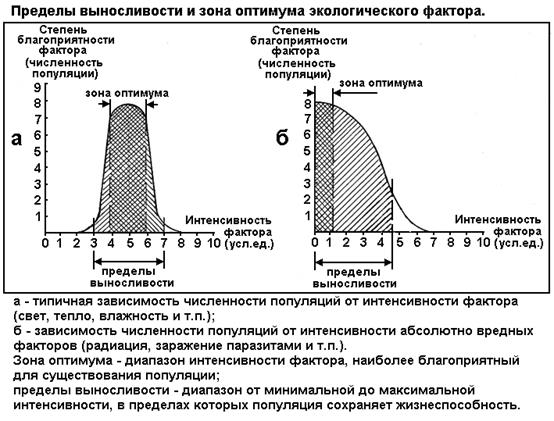 Каждый биологический вид по каждому экологическому фактору имеет свои пределы. Бурый медведь по многим факторам имеет широкие пределы выносливости: переносит большие колебания температуры, влажности, неприхотлив в выборе пищи - всеяден. Арктические рыбы, напротив, имеют очень узкие температурные пределы - от -2 до +2 градусов Цельсия. Среди растений есть светолюбивые, световыносливые и тенелюбивые. Каждый биологический вид по каждому экологическому фактору имеет свои пределы. Бурый медведь по многим факторам имеет широкие пределы выносливости: переносит большие колебания температуры, влажности, неприхотлив в выборе пищи - всеяден. Арктические рыбы, напротив, имеют очень узкие температурные пределы - от -2 до +2 градусов Цельсия. Среди растений есть светолюбивые, световыносливые и тенелюбивые.
Выяснив природу экологических факторов и характер их воздействия на организмы, можно перейти к рассмотрению сути вопроса - об экологической саморегуляции.
Находясь под действием самых разнообразных экологических факторов, хорошо сбалансированный по составу биоценоз тем не менее саморегулируется
и поддерживает внутреннее постоянство - гомеостаз
.
Состояние гомеостаза выражается в том, что:
организмы нормально размножаются
, поэтому:
численность различных популяций в сообществе поддерживается на определенных уровнях
, хотя и в колебательном режиме;
биоценоз
сохраняет устойчивость
и самовоспроизводится
даже при колебаниях климатических условий
.
Рассмотрим подробнее эти закономерности.
Элементарная саморегуляция
осуществляется на уровне отдельных популяций конкретных видов животных, растений, грибов, бактерий. Численность популяции зависит от противодействия двух начал:
репродуктивного потенциала популяции
и сопротивления среды
, между которыми устанавливаются прямая и обратная связи (рис. 24). Поясним это конкретным примером. Когда европейцы завезли в Австралию кроликов, последние, не встретив хищников, быстро расселялись по богатым растительностью территориям и их численность быстро возрастала. Этому способствовал высокий репродуктивный потенциал
(плодовитость) кроликов. Но вскоре пищи стало не хватать, возник голод, распространились болезни - численность кроликов пошла на убыль. Сработал фактор сопротивления среды
, который и выступил в качестве обратной отрицательной связи
. Пока популяция кроликов пребывала в угнетенном состоянии, среда (растительность) восстановилась и процесс пошел на новую волну. Через несколько циклов амплитуда колебаний численности кроликов сократилась, и установилась некоторая средняя плотность популяции. Этот показатель обозначают в экологии как поддерживающая емкость среды
.
Рис. 24
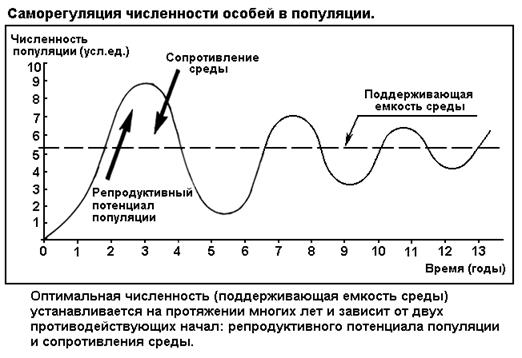
На самом деле в биоценозе все сложнее
, так как он состоит из нескольких взаимодействующих сообществ (зооценозы, фитоценозы, микробоценозы), а сообщества включают разные популяции конкретных видов. Все это взаимодействует на основе многочисленных прямых и обратных связей. Прежде всего важны трофические (пищевые) связи
, которые можно определить и как энергетические
, поскольку с пищей между организмами переносится энергия. По положению в пищевых отношениях все организмы делятся на три большие группы: продуценты, консументы и редуценты.
Продуценты
- первичные производители органических веществ (прежде всего глюкозы и аминокислот) из неорганических веществ неживой природы: Н2О, СО2, NН3. Это автотрофные организмы - растения и некоторые (хемосинтетические) бактерии, использующие энергию солнечного света и хемоэнергию для первичного синтеза глюкозы (см. сегмент 13). Таким образом, внешняя энергия фиксируется для собственных потребностей продуцентов и для дальнейшего использования животными. Значительная часть энергии выводится из оборота, так как сохраняется в ископаемых остатках растительного происхождения: каменном угле (минерализованная древесина), янтарях (застывшие растительные смолы).
Консументы
- потребители первичной продукции. Это животные организмы - гетеротрофы, в свою очередь выстроенные в пищевой ряд: травоядные (многие группы моллюсков, насекомых, рыб, птиц, копытные млекопитающие, грызуны), всеядные (есть в большинстве групп) и плотоядные - хищники (также имеются в разных группах беспозвоночных и позвоночных животных). Консументы поэтапно изменяют первичное органическое вещество и извлекают из него энергию. Часть этой энергии тратится на собственную жизнедеятельность, часть в виде тепла уходит во внешнюю среду, и третья часть сохраняется в мертвых остатках. Энергия, заключенная в минерализованных остатках (донные меловые отложения раковин фораминифер, моллюсков и других животных), фактически пропадает для дальнейшего использования, а энергия мягких тканей передается на следующий уровень.
Редуценты
- разрушители органического вещества. Сюда относятся многие бактерии, грибы, а из животных - некоторые черви (дождевой и др.), насекомые (термиты, навозники, личинки мух) и другие. Все они гетеротрофы, так как питаются органическим, хотя и мертвым веществом - отмершими растениями, животными и продуктами их выделений. Редуценты доводят распад биомассы до неорганических веществ: Н2
О, СО2
, NН3
и выделяют их во внешнюю среду - почву, воду, воздух. Перехваченная энергия используется редуцентами на их жизнедеятельность и в итоге рассеивается в разных оболочках биосферы - литосфере, гидросфере, атмосфере, а освобожденные неорганические вещества вновь поступают к продуцентам.
Таким образом, все организмы связаны переносом вещества и энергии
, через них и через внешнюю среду совершается глобальный круговорот материи
на Земле
. Основным донором энергии для поддержания этого круговорота выступает Солнце - его световая энергия обеспечивает фотосинтез глюкозы у растений. Пути передачи вещества и энергии через пищевые отношения организмов обозначаются как цепи питания, или пищевые цепи
. Эти цепи имеют одностороннюю направленность: от автотрофной биомассы продуцентов - в основном зеленых растений - к гетеротрофным консументам и далее к редуцентам. Значительная часть вещества возвращается в круговорот, но энергия, полученная от солнца, для живых организмов безвозвратно теряется, она либо аккумулируется в новых минералах почвы и донных осадков (каменные угли, мел и другие ископаемые), либо накапливается в виде тепла в оболочках Земли (разогрев атмосферы), либо рассеивается в Космос. Примеры пищевых цепей приведены на рис. 25 и 26.
Рис. 25
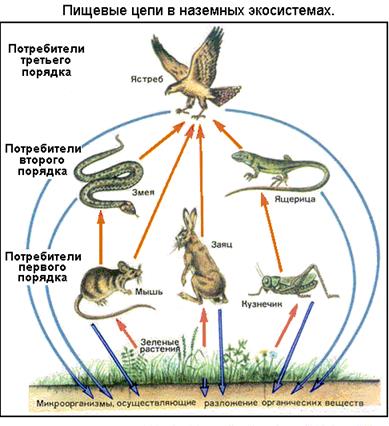
Рис. 26

Цепи питания имеют разную сложность, число звеньев в каждом из трех уровней может быть различным. Допустим, вариант короткой цепи: растения - заяц - волк - черви, бактерии. Длинная цепь: растения - травоядные насекомые (саранча, лесной клоп и др.) - хищные насекомые (жужелица, личинка стрекозы, водяной клоп и др.) - насекомоядные птицы (ласточки, мухоловки и др.) - хищные птицы (орел, коршун и др.) - черви, бактерии. Морская цепь: фитопланктон - мелкие ракообразные, черви - рыбы, питающиеся рачками и червями - хищные рыбы - хищные птицы ... В любой цепи возможны многочисленные ответвления и запасные пути. Если какой-то член выпадает, поток вещества идет по другим каналам. Допустим, выпадение личинок стрекоз компенсируется водными клопами - те и другие водные хищники. Если исчезает основной вид пищевой растительности, травоядные животные переходят на второстепенные корма. Если пропадают бабочки, ласточки ловят мух. От травоядных насекомых цепь может пойти вообще по другому пути: лягушка- цапля - лиса и т. д. Особенно большую путаницу в пищевые цепи вносят всеядные животные и, конечно, человек, так как они «встраиваются» в цепи в самых разных звеньях. Так что на самом деле существуют не цепи, а пищевые сети
- каждый трофический уровень образован многими видами.
Такое положение стабилизирует потоки вещества и энергии через живые сообщества, увеличивает устойчивость биоценозов
. Тем не менее общее направление трофического потока неизменное - продуценты - консументы нескольких порядков - редуценты.
Важно учитывать, что каждый трофический уровень передает на другой уровень энергию
, заключенную в макромолекулах организмов. Причем эта энергия составляет только часть
полученной от предыдущего уровня энергии, так как основная ее доля тратится на жизнедеятельность организмов данного уровня (биосинтезы, движение, транспорт ионов и др.), а так же теряется в виде тепла или построенных минералов. Эти же пропорции можно выразить и через понятие биомассы. Выстраивается так называемая экологическая пирамида
- при переходе с низших трофических уровней на более высокие количество внутренней (свободной, заключенной в организмах) энергии и общая биомасса организмов уменьшаются
. Подсчитано, что с уровня на уровень переходит около 10 % энергии, а от растительной массы до хищников и от них к редуцентам доходит всего 0,01 % энергии, полученной растениями от солнца. Наглядный пример пищевой пирамиды, построенной по численности обитателей на 1 гектаре земли приведен ниже:
3 птицы
350 тысяч хищных насекомых
700 тысяч растительноядных насекомых
9 миллионов растений.
Пищевая пирамида экосистемы, как отражение ее структуры, сохраняет саморегуляцию и устойчивое развитие
. Характерная пропорция разных обитателей биоценоза устанавливается сама по себе, в результате процессов саморегуляции и отражает в целом поддерживающую емкость среды. Приведенные цифры являются средними, но реально во всех популяциях происходит колебание численности особей
, причем колебания на низшем уровне неизменно ведут к таким же колебаниям на следующем уровне, а в целом система поддерживает равновесное состояние.
Рис. 27
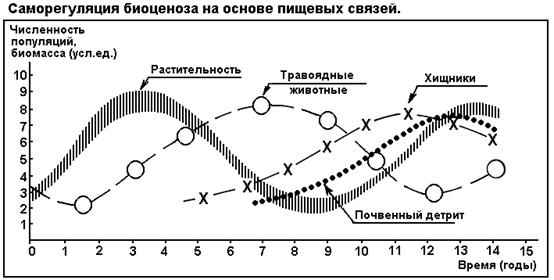
На рис. 27 приведен пример саморегулирующегося биоценоза
из 4 трофических уровней. Проследим, как проявляются прямые и обратные связи в динамике численности организмов разных трофических уровней. В зависимости от колебаний погодно-климатических условий (солнечная активность, количество осадков и др.) год от года варьирует урожай кормовых растений - продуцентов. Вслед за ростом зеленой биомассы увеличивается численность травоядных животных - консументов первого порядка (прямая положительная связь), но уже на следующий год это отрицательно скажется на урожае растений, так как большинство из них не успеет дать семян, поскольку будет съедено (обратная отрицательная связь). В свою очередь увеличение числа травоядных создаст условия для хорошего питания и размножения хищников - консументов второго порядка, их численность начнет возрастать (прямая положительная связь). Но следом пойдет на убыль численность травоядных (обратная отрицательная связь). К этому времени в почве успеет разложиться до минеральных веществ травяной опад от первой волны урожая и экскременты травоядных животных и, в меньшей степени, хищников, что создаст благоприятные условия для роста растений. Начнется вторая волна урожая, и цикл повторится. Год от года численность особей разных трофических уровней будет варьировать, но в среднем на протяжении многих лет биоценоз будет сохранять устойчивое состояние. Это и есть гомеостаз.
Как отмечено в начале, биоценоз должен не просто саморегулироваться (судя по приведенной схеме, это не так уж и сложно), но он должен иметь устойчивость к изменениям внешних (абиотических, погодно-климатических) факторов
, так сказать - запас прочности на случай неблагоприятных условий среды. Поддержанию высокой устойчивости биоценоза будет способствовать ряд условий
:
высокий, но сбалансированный репродуктивный потенциал
отдельных популяций - на случай массовой гибели особей;
адаптации
(приспособления) отдельных видов к переживанию неблагоприятных условий;
разнообразие
сообществ и разветвленные пищевые сети - исчезнувший объект должен заменяться другим, в норме второстепенным.
Таким образом, устойчивость экосистемы, ее саморегуляция, или гомеостаз, проявляется в ее самовоспроизведении
, саморегуляции численности и устойчивости к экстремальным факторам среды
.
Но устойчивость экосистемы относительна, не беспредельна
. Она нарушается в основном в двух случаях:
при сильных изменениях
внешней среды
- пожары, наводнения, продолжительные засухи, оледенения и другие природные катаклизмы;
при резких изменениях
состава сообществ
- обычно человеком, например, в результате массового отстрела хищников, заселения новых видов, как было с кроликами в Австралии, вырубки лесов, распахивания степей и т. д.
При этом происходит смена экосистем
, их переход в новое качество
, что означает новый цикл развития
в направлении повышения устойчивости
. Идет восстановление биоценоза, но уже с новыми сообществами организмов, с новыми прямыми и обратными связями. Этот процесс смены экосистемы и ее развития к новому состоянию устойчивости происходит поэтапно и очень медленно и обозначается понятием сукцессия
(от латинского successio - преемственность, наследование). Особо подчеркнем, что сукцессия представляет уже не саморегуляцию
, а ее противоположность - самоорганизацию, развитие
, так как при перестройке системы преобладают обратные положительные связи, вместо обратных отрицательных, и изменяются ее количественные и качественные характеристики
.
Различают первичную и вторичную сукцессии.
Первичная сукцессия
- самоорганизация экосистемы на свободном первичном субстрате: скальная порода, образовавшаяся в ходе геологических разломов земной коры; дно высохшего водоема, например, Аральского или Каспийского морей; пустыня, образовавшаяся после отступления ледника; пустой карьер после выработки полезных ископаемых и т. д. Ниже приведена типичная динамика первичной сукцессии
.
Первичный субстрат (скала, песок, ил, щебень или др.);
выветривание, эрозия породы (разрушение под действием ветра, воды, перепада температур);
заселение бактериями, водорослями, микроскопическими грибами, что ведет к фиксации азота и формированию почвы;
поселение мхов и лишайников, увеличение слоя почвы;
поселение трав, формирование лугов и степей; одновременное заселение мелких животных - червей, насекомых, грызунов; вытеснение мхов и лишайников в результате конкуренции;
поселение кустарников, увеличение разнообразия животных (появление птиц, крупных млекопитающих);
поселение деревьев, формирование лесного многоярусного сообщества с разветвленной пищевой сетью.
Это конец развития
- сформирована новая устойчивая саморегулирующаяся экосистема, с новым экологическим гомеостазом
. Такое законченное, сбалансированное сообщество животных, растений, грибов, микроорганизмов называется климаксным сообществом
. Однако конечный состав экосистемы зависит от географической широты, климата. Устойчивой может стать и тундра, саванна, даже пустыня - каждая со своим набором приспособленных к этим условиям организмов. На формирование первичной сукцессии обычно уходит несколько тысяч лет.
Вторичная сукцессия
- процесс восстановления экосистемы после повреждений, причиненных внешним воздействием: после бури, пожара, вырубки леса, выпаса скота и т. п. Вторичные сукцессии обычно бывают неполные, упрощенные, обедненные видовым составом сообществ. Роль вторичных сукцессий возрастает с увеличение человеческого населения Земли, особенно с развитием городов, промышленного производства и интенсивного земледелия. Влияние человека на состояние экосистем стало сегодня решающим. Вторичные сукцессии развиваются в течение нескольких лет или десятилетий.
Таким образом, несмотря на саморегуляцию в экологических системах, природа закономерно и необратимо изменяется
. Это естественный биогеохимический процесс, идущий независимо от воли и деятельности человека. Когда он протекает без резких отклонений, говорят об устойчивом развитии
экосистем и в целом биосферы Земли. В этом определении отражено единство противоположностей: устойчивость, гомеостаз, с одной стороны, и развитие, необратимое изменение - с другой. Нарушение устойчивого развития означает наступление экологического кризиса
, чреватого таким изменением биосферы, которое станет несовместимо с жизнью человечества. Основной причиной необратимых изменений биосферы в XX столетии стала хозяйственная деятельность человека. Задача науки экологии - понять движущие силы устойчивого развития, спрогнозировать экологический кризис, а задача общества - воспринять новое экологическое мышление и принять своевременные меры к недопущению кризисных состояний.
ЗАКЛЮЧЕНИЕ ПО ТЕМЕ 4
Саморегуляция и поддержание устойчивого состояния - гомеостаз
- обязательное свойство живых систем, не зависимо от уровня их сложности. Регулируется и поддерживается относительное постоянство физико-химических параметров клетки. Сохраняется в пределах физиологической нормы состояние тканей и органов многоклеточного организма. Воспроизводится состав живых сообществ в биоценозах. В основе поддержания гомеостаза лежит универсальный принцип обратной отрицательной
связи.
В то же время живые системы направленно и необратимо изменяются, самоорганизуются, что составляет сущность их развития
. Клетки дифференцируются, работают и умирают. Организмы растут, размножаются, стареют и умирают. Биоценозы подвергаются сукцессиям и так же необратимо изменяются с изменением климата на Земле. Направленное изменение биосистемы по сути противоположно гомеостазу, оно происходит на основе обратных положительных
связей.
Устойчивость, неизменность биосистем, с одной стороны, и их постепенное изменение, развитие - с другой - представляют диалектическое единство противоположностей, что и выражается понятием устойчивое развитие
. При естественном и сбалансированном течении этих процессов клетки нормально функционируют на протяжении всей жизни организма, человек в полном здравии и уме доживает до 100-150 лет, биосфера Земли сохраняет перспективу жизнеспособности на миллионы лет. Однако разум человека, ставший частью биосферы и превративший ее в качественно новое образование - ноосферу, способен в короткое время разрушить процесс устойчивого развития. Мы проделываем это со своим собственным организмом, когда курим, злоупотребляем алкоголем, принимаем наркотики, не предостерегаемся от инфекций, много работаем и мало отдыхаем - в итоге сокращаем свою жизнь вдвое. Мы напрягаем до предела и вот-вот сломаем равновесие своей экологической системы. Отравленные воды, непригодные для питья; озоновые дыры в атмосфере, несущие губительные ультрафиолетовые лучи; парниковый разогрев земной поверхности от избытка в атмосфере углекислого газа и быстрое таяние полярных льдов; накопленные арсеналы ядерного, химического и биологического оружия, способные многократно уничтожить все живое на земле ... - вот далеко не полный перечень итогов «разумной» деятельности человека в своем общем доме. Сумеет ли человечество осознать происходящее и принять меры прежде, чем эти изменения войдут в кризисную стадию? Есть ли еще шанс сохранить устойчивое развитие биосферы или мы доживаем последнее столетие? Увы, но сейчас никто не возьмет на себя смелость дать положительные ответы на эти вопросы.
ТЕМА 5. КОНЦЕПЦИЯ САМООРГАНИЗАЦИИ И БИОЛОГИЧЕСКОЙ ЭВОЛЮЦИИ. ФИЛОГЕНЕЗ
Сегмент 30. Проблема самоорганизации и наука синергетика
Сегмент 31. Возникновение жизни на земле
Сегмент 32. Этапы развития жизни на земле и современное биоразнообразие
Сегмент 33. Факторы биологической эволюции
Сегмент 34. Происхождение и эволюция человека
Заключение к теме 5
СЕГМЕНТ 30. ПРОБЛЕМА САМООРГАНИЗАЦИИ И НАУКА СИНЕРГЕТИКА
В предыдущей теме, рассматривая закономерности и механизмы саморегуляции
живых систем, мы вынуждены были затронуть и проблему самоорганизации
. Несмотря на созвучность терминов и их кажущуюся однозначность, на самом деле они выражают альтернативные понятия. Как мы только что показали на примере экологических систем, саморегуляция означает поддержание стабильного состояния системы, ее гомеостаз на основе обратных отрицательных связей, тогда как самоорганизация - это необратимое изменение, развитие системы на основе обратных положительных связей. В соответствии с законами диалектики эти две противоположности взаимодействуют, дополняют друг друга, так что вместе обеспечивают процесс
устойчивого развития
биосистем
.
В этой теме мы должны более основательно рассмотреть вопросы происхождения и исторического развития жизни на Земле
, эволюцию живых форм, причины и движущие силы этих глобальных процессов. Ключевым понятием в проблеме эволюции сегодня выступает
понятие самоорганизации
как основы любого процесса развития
. В кругу этих проблем на стыке интересов физики, химии, биологии, а также социологии и философии во второй половине XX века возникла новая наука синергетика
(от греческого synergos - совместно действующий) - наука о самоорганизации физических, биологических и социальных систем.
До недавнего времени проблема эволюции жизни оставалась чисто биологической, так как еще в XIX веке эволюция в неживых системах понималась физиками иначе, чем в биологии. Обращаясь с системами закрытого типа, теплофизика считала, что их самопроизвольное изменение, то есть эволюция, протекает путем дезорганизации и разрушения систем. При этом доля свободной энергии, способной к совершению работы, в системе убывает, а энтропия системы - деградированная, отработанная энергия - растет и стремится к максимальному значению. Этот закон был сформулирован как второе начало термодинамики
, о чем мы уже говорили в сегменте 12. Однако оказалось, что реальные системы в природе являются открытыми
. Это означает, что они обмениваются с внешней средой веществом, энергией и информацией. При поглощении внешней энергии в них возникают процессы самоорганизации, усложнения материи, но при этом происходит диссипация (рассеяние) использованной энергии, которая становится непригодной к производству работы
. Можно сказать, что открытая развивающаяся система производит энтропию, но не накапливает ее, а рассеивает во внешнюю среду
. Таким образом, интерес ученых сместился к изучению открытых диссипативных систем и принципов их взаимодействия с внешней средой, так как в этом взаимодействии и виделся ключ к пониманию универсальных законов эволюции.
Диссипативные системы
- способные к поглощению и диссипации энергии и поддерживающие за этот счет свою собственную структуру и самоорганизацию
- существуют на разный уровнях организации материи. Мы уже видели это на примере жизнедеятельности элементарной живой системы - клетки (см. сегмент 12 и рис. 4). За счет солнечной энергии или энергии экзотермических химических реакций клетка строит из простых неорганических веществ сложные органические вещества, поддерживает свою целостность и развитие, тем самым противодействуя росту энтропии.
Оказалось, что диссипативные процессы самоорганизации происходят и в неживой природе
. В 60-70-е годы XX века физиками открыты кооперативные резонансные процессы элементарных частиц в лазере, происходящие под действием внешнего света, а в химии открыты колебательные реакции, идущие по принципу «химических часов». Причем движущей силой самоорганизующихся реакций, пружиной химических часов может выступать такая незаметная на первый взгляд сила, как гравитационное поле Земли. Колебательная химическая система, названная брюсселятором, изучена отечественными учеными радиохимиком Б. П. Белоусовым и биофизиком А. М. Жаботинским. При свободном поступлении в такую систему химических субстратов и при наличии в ней катализаторов происходит реакция, продукты которой удаляются, освобождая место для поступления новой порции субстрата. Реакция идет по замкнутому циклу и в результате изменения концентрации реагирующих веществ сопровождается образованием характерных пространственных структур - в виде расходящихся колец на реакционной поверхности. Создается впечатление пульсирующей, «живущей» химической системы.
Теоретическое объяснение и математическую модель процессов самоорганизации диссипативных структур предложил бельгийский физико-химик И. Р. Пригожин, получивший в 1977 г. за эту работу Нобелевскую премию. Назовем основные положения синергетики
, объясняющие механизм самоорганизующихся процессов. С некоторыми из них мы уже хорошо знакомы.
1. Самоорганизующаяся система должна быть открытой
- доступной для обмена веществом, энергией и информацией с внешней средой.
2. Система должна быть неравновесной
, то есть находиться достаточно далеко от точки термодинамического равновесия (точка дезорганизации с максимальной энтропией), так как вблизи этой точки наступает необратимое скатывание к равновесному состоянию.
3. Образование нового порядка через флуктуации
. В системе всегда возникают флуктуации - случайные отклонения от среднего положения. По законам саморегуляции они устраняются, но при достаточной неравновесности системы за счет свободной энергии отклонения усиливаются, наступает момент бифуркации
- переломная точка в развитии системы, за которой возможно устойчивое отклонение от прежнего состояния. Прежний порядок исчезает, возникает и закрепляется новый порядок элементов в системе.
4. Самоорганизация ведет к новому порядку согласно принципу
обратной положительной связи
, по которому отклонения в системе не устраняются, а напротив, закрепляются и усиливаются.
5. Самоорганизация ведет к нарушению симметрии
- структура и
свойства системы до и после точки бифуркации не симметричны, то есть различаются в следствие необратимости процессов развития.
6. Самоорганизация возможна при некотором критическом количестве элементов в системе
, достаточном для возникновения их кооперативного поведения. Путь к новому качеству возможен через изменение количества.
Разумеется, здесь приведены лишь самые необходимые условия самоорганизации. В зависимости от уровня сложности развивающихся систем, могут появляться и другие, частные факторы, необходимые и достаточные для полноценной эволюции. Наша дальнейшая задача - найти эти факторы в процессах исторического развития жизни на Земле. И первый вопрос - о происхождении жизни как таковой.
СЕГМЕНТ 31. ВОЗНИКНОВЕНИЕ ЖИЗНИ НА ЗЕМЛЕ
От античных времен до средних веков многие философы и ученые считали, что живые организмы могут возникать из неживой материи. Но эта принципиально важная мысль не получала доказательств. Представления о происхождении лягушек и рыб из ила, а мух из гнилого мяса были опровергнуты в 1661 г. итальянским врачом Франческо Реди, который в простых опытах показал, что мухи возникают не из мяса, а из яиц, отложенных в мясо другими мухами. Через 200 лет выдающийся французский микробиолог Луи Пастер поставил точку в спорах о самозарождении жизни, доказав, что даже микробы - мельчайшие свободноживущие клетки - не могут возникать из мертвой, прокипяченной питательной среды. При этом Пастер убедил даже виталистов, которые считали, что для возникновения жизни нужна особая «жизненная сила». Он придумал знаменитую колбу с длинным S-образным горлышком, в которой после кипячения бульона, несмотря на то, что горлышко оставалось открытым и доступным для «жизненной силы», микробы тем не менее не заводились. В 1862 г. Пастер по этому поводу выиграл специальную премию Французской академии наук, объявленную тому, кто разрешит проблему самозарождения жизни. Восторжествовал закон: все живое - от живого. Но значило ли это, что жизнь не могла зародиться на Земле из неживого субстрата в доисторические времена, когда еще не было никаких организмов, да и состав земной поверхности, вероятно, был другим, нежели в современную эпоху? Тогда откуда и как появились первые живые существа, хотя бы те же бактерии? Убедительного ответа на этот вопрос нет до сих пор, хотя имеются более или менее обоснованные гипотезы.
Так называемая «гипотеза» креационизма
- о сотворении жизни сверхъестественным, божественным существом - не имеет никаких доказательств. Она основана на вере и является по сути не научной, а религиозной идеей, поэтому нами рассматриваться не будет.
Более интересна гипотеза вечности жизни
, которая в представлениях шведского физико-химика конца XIX века Аррениуса известна как теория панспермии
(от греческих pan - всеохватывающий и sperma - семя). Согласно этой гипотезе жизнь всегда присутствует в космосе в виде мельчайших спор микроорганизмов, которые мигрируют между планетами, галактиками в составе комет, метеоритов и других космических тел, пересекающих большие пространства. Когда-то жизнь попала и на Землю, а потом развивалась, совершенствовалась. Выполнен ряд работ с метеоритными телами, в которых найдены органические вещества (аминокислоты, спирты, углеводороды), а также мельчайшие замурованные пузырьки, похожие на отпечатки клеток или спор. В некоторых метеоритах, упавших на Землю в 40-50-х годах и недавно извлеченных, найдены даже бактерии. Кроме того, астрофизики на основании спектральных анализов удаленных космических тел утверждают, что и в космосе присутствуют органические вещества; особенно много их в кометах. Однако, все эти наблюдения не являются доказательствами присутствия жизни. Органическое вещество - соединения на основе углерода - вполне может быть неживого происхождения, пузырьки в метеоритах скорее всего были заполнены газами, а встречаемые в метеоритных останках бактерии, очевидно, проросли за десятилетия из почвы. В космосе ни вирусы, ни бактерии пока что не обнаружены. К таким выводам сходится большинство ученых, озабоченных проблемой происхождения жизни.
Для полноты картины добавим, что существует еще теория направленной панспермии
, согласно которой жизнь в виде простейших организмов на Землю занесли инопланетяне, опередившие нас по уровню развития на миллиарды лет. Эта идея подогревается наблюдениями различных НЛО, спекуляциями по поводу падающих «летающих тарелок» и захваченных, но утаиваемых от общественности, инопланетян. Оставим эти домыслы журналистам и досужим обывателям, поскольку в научной среде факты о посещении Земли какими-либо существами отсутствуют.
Даже если жизнь «вечна» и на Землю попала из космоса, остается вопрос о происхождении первых организмов - где бы и когда бы они не возникли. Поскольку современные космологические теории, в частности теория большого взрыва, выводят Вселенную из материального хаоса, ни о каком присутствии в Космосе живых организмов до появления Вселенной не может быть и речи. Поэтому обратимся к тем гипотезам, которые объясняют абиогенное возникновение жизни
из первичных неорганических субстратов, причем ни где-нибудь в космических далях, о которых мы мало что знаем, а у нас на Земле.
Исходя из макромолекулярной специфики живой материи - как помним, ее основу составляют белки и нуклеиновые кислоты - любая теория абиогенного (не из живого) возникновения жизни в первую очередь должна объяснить происхождение этих сложных нерегулярных полимеров и, более того, предложить механизм возникновения генетического кодирования первичной структуры белков через структуру ДНК и РНК (см. центральную догму молекулярной биологии - сегмент 22). Кроме того, надо понять как возникли липидные мембраны и тогда несложно прийти к формированию протоклеток - простейших комочков живой плазмы, ограниченных мембраной. В современной биологии сложилось два подхода к объяснению этих механизмов и соответственно две модели абиогенного происхождения жизни: бульонно-коацерватная и твердоматричная модели.
Бульонно-коацерватная модель
исходит из того, что сложные органические вещества возникли в растворах (в «бульоне»), из которых формировались коацерваты - прямые предшественники протоклеток.
Уже Ламарк в 1802 г. высказал идею о самопроизвольном зарождении живого под действием «флюидов» - теплоты и электричества. Позднее Чарльз Дарвин в одном из частных писем высказывал мысль о самозарождении жизни на Земле в каком-нибудь «маленьком теплом водоеме». Он допускал, что жизнь возникла из молекул химических веществ, которые под действием света, тепла и электричества взаимодействовали, давая сложные соединения. При этом Дарвин замечал, что такое было возможно только в эпоху первобытной Земли, так как в современных условиях всякий новый организм становился бы жертвой конкуренции или хищничества со стороны уже существующих организмов.
В 20-е годы XX века гипотеза возникновения жизни на основе химической эволюции была детально разработана российским академиком А. О. Опариным и, независимо от него, американцем Дж. Холденом, а позднее получила некоторые экспериментальные подтверждения. В современном толковании, согласно этой гипотезе, жизнь возникла из неорганических веществ в несколько этапов, причем химическая эволюция перешла в биологическую эволюцию. Как же это могло быть?
Возраст Земли определяют в 5-7 млрд лет. В этот ранний период наша планета представляла раскаленное газо-пылевое облако. Около 4 млрд лет назад образовалась кора. Примерно 3,6 млрд лет назад уже возникла жизнь. По геологическим данным первые организмы - бактерии и сине-зеленые водоросли - населяли воды мирового океана: моря, лагуны, ванны, гидротермы (места выхода горячих газов). Но появлению микроорганизмов предшествовала длительная химическая эволюция, в ходе которой на первом этапе
из неорганических веществ синтезировались органические биополимеры
.
По Опарину 4 млрд лет назад атмосфера земли состояла из аммиака (NH3
), метана (CH4
), углекислого газа (CO2
) и паров воды (H2
O). По современным данным аммиак и метан, возможно, отсутствовали, но не исключается наличие водорода (Н2
), хлора (Cl2
), азота (N2
), сероводорода (H2
S). Газы вырывались из остывающего центра земного «облака». Кислород не доходил до атмосферы, окисляя по пути различные вещества. Когда температура поверхности упала ниже 100 градусов, начался период горячих дождей, сформировался мировой океан, многочисленные моря и мелкие водоемы. Начался первый этап
возникновения жизни - синтез органических веществ
. В горячей воде хорошо растворялись атмосферные газы, а также газы, минеральные соли и другие вещества, приносимые горячими источниками и вулканами. Атмосфера и поверхность земли подвергались действию космического излучения, включавшего ультрафиолетовый свет от солнца, радиоактивные частицы, гамма-излучение. Были частые и сильные грозы, дававшие мощное электромагнитное излучение. В этих условиях из газов в воде возникали простые органические соединения: HCN, CH3
-CH3
, CH2
=CH2
, CH2
=O (формальдегид), а из них и более сложные: полимерные углеводороды типа СН3
-СН2
-СН2
-...-СН3
, карбоновые кислоты СН3
-СООН, СН3
-СН2
-СН2
-...-СООН. Путем присоединения к этим веществам азота в форме аминогрупп (-NH2
) легко могли сформироваться аминокислоты: NH2
-CH®-COOH, где R - изменчивый радикал, а группы -NH2
и -СООН представляют боковые «ручки» для полимеризации аминокислот в белок. Аналогично можно вывести из простых органических предшественников мочевину, глицерин, липиды, углеводы, азотистые основания и, далее, через полимеризацию нуклеиновые кислоты - ДНК и РНК.
В 1953 г. молодой американский аспирант С. Миллер сумел воспроизвести в колбе искусственный абиогенный синтез органических веществ. Из метана, аммиака, водорода и воды при температуре 80 градусов Цельсия, при высоком давлении и при пропускании электрических разрядов напряженностью 60 тысяч вольт он получил жирные кислоты, мочевину, уксусную кислоту и, самое главное, прямые предшественники белка - аминокислоты (глицин и аланин - 2 аминокислоты из 20, входящих в состав белков). Позднее американец С. Фокс при нагревании смеси аминокислот синтезировал и сам белок - в виде простейшего полипептида. Были получены также и нуклеотиды - предшественники ДНК и РНК.
Таким образом, теория Опарина-Холдена совершала триумфальное шествие в лабораториях биохимиков середины XX века. Однако появилась и альтернативная версия о том, что органические вещества могли приноситься на Землю в составе комет и метеоритов. Но как бы там ни было, от органических веществ до простейшей клетки - дистанция большого размера. Как представлял себе Опарин, на втором этапе
химической эволюции происходило концентрирование органических веществ в плотных капельках - коацерватах
. В присутствии электролитов (растворенных солей) органические вещества отделяются от общего раствора в виде геля - более концентрированного раствора с гидрофобными, водородными и другими дополнительными связями между молекулами. Эти капли - коацерваты - получали искусственно. Некоторые их свойства были сравнимы с функциями живых клеток.
Искусственные коацерваты могли поглощать из окружающего раствора различные вещества, имитируя питание и рост
. Эти вещества преобразовывались в результате химических реакций, а продукты выделялись во внешнюю среду. Таким образом, происходил примитивный обмен веществ
. При встряхивании коацерваты делились
, а потом снова росли путем самосборки
. Опарин даже считал, что между коацерватами идет борьба за существование,
и «выживают» более устойчивые, приспособленные к данной среде. Эту же мысль с позиций теории самоорганизации позднее развивал немецкий ученый М. Эйген. Он также считал, что происхождение жизни есть результат отбора и адаптаций на уровне органических макромолекул при том, что молекулярная система открыта для обмена веществом и энергией, способна к автокатализу и мутациям.
Но даже сложные коацерваты, если они действительно формировались в первобытных морях, еще не представляли живые системы; это могла быть лишь стадия преджизни. Необходим третий этап -
возникновение механизма генетического кодирования и воспроизведения жизни
. Классическая версия предполагает возникновение и включение в коацерваты нуклеиновых кислот и белков-ферментов. Причем некоторые ученые считают, что первой возникла РНК (сейчас доказано, что она способна к ауторепликации без ферментов), а уже потом появилась способность к синтезу белков с матрицы РНК и еще позже - с ДНК. Под действием излучений в нуклеиновых кислотах накапливались ошибки репликации, то есть мутации, которые вели с появлению молекулярного разнообразия. С появлением в среде липидов возникли мембраны (как образуются тончайшие масляные пленки на поверхности воды) и появилась возможность более надежной изоляции коацерватных капель. Это надмолекулярное образование уже можно было бы назвать простейшей клеткой.
Однако бульонно-коацерватная модель возникновения жизни имеет и большие трудности
. Все рассуждения о предполагаемых событиях третьего этапа остаются спекулятивными, к ним практически нет реальных оснований. Причем они не объясняют самого главного - как возник генетический код, триплетное соответствие первичных структур нуклеиновых кислот, с одной стороны, и белков - с другой
. А ведь именно это соответствие представляет суть центральной догмы молекулярной биологии. Трудный вопрос: откуда взялся для построения ДНК фосфор - редкий элемент земной коры. В 1990 г. синтезировано органическое соединение AATE (AminoAdenosinTriacidEster), состоящее из двух частей - со свойствами белка и нуклеиновой кислоты. Вещество оказалось способно к аутокаталитическому воспроизведению (в хлороформе). Но выход на гены и белки отсюда тоже не виден. Попытки объяснить происхождение кодированного соответствия ДНК (РНК) - белок содержатся в другой модели абиогенного возникновения жизни.
Твердоматричная модель
происхождения жизни исходит из того, что органические вещества образовались не в растворе, а на твердых фазах минералов. Минеральная поверхность может служить катализатором, то есть резко ускорять реакции синтеза, и одновременно образцом (матрицей) для той химической структуры, которая на ней синтезируется.
На Западе популярна версия о происхождении жизни на минерале пирите
- FeS2
. Пирит повсеместно распространен, в том числе в гидротермальных источниках. Поверхность кристалла пирита несет положительный заряд, и с ним могут связываться молекулы органических веществ. При образовании пирита из железа (Fe) и серы (S) выделяются электроны и энергия (!), которая может идти на синтез сложных органических соединений. Немецкий химик-органик Вехтершойзер считает, что первые живые клетки представляли собой кристаллические пиритовые «зерна», окруженные мембраной из органических веществ. Размножались они почкованием кристалла вместе с мембраной.
Другой вероятный кандидат на роль матрицы жизни - кристаллическая глина
. Она обладает большей структурной сложностью, чем пирит, а это хорошая основа для мутаций и эволюции образующихся макромолекул. Обсуждается также возможная роль кремнезема, слюды, кварца.
Совершенно особая ситуация возникла вокруг минерала апатита
. Его большие залежи имеются в горах Хибинах, что на Кольском полуострове. Апатит добывают ради производства фосфатных удобрений, так как в его кристаллической решетке содержится много фосфора. Кроме того, кристаллический апатит присутствует в живых системах - например, в составе зубов и костей. Кристалл имеет форму бесконечно длинной колонки очень малого диаметра. Самое парадоксальное оказалось то, что периодичность повторяющихся элементарных ячеек в кристалле апатита в 3,4 ангстрема (1 ангстрем = 0,0001 мм) точно совпадает с расстоянием между последовательными парами мономеров (нуклеотидов) в молекуле ДНК, а один шаг двойной спирали ДНК соответствует циклу в 6 ячеек апатита. Атомарная структура кристаллической решетки апатита также имела сходство со структурой ДНК. На этих основаниях российским биохимиком Э. Я. Костецким в 1981 г. выдвинута гипотеза абиогенного синтеза нуклеопротеидов, то есть комплекса ДНК и белка, на матрице апатита (рис. 28).
Рис. 28
Было проведено сопоставление данных рентгеноструктурного анализа апатита, с одной стороны, и цепей ДНК и простых белков - с другой. В результате построена пространственная модель апатита с включенными фрагментами ДНК и белков. Как видно на рис. 28, в структуре кристалла апатита легко вписываются ДНК и несколько белковых цепей. При этом источником неорганического фосфата для ДНК выступал фосфат апатита без изменения его положения в структуре кристалла, а азот включался из синильной кислоты HCN и аммиака NH3
. Согласно модели, апатит земной коры, находясь в безводной среде, подвергался постоянному и длительному воздействию глубинных газов NH3
, CH4
, HCN, CO и др. при высоких значениях давления и температуры. В таких условиях ослабляется кристаллическая решетка апатита, облегчается диффузия газовых компонентов вдоль оси кристалла при сохранении его структуры. Ионы -PO4
, присутствующие в кристалле, остаются на своих местах и определяют диаметр будущей молекулы ДНК, а другие, в частности избыточный кальций, замещаются на новые атомы из состава газовой смеси. Таким образом, при участии элементов газовой фазы и апатита внутри кристалла, а не на его поверхности, мог осуществляться одновременный синтез ДНК и белка с постепенной заменой части минеральной матрицы органической основой. На смешанных кристаллах, имеющих с апатитом совпадающие пропорции, (кальцит, арагонит, кристобалит), видимо, происходил синтез РНК, а также скелетных белков будущих клеток. Источником энергии для проведения синтезов служили силовое поле апатита, тепло глубинных процессов, энергия фосфатных связей апатита и элементов газовой фазы.
Принципиально важным и революционным в апатитовой модели является то, что в неорганическом минерале уже была заложена структурно-химическая основа ДНК и параллельно - спираль в спираль - основа полипептидных (белковых) цепей. ДНК и белок возникли одновременно и в одной связке, поэтому и их генетическое соответствие родилось в кристалле по принципу стереоспецифической комплементарности (пространственной взаимной дополнительности).
Отсюда легко выводится и РНК как посредник между ДНК и белком. Объясняется и возникновение генетического кода - через естественные множественные нарушения микроструктуры кристалла, которые закреплялись как неоднородности нуклеотидного состава ДНК. Модель предусматривает и образование липидных мембран, а также целых протоклеток про- и эукариотического типа.
Как видно, в решении проблемы происхождения жизни имеются весьма надежные основания. Современная наука уже располагает знаниями, достаточными, чтобы утверждать: жизнь возникла на первобытной Земле вполне естественным путем как процесс самоорганизации, на основе преобразования неорганических субстратов в органические макромолекулярные комплексы и, далее, в протоклетки
. Что касается конкретных механизмов образования органических комплексов, то, как показывает последний материал, будущее, очевидно, за твердоматричными моделями, среди которых наиболее продуктивна модель апатитовой матрицы
.
СЕГМЕНТ 32. ЭТАПЫ РАЗВИТИЯ ЖИЗНИ НА ЗЕМЛЕ И СОВРЕМЕННОЕ БИОРАЗНООБРАЗИЕ
Как уже было отмечено выше, первые следы жизни на Земле датируются возрастом примерно 3,6 млрд, или 3600 млн, лет. Таким образом, жизнь возникла вскоре после образования земной коры.
Историю развития жизни изучают по данным геологии и палеонтологии
, поскольку в структуре земной поверхности сохранилось много ископаемых остатков, произведенных живыми организмами. Это - так называемые осадочные породы, содержащие огромные пласты мела, песчаников и других пород, представляющих донные осадки известковых раковин одноклеточных фораминифер, кораллов, моллюсков, кремниевые скелеты и другие твердые остатки древних организмов. Например, на современной территории Евразии был когда-то океан, Уральские годы были его срединным хребтом. Поэтому любой естественный разлом, горная осыпь, крутой берег реки или обрыв карьера могут служить источником информации о прошлых жителях Земли. Ценные сведения дают также результаты бурения; например, сверхглубокая скважина на Кольском полуострове прошла уже более 12 км вглубь Земли и в ней также обнаружены окаменевшие остатки организмов. Имеются и надежные методы определения возраста земных пород, в частности радиоуглеродный метод, основанный на измерении содержания радиоактивных изотопов в составе углерода, которое закономерно изменяется во времени. Так что любую находку окаменелых остатков жизни можно датировать по времени ее происхождения.
Сразу отметим, что изменение форм жизни на Земле шло параллельно с геологической перестройкой структуры и рельефа земной коры, состава атмосферы, границ мирового океана и другими геологическими процессами. Эти изменения и обусловливали в решающей степени направление и динамику биологической эволюции
.
В соответствии с наиболее значительными событиями гео-биологической эволюции в истории Земли выделяют крупные интервалы времени - эры, внутри них - периоды, в пределах периодов - эпохи и т. д. Проследим и мы наиболее важные события, менявшие шаг за шагом биосферу нашей планеты. Для большей наглядности изобразим календарь жизни в виде условного годового цикла, в котором 1 месяц соответствует 300 млн лет реального времени (рис. 29). Тогда весь период развития жизни на Земле как раз и составит один условный год нашего календаря - от 1 января (3600 млн лет тому назад), когда образовались первые протоклетки, до 31 декабря (0 лет), когда живем мы с вами. Как видим, отсчет геологического времени принято вести в обратном порядке.
Рис. 29
Условный календарь развития жизни

Архейская эра
(эра древнейшей жизни) -
от 3600 млн лет до 2600 млн лет, протяженность 1 млрд лет - примерно четверть всей истории жизни (на нашем условном календаре это - январь, февраль, март и несколько дней апреля).
3600 - 3500 млн лет - отведем это время на те события предбиологической эволюции
, которые были описаны в предыдущем сегменте. На самом деле они могли начаться гораздо раньше, как только сформировалась земная кора.
3500-3300 млн лет - возникновение в водах мирового океана анаэробных гетеротрофных бактерий и сине-зеленых водорослей, формирование сообществ прокариотных (доядерных) одноклеточных организмов
. В атмосфере Земли еще не было кислорода, но в воде были свободные органические вещества, поэтому первые организмы - бактерии были анаэробами, а питались гетеротрофно - поглощали готовую органику и за счет брожения получали энергию (см. сегмент 13). В гидротермальных источниках, богатых выделениями сероводорода (H2
S), при температурах до 120 градусов могли жить аутотрофные хемосинтетические бактерии. По мере истощения первичных запасов органического вещества возникли аутотрофные фотосинтетические клетки - сине-зеленые водоросли. Аутотрофы, в отличие от гетеротрофов сами создают органические вещества из неорганических с использованием энергии солнца или химических реакций.
3300 млн лет - выход бактерий и сине-зеленых водорослей на сушу и начало образования почвы
. Почва представляет так называемое биокосное тело - продукт взаимодействия живых организмов с неорганическими соединениями верхнего слоя коры.
3300 - 3000 млн лет - появление в воде и атмосфере свободного кислорода
от фотосинтетических водорослей и цианобактерий. Идет также накопление углекислого газа при снижении концентрации других газов - аммиака, метана, водорода. Все это создает возможность для развития аэробных гетеротрофных бактерий
, с гораздо более продуктивным энергетическим обменом, чем у анаэробов.
3000-2700 млн лет - появление первых эукариотных клеток
, имеющих сложную структуру с настоящим ядром и органоидами (о различиях про- и эукариотных клеток см. сегмент 8 и рис. 3) . Эти клетки могли появиться в результате симбиотического объединения различных прокариотных клеток или путем постепенного усложнения последних. Во всяком случае это были значительно более крупные клетки с комбинированным энергетическим обменом, из которых в последствие и развивались основные стволы эволюции - растения, грибы и животные.
2700-2600 млн лет - в конце Архея, вероятно, появились уже и примитивные многоклеточные организмы
наподобие нитчатых водорослей и гидроидных полипов, которым предшествовали колониальные объединения одноклеточных.
Таким образом, в Архейскую эру возникли про- и эукариотные клетки с разным типом питания и энергетического обеспечения и наметился переход к многоклеточным организмам
.
Протерозойская эра
(эра ранней жизни) - от 2600 до 570 млн лет - самая протяженная эра, охватывающая около 2 млрд лет, то есть более половины всей истории жизни (на нашем условном календаре это период от апреля до начала ноября - см. рис. 29). Геологическим маркером начала новой эры стали интенсивные процессы горообразования
, изменившие соотношение океана и суши, изменившие в целом лик планеты. Многие пионерские группы организмов, сделав свое дело, вымирали, на их смену приходили новые. Но в целом биологические преобразования совершались очень медленно и постепенно.
Хотя уже в Архее появились клетки высшего - эукариотного - типа, тем не менее первая половина Протерозоя проходила при полном расцвете прокариот - бактерий и сине-зеленых водорослей
. В это время железобактерии мирового океана, осаждаясь поколение за поколением на дно, формируют огромные залежи осадочных железных руд. Крупнейшие из них до сих пор разрабатываются под Курском и Кривым Рогом. Эукариоты были представлены в основном водорослями - зелеными, красными, бурыми - преимущественно одноклеточными, наподобие современных эвглены или хлореллы. Многоклеточные организмы были малочисленны и весьма примитивны.
1500 млн лет - к середине протерозоя наблюдается расцвет зеленых водорослей
. Их фотосинтетическая активность заметно увеличивает содержания кислорода в воде и атмосфере. Примерно через 200 млн лет появляются низшие грибы
. Подобно животным они являются гетеротрофами - питаются органической пищей и используют кислород для ее окисления и получения энергии.
1000 млн лет - водоросли господствуют на Земле, темп накопления кислорода быстро возрастает
. Этому способствует также завершение окисления железа земной коры, которое до сих пор поглощало основную массу кислорода. В результате начинается бурное и независимое развитие аэробных простейших (одноклеточных) и многоклеточных животных
. Последняя четверть Протерозоя известна как «век медуз», так как эти и им подобные кишечнополостные животные составляют доминирующую и наиболее прогрессивную на то время форму жизни.
700 млн лет - прогрессивное развитие животных приобретает взрывной характер
. Особенно бурные события происходили на протяжении 40 млн лет в так называемый Вендский период (690-650 млн лет). В результате быстрых эволюционных изменений закладываются все основные типы животных. Появляются разные типы червей. От кольчатых червей (современные представители - морской пескожил, дождевой червь) ответвляются моллюски и членистоногие, похожие на мокриц, многоножек, ракообразных. К концу протерозоя закладываются морские иглокожие и примитивные хордовые. Жизнь сконцентрирована в морях, донные растения и животные образуют сложные сообщества - биоценозы. В них преобладают многоклеточные: густые заросли придонных водорослей, а среди животных - губки, медузы, актинии, черви. Моллюски, членистоногие и другие типы высших животных только начинают свою эволюцию, но общий план будущего биоразнообразия уже заложен
.
Конец Протерозоя ознаменовался и очень важным геологическим событием, которое сильно повлияло на дальнейшую биологическую эволюцию. Около 620 млн лет тому назад часть накопленного в атмосфере кислорода преобразуется из двухатомного состояния О2 в трехатомное О3. Это озон, хорошо поглощающий ультрафиолетовые лучи солнечного света. Таким образом, сформировался озоновый экран
, уровень солнечной радиации резко упал, снизилась и мутагенность солнечного излучения. Этот фактор обусловил иной характер эволюции на следующие эры. Дальнейшие новообразования будут многочисленны и разнообразны, но они будут носить все менее радикальный характер - в пределах уже сформированных биологических царств (бактерии, растения, грибы, животные) и типов.
Итак, в течение Протерозойской эры господство прокариот сменилось господством эукариот, произошел радикальный переход от одноклеточности к многоклеточности, сформировались все основные типы животного царства
. Но эти сложные формы жизни существовали исключительно в морях
. Земная суша медленно покрывалась почвой, в ее сырых низинах селились бактерии, низшие водоросли, простейшие одноклеточные животные, которые по-прежнему прекрасно существовали в своих экологических нишах. Суша еще ждала своих завоевателей. А на нашем историческом календаре уже было начало ноября. До «Нового года» - до наших с вами дней - оставалось меньше двух месяцев, всего 570 млн лет.
Палеозойская эра
(эра древней жизни) - от 570 до 230 млн лет - общая протяженность 340 млн лет. Очередной период интенсивного горообразования
, по которому и проводят разделение Протерозоя и Палеозоя, привел к изменению рельефа земной поверхности. Посреди бывшего океана встали материки, бывшие участки суши оказались под водой. В этих условиях судьба большинства групп организмов решалась радикально - быть или не быть. Некоторые группы вымерли, но другие приспосабливались и осваивали новые среды обитания вслед за изменениями земной поверхности и климата
.
Палеозойская эра делится на 6 периодов. Об этом времени имеются богатые палеонтологические данные, так как многие животные в процессе эволюции приобрели наружный или внутренний скелет, а растения - плотные клеточные оболочки, что и позволило им сохраниться в виде ископаемых остатков. Мы сможем дать лишь самую общую характеристику событий этого времени, отметив наиболее значимые новообретения жизни. Общий ход эволюции, начиная с этого времени, отражен на рис. 30. Обратите внимание на то, что большинство групп организмов, зародившихся в прежние эпохи, продолжают сосуществовать с вновь появляющимися молодыми группами, хотя многие сокращают свой объем. Природа расстается с теми, кто не соответствует меняющимся условиям, но максимально сохраняет удачные варианты, отбирает и развивает из них наиболее приспособленные и, кроме того, создает новые формы.
Рис. 30 Эволюционное развитие живого мира
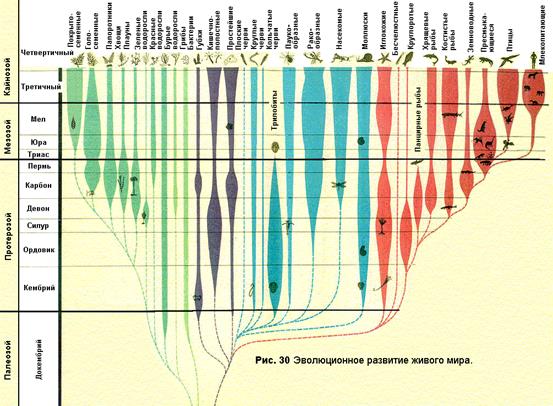
Кембрий
- 570 - 500 млн лет. Жизнь по-прежнему связана в основном с морским дном. Процветают различные водоросли - зеленые, бурые, красные. Но появляются высшие растения
- завоеватели суши
. Их тело разделено на корень и стебель, что позволяет хорошо закрепляться на почве и доставать из нее влагу и минеральные вещества. Внутри стебля возникают сосуды для транспорта веществ. Это первые сосудистые растения - хвощи и плауны.
Среди морских животных идут процессы видообразования в пределах ранее возникших групп. Заметным событием становится образование коралловых рифов - нагромождения сидячих форм мелких кишечнополостных животных, родственников обыкновенных гидр и медуз, имеющих твердый известковый скелет. Но существуют и подвижные формы - двустворчатые, брюхоногие (улитковые) и головоногие моллюски, черви, примитивные ракообразные.
Ордовик
- 500 - 440 млн лет. Сохраняется умеренный теплый климат, но значительно увеличивается площадь морей. Морская фауна и флора бурно эволюционирует в сторону увеличения разнообразия видов внутри сформированных типов и классов
. Заметное место начинают занимать иглокожие (морские лилии, голотурии) и хордовые (круглоротые) животные.
Силур
- 440 - 410 млн лет - новый период поднятия суши и горообразования. Теплые мелководные моря отступают, климат в целом становится более сухим. В растительном царстве прежнее обилие водорослей, на земле доминируют псилофиты - растения без листьев, внешне похожие на своих морских предшественников. В морях богатая фауна трилобитов (примитивных ракообразных), головоногих моллюсков, кораллов. Появляются панцирные «рыбы» - бесчелюстные хрящевые позвоночные, впрочем, вымершие уже в следующие периоды. На сушу выходят первые членистоногие животные
- многоножки, пауки, ракоскорпионы, бескрылые насекомые. В конце силура происходит очередной процесс горообразования.
Девон
- 410 - 350 млн лет. Идет новое поднятие суши и сокращение морей. Климат становится более контрастным. На земле появляются мхи, папоротники, грибы
. Формируются первые леса, состоящие из гигантских папоротников, хвощей и плаунов. В морях наблюдается расцвет рыб
, особенно хрящевых (акулообразные). Появляются двоякодышащие (с жабрами и легкими), кистеперые и лучеперые рыбы - непосредственные предшественники наземных позвоночных. На суше в леса продвигаются членистоногие с воздушным дыханием - многоножки, насекомые. Наконец, появляются первые земноводные позвоночные, или амфибии
, среди их ископаемых форм наиболее известны стегоцефалы. Заметим, что первые покорители суши - как растительные, так и животные - занимали прибрежные области, так как вода оставалась средой их размножения.
Карбон
- 350 - 280 млн лет. Наступает потепление и увлажнение климата. Кругом распространены болотистые леса из громадных (до 40 м) древовидных папоротников, хвощей и плаунов
. К ним добавляются семенные папоротники - первые голосеменные растения. Именно эти леса оставили нам огромные залежи каменного угля («каменноугольные леса»
). Среди наземных животных кроме разнообразных стегоцефалов появляются крылатые насекомые - тараканы, стрекозы. В конце карбона идет поднятие суши и похолодание, исчезают псилофиты и появляются первые пресмыкающиеся животные
, окончательно освободившиеся от водной зависимости.
Пермь
- 280 - 230 млн лет. Очередное поднятие суши сопровождалось уменьшением влажности воздуха и похолоданием. Сужается к экватору зона пышных лесов. Вслед за псилофитами вымирают древовидные хвощи, плауны, папоротники. Наступает расцвет семенных и травянистых папоротников, появляются древние голосеменные растения
- предшественники наших хвойных. Из-за сухости климата вымирают и крупные земноводные - стегоцефалы. Зато быстро прогрессируют пресмыкающиеся и насекомые
- их яйца защищены от высыхания плотными оболочками, кожа покрыта чешуей или хитином, усложняются легкие или трахеи. В двух совершенно не родственных группах происходит параллельная эволюция на сходные признаки.
Общий итог Палеозоя - заселение суши растениями, грибами и животными
. При этом и те, и другие, и третьи в процессе своей эволюции усложняются анатомически, приобретают новые структурные и функциональные приспособления для размножения, дыхания, питания, способствующие освоению новой среды обитания. Эволюция идет по пути морфофизиологического прогресса
. Завершается Палеозой, когда на нашем календаре 7 декабря. Природа торопится, темп эволюции в группах высок, сжимаются сроки преобразований, но на сцену только выходят первые пресмыкающиеся, а время птиц и млекопитающих еще далеко впереди.
Мезозойская эра
(эра средней жизни) - от 230 до 67 млн лет - общая протяженность 163 млн лет. Поднятие суши, начавшееся в предыдущем периоде, продолжается. Существует единый материк. Его общая площадь очень велика - значительно больше, чем в настоящее время. Континент покрыт горами, сформированы Урал, Алтай и другие горные массивы. Климат становится все более засушливым.
Триас
- 230 -195 млн лет. Закрепляются тенденции, заложенные в пермском периоде. Вымирает большинство первобытных земноводных, почти исчезают хвощи, плауны, папоротники. Преобладают голосеменные древесные растения, так как их размножение не связано с водной средой. Среди животных на суше начинают свое триумфальное шествие травоядные и хищные пресмыкающиеся - динозавры
. Среди них есть уже и современные виды: черепахи, крокодилы, гаттерия. В морях по-прежнему живут земноводные, разнообразные головоногие моллюски, появляются костистые рыбы вполне современного вида. Это обилие пищи привлекает в море хищных пресмыкающихся, отделяется их специализированная ветвь - ихтиозавры. В конце триасового периода от каких-то ранних пресмыкающихся обособилась небольшая группа, давшая начало млекопитающим животным
. Они еще размножаются с помощью яиц, как современные ехидна и утконос, но уже имеют важную особенность, которая даст им преимущества в дальнейшей борьбе за существование. Млекопитающие, как и птицы, ведущие свое начало также от пресмыкающихся, являются теплокровными животными - они впервые приобретают механизм температурной саморегуляции. Но их время еще впереди, а пока земные пространства продолжают осваивать динозавры.
Юра
- 195 - 137 млн лет. В лесах преобладают голосеменные, среди них живет уже секвойя, дошедшая и до наших дней. Появились первые покрытосеменные (цветковые) растения. Господствуют гигантские пресмыкающиеся, освоившие все среды обитания
. На суше это - травоядные и хищные динозавры, в море - ихтиозавры и плезиозавры, в воздухе - летающие ящеры, охотящиеся на многочисленных насекомых и своих же более мелких собратьев. От каких-то из них обособились первоптицы - археоптериксы
. Они имели скелет ящеров, хотя и сильно облегченный, но уже были покрыты перьями - видоизмененными кожными чешуями. В теплых морях Юрского периода, кроме морских пресмыкающихся, процветают костистые рыбы и разнообразные головоногие моллюски - аммониты и белемниты, похожие на современных наутилуса и кальмаров.
В Юрском периоде происходит раскол единого материка и начинается расхождение материковых плит к их современному состоянию
. Это привело к изоляции и относительно независимому развитию фауны и флоры на разных материках и островных системах. Особенно быстро и радикально обособилась Австралия, где животный и растительный состав в итоге сильно отличался от обитателей других материков.
Мел
- 137 - 67 млн лет. Руководящей формой в палеонтологических пробах выступают фораминиферы - раковинные простейшие животные, подвергшиеся массовому вымиранию в этот период и оставившие огромные осадочные пласты мела
. Среди растительности быстро распространяются и доминируют покрытосеменные
, многие из них вполне современного вида и уже имеют настоящий цветок
. Гигантские пресмыкающиеся вытесняются новыми динозаврами
, передвигающимися на задних ногах. Вполне обычны первоптицы, но появляются и настоящие теплокровные птицы
с характерным клювом, без длинного хвоста. Встречаются и мелкие млекопитающие; кроме сумчатых появились и плацентарные
, длительно вынашивающие детенышей в материнской утробе в контакте с кровью через плаценту. Насекомые осваивают цветок
, что принесло выгоду и насекомым и цветковым растениям.
Конец мелового периода был ознаменован значительным общим похолоданием
. Сложная пищевая цепь пресмыкающихся, построенная на ограниченном круге продуцентов, рухнула «в одночасье» (по меркам нашего условного календаря). За какие-то несколько миллионов лет основные группы динозавров вымерли
. Есть разные версии причин происшедшего в конце мелового периода, но, по-видимому,
дело прежде всего в изменении климата и разрушении пищевых цепей
. В похолодевших морях исчезли крупные головоногие моллюски - основная пища морских ящеров. Естественно, это привело к вымиранию последних. На суше происходило сокращение зоны произрастания и биомассы мягкой сочной растительности, что привело к вымиранию растительноядных, а за ними и хищных динозавров. Сократилась кормовая база и для крупных насекомых, а за ними стали исчезать летающие ящеры - как насекомоядные, так и их хищные собратья. Надо иметь в виду и то обстоятельство, что пресмыкающиеся были холоднокровными животными и оказались не приспособленными к существованию в новом, значительно более суровом климате
. В этой всемирной биологической катастрофе выжили и получили дальнейшее развитие мелкие пресмыкающиеся - ящерицы, змеи; а крупные - такие как крокодилы, черепахи, гаттерия - сохранились лишь в тропиках, где остались необходимая кормовая база и относительно теплый климат.
Таким образом, мезозойская эра полноправно называется эрой пресмыкающихся. За 160 млн лет они пережили свой расцвет, широчайшую дивергенцию по всем средам обитания и вымерли в борьбе с неизбежной стихией. На фоне этих событий
огромные преимущества получили теплокровные организмы - млекопитающие и птицы, перешедшие к освоению освобожденных экологических сфер
. Но это уже была новая эра. До «Нового года» оставалось 7 дней.
Кайнозойская эра
(эра новой жизни) - от 67 млн лет до настоящего времени. Это эра цветковых растений, насекомых, птиц и млекопитающих. В эту эру появился и человек.
Третичный период
делится на палеоген
(67 - 25 млн лет) и неоген
(25 - 1,5 млн лет). Отмечается широкое распространение цветковых растений, особенно травянистых
. Формируются обширные степи - результат отступления тропических лесов из-за похолодания. Среди животных доминируют млекопитающие, птицы, насекомые
. Продолжают исчезать отдельные группы пресмыкающихся и головоногих моллюсков. Около 35 млн лет назад в классе млекопитающих появляется отряд приматов (лемуры, долгопяты), давший в последствие обезьян и человека
. Первые люди появились около 3 млн лет тому назад (за 7 часов до «Нового года») в восточном средиземноморье.
Четвертичный период
, или антропоген
включает последние 1,5 млн лет развития жизни. Сформирован современный растительный и животный мир. Идет бурная эволюция и господство человека
. Происходит четыре периодических оледенения северного полушария Земли. За это время вымерли мамонты, многие крупные звери, копытные животные. Большую роль в этом сыграли люди, которые активно занимались охотой и земледелием. Периодические замерзания и таяния воды изменяли уровень морей, то наводя, то разрушая мосты между Азией и Северной Америкой, Европой и Британией, Индокитаем и Островами. Эти обстоятельства позволяли мигрировать животным и растениям, поддерживая их эволюционные изменения по мелким адаптивным признакам. Австралия имеет полную изолированность от других материков, что создало там особые направления и темпы эволюции. Отсутствие хищников позволило сохраниться древним сумчатым и яйцекладущим млекопитающим, давно вымершим на других континентах. Происходили изменения и в семействе людей, но о них мы расскажем в отдельной теме. Здесь же отметим, что человек современного типа сформировался всего лишь 50 тысяч лет назад (в 23 часа 53 минуты 31 декабря нашего условного года развития жизни на Земле; мы существуем в этом году всего-то его последние 7 минут!).
Как итоговое отражение эволюции - с взлетами одних групп и крушениями других - выступает современная картина биоразнообразия
,
представленная на рис. 31.
Рис. 31 Схема современного биоразнообразия на Земле

Еще раз обратим внимание на то, что древние группы организмов - дробянки (бактерии и сине-зеленые водоросли), водоросли, одноклеточные (простейшие) животные не исчезли с биологического поля. Конечно, многие их представители, и даже целые большие группы, вымерли и уступили место другим. Но в целом эволюция не теряла заложенные когда-то планы строения животных и растений. В каждой крупной группе (ранга царства или типа), как правило, возникали новые вариации, которые занимали свои экологические ниши. Поэтому мы и имеем на сегодня столь широкий спектр биоразнообразия - от бактерий и вирусов до цветковых растений, насекомых и млекопитающих.
Несколько слов о царстве вирусов
. Никто не знает, каково их настоящее происхождение. По уровню организации они отличаются от всех прочих организмов тем, что не имеют клеточного строения. По этому признаку и выделены две империи, одна из них за вирусами. Есть мнение, что вирусы - «осколки жизни», блуждающие генетические элементы, мигрирующие между разными организмами. Но кто и когда породил эти осколки - науке не известно.
И последнее замечание - о движущих силах биологической эволюции
. Их надо видеть в двух взаимосвязанных плоскостях - геологической и собственно биологической
. Каждая очередная крупномасштабная перестройка земной поверхности влекла за собой неизбежные эволюционные преобразования в живом мире. Каждое новое похолодание приводило к вымиранию неприспособленных. Дрейф материков определил различие темпов и направлений эволюции в изолятах. С другой стороны, прогрессивное развитие и размножение групп растений и животных сказывалось и на самой геологической эволюции. Вспомним, как накопление кислорода в конце Протерозоя привело к образованию озонового экрана, что оградило живые организмы от солнечной радиации и изменило уровень эволюционных преобразований. Многие продукты жизнедеятельности организмов оставались навсегда в земных недрах, преобразуя их необратимо. Это и органогенные железные руды, и залежи серы, мела, каменного угля и многое другое. Живое, порожденное из неживой материи, эволюционирует вместе с ней, в едином биогеохимическом потоке вещества и энергии. Что же касается внутренней сущности и непосредственных факторов биологической эволюции, мы рассмотрим их в следующем сегменте.
СЕГМЕНТ 33. ФАКТОРЫ БИОЛОГИЧЕСКОЙ ЭВОЛЮЦИИ
Процесс необратимого исторического развития мира живых организмов - как в целом, так и отдельных таксономических групп - называется филогенезом
(от греческих phylon - род, племя и genesis - происхождение, возникновение).
Как мы уже видели, развитие - важнейшее свойство жизни. Но развитие, самоорганизация присущи материи вообще. В философском понимании развитие - это движение, изменение в пространстве и времени. Движение как способ существования материи не возникает и не исчезает, существует вечно - без начала и конца. В то же время движение - векторный, направленный, необратимый процесс, а значит на отдельных отрезках имеет начало и конец: ........ - ........ - ........ - ........ .
Таким образом, развитие как движение бесконечно и вместе с тем существует всякий раз как отдельный конечный процесс. Эти кажущиеся противоречия разрешаются при диалектическом взгляде на процесс развития. Развитие любой системы: физической, биологической, социальной - подчиняется общим законам диалектики и синэргеники. Развитие совершается по спирали и бифуркационно
(рис. 32).
Рис. 32
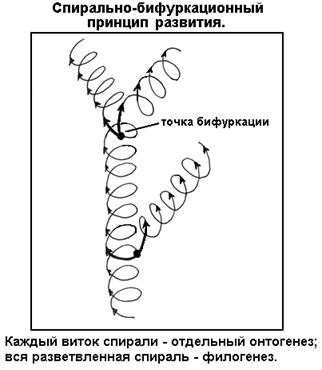
Один виток спирали - некоторый конечный процесс. В схеме исторического развития виток представляет онтогенез
(индивидуальное развитие) отдельной особи.
Вся спираль с ее бифуркационными разветвлениями - бесконечный процесс. Бифуркационная спираль представляет филогенез
- совокупность онтогенезов с постепенными изменениями, ведущая к их необратимой эволюции.
Таким образом, любые филогенетические преобразования происходят посредством перестройки онтогенезов особей; при этом приспособительную ценность могут иметь изменения любой стадии индивидуального развития. Можно сказать, что филогенез представляет собой преемственный ряд онтогенезов последовательных поколений. Это важнейшее концептуальное положение биологии наглядно проявляется в так называемом биогенетическом законе
, согласно которому в онтогенезе высшего организма частично сохраняются (повторяются) основные черты предковых низших организмов, то есть отражается его филогенез.
Действительно, при сравнительно-эмбриологическом изучении родственных групп организмов обнаруживается, что их зародыши более сходны, чем взрослые особи. При этом чем более ранние стадии сравниваются, тем больше сходства выявляется. Например, у всех представителей позвоночных прослеживаются одинаковые эмбриональные стадии и признаки: бластула, гаструла, трехслойность, закладка хорды и ее преобразование в позвоночник, кровеносная система с одним кругом кровообращения, закладка жаберных карманов, хвоста. На этой стадии все зародыши имеют типичные рыбьи признаки. Далее появляются различия: постепенно можно обнаружить новые признаки классов, потом отрядов и т. д. до видовых различий, которые проявляются перед самым рождением. Жабры развиваются только у рыб и амфибий (на стадии головастика); у пресмыкающихся, птиц и млекопитающих жаберные щели зарастают. У зародышей млекопитающих, особенно у приматов, быстро развивается головной мозг. Таким образом, по ходу эмбрионального развития идет расхождение (дивергенция) признаков у зародышей, отражающее их филогенетическую дивергенцию. Эту закономерность уже в середине XIX века отмечал Дарвин: эмбриональное развитие вида отражает его эволюцию. А вскоре Мюллер и Геккель сформулировали ее в форме биогенетического закона. На основании биогенетического закона путем изучения эмбрионального развития организмов можно устанавливать степень их филогенетического родства.
Какие же факторы и каким образом движут филогенетическое развитие? Почему возникают изменения в индивидуальных онтогенезах? Что происходит в точке бифуркации, за которой один биологический вид превращается в два или более видов с новыми признаками? На эти вопросы дает ответ синтетическая теория эволюции
. Ее формирование имеет свою сложную историю.
Еще в античные времена существовало два противоположных взгляда по вопросу происхождения современного разнообразия животных и растений. Креационисты
утверждали, что мир сотворен Богом; животных и растений столько, сколько их создал Творец. Они изначально целесообразны и неизменны. В понятиях философии это - чистейшая метафизика (от греческого «Meta ta physika» - «После физики»; так называлось философское сочинение Аристотеля, помещенное после его трактатов по физике). Даже в XVIII веке этот взгляд разделял великий шведский натуралист, основатель современной систематики Карл Линней. Другая точка зрения - трансформизм
: мир изменяется, исторически развивается. Применительно к биологии это означало историческое изменение организмов, постепенное усложнение их организации. В XVIII веке такие мысли выражали Бюффон во Франции, Эразм Дарвин (дед Чарльза Дарвина) в Англии, Гёте в Германии, Ломоносов в России). Но первые эволюционные догадки были умозрительные, они не имели достаточной фактической основы. Только в XIX веке сформировалось научное эволюционное учение.
Первым настоящим эволюционистом считается французский биолог Ж.Б. Ламарк
. В своей известной книге «Философия зоологии» (1809) он предложил первую целостную теорию эволюции живого мира, отвергнув метафизическую идею о постоянстве и неизменности видов. Учение Ламарка основано на новом видении системы - от простых организмов к сложным, оно утверждало прогрессивное развитие жизни на Земле. Но Ламарк ошибался в оценке движущих сил (факторов) эволюции. Совершенно справедливо признавая ведущую роль в изменении видов за внешними условиями (климат, почва, пища, свет, теплота и т. д.), он ошибочно считал, что упражнение органов у отдельной особи ведет к их целенаправленному изменению, совершенствованию, и эти изменения автоматически закрепляются в потомстве, передаются по наследству (наследование благоприобретенных признаков).
Этот постулат Ламарка стал темой долгих дискуссий в биологии, а в последствие и в генетике. В Советском Союзе в 30-50-е годы Ламаркизм был использован в обоснование антинаучных, антигенетических построений группы Т. Д. Лысенко, захватившей при поддержке Сталина ключевые посты в Академии наук и фактически учинившей разгром отечественной школы генетиков. Развитие биологии в Союзе было задержано на десятилетия.
Тем не менее эволюционную идею Ламарка разделяли почти все биологи его времени. Но причины и движущие силы филогенетических процессов верно вскрыл только Дарвин.
Чарльз Дарвин
(1809-1882) - великий английский натуралист, создатель учения о происхождении видов путем естественного отбора. С детства увлекался коллекционированием и изучением растений, животных и минералов. В 1831-1836 гг. в качестве натуралиста совершил кругосветное путешествие на корабле «Бигль», в ходе которого собрал большой биологический, географический, геологический и палеонтологический материал, легший в основу его учения. Досконально изучил историю селекции домашних животных и растений и пришел к выводу, что искусственный отбор играет такую же важную роль при создании пород животных и сортов растений, какую естественный отбор играет в дикой природе. Основные эволюционные труды: «Происхождение видов путем естественного отбора...» (1859), «Прирученные животные и возделанные растения» (1868), «Происхождение человека и половой отбор» (1871), «Выражение эмоций у человека и животных» (1872). В знак больших заслуг и всеобщего признания похоронен в Вестминстерском аббатстве рядом с И. Ньютоном.
Синтетическая теория эволюции сложилась к середине XX века на основе классической теории эволюции Дарвина, генетики и экологии. По сути она развивала на новом уровне знания идею естественного отбора, обоснованную Дарвином за 100 лет до этого синтеза. Какие же базовые положения были привнесены в синтетическую теорию из ее трех источников?
Дарвинизм
остался фундаментом новой теории. В классическом произведении Дарвина «Происхождение видов путем естественного отбора...» обозначены главные факторы, или движущие силы, эволюционного процесса. Согласно теории, (1) виды способны к неограниченному размножению, при этом им свойственна неопределенная наследуемая изменчивость (ненаправленная, случайная - в отличие от представлений Ламарка); (2) из-за ограниченности ресурсов (пища, пространство и др.) особи и виды вступают в борьбу за существование, в ходе которой большая часть гибнет и не дает потомства; (3) выживают и оставляют потомство наиболее приспособленные к данным условиям среды, в чем и проявляется естественный отбор; (4) отбор накапливает у разных особей одного вида разные приспособительные признаки, что ведет к их дивергенции и образованию новых видов. Таким образом, наследственная изменчивость
представляет предпосылку эволюции, борьба за существование и естественный отбор -
ее процесс, а видообразование
- результат.
Генетика
уже в новом, XX веке дала понимание природы наследственности и механизмов изменчивости. Открытые в 1865 г. и переоткрытые в 1900 г. законы Менделя о независимом наследовании отдельных признаков и их сохранении в скрытом (рецессивном) состоянии позволили объяснить то, что Дарвин не понимал и не мог понимать, так как природа наследственности в его время была неизвестна. Самая суть генетической основы эволюции состоит в том, что каждый признак кодируется двумя аллельными генами - от двух родителей (хромосомный набор диплоидный). Один из них может быть сильнее другого - доминантный, тогда другой - слабый, мутантный ген проходит как рецессивный и может не проявиться в нескольких поколениях, пока не сойдется в паре с таким же рецессивным геном.
Допустим, некий ген А кодирует желтый пигмент у семян гороха, тогда ген а представляет его рецессивную мутацию. Растения, у которых оба гена доминантные - АА (доминантная гомозигота), будут иметь безусловно желтые семена. При комбинации Аа (гетерозигота) семена также будут желтые, так как для проявления признака достаточно и одного гена - пигмент все равно будет вырабатываться. Но такие генетически смешанные особи несут потенциальную способность дать искажение признака в следующих поколениях. Дело в том, что гетерозиготные родители производят разные половые клетки (они гаплоидные) - либо А, либо а. При их скрещивании случайные комбинации гамет (половых клеток) могут дать разные генотипы: АА, Аа. аа. Растения с генотипом аа (рецессивная гомозигота) будут иметь не желтый, а простой зеленый или бесцветный фенотип, так как желтый пигмент у них не вырабатывается - ген дефектный в обеих хромосомах. Вот почему некоторые признаки у детей наследуются не от родителей, а от бабушек и дедушек или даже более далеких предков.
Российский генетик С. С. Четвериков в 20-х годах разработал очень важное положение о генофонде вида, популяции, под которым понимается совокупность всех генов, участвующих в свободном скрещивании. Дело в том, что в половых клетках организмов могут возникать спонтанные или индуцированные внешними факторами мутации (допустим - d) - искажения молекулы ДНК, несущие потенциально новый признак. Пока мутация сохраняется в гетерозиготном состоянии (Dd) - под прикрытием нормального гена, она никак не проявляется. Но если мутация при скрещивании выходит в гомозиготу (dd), прежде скрытый признак проявляет себя фенотипически (то есть выявляется внешне - как новый цвет, новая форма, новая ферментная активность и т. п.) и проверяется путем естественного отбора на свою полезность, бесполезность или вредность. Таким образом, гетерозиготные особи в популяциях несут скрытый резерв наследственной изменчивости. Чем более популяция насыщена гетерозиготами, тем выше у нее эволюционный потенциал.
Наконец, современная экология
дала для синтетической теории эволюции понимание того, что элементарные процессы изменчивости, скрещивания и наследования происходят не у отдельных особей, как считал Дарвин, а в рамках целой популяции организмов. Популяция - относительно изолированная группа особей одного вида, длительное время населяющих определенный ареал и способных к свободному скрещиванию. Генетические изменения на уровне популяций изучает популяционная генетика, основателем которой и был Четвериков. Кроме того, из экологии используются знания о механизмах возникновения изоляций внутри популяции или между близкими популяциями одного вида. Изоляции крайне важны на заключительных этапах видообразования, так как с их помощью закрепляются возникшие новые признаки.
Теперь мы готовы сформулировать 10 основных положений синтетической теории эволюции
, практически исчерпывающих всю проблему филогенеза. Это самая крупная и значимая биологическая теория.
1. Единицей существования, воспроизводства, изменчивости и эволюции вида является популяция
. Главное условие эволюционных изменений в популяции - способность ее членов к свободному скрещиванию.
2. Необходимым условием, предпосылкой эволюции является наследственная изменчивость
отдельных особей в популяции. Изменчивость бывает двух видов. Мутационная изменчивость особей возникает в результате спонтанных или индуцированных мутаций генов в половых клетках их родителей и несет в рецессивном состоянии потенциальные новые признаки. Комбинативная изменчивость является результатом случайных комбинаций отцовских и материнских генов в половом процессе: в ходе мейоза (кроссинговер и независимое расхождение хромосом половых клеток) и при образовании зиготы в результате оплодотворении яйца сперматозоидом. Комбинативная изменчивость не создает новых генов, но в результате комбинаций внешние признаки могут сильно варьировать.
Наследственная изменчивость отдельных особей ведет к возрастанию гетерозиготности генофонда популяции. В результате скрещивания гетерозиготных особей появляются гомозиготы с новым признаком, которые подлежат отбору на пригодность в данных условиях. Чем выше гетерозиготность, тем чаще возникают гомозиготы и больше предложений для отбора. Таким образом, высокая гетерозиготность - скрытый резерв эволюции.
3. Концентрация мутаций и, соответственно, гетерозигот повышается при усилении действия мутагенных факторов
. Естественными мутагенами являются: ионизирующая радиация космоса, Солнца и земных недр, химические пищевые вещества (особенно свободные активные радикалы), повышенная температура, вирусы и др. Интенсивность природных мутагенных факторов меняется в определенные периоды развития Земли, обусловливая изменение темпов и масштабов эволюции. Мутации возникают также спонтанно - как обычные ошибки в ходе репликации ДНК при подготовке к делению клетки, поэтому их минимальный уровень обеспечен в любое время исторического развития.
4. Концентрация мутаций и гетерозигот в популяции организмов может случайно повышаться в результате резких колебаний численности особей. Такие колебания - популяционные волны
- регулярно совершаются по экологическим причинам: при саморегуляции в пищевых цепях; при колебаниях численности популяций в лунных (месячных), сезонных (годичных), солнечных (11-летних) и других циклах; в результате природных или техногенных катаклизмов - пожаров, засух, наводнений, оледенений и т. п. В малых популяциях вероятность встречи одноименных гетерозигот и их выхода в гомозиготу резко возрастает. Такое смещение частоты гетерозигот в пользу определенных генотипов называется дрейфом генов
.
5. Случайные изменения генофонда популяции закрепляются благодаря изоляциям
. Изоляция - это возникновение разнообразных препятствий к свободному скрещиванию особей внутри популяции, а также между близкими популяциями, которые еще сохраняли генетический обмен на краях своих ареалов. При возникновении изоляции размножение идет в пределах изолятов - обособленных групп, так что между ними прекращается обмен генами. Возникшие ранее изменения генотипа закрепляются в потомстве и усиливаются, что ведет к расхождению признаков особей (дивергенции) в пределах бывшей однородной популяции.
Изоляции могут быть пространственные (географические, экологические) и биологические (анатомические, физиологические, поведенческие, цитогенетические и др.). Пространственные изоляции появляются в результате геологических процессов (горообразование, поднятие и опускание уровня воды и т. п.), а биологические - как результат закрепления мутаций в пространственных изолятах.
Все уже рассмотренные факторы эволюции носят случайный, не направленный характер - они повышают или понижают концентрацию разных мутаций в популяции. Сами они не создают направленный процесс эволюции. Единственный направляющий фактор эволюции - естественный отбор.
6. Борьба за существование и естественный отбор
- процесс, направленный на избирательное выживание и возможность оставления потомства с полезными в данных условиях признаками. Отбирающим фактором выступают условия среды, борьба за существование. Главная причина
борьбы за существование - ограниченность пищевых ресурсов для молоди, при том что организмы стремятся к беспредельному размножению.
Различаются: (1) внутривидовая борьба, хотя существуют и ее ограничения - разделение территорий, отпугивающие метки, семейная и стайная взаимопомощь и др.; (2) межвидовая борьба - при перекрытии ареалов, совпадении пищевых объектов; (3) «борьба» с неблагоприятными условиями среды - способность выживать при появлении таких условий (похолодание, засуха и др.).
Таким образом, борьба за существование
есть непосредственная причина
и механизм естественного отбора, или: естественный отбор
- следствие
борьбы за существование. Материалом
для отбора является генетическая неоднородность
популяций. Эффективность отбора
тем выше, чем больше гетерозиготность
в популяции и чем активнее идут процессы изоляции
.
7. Эволюция носит приспособительный характер
. Закрепляемые признаки адаптивны (имеют приспособительное значение) в данных условиях, в данное время. Адаптации относительны, так как в других условиях и в другое время они теряют смысл. Некоторые признаки могут быть нейтральными, не имеющими приспособительного значения. Их гены находятся в хромосомах вблизи с генами полезных признаков и наследуются сцепленно с ними.
8. Расхождение признаков в популяциях - дивергенция - постепенно ведет к образованию новых биологических видов
. Этот процесс называется микроэволюцией
. Главное в видообразовании - прекращение скрещивания между отдельными изолятами внутри популяции или между популяциями исходного вида.
Биологический вид
- это совокупность особей (популяций), обладающих наследственным сходством по морфологическим, физиологическим, генетическим и эколого-географическим признакам, способных свободно скрещиваться и давать плодовитое потомство; вид - это наименьшая генетически устойчивая надорганизменная система.
Видообразование, или микроэволюция, идет и в настоящее время - в так называемых сложных видах, где велика внутривидовая изменчивость и выделяются несколько подвидов или рас.
9. Наиболее радикальные мутации или их сочетания, затрагивающие ранний онтогенез и дающие значительные отклонения в организации особей, ведут к макроэволюции
- возникновению таксонов выше видового ранга. Так возникали роды, семейства, отряды, классы, типы... Из наиболее крупных макроэволюционных новообразований (их называют ароморфозами) можно вспомнить возникновение проводящих сосудов или цветка у растений, развитие легких у наземных животных, формирование теплокровности у млекопитающих и птиц.
В целом микро- и макроэволюционные преобразования ведут к биологическому прогрессу
. Признаки прогресса - возрастание приспособленности организмов к окружающей среде, ведущее к увеличению их численности и расширению ареала. Обратные процессы характеризуются как биологический регресс
, ведущий к вымиранию группы. Биологическая эволюция - это сочетание
прогресса одних групп с регрессом других.
10. Эволюция организмов полностью зависит от геологической эволюции Земли
. Изменения геологических условий среды происходили периодично и в целом необратимо. Часто они проявлялись в форме крупномасштабных катаклизмов. По этой причине в истории жизни отмечается изменчивость темпов эволюции, смена форм естественного отбора. В периоды быстрых изменений среды темп эволюции возрастает, идет так называемый движущий отбор - на закрепление новых признаков. В периоды относительного спокойствия среды темп эволюции снижается, а отбор приобретает стабилизирующий характер - на консервацию имеющихся признаков и отсечение всяких новообразований.
Биологическая эволюция, как и зарождение жизни, - естественный материальный процесс развития Земли
.
СЕГМЕНТ 34. ПРОИСХОЖДЕНИЕ И ЭВОЛЮЦИЯ ЧЕЛОВЕКА
Вопрос о происхождении человека принципиально важен, поскольку вокруг него скрещиваются интересы многих наук, религии, культуры. Кто он - человек? Хозяин природы или ее часть? Животное или особое одухотворенное создание? Когда и откуда появился на Земле?
Уже Карл Линней, которого трудно заподозрить в атеизме, выделяя в системе животных группу приматов, поместил туда и человека вместе с обезьянами. Ламарк прямо выводил человека из обезьяноподобных предков. Дарвин в «Происхождении человека...» (1871) пришел к выводу, что человек имеет дальних предков с человекообразными обезьянами, что это последнее звено в цепи развития живых существ (в своей ветви эволюционного древа).
Действительно, имеется масса доказательств
животного
происхождения человека
. По всем анатомо-физиологическим признакам род Homo (человек) относится к типу хордовых, подтипу позвоночных, классу млекопитающих, отряду приматов, семейству людей.
Признаки млекопитающих
: общее эмбриональное развитие; общее строение скелета и внутренних органов; разделение зубов на коренные, клыки и резцы; млечные железы; внутриутробное развитие; диафрагма, обеспечивающая легочное дыхание; кожа с волосом; 4-камерное сердце; рудименты (копчик, аппендикс, третье веко) и атавизмы (появляющиеся у отдельных индивидуумов хвост, волосатость лица, дополнительные соски, мощные клыки).
Признаки приматов
: общая анатомия, физиология, биохимия и молекулярно-генетические признаки; в частности, сильно развит головной мозг, смена молочных зубов на постоянные, передняя конечность хватательного типа, наличие ногтей, одна пара сосков молочных желез, глаза в одной плоскости и др.
Человек относится к семейству гоминид, или людей
. Наибольшее его сходство проявляется с семейством антропоидов, или человекообразных обезьян, таких как горилла, орангутан, шимпанзе и близкие к ним гиббоны
. Общие признаки: пропорции тела; исчезновение хвостового отдела позвоночника; способ передвижения - прямохождение и ведущая роль задних конечностей; большой объем головного мозга; одинаковые группы крови по системе AB0 и резус-фактор; общие паразиты (головная вошь и др.) и болезни (грипп, оспа, холера, брюшной тиф); близость хромосомного набора (у антропоидов 2n=48, у человека 2n=46 - из-за слияния двух хромосом в одну); высокий уровень молекулярно-генетического сходства - гомология по ДНК более 95%); общее выражение эмоций с развитием мимической мускулатуры; хорошая память; сходная высшая нервная деятельность.
Однако человек приобрел и важные отличия от своих ближайших родственников, на которые мы укажем ниже.
Что же способствовало выделению из состава приматов и специфическому развитию семейства людей? Каковы предпосылки
возникновения человека?
Прежде всего надо иметь в виду, что современные человекообразные обезьяны не являются прямыми предшественниками человека, но имеют с ним общих предков (наши «двоюродные братья и сестры»). Имеется много ископаемых остатков наших предшественников - полулюдей - полуобезьян, с признаками, промежуточными между человеком и обезьяной.
Первая важнейшая предпосылка антропогенеза - переход обезьян с деревьев на землю
и вынужденное передвижение на задних конечностях, помогая передними. Это способствовало, в свою очередь, выпрямлению туловища, изменению соотношения между внутренними органами, развитию новых групп мышц, образованию сводчатой стопы и S-образного позвоночника - для лучшей амортизации при ходьбе. Но самое главное - переход на задние конечности высвободил передние для добывания пищи, а впоследствии и для трудовой деятельности. Произошло некоторое укорочение передних конечностей, увеличилось разнообразие их движений, сформировалась гибкая кисть с оттопыренным большим пальцем. Так возникла рука человека.
Вторая предпосылка - стадный (социальный) образ жизни
. Многие животные живут стадами, стаями, иными общественными группировками. Социальность всегда способствовала прогрессу вида и его отдельных индивидуумов. Та группа предковых обезьян, которая выбилась в отдельную ветвь людей, вероятно, обладала наиболее выраженным общественным поведением, что дало ей серьезные преимущества в борьбе за существование. Во-первых, это совместная оборона от зверей и, одновременно, более удачливая охота. Во-вторых, передача опыта и быстрейшее научение молодняка, развитие средств общения. В-третьих, лучшая забота о потомстве на основе разделения функций в стаде.
Как следствие
этих новых возможностей происходит быстрое развитие мозга, психики, возникновение речи и понятийного (абстрактного) мышления
, отсутствовавших у обезьян. Обезьяны, как и многие другие животные, обладают предметным (конкретным) мышлением, основанным на восприятии предмета как такового через органы чувств. Человек может узнать предмет не видя его, по понятиям (большое, черное, лохматое - это медведь). Становится возможным использование и изготовление предметов - орудий защиты и охоты, а потом и труда. Совершенствуется вся сфера социальных отношений
.
Таким образом, главные отличия человека от обезьян
, сделавшие его таковым, состоят в следующем:
речь, понятийное (абстрактное) мышление, труд, изготовление и совершенствование орудий труда, развитие социальных отношений
. Появление речи и развитие психических функций коррелировали с изменениями объема и структуры головного мозга. У человека мозг составляет 1000-1800 мл, у антропоидов всего 600-700 мл, у обычных обезьян еще меньше. В составе мозга человека сильно развивается кора больших полушарий, особенно ее теменные, височные и лобные доли, ответственные за речь и психические функции. Соответственно, изменены и пропорции черепа: мозговая часть преобладает над лицевой, надбровные дуги разделены, развит подбородочный выступ, где крепятся мимические и речевые мышцы.
Рассмотрим теперь ступени развития человека
. Имеется очень много ископаемых останков древних людей и их предшественников. Современный уровень развития сравнительной анатомии позволяет восстанавливать скелет по его частям, внешний облик по скелету, образ жизни и тип питания по нескольким сохранившимся зубам. Окружающие находку другие ископаемые объекты и минеральные компоненты позволяют определить время жизни предмета вашего исследования. Так что летопись человека сегодня достаточно полная и надежная, хотя и в ней есть белые пятна и спорные вопросы. Общая схема филогении человека и его предшественников приведена на рис. 33.
Рис. 33
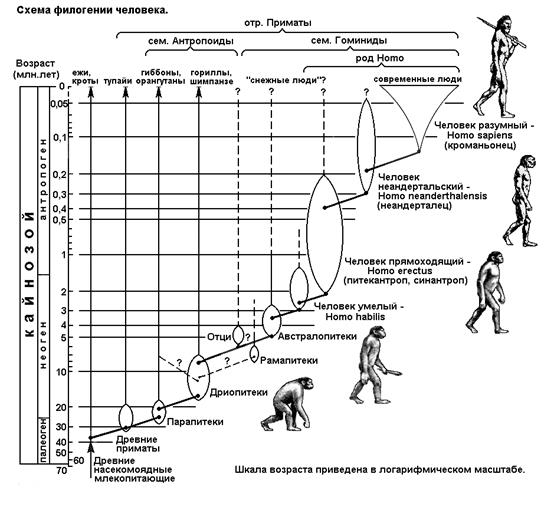
Ветвь древних приматов
зарождается от примитивных насекомоядных млекопитающих 35 млн лет тому назад, примерно в середине Кайнозойской эры. Большая часть древних приматов вымерла, но некоторые группы сохранились и даже получили прогрессивное развитие. Из современных приматов к их древним формам наиболее близки тупайи.
Около 30 млн лет назад состоялось отделение ветви парапитеков
- древесных обезьян, имевших смешанную пишу из растений и насекомых. Челюсти и зубы у них были уже как у человекообразных обезьян. Парапитеки дали современных гиббонов и орангутанов, а также ветвь дриопитеков
, отделившуюся в Неогене 20 млн лет назад. Дриопитеки населяли тропические леса восточной Африки (современные Кения, Египет, Эфиопия), их остатки найдены также в западной Европе и южной Азии. Вели стадный, полудревесный-полуназемный образ жизни, имели хорошо развитые передние конечности, относительно большой головной мозг, бинокулярное зрение. Орудия труда не использовали. Большая часть видов вымерла к 8 млн лет.
Необходимо отметить, что еще в конце Палеогена - от 30 до 25 млн лет назад - прошло очередное похолодание
, после чего началось вытеснение тропических лесов к югу, формирование степей, саванн. К 10-8 млн лет этот процесс обрел критический для дриопитеков характер. Большая их часть вымерла, а наиболее приспособленным предстояло буквально судьбоносное расселение. Одна часть дриопитеков последовала за отступающими тропическими лесами
, с их влажным климатом и обильной зеленой массой. От них и остались современные нам гориллы и шимпанзе - наиболее продвинутые человекообразные обезьяны. Другие дриопитеки вышли на освоение открытых пространств
, что и поставило их на ноги и повело всю дальнейшую эволюцию по пути постепенного перехода к прямохождению и освобождения рук.
Возрастом около 5 млн лет датируются ископаемые находки в горах Тироля скелета обезьяночеловека, названного Отци
. Его строение близко формам современного человека, однако сильно скошенный лоб и оттопыренный большой палец на стопе (на задних конечностях) явно указывают на его недостаточный интеллект и древесный образ жизни. Вероятно, это была одна из попыток очеловечивания, но в условиях старой среды обитания закончившаяся неудачно.
В целом надо сказать, что на вероятный путь к человеку вступали многие популяции и виды обезьян, но погибали в борьбе за существование - и с хищниками, и с такими же обезьянами. Их естественный отбор шел на способность к трудовой деятельности - освобождение рук, владение орудиями и их изготовление.
Другой, вероятно параллельной, ветвью от дриопитеков 5 млн лет назад отделились австралопитеки
(дословно - южная обезьяна), впервые найденные в южной Африке. Теперь скелеты австралопитеков найдены по всей Африке. Они имели рост 120 -150 см, вес 20 - 50 кг, мозг объемом 650 мл, ходили на двух ногах довольно прямо, руки были свободны. Вели стадный образ жизни, занимались охотой и собирательством, были всеядны, начинали осваивать огонь. Использовали подручные предметы как орудия труда: камни, палки, кости, но изготавливать, видоизменять орудия не умели. Австралопитеков уже относят к семейству гоминид. Однако основная их группа вымерла около 1 млн лет назад, но какая-то ветвь, по-видимому, и стала прямым предшественником человека. Хотя существует мнение, что другие обезьянолюди, наподобие уже упомянутого Отци, были нашими предками. Тогда люди представляют независимую, параллельную с австралопитеками ветвь эволюции высших приматов.
Итак, около 3 млн лет назад от боковой прогрессивной ветви австралопитеков или от других подобных предшественников отделился род Homo - человек
. Всего в истории собственно человека условно выделяют три стадии: древнейшие люди, древние и современные. По биологической номенклатуре они делятся на 4 разных вида, существовавшие практически на одном ареале и сменявшие друг друга в результате острой конкурентной борьбы за существование.
Древнейшие люди (архантропы)
известны по нескольким находкам в разных местах земного шара, основные - в восточной Африке. Существовали очень долго параллельно с австралопитеками. Выделяют два вида, сменившие друг друга.
Homo habilis - человек умелый
. Жил 3 - 1,5 млн лет назад, соседствуя с австралопитеками. Рост 150 см, мозг 800 мл, зубы человеческого типа, первый палец стопы не оттопырен. Изготовлял примитивные орудия труда из гальки, вел стадный образ жизни. Был расселен по Африке, Средиземноморью, в Азии. Как создатель первой, хоть и примитивной (галечной), культуры человек умелый преодолел грань, отделяющую ископаемых человекообразных обезьян от древнейших людей
.
Homo erectus - человек прямоходящий
. Сменил человека умелого около 1,5 млн лет назад, хотя в это время еще существовали поздние австралопитеки. Последние находки датируются возрастом 200 тыс. лет. Встречался изолированными группами в Африке и Евразии. Известно несколько ископаемых форм человека прямоходящего, которые по строению относятся все же к одному виду. Это - питекантроп (дословно - обезьяночеловек), найденный в 1891 г. на острове Ява (Индонезия), и синантроп (китайский человек), поселения которого изучены в пещерах близ г. Пекина. Рост 160 см и больше, но формы еще не современные. Головной мозг 800 - 1400 мл, левое полушарие преобладает - значит развита праворукость. Совершенствовал каменные орудия труда, охотился, использовал огонь от лесных пожаров и умел поддерживать длительное время. Развиты мышление и примитивная речь, то есть понятийное общение как следствие трудовой деятельности. Хотя уже действует социальный фактор, но продолжается и биологическая эволюция. Естественный отбор идет на физические качества, прямохождение, развитие мозга.
Древние люди (палеоантропы)
жили в ледниковую эпоху - от 250 до 35 тыс. лет тому назад. Первые находки человека этого типа сделаны у реки Неандерталь в Германии в 1856 г., поэтому его и назвали неандертальцем. Специалисты относят этого древнего человека к новому биологическому виду - Homo neanderthalensis - человек неандертальский
. Был расселен по Африке, Европе, Азии. Рост 155-165 см, мозг до 1400 мл, есть подбородочный выступ - свидетель речевого общения. Использовал совершенные каменные орудия. Огонь не только поддерживали, но и умели добывать, что позволило выжить в условиях ледникового похолодания, использовать пещеры. Отмечается углубление социальных отношений: забота о ближних, передача опыта, совместный труд и охота. Продолжается и отбор на совершенствование физических характеристик, формы скелета, особенно черепа.
Современные, или новые, люди (неоантропы) - Homo sapiens - человек разумный
.
Эволюционировали от прогрессивной ветви ранних неандертальцев по разным оценкам от 200 до 50 тыс. лет тому назад. Ископаемые новые люди известны как кроманьонцы - по первой находке в гроте Кроманьон во Франции в 1868 г. Обособились как новый вид в Восточном Средиземноморье - Передней Азии, откуда расселились по всей Земле.
Кроманьонец имеет все физические свойства современного человека: рост 180 см, мозговая часть черепа больше лицевой, надглазничный валик разделен на две части (бровные дуги), развитый подбородочный выступ, развитые лобные доли мозга (речь и мышление). Создает и совершенствует кремниевые орудия труда, использует так же кость и рог. Характерна сложная трудовая деятельность. Используется одежда из шкур. Создает рисунки на скалах, в пещерах, украшения и культовые предметы в виде статуэток - зарождается современная культура. Биологическая эволюция завершилась, но продолжается эволюция социальная
.
Современная биологическая изменчивость внутри вида Homo sapiens выражена в форме человеческих рас
. Раса - категория систематическая, но это подразделения внутри единого полиморфного вида. Человеческие расы - результат расселения и географической изоляции разных популяций неоантропов. Расселение человечества совершалось постоянно. Первичное расселение охватывает период от 500 до 40 тыс. лет, вторичное состоялось около 10 тыс. лет назад. В ходе расселения происходили морфологические и функциональные адаптации к разным климатическим условиям.
В процессе миграции людей в трех генеральных направлениях сформировались и три основные расы:
1) европеоидная (евразийская) раса
- население Европы, южной Азии, северной Африки (до эпохи великих географических открытий и европейской колонизации, после чего началось смешение рас);
2) негроидная (австрало-негроидная; экваториальная) раса
- население центральной и южной Африки и Австралии;
3) монголоидная (азиатско-американская) раса
- коренное население центральной и восточной Азии, Сибири, Северной и Южной Америки.
Ни одна из рас не достигла видового обособления
, так как с самого начала шло постоянное смешение рас на краях ареалов разных популяций. По этой же причине возникло множество промежуточных (малых) рас и подрас - всего их насчитывают 22. (Человеческие расы по генетическому и морфологическому статусу подобны породам и сортам в селекции животных и растений). Все расы свободно скрещиваются, обладают одинаковым интеллектуальным потенциалом. Различия уровней культуры обусловлены социальными условиями развития общества.
ЗАКЛЮЧЕНИЕ К ТЕМЕ 5
Живой мир возник на планете Земля вскоре после ее формирования - около 3,5 млрд лет тому назад - в результате самоорганизации из неживых химических систем. С тех времен живые организмы в составе популяций, сообществ, биогеоценозов претерпевают необратимое историческое развитие - филогенез. Основными движущими силами биологической эволюции являются наследственная изменчивость и естественный отбор популяций организмов, наиболее приспособленных к меняющимся условиям среды. Биологическая эволюция совершается параллельно и взаимообусловленно с геологической перестройкой Земли.
Сформированное биоразнообразие включает эволюционно молодые группы, а также и группы древние, создавшие такие виды, которые были способны выиграть борьбу за существование с новыми, более сложными организмами и занять свободные экологические ниши. Поэтому на современной Земле благополучно сосуществуют и взаимодействуют в сложных биогеоценозах самые разные организмы - от бактерий и вирусов до высших растений и животных.
Человек тоже является продуктом биологической эволюции. Он относится к царству животных, типу хордовых, подтипу позвоночных, классу млекопитающих, отряду приматов, семейству людей, роду человек. Все человечество на Земле - единый биологический вид - Homo sapiens, представленный многочисленными расами и подрасами, возникшими как адаптивные модификации в ходе географического расселения вида.
Полагают, что биологическая эволюция человека прекратилась, так как ее ограничивает все более прогрессирующая социальная эволюция. Действительно, эволюция человечества сопровождалась постепенным сужением действия естественного отбора в силу возникновения и развития общественных законов и создания новой, «искусственной» среды обитания. Эволюция человека превратилась в эволюцию разума. Разум Человека стал частью Биосферы Земли и движущим фактором ее дальнейшего развития. В какую сторону?
РЕКОМЕНДУЕМАЯ ЛИТЕРАТУРА
Основная:
1. Биология для поступающих в вузы (под ред. В.Н. Ярыгина). М.: Высшая школа, 1995.
2. Грин Н., Стаут У., Тейлор Д. Биология: в 3 томах. М.: Мир, 1990.
3. Кэмп П., Армс К. Введение в биологию. М.: Мир, 1998.
4. Общая биология. Учебник для 10-11 классов с углубленным изучением биологии в школе (под ред. В.К. Шумного, Г.М. Дымшица, А.О. Рувинского). М.: Просвещение, 1995.
Дополнительная:
1. Биологический энциклопедический словарь (гл. ред. М.С.Гиляров). М.: Советская энциклопедия, 1986.
2. Горелов А.А. Концепции современного естествознания. М: Центр, 1997.
3. Заренков Н.А. Теоретическая биология. М.: МГУ, 1988.
4. Кузнецов В.И., Идлис Г.М., Гутина В.Н. Естествознание. М., 1996.
5. Медников Б.М. Аксиомы биологии. М.: Знание, 1982.
6. Потеев М.И. Концепции современного естествознания. С.-Петербург: Питер, 1999.
7. Пригожин И.Р., Стенгерс И. Порядок из хаоса. М., 1986.
8. Рузавин Г.И. Концепции современного естествознания. М.: Культура и спорт, 1997.
9. Эткинс П. Порядок и беспорядок в природе. М.: Мир, 1987.
|
