Федеральное агентство по образованию
Государственное образовательное учреждение высшего профессионального образования
«Тамбовский государственный университет им. Г.Р. Державина»
Академия экономики и предпринимательства
Кафедра: Финансы
Специальность 080107 налоги и налогообложение
РЕФЕРАТ
по концепции современного естествознания
на тему: « Роль мутации в эволюции живого»
Выполнила:
Студентка 1 курса 107 группы
Юдина Татьяна Михайловна
Проверил:
Чиванов Андрей Викторович
Тамбов
2009
Содержание:
Стр.
1. Введение 3
2. Развитие учения об эволюции и современные взгляды на нее.
2.1.Развитие учения об эволюции. 4
2.2.Синтетическая теория эволюции. 8
3.
Эволюционные процессы.
3.1. Мутации, их типы и принципы классификации. 9
3.2. Давление отбора на процессы преобразования генетических структур в популяциях. 13
3.3. Принципы динамического равновесия между мутационным процессом и естественным отбором. 15
3.4. Генотипическая изоляция и адаптация – один из путей видообразования. 16
4.
Методологическая часть.
4.1. Искусственный (индуцированный) мутагенез. 18
4.2. Генетическая токсикология. 22
5. Выводы 25
6.Список использованной литературы 27
1. Введение
Наука делает грандиозные шаги вперёд. Появляются и получают развитие многие её отрасли. Одна из таких отраслей, которая заслуживает внимания-генетика. Проблема данного исследования до сих пор носит актуальный характер и представляет научный и практический интерес. Актуальность ее не подлежит сомнению, так как людям необходимо знать что и как влияет на окружающий их мир. Это нужно для того, чтобы контролировать свое воздействие на природу, создавать новые виды животных и растений путем селекции.
Эта работа рассматривает перестройку генома организма под влиянием окружающей среды. Цель
этой работы
– выявить роль мутаций и окружающей среды в эволюции живого. Ее задачи
включают следующие пункты:
1. раскрыть развитие учения об эволюции;
2. изучить синтетическую теорию эволюции;
3. разобрать типы мутаций и принципы классификации;
4. исследовать давление отбора на процессы преобразования генетических структур в популяциях;
5. выявить принципы динамического равновесия между мутационным процессом и естественным отбором;
Реклама

6. проанализировать генотипическую изоляцию и адаптацию, как один из путей видообразования;
7. рассмотреть искусственный (индуцированный) мутагенез;
8. изучить генетическую токсикологию.
Гипотезой
в этой работе является предположение, что эволюция вида не может происходить только за счет мутационных процессов, она обусловлена множеством факторов.
Хотя данная проблема далеко не нова, она постоянно получает новую подпитку извне, так как генетика и связанные с ней биотехнологии являются одними из наиболее перспективных отраслей науки и горизонты их развития в ближайшем будущем необозримы.
2.
Развитие учения об эволюции
и современные взгляды на нее.
2.1. Развитие учения об эволюции.
Идея эволюции имеет очень древнюю историю: в разной форме мысли о последовательном появлении на Земле живых организмов с постепенно усложняющейся их организацией содержатся и в Книге Бытия, и в «Ригведе», и в произведениях философов древнего Китая и Греции. Истоки эволюционных воззрений восходят к глубокой древности. Философы Древней Греции и Древнего Рима (Демокрит, Анаксагор, Аристотель, Лукреций, Кар и др.) высказывали различные предположения о развитии и превращениях организмов, а также о движущих силах этих явлений. Но их выводы носили характер догадок и не опирались на конкретные научные факты.
В средние века господство религиозного догматизма и схоластики привело к проповедованию абсолютного постоянства природы: все виды, однажды появившиеся в результате божественного акта творения, навечно остаются неизменными.
В 15-18 в.в. в связи с великими географическими открытиями произошло накопление фактического материала о многообразии животного и растительного мира. Исследование такого многообразия способствовало развитию сравнительного метода, позволяющего изучать животные и растительные организмы по признакам сходства и различия. Это привело к становлению систематики и сравнительной анатомии, сыгравших огромную роль в формировании эволюционного учения.
Первую попытку создания целостного учения об эволюции живых существ принадлежит французскому зоологу Ж.Б.Ламарку. В начале своей долгой научной карьеры Ламарк занимался преимущественно ботаникой, а затем – зоологией. К эволюционным представлениям он пришел во второй половине жизни и изложил их в капитальном труде «Философия зоологии» (1809 г.). В основе эволюционных представлений Ламарка лежат три гипотетических принципа:
1) общее внутреннее стремление всего живого к совершенству;
Реклама

2) внутреннее ощущение определенных потребностей, приводящее к возникновению или совершенствованию надлежащих органов; упражнение усиливает развитие органов, а не упражнение приводит к деградации и исчезновению их;
3) передача по наследству признаков, приобретенных в индивидуальной жизни.
Теория Ламарка показалась его современникам слишком спекулятивной, неправдоподобной. Особенный скептицизм вызывал второй постулат. Как могли, например, бесхоботные предки слонов ощущать потребность
в хоботе? Как мог прародитель орхидей ощущать потребность в развитии того удивительного разнообразия строения и окраски цветов, которое свойственно орхидеям? В результате теория Ламарка встретила холодный прием и значительного распространения не получила1
.
Противоположный прием ожидал опубликованную через 50 лет после «Философии зоологии» Ламарка теорию Чарльза Дарвина. Она сразу нашла многочисленных горячих приверженцев и пропагандистов, буквально перевернув при этом мышление биологов (и не только их!). Успех теории Дарвина, конечно, нельзя объяснить только тем, что он собрал большое количество фактов из разных областей биологии, которые говорили в пользу признания эволюции. Подобных фактов достаточно много было известно и Ламарку, и даже до него. Признание эволюции задерживалось не из-за недостатка иллюстраций возможностей эволюции, а из-за отсутствия убедительного объяснения, каким образом эволюция осуществляется, какой механизм ее двигает. И только Дарвин выявил этот механизм.
Дарвиновский механизм элементарно прост: он сформулирован в заглавии его знаменитой книги «Происхождение видов путем естественного отбора, или сохранение благоприятствуемых рас в борьбе за жизнь» (под «благоприятствованием» Дарвин понимал большую приспособленность к условиям жизни). Согласно Дарвину, для того, чтобы эволюция происходила, нужна способность организмов производить избыток потомства, т. е. больше, чем может просуществовать на том же пространстве, где существуют родительские организмы, и, кроме того, потомство должно обладать известным разнообразием, прежде всего, в своем отношении к среде обитания.
При наличии этих двух предпосылок неизбежно будет из поколения в поколение будет происходить естественный отбор в пользу особей, обладающих лучшей приспособленностью к данным условиям. И если результаты отбора, произошедшего в каждом поколении, будут в какой-то степени наследоваться потомством, должна происходить эволюция. А так как приспособляться к условиям обитания можно разнообразнейшими путями и борьба за жизнь непременно заставит искать эти разные пути, то эволюция неизбежно должна носить не только приспособительный (адаптивный), но и дивергентный (расходящийся) характер.
Таким образом, выдвинув теория естественного отбора, Дарвин сразу «убил двух зайцев»: дал естественное, материальное объяснение не только возникновения и многообразия органического мира, но и целесообразности, свойственной этому миру. Идея естественного отбора – сердцевина дарвиновской теории, это самое главное, что Дарвин внес в биологию и что обозначило крупнейший поворотный пункт в развитии биологии.
Выдвигая свою теорию, Дарвин не представил прямых экспериментальных доказательств, подтверждающих ее; вся аргументация носила косвенный характер. Но в отличие от теории Ламарка дарвиновская
1Четвериков С. С. О некоторых моментах эволюционного процесса с точки зрения современной генетики // Классики современной генетики. М.: 1968.
теория не подразумевала никаких надуманных, отвлеченных, внематериальных принципов и поэтому для современников была если и не вполне доказанной, то во всяком случае вполне правдоподобной. Однако хотя Дарвин и сказал самое главное, он не мог раскрыть всего, что нужно для построения цельной, достаточной и внутренне не противоречивой теории. Достройка теории затянулась еще более чем на полвека.
Первое серьезное затруднение, которое должна была преодолеть теория, состояло в следующем. Допустим, среди множества особей, принадлежащих к какому-либо одному виду животных или растений, наметилась дивергенция. Но ведь, принадлежа к одному виду, разные особи между собой будут скрещиваться, и уже в ближайших следующих поколениях вся дивергенция сойдет на нет. Это затруднение удалось преодолеть, признав в качестве необходимого условия эволюционной дивергенции изоляцию. Чтобы дивергенция сохранилась и нарастала далее, необходима изоляция дивергирующих групп друг от друга. А из этого в свою очередь следует, что дивергентную эволюцию могут проделывать только целые группы совместно существующих особей (т. е. то, что мы теперь называем популяциями). Значение изоляции как необходимого фактора эволюции стало ясным особенно в результате работ немецкого зоолога М. Вагнера.
Второе (в исторической последовательности) затруднение, преодоленное теорией, было в основном логического порядка. В эпоху Дарвина имела общее распространение уверенность в так называемом наследовании приобретенных признаков, т. е. передаче потомству свойств, приобретенных родителями в течение их индивидуальной жизни в качестве реакции на воздействие различных внешних факторов. Как правило, организмы реагируют на внешние воздействия целесообразно, приспособительно. А если эта приспособительность наследуется, то, следовательно, адаптивная эволюция возможна и без всякого естественного отбора. Отбор оказывается необязательным, а значит, и ненужным элементом теории. Эта логическая несообразность не была достаточно ясна ни самому Дарвину, ни его ближайшим последователям. И только биолог-мыслитель Август Вейсман в 80-х годах 19 в. понял, насколько эта несообразность нарушает стройность теории, и смог ее устранить. Логически проанализировав простые и общеизвестные биологические факты, Вейсман пришел к выводу о ненаследуемости приобретенных признаков.
Устранение затруднения с проблемой ненаследуемости признаков привело к другой трудности: если не наследуются изменения, приобретенные родителями, то откуда же вообще берется адаптивная наследственная изменчивость?
Дарвин отмечал, что, наряду с проявлениями индивидуальной изменчивости, возникающими как реакция на определенные воздействия в течение жизни одной особи и имеющие явно адаптивный характер («определенная изменчивость» по Дарвину, или модификация по современной терминологии), существует также изменчивость
«неопределенная», в которой непосредственно адаптивного смысла усмотреть не получается. Это то, что уже давно в животноводстве и особенно в садоводстве под названием «спортов» (появляющиеся без какой-либо видимой причины стойкие наследственные уклонения непредсказуемого характера). Дарвин сильно колебался в оценке значения «неопределенной» изменчивости для эволюции, но так и не решился сделать на нее основную ставку.
На рубеже 19-20 вв. проявления неопределенной изменчивости стали предметом усиленного внимания исследователей и получили название «мутации». С. И. Коржинский и голландец Г. Де Фрис предположили, что только мутации определяют и делают эволюцию, а естественный отбор если и имеет какое-либо значение, то лишь консервативное, охраняющее некоторую норму. Предполагалось, что мутация может дать сразу начало новому виду. Народившаяся в начале 20 в. генетика также стала в оппозицию дарвиновскому представлению о роли отбора. Важнейшие достижения раннего этапа развития генетики – установление стабильности генов и выявление того факта, что признаки гомозиготной чистой линии никакой отбор не в силах изменить – поначалу истолковывались как несовместимые с дарвиновской теорией. Казалось, дарвинизм зашел в тупик и вот-вот будет окончательно отброшен2.
Однако в начале 20 века полечила развитие не только генетика. Набирает силу новое научное направление – экология. Принципиальный прогресс был достигнут и в систематике. На смену господствовавшему в 19 в. представлению, что единство вида заключается в единообразии составляющих его организмов и, следовательно, вид есть некоторый морфофункциональный тип (типологическая концепция вида), пришло представление о виде как о реальном множестве особей, единство которого заключается в специфичности занимаемой им экологической ниши и в генетическом родстве между особями (популяционная концепция вида).
Вполне понятно, что ни натуралисту-экологу, ни натуралисту-систематику, стоявшему на позициях популяционной концепции вида, никак не могла импонировать замена дарвиновских представлений об адаптивном характере эволюции на лишенные всякого экологизма и по существу типологические идеи мутационной теории. Для натуралиста мутационная теория выглядели тупиком, вряд ли намного более привлекательным, нежели идея божественного творения.
Тупик на одном направлении, тупик на другом… Но выход был найден в лице синтетической теории эволюции.
______________________________________________________
2Вяткин Ю. С., Журавлев В. Б., Киселев В. Д. Эволюционная теория Дарвина и современность // На сайте Алтайского государственного университета (www.asu.ru)
2.2. Синтетическая теория
эволюции.
Взгляды родоначальников генетики на движущие факторы эволюционного
процесса, в особенности на изменчивость и наследственность, существенно отличны от воззрений Ламарка и Дарвина.
Тем не менее в 30-50-е годы трудами Добжанского, Симпсона, Майра и других была предпринята попытка примирить генетику с дарвинизмом. Так появилась синтетическая теория эволюции, создатели которой стремились обобщить все накопленные генетикой факты и в приложении к эволюционному учению объединить их с позиций Дарвина.
Объединение дарвинизма и генетики было сделано Г. Харди и В. Вайнбергом, которые в 1908 г. показали, что в популяции при свободном скрещивании, отсутствии мутаций данного гена и отбора по данному признаку соотношение генотипов АА, Аа и аа остается постоянным. Это позволило им сформулировать закон, который стал основой построения синтетической теории эволюции. Содержание закона сводится к тому, что частоты встречаемости генов в бесконечно большой панмиктической популяции (свободное скрещивание особей в пределах популяции) без давления со стороны каких-либо внешних факторов стабилизируются уже после одной смены поколений. Однако бесконечно больших панмиктических популяций, как известно в природе не существует. Поэтому значение закона Харди-Вайнберга заключается в том, что накопленные наследственные изменения в генофонде популяций бесследно не исчезают, т. е. частоты встречаемости генов постоянны.
В процессе следствия закона Харди-Вайнберга углублялись и конкретизировались. Так, при рассмотрении роли изменчивости в эволюционном процессе, что именно исходя из закона Харди-Вайнберга отечественный генетик С.С. Четвериков в 1926 г., исходя из этого закона и учитывая влияние отбора и возникновение новых мутаций, показал, что в результате спонтанного мутационного процесса во всех популяциях создается гетерогенность. Другими словами, в популяции всегда есть мутации. С. С. Четвериков первым продемонстрировал, что популяция насыщена мутациями, как губка, и что мутации служат основой (материалом) эволюционного процесса, идущего под действием естественного отбора. Однако, когда популяции малы, в них возникают процессы, получившие название генетико-автоматических, т. е. в малых популяциях происходит дрейф генов, в результате которого изменяются частоты встречаемости генов, устраняются гетерозиготыи появляются гомозиготы. Изолированная популяция становится доминантной гомозиготной или рецессивной гомозиготной.
Если дрейфует мутантный летальный ген, это ведет к вымиранию организмов. Таким образом, структура популяции зависит не только от возникновения новых мутаций, но и от простого изменения частоты встречаемости данного гена.
Современное эволюционное учения представляет собой синтез достижений дарвинизма, физико-химической биологии, генетики, систематики, морфологии, физиологии и других наук. В наше время эволюционное учение, основу которого составляет дарвинизм, – это наука об общих законах развития органической природы. Эволюционное учение стало методологической базой всех специальных биологических дисциплин3.
3.
Эволюционные процессы.
3.1. Мутации, их типы и принципы классификации.
Действие мутагенов на наследственные структуры клеточного ядра неодинаково, поэтому возникают различные мутации.
Мутации
(от лат. mutatio — изменение, перемена), возникающие естественные (спонтанные) или вызываемые искусственно (индуцированные) стойкие изменения наследственных структур живой материи, ответственных за хранение и передачу генетической информации. Мутация-основа наследственной изменчивости в живой природе.
Существует несколько классификаций мутации.
Классификация по изменению генотипа.
По характеру изменения генетического аппарата мутации делят на геномные, хромосомные и генные, или точковые.
Геномные
: — полиплоидизация (образование организмов или клеток, геном которых представлен более чем двумя (3n, 4n, 6n и т. д.) наборами хромосом) и анеуплоидия (гетероплоидия) — изменение числа хромосом, не кратное гаплоидному набору. В зависимости от происхождения хромосомных наборов среди полиплоидов различают аллополиплоидов, у которых имеются наборы хромосом, полученные при гибридизации от разных видов, и аутополиплоидов, у которых происходит увеличение числа наборов хромосом собственного генома, кратное n.
При хромосомных мутациях
происходят крупные перестройки структуры отдельных хромосом. В этом случае наблюдаются потеря (делеция) или удвоение части (дупликация) генетического материала одной или нескольких хромосом, изменение ориентации сегментов хромосом в отдельных хромосомах (инверсия), а также перенос части генетического материала с одной хромосомы на другую (транслокация) (крайний случай — объединение целых хромосом, т. н. Робертсоновская транслокация, которая является переходным вариантом от хромосомной мутации к геномной).
Воронцов Н. Н. Синтетическая теория эволюции: ее источники, основные постулаты и нерешенные проблемы // Журн. Всес. хим. о-ва им. Д. И. Менделеева. 1980. Т. 25. N 3. С. 293—312.
На генном
уровне изменения первичной структуры ДНК генов под действием мутаций менее значительны, чем при хромосомных мутациях, однако генные мутации встречаются более часто. В результате генных мутаций происходят замены, делении и вставки одного или нескольких нуклеотидов, транслокации, дупликации и инверсии различных частей гена. В том случае, когда под действием мутации изменяется лишь один нуклеотид, говорят о точковых мутациях. Поскольку в состав ДНК входят азотистые основания только двух типов — пурины и пиримидины, все точковые мутации с заменой оснований разделяют на два класса: транзиции (замена пурина на пурин или пиримидина на пиримидин) и трансверсии (замена пурина на пиримидин или наоборот). Возможны четыре генетических последствия точковых мутаций: 1) сохранение смысла кодона из-за вырожденности генетического кода (синонимическая замена нуклеотида), 2) изменение смысла кодона, приводящее к замене аминокислоты в соответствующем месте полипептидной цепи (миссенс-мутация), 3) образование бессмысленного кодона с преждевременной терминацией (нонсенс-мутация). В генетическом коде имеются три бессмысленных кодона: амбер — UAG, охр — UAA и опал — UGA (в соответствии с этим получают название и мутации, приводящие к образованию бессмысленных триплетов — например амбер-мутация), 4)обратная замена (стоп-кодона на смысловой кодон).
Классификация мутаций по их действию и влиянию на организм.
Действие, проявление и влияние мутаций на организм очень многообразны.
По действию мутации делят на морфологические, физиологические
и биохимические.
Они могут изменять проявление любого внешнего признака, влиять на функции отдельных органов, рост и развитие организма, вызывать различные изменения химического состава клеток и тканей и т. д.
По относительному влиянию на жизнеспособность и плодовитость организма мутации делятся на полезные, нейтральные
и вредные
.
Полезные мутации повышают устойчивость организма к неблагоприятным внешним условиям, вредные тормозят нормальный ход жизненных процессов, понижают жизнеспособность организмов. К вредным относят летальные мутации, обычно вызывающие гибель организма. Они могут быть и доминантными и рецессивными. Доминантные летальные мутации в результате непосредственного проявления быстро удаляются естественным отбором, а рецессивные могут накапливаться в генотипе и проявляться в последующих поколениях.
Совокупность всех мутаций, возникающих у организма под действием определенного мутагена, называют спектром мутаций. При большом их разнообразии говорят о широком спектре; однотипные мутации характеризуются узким спектром.
Генеративные и соматические мутации.
Мутационная изменчивость происходит на различных этапах развития организма и во всех его клетках. Мутации, возникающие в гаметах и клетках, из которых они образуются, называются генеративными. Мутации, происходящие в соматических клетках организма, называются соматическими. По своей природе генеративные и соматические мутации ничем не отличаются. И те, и другие связаны с изменением структуры хромосом, по характеру проявления и значимости для эволюции и селекции различия между этими видами мутаций очень существенны.
Генеративные мутации при половом размножении передаются следующим поколениям организмов. Доминантные мутации проявляются уже в первом поколении, а рецессивные – только во втором и последующих поколениях, при переходе их в гомозиготное состояние. Соматические мутации возникают в диплоидных клетках, поэтому проявляются только по доминантным генам или по рецессивным генам в гомозиготном состоянии.Они имеют большое значение для эволюции организмов, у которых возможно вегетативное размножение.
Очень многие растения, например плодовые и ягодные культуры, размножаются вегетативным путем. У них любая соматическая мутация, возникшая в тканях, из которых может развиться новое растение, будет передана дальнейшим поколениям. У плодовых растений хорошо изучены мутации, происходящие в клетках точек роста, так называемые почковые мутации.
Первый, созданный в 1888 г. И. В. Мичуриным сорт яблони Антоновка шестисотграммовая ведет свое начало от почковой мутации, обнаруженной у сорта Антоновка могилевская белая.
По причине возникновения мутации делятся на спонтанные и индуцированные
. Спонтанными
называют мутации, появление которых не контролируется человеком. Строго говоря, назвать их случайными нельзя, так что формулировка "неконтролируемые человеком" корректнее. Как уже отмечалось выше, мутагенез бывает управляемым внутри- и трансклеточным самомутагенезом, программа которого заложена в первичной структуре ДНК (он опосредован центриолями и МГЭ), и истинно случайным, например, действие УФ-излучения или жестких рентгеновских лучей.
Причины индукции спонтанных мутаций не ясны. Долгое время полагали, что к числу индуцирующих факторов относится естественный фон ионизирующих облучений, образуемый доходящими до поверхности земли космическими лучами..., поступающими в малых количествах в организм из окружающей среды. Однако, как показали расчеты, у дрозофилы естественный радиационный фон может быть ответственен только за 0.1% спонтанных мутаций. Хотя, по мере увеличения продолжительности жизни организма воздействие естественного фона может накапливаться, и у
Второй причиной спонтанных мутаций являются случайные повреждения хромосом и генов в ходе нормальных метаболических (обменных) процессов, происходящих в клетке. По многочисленным данным спонтанные мутации возникают во время деления хромосом и репликации ДНК. Считают вероятным, что спонтанные мутации представляют чаще всего следствие случайных ошибок в функционировании молекулярных механизмов.
Третьей причиной спонтанных мутаций является перемещение по геному мобильных элементов, которые могут внедриться в любой ген и вызвать в нем мутацию. По расчетам американского генетика Мелвина Грина около 80% спонтанных мутаций приходится на счет перемещений мобильных элементов.
Под индуцированными мутациями понимают мутации, вызванные искусственно, сознательными действиями человека. В отличии от управляемого мутагенеза (центриоли и МГЭ), в этом случае, по видимому, вероятность замены одного нуклеотида на другой не зависит от его локализации в молекуле ДНК. Это случай так называемого неуправляемого мутагенеза. Мутагенов весьма много: жесткие рентгеновские лучи, УФ и ИК-лучи, иприт, этиловый спирт, никотин, наркотики, гормоны, вырабатываемые при стрессе и т. д. 4
Итальянские учёные в течение 3 лет проводили опыты на животных и выяснили, что кокаин и экстази не только вызывают наркозависимоть, повышают риск возникновения рака, но и вызывают генетические мутации.
"Оказывается, экстази и кокаин более опасны, чем мы думали, — заявил Джорджио Бронзетти (Giorgio Bronzetti), главный учёный Национального центра биотехнологических исследований (Consiglio Nazionale delle Ricerche — CNR). — Эти наркотики на вершине их токсикологического воздействия атакуют ДНК, провоцируя мутации и изменения генетического материала".
Строго говоря, грань между индуцированными и спонтанными мутациями условна. Все живые организмы в какой-то мере подвергаются действию мутагенов, например, получают небольшие дозы природного (солнечного) УФ-излучения, вдыхают радон, что, по моему мнению, является одной из причин деградации первичной структуры ДНК, и, как следствие, старения.
Изучение мутагенного действия ионизирующих излучений показало, что у всех исследованных организмов они вызывают многочисленные генные мутации и перестройки хромосом и что частота индуцированных мутаций зависит в основном от дозы радиации.
При этом не имеет большого значения, в один ли прием дана та или иная доза или она разбита на дробные порции, разделенные во времени - мутагенный эффект в целом соответствует общей дозе облучения.
_____________________________________________
4
Гершензон, 1983, с. 237
Комментарии - указанный эффект означает, например, что люди, получившие большую дозу облучения на Чернобыльской АЭС, и погибшие вследствие этого, и люди, умершие "естественной" смертью в преклонном возрасте, умерли от одной и той же причины - необратимого накопления мутаций первичной структуры молекул ДНК их клеток. Разница состоит лишь в том, что люди, облучившиеся на Чернобыльской АЭС, получили за несколько часов ту дозу, которую остальные получают за несколько десятков лет. Это грубый пример, не учитывающий управляемый самомутагенез, тем не менее, хорошо отражает качественную сторону явления.
В частности, это означает, что естественной смерти не бывает. Смерть от старости естественна не более, чем смерть от ДДП.
3.2. Давление отбора на процессы
преобразования генетических
структур в популяциях.
Мутационный процесс неустанно ведет к возникновению широкого спектра внутрипопуляционной наследственной изменчивости. В этом спектре важнейшее место занимают вредные мутации вплоть до леталей, отметаемых отбором. Вместе с тем естественный отбор, опираясь на полезные мутации, перестраивает наследственные структуры популяций и видов и одновременно сохраняет наследственные особенности вида от разрушающего действия мутаций.
Дарвиновская теория естественного отбора основана на том принципе, что количество оставляемых потомков зависит от качественных особенностей родителей. Генетическая теория естественного отбора также признает, что организмы разных наследственных типов в популяции имеют дифференциальное переживание и оставляют различное количество потомков. Для изучения интенсивности отбора введено понятие о коэффициенте селекции s
, который измеряет для того или иного наследственного типа в популяции данного поколения степень преимущественности или неуспеха при оставлении потомков следующего поколения. Предположим, что в популяции генотипы с
оставляют одинаковое количество потомков. В то же время гомозиготы по рецессивному аллелю аа
испытывают давление отрицательного отбора с интенсивностью s
=0,01, т. е. вместо ожидаемой 1000 они составляют 999 потомков. В таких условиях частота аллеля А
в популяции будет возрастать, а концентрация аллеля а
будет снижаться в процессе смены поколений. Для генотипов АА
и Аа
можно сказать, что их адаптивная ценность равна 1, в то время как для генотипа аа
она равна 0,999. В случае одинаковой жизнеспособности, т. е. когда адаптивная ценность генотипов АА, Аа
и аа
равна 1, коэффициент селекции равен 0. В случае стерильности или летальности гомозигот аа
их адаптивная ценность равна 0, а коэффициент селекции равен 1.
Процесс отбора наиболее эффективен, когда он направлен против доминантных мутаций, при условии их полного проявления и выражения. В этих случаях при коэффициенте селекции, равном 1, популяция может избавиться от доминантных изменений всего лишь за одно поколение. Так, у человека имеется немало доминантных наследственных заболеваний, которые не позволяют из носителям иметь детей. Это в значительной мере касается, например, заболевания ахондроплазии (карликовость). Все мутантные особи этого рода оказывают очень малое влияние на генотипический состав популяции человека в следующем поколении.
В тех случаях, когда проявление вредного доминантного гена запаздывает и его носители успевают оставить потомство, они могут оказывать значительное неблагоприятное влияние на состав будущего поколения, полностью передавая наследственное заболевание своим потомкам. Для такого рода болезней возможно применение генетической профилактики. Зная наследствование этих заболеваний и зная трагическую участь детей, можно советовать отказаться от их рождения.
Отбор против рецессивных мутаций оказывается более трудным делом. Рассмотрим пример с аллелем альбинизма у человека. По статистическим данным, в Великобритании среди 20000 детей рождается один альбинос (0,00005). Концентрация аллеля альбинизма, следовательно, равна 0,007. Концентрация гена нормальной пигментации в этой популяции составляет 0,993. Эти данные позволяют рассчитать число гетерозигот, которое равно 1,38%. В такой популяции число гетерозигот (Аа
) в 276 раз больше числа гомозигот.
Таким образом, характерным различием между отбором против доминантов и против рецессивов служит то, что в первом случае все аллели подвержены отбору, во втором только их небольшая часть. Для рецессивных мутаций существует обширная зона гетерозигот, пребывая в которой они ускользают от действия отбора. Поскольку зона гетерозигот гораздо шире зоны гомозигот по рецессивам, эффект отбора в этом случае оказывается очень затруднительным.
Рассмотрим наиболее благоприятный случай отрицательного отбора, когда коэффициент селекции равен 1, т. е. гомозиготы или гибнут или оказываются полностью стерильными. Возьмем в качестве исходной популяцию, где концентрация рецессивного аллеля составляет 0,5. Очевидно, что распределение генотипов в такой популяции будет иметь вид 25%АА+
50%с+
25%аа.
Гетерозиготы (Аа)
в этом случае встречаются в популяции в два раза чаще гомозигот (аа, АА)
.
При коэффициенте селекции, равном 1, потомство в такой популяции будет получаться только от особей АА
и Аа.
В табл. 1 показано, что такой отбор приведет во втором поколении к тому, что концентрация аллеля а
упадет до 33%, а распределение генотипов примет вид: 44,44% АА+
44,44% Аа+
11,12% аа.
В такой популяции число гетерозигот (Аа
) в 4 раза превышает число гомозигот (аа
).
Вначале действие отбора на такую популяцию, благодаря высокой исходной концентрации аллеля а,
осуществляется вполне эффективно. За 9 поколений отбора (табл. 1) концентрация аллеля а
уменьшается в 5 раз, с 50 до 10%. Однако возрастает зона недопустимости аллеля для отбора. На 9 поколении количество гетерозигот уже в 18 раз больше количества рецессивных гомозигот, в то время как в исходном поколении оно было больше всего лишь в два раза. Это резко затрудняет деятельность отбора. Так, для того чтобы уменьшить концентрацию аллеля а
в два раза (от 0,020 до 0,010), необходимо уже 50 поколений отбора. В последнем из рассматриваемых, сотом поколении, количество гетерозигот (Аа
) в 196 раз превышает число гомозигот (аа
).
Разительные различия в эффективности отбора против доминантных мутаций и против рецессивных показаны на рис. 1. Как уже указывалось выше, при коэффициенте селекции, равном 1, все доминанты выбрасываются из популяции. При том же коэффициенте селекции против рецессивных гомозигот после 10 поколений отбора их число падает всего лишь в 4 раза. На рис. 1 показан ход падения числа гомозигот (аа
) по всем 10 поколениям отбора.
Рассматривая процессы отбора по изменениям концентраций отдельных генов, мы упрощаем процессы, идущие при отборе в природе и при искусственной селекции. Реальный процесс отбора идет на уровне генотипов в целом (особей), а не на уровне отдельных аллелей. Эффекты такой селекции, которая может быть оценена по количественным признакам, более сложны и редко модифицируются средой. В этих случаях эффективность отбора во многом определяется степенью наследуемости признака. Во многих случаях выраженность количественных признаков (продуктивность, урожайность и т. д.) зависит от положительного влияния внешней среды. Та компонента признака, которая вызвана внешней средой и является модификационной, не передается по наследству. Такие модификации могут маскировать генотипические свойства особи и этим тормозить эффективный отбор. На рис. 2 представлен результат отбора в потомстве двух фенотипически одинаковых популяций. На левой части рисунка дан ход селекции при полной наследуемости признака. Правая половина рисунка иллюстрирует такой же отбор, но при 20%-ной наследуемости признака. Во всех поколениях отбирались особи с максимальным проявлением признака. Мы видим, насколько различны итоги селекции даже в пределах всего лишь первых 4 поколений.
3.3. Принципы динамического равновесия
между мутационным процессом и
естественным отбором.
Изменения в генетических структурах популяций всегда происходят под влиянием сложного комплекса эволюционных факторов. Очень важное значение имеет соотношение отбора и давления мутаций. Если данный аллель поддерживается отбором, тогда носители этого аллеля, будучи более приспособленными, характеризуются преимущественным размножением. В результате отбор вытесняет все другие аллели. Однако, как показывает элементарная теория, в этих условиях, но при наличии мутаций отбор не в состоянии целиком утвердить в популяции благоприятный аллель, т. е. концентрация такого аллеля никогда не достигнет 100%. В такой популяции, где мутации противоборствуют отбору, концентрация данного аллеля установится на том или ином равновесном уровне, отражая взаимодействие обоих факторов.
При наличии отбора против постоянно возникающих доминантных аллелей равновесие устанавливается на уровне, равном u/
s
(p=
u/
s
). Для рецессивных аллелей состояние равновесия достигается на уровне q-4/
s.
*
У человека скорость мутирования по доминантному гену ахондроплазии (карликовости) u
=4.2´10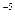 . Но, поскольку не все карлики доживают до репродуктивного возраста и оставляют потомство, аномальный ген передается следующему поколению лишь с невысокой частотой. В Дании, например, она составляет 0,125. Эффективность передачи нормального аллеля у непораженных сибсов соответствует 0,64. Следовательно, селективная ценность аллеля ахондроплазии равна 0,125; 0,64 или ~ 20%. Очевидно, что в этом случае коэффициент селекции равен 0,8 (s
= 0.8). . Но, поскольку не все карлики доживают до репродуктивного возраста и оставляют потомство, аномальный ген передается следующему поколению лишь с невысокой частотой. В Дании, например, она составляет 0,125. Эффективность передачи нормального аллеля у непораженных сибсов соответствует 0,64. Следовательно, селективная ценность аллеля ахондроплазии равна 0,125; 0,64 или ~ 20%. Очевидно, что в этом случае коэффициент селекции равен 0,8 (s
= 0.8).
В каждой популяции содержится два класса генных мутаций. Одни из них – это мутации, заново появившиеся в этом поколении. Вторые – мутации, перешедшие из предыдущего или предыдущих поколений. Для гена А (ахондроплазия) 0,2 от u
(0.2u
) мутантов являются переданными из предыдущего поколения. Количество новых мутантов равно единице. В целом в популяции следует ожидать число гетерозигот (Аа
), равное 1,2и
. В каждом следующем поколении число гетерозигот должно возрастать за счет увеличения доли, передаваемой из предыдущего поколения. Однако это может происходить только до тех пор, пока число гибнущих при отборе генов не станет равным числу возмещения мутаций. Так возникнет генетическое равновесие.
3.4.Генотипическая изоляция и адаптация –
один из путей видообразования
.
Конечно, внешняя среда играет важную роль в биологической дифференциации. Можно даже сказать, что если бы среда повсюду была совершенно одинаковой, то Земля, вероятно, была бы населена одним-единственным видом живых организмов. Однако, поскольку среда чрезвычайно разнообразна как в отношении климата, так и в отношении условий обитания, то это вызвало дифференциацию организмов и их приспособление к различной и изменяющейся среде.
Эта дифференциация подразумевает не только индивидуальное приспособление к среде, но также генотипическое приспособление, которое прежне всего осуществляется путем рекомбинации генов. Разделение на два разных пола и многие другие специальные механизмы обеспечивают постоянное поддержание рекомбинационной изменчивости.
Другое важное достижение генетики – установление того, что наряду с выраженным постоянством генов каждому организму свойственна определенная частота спонтанных мутаций. Новые гены или аллели возникают непрерывно и служат материалом для рекомбинаций.
Очень многие из мутантных генов не имеют значения для эволюции, т. к. их носители погибают в борьбу за существование. Однако некоторые из них оказываются благоприятными в том или ином отношении и могут после рекомбинаций дать организму преимущество для приспособления к новым условиям окружающей среды. Приспособление может состоять в том, что вид окажется в состоянии расширить область своего распространения, или же в том, что ареал вида сохраняет свои прежние границы, но вид становится выжить при измененных условиях среды, например при ухудшении климата. Этот процесс приспособления прежде всего связан с дифференциацией экотипов внутри видов. Настоящее образование новых видов начнется только в том случае, когда эта экологическая дифференциация будет сопровождаться возникновением преград между различными группами биотипов.
В первую очередь такими преградами служат всевозможные препятствия к скрещиванию, которые затрудняют или делают невозможным образование гибридных форм. Кроме того, если гибриды все же образуются, то они оказываются частично или полностью стерильными.
Для возникновения преграды между видами имеет первостепенное значение, чтобы часть популяции данного вида оказались тем или иным способом изолированной от основной массы вида. После того как это произойдет, дифференциация начнется почти что автоматически. Эта дифференциация начнется хотя бы потому, что в обособившейся группе по чистой случайности окажутся не все те гены, которые имеются в исходной популяции. Кроме того, обособившаяся группа обычно располагается на границе ареала исходного вида, где условия отличаются от условий в центре ареала. Поэтому отделившаяся группа с самого начала представляет собой известного рода экотип, который в результате постепенного возникновения биологической изоляции становится настоящим видом.
Таким образом, мы подходим к важному заключению, что видообразование – медленный процесс, по существу аналогичный процессу дифференциации генотипов в пределах вида. Поэтому становится понятным, почему в природе мы часто встречаем различные группы биотипов, которые так сильно обособились друг от друга, что бывает трудно решить, представляют ли они собой отдельные виды, подвиды или разные экотипы одного и того же вида.
4
. Методологическая часть.
4.1.Искусственный (индуцированный) мутагенез.
Природа мутаций, причины их появления почти четверть века после провозглашения Де-Фризом мутационной теории оставались загадочными. Считалось, что мутации происходят под влиянием неизвестных внутренних причин, заложенных в самой природе организмов. Появление мутаций сравнивалось с процессом самопроизвольного распада атомов радия. Это извращало действительную природу мутационного процесса и закрывало пути к разработке методов управления наследственной изменчивостью организмов. Но в 1925 г. ученые Ленинградского радиевого института Г. П. Надсон и Г. С. Филиппов впервые в мире получили мутации у дрожжевых грибов под влиянием лучей радия. Через два года, в 1927 г., американский генетик Г. Мёллер опубликовал работу «Искусственные трансмутации гена», в которой сообщалось о большом повышении частоты мутаций у дрозофилы при облучении ее лучами Рентгена. В 1928 г. В США Л. Стадлер получил рентгеномутации у ячменя и кукурузы.
Эти открытия доказывали, что наследственные изменения – мутации у растений, животных и микроорганизмов – можно вызвать в эксперименте, воздействуя внешними условиями. Тем самым устанавливались причины возникновения мутаций и открывались возможности для получения нужных наследственных изменений. Процесс возникновения мутаций – мутагенез* становится одной из важнейших проблем генетики.
Как в природе, так и в опытах мутации возникают под влиянием различных воздействий, называемых мутагенными факторами,
или мутагенами.
Применяемые для искусственного получения мутаций мутагены делятся на физические и химические. К физическим мутагенам относятся: радиация, высокая и низкая температура, механические воздействия, ультразвук. В качестве химических мутагенов используют различные органические и неорганические соединения. В данной работе я остановлюсь на более подробном рассмотрении физических мутагенов.
Физические мутагены.
К ним относятся радиационные излучения, все виды которых можно разделить на две категории: электромагнитные, или волновые, и корпускулярные.
Электромагнитные излучения
возникают в результате перехода электронов с орбиты на орбиту в пределах внешней оболочки атома (ультрафиолетовые лучи) или перемещения электронов между внутренними оболочками или внутренними и внешними оболочками (лучи Рентгена и гамма-излучение). Лучи Рентгена имеют длину волны 0,000005—0,001 мкм, гамма-излучение — менее 0,000005 мкм, и поэтому проникающая способность их выше. Электромагнитные излучения представляют собой
дискретные частицы—фотоны высокой энергии, распространяющиеся со скоростью света (300000км/с).
Лучи Рентгена
получают путем торможения быстрых электронов в аноде рентгеновской трубки. Для облучения используют медицинские или специальные промышленные рентгеновские аппараты. Для гамма-излучения
используют специальные передвижные или стационарные установки. В качестве источника радиации в них чаще всего используют изотопы 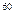 Co или Co или 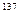 Сs. При больших масштабах работы и необходимости изучения действия гамма-излучения в разные фазы роста растений создают так называемые гамма-поля. Сs. При больших масштабах работы и необходимости изучения действия гамма-излучения в разные фазы роста растений создают так называемые гамма-поля.
Гамма-поле (рис.3) представляет собой участок чаще всего в форме окружности, в центре которого в стальном передвижном цилиндре помещается источник излучения Co (кобальтовая пушка) с дистанционным управлением. При включении установки источник облучения поднимается над поверхностью, при выключении – опускается глубоко под землю. Интенсивность излучения регулируется величиной расстояния объекта от источника излучения.
Ультрафиолетовые лучи
относят к электромагнитным колебаниям, но ионизации они не производят, их действие на организм связано с образованием в облученных тканях возбужденных молекул и атомов. Возбуждение представляет собой процесс поглощения энергии, сопровождающийся перемещением электронов с ближней на более далекую от ядра атома оболочку. Наиболее эффективен этот вид излучения в отношении облучения пыльцы, бактерий и спор грибов. В качестве источника ультрафиолетовых лучей для получения мутаций обычно используют ртутные лампы.
Корпускулярные излучения
распространяются со скоростью меньшей, чем скорость света. Они возникают в результате естественной и искусственной радиоактивности (a-частицы, электроны - b-частицы, протоны, дейтроны, нейтроны).
Для облучения нейтронами используют специальные камеры ядерных реакторов. Так как нейтроны лишены заряда и поэтому не взаимодействуют с электронной оболочкой атомов, их воздействие на живую материю носит более сложный характер. Обладая огромной энергией, они проникают в ядра атомов вещества и выбивают из них положительно заряженные частицы – протоны (ядра атомов водорода), которые являются сильно ионизирующими частицами: благодаря большой массе они обеспечивают глубоко проникающую и очень плотную ионизацию. Она приблизительно в 25 раз больше, чем у лучей Рентгена и гамма-лучей.
Дозы излучения и поглощения.
Действие радиации на живые организмы определяется количеством энергии, поглощаемой клетками облучаемых тканей. Поэтому необходимо измерять дозу излучения, падающую на облучаемый объект, и дозу энергии, поглощенной тканями организма. Величина их различна. Доза излучения измеряется величиной ионизации воздуха. Единица дозы облучения
– один рентген (Р). Рентген
– количество излучения, вызывающее образование в 1 см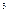 сухого воздуха (0,001293 г) при 0 сухого воздуха (0,001293 г) при 0 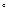 C и давлении 760 мм ртутного столба около двух миллиардов (2,1*10 C и давлении 760 мм ртутного столба около двух миллиардов (2,1*10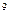 ) пар ионов. ) пар ионов.
Единица поглощенной дозы
– рад.
Один рад поглощенной радиации вызывает образование в 1 мкм
ткани в среднем около двух ионизированных и двух возбужденных молекул.
Действие ионизирующей радиации на живые организмы.
Излучения, электромагнитные (лучи Рентгена и гамма-лучи) и корпускулярных частиц (протоны, нейтроны и др.), попадая в ткани организма, теряют свою энергию, а ткани организма претерпевают сложные изменения. Основным первичным физическим процессом такого взаимодействия являются ионизация и образование возбужденных атомов и молекул, заключающиеся в том, что квант энергии электромагнитных излучений или ядерная частица вырывает электрон из внешней оболочки атома или молекулы. Потеряв электрон они становятся положительно заряженными ионами. Оторвавшийся электрон, несущий отрицательный заряд, присоединяется к другому атому или молекуле, которые также превращаются в отрицательно заряженные ионы. Так возникает пара ионов. Отсюда и название этой радиации – ионизирующая, т. е. вызывающая образование ионов при прохождении излучений через вещество. Пары ионов возникают на всем пути пробега кванта или частицы, и нейтральные атомы становятся заряженными.
Предполагается, что ионизирующие излучения могут действовать на наследственные структуры клеточного ядра двумя путями: непосредственно, ионизируя и возбуждая атомы и молекулы ДНК и белков, и через воздействие на них ионизированных молекул воды. И в том и в другом случае действие энергии излучений на хромосомы вызывает цепь радиационно-химических реакций, в результате которых изменяется нуклеотидный состав ДНК.
Прямое действие ионизирующих излучений.
Ионизирующие излучения могут непосредственно поражать в клетке молекулы нуклеиновых кислот и белков, которые будут претерпевать первичные изменения, связанные с ионизацией и возбуждением атомов и молекул. В ряде опытов на дрозофиле, ячмене и многих других объектах было показано, что число точковых рецессивных летальных мутаций и мелких разрывов хромосом возрастает прямо пропорционально дозе. Иными словами, зависимость частоты мутаций от дозы носит линейный характер.
Косвенное действие ионизирующих излучений.
Ионизации в живом организме прежде всего могут подвергаться молекулы воды – основного компонента цитоплазмы. Процесс разложения молекул воды на атомы кислорода и водорода с образованием свободных радикалов H и OH называют радиолизом воды *. Свободные радикалы существуют всего две десятимиллионные (2*10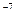 ) доли секунды, но они могут непосредственно реагировать с белками и нуклеиновыми кислотами, оказывая на них сильное действие и изменяя наследственные структуры. Очевидно, действие свободных радикалов на хромосомы может быть различным в зависимости от того, каким запасом энергии они обладают. Если энергия их мала, они вызывают преимущественно точковые (генные) мутации; когда они наделены большой энергией, происходят крупные перестройки хромосом, сопровождаемые множеством летальных мутаций. ) доли секунды, но они могут непосредственно реагировать с белками и нуклеиновыми кислотами, оказывая на них сильное действие и изменяя наследственные структуры. Очевидно, действие свободных радикалов на хромосомы может быть различным в зависимости от того, каким запасом энергии они обладают. Если энергия их мала, они вызывают преимущественно точковые (генные) мутации; когда они наделены большой энергией, происходят крупные перестройки хромосом, сопровождаемые множеством летальных мутаций.
Продленный мутагенез.
По современным представлениям, появление мутации не одномоментный акт реакции мутагена с участком хромосомы или геном, а очень сложный процесс. Он состоит из трех этапов. На первом этапе в результате взаимодействия мутагена и молекулы ДНК возникают первичные молекулярные повреждения в хромосоме. Второй этап, получивший название предмутационного состояния, связан с изменением структуры ДНК. На третьем этапе появляется собственно мутация как следствие фиксации потенциального изменения.
Первый этап обычно бывает очень кратковременным, иногда он длится всего 10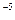 – 10 – 10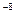 секунд, второй может быть длительным, охватывая ряд клеточных циклов, а у одноклеточных организмов – и несколько поколений особей. Потенциальные изменения бывают короткоживущими, длительными и сверхдлительными. В последнем случае они реплицируются и могут проходить через ряд синтезов ДНК. В соответствии с этим наблюдается разнообразие типов продленного мутагенеза. секунд, второй может быть длительным, охватывая ряд клеточных циклов, а у одноклеточных организмов – и несколько поколений особей. Потенциальные изменения бывают короткоживущими, длительными и сверхдлительными. В последнем случае они реплицируются и могут проходить через ряд синтезов ДНК. В соответствии с этим наблюдается разнообразие типов продленного мутагенеза.
Модифицирование мутагенного эффекта.
Частоту и спектр мутаций, индуцируемых определенными мутагенами у данного объекта, можно существенно менять, т. е. модифицировать. Это достигается путем изменения режима мутагенной обработки или применением до, во время или после воздействия некоторых специфических агентов.
Повреждающее влияние ионизирующих излучений, в частности лучей Рентгена, на хромосомный аппарат и ростовые процессы клеток можно в значительной степени снижать путем воздействия многими факторами, например химическими веществами, температурой и т. д. Агенты, обладающие способностью понижать частоту спонтанных или индуцированных мутаций, называются антимутагенами*
.
В опытах по облучению дрозофилы и некоторых растений было установлено, что эффект ионизирующей радиации в сильной степени зависит от того, в присутствии кислорода или без него он проводится. Оказалось, что бескислородная среда является защитным средством против ионизации, облучение же в атмосфере чистого кислорода резко увеличивает процент мутаций. Возрастание частоты мутаций при действии ионизирующей радиации в присутствии кислорода получило название кислородного эффекта
5
.
______________________________________
5
Рапопорт И. А. Гены, эволюция, селекции: Избр. труды – М., Наука, 1996
4.2.Генетическая токсикология.
Следует отметить, что загрязнение окружающей среды опасно не только ныне живущему поколению, но часто представляет опасность для грядущих поколений, поскольку многие загрязнители мутагенны. Выявление и устранение генетически активных факторов из среды обитания человека – задача генетической токсикологии, которая представляет собой наиболее активно развивающийся раздел экологической генетики. Это объясняется ее огромным прикладным значением.
Парадоксально, но факт, что открытие индуцированного мутационного процесса потребовало значительных усилий от исследователей: вспомним, что Г. Дж. Меллер получил Нобелевскую премию за открытие мутагенного действия рентгеновских лучей. Теперь же мутагены обнаруживаются на каждом шагу. Многие продукты производственной деятельности человека, появляющиеся как результат так называемого технического прогресса, обладают генетической активностью. При этом мы не говорим об отходах производства. Это могут быть лекарства, консерванты, пищевые добавки и красители, косметика, не говоря уже о дыме сигарет и излучениях, сопровождающих «мирный атом», тем более оружие массового уничтожения – ядерное и химическое.
Генетически активные факторы.
В генетической токсикологии принято говорить не только о мутагенах, но и, более широко, о генетически активных факторах. Не всегда удается определить непосредственный мутагенный эффект того или иного воздействия, но можно показать его влияние на кроссинговер, т. е. рекомбинацию генов или индукцию репаративного синтеза ДНК, сопровождающего многие повреждения генетического материала.
Таким образом, мутагенез, рекомбинагенез и индукция репаративного синтеза ДНК – это показатели генотоксичности* или генетической активности исследуемого фактора.
Генетически активные факторы делятся на физические, химические и биологические. К физическим факторам относятся температура, ионизирующая радиация, ультрафиолетовый свет и т. д. Химические генетически активные факторы гораздо труднее поддаются перечислению и классификации. Достаточно сказать, что к ним относятся любые вещества, прямо или косвенно нарушающие структуру и воспроизведение молекул ДНК. Выхлопные газы автотранспорта и выбросы в атмосферу производственных предприятий содержат алкилирующие соединения (их называют радиомиметиками), органические соединения ртути, полициклические углеводороды, обладающие генетической активностью. Многие химические соединения сами по себе не проявляют генетической активности, но их легко активизируют внутриклеточные метаболиты, а иногда и соединения, находящиеся в окружающей организм среде. Например, распространенные соли азотной кислоты легко превращаются в нитраты (соли азотистой кислоты) – мутагены, дизаминирующие основания ДНК.
С некоторых пор ( у нас с 1979 года) все новые химические соединения (а всего их в обиходе более 5,4 млн.) проходят проверку на генетическую активность. Это своеобразная служба генетической безопасности, использующая богатый арсенал различных тест-систем для выявления генетической активности. Эти системы позволяют учитывать мутации генов, их рекомбинации, потери и другие аберрации хромосом, нарушение деления ядра, индукцию репарации ДНК и т. д. При этом используются различные объекты: бактерии, дрожжи и другие низшие грибы, плодовая мушка-дрозофила, растения, культура клеток животных и человека.
Наибольший интерес представляет генетическая активность исследуемых агентов для человека. Поскольку прямое исследование их действия на человека невозможно, приходится ограничиваться результатами, получаемых на модельных объектах. Эти результаты в значительной степени справедливы и для человека из-за биологической универсальности свойств генетического материала – это всегда ДНК. Тем не менее экстраполяция получаемых результатов на человека всегда представляем некоторые сложности, так как наряду с принципом биологической универсальности следует учитывать и специфику объектов, имеющих свои особенности реагирования на мутагены.
В качестве примера расскажем только об одной тест-системе, получившей широкое распространение при первичном выявлении генетической активности. Эта система, разработанная в 60-е годы 20 века американским исследователем Б. Эймсом, который длительное время изучал мутации в генах, контролирующих биосинтез гистидина у Salmonella
typhimuium.
Работа Эймса, прекрасный пример того, как первоначально чисто теоретическое исследование, направленное на выяснение структуры и функции гена, приобрела сугубо практическое значение. Имея в своем распоряжении подробно охарактеризованные мутанты сальмонеллы, нуждающиеся в гистидине, зная молекулярную природу мутационных изменений: замены, вставки или выпадения пар оснований в ДНК гена или более крупные перестройки генетического материала, Эймс предложил изучить реверсии гистидиновых мутантов, то есть восстановление у них способности синтезировать гистидин, и, следовательно расти на среде без гистидина в результате воздействия различных мутагенов.
Тест очень прост: достаточно засеять среду без гистидина мутантом сальмонеллы, нуждающемся в гистидине (который естественно не растет на такой среде), и нанести в центр используемой для этого чашечки Петри испытуемое химическое соединение. Через 2-3 суток можно видеть появление колонии мутантов (в данном случае реверантов) вокруг пятна нанесенного вещества, если оно обладает генетической активностью (рис. 4).
С применением теста Эймса впервые были показаны мутагенные эффекты сигаретного дыма, некоторых пищевых красителей, консервантов и т. д.
Особый интерес представляют биологические генетически активные факторы, поскольку их существование указывает на генетическую активность синэкологических отношений. В конце 30-х годов С. М. Гершензон установил мутагенный эффект ДНК и вирусов. Позже было выяснено, что хромосомные аберрации в соматических клетках вызывают вирусы оспы, кори, ветряной оспы, гриппа, гепатита. Привести подобных примеров можно множество.
Под руководством М. Е. Лобашева еще в 60-е годы на кафедре генетики и селекции Ленинградского университета были начаты эксперименты, доказывающие роль нервной системы в контроле частоты хромосомных аберраций в соматических клетках (роговице глаза) у мышей. Развивая это направление ученики Лобашева (Цапыгина, Новоков, Даев) показали мутагенный эффект фермонального стресса у мышей. При этом важно, что речь уже идет не о мутациях в соматических, а в генеративных клетках – при сперматогенезе. Схема проделанного эксперимента представлена на рис. 5
Известно, что запах во взаимоотношении мышей выполняет функции своеобразного языка. Феромоны, летучие вещества, содержащиеся в моче этих животных, играют роль сигналов, вызывающих реакцию подчинения, агрессии и т. д. Пользуясь этими сигналами старые самцы держат в подчинении самок и молодых самцов. Оказалось, что запах старого самца при однократном воздействии повышает частоту цитологических нарушений в сперматогенезе у молодых самцов, увеличивает частоту аномальных сперматозоидов и доминантных летальных мутаций, выявляемых после их спаривания с самками, не подвергавшимися воздействию.
Таким образом, синэкологические отношения* могут быть источником наследственной изменчивости и тем самым служить дополнительным фактором эволюции.
5.Выводы.
В этой работе я рассмотрела основные факторы, которые влияют на организмы, приводя к изменению их генотипа. А, как известно, мутировавшие организмы являются основой для естественного отбора, который может привести к образованию новых видов.
Проделав данную работу, я пришла к следующим выводам:
1. Эволюционное учение прошло несколько этапов в своем развитии: от эволюционных представлений Ламарка, где главенствующая роль отводилась стремлениям организмов к самосовершенствованию, наука перешла к эволюционной теории Дарвина. Но и в ней нашлись слабые места (представления о наследственности). В данное время в науке господствует синтетическая теория эволюции.
2. Синтетическая теория эволюции утверждает, что структура популяции зависит не только от возникновения новых мутаций, но и от простого изменения частоты встречаемости данного гена.
3. Существует множество принципов классификации мутаций. Я рассмотрела классификацию по их действию и влиянию на организм. А также виды и типы мутаций.
4. При изучении данного вопроса я пришла к выводу, что наиболее эффективен естественный отбор, направленный против доминантных мутаций, в то время как рецессивные мутации исключаются естественным отбором очень медленно.
5. В отдельных популяциях никогда данный признак не может быть истреблен полностью (вне зависимости от того является ли он рецессивным или доминантным).
6. В природе невозможно появление новых видов без генотипической изоляции и адаптации. Если данная группа организмов окажется изолированной от основной части популяции, то в ней выживут только наиболее приспособленные особи, генотип которых, в силу отличия окружающей природы в данной местности от природы ареала, где обитала исходная популяция, отличается от генотипа предков.
7. Ученые выявили множество мутагенов – факторов, влияющих на возникновение мутаций. В настоящее время они применяются для искусственного получения мутаций. К физическим мутагенам относят электромагнитные и корпускулярные излучения, которые являются радиационными.
8. Окружающая среда постоянно загрязняется человеком, причем в нее попадает все то, что человек использовал в своей деятельности. И, к сожалению, очень редко мы задумываемся о том, не несет ли нам это угрозу. Генетическая токсикология занимается изучением генетически активных факторов (мутагенов), которые могут повлиять на изменения генотипа организма.
Я считаю, что гипотеза этой работы получила полное подтверждение: невозможно образование новых видов, а как следствие и эволюция, только результате мутационного процесса. С течением времени (без изоляции) данный признак (полученный в результате мутации) «растворяется» в популяции. Или, например, если бы отсутствовал естественный отбор, то говорить о существовании видов, а тем более эволюции не было бы смысла, т. к. в данном случае все мутации сохранялись бы.
6. Список использованной литературы.
1. Гуляев С.А., Жуковский В.М., Комов С.В. «Основы естествознания», Екатеринбург, 1997 г.
2. Дубнищева Т.Я. «Концепции современного естествознания», Новосибирск, «Изд-во ЮКЭА», 1997 г.
3. Петровский Б.В. «Популярная медицинская энциклопедия», М., «Советская энциклопедия», 1997 г.
4. Айала, Франциско Дж. Современная генетика – М., 1988.
5. Гуляев Г. В. Генетика – М., Колос, 1984.
6. Дубинин Н. П. Общая генетика: изд. 2-е, перераб. и доп. – М., 1976.
7. Иорданский Н. Н. Эволюция жизни: Учеб. Пособие для вузов – М. Академия, 2001
8. Мюнтцинг А. Генетика. Общая и прикладная – М., 1967
9. Приходченко Н. Н., Шкурат Т. П. Основы генетики человека – Ростов-на-Дону, Феникс, 1997
10. Рапопорт И. А. Гены, эволюция, селекции: Избр. труды – М., Наука, 1996
11. Скворцов А. К. Микроэволюция и пути видообразования – М., знание, 1982.
12. Воронцов Н. Н. Синтетическая теория эволюции: ее источники, основные постулаты и нерешенные проблемы // Журн. Всес. хим. о-ва им. Д. И. Менделеева. 1980. Т. 25. N 3. С. 293—312.
13. Вяткин Ю. С., Журавлев В. Б., Киселев В. Д. Эволюционная теория Дарвина и современность // На сайте Алтайского государственного университета (www.asu.ru).
14. Грант В., "Эволюционный процесс: Критический обзор эволюционной теории": Пер. с англ. - М.: Мир, 1991. ISBN 5-03-001432-2
15. Иорданский Н. Н. Эволюция жизни. М., 2001.
16. Красилов В. А. Теория эволюции: необходимость нового синтеза // Эволюционные исследования. Макроэволюция. Владивосток: 1984.
17. Четвериков С. С. О некоторых моментах эволюционного процесса с точки зрения современной генетики // Классики современной генетики. М.: 1968.
18. Шмальгаузен И. И. Пути и закономерности эволюционного процесса. — 2-е изд. — М., 1983. — (Сер. Избр. труды).
*
u
–
общий объем мутаций
*s
–
коэффициент селекции
*p
–
концентрация доминантного аллеля
*q - концентрация аллеля
|
