Предмет и задачи Микробиологии. Разделы микробиологии. Основные перспективные направления науки.
После открытия микроорганизмов прошло три столетия, и наука занимающиеся их изучением - МИКРОБИОЛОГИЯ - заняла достойное место среди других биологических и медицинских наук. Микроорганизмы широко распространены в природе. Они находятся в воздухе , почве , пище , на окружающих нас предметах , на поверхности и внутри нашего организма . Такое широкое распространение микробов свидетельствует об их значительной роли в природе и жизни человека. Микроорганизмы обуславливают круговорот веществ в природе, осуществляют расщепление органических соединений и синтез белка. С помощью микроорганизмов происходят важные производственные процессы : хлебопечение, производство ферментов, гормонов антибиотиков и других в-вв.
Наряду с полезными микроорганизмами существует группа патогенных микробов - возбудители различных заболеваний человека , животных , растений . Микроорганизмы были открыты в конце 18 века, но микробиология как наука сформировалась только в начале 19 века , после гениальных открытий французского ученого Луи Пастера .
В связи с огромной ролью и задачами микробиологи не может справится со всеми вопросами в пределах одной дисциплины и в следствие этого происходит ее дифференцировка в различные дисциплины .
Общая микробиология - изучает морфологию , физиологию , биохимию микроорганизмов , их роль в круговороте в-в и распространение в природе.
Техническая микробиология - входит изучение микробов участвующих в производстве антибиотиков , спиртов , витаминов , также разработка методов защиты материалов от воздействия микроорганизмов .
Сельскохозяйственная микробиология - изучает роль и значение микробов в формирование структуры почвы , ее плодородия , минерализация и питание растений .
Ветеринарная микробиология - изучает возбудители заболеваний у животных , разрабатывает методы специфической профилактики и терапии инфекционных заболеваний .
Медицинская микробиология - рассматривает свойства патогенных и условно - патогенных микробов , их роль в развитие инфекционного процесса и иммунного ответа , разрабатывает методы лабораторной диагностики и специфической профилактики и терапии инфекционных заболеваний .
Реклама

Вирусология - изучает неклеточные микробы - вирусы , их природу ,химический состав , взаимоотношение с клеткой хозяина , механизмы внутриклеточного паразитизма и т.д.
Важнейшими задачами медицинской микробиологии , вирусологии , иммунологии является дальнейшие изучение роли отдельных видов патогенных агентов в этиологии и патогенезе различных заболеваний людей , в том числе в возникновение опухолей , а также механизмов формирования наследственного и приобретенного иммунитета , разработка методов лечения и профилактики инфекционных заболеваний при помощи иммунологических и химиотерапевтических средств и методов специфической диагностики , в том числе экспресс-методов.
Большое значение в народном хозяйстве приобретает использование мик-роорганизмов как продуцентов множества полезных веществ, как–то: кормового белка, ферментов, антибиотиков, витаминов. Активно разрабатываются способы рационального использования биохимической активности микроорганиз-мов для повышения плодородия почв, добычи полезных ископаемых, восполнения энергетических ресурсов и очистки окружающей среды от многих загрязняющих веществ.
Вместе с тем остается необходимость изыскивать эффективные способы борьбы с некоторыми микроорганизмами, вызывающими заболевания человека, животных и растений, а также порчу промышленных изделий и нежелательные изменения окружающей среды.
История развития микробиологии. Основные открытия. Достижения русских ученых в развитии микробиологии. Развитие современной науки.
первым конструктором микроскопа считается Г. Галилей (1564 – 1642)
первым исследователем, наблюдавшим при помощи сильной лупы простейшие организмы в испорченном мясе, молоке и других продуктах, был Афанасий Кирхер (1601 – 1680).
Морфологический:
Период наблюдений и описаний, период первых в микробиологии терминов, рисунков, статей. Антони ван Левенгук (Leeuwenhoek) (1632-1723) - нидерландский натуралист, один из основоположников научной микроскопии. Изготовив линзы с 150-300-кратным увеличением, впервые наблюдал и зарисовал простейших, сперматозоиды, бактерии, эритроциты и их движение в капиллярах.
Физиологический:
Период экспериментов, поиска новых методов изучения, изобретения новых микроскопов, период открытий в микромире. Работы Пастера по оптической асимметрии молекул легли в основу стереохимии. Открыл природу брожения. Опроверг теорию самозарождения микроорганизмов. Изучил этиологию многих инфекционных заболеваний. Разработал метод профилактической вакцинации против куриной холеры (1879), сибирской язвы (1881), бешенства (1885). Ввел методы асептики и антисептики.
Реклама

Во 2-й половине XIX в. в России и в мире микробиология разделилась на два направления:
- Общая: Основоположник Л.С. Ценковский (1822 – 1887)
-Медицинская: Основоположник Роберт Кох (1843 – 1910)
Ивановский Д. И. (1864 – 1920) Основоположник вирусологии,
Мечников И. И. (1845-1916) Основоположник иммунологии
Виноградский С. Н. (1856—1953) Основоположник Почвенной микробиологии
Гамалея Н. Ф. врач бактериолог Его труды касаются общей бактериологии, бешенства и многих патогенных микробов.
- Появление электронного микроскопа стало возможным после ряда физических открытий конца XIX — начала XX векая:
- 1897 г. открытие электрона Дж. Томсон
- 1926 г. экспериментальное обнаружение волновых свойств электрона К. Дэвиссон, Л. Гермер
- 1926 г. X. Буш создание магнитной линзы, позволяющую фокусировать электронные лучи
- 1931 г. Р. Руденберг собрал просвечивающий электронный микроскоп
- 1932 г. М. Кнолль и Э. Руска построили первый прототип современного прибора.
- Использование электронного микроскопа для научных исследований было начато в конце 1930-ых годов и тогда же появился первый коммерческий прибор, построенный фирмой Siemens.
- В 1930—1940 годах появились первые растровые электронные микроскопы. Массовое применение этих приборов в научных исследованиях началось в 1960-ых годах, когда они достигли значительного технического совершенства.
Распространение микроорганизмов в природе. Участие в производственных процессах.
В природе микроорганизмы
заселяют практически любую среду (почва, вода, воздух) и распространены гораздо шире, чем другие живые существа. Благодаря разнообразию механизмов утилизации источников питания и энергии, а также выраженной адаптации к внешним воздействиям, микроорганизмы могут обитать там, где другие формы жизни не выживают.
Естественные среды обитания
большей части организмов — вода, почва и воздух. Число микроорганизмов, обитающих на растениях и в организмах животных, значительно меньше. Широкое распространение микроорганизмов связано с лёгкостью их распространения по воздуху и воде; в частности, поверхность и дно пресноводных и солёных водоёмов, а также несколько сантиметров верхнего слоя почвы изобилуют микроорганизмами, разрушающими органические вещества. Меньшее количество микроорганизмов колонизирует поверхность и некоторые внутренние полости животных (например, ЖКТ, верхние отделы дыхательных путей) и растений.
В зонах обитания микроорганизмы образуют биоценозы
— сложные ассоциации со специфическими и часто необычными взаимоотношениями. Каждое микробное сообщество в конкретном биоценозе образуют специфичные аутохтонные микроорганизмы
, то есть микробы, присущие конкретной области. В состав этих сообществ могут внедрятся аллохтонные микробы (
например, паразитические), обычно в них не встречающиеся. В природных биоценозах (почва, вода, воздух) выживают и размножаются лишь те микроорганизмы, которым благоприятствует окружающая среда; их рост прекращается, как только меняются условия окружающей среды.
В природе большую часть бактерий
поедают хищные простейшие, но часть клеток каждого вида выживает; при наступлении благоприятных условий они дают начало новым клонам микроорганизмов.
Неклеточные формы жизни. Морфология и размножение вирусов. Отличительные черты прионов.
Живые организмы делятся на клеточные (Прокариоты и эукариоты) и неклеточные (прионы и вирусы).
Прионы
– возбудители медленных неконвекционных инфекций. Состоят из набора специфичных белков и аномальных изоформных клеточных белков с молекулярной массой от 20000 до 37000 единиц. (Заболевания: Куру, Болезнь Кройтцфельда – Якоба, Амниотрофический лейкоспорангиоз)
Морфология и размножение вирусов
.
По внешнему виду вирусы делят на сферические, или шарообразные, кубические, палочковидные, или нитевидные, и сперматоподобные.
При некоторых вирусных инфекциях (бешенство, оспа и др.) в цитоплазме или ядре пораженной вирусом клетки образуются особые, специфические для каждой инфекции внутриклеточные включения, значительно превосходящие по величине вирус и видимые в световой микроскоп. Это колонии вирусов. Обнаружение их в клетке имеет большое значение при диагностике бешенства, оспы и других инфекций
Отдельные виды вирусов, преимущественно вирусы растений, образуют в клетках кристаллические образования (кристаллы Ивановского). Их можно растворить, и из раствора выделяется вирус в аморфном, не кристаллическом состоянии, обладающий инфекционными свойствами. В каждом кристалле содержится до 1 млн. вирионов. Из зоопатогенных вирусов в кристаллическом виде пока получен чирус полиомиелита.
Размеры вирусов колеблются в широких пределах. Мельчайшие из них (вирусы полиомиелита, ящура, энцефалитов) имеют в диаметре около 20—30 тр. (миллимикрон) и приближаются по величине к белковым молекулам, а крупные вирусы (вирусы оспы, герпеса, плевропневмонии) по рамерам близки к мельчайшим бактериям. Размер вирусов определяют ультрафильтрацией, ультрацентрифугированием и электроноскопйей. Каждым из этих методов получены более или менее сходные результаты, однако наиболее точным является электроноскопия высокоочищенного вируса.
Размножение
вирусов включает в себя три процесса: репликацию вирусной нуклеиновой кислоты, синтез вирусных белков и сборку вирионов.
После проникновения вирусов в клетку и раздевания вирусный геном и связанные с ним вирусные белки оказываются в цитоплазме. Внутри зараженной клетки происходят репликация вирусного генома и синтез структурных белков, из которых собираются новые вирусы. Существует определенный порядок
транскрипции вирусных
мРНК, которые затем транслируются с образованием белка. Репликация генома и сборка
нуклеокапсидов большинства
РНК-содержащих вирусов происходят в цитоплазме, а большинства
ДНК-содержащих вирусов - в ядре.
Сборка вирионов высокоспецифический процесс взаимодействия белковых и нуклеиновых молекул, приводящий к образованию вирионов
. У простых РНК-геномных вирусов с кубической или спиральной симметрией сборка вирионов состоит в ассоциации вирусного генома с капсидными белками с помощью репликативного комплекса. У сложных РНК-геномных вирусов нуклеокапсид образуется так же, как у простых вирусов. Формирование суперкапсида - сложный многоступенчатый процесс, к-рый протекает в цитоплазматической мембране или специальных мембранных структурах («фабриках» вируса). У сложных ДНК-геномных вирусов сначала раздельно образуются капсид и нуклеоид, затем нуклеоид вносится в пустой капсид. Дальнейшая достройка вириона происходит в цитоплазматической мембране или эндоплазматическом ретикулуме. У поксвирусов все этапы размножения, включая С., протекают в транскриптазо-рибосомальных комплексах цитоплазмы.
Отличия вирусов от бактерий. Характеристика бактериофагов, их биологическое значение.
Вирусы отличаются от бактерий следующими особенностями:
- содержат нуклеиновую кислоту только одного типа – или ДНК, или РНК; - для репродукции необходима только одна нуклеиновая кислота;
- не способны размножаться вне живой «хозяйской» клетки;
- Не дышат, не питаются, не двигаются, не выделяют
- Вирусы – абсолютные паразиты
- Вне клетки вирус существует в виде вирусной частицы (вириона
)
- Вирион состоит из нуклеиновой кислоты и белковой оболочки – капсида
.
- Развитие вируса приводит к гибели клетки-хозяина
Существуют РНК-вирусы (пикорновирусы, флавивирусы, ортомиксовирусы, парамиксовирусы, реовирусы, ретровирусы, рабдовирусы) и ДНК-вирусы (бактериофаги, паповавирусы, аденовирусы, герпесвирусы, поксвирусы, онкорновирусы)
Бактериофаги
- Вирусы микроорганизмов (актиномицетов, бактерий) получили название фагов
(актинофаги и бактериофаги).
- Размеры их колеблются от 40 до 150 нм.
- Фаги существуют в двух формах: внутриклеточной (профаг, ДНК) и внеклеточной (вирион).
По морфологии фаги подразделяются:
- фаги с длинным отростком, чехол которого сокращается;
- фаги с длинным отростком, чехол которого не сокращается;
- фаги с короткими отростками;
- фаги с аналогом отростка;
- фаги без отростка;
- нитевидные фаги
Существует 2 типа взаимодействия фагов с клеткой:
Литический
(продуктивная вирусная инфекция) фаги литические
или вирулентные
- адсорбируется на клеточной стенке, проникает
- впрыскивается нуклеиновая кислота фага
- происходит репродукция фага
- подавляется синтез клеточных компонентов
- образуются ферменты для синтеза ДНК фага – «ранние белки»
- синтезируется нуклеиновая кислота фага
- прекращается синтез ДНК бактерии
- на рибосомах - синтез белка, образуются белки оболочки, лизоцимы и эндолизины – «поздние белки»
- происходит созревание, образуются новые частицы фагов
- вирионы выходят из клетки
- происходит лизис бактерии
Лизогенный
фаги умеренные
- адсорбируется на клеточной стенке, проникает
- впрыскивается нуклеиновая кислота фага
- происходит интеграция в геном клетки
- наблюдается длительное сожительство фага с клеткой без ее гибели
- при изменении внешних условий могут происходить выход фага из интегрированной формы и развитие продуктивной вирусной инфекции
Биологическое значение бактериофагов: они являются причиной гибели бактерий-продуцентов
Клеточные формы жизни. Основные отличия прокариот от эукариот.
| Признак
|
Прокариоты
|
Эукариоты
|
| Особенности строения клеток
|
| Наличие ядра |
Обособленного ядра нет |
Обособленное ядро, отделенное от цитоплазмы двойной мембраной. |
| Число хромосом и их строение |
У бактерий – одна кольцевая хромосома, прикрепленная к мезосоме, двуцепочечная ДНК, не связанная с белками гистонами. У цианобактерий – несколько хромосом в центре цитоплазмы. |
Определенное число хромосом для каждого вида. Хромосомы линейные, двуцепочечная ДНК, связанная с белками гистонами. |
| Плазмиды |
Имеются. |
Имеются у митохондрий и у пластид. |
| Ядрышки |
Отсутствуют. |
Имеются. |
| Организация генома |
До 1,5 тысяч генов. Большинство генов в единственной копии. |
В зависимости от вида от 5 до 200 тысяч генов. У человека около 100 тысяч генов. Доля генов, представленных в нескольких копиях достигает 45 %, число копия одного гена может достигать нескольких тысяч. |
| Признак
|
Прокариоты
|
Эукариоты
|
| Рибосомы |
Мельче, чем у эукариот – от 70 S. Распределены в цитоплазме. Обычно свободные, редко связанные с мембранными структурами. Составляют до 40 % массы клетки. |
Крупные, от 80 S. Находятся в цитоплазме в свободном состоянии или связаны с мембранами ЭПР. В пластидах и митохондриях содержатся рибосомы 70 S. |
| Одномембранные замкнутые органеллы |
Отсутствуют. Их функции выполняют выросты клеточной мембраны. |
Многочисленны: ЭПР, АГ, вакуоли, лизосомы. |
| Двухмембранные органеллы |
Отсутствуют. |
Митохондрии у всех эукариот, пластиды – у растений. |
| Клеточный центр |
Отсутствует. |
Имеется в клетках животных, грибов, растений (мхи и водоросли). |
| Мезосомы |
Имеются у бактерий. |
Отсутствуют. |
| Клеточная стенка |
У бактерий содержит муреин, у цианобактерий – целлюлозу, пектиновые вещества, немного муреина |
У растений – целлюлоза, у грибов – хитин, у животных клеточных стенок нет. |
| Признак
|
Прокариоты
|
Эукариоты
|
| Капсула и слизистый слой |
Имеются у некоторых бактерий. |
Отсутствуют. |
| Жгутики |
Простое строение, не содержат микротрубочек. Диаметр 20 нм. |
Сложного строения, содержат микротрубочки. Диаметр 200 нм. |
| Размер клеток |
Диаметр 0,5 – 5 мкм. |
Диаметр до 50 мкм. Объем может превышать объем прокариотической клетки в тысячу и более раз. |
| Особенности жизнедеятельности клеток
|
| Движение цитоплазмы |
Отсутствует. |
Наблюдается часто. |
| Аэробное дыхание |
У бактерий – в мезосомах, у цианобактерий – на ЦПМ. |
В митохондриях. |
| Фотосинтез |
Хлоропластов нет. У фотосинтезирующих бактерий и цианобактерий происходит на мембранах. |
В хлоропластах у растений. |
| Фагоцитоз и пиноцитоз |
Отсутствует из-за наличия жесткой клеточной стенки. |
Только у животных. |
| Спорообразование |
Некоторые бактерии обладают способностью образовывать спору, предназначенную для перенесения неблагоприятных условий среды. |
Свойственно растениям и грибам, предназначены для размножения. |
| Способы деления клетки |
Равновеликое бинарное деление, редко почкование. Митоз и мейоз отсутствуют. |
Митоз, мейоз, амитоз. |
Характеристика эукариотических микроскопических организмов. Морфология дрожжей
Микроскопические грибы
- большая группа эукариотических организмов
- бесхлорофильные низшие организмы с гетеротрофным типом питания, неподвижные, размножаются различными способами
- наиболее широко в природе среди микроскопических форм распространены дрожжи и плесневые грибы.
Дрожжи
– одноклеточные неподвижные организмы, относятся к классу сумчатых грибов Ascomycetes
. Классификация основана на особенностях их размножения и других морфологических признаках.
Особенности дрожжей:
-
Дрожжи имеют разнообразную форму: круглую, эллиптическую, овальную, реже лимонообразную и цилиндрическую, иногда сильно вытянутую в виде гиф. Форма и структура клеток непостоянна, она может изменяться в связи с изменением условий культивирования.
-
Жесткая клеточная стенка (хитин)
-
Развитый мембранный аппарат
-
Вакуоли
-
Запас питательных веществ и ферментов
-
Различные способы размножения. (Сумки со спорами обычно повторяют форму и размер вегетативных клеток. Форма и размер спор у различного рода спорегенных дрожжей отличаются.)
Характеристика эукариотических микроскопических организмов. Морфология плесеней.
Микроскопические грибы
- большая группа эукариотических организмов
- бесхлорофильные низшие организмы с гетеротрофным типом питания, неподвижные, размножаются различными способами
- наиболее широко в природе среди микроскопических форм распространены дрожжи и плесневые грибы.
Группа плесневых грибов
относятся к несовершенным грибам из класса Phicomycetes
(представители – Muc
о
r
,
Rhizopus
,
Fhamnigium
) и класса Ascomycetes
семейства Aspergillaceae
(представители Aspergillus
и Penicillium
)
Особенности плесеней:
-
Клетки-гифы, мицелий
-
Одноклеточность и многоклеточность
-
Многоядерность
-
Устойчивость к воздействию температур
-
Токсинообразование, антибиотикообразование
-
Многообразие способов размножения
Тело плесневых грибов имеет большие линейные размеры, а гифы — относительно большую толщину. Поэтому их микроскопирование не требует предварительной окраски препарата. Обычно изучение ведется в препарате «раздавленная капля».
Характеристика эукариотических микроскопических организмов. Отличительные черты простейших, вызывающих инфекционные заболевания.
Тип Protozoa
насчитывает свыше 30 000 видов и подразделяется на четыре класса: 1) жгутиковые; 2) саркодовые; 3) споровики; 4) ресничные.
Простейшие – одноклеточные эукариотные животные организмы, более высоко организованные по сравнению с бактериями. Имеют цитоплазму, дифференцированное ядро, различную по своим оптическим свойствам оболочку, примитивные органоиды, часто - органы движения, лишены клеточной стенки
Кожный лейшманиоз.
Класс жгутиковых, семейство Trypanosomidae
.
Лейшмании (Leishmania
tropica
) в пораженных тканях (клетках) представляют собой круглые или овальные неподвижные образования. Длина 2–6 мкм, ширина 2–3 мкм. В организме беспозвоночных и культурах образуются лептомонадные (жгутиковые) формы; размеры паразитов увеличиваются до 20 мкм в длину и 3 мкм в ширину. Заражение лейшманиозом осуществляется переносчиком – комаром рода Phlebotomus
.
Трипаносомоз.
Возбудителями являются два вида: Trypanosoma
gambiense
(Даттон Дж., 1902) и Trypanosoma
rhodosiense
(Фентем Г., 1910). Класс жгутиковых, семейство Trypanosomidae
, род Trypanosoma
.
Возбудители имеют вид веретенообразной клетки с мембраной, заостренными жгутиками на концах; подвижны, длина их 25–40 мкм, ширина 20 мкм. Трипаносомоз передается через укусы мух Glossina
palpalis
(це-це).
Трихомониаз.
В организме человека паразитирует три вида четырехжгутиковых трихомонад, относящихся к классу жгутиковых, семейству Trichomonadidae
.
-
Trichomonas
vaginalis
(Донне А., 1837). Размер 20–36 мкм, обитает в основном в нижних отделах половой системы и развивается у женщин в возрасте 18–45 лет.
-
Trichomonas
intestinalis
(Давен К., 1860) размером 10–17 мкм, живет в толстом отделе кишечника человека.
-
Trichomonas
hominis
(Мюллер О., 1873), размерами 10–17 мкм встречается главным образом во рту у пожилых людей с плохими зубами, пародонтозом.
Лямблиоз.
Класс жгутиковых, Lamblia
intestinalis
, открытая Д. Ф. Лямблем в 1859 г.
Имеют вид двустороннего симметричного грушевидной формы организма с вытянутым задним концом, с двумя симметрично расположенными ядрами, длина 10–18 мкм и ширина 8–10 мкм.
Паразиты образуют цисты овальной формы длиной 10–14 мкм и шириной 7,5–9 мкм. Лямблии имеют четыре ядра.
Малярия.
Класс Sporozoa
. Обладают способностью инифицировать эритроциты и другие клетки позвоночных. В 1880 г. А. Лаверан открыл возбудителя четырехдневной малярии, в 1890 г. В. Грасси и Р. Фелетти – возбудителя трехдневной малярии, в 1897 г. У. Уэлч – возбудителя тропической малярии, в 1922 г. Ж. Стивене – возбудителя трехдневной малярии (Plasmodium
ovale
).Роль комара в эпидемиологии малярии птиц установлена в 1895 г. Р. Россом, а в эпидемиологии малярии человека – в 1898 г. П. Мансоном. Промежуточный хозяин - комар из рода Anopheles
Токсоплазмоз.
Возбудитель токсоплазмоза – Toxoplasma
gondii
– выделен у грызуна Ctenodactilis
gondii
в 1908 г. Ш. Николем и Л. Мансо в Алжире. В последующие годы этот паразит был обнаружен у разных видов домашних и диких животных. Он относится к классу споровиков и имеет сходство с плазмодиями малярии. Toxoplasma
gondii
имеет форму полумесяца, овала; концы ее иногда заострены; длина 4–7 мкм и ширина 5 мкм.
Морфология бактерий. Разнообразие форм. Размеры МО. Методы изучения морфологии бактерий. Виды микроскопов.
Бактерии
– одноклеточные прокариоты микроскопических размеров, размножающиеся преимущественно путем деления.
Форму и размер бактерий
изучают в убитом и живом состоянии при помощи микроскопии в окрашенных и неокрашенных препаратах.
Палочковидные: палочки, образующие споры, называют бациллами (от лат. bacillus
– палочка), а палочки, необразующие споры, – бактериями (от греч. bacteria
– палочка). Палочковидные бактерии могут иметь цилиндрическую форму с ровными концами или овальную с закругленными концами. (бациллы, диплобациллы, стрептобациллы)
Извитые, или спиралевидные, формы.
Одни из них имеют от 6 до 15 и более витков, другие представляют лишь часть витка. (спирохеты, спириллы, вибрионы)
Кокковидные:
имеют правильную форму шара, но некоторые виды напоминают форму кофейного зерна или вытянуты наподобие пламени свечи и ланцета (микрококки, диплококки, стрептококки, тетракокки, сарцины, стафилококки)
И другие самые разнообразные неклассифицированные формы бактерий
Размеры бактерий.
-
Стафилококки и стрептококки обычно имеют диаметр от 0,75 до 1,25 мкм.
-
Средних размеров палочковидные бактерии имеют ширину от 0,5 до 1 мкм и длину от 2 до 3 мкм (Е. соli, Shigelladisentheria и др.)
-
Мелкие палочки имеют ширину 0,2–0,4 мкм и длину 0,7–1,5 мкм (Bordetellapertussis).
-
Среди палочковых бактерий имеются относительно крупные. Bacillusanthracis имеет поперечник 1–1,25 мкм и длину от 3 до 8–10 мкм.
Микоплазмы - величина находится на грани видимости в микроскопы (0,1–0,2 мкм).
Длина клетки спирохеты может достигать 500 мкм.
Масса бактериальной клетки очень мала и составляет приблизительно 4×10-13 г.
Методы изучения морфологии бактерий.
1.
Световая микроскопия.
-
Обычная микроскопия
-
фазово-контрастная микроскопия
-
микроскопия с высоким разрешением
-
люминесцентная микроскопия
-
иммерсионные масла
2. Электронная микроскопия
-
просвечивающая
-
сканирующая
Морфология бактерий. Химический состав бактериальной клетки.
| Компонент
|
Общее количество, % от сухих веществ клетки
|
Молекулярная масса, Да
|
Количество молекул в клетке
|
Число разных видов молекул в клетке
|
| Белок |
55,0 |
4,7·104
|
2 350 000 |
1850 |
| РНК |
20,5 |
| тРНК |
2,5·104
|
198 000 |
60 |
| иРНК |
1,0·106
|
1 380 |
600 |
| ДНК |
3,1 |
2,5·109
|
2 |
1 |
| Липиды |
9,1 |
705 |
22 000 000 |
1 |
| Липополисахариды |
3,4 |
4070 |
1 430 000 |
1 |
| Пептидогликан |
2,5 |
(904)n |
1 |
1 |
| Гликоген |
2,5 |
1,0·106
|
4300 |
1 |
| Полиамины |
0,4 |
1 |
| путресцин |
88 |
5 600 000 |
1 |
| спермидин |
145 |
1 100 000 |
1 |
| Метаболиты, ионы |
3,5 |
800 |
- Осевшая клеточная масса содержит 70–85% воды
- сухая биомасса составляет 15–30% от сырой массы
Сухое вещество бактерий: белки (50%), компоненты клеточной стенки (10–20%), РНК (10–20%), ДНК (3–4%), липиды (10%)
Десять важнейших химических элементов: углерод – 50%, кислород – 20%, фосфор – 3%, азот – 14%, водород – 8%, сера – 1%, калий – 1% кальций – 0,5%, магний – 0,5%, железо – 0,2%
Морфология бактерий. Строение и химический состав внешних слоев. Капсула, слизистые слои, чехлы.
Клетка бактерий окружена трехслойной оболочкой: Слизистый слой, Клеточная стенка, Цитоплазматическая мембрана
Слизистый слой.
Капсула
– защитное приспособление, вырабатываемое слизистым слоем. Состоит из гликопротеидов муцина и мукоидов
Зооглея –
капсулы отдельных клеток сливаются в слизистые массы, в которые вкраплены бактериальные клетки. Чехлы
–
слизистые структуры, отличающиеся от капсулы более тонкой структурой, часто в несколько слоев.S
-слои
(от англ. surface
– поверхность) – это простейший тип биологических мембран, возникших в ходе формирования первых клеток и входящих в состав поверхностей многих бактерий и почти всех архей.

Схема расположения внешних слоев бактериальной клетки: 1 – микрокапсула; 2 – клеточная стенка; 3 – цитоплазматическая мембрана; 4 – слизистый слой; 5 – капсула.
Различают микро- и макрокапсулу. Макрокапсулу можно выявить, используя специальные методы окраски, сочетая позитивные и негативные методы. Микрокапсула – утолщение верхних слоев клеточной стенки. Такую капсулу можно обнаружить только методом электронной микроскопии.
Среди бактерий различают:
- истиннокапсульные
бактерии (род Klebsiella
) – сохраняют капсулообразование и при росте на питательных средах, и в макроорганизме;
- ложнокапсульные
– образуют капсулу только при попадании в макроорганизм.
Значение капсулы:
слизь предохраняет клетки от неблагоприятных условий, защищает клетку от механических повреждений и высыхания, создает дополнительный осмотический барьер, служит препятствием для проникновения фагов, может являться источником запасных питательных веществ
Зооглея
Ослизнение клеточных стенок иногда бывает настолько сильным, что капсулы отдельных клеток сливаются в слизистые массы, в которые вкраплены бактериальные клетки
Образуемые некоторыми бактериями слизистые вещества не удерживаются в виде компактной массы вокруг клеточной стенки, а диффундируют в окружающую среду
Чехлы
могут иметь более сложную химическую организацию: белки – до 27 %, углеводы – до 36 %, гексозамин – до 11 %, липиды – около 5 %, фосфора – 0,5 %.
Чехол имеет поры диаметром 3 – 5 нм, которые позволяют проникать растворимым соединениям. Чехлы некоторых архей имеют особенности строения. Так, чехол клеток Methanospirillumhungateii образован плотным гибким слоем белковых или гликопротеиновых молекул толщиной около 10 нм, в состав которых входят 18 аминокислот (65 – 72 % от общей массы чехла) и 5 нейтральных сахаров (3 – 5 % общей массы).
Поверхностные
S
-слои.
-
выстилают наружную поверхность клеточной стенки равномерно упакованными образованиями правильной формы подобно паркетным дощечкам из белковых или гликопротеиновых молекул.
- у грамотрицательных бактерий S-слои прилегают непосредственно к внешней мембране, у грамположительных – ассоциированы с поверхностью муреина.
- S-слои архей могут быть единственным слоем клеточной стенки кроме ЦПМ, они более высоко организованы, чем бактериальные, располагаются над ЦПМ, образуя между слоями и ЦПМ небольшое пространство, связаны ковалентно с ЦПМ специфическими выступами.
- легко теряют регулярную структуру при приготовлении препаратов для микроскопирования.
Выполняют следующие функции:
- Защищают клетку при изменениях рН и концентраций других ионов, осмотического стресса, от действия ферментов, бактерий-хищников, фагов.
- Помогают удерживать определенную форму клетки, способствуют адгезии клеток к поверхностям и с ферментами.
- Участвуют в делении клеток, могут взаимодействовать с вирусами.
- Предотвращают неспецифическую адсорбцию макромолекул, помогают патогенным бактериям справиться с защитными факторами макроорганизма, избежать фагоцитоза и действия комплемента, что повышает вирулентность отдельных возбудителей.
Морфология бактерий. Клеточная стенка грамположительных и грамотрицательных бактерий. Окраска по Граму.
Клеточная стенка
- важный и обязательный структурный элемент большинства бактерий. Она служит механическим барьером между протопластом и окружающей средой, придает клетке определенную форму, большинство прокариот обладает ригидной клеточной стенкой.
Состав клеточной стенки:
-
У эубактерий - основа – полимер – гликопептид (пептидогликан, мукопептид) муреин
-
У архей – псевдомуреин
-
У уксуснокислой бактерии Acetobacter
xylinum
- целюллоза
Кроме муреина содержатся: Белки, липиды, липопротеиды, у некотороыз бактерий – тейхоевые кислоты.
Муреин состоит из цепочек N-ацетилглюкозамина и N-ацетилмурамовой кислоты, соединенных b-1,4-гликозидными связями, является опорным каркасом клеточной стенки, к которому прикрепляются другие вещества
Лизоцим бактерицидный фермент, активно растворяющий клеточную стенку,
содержится в слезной жидкости, носовой слизи, молоке, яичном белке,содержится в капсидах бактериофагов, хим. природа – ацетилмурамидаза, разрывает в муреине гликозидную связь
Муреиновый мешок: эластичен, способен растягиваться под воздействием внешних факторов, способен сжиматься до четырех раз, имеет отверстия размером до 6 нм, обладает избирательностью, имеет отверстия для выхода фимбрий и жгутиков
Окрашиваение по Граму:
Грамположительные окрашиваются в фиолетовый цвет, Грамотрицательные окрашиваются в розовый цвет. Окраска по Граму служит важным таксономическим признаком, с которым коррелируют другие свойства бактерий. Метод предложил в 1884 году Ганс Кристиан Йоахим Грам (1853 – 1938)
Грамположительные:
Толстый слой муреина, Содержание муреина 95 %, Содержание липидов около 5 %, Присутствуют тейхоевые кислоты
Грамотрицательные:
Тонкий слой муреина, Содержание муреина 5 – 10%, Содержание липидов около 75 %, Отсутствуют тейхоевые кислоты
Этапы окраски по Граму:
Фиксация препарата, Воздействие генциан-виолетом, Воздействие Люголя (Йод), Смывание спиртом, Смывание водой, Докраска сафранином или фуксином
Механизм образования окрашенного комплекса:
Муреин + тейхоевые кислоты + Йод + Генциан-виолет = фиолетовый стойкий окрашенный комплекс (Гр+)
Муреин + Йод + Генциан-виолет = нестойкий комплекс, легко смываемый спиртом (Гр-),
докрашиваются бактерии сафранином
Категории бактерий:-
грациликуты (gracilicutes– от лат. gracilis – тонкий, стройный, cutes – кожа) – бактерии с тонкой клеточной стенкой, грамотрицательные, к ним относятся различные извитые, палочковидные, кокковые формы бактерий, а также риккетсии и хламидии;
-
фирмикуты (firmicutes– от лат. firmis – крепкий, cutes – кожа) – бактерии с толстой клеточной стенкой, грамположительные, к ним относятся палочковидные, кокковые формы бактерий, а также актиномицеты, коринебактерии и микобактерии;
- тенерикуты или молликуты (mollicutes– от лат. mollis – мягкий, cutes – кожа) – бактерии без ригидной клеточной стенки (микоплазмы);
- мендозикуты(mendosicutes) – архебактерии, отличающиеся дефектной клеточной стенкой, особенностями строения рибосом, мембран и рРНК.
Морфология бактерий. Явление
L
-трансформации. Биологическая роль.
-
особая форма изменчивости или способность адаптироваться к окружающим условиям
-
проявляется в колониях чистых культур при исследовании их морфологии
-
может быть обратимой или необратимой
-
обнаруживаются необычные для данного вида бактерий формы: нитевидные, волокнистые, колбасовидные, шаровидные образования и мелкие гранулы размером 0,1-0,15 мкм (фильтрующиеся формы бактерий).
Обратимая
L
-трансформация:
- генетический контроль синтеза клеточной стенки сохраняется
- L-формы при благоприятных условиях могут возвращаться в исходную бактериальную форму
-
восстановление всех основных биологических свойств
Необратимая
L
-трансформация:
-
генетический контроль синтеза клеточной стенки нарушен необратимо
-
L-трансформация приобретает необратимый характер,
-
L-трансформанты по своим морфологическим, культуральным и иным свойствам становятся неотличимыми от микоплазм
Особенности
L
-форм:
-
Сходство морфологических изменений: образование нитевидных, волокнистых, колбасовидных, шаровидных, гранулярных и микоплазменных форм.
-
Сходные культуральные свойства: рост на плотных средах в виде характерных колоний двух типов – А и В.
-
Колонии типа А растут на поверхности агара, имеют очень мелкие размеры. Они состоят главным образом из гранулярных структур, лишенных клеточной стенки, и очень похожи на микоплазмы.
- Колонии типа В состоят из центральной зоны, врастающей в агар, и прозрачной фестончатой периферической зоны. В этих колониях обнаруживаются крупные тела, содержащие компоненты клеточной стенки, сходные со стенкой родительских бактерий, но лишенные ригидности.
- Образование стабильных и нестабильных L-форм (в зависимости от степени полноты утраты способности синтезировать клеточную стенку).
- Изменение антигенных свойств (утрата К- и О-антигенов как следствие нарушения синтеза клеточной стенки).
- Снижение вирулентности по сравнению с исходными родительскими формами в связи с утратой различных факторов патогенности.
- Способность длительно персистировать в организме. Утрата клеточной стенки делает L-формы нечувствительными к различным химиопрепаратам и антителам.
- Способность при неполной утрате синтеза клеточной стенки возвращаться в исходную бактериальную форму.
Морфология бактерий. Бактериальная мембрана. Строение мезосом, рибосом. Химический состав цитоплазмы.
Цитоплазматическая мембрана
(ЦПМ) плотно прилегает к клеточной стенке и отделяет от клеточной стенки содержимое клетки. О
бязательная структура любой клетки. Нарушение целостности мембраны приводит к потере клеткой жизнеспособности
На долю цитоплазматической мембраны приходится 8–15 % сухого вещества клетки. Толщина ее 7–10 нм. Представлена в виде трехслойной структуры – одного липидного слоя и двух примыкающих к нему с обеих сторон белковых слоев. Часто образует впячивания внутрь клетки или спиралевидные тельца – мезосомы.
Цитоплазматическую мембрану с мезосомами называют бактериальной мембраной.
Основные компоненты: белки (50–75 %), фосфолипиды (15–45 %), связанная вода (25 % сухой массы), небольшое количество углеводов, различные ферменты. Стеролы и стерины в бактериальной мембране большинства прокариот отсутствуют, поэтому бактерии проявляют устойчивость по отношению к некоторым антибиотикам, которые способны связываться со стеролами мембран эукариот. Так, например, нистатин и амфотерицин связывается со стеролами мембраны и увеличивает пропускную способность ЦПМ грибной клетки, в результате чего клетка гибнет.
Бактериальная мембрана закрепляет ферменты, выполняет штативную функцию, транспортирует вещества, принимает участие в обмене веществ между клеткой и окружающей средой, является главным осмотическим барьером клетки, участвует в репликации генома.
Мезосомы
локализуют ферменты, участвующие в энергетических процессах, в снабжении клетки энергией, осуществляют синтез материала клеточной стенки, принимают участие в распределении ДНК между новообразующимися особями при делении клетки. Хорошо развитые мезосомы обнаружены только у грамположительных бактерий, у грамотрицательных они развиты слабо и имеют более простое строение.
3 вида мезосом: ламеллярные (пластинчатые); везикулярные (в форме пузырьков); тубулярные (трубчатые). Часто у бактерий можно наблюдать мезосомы смешанного типа: состоящие из ламелл, трубочек, пузырьков.
По расположению в клетке различают: мезосомы, образующиеся в зоне клеточного деления и формирования поперечной перегородки (септы); мезосомы, к которым прикреплен нуклеоид; мезосомы, сформированные в результате инвагинации периферических участков ЦПМ.
Рибомосмы
рассеяны в цитоплазме в виде мелких гранул размером 20–30 нм, нуклеопротеиды, содержащие примерно 60 % РНК и 40% белка, количество рибосом в клетке зависит от условий ее культивирования и возраста (от 5000 до 50 000), имеют константу седиментации 70S, поэтому называются 70S-частицы, ответственны за синтез белка клетки.
Цитоплазма
полужидкая коллоидная система,
степень вязкости зависит от возраста и физиологического состояния клетки, обладает круговой подвижностью, обеспечивающую интенсивный перенос продуктов обмена от одного органоида к другому. Неоднородна, содержит различные мембраны, связанные системой канальцев и пузырьков. Содержит воду (в количестве 70–80 % от общей массы клетки), ферментные белки, аминокислоты, рибонуклеиновые кислоты, углеводы, липиды и различные низкомолекулярные вещества. Включает ядерный аппарат, рибосомы и гранулы запасных веществ различной природы.
В состав цитоплазмы входят все виды органических и неорганических веществ. В ней присутствуют также нерастворимые отходы обменных процессов и запасные питательные вещества. Основное вещество цитоплазмы — вода.
Морфология бактерий. Запасные включения бактериальной клетки.
Цитоплазматические включения очень разнообразны. Это питательные вещества. откладываются в клетке, когда она развивается в условиях избытка питательных веществ в среде, и потребляются, когда клетки попадают в условия голодания.
| Запасное вещество
|
Структурные характеристики
|
Химический
состав
|
Функции
|
Распространение
|
Гранулы
гликогена
(а-гранулы)
|
Сферической формы, диаметр 20–100 нм |
Высокомолекулярные полимеры глюкозы |
Источник углерода и энергии |
Широко распространенный тип запасных веществ |
| Гранулы поли-p-оксимасляной кислоты (до 50 % сухого вещества клетки) |
Диаметр 100–1000 нм; окружены однослойной белковой мебраной 2–3 нм толщиной |
98 % полимера поли-3- оксимасляной кислоты, 2 % белка |
Источник углерода и энергии |
Широко распространены только у прокариот |
| Гранулы или капли жира |
В жидком состоянии в виде капель |
Триацилглицериды |
Источник углерода и энергии |
Широко распространенный тип запасных веществ |
Гранулы
полифосфата
|
Диаметр 500 нм, зависит от объекта и условий роста |
Линейные полимеры ортофосфата |
Источник фосфора иэнергии |
Распространенный тип запасных гранул |
Аэросомы
– газовые вакуоли, имеющиеся у 15 таксономических групп почвенных и водных бактерий в цитоплазме. Аэросомы – это сложно организованные структуры, состоящие из множества регулярно расположенных газовых пузырьков, имеющих форму вытянутого цилиндра с заостренными концами длиной 200-1200 и диаметром 65-115 нм. Пузырьки окружены белковой мембраной и заполнены азотом, количество их может достигать 40–60 шт. Мембрана аэросом проницаема для газов, но не для воды.Регулируя степень наполнения вакуолей газом, клетка осуществляет механическое передвижение снизу вверх в водоемах или капиллярах почвы. Аэросомы присущи безжгутиковым формам и являются альтернативным видом движения.
Хлоросомы
– структуры, присущие фотосинтезирующим зеленым бактериям, локализующие пигменты – хлорофиллы с, d и е, поглощающие кванты света и передающие их в реакционные центры клетки. Хлоросомы имеют форму продолговатых пузырьков длиной 90-120 и шириной 25-70 нм, окруженных однослойной белковой мембраной толщиной 2-3 нм. Хлоросомы расположены вблизи от ЦПМ.
Фикобилисомы
– структуры цианобактерий, расположенные правильными рядами на внешних поверхностях фотосинтетических мембран, локализуют фикобилипротеины, имеют вид гранул диаметром 28-55 нм.
Карбоксисомы
– полиэдральные тела клеток некоторых фототрофных и хемолитотрофных эубактерий. Имеют форму многогранника с 4-6 сторонами и диаметром 90-500 нм.
Структуры белковой природы толщиной 3 нм заполнены гранулярным содержимым, включающим рибулозодифосфаткарбоксилазу, принимают участие в восстановительном пентозофосфатном цикле.
Карбоксисомы обеспечивают защиту и консервирование ключевого фермента рибулезофосфатного метаболического пути, предотвращая его разрушение внутриклеточными протеазами.
Магнитосомы
– цитоплазматические включения магнитных бактерий, заполненные железом в форме магнетита (Fe3O4) и выполняющие функцию магнитной стрелки. На долю магнетита может приходиться до 4 % сухого вещества таких бактерий. Магнитосомы для бактерий имеют приспособительное значение.
| Запасное вещество
|
Структурные характеристики
|
Химический
состав
|
Функции
|
Распространение
|
| Цианофициновые гранулы |
Размер и форма различны; могут достигать в диаметре 500 нм |
Полипептид, содержащий аргинин и аспарагиновую кислоту (1: 1),
М=(25–100)·103
Да
|
Источник азота |
Обнаружены у многих видов цианобактерий |
| Гранулы серы |
Диаметр 100–800 нм; окружены мембраной |
Включения жидкой серы |
Донор электронов или источник энергии |
Пурпурные серобактерии, бесцветные бактерии, окисляющие H2
S |
| Углеводородные гранулы |
Диаметр 200–300 нм; окружены белковой оболочкой 2–4 нм толщиной |
Углеводороды того же типа, что и в среде |
Источник углерода и энергии |
Представители родов Arthrobacter
,
Acinetobacter
, Мусо
bacterium
,
Nocardia
и другие прокариоты, использующие углеводороды |
Движение бактерий. Строение жгутика, толщина, длина, химический состав. Приготовление фиксированных препаратов и препаратов живых клеток МО.
Жгутики
– это тонкие, спирально закрученные нити белковой природы, которые могут осуществлять вращательные движения. Они прикрепляются к цитоплазматической мембране с помощью двух пар дисков и через поры в мембране выходят наружу. Толщина их от 10 до 20 нм, длина может достигать 200 мкм. У одних бактерий жгутик может быть полым, у других он заполнен веществом липидной природы.
Сам жгутик спирально закручен и состоит из порядка 20000 молекул сократительного белка флагеллина общей молекулярной массой от 20 до 60 кДа, синтез которого кодируется более чем 30 генами.
Флагеллин не обладает ферментативной активностью, однако, способен выполнять ритмические сокращения, в результате которых жгутик совершает спиральные волнообразные движения
В белках бактериальных жгутиков отсутствуют серосодержащие аминокислоты, мало ароматических и циклических аминокислот.
Кислые аминокислоты составляют до 20 % всех аминокислот филамента. Вследствие отсутствия цистеина полая структура жгутика защищена от изменений, вызванных изменением потенциала в окружающей среде.
Жгутики хорошо функционируют при рН среды от 3,0 до 11,0, при наступлении неблагоприятных условий жгутики дезагрегируют, движение затрудняется.
Жгутик состоит из трех частей. Основную массу жгутика составляет длинная спиральная нить (фибрилла, филамент), у поверхности клеточной стенки переходящая в утолщенную изогнутую структуру – крюк. Нить с помощью крюка прикреплена к базальному телу, вмонтированному в ЦПМ и клеточную стенку.
У большинства прокариот нить состоит только из одного типа белка флагеллина. Белковые субъединицы уложены в виде спирали, внутри которой проходит полый канал. Наращивание жгутика происходит с дистального конца, куда субъединицы поступают по внутреннему каналу. У некоторых видов жгутик снаружи дополнительно покрыт чехлом особого химического строения или же являющимся продолжением клеточной стенки и, вероятно, построенным из того же материала.
Базальное тело содержит 9 – 12 различных белков и представляет собой систему из двух или четырех колец, нанизанных на стержень, который является продолжением крюка. Базальное тело – это самая сложно устроенная часть жгутика, напоминающая ось с нанизанными на нее колесами.
Два внутренних кольца (М и S) – обязательные составные части базального тела, в то время как наружные кольца (Р и L) отсутствуют у грамположительных эубактерий и, следовательно, не необходимы для движения. М-кольцо локализовано в ЦПМ, S-кольцо располагается в периплазматическом пространстве грамотрицательных или в пептидогликановом мешке грамположительных эубактерий. Кольца Р и L, имеющиеся только у грамотрицательных эубактерий, локализованы соответственно в пептидогликановом слое и во внешней мембране.
Особенности строения базального тела определяются строением клеточной стенки. Интактность последней необходима для движения жгутиковых бактерий. Обработка клеток лизоцимом, приводящая к удалению пептидогликанового слоя клеточной стенки, вызывает и потерю способности бактерий к движению, хотя жгутики остаются при этом неповрежденными.
Движение бактерий. Виды расположения жгутиков. Функции фимбрий и пилей.
Виды расположения жгутиков: - Монотрихиальное
жгутикование
(Располагаются на поверхности тела бактерий по одиночке)
- Лофотрихиальное
жгутикование
(пучком на одном или обоих концах клетки)
- Перитрихиальное
жгутикование
(они могут находиться на всей поверхности клетки)
Фимбрии (от лат. firnbriac – бахрома), длинные, тонкие, прямые выросты, состоящие из гидрофобного белка и находящиеся в большом количестве (иногда до нескольких тысяч) на поверхности клеток грамотрицательных бактерий. Длина Ф. – до 12 мкм, толщина – не более 100 Å. Они значительно тоньше и короче жгутиков. "Мужские" клетки бактерий (доноры) могут иметь 1–3 половые Ф. (пили), образующие между ними и "женскими" клетками (реципиентами) полые мостики, через которые при конъюгации бактерий передаётся ДНК. Ф. могут быть как у подвижных, так и у неподвижных бактерий; возникают обычно из базального тельца, находящегося в цитоплазматической мембране, и проходят через клеточную стенку наружу. Ф. придают бактериальной клетке способность неспецифически "прилипать" к плотной поверхности клеток, тканей и т.п.
Пили (Pili), Фимбрии (Fimbriae)
- это:
тонкие полые нитевидные отростки, имеющиеся на поверхности ряда бактерий. Считается, что они способствуют прилипанию этих бактерий к поверхности других клеток. У некоторых бактерий выделяют специализированные фимбрии (F-фимбрии) (ред.), участвующие в процессе передачи ДНК во время конъюгации
Движение бактерий. Характер движения бактериальной клетки. Виды таксисов.
Различные вида таксисов:
Хемотаксис
– движение за счет влияния химических или питательных веществ.(Аттрактант –
привлекающий фактор, Репеллент –
отталкивающий фактор)
Фототаксис
– движение в зависимости от света, положительный фототаксис свойственен для фотосинтезирующих бактерий
Аэротаксис
– движение за счет воздуха (Аттрактантом для аэробных и репеллентом для анаэробных прокариот является молекулярный кислород.)
Магнитотаксис
– движение под влиянием соединений железа
Вискозитаксис
– способность реагировать на изменение вязкости раствора и перемещаться в направлении ее увеличения или уменьшения
Выделяют следующие виды движения: Плавание, Дрожание или кувыркание, Катание по слизи
Подвижные бактерии оставляют на субстрате особый налет – «роение». Это объясняется наличием у подвижных бактерий Н-антигена (от нем. Hauch
– налет), содержащегося в жгутиках. Неподвижные бактерии имеют только соматический антиген О-антиген (от нем. ohne
Hauch
– без налета).
У бактерий с перитрихиальным жгутикованием выявлены два вида двигательного поведения: прямолинейное движение и кувыркание, т.е. периодические и случайные изменения направления движения.
Если в клетке много жгутиков, все они при движении собираются в пучок, вращаясь в одном направлении.
Вращение жгутиков передается клетке, начинающей вращаться в противоположном направлении, и обеспечивает эффективное движение (плавание) в жидкой среде и более медленное перемещение по поверхности твердых сред.
Плавающее движение осуществляется клеткой, когда вращение жгутиков синхронизировано.
Если жгутики не синхронизированы, то движение бактерий напоминает кружение на одном месте, дрожание или кувыркание.
Обычно дрожание и плавание чередуются в зависимости от наличия аттрактанта, при увеличении концентрации аттрактанта дрожание подавляется, сменяясь плаванием.
Движение бактерий, имеющих жгутики, носит свободно-плавающий
характер: - плавание - синхронизированное, в сторону аттрактанта
- кувыркание – не синхронизировано, в сторону от репеллента
Движения спирохет весьма активны. Характер движения – вращательно-спиральный,
обусловленный сокращением осевой нити (аксостиля) клетки
Характер движения миксобактерий – скользящий,
что определяется соприкосновением выбрасываемой ими слизи с субстратом.
Цианобактерии
Лишены жгутиков, однако они тоже способны к движению за счет энергии градиента. Моторы сине-зеленых водорослей вращают белковые тяжи, спрятанные в периплазме
Жгутики есть во всех группах архей
, даже у группы термоплазм, лишенных клеточной стенки. Археи живут в экстремальных условиях, поэтому их жгутики устойчивы к экстремальным внешним воздействиям.
Бактериальное ядро. Строение, состав. Характеристика ДНК.
Бактериальное ядро представлено молекулами, имеющими наименьшее для клеточных организмов количество ДНК (0,4–0,8)•109
Наибольшее содержание ДНК среди прокариот обнаружено у нитчатых цианобактерий (8,5•109).
Каждая прокариотная клетка содержит 1 хромосому
При действии вредных факторов (температуры, рН среды, ионизирующего излучения, солей тяжелых металлов, некоторых антибиотиков) происходит образование множества копий хромосомы
При устранении воздействий, после перехода в стационарную фазу в клетках обнаруживается по одной копии хромосомы.
Из изложенного выше следует, что термины «нуклеоид» и «хромосома» не всегда совпадают. В зависимости от условий нуклеоид прокариотной клетки может состоять из одной или некоторого числа копий хромосомы.
Характеристика ДНК.
ДНК прокариот построена так же, как и эукариот. Молекула ДНК несет множество отрицательных зарядов, поскольку каждый фосфатный остаток содержит ионизированную гидроксильную группу. У эукариот отрицательные заряды нейтрализуются образованием комплекса ДНК с основными белками – гистонами.
В клетках подавляющего большинства прокариот не обнаружено гистонов
Нейтрализация зарядов осуществляется взаимодействием ДНК с полиаминами (спермином и спермидином), а также с ионами Mg2+.
У некоторых архебактерий и цианобактерий обнаружены гистоны и гистоно-подобные белки, связанные с ДНК.
Содержание пар оснований А+Т и Г+Ц в молекуле ДНК является постоянным для данного вида организма и служит важным диагностическим признаком. У прокариот молярная доля ГЦ в ДНК колеблется в очень широких пределах: от 23 до 75 %.
В среднем каждый ген состоит примерно из 1000 пар нуклеотидов, а вес одного нуклеотида ДНК составляет около 500 дальтон.
геном вируса гепатита В из четырех генов (S, С, Р и X)
геном вируса ВИЧ из 9 генов
геном бактериофага 0X174 из 9 генов
у бактериофага Т4 – из 200 генов
у F-плазмиды – из 90 генов (94,5 тыс. пар нуклеотидов), у хламидий – из 400-600 генов, у риккетсий – из 1000 генов
Бактериальное ядро. Особенности генетической системы бактерии. Типы репликации ДНК бактерии.
Хромосомы бактерий (и плазмид) располагаются свободно в цитоплазме, не отграничены от нее никакими мембранами, но связаны с определенными рецепторами на цитоплазматической мембране.
Хромосома особым компактным образом в ней упакована, ДНК находится в суперспирализованной форме и свернута в виде петель. Число петель 12-80 на хромосому, петли в центре нуклеоида объединяются за счет связывания ДНК с сердцевинной структурой, представленной молекулами особого класса РНК. Данная упаковка обеспечивает возможность транскрипции отдельных оперонов хромосомы, не препятствует ее репликации, а также петли упакованной хромосомы способствуют компартментализации рибосом.
Бактерии являются гаплоидными организмами, имеют один набор генов. Содержание ДНК у них непостоянно, оно может достигать значений, эквивалентных по массе от двух до восьми хромосом. Регулируя содержание копий своих генов, бактерии одновременно приспосабливают скорость своего размножения к условиям роста. Наряду с увеличением содержания ДНК у бактерий в этом случае существенно возрастает и количество рибосом. Благодаря этому возрастает суммарная скорость биосинтеза всех субклеточных и клеточных структур и, соответственно, скорость размножения бактерий.
У бактерий в естественных условиях передача генетической информации происходит не только по вертикали, т. е. от родительской клетки дочерним, но и по горизонтали с помощью различных механизмов: конъюгации, сексдукции, трансдукции, трансформации. У бактерий часто помимо хромосомного генома имеется дополнительный плазмидный геном, наделяющий их важными биологическими свойствами, нередко – специфическим иммунитетом к антибиотикам и химиопрепаратам.
Эти особенности определяют возможность накопления в окружающей среде большого запаса клеток данного вида, что является причиной высоких адаптационных способностей и существования бактерий в природе многие миллионы лет.
Типы репликации ДНК.
Вегетативная
репликация хромосомной и плазмидной ДНК обусловливает передачу генетической информации по вертикали, контролируется хромосомными и плазмидными генами.
Конъюгативная
репликация осуществляется при конъюгации, контролируется плазмидными генами.
Репаративная
репликация – механизм, посредством которого осуществляется устранение из ДНК структурных повреждений или заключительный этап генетической рекомбинации, контролируется хромосомными и плазмидными генами
Стабильная
репликация так названа потому, что происходит независимо от наличия или отсутствия синтеза белка.
-
Деление молекулы ДНК (репликация) происходит по полуконсервативному механизму и в норме всегда предшествует делению клетки. Репликация ДНК начинается в точке прикрепления кольцевой хромосомы к ЦПМ, где локализован ферментативный аппарат, ответственный за репликацию. Контакт ДНК с ЦПМ осуществляется посредством мезосом. . Репликация, начавшаяся в точке прикрепления, идет затем в двух противоположных направлениях, образуя характерные для кольцевой хромосомы промежуточные структуры.
Возникающие дочерние хромосомы остаются прикрепленными к мембране. Репликация молекул ДНК происходит параллельно с синтезом мембраны в области контакта ДНК с ЦПМ. Это приводит к разделению (сегрегации) дочерних молекул ДНК и оформлению обособленных хромосом
Бактериальное ядро. Виды деления бактериальной клетки. Процесс деления.
Виды деления бактериальной клетки:
Неравновеликое бинарное деление, или почкование
. При почковании на одном из полюсов материнской клетки образуется маленький вырост (почка), увеличивающийся в процессе роста. Постепенно почка достигает размеров материнской клетки, после чего отделяется от последней. Клеточная стенка почки полностью синтезируется заново. В процессе почкования симметрия наблюдается в отношении только продольной оси. При почковании материнская клетка дает начало дочерней клетке, и между ними можно в большинстве случаев обнаружить морфологические и физиологические различия: есть старая материнская клетка и новая дочерняя.
Размножение путем множественного деления
, характерное для одной группы одноклеточных цианобактерий, в результате образуются мелкие клетки, получивших название баеоцитов (греч. bae
– маленькая, cyto
– клетка), число которых у разных видов колеблется от 4 до 1000. Освобождение баеоцитов происходит путем разрыва материнской клеточной стенки. В основе множественного деления лежит принцип равновеликого бинарного деления. Отличие заключается в том, что в этом случае после бинарного деления не происходит роста образовавшихся дочерних клеток, а они снова подвергаются делению.
Процесс деления.
Процесс деления бактериальной клетки начинается с репликации хромосомной ДНК. В точке прикрепления хромосомы к цитоплазматической мембране (точке-репликаторе) действует белок-инициатор, который вызывает разрыв кольца хромосомы, и далее идет деспирализация ее нитей. Нити раскручиваются, и вторая нить прикрепляется к цитоплазматической мембране в точке-прорепликаторе, которая диаметрально противоположна точке-репликатору. За счет ДНК-полимераз по матрице каждой нити достраивается точная ее копия. Удвоение генетического материала – сигнал для удвоения числа органелл. В септальных мезосомах идет построение перегородки, делящей клетку пополам.
Двухнитевая ДНК спирализуется, скручивается в кольцо в точке прикрепления к цитоплазматической мембране. Это является сигналом для расхождения клеток по септе. Образуются две дочерние особи.
Размножение бактерий определяется временем генерации. Это период, в течение которого осуществляется деление клетки. Продолжительность генерации зависит от вида бактерий, возраста, состава питательной среды, температуры и др.
Фазы размножение бактериальной клетки на жидкой питательной среде:1) начальная стационарная фаза; то количество бактерий, которое попало в питательную среду и в ней находится;2) лаг-фаза (фаза покоя); продолжительность – 3–4 ч, происходит адаптация бактерий к питательной среде, начинается активный рост клеток, но активного размножения еще нет; в это время увеличивается количество белка, РНК;3) фаза логарифмического размножения; активно идут процессы размножения клеток в популяции, размножение преобладает над гибелью;4) максимальная стационарная фаза; бактерии достигают максимальной концентрации, т. е. максимального количества жизнеспособных особей в популяции; количество погибших бактерий равно количеству образующихся; дальнейшего увеличения числа особей не происходит;5) фаза ускоренной гибели; процессы гибели преобладают над процессом размножения, так как истощаются питательные субстраты в среде. Накапливаются токсические продукты, продукты метаболизма. Этой фазы можно избежать, если использовать метод проточного культивирования: из питательной среды постоянно удаляются продукты метаболизма и восполняются питательные вещества.
Бактериальное ядро. Формы обмена генетической информацией у бактерий. Изменчивость бактерий.
Формы обмена генетической информацией у бактерий по горизонтали
- Трансформация
- перенос генетического материала, заключающийся в том, что бактерия-реципиент захватывает (поглощает) из внешней среды фрагменты чужеродной ДНК.
Трансформация бывает:
- Индуцированная (искусственно получаемая) трансформация происходит при добавлении к культуре бактерий очищенной ДНК, полученной из культур тех бактерий, генетические признаки которых стремятся передать исследуемой культуре.
- Спонтанная трансформация происходит в естественных условиях и проявляется в возникновении рекомбинантов при смешивании генетически различающихся клеток. Она протекает за счет ДНК, выделяющейся клетками в окружающую среду вследствие их лизиса или в результате активного выделения ДНК жизнеспособными клетками-донорами.
Трансфекция
– вариант трансформации бактериальных клеток, лишенных клеточной стенки, осуществляемый вирусной (фаговой) нуклеиновой кислотой.
С помощью трансфекции удается вызвать у таких бактерий (без клеточной стенки) вирусную инфекцию. Трансфекцию можно осуществить и с другими (не бактериальными) клетками, если ввести в них чужеродную ДНК, способную рекомбинировать с ДНК этих клеток, или воспроизводить вирионы, или самостоятельно реплицироваться.
Трансдукция-
перенос генетического материала от клетки-донора клетке-реципиенту с помощью бактериофагов. Различают трансдукцию неспецифическую и специфическую.
Трансдукция бывает:
Неспецифическая трансдукция
- случайный перенос фрагментов ДНК от одной бактериальной клетки к другой.
Специфическая трансдукция
осуществляется только умеренными фагами, способными включаться в строго определенные участки хромосомы бактериальной клетки и переносить определенные гены.
Конъюгация.
Под конъюгацией понимают процесс обмена генетическим материалом (хромосомным и плазмидным), осуществляемый при непосредственном контакте клеток донора и реципиента.
-
Этот процесс контролируется только конъюгативными плазмидами, имеющими совокупность генов, называемую tra-опероном (tra – от англ.,
transfer
– перенос). Этот оперон контролирует синтез аппарата переноса, конъюгативную репликацию и явление поверхностного исключения. Аппаратом переноса являются специальные донорные ворсинки, с помощью которых устанавливается контакт между конъюгирующими клетками. Донорные ворсинки представляют собой длинные (1-20 мкм) тонкие трубчатые структуры белковой природы с внутренним диаметром около 3 нм.
Стадии конъюгации
1.
установление контакта между донором и реципиентом
2.
протаскивание нити ДНК от донора к реципиенту
3.
достройка перенесенной нити ДНК комплементарной ей нитью в реципиентной клетке
4.
рекомбинация между переданной хромосомой (ее фрагментами) и хромосомой клетки-реципиента
5.
размножение мерозиготы
6. образование клеток, несущих признаки донора и реципиента
Изменчивость бактерий
Бактерии в силу относительной простоты их организации и короткого срока жизни подвергаются изменчивости быстрее, чем многие другие организмы. В основе их изменчивости лежат мутации и генетические рекомбинации, особенно протекающие с участием транспонируемых элементов.
Мутации – изменения в генотипе, которые стабильно наследуются.
Мутации могут быть спонтанными или индуцированными.
Индуцированные мутации происходят с гораздо большей частотой, они возникают в результате воздействия различных мутагенов – физических и химических факторов, повреждающих ДНК: ионизирующая радиация, УФ облучение, различные аналоги оснований, ДНК алкилирующие соединения, акридины, антибиотики
Точечные мутации могут быть обусловлены: заменой оснований, выпадением (делецией) основания, появлением дополнительного основания (вставки)
Нарушение генома может быть следствием: протяженных делеций, инверсии (поворот сегмента хромосомы на 180°), транслокации (перемещение участка хромосомы из одной позиции в другую). Все это также будет приводить к изменению и нарушению различных функций клетки (организма).
- Большая роль в изменчивости бактерий и других организмов принадлежит так называемым транспонируемым генетическим элементам, то есть генетическим структурам, способным в интактной форме перемещаться внутри данного генома или переходить от одного генома к другому, например от плазмидного генома к бактериальному и наоборот. Различают три класса транспонируемых элементов: IS
-элементы, транспозоны
и эписомы
.
- Вставочные последовательности
(от англ,
insertionsequence), имеют обычно размеры, не превышающие 2 тыс. пар оснований, или 2 к.б. (килобаза – тысяча пар оснований).
- IS-элементы несут только один ген, кодирующий белок транспозазу, с помощью которой IS-элементы встраиваются в различные участки хромосомы. Их обозначают цифрами: IS1, IS2, IS3 и т. д.
Транспозоны
- представляют собой более крупные сегменты ДНК, фланкированные инвертированными IS-элементами.
- способны встраиваться в различные участки хромосомы или переходить из одного генома в другой, т. е. ведут себя как IS-элементы
- помимо генов, обеспечивающих их перемещение, они содержат и другие гены, например гены лекарственной устойчивости.
- Транспозоны обнаружены в геномах плазмид, вирусов, прокариот и эукариот и их, как и IS-элементы, обозначают порядковым номером: Tп1, Тп2, ТпЗ и т. д.
Бактериальное ядро. Плазмиды. Биологическая роль, отличия от вирусов, виды плазмид.
Плазмида
или R-фактор (от англ. resistance
– устойчивость) (экстрахромосомный генетический элемент) представляет собой репликон, который стабильно наследуется в экстрахромосомном состоянии.
Плазмиды имеют собственные гены, которые наделяют их специфическими наследственными признаками и способностью к размножению, должны быть отнесены к живым организмам.
Плазмиды
– наипростейшие организмы, лишенные оболочки, собственных систем синтеза белка и мобилизации энергии и представляющие собой особый класс абсолютных внутриклеточных паразитов, наделяющих своих бактерий-хозяев полезными для них свойствами
Сходство с вирусами
-
Плазмиды не имеют собственной белоксинтезирующей системы
-
Не имеют собственной системы мобилизации энергии;
-
Плазмиды не способны к росту и бинарному делению
-
Плазмиды размножаются путем воспроизведения себя из собственного генома (путем его саморепликации)
-
Являются абсолютными внутриклеточными паразитами
Отличия от вирусов:
| Плазмиды |
Вирусы |
Геном представлен
только двунитевой ДНК
|
Имеется более 10 вариантов
РНК- и ДНК-геномов
|
Не имеют оболочки
– представляют собой
«голые» геномы
|
Имеют белковую оболочку –
капсид или суперкапсид
|
Размножение только
путем саморепликации
|
Репродукция требует
дополнительной системы
синтеза и сборки белка
|
Средой обитания - бактерии
|
Среда обитания – любые клетки
|
Наделяют хозяина важными
свойствами, повышающими
жизнеспособность
|
Инфицируют,
используют, уничтожают хозяина
|
КЛАССИФИКАЦИЯ ПЛАЗМИД
| Группа плазмид
|
Свойства
|
| F-плазмиды |
Донорные функции |
| R-плазмиды |
Устойчивость к лекарственным препаратам |
| Col-плазмиды |
Синтез колицинов |
| Ent-плазмиды |
Синтез энтеротоксннов |
| Hly-плазмиды |
Синтез гемолизинов |
| Биодеградативные плазмиды |
Разрушение различных органических и неорганических соединений, в том числе содержащих тяжелые металлы |
| Криптические плазмиды |
Неизвестны |
- Саморегулируемая репликация
. Эта функция свойственна всем живым организмам. В составе плазмидных ДНК имеются фиксированная точка ori (точка начала репликации) и соответствующие гены, контролирующие репликацию.
- Явление поверхностного исключения
. Этот механизм не позволяет проникнуть в клетку, уже содержащую плазмиду, другой родственной ей плазмиде. Поверхностное исключение обеспечивается синтезом под контролем генов плазмиды особых белков наружной мембраны, которые препятствуют установлению контакта этой клетки с клеткой, несущей такую же плазмиду, или подавляют конъюгативный метаболизм ДНК этой плазмиды
- Явление несовместимости
. Суть его заключается в том, что две близкородственные плазмиды не могут стабильно сосуществовать в одной клетке, одна из них подвергается элиминации (удалению).
Контроль числа копий плазмиды на хромосому клетки
. Различают малокопийные (1-4 копии) и многокопийные плазмиды (12-38 копий). Наличие собственных генов позволяет плазмиде осуществлять репликацию независимо от клеточного цикла клетки-хозяина. Информация, необходимая для осуществления репликации плазмиды, обычно заключена в небольшой участок ее ДНК, получивший название основного, или базового репликона
- Контроль стабильного сохранения плазмид в клетке-хозяине (контроль стабильного поддержания
).
- Контроль равномерного распределения дочерних плазмид в дочерние бактериальные клетки
.
- Распределение плазмид между дочерними клетками происходит нерандомически, т. е. не по принципу случайности, а существует генетический механизм контроля равномерного распределения (сегрегации) вновь синтезированных плазмид при клеточном делении.
- Носительство плазмид для клетки-хозяина становится генетически необходимым, благодаря этому обеспечивается существование плазмид как организмов.
- Способность к самопереносу
(у конъюгативных плазмид).
- Способность к мобилизации на перенос
(у неконъюгативных плазмид).
- Способность наделять клетку-хозяина дополнительными важными для нее биологическими свойствами, способствующими выживанию бактерий, а следовательно, и плазмид в природе.
фертильность; антибиотикорезистентность;устойчивость к тяжелым металлам (кадмий, ртуть); способность к продукции бактериоцинов, антибиотиков, токсинов; способность к утилизации необычных источников углерода (камфора, октан, октанол).
Морфологическая дифференцировка прокариот. Формы клеток. Покоящиеся формы. Процесс поддержания состояния покоя.
Покоящиеся формы (обеспечение выживания в течение длительного времени в неблагоприятных условиях)
- эндоспоры
(у грамположительных эубактерий),
- цисты
(азотобактера, спирохет, миксобактерии, риккетсий),
- акинеты
(у цианобактерий),
- миксоспоры
(у миксобактерий),
- экзоспоры
(у некоторых бактерий, грибов)
Для всех покоящихся форм характерна повышенная по сравнению с вегетативными клетками устойчивость к действию разнообразных повреждающих факторов: высоких и низких температур, обезвоживанию, высокой кислотности среды, радиации, механических воздействий и др. В наибольшей степени устойчивость проявляется у эндоспор
| Тип покоящихся клеток
|
Повреждающий фактор
|
| Высокая температура
|
Высушивание
|
| Миксоспоры миксобактерий |
Гибель 90% после выдерживания при 50 о
С в течение 20 мин |
Гибель 50% после хранения в течение 6 сут |
| Цисты азотобактера |
100%-я гибель после выдерживания при 60 °С в течение 15 мин |
100%-я жизнеспособность при хранении в течение 12 сут |
| Акинеты цианобактерий |
Гибель 95% после выдерживания при 40 о
С в течение 10 мин |
95%-я жизнеспособность после хранения в течение 15мес при 4 °С |
| Эндоспоры некоторых эубактерий |
Гибель 90% после выдерживания при 100 о
С в течение 11 мин |
Жизнеспособность сохранялась в течение приблизительно 1000 лет |
| Эндоспоры актиномицетов |
Гибель 99% после выдерживания при 75 °С в течение 70 мин |
Жизнеспособность сохранялась в течение 14 лет |
Факторы обеспечивающие устойчивость
- нахождение споровой цитоплазмы в обезвоженном состоянии
- термостойкость споровых ферментов
- наличие дипиколиновой кислоты
- большое количества двухвалентных катионов
- поверхностные структуры, механически защищающие содержимое споры от проникновения извне агрессивных веществ.
Факторы, приводящие к образованию покоящихся форм
- Формированию цист у миксобактерий способствует наличие в среде глицерина, аминокислот.
- Количество цист азотобактера возрастает при добавлении к среде параоксимасляной кислоты и повышении концентрации двухвалентных катионов.
- В качестве факторов, индуцирующих формирование акинет цианобактерий, отмечены низкая температура, высушивание, отсутствие в среде связанного азота и увеличение содержания глутаминовой кислоты.
Гипотезы, объясняющие механизмы, поддержания специализированных клеток в состоянии покоя:
1. В покоящихся клетках имеются вещества, ингибирующие ферменты, блокирующие метаболизм и предотвращающие прорастание спор.
2. Особая структура спор обеспечивает поддержание ее сердцевины в обезвоженном состоянии.
3. Ферменты спор находятся в неактивной форме. Изменения их конфигурации, приводящие к активированию ферментов, выводят покоящуюся клетку из этого состояния.
Морфологическая дифференцировка прокариот. Строение эндоспоры. Химический состав, слои. Бактерии, способные к спорообразованию.
Факторы, приводящие к образованию эндоспор
- наличие или отсутствие определенных питательных веществ среды
- Неприемлемая температура
- Неблагоприятная кислотность среды
- Неблагоприятные условия аэрирования
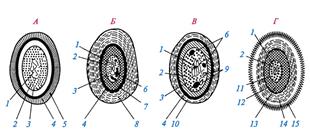
Строение покоящихся форм прокариот:
А - миксоспоры миксобактерий;
Б - цисты азотобактера;
В - акинеты цианобактерий;
Г - эндоспоры;
1 - нуклеоид; 2 - цитоплазма; 3 -
ЦПМ ; 4 -клеточная стенка; 5 - капсула; 6 - гранулы запасных веществ; 7 - внутренние покровы (интина); 8 - внешние покровы (экзина); 9 -
тилакоиды ; 10 -
чехол ; 11 - внутренняя мембрана споры; 12 - наружная мембрана споры; 13 - кортекс; 14 - покровы споры, состоящие из нескольких слоев; 15 - экзоспориум.
Экзоспориум состоит из липидов и белков и, вероятно, выполняет функцию дополнительного барьера, защищающего спору от внешних воздействий, а также регулирующего проникновение в нее различных веществ. Однако, никаких данных, подтверждающих эти предположения, пока нет. Механическое удаление экзоспориума не приводит к какому-либо повреждению споры. Они обнаруживают такую же способность к прорастанию, как и споры с неудаленным экзоспориумом.
Споровые покровы в основном состоят из белков и в небольшом количестве из липидов и гликолипидов. Белки покровов обладают высокой устойчивостью к неблагоприятным условиям и обеспечивают спорам защиту от действия литических ферментов, других поврждающих факторов, а также предохраняют от преждеременного прорастания. Оказалось, что споры мутантов, лишенные покровов, прорастают сразу же после выхода из иатеринской клетки, даже если условия для последующего роста неблагоприятны. Кортекс построен в основном из молекул особого типа пептидогликана. При прорастании споры из части кортекса, прилегающей к внутренней споровой мембране, формируется клеточная стенка вегетативной клетки.
Бактерии, способные к спорообразованию:
Bacillus, Clostridium, Sulfobacillus, Sporosarcina, Thermoactinomyces, Sporospirillum
Морфологическая дифференцировка прокариот. Биохимические и физиологические изменения в процессе прорастания эндоспоры. Факторы устойчивости эндоспор в окружающей среде.
Прорастание – это процесс, сопровождающийся сложными физиологическими и биохимическими изменениями.
Начинается он с интенсивного поглощения спорой воды и набухания.
Происходит активация литических ферментов.
Резко возрастает дыхание, т.е. мобилизуется энергия. Происходят изменения в химическом составе (удаляется дипиколиновая кислота). Идет активный синтез белка и РНК.
Происходят процессы репарации повреждений ДНК, произошедших в период покоя споры. За время прорастания споры теряют до 1/3 первоначальной массы. Последующие этапы состоят в разрушении кортекса, выхода ростовой трубочки, достраивания клеточной стенки и последующим делении.
Факторы обеспечивающие устойчивость
- нахождение споровой цитоплазмы в обезвоженном состоянии
- термостойкость споровых ферментов
- наличие дипиколиновой кислоты
- большое количества двухвалентных катионов
- поверхностные структуры, механически защищающие содержимое споры от проникновения извне агрессивных веществ.
Морфологическая дифференцировка прокариот. Формирование споры, слои эндоспоры.
Образование эндоспор – уникальный процесс, имеющий место только в мире прокариот. Бактериальные эндоспоры – это особый тип покоящихся клеток округлой или эллиптической формы грамположительных эубактерий, формирующихся эндогенно, т. е. внутри цитоплазмы «материнской» клетки (спорангия)
Эндоспоры обладают специфическими структурами и свойствами:
многослойные белковые покровы
наружная и внутренняя мембраны
кортекс
устойчивость к высоким температурам и дозам радиации, летальным в норме для вегетативных клеток
особое физическое состояние протопласта
Схема строения зрелой бактериальной споры:
1 – экзоспориум;
2 – наружная оболочка споры;
3 – внутренняя оболочка споры;
4 – кортекс; 5 – клеточная стенка зародыша;
7 – цитоплазматическая мембрана;
7 – цитоплазма с ядерным веществом
Способностью образовывать внутри клетки особое тельце – спору
(эндоспору) обладают только палочковидные бактерии, относящиеся к родам Bacillus
,
Clostridium
и др. Исключением являются лишь сарцина, разлагающая мочевину (Sporosarcina
urea
), и некоторые извитые бактерии, способные также образовывать споры. В каждой бактериальной клетке, как правило, формируется одна эндоспора.
Спорообразование: стадии
1.образование проспоры
– структуры, расположенной внутри материнской клетки и полностью отделенной от нее двумя элементарными мембранами: наружной и внутренней
2. формируется впячивание внутрь клетки ЦПМ
3. обособление от клеточного содержимого с помощью перегородки
4. у одного из полюсов клетки происходит уплотнение цитоплазмы
5. образование тяжа вдоль длинной оси спорулирующей клетки
6. изменение морфологии ядерного вещества вегетативной клетки
7. мембрана нарастает от периферии к центру
8. образование споровой перегородки
9. «обрастание» отсеченного участка клеточной цитоплазмы с ядерным материалом
- Выделяют 3 типа спорообразования:
- бациллярный
(спора уложена посредине клетки, расширяя ее центральную часть);
- клостридиальный
(спора значительно расширяет центральную часть клетки, которая имеет зауженные концы, из-за чего спороносная клетка похожа на ветерено);
- плектридиальный
(спора занимает один из концов клетки, клетка похожа на теннисную ракетку)
После образования проспоры дальнейшие этапы спорообразования необратимы. Между наружным и внутренним мембранными слоями просторы начинается формирование кортикального слоя (кортекса
).
Затем поверх наружной мембраны проспоры синтезируются споровые покровы, состоящие из нескольких слоев, число и строение которых различны у разных видов спорообразующих бактерий. В формировании слоев споровых покровов принимают участие наружная мембрана споры и протопласт материнской клетки.
У многих бактерий поверх покровов споры формируется еще одна структура – экзоспориум
, строение которой различается в зависимости от вида бактерий. Часто экзоспориум многослоен, с характерной для каждого слоя тонкой структурой.
Все слои, окружающие протопласт эндоспоры, находятся внутри материнской клетки. На их долю приходится примерно половина сухого вещества споры.
После сформирования споры происходит разрушение (лизис) «материнской» клеточной стенки и спора выходит в среду.
Спорообразование сопровождается активным синтезом белка. Белки эндоспор в отличие от белков вегетативных клеток богаты цистеином и гидрофобными аминокислотами, с чем связывают устойчивость спор к действию неблагоприятных факторов.
Содержание ДНК в споре несколько ниже, чем в исходной вегетативной клетке, поскольку в спору переходит лишь часть генетического материала материнской клетки. Генетический материал поступает в спору в виде полностью реплицированных молекул ДНК. Споры некоторых видов содержат по 2 или 3 копии хромосомы. Содержание РНК в спорах ниже, чем в вегетативных клетках, и РНК в значительной степени при спорообразовании синтезируется заново. Одним из характерных процессов, сопровождающих образование эндоспор, является накопление в них дипиколиновой кислоты и ионов кальция.
Классификация и систематика бактерий. Классификация бактерий по Берджи. Признаки, используемые при описании бактерий. Характеристика основных групп бактерий по классификатору Берджи.
Для растений, животных и бактерий применяется бинарная номенклатура:
родовое и видовое название. Классификация микроорганизмов, причисляемых к прокариотам, учитывает в первую очередь практические моменты и служит для распознавания описанных форм. Классификация
– это распределение единиц по группам более высокого порядка. Она осуществляется по иерархической схеме.
Основной единицей является чистая культура выделенной бактерии – «штамм». Штамм
– это различные культуры одного вида микроорганизмов, отличающиеся по незначительным признакам и выделенные из разных мест обитания. Штаммы объединяются в виды (species
). Вид
– совокупность микроорганизмов, имеющих единое происхождение и генотип, сходных по морфологическим и биологическим свойствам, обладающих наследственно закрепленной способностью вызывать в среде естественного обитания качественно определенные специфические процессы.
Виды объединяются в роды (genus
, множественное число genera
), а роды в семейства (латинские названия семейств оканчиваются на -асеае
). Кроме этого, роды могут быть объединены в подгруппы, порядки. Высшая таксономическая единица – группа. Основой для классификации служит адекватное описание штаммов, в соответствии с которым проводят сравнение и разграничение рассматриваемых единиц.
Установлением соподчинения отдельных групп организмов занимается таксономия.
Следует различать два вида классификаций: филогенетические, или «естественные», и искусственные. Построение естественной
классификации – конечная цель таксономии бактерий, которая состоит в том, чтобы объединить родственные формы, связанные общностью происхождения, и на этой основе создать филогенетическое древо бактерий.
Искусственная классификация
ставит перед собой более скромные цели, чем филогенетическая. Она довольствуется объединением организмов в отдельные группы на основе их сходства и используется для идентификации и распознавания (определения) организмов. Искусственная система рассчитана на использование ее в качестве ключа для определения. В современной микробиологии принята Международная классификация, представленная в определителе бактерий «Berdgy'sManualofdeterminativebacteriology», который был подготовлен международным коллективом авторов (9-е издание, 1994 года, выпущено в 1997 г. в России).
В соответствии с новым кодексом номенклатуры бактерий, действующим с 1 января 1980 года, существуют следующие классификационные категории – Царство Прокариоты, далее бактерии делятся: отдел – класс – порядок – семейство – род – вид. В девятом издании определителя бактерий Берджи бактерии подразделяются по фенотипическим признакам на четыре основные категории, содержащие в себе 35 групп.
Для классификации используют следующие признаки бактерий:
морфологические
(форма, размеры клетки, подвижность, характер образования жгутиков, способность к спорообразованию, окраска по Граму);
физиологические
(отношение к различным углеводам, кислороду, тип питания, способы получения энергии, характер образующихся продуктов в процессе обмена);
 культуральные
(характер роста на различных питательных средах: наличие пленки, мути, осадка на жидких средах, тип колоний, их особенности на плотных средах и т. п.). культуральные
(характер роста на различных питательных средах: наличие пленки, мути, осадка на жидких средах, тип колоний, их особенности на плотных средах и т. п.).
биохимические (генотипические)
признаки, т. е. особенности нуклеотидного состава ДНК. Особи одного и того же вида имеют одинаковый состав оснований ДНК, а у видов, принадлежащих к одному роду, нуклеотидный состав ДНК имеет близкие значения.
При описании указывается:
отношение к кислороду, т.е. растут ли клетки в аэробных или в анаэробных условиях, либо в тех и других;
каким образом они получают необходимую энергию – в процессе дыхания, брожения или фотосинтеза;
зависимость роста от температуры и рН, с указанием оптимума и пределов устойчивости;
усваиваемые клетками питательные вещества;
места обитания;
наличие симбиотических или паразитических взаимоотношений;
клеточные включения, пигментация и состав капсулы;
компоненты клеточной стенки (пептидогликановый остов, липополисахариды, тейхоевая кислота);
серологическая дифференциация (поверхностные антигены, гомологичные белки);
состав оснований ДНК;
гибридизация ДНК, трансформируемость при межвидовом переносе ДНК;
последовательность нуклеотидов в 16S или 5S рРНК;
чувствительность к антибиотикам.
Категории бактерий:
Грациликуты (Gracilicutes
(от лат. cutes
– кожа, gracilis
– тонкий, стройный)).
Фирмикуты (Firmicutes
(от лат. firmis
– крепкий, cutes
– кожа)).
Тенерикуты (Tenericutes
(
от лат.Tener
–
отсутствую))
Мендозикуты (Mendosicutes
относят необычные микроорганизмы класса Archaebacteria
.)
Классификация и систематика бактерий. Категории бактерий. Особенности эубактерий и архибактерий.
Категория
I
– Грамотрицательные (грам -) эубактерии, имеющие клеточные стенки
Прокариоты со сложным (грамотрицательного типа) строением клеточных стенок. С
тенки состоят из наружной мембраны, тонкого внутреннего слоя пептидогликана и дополнительно различных других компонентов снаружи или между этими двумя слоями. Обычно окрашиваются по Граму отрицательно. Клетки могут быть сферической или овальной формы, в виде прямых или изогнутых палочек, спиралей либо нитей; некоторые из этих форм могут быть окружены чехлом или капсулой. Размножение происходит путем бинарного деления, однако для некоторых групп характерно почкование и у редких организмов (
Pleurocapsales
)
имеет место множественное деление.
Миксобактерии могут образовывать плодовые тела и миксоспоры. Возможна подвижность – «плавание», скользящее движение – либо неспособность к движению. В эту категорию входят фототрофы и нефототрофы (как литотрофы, так и гетеротрофы); среди них известны аэробы, анаэробы, факультативные анаэробы и микроаэрофилы. Н
екоторые представители – облигатные внутриклеточные паразиты.
Класс 1
– Scotobacteria
. К классу Scotobacteria
(от греч. scotos
– темнота), относят грамотрицательные нефотосинтезирующие бактерии. Данный класс подразделяется на 10 групп.
Группа 1 – спирохеты. Эти микроорганизмы объединены в порядок Spirochaetales
, семейства Spirochaetaceae
и Leptospiraceae
.
Группа 2 – аэробные спиральные и виброидные грамотрицательные бактерии, включает семейство Spirillaceae
.
Группа 3 – аэробные грамотрицательные палочки и кокки. В группу входят семейства Pseudomonadaceae
,
Azotobacteriaceae
,
Rhizobiaceae
,
Methylococcaceae
,
Acetobacteriaceae
,
Neisseriaceae
.
Группа 4 – факультативно анаэробные грамотрицательные палочки. Эта группа грамотрицательных бактерий включает традиционные семейства Enterobacteriaceae
,
Vibrionaceae
и Pasteurellaceae
, а также несколько других родов факультативно анаэробных палочек.
Группа 5 – анаэробные грамотрицательные прямые, изогнутые и спиральные палочки представлена семейством Bacterioides
и бактериями, осуществляющими диссимиляционное восстановление сульфата или серы.
Группа 6 – грамотрицательные хемолитотрофные бактерии, объединенные в два семейства Nitrobacteriaceae
и Siderocapsaceae
и 15 родов.
Группа 7 – скользящие бактерии. К группе относят два порядка – Myxobacteriales
и Cytophagales
. К первому порядку относятся миксобактерии, образующие плодовые тела. Ко второму порядку относят скользящих бактерий, не способных к образованию плодовых тел.
Группа 8 – хламидобактерии. Это бактерии, имеющие чехлы или влагалища. В группу отнесены семь родов.
Группа 9 – почкующиеся и/или стебельковые бактерии. Эти бактерии образуют особые выросты – простеки, стебельки, имеют клетки причудливых необычных форм – звездчатая, веретеновидная, яйцевидная и другие. Как правило, имеют некоторые особенности в жизненном цикле.
Группа 10 – риккетсии и хламидии. Группа представлена двумя порядками – Rickettsiales
и Chlamydiales
.
Эти порядки объединяют большое количество внутриклеточных паразитов, вызывающих инфекции человека и животных.
Класс 2
–
Anoxyphotobacteria
. Класс представлен аноксигенными фототрофными бактериями, разделенными на два порядка: Rhodospirillales
(пурпурные бактерии) и Chlorobiales
(зеленые бактерии).
Класс 3
–
Oxyphotobacteria
–
организмы, у которых фотосинтез сопровождается выделением молекулярного кислорода. Порядок Cyanobacteriales
– широко распространенная и основательно изученная группа сине-зеленых водорослей. Порядок Prochlorales
– одноклеточные прокариоты, отличающиеся от цианобактерий набором фотосинтетических пигментов.
Категория
II
– Грамположительные (грам+) эубактерии, имеющие клеточные стенки
Прокариоты с клеточными стенками грамположительного типа; окраска по Граму обычно, хотя и не всегда, положительная.
Форма клеток может быть сферической, палочковидной или нитевидной; палочки и нити могут быть неветвящимися, но у многих организмов имеется настоящее ветвление.
Размножение обычно происходит путем бинарного деления; некоторые представители образуют споры в качестве покоящихся форм (эндоспоры или споры на гифах).
В большинстве нефотосинтезирующие организмы; обычно это хемосинтезирующие гетеротрофы, среди которых есть аэробы, анаэробы, факультативные анаэробы и микроаэрофилы.
К этой категории относятся простые аспорогенные и спорообразующие бактерии, а также актиномицеты и родственные им организмы.
Недавно был выделен род Heliobacterium
, представители которого –
грамположительные гетеротрофные почвенные аэробы, способные к азотфиксации и фотосинтезу.
Класс 1
–
Firmibacteria
.
Класс 2
–
Tallobacteria
. Включает актиномицеты и родственные организмы с достаточно разнообразными характеристиками.
Категория
IV
–
Архебактерии
Архебактерии – это преимущественно почвенные или водные микроорганизмы, которые встречаются в анаэробных условиях или в гиперсоленых, либо гидро- и геотермально нагретых средах и, кроме того, как симбионты в пищеварительном тракте животных.
В эту группу входят аэробы, анаэробы и факультативные анаэробы, способные расти как хемолитоавтотрофы, гетеротрофы или факультативные гетеротрофы.
Архебактерии могут быть мезофилами или термофилами, причем некоторые виды способны расти даже при температуре выше 100 °С.
Результаты окрашивания по Граму могут быть положительными или отрицательными внутри одного порядка, поскольку типы клеточных оболочек сильно различаются.
Форма клеток может быть разнообразной – сферической, спиральной, пластинчатой или палочковидной. Встречаются также одноклеточные и многоклеточные формы в виде нитей или агрегатов.
Диаметр клеток варьирует в пределах 0,1–15 мкм, а длина нитей может достигать 200 мкм.
Размножение происходит путем бинарного деления, почкования, перетяжки, фрагментации или неизвестными способами.
|
