Министерство образования Республики Беларусь
Учреждение образования
«Гомельский государственный университет
им. Ф. Скорины»
Биологический факультет
Генетико-эволюционные и таксономические взаимоотношения у видов-двойников
Drosophila
группы
virilis
Палеарктики
Курсовая работа
Исполнитель:
Студентка группы К-42 ____________ Лапухова А.Ю.
Научный руководитель: Загрушевская Т.Е.
Гомель 2005
Содержание
Введение
1 Материалы и методы
2 Результаты и обсуждение
3 Электрофоретический ключ для типировки взрослых особей
Заключение
Литература
Введение
Более полувека виды-двойники Drosophila группы virilis являются одними из основных объектов для различных генетических исследований. Представители этой группы двойниковых видов успешно использовались в качестве модельной системы для изучения процессов видообразования (Patterson, Stone, 1952; Майр,1963; Гончаренко и др., 1989), генетики развития (Корочкин и др., 1975), молекулярной эволюции (Nei, 1971; Throckmorton, 1982; Spicer, Bell, 2002), а также таксономии и систематики (Гончаренко, Емельянов, 1990; Goncharenko, Emeljanov,1992). В пределах группы virilis в настоящее время насчитывается 14 видов-двойников, которые подразделяются на две филады – филада virilis и montana. Несмотря на то, что виды Drosophila группы virilis широко использовались в генетических экспериментах многие вопросы касающиеся таксономических и эволюционно-филогенетических взаимоотношений между ними остаются предметом дискуссий.
Целью данной работы является анализ генетико-эволюционных и таксономических взаимоотношений у видов-двойников Drosophila группы virilis, обитающих в природных популяциях Палеарктики с использованием генов, кодирующих различные изоферменты.
1. Материалы и методы
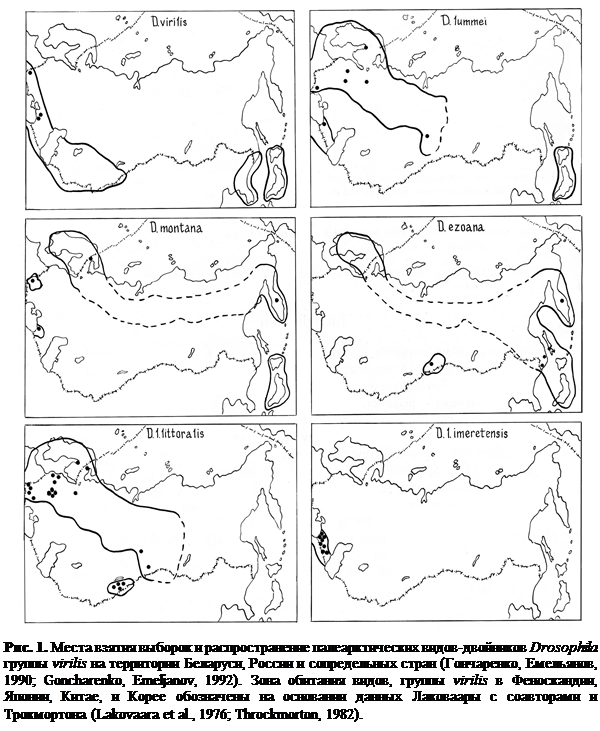
Ареалы распространение палеарктических видов-двойников Drosophila группы virilis обитающих в природных популяциях на территории России, Беларуси и сопредельных стран, а также месторасположение проанализированных популяций показаны на рис. 1. D. virilis Sturtevant встречается только в южных районах Палеарктики на винных и соковых заводах. По данному виду проанализированы особи из 3 популяций (Украина – Закарпатье, Россия – Краснодарский край). Другие 4 вида обитают в естественных лесах, вблизи незагрязненных рек и водоемов. D. lummei Hackman распространена севернее D. virilis, однако ареалы этих двух видов отчасти перекрываются в районах Закарпатья, Кавказа и Японских островов. Особи D. lummei взяты для анализа из 7 популяций (Молдавия, Беларусь, Финляндия, Россия – Краснодарский край, Московская и Новосибирская обл.). D. montana Stone, Griffen, Patterson, по-видимому, имеет типичный бореомонтанный ареал. Выборка особей этого вида была сделана в 4 популяциях (Украина – Карпаты, Россия – Краснодарский край, Карелия, Камчатка). Представители вида D. ezoana Takada, Okada отловлены и проанализированы нами в 5 популяциях (Алтай, Приморский край, Камчатка, Сахалин). Подвид D. littoralis littoralis Meigen исследован в 20 популяциях (Украина, Беларусь, Финляндия, Кыргызстан, Россия – Карелия, Алтай, Московская и Новосибирская обл.,). Особи другого подвида – D. littoralis imeretensis Goncharenko, Mitrofanov, Sokolov, обитающего только на Кавказе, исследованы в 8 популяциях (Россия – Краснодарский край, Грузия).
Реклама

Название и расположение популяций
Drosophila
группы
virilis
*. D.virilis: г. Краснодар, КК, 1981**; г. Северский, КК, 1981; г. Мукачево, Укр, 1986. D. lummei: вблизи г. Тирасполь, Мл, 1984; вблизи г. Геленджик, КК, 1982; вблизи г. Гомель, Бл, 1983-1986; вблизи г. Орша, Бл, 1985; вблизи г. Москва, МО; вблизи г. Оулу, Фн; р. Обь в 100 км южнее г. Новосибирск, НО, 1985-1988. D. montana; вблизи г. Геленджик, КК, 1982; вблизи г. Мукачево, Укр, 1986; Карельское побережье Белого моря, Кр, 1985; р. Камчатка в 200 км от устья, КО, 1988. D. ezoana: р. Катунь, АК, 1988; Уссурийский заповедник, ПК, 1987; заповедник "Кедровая Падь", ПК, 1987; р. Камчатка в 200 км от устья, КО, 1988; вблизи г. Южносахалинск, СО, 1990. D. l. littoralis: вблизи г. Мукачево, Укр, 1986; р. Латорица в 10 км восточнее Мукачево, Укр, 1985; г. Воловец, Укр, 1986; вблизи г. Черновцы, Укр, 1985; Галые болота восточнее г. Гомель, Бл, 1981-1984; ручей в 10-км южнее г. Гомель, Бл, 1984-1987; вблизи г. Речица, Бл, 1985; п. Гаголи, недалеко от устья р. Березина, Бл, 1985; вблизи г. Орша, Бл, 1985; вблизи г. Цесие, Лт, 1989; национальный парк "Гауя", Лт, 1989; вблизи п. Кропотово, МО; вблизи г. Оулу, Фн; Карельское побережье Белого моря, КР, 1985; вблизи г. Талас, Кз, 1986-1987; Таш-Арык в 15 км восточнее г. Талас, Кз, 1987; Киргизский хребет р. Кен-Кол, Кз, 1986; Чаткальский хребет в верховьях р. Чаткал, Кз, 1988; р. Обь в 100 км южнее г. Новосибирск, НО,1887-1989; р. Катунь, АК, 1989. D. l. imeretensis: вблизи г. Геленджик, КК, 1982; п. Лермонтовка побережье Черного моря, КК, 1981; п. Мирный в 45 км севернее п. Лермонтовка, КК, 1981; п. Убинка в 60 км севернее п. Лермонтовка, КК, 1982; р. Безымянка в 30 км севернее п. Лермонтовка; вблизи п. Гумати, Гр; вблизи п. Мцхета, Гр; п. Кеда восточное побережье Черного моря, Гр.
Реклама

Отловленных взрослых самок отсаживали в отдельные пробирки для откладки яиц, а самцов подвергали электрофоретическому исследованию. Через несколько дней после откладки яиц самок также анализировали на изоферментный состав. Для определения генотипа самца, оплодотворившего самку, исследовали от 2 до 10 взрослых потомков от каждой самки. Данные по популяциям Грузии, Московской обл. и Финляндии получены на основании анализа лабораторных линий. В этих случаях считалось, что каждая линия несет только один геном дикого типа.
Взрослые особи палеарктических видов Drosophila группы virilis исследовались методом электрофореза. Каждая особь гомогенизировалась в 25 мкл дистиллированной воды или гелевого буфера. Электрофоретическое фракционирование гомогенизированных экстрактов индивидуальных особей проводилось нами по 11 ферментам в 13-14% крахмальном геле с использованием двух буферных систем: А
) трис-ЭДТА-боратная, рН 8.6; В
) трис-цитрат, рН 6.2. Все параметры электрофоретического фракционирования, а также экстракция и гистохимическое выявление ферментов подробно приведены нами ранее (Гончаренко и др., 1984; Гончаренко, 1987; Goncharenkoetal., 1985). Название ферментов, их кодовый номер, предпочитаемая для анализа буферная система, а также количество используемых локусов приведены в табл.1. Каждая муха была исследована по 15 генам, кодирующим 11 ферментов (табл.1.). Обозначение выявленных электрофоретических вариантов дано по общепринятой номенклатуре Пракаша с соавторами (Prakash et al., 1969), в соответствии с которой наиболее часто встречающийся электроморф по каждому локусу у D. virilis и кодирующий его аллель обозначались символом 1.00, а все другие аллельные варианты, встреченные у проанализированных нами видов Drosophila группы virilis обозначены цифровыми символами в зависимости от их электрофоретической подвижности относительно 1.00. Нулевые аллели обозначены символом 0.
Таблица 1
. Ферменты и количество локусов, использованные для анализа видов-двойников Drosophila группы virilis Палеарктики
| Фермент |
Аббревиатура |
Буферная система |
Кодовый номер |
Количество локусов |
| Алкогольдегидрогеназа |
ADH |
А |
1.1.1.1. |
1 |
| Гексокиназа |
HK |
А |
2.7.1.1. |
2 |
| Изоцитратдегидрогеназа |
IDH |
В |
1.1.1.42. |
1 |
| Кислая фосфатаза |
ACPH |
А |
3.1.3.2. |
1 |
| Малатдегидрогеназа |
MDH |
В |
1.1.1.37. |
2 |
| Малик |
ME |
А |
1.1.1.40 |
1 |
| Октанолдегидрогеназа |
ODH |
А |
1.1.1.1. |
1 |
| Фосфоглюкомутаза |
PGM |
А |
2.7.5.1. |
1 |
| Эстераза |
EST |
А |
3.1.1.1. |
3 |
| Фумараза |
FUM |
А |
4.2.1.2. |
1 |
| α-Глицерофосфатдегидрогеназа |
GPDH |
В |
1.1.1.8. |
1 |
Для оценки генетической близости и дифференциации
среди представителей Drosophila группы virilis использовался коэффициент генетической дистанции Неи, DN
(Nei, 1972), который учитывает различия в аллельных частотах всех проанализированных локусов:
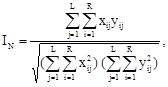
DN
= – lnIN
,
где xij
и yij
— частоты i-того аллеля j-того локуса сравниваемых таксонов.
Если DN
равно 0, то таксоны идентичны. Чем больше значение DN
, тем менее они родственны. Считается, что коэффициент дистанции Неи дает наиболее точные оценки генетической дифференциации, и поэтому он широко используется практически всеми исследователями. Дендрограмма отражающая картину генетических взаимоотношений между исследованными видами Drosophila группы virilis на основании коэффициентов DN
была построена путем невзвешенного парногруппового метода кластерного анализа
(UPGMA)
(Sneath, Sokal, 1973).
Еще одним достоинством коэффициента генетической дистанции Неи (DN
) является то, что он позволяет рассчитывать время дивергенции (t)
таксонов различного ранга исходя из следующего соотношения:
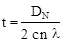
где с – доля аминокислотных замещений, которая может быть определена посредством электрофореза, n – общее количество кодонов, связанных с синтезом полипептида, и λ – средняя скорость аминокислотных замещений (Nei, 1971). Согласно Неи (1971), t = 7.4 ´ 105
DN
. Имеется также другая оценка временной шкалы, предложенная Неи в более поздней работе, где t = 5 ´ 106
DN
(Nei, 1975).
2. Результаты и обсуждение
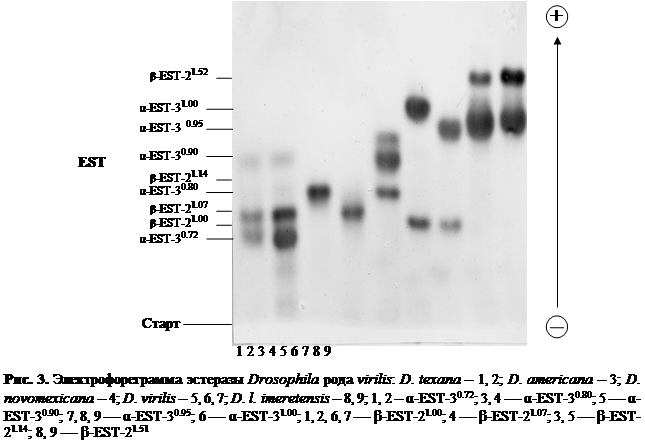
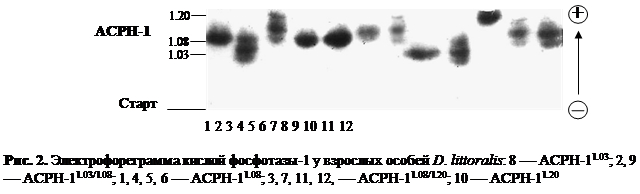
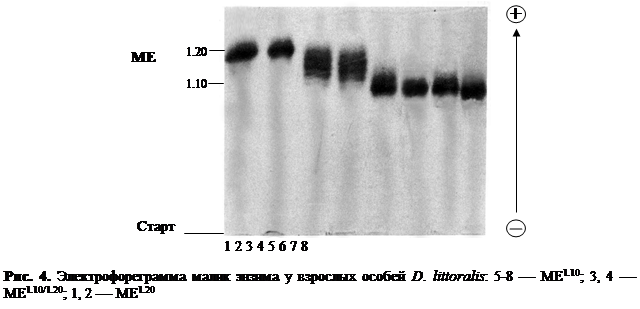
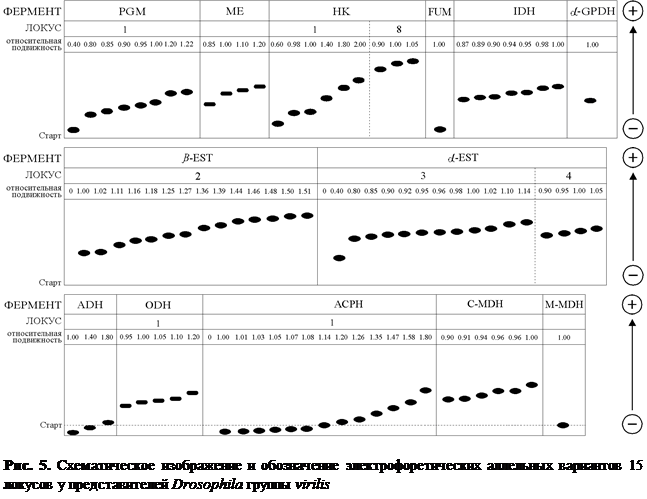
В ходе электрофоретического исследования особей пяти видов Drosophila группы virilis из 47 природных популяций удалось выявить 89 различных электрофоретических вариантов. В результате проведенного нами всестороннего генетического анализа было установлено, что эти 89 электрофоретических вариантов, выявленных по 11 ферментным системам у представителей Drosophila группы virilis, находятся под генетическим контролем 15 локусов. Наглядное изображение электрофоретических спектров диагностических локусов кислой фосфотазы-1, эстеразы и малик энзима у исследованных видовDrosophila группы virilis с выявленными электрофоретическими вариантами приведено на рис. 2, 3 и 4. Все обнаруженные нами 89 аллельных вариантов шести видов-двойников Drosophila группы virilis из природных популяций и их относительная электрофоретическая подвижность схематически изображены на рис. 5.
Аллельные частоты по 15 локусам для всех шести исследованных представителей
Drosophilaгруппы virilisотображены в табл. 2. Следует подчеркнуть, что в популяционных исследованиях использованы только локусы с установленной нами генетической детерминацией. При анализе особей шести видов оказалось, что локусы Fum, α-Gpdh, m-Mdh являются мономорфными поскольку по этим генам у всех видов группы virilis найден только один аллель. Наибольшая изменчивость обнаружена по генам, кодирующим α-эстеразу-3, ß-эстеразу-2, кислую фосфотазу-1. В каждом из этих локусов встретилось более 10 аллелей. Из таблицы 2 следует, что самыми близкими генетическими структурами обладают D. l. littoralis и D. l. imeretensis. Эти аллопатрические подвиды (Гончаренко и др., 1989) существенно различаются по трем генам Me, ß-Est-2, α-Est-3, хотя даже в этом случае качественного уровня разница между подвидами достигла только по локусу Me(табл.2). Качественно различающиеся локусы, по которым можно определять видовую принадлежность особи с ошибкой менее 5%, Айалой, Пауэллом и Левонтиным было предложено называть диагностическими (Ayala, Powell, 1972; Левонтин 1978).
Из табл. 2 видно, что ни один ген не является диагностическим для всех пяти видов. Наиболее информативным оказался локус Me, по которому высоко диагностические различия в аллельных частотах имеются даже у аллопатрических подвидов D. l. littoralis и D. l. imeretensis. Достаточно удобными для диагностики видов-двойников группы virilis являются гены Acph-1, Pgm, ß-Est-2 и α-Est-3. Что касается гена α-Est-4, то этот локус имеется только у D. montana. Однако он позволяет легко отнести каждую исследуемую особь к D. montana или к другим видам двойникам группы virilis.
В табл. 3 ниже диагонали приведены числа высоко диагностических локусов по каждой паре исследованных видов. Как и следовало ожидать, наименьшее количество высоко диагностических локусов (один) обнаружено у подвидов D. l. littoralis и D. l. imeretensis. Виды D. virilis и D. lummei, которые входят в филаду virilis (Throckmorton, 1982), различаются только по трем диагностическим генам Pgm, Acph-1, Idh, (табл. 3). Подвиды D. l. imeretensis, D. l. littoralis и виды D. montana, D. ezoanaвходящие в другую филаду двойниковых видов – филаду montana, отличаются между собой несколько больше. Здесь различия найдены по 5 – 6 высоко диагностическим генам. В целом разница между видами из разных филад достигает семи диагностических генов и превышает внутрифиладные различия. Если экстраполировать результаты, полученные по 15 локусам на весь геном, то можно предположить, что у видов-двойников группы virilis из разных филад более 60% структурных генов несут видоспецифические аллели. В настоящее время это самый большой процент диагностических различий, найденный у двойниковых видов Drosophila (Kojimaetal.,1970; Ayala, Powell, 1972; Andersonetal., 1977; Pinsker, Buruga, 1982).
Таблица 2.
Аллельные частоты по 15 локусам у D. virilis, D. lummei, D. l. littoralis, D. l. imeretensis, D. montana и D. ezoana
| Локус |
Аллели |
D.virilis |
D. lummei |
D. littoralis |
D. imeretensis |
D. montana |
D. ezoana |
|
P
gm |
n1)
|
196 |
99 |
570 |
819 |
32 |
44 |
| 0.40 |
.000 |
.000 |
.018 |
.011 |
.000 |
.000 |
| 0.80 |
.000 |
.929 |
.958 |
.879 |
.000 |
.251 |
| 0.85 |
.000 |
.000 |
.000 |
.001 |
.000 |
.000 |
| 0.95 |
.000 |
.000 |
.000 |
.006 |
.000 |
.000 |
| 1.00 |
.964 |
.071 |
.019 |
.097 |
1.000 |
.704 |
| 1.20 |
.036 |
.000 |
.005 |
.002 |
.000 |
.045 |
| 1.22 |
.000 |
.000 |
.000 |
.004 |
.000 |
.000 |
| Me
|
n |
172 |
98 |
605 |
487 |
38 |
42 |
| 0.85 |
.000 |
.000 |
.000 |
.000 |
.974 |
.000 |
| 1.00 |
1.000 |
1.000 |
.000 |
.000 |
.026 |
1.000 |
| 1.10 |
.000 |
.000 |
.002 |
.990 |
.000 |
.000 |
| 1.20 |
.000 |
.000 |
.998 |
.010 |
.000 |
.000 |
| Hk-1
|
n |
196 |
96 |
539 |
819 |
32 |
44 |
| 0.60 |
.005 |
.000 |
.000 |
.000 |
.000 |
.000 |
| 0.98 |
.000 |
.000 |
.000 |
.000 |
.000 |
.978 |
| 1.00 |
.995 |
1.000 |
.011 |
.001 |
1.000 |
.000 |
| 1.40 |
.000 |
.000 |
.978 |
.991 |
.000 |
.022 |
| 1.80 |
.000 |
.000 |
.007 |
.002 |
.000 |
.000 |
| 2.00 |
.000 |
.000 |
.004 |
.006 |
.000 |
.000 |
| Hk-
8
|
n |
192 |
96 |
539 |
819 |
30 |
44 |
| 0.90 |
.000 |
.000 |
.000 |
.000 |
.266 |
.000 |
| 1.00 |
1.000 |
1.000 |
.998 |
1.000 |
.734 |
1.000 |
| 1.05 |
.000 |
.000 |
.002 |
.000 |
.000 |
.000 |
| ß-Est-2
|
n |
192 |
100 |
609 |
821 |
37 |
60 |
| 0 |
.000 |
.050 |
.000 |
.000 |
.000 |
.000 |
| 1.00 |
.578 |
.000 |
.000 |
.000 |
.000 |
.000 |
| 1.02 |
.000 |
.870 |
.000 |
.000 |
.000 |
.000 |
| 1.11 |
.250 |
.010 |
.000 |
.000 |
.000 |
.000 |
| 1.16 |
.000 |
.070 |
.000 |
.000 |
.000 |
.000 |
| 1.18 |
.172 |
.000 |
.000 |
.000 |
.000 |
.000 |
| 1.25 |
.000 |
.000 |
.000 |
.000 |
.000 |
.350 |
| 1.27 |
.000 |
.000 |
.000 |
.000 |
.054 |
.000 |
| 1.36 |
.000 |
.000 |
.002 |
.000 |
.027 |
.000 |
| 1.39 |
.000 |
.000 |
.002 |
.000 |
.054 |
.567 |
| 1.44 |
.000 |
.000 |
.074 |
.001 |
.135 |
.083 |
| 1.46 |
.000 |
.000 |
.021 |
.000 |
.000 |
.000 |
| 1.48 |
.000 |
.000 |
.575 |
.008 |
.297 |
.000 |
| 1.50 |
.000 |
.000 |
.000 |
.000 |
.352 |
.000 |
| 1.51 |
.000 |
.000 |
.326 |
.991 |
.081 |
.000 |
| α-Est-3
|
n |
192 |
95 |
594 |
782 |
37 |
21 |
| 0 |
.000 |
.042 |
.000 |
.000 |
.000 |
.000 |
| 0.84 |
.453 |
.000 |
.000 |
.000 |
.000 |
.000 |
| 0.85 |
.000 |
.000 |
.000 |
.076 |
.000 |
.239 |
| 0.90 |
.151 |
.905 |
.002 |
.235 |
.000 |
.285 |
| 0.92 |
.000 |
.000 |
.000 |
.000 |
.162 |
.000 |
| 0.95 |
.000 |
.053 |
.178 |
.492 |
.108 |
.476 |
| 0.96 |
.042 |
.000 |
.000 |
.000 |
.000 |
.000 |
| 0.98 |
.182 |
.000 |
.000 |
.000 |
.108 |
.000 |
| 1.00 |
.172 |
.000 |
.052 |
.000 |
.244 |
.000 |
| 1.02 |
.000 |
.000 |
.332 |
.087 |
.081 |
.000 |
| 1.10 |
.000 |
.000 |
.296 |
.110 |
.297 |
.000 |
| 1.14 |
.000 |
.000 |
.140 |
.000 |
.000 |
.000 |
| α-Est-4
|
n |
128 |
95 |
558 |
782 |
34 |
21 |
| 0.90 |
.000 |
.000 |
.000 |
.000 |
.029 |
.000 |
| 0.95 |
.000 |
.000 |
.000 |
.000 |
.147 |
.000 |
| 1.00 |
.000 |
.000 |
.000 |
.000 |
.677 |
.000 |
| 1.05 |
.000 |
.000 |
.000 |
.000 |
.147 |
.000 |
| Acp
h-1
|
n |
172 |
98 |
603 |
785 |
34 |
77 |
| 0 |
.000 |
.000 |
.000 |
.001 |
.000 |
.000 |
| 1.00 |
.994 |
.000 |
.000 |
.000 |
.000 |
.259 |
| 1.01 |
.000 |
.020 |
.000 |
.000 |
.000 |
.000 |
| 1.03 |
.000 |
.000 |
.017 |
.006 |
.000 |
.000 |
| 1.05 |
.006 |
.000 |
.000 |
.000 |
.000 |
.129 |
| 1.07 |
.000 |
.051 |
.000 |
.000 |
.000 |
.000 |
| 1.08 |
.000 |
.643 |
.869 |
.979 |
.000 |
.611 |
| 1.14 |
.000 |
.000 |
.071 |
.010 |
.000 |
.000 |
| 1.20 |
.000 |
.000 |
.053 |
.004 |
.000 |
.000 |
| 1.26 |
.000 |
.276 |
.000 |
.000 |
.000 |
.000 |
| 1.35 |
.000 |
.000 |
.000 |
.000 |
.883 |
.000 |
| 1.45 |
.000 |
.000 |
.000 |
.000 |
.029 |
.000 |
| 1.58 |
.000 |
.000 |
.000 |
.000 |
.088 |
.000 |
| 1.80 |
.000 |
.010 |
.000 |
.000 |
.000 |
.000 |
| A
dh
|
n |
192 |
96 |
535 |
717 |
34 |
22 |
| 1.00 |
1.000 |
1.000 |
.002 |
.001 |
.000 |
.000 |
| 1.40 |
.000 |
.000 |
.998 |
.994 |
1.000 |
1.000 |
| 1.80 |
.000 |
.000 |
.000 |
.005 |
.000 |
.000 |
| L-Gpdh
|
n |
38 |
66 |
34 |
62 |
30 |
26 |
| 1.00 |
1.000 |
1.000 |
1.000 |
1.000 |
1.000 |
1.000 |
| Odh
|
n |
126 |
32 |
133 |
180 |
4 |
0 |
| 0.95 |
.000 |
.031 |
.015 |
.005 |
.000 |
.000 |
| 1.00 |
.976 |
.969 |
.917 |
.941 |
1.000 |
.000 |
| 1.05 |
.024 |
.000 |
.023 |
.016 |
.000 |
.000 |
| 1.10 |
.000 |
.000 |
.038 |
.033 |
.000 |
.000 |
| 1.20 |
.000 |
.000 |
.007 |
.005 |
.000 |
.000 |
| Fum
|
n |
128 |
44 |
525 |
819 |
28 |
4 |
| 1.00 |
1.000 |
1.000 |
1.000 |
1.000 |
1.000 |
1.000 |
| C-Mdh
|
n |
92 |
90 |
188 |
4 |
40 |
26 |
| 0.90 |
.000 |
.044 |
.000 |
.000 |
.000 |
.000 |
| 0.91 |
.000 |
.000 |
.952 |
1.000 |
.000 |
.000 |
| 0.94 |
.000 |
.000 |
.048 |
.000 |
1.000 |
.000 |
| 0.96 |
.000 |
.056 |
.000 |
.000 |
.000 |
.000 |
| 0.98 |
.000 |
.000 |
.000 |
.000 |
.000 |
1.000 |
| 1.00 |
1.000 |
.900 |
.000 |
.000 |
.000 |
.000 |
| M-Mdh
|
n |
92 |
90 |
188 |
4 |
40 |
26 |
| 1.00 |
1.000 |
1.000 |
1.000 |
1.000 |
1.000 |
1.000 |
| Idh
|
n |
4 |
90 |
34 |
0 |
40 |
56 |
| 0.87 |
.000 |
.000 |
.000 |
.000 |
.975 |
.000 |
| 0.89 |
1.000 |
.000 |
.000 |
.000 |
.000 |
.000 |
| 0.90 |
.000 |
.000 |
.000 |
.000 |
.025 |
.000 |
| 0.94 |
.000 |
.000 |
.000 |
.000 |
.000 |
1.000 |
| 0.95 |
.000 |
.011 |
.000 |
.000 |
.000 |
.000 |
| 0.98 |
.000 |
.000 |
.912 |
.000 |
.000 |
.000 |
| Примечание:1)
n – число проанализированных геномов. |
|
|
1.00 |
.000 |
.989 |
.088 |
.000 |
.000 |
.000 |
Таблица 3.
Число диагностических генов для каждой пары шести представителей Drosophila группы virilis Палеарктики
| № вида |
Виды |
№ вида |
| 1 |
2 |
3 |
4 |
5 |
6 |
| 1 |
D. virilis |
4 |
7 |
7 |
7 |
6 |
| 2 |
D. lummei |
3 |
4 |
5 |
8 |
7 |
| 3 |
D. l. littoralis |
6 |
3 |
3 |
7 |
6 |
| 4 |
D. l. imeretensis |
7 |
5 |
1 |
7 |
6 |
| 5 |
D. montana |
6 |
5 |
6 |
6 |
6 |
| 6 |
D. ezoana |
5 |
5 |
4 |
6 |
5 |
Примечание. Ниже диагонали – гены, по каждому из которых ошибка в определении вида не превышала 1 %; выше диагонали – все гены, по которым имеется возможность установить видовую принадлежность особей (включая высокодиагностические)
Некоторые гены еще не достигли уровня диагностических, хотя и по ним с высокой вероятностью можно определить, к какому виду относится та или иная особь группы virilis (табл. 3). Количество таких генов вместе с диагностическими по каждой паре видов приведены в табл. 3 выше диагонали.
Для точной оценки уровня генетической дифференциации видов-двойниковDrosophila группы virilis мы использовали коэффициент генетической дистанции Неи (Nei, 1972), который учитывает различия по всем исследованным локусам, а не только по диагностическим и сильноразличающимся. Значения коэффициентов генетической дистанции полученные нами для 5 палеарктических видов группы virilis приведены в табл. 4. Как и следовало ожидать исходя из сравнения аллельных частот, наименьшие значения коэффициентов дистанции Неи выявлены между аллопатрическими подвидами D. l. littoralis и D. l. imeretensis средняя величина DN
для которых равна 0.141 (табл. 4). Дистанция Неи между D. virilis и D. lummei входящих в одну филаду составила 0,334. Наибольшие значения коэффициентов дистанции Неи у видов из разных филад – D. virilis и D. l. littoralisсредняя величина DN
равна 0.909. Следовательно, у двух видов двойников произошло более 90 аллельных замен на 100 локусов.
Таблица 4.
Значения коэффициентов генетической дистанции Неи для шести представителей Drosophila группы virilis Палеарктики
| Виды |
D. lummei |
D. l. littoralis |
D. l. imeretensis |
D. montana |
D. ezoana |
| D. virilis |
0.334 |
0.909 |
0.843 |
0.608 |
0.627 |
| D. lummei |
0.645 |
0.580 |
0.764 |
0.636 |
| D. l. littoralis |
0.141 |
0.687 |
0.617 |
| D. l. imeretensis |
0.668 |
0.511 |
| D. montana |
0.684 |
Для наглядного изображения полученных результатов на базе данных табл. 4 с использованием невзвешенного парно-группового метода кластерного анализа (UPGMA) построена дендрограмма, иллюстрирующая степень генетической дифференциации у 6 исследованных видов-двойниковDrosophila группы virilis (рис. 6). Из дендрограммы хорошо видно, что D. ezoana, D. montana, и D. littoralis, которые входят в филаду montanaзамыкаются в собственный кластер, отделённый от представителей филады virilis. Дистанция Ней как между видами филады montana, так и видами из разных филад, превышает значение 0,55 (табл. 4). Иными словами разница в генотипах между этими видами превысила 50%. Вместе со столь внушительной разницей появилась и практически полная репродуктивная изоляция, так как среди немногочисленных межвидовых гибридов фертильны только некоторые самки. Причём, особи D. littoralis, D. montana, и D. ezoana в природных популяциях часто встречаются вместе, однако нам ни разу не удалось обнаружить межвидовых гибридов. Это говорит о том, что наряду с постзиготической репродуктивной изоляцией между этими видами уже сформировались и действуют презиготические изолирующие механизмы.

Сходная ситуация имеет место и при сравнении видов и разных филад. В этом случае разница в геномах превышает 70% (табл. 4) и среди межвидовых гибридов фертильны только отдельные самки. Следовательно, мы наблюдаем 5-ый уровень генетической дифференциации, когда после завершения процесса видообразования генетические различия и репродуктивный барьер продолжают углубляться, но морфологические отличия ещё крайне не значительны и визуально не обнаруживаются.
Установленные нами на основании генетической близости филогенетические взаимоотношения 5 видов-двойников Drosophila группы virilis (рис.6) в целом согласуются с результатами цитологического анализа (Throckmorton, 1982), по которым виды группы virilis распадаются на две филады и возникают путём классической дивергентной эволюции. В тоже время, исходя из наших данных (рис. 6), распад общей предковой формы произошёл около миллиона лет назад сразу на 3 ветви: ветвь virilis, которая затем претерпела собственную эволюцию на территории Евразии; ветвь littoralis, позднее распавшуюся на D. littoralis и D. ezoana и широко распространившиеся по всей Полеарктике; и, наконец, ветвь montana, которая дала D. montana, обитающую как в Еразии, так и в Северной Америке.
В ходе исследования природных популяций Drosophila группы virilis выявлено 5 уровней генетической дифференциации табл.5. На первом уровне находятся географически связанные популяции, поток генов между которыми нивелирует все генетические различия; на втором уровне находятся географически изолированные популяции одного вида; на третьем – кавказский и северный подвиды D. littoralis, у которых появились первые признаки репродуктивного барьера; на четвертом – двойниковые виды D. virilis и D. lummeiиз одной филады, меду которыми ещё возможны фертильные гибриды обоего пола; и, наконец, на пятом уровне находятся виды-двойники из разных филад, репродуктивный барьер между которыми сформировался почти полностью. Показано, что эволюционный процесс у евроазиатских видов группы virilis начался более миллиона лет назад с распада сразу на 3 ветви: virilis, littoralis и montana, которые затем эволюционировали самостоятельно.
Таблица 5.
Генетическая дифференциация между популяциями Drosophila группы virilis, находящихся на разных уровнях эволюционной дивергенции. Уровни 3 и 4 отвечают значениям коэффициентов генетической дистанции Неи для шести представителей Палеарктики
| Уровни сравнения |
Значения дистанции Неи |
| 1. |
Географически связанные популяции |
0.022 |
| 2. |
Географически разделенные популяции |
0.065 |
| 3. |
Подвиды |
0.141 |
| 4. |
Молодые виды-двойники одной филады |
0.334 |
| 5. |
Виды-двойники разных филад |
0.682 |
3. Электрофоретический ключ для типировки взрослых особей
Изоферментные маркеры постепенно начали применяться и для выяснения ряда таксономических вопросов (Ayala, Powell, 1972; Pinsker, Buruga, 1982; Geiger, Scholl, 1985). Ранее предпринимались попытки построения электрофоретических ключей, которые позволяют быстро и надежно устанавливать видовую принадлежность отдельных особей (Miles, 1979; Berlocher, 1980; Hovard, Furth, 1986; Menken, Ulenberg, 1986).
На основании представленного выше электрофоретического анализа изоферментов нами составлен точный и относительно недорогой определительный ключ, позволяющий легко и быстро идентифицировать особей пяти палеарктических видов-двойников Drosophila группы virilis.
Из таблицы аллельных частот (табл. 2) хорошо видно, что локус Adh делит виды на две группы. К первой относятся виды, входящие в филаду virilis – D. lummei и D. virilis, у которых встречается только аллель 1.00. Вторую группу составляют три вида филады montana, практически все особи которых несут аллель 1.40.
Ген, кодирующий фосфоглюкомутазу, является наиболее изменчивым из трех локусов, тем не менее он позволяет нам легко отличить особей D. virilis, гомозиготных или гетерозиготных только по аллелям 1.00 и 1.20, от представителей D. lummei, гомозиготных по аллелю 0.80 или гетерозиготных по аллелям 0.80/1.00.
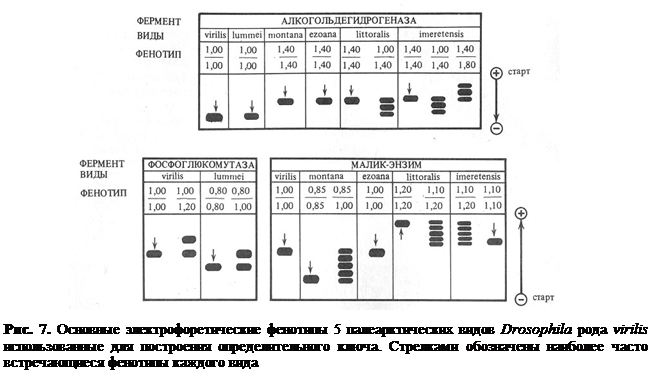
По ME можно легко протипировать особей D. montana, имеющих "медленные" электрофоретические фенотипы, а также особей D. l. littoralis и D. l. imeretensis, которые отличаются своими аллелями не только от остальных видов группы, но и между собой.
На рис. 7 дана схема, показывающая основные электрофоретические фенотипы исследованных нами видов-двойников группы virilis. Представлены фенотипы, которые встречаются в природных популяциях не реже чем один на тысячу особей. На основании данных содержащихся в табл. 2 и рис. 7 нами составлен электрофоретический ключ, позволяющий быстро и надежно определять особей пяти видов-двойников. Необходимым условием использования данного ключа является наличие особей вида-стандарта. В нашем ключе стандартом является D. virilis, которая легко культивируется в лабораторных условиях.
На начальном этапе использования ключа анализируется фермент ADH (табл. 6). На одном электрофоретическом блоке вместе с образцами мух неизвестных видов ставится стандартный образец. Если электрофоретический фенотип определяемой мухи совпадает со стандартом, то данная особь относится либо к D. virilis, либо к D. lummei. Следует отметить, что стандартный фенотип по ADH может встретиться также и у D. littoialis (см. табл. 2), однако вероятность этого события не превышает 4 х 10-6
. Иными словами, при определении видовой принадлежности одного миллиона особей D. littoralisна данном этапе ошибочно будут определены только 4 мухи (т.е. ошибка менее 0,0004%). Для того чтобы точно установить видовую принадлежность особей, стандартных по ADH, следует проанализировать их фенотип по фосфоглюкомутазе. Если электрофоретический фенотип по PGM у таких особей совпадает или выше стандарта, то это D. virilis, а если ниже стандарта, то – D. lummei. При этом ошибка определения на данном этапе не превышает 0,05%.Особи, оказавшиеся нестандартными по ADH, должны быть исследованы по ферменту малик-энзиму. Мухи, стандартные по ME, относятся к D. ezoana, показавшие более медленный фенотип – к D. montana, а более быстрый – к D. littoralis. Причем особи, относящиеся к подвиду D. l. littoralis, отличаются от D. l. imeretensis более быстрым электрофоретическим фенотипом по малик-энзиму.
Таблица 6
. Электрофоретический ключ для палеарктических видов группы virilis
№ этапа определения
|
Фермент |
Подвижность относительно стандарта |
Диагноз |
Максимальная ошибка диагноза. %
|
1
|
ADH |
стандарт
|
2
|
0,0004
|
| ADH |
быстрее стандарта
|
3
|
0,0000
|
| 2 |
PGM |
стандарт, или быстрее стандарта |
D. virilis |
0,0441 |
| PGM |
медленнее стандарта
|
D. lummei
|
0,0445
|
3
|
ME |
стандарт
|
D. ezoana
|
0,1225
|
| ME |
не стандарт
|
4
|
0,0000
|
| 4 |
ME |
медленнее стандарта |
D. montana |
0,1225 |
| ME |
быстрее стандарта |
5 |
0,0000 |
5
|
ME |
самый быстрый электрофоретический фенотип в группе virilis |
D. l.littoralis |
0,4100 |
| ME |
быстрее стандарта, но медленнее D. l. littoralis |
D. l.imeretensis |
0,4096
|
Заключение
Таким образом, проведен анализ генетико-эволюционных и таксономических взаимоотношений у видов-двойников Drosophila группы virilis, обитающих в природных популяциях Палеарктики, с использованием генов, кодирующих различные изоферменты, который показал, что, в ходе электрофоретического исследования особей шести представителей Drosophila группы virilis из 47 природных популяций удалось выявить по 11 ферментным системам 89 различных электрофоретических вариантов, находящихся под генетическим контролем 15 локусов. Проанализированы диагностические гены. Наиболее информативными оказались локусы Me, Acph-1, Pgm, ß-Est-2 и α-Est-3. Для точной оценки уровня генетической дифференциации видов-двойниковDrosophila группы virilis использован коэффициент генетической дистанции Неи. Составлен точный и относительно недорогой электрофоретический ключ, позволяющий легко и быстро идентифицировать особей пяти палеарктических видов-двойников Drosophila группы virilis.
Литература
1. Гончаренко Г.Г. Аллозимная диагностика видов-двойников Drosophila группыvirilis // ДАН СССР. – 1987. – Т. 295. N 4. – С. 976-980.
2. Гончаренко Г.Г., Емельянов И.М. Электрофоретический ключ для типировки взрослых особей двойниковых видов Drosophila группы virilis, обитающих в Палеарктике // Докл. АН СССР. – 1990. – Т. 313. N 2. – С. 448-452.
3. Гончаренко Г.Г., Митрофанов В.Г., Катохин А.Н. Изучение биохимического полиморфизма у Drosophila imeretensis в природных популяциях Краснодарского края // Генетика. – 1984. – Т. ХХ. № 4. – С. 620-627.
4. Гончаренко Г.Г., Митрофанов В.Г., Корочкин Л.И., Савицкий Б.П. Первый этап видообразования у двух подвидов Drosophila группыvirilis // ДАН СССР. – 1989. – Т. 304. № 2. – С. 448-451.
5. Левонтин Р. Генетические основы эволюции: Пер. с англ. М.: Мир, 1978. – 352 с.
6. Майр Э. Зоологический вид и эволюция. Пер. с англ. М.: Мир, 1968. – 462 с.
7. Anderson W.W., Ayala F.J., Michod R.E. Chromosomal and allozymic diagnosis of three species of Drosophila.. // J. Hered., 1977. – vol. 68, p. 71-74.
8. Ayala F.J., Powell J.R. Allozymes as diagnostic characters of sibling species of Drosophila// Proc. Nat. Acad. Sci. USA. 1972. V. 69. – P. 1094–1096.
9. Berlocher S.M. An electrophoretic key for distinguishing species of the genus Rhagoletic (Diptera; Tephritidae) as larvae, pupae or adults // Ann. Entomol. Sos. Amer. 1980. – Vol.73. – P.131-137.
10. Geiger H., Scholl A. Systenatic and evolution of Holarctic Pierinae (Lepidoptera). An enzyme electrophoretic approach // Experientia. – 1985, vol. 41, p. 24-29.
11. Goncharenko G.G., Emelianov I.M. An electrophoretic key to adult members of the sibling species belonging to the Drosophilavirilis group (Diptera, Drosophilidae) inhabiting Soviet Union and adjacent countries // Z. zool. Syst. Evolut.-forsch. – 1992. – V. 30. – P. 281-286.
12. Goncharenko G.G., Mitrofanov V.G., Korochkin L.I. Localization of genes coding for biochemical traits on the second chromosome of Drosophila imeretensis Sokolov (D. littoralis Meigen)// Biochemical Genetics. – 1985. – V. 23. N 3-4. – P. 329-335.
13. Hovard D.J., Furth D.G. Review of the Allonemobius fasceatus (Orthoptera: Grillidae) comlex with the description of two new species separated by electrophoresis, songs and morphometrics. Ann. Entomol. Soc. Am. 79, 1986. – p. 472-481.
14. Kojima K., Gillespie J., Tobari Y.N. A prophile of Drosophila species enzymes assayed by electrophoresis. I. Nurmber of alleles, heterozygosites and lincage disequilibrium in glucose-metabolizing systems and some other enzymes. – Biochem. Genet. – 1970. – Vol.4. – N4. – P.627.
15. Korochkin L.I. Genetic control and development expression of esterase isozymes in Drosophila of the virilis group. In C. Markert [ed.], 3.ISOZYMES: Developmental biology. Academic Press Inc., London. – 1975. – P. 99-117.
16. Lakovaara S., Saura A., Lankinew P., Pohjola L., Lokki P. The use of isoenzymes in tracing evolution and in classifying Drosophilidae // Zool. Scr., 1976. V.5. – P.173-179.
17. Menken S., Ulenberg S. Allozymatic diagnosis of four economically important Liriomyza species (Diptera, Agromyzidae). – Ann. appl. Biol., 1986, vol. 109. – p. 41-47.
18. Miles S. A biochemical key to adult members of the Anopheles gambiae group of species (Diptera: Culicidae). J. Med. Entomol. 15, 1979. – p. 297-299.
19. Nei M. Interspecific gene differences and evolutionary time estimated from electrophoretic data on protein identity // Am. Nat. 1971. V. 105. – P. 385–398.
20. Nei M. Genetic distance between populations // Am. Nat. 1972. V. 106. – P. 283–292.
21. Nei M. Molecular Population Genetics and Evolution // Amsterdam: Holland Press, 1975. – 278 p.
22. Patterson S.T., Stone W.S. Evolution in the genus Drosophila N. Y.: McMillan. – 1952 – P.610.
23. Pinsker W., Buruga J. Comparative study of allozyme variation in six species of the Drosophila obscura group. – 1982, vol.20, p.53-63.
24. Prakash S., Lewontin R. C., Hubby J. L. A molecular approach to the study of genic heterozygosity in natural populations. IV. Patterns of genic variation in central, marginal and isolated populations of Drosophila pseudoobscura // Genetics. 1969. V. 61. – P. 841–858.
25. Sneath P. H. A., Sokal R. R. Numerical Taxonomy: the Principles and Practice of Numerical Classification. San Francisco: W. H. Freeman, 1973. – 573 p.
26. Spicer G. S., Bell C.D. Molecular phylogeny of the Drosophilavirilis species group(Diptera: Drosophilidae) inferred from mitochondrial 12S and 16S ribosomal RNA gene // GenesAnn. Entomol. Soc. Am. 95(2). – 2002. – P. 156-161.
27. Throckmorton L. H. The virilis species group In M. Ashburner and E. Novistky [eds,], The genetics and biology of Drosophila, vol. 3B. Academic, London. – 1982. – P. 227-297.
|
