ЗМІСТ
ВСТУП
1. ОСНОВНІ ТЕОРЕТИЧНІ ВІДОМОСТІ ПРО ЕРГОСТЕРИН
1.1 Ергостерин
1.2 Продуценти ергостерину
1.3 Біосинтез ергостерину
1.4 Умови утворення ергостерину дріжджами
2. ТЕХНОЛОГІЯ БІОСИНТЕЗУ ЕРГОСТЕРИНУ
2.1 Технологічні аспекти отримання ергостерину
2.2 Накопичення біомаси дріжджів
2.3 Виробництво концентрату вітамінів групи В
2.4 Виробництво концентрату провітаміна D2 та технологія його трансформації у вітамін D2
3. РОЗРАХУНОК ФЕРМЕНТЕРА МАРКИ Б-50
3.1 Розрахунок габаритних розмірів ферментера
3.2 Розрахунок товщини стінки ферментера
ВИСНОВКИ
СПИСОК ВИКОРИСТАНОЇ ЛІТЕРАТУРИ
Вступ
У 1912 р. для позначення додаткових харчових факторів, що у малих кількостях ефективні для лікування ряду захворювань, К. Функ ввів термін вітамін. Тоді виділяли лише 2 вітаміни — А (жиророзчинний), В (водорозчинний), сьогодні їх кількість сягає десятків.
Вітаміни - це біологічно активні низькомолекулярні природні, органічні сполуки, різної хімічної структури, учасники обміну речовин і беруть участь в перетворенні енергії, переважно як компоненти ферментів.
Існування і значення вітамінів відкрив російський лікар М. Лунін у кінці ХІХ ст.. Польський хімік К. Функ назвав біологічно активну речовину вітаміном, бо вона містила у своїй молекулі аміногрупу. Вітаміни мають надзвичайно велике значення для нормального обміну речовин і життєдіяльності, перебуваючи у дуже малих кількостях в продуктах харчування. За нормального раціону і зорового способу життя потреба у вітамінах задовольняється природним шляхом.
За нестачі вітамінів групи D, у дітей переважно перших трьох років життя з'являються ознаки рахіту. В дорослих (особливо у вагітних жінок), які мало перебувають на сонці, не вживають достатньо повноцінної їжі, кісткова тканина втрачає кальцій і кістки розм'якшуються. В цих випадках таке явище називають остеопорозом. Недостатність вітамінів групи D може розвинутись і у дітей старших трьох років, особливо в періоди інтенсивного росту, якщо їх білкове харчування є недостатнім, а також має місце різка зміна кліматичних умов. Крім того, до розвитку D-вітамінної недостатності ведуть хронічна ниркова недостатність, хвороби печінки, тривалий прийом протисудомних препаратів, синдром мальабсорбції (порушеного всмоктування в кишечнику) різного генезу.
Реклама

Тому важливе значення має пошук нових безпечних джерел надходження вітамінів у організм. У випадку із вітаміном D2
, використання препаратів мікробіологічного синтезу має значні переваги перед синтетичними аналогами, і з точки зору сприйнятливості організмом, і з точки зору економії енергоресурсів, затрачених на процес синтезу. Використання провітаміну D2
(ергостерину) також є більш доцільним, аніж використання готового препарату D2
, оскільки організм людини здатний самостійно перетворювати провітамін у вітамінну форму, за умови достатньої кількості субстрату та сонячного світла.
На сьогоднішній день перспективним є пошук нових, більш ефективних продуцентів, а також нових технологічних схем виділення та очистки кінцевого продукту.
1. Основні теоретичні відомості про ергостерин
1.1 Ергостерин
Природні джерела вітаміну D досить обмежені. Відомо, що вітамін D зустрічається лише в деяких тваринних та рибних продуктах (риб’ячий жир, печінка риб, вершкове масло, молоко, ікра), а також в яєчному жовтку. Рослини, як правило, не містять вітамінів групи D, проте серед стеринів рослинних жирів виявлено провітамін D – ергостерин.
Багаті на вітамін D лише печінкові жири риб. Деякі з них, наприклад печінковий жир тунця, містить в 1 г від 40 до 60 од. вітаміну D2
.
В молоці, вершковому маслі і яйцях вміст вітаміну D2
обмежений. Наприклад, в 100 г молока міститься лише 1 од. D2
, у вершковому маслі – 40 – 320 од.
Незначний вміст вітаміну D2
в природніх продуктах змушує шукати багаті природні джерела провітаміну D2
– ергостерину.
В 30-х роках ХХ ст. рядом дослідників було виявлено, що хлібопекарські дріжджі являються багатим джерелом ергостерину. Пізніше було встановлено, що деякі мікроорганізми можуть слугувати джерелом вітаміну D[2].
Ергостерин – ергоста-5,7,22-трієн-3β-ол – вихідний продукт виробництва жиророзчинного вітаміну D2
і кормових препаратів, збагачених вітаміном D2
. В групу вітамінів Dоб’єднують споріднені сполуки, найважливішими з яких є вітаміни D2
і D3
, що мають антирахітичну дію. Вітамін D2
(ергокальциферол) утворюється під час опромінення ультрафіолетовим випромінюванням ергостерину, вітамін D3
(холекальциферол) утворюється із 7-дегідрохолестерину. В організмі людини і тварин ці сполуки регулюють засвоєння кальцію та фосфору із їжі і відкладання їх в кістковій тканині [1].
Реклама

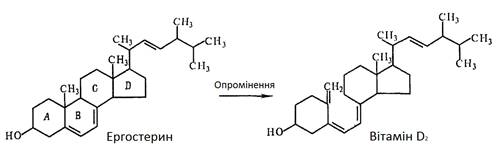
Для прикладної медицини найбільше значення мають вітаміни D2
та D3
. Нестача вітаміну D2
погіршує абсорбцію кальцію в кишківнику, у зв’язку з чим його вміст в крові також зменшується. Це викликає підвищену активність паращитовидних залоз, завдяки чому рівень кальцію в крові утримується на невисокому рівні за рахунок мобілізації його із запасів організму. Одночасно спостерігається підвищене виділення нирками фосфору, оскільки гіперпродукція паратиреоїдного гормону знижує реабсорбція фосфору в ниркових каналах. Таким чином вітамін D2
відграє важливу роль в забезпеченні організму кальцієм, перш за все, за рахунок регуляції його засвоєння з їжі. Загальновідомо, що нестача вітаміну D2
в організмі відбувається порушення мінералізації в процесі кісткоутворення, що призводить до серйозних змін в кістковому скелеті.
В основі структури ергостерину і вітаміну Dлежать чотири вуглецевих цикла (A, B, C, D). В молекулі вітаміну Dкільце В розімкнуте. Вуглеводнева структура ергостерину та вітаміну Dвизначають їхні ліпофільні властивості [1].
1.2 Продуценти ергостерину
Джерелом ергостерину є фітопланктон, бурі і зелені водорості, але особливо багаті на ергостерин дріжджі та плісняві гриби, які слугують сировиною для його промислового виробництва. Ергостерин – основний стерин дріжджів, на який припадає 60 – 90 % від інших стеринів: вміст ергостерину складає 0,2 – 0,5 %, але в деяких випадках досягає 10 % від сухої біомаси.
Культурні раси дріжджів завжди містять більше стеринів, ніж дикі. Найбільшу кількість стеринів містять пекарські та пивні дріжджі. У відношенні ергостеролсинтезуючої здатності (% ергостеролу в абсолютно сухих дріжджах) дріжджі під час поверхневого культивування розташовуються в наступному порядку: Saccharomyces
carlsbergensis
(0,49 – 4,3), S
.
ellipsoideus
(1,2 – 1,5), Rhodotorula
glutinis
(0,7 – 0,9), Candida
utilis
(0,4 – 0,6), C
.
tropicalis
(0,2 – 0,3). В міцелії грибів Aspergillus
і Penicillium
вміст стеринів може досягати 1,2 – 1,4 % (P
.
westlingii
близько 2,2 %) в розрахунку на сухий міцелій.
Бактерії, як правило, синтезують дуже малі кількості стеринів. Зазвичай, вміст стеринів в їхніх клітинах складає 0,001 – 0,1 мг/г сухої біомаси. Стерини виявлено в Lactobacillus arabinosus
, L. pentosus
, Escherichia coli
, Azotobacter chroococcum
, Micromonospora sp.
, Streptomyces griseus
, Sphaerotillis natans
, Rhodospirillum rubrum
. Але відомо два представника бактерій: Halobacterium cutirubrum
і Methylococcus capsulatus
, що синтезують високі кількості сквалену (1,0 і 5,5 мг відповідно на грам сухих клітин). Сквален і його чотири гідроформи виділені із метанутворюючої бактерії Methanobacillus
kuzneceovii
[1].
1.3 Біосинтез ергостерину
Стерини, каротиноїди, сполуки груп Q-коферментів належать до терпенів і мають спільний шлях біосинтезу за «ізопреновим правилом». У відповідності до цього правила, каротиноїди (політерпени), стерини (тритерпени), а також убіхінони і гіберелінова кислота синтезуються із ізопренових одиниць в результаті проходження чотирьох стадій: 1) утворення мевалонату із ацетил-КоА або лейцину; 2) дегідратація і декарбоксилювання мевалонілпірофосфату з утворенням «активного ізопрену» - ізопентенілпірофосфату і конденсація ізопренових ланок з утворенням ациклічних терпенів різної довжини; 3) циклювання ациклічних структур; 4) подальша модифікація циклічної структури.
Інтермедіатами синтезу стеринів є ацетат, мевалонова кислота, сквален, ланостерин. Сквален – загальний попередник стеринів рослинного і тваринного походження, накопичується в дріжджах і під час аерування перетворюється в стерин.
Стартовою сполукою в біосинтезі стеринів є ацетат. Дві молекули ацетил-КоА конденсуються із утворенням ацетоацетил-КоА, який в свою чергу конденсується з ще одною молекулою ацетил-КоА, утворюючи β-гідрокси-β-метилглутарил-КоА. Під час відновлення даної сполуки утворюється мевалонова кислота (МВК), котра за наявності АТФ фосфорилюється із утворенням пірофосфату МВК. За наявності АТФ шляхом декарбоксилювання і дегідрування пірофосфат МВК перетворюється в 5-вуглеводневу ізопренову одиницю – ізопентенілпірофосфат. Ізопентенілпірофосфат (ІПФ) ізомерується до стадії деметилалілпірофосфату (ДМАПФ). Потім відбувається конденсація ІПФ і ДМАПФ з утворенням геранілпірофосфату. Ці сполуки, що містять 10 атомів вуглецю, конденсуються з ІПФ з утворенням фарнезілпірофосфату
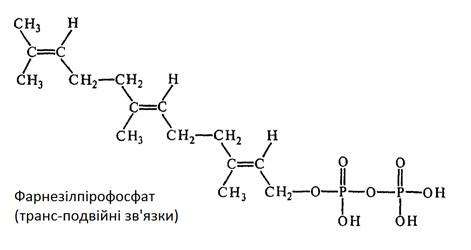
Фарнезілпірофосфат далі димеризується із утворенням сквалену С15
+ С15
= С30
:
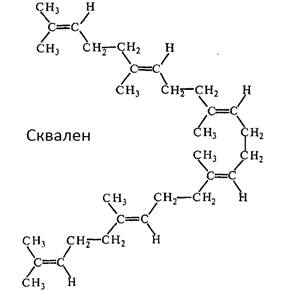
Під час циклювання і відщіплення протону утворюється ланостерин, попередник холестерину і ергостерину:
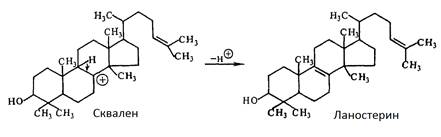
Перетворення ланостерину в ергостерин відбувається в результаті наступних стадій:
1) деметилювання ланостерину;
2) трансалкілування з утворенням 24(28)-метиленової групи і одночасним відновленням С-24(25) подвійного зв’язку;
3) десатурація бічного ланцюга з утворенням С-22(23) подвійного зв’язку;
4) ізомеризація ∆8
∆7
;
5) дегідрування з утворенням ∆5
;
6) відновлення 24(28) метиленової групи до метильної [1].
1.4 Умови утворення ергостерину дріжджами
Найбільш високу кількість стеринів синтезують штами Saccharomyces
carlsbergensis
. Їх біомаса може містити більш ніж 10 % ергостерину.
Важливою умовою синтезу ергостерину дріжджами є достатнє аерування. В анаеробних умовах в клітинах дріжджів накопичується попередник ергостерину – сквален. Доведено, що кисень індукує синтез стеринів, надаючи активуючу дію на епоксидазу сквалену – перший фермент біосинтетичного шляху. Індукція синтезу ергостерину починається за 0,03 %-го вмісту кисню в газовій фазі і досягає максимуму за 2 %-ї концентрації.
Для біосинтезу стеринів є важливим, щоб середовище містило великий надлишок вуглеводів і мало азоту. Дріжджі багаті на білок, як правило, містять мало стеринів. Ці дані, головним чином, стосуються пекарських дріжджів. У випадку дріжджів роду Candida
високе співвідношення C/N в середовищі призводить до накопичення ліпідів, а не ергостерину.
Н
-алкани, якщо вони можуть споживатись дріжджами, являються найкращим джерелом вуглецю для синтезу ергостерину, аніж вуглеводи. Можливо, це пов’язано з утворенням із алканів ацетату (попередника ергостерину) в результаті β-окиснення алканів. Для пекарських дріжджів ацетат також являється гарним джерелом вуглецю в біосинтезі стеринів.
Стимулюючу дію на утворення стеринів дріжджами спричиняють інгібітори гліколізу та роз’єднувачі окисного фосфорилювання і дихання, а також забезпеченість дріжджів вітамінами і, перш за все, пантотеновою кислотою, яка у складі КоА бере участь у побудові молекули ергостерину.
Під час дії на дріжджі рентгенівського випромінювання вміст ергостерину збільшується в 2 – 3 рази, що пояснюється пригніченням процесу амінування, який супроводжується підвищеним синтезом ліпідів. Подібно до іонізуючого випромінювання діють радіоміметричні речовини, порушуючи метаболізм клітини і стимулюючи ліпідний обмін. Наприклад, під час комбінованої дії на клітини дріжджів радіоміметричної речовини (ембіхіну) і рентгенівського випромінювання вихід стеринів Sacch
.
cerevisiae
збільшується на 109 % по відношенню до контролю.
Полієнові антибіотики ністатин і філіпін, взаємодіючи з мембраною дріжджів, підвищує рівень стеринів на 50 – 60 % по відношенню до контролю.
Синтез стеринів не пов’язаний із ростом дріжджів. Вміст стеринів підвищується по мірі старіння культури і стериноутворення продовжуються після зупинки росту дріжджів.
Роль стеринів в метаболізмі продуцентів не зовсім зрозуміла. Спостерігається зв’язок між дихальною активністю дріжджів і утворенням ергостерину. В анаеробних умовах дріжджі містять багато сквалену і мало ергостерину. Роль ергостерину, як структурного компоненту мембран, пов’язують із проникністю клітинних мембран дріжджів.
Найбільш збагачена стеринами у дріжджів фракція мітохондрій. Вірогідно, що мітохондрії беруть участь у біосинтезі стеринів, а стерини, в свою чергу, приймають участь в утворенні мітохондріальних структур і впливають на їхню функціональну активність.
Стерини містять в молекулі ОН-групу і в клітині значною мірою присутні як ефіри жирних кислот. Тому існує твердження, що утворення стеринів – це реакція детоксикації, що захищає організм від надлишкової продукції жирних кислот. Стерини використовуються для транспорту ненасичених кислот і, можливо, інших сполук в клітині. Мікоплазми включають стерини клітинні мембрани. Дріжджі Sacch
.
cerevisiae
в умовах низького вмісту розчиненого кисню в середовищі поглинають екзогенні стерини, при цьому частина їх піддається трансформуванню із утворенням комерційно вигідних стероїдів. Наприклад, десмостерин дріжджі перетворюють в 24 β-метилхолестерин, а 24,25-дигідроланостерин – в 7-дегідрохолестерин, який є інтермедіатом під час синтезу вітаміну D3
[1].
2. Технологія біосинтезу ергостерину
2.1 Технологічні аспекти отримання ергостерину
В промисловості ергостерин отримують, використовуючи дріжджі Sacch
.
cerevisiae
, Sacch
.
carlsbergensis
, а також міцеліальні гриби. Засів проводять великою кількістю інокуляту. Культивування проводять за високої температури і сильного аерування середовища, що містить великий надлишок джерел вуглецю по відношенню до джерела азоту.
Максимум поглинання ергостерину знаходиться за довжини хвилі світла 280 нм. Саме це випромінювання збуджує окремі зв’язки кілець А і В в молекулі ергостерину і викликає його перетворення у вітамін D2
. Опромінення проводять ультрафіолетовими лампами із довжиною хвилі 280 – 300 нм. Опромінюють або сухі дріжджі, або тонкий шар їхньої 5 %-ї суспензії. Під впливом більш коротко- або довгохвильових випромінювань збільшується вихід інших сполук стеринової природи.
На вихід вітаміну D2
(і утворення інших сполук) впливають тривалість опромінення, температура, наявність домішок. Тому опромінення ергостерину, що використовується в якості харчових добавок, проводять з більшою обережністю.
Провітамін D2
отримують із маси, що залишається після фільтрування культуральної рідини. Біомасу промивають, висушують, розмелюють і двічі оброблюють за 78 °С трикратним об’ємом спирту.
Спиртові екстракти згущують до 70 %-го вмісту сухих речовин. Таким чином отримують ліпідний концентрат. Його омилюють розчином гідроксиду натрію, стерини залишаються в неомиленій фракції. Кристали ергостерину випадають із розчину за 0 °С. Очищення кристалів проводять шляхом перекристалізації, послідовним промиванням 69 %-м етиловим спиртом, сумішшю етилового спирту і бензолу (4:1) і повторним перекристалізуванням. Отримані кристали ергостерину висушують, розчиняють в ефірі і опромінюють, Після цього ефір випаровують, а розчин вітаміну концентрують і кристалізують.
Для отримання олійного концентрату розчин вітаміну після фільтрування розводять олією до стандартного рівня.
Джерелом отримання ергостерину також може слугувати міцелій грибів, що залишається як відхід антибіотичної промисловості і виробництва лимонної кислоти [1].
Крім провітаміну D (ергостерину) в дріжджах містяться вітаміни групи В: В3
, В6
, Н, Вс, холін, інозит, р-амінобензойна кислота. За виключенням ергостерину, що розчинний в жирах, всі інші вітаміни, що містяться в дріжджах розчиняються у воді. В дріжджах також містяться фосфоліпіди, ферменти і нуклеїнові кислоти. Відповідно, раціональна технологічна схема переробки дріжджів повинна базуватися на процесах, що забезпечують повну екстракція вітамінів, що містяться у дріжджах, та збереження дріжджового білку.
Проте, екстрагування із дріжджів ергостерину та водорозчинних вітамінів пов’язано із значними складнощами, оскільки вітаміни міцно зв’язані із білковою частиною дріжджової клітини. Тож для вилучення вітамінів необхідно руйнувати білкові комплекси клітин, що може здійснюватися різними методами, зокрема гідролізом або автолізом дріжджової біомаси.
Гідроліз білку може здійснюватись шляхом нагрівання і кип’ятіння дріжджів, розведених і підкислених молочною або соляною кислотою до рН 5, під невеликим тиском в автоклаві. За допомогою цього методу можливим є комплексне використання дріжджів з отриманням концентрату вітамінів групи В, кристалічного ергостерину і білкової кормової сировини.
За методом ферментативного гідролізу дріжджову біомасу піддають автолізу за температури 45 °С. В процесі автолізу фермент, що міститься в дріжджах, руйнує білкові речовини протоплазми, звільнюючи вітаміни, що входять в їх склад.
Автолізовані або гідролізовані дріжджі обробляють спиртом для вилучення вітамінів групи В і для забезпечення належної здатності до фільтрації. Спиртом або іншим розчинником екстрагують стерини, а дріжджовий шріт після вилучення спирту використовують як кормовий продукт.
Для зневоднення дріжджових клітин та коагуляції білку біомасу обробляють спиртом. Дріжджові клітини стають щільними, легко осідають на дно і добре фільтруються.
Тож, для максимального вилучення вітамінів і використання цінних поживних властивостей дріжджового шроту, технологічну схему варто будувати наступним чином:
1. Для вилучення вітамінів, міцно зв’язаних із білково-ферментними комплексами, і збільшення таким чином їхнього виходу, необхідно піддавати дріжджі гідролізу і автолізу.
2. Для забезпечення належної консистенції необхідно автолізовану (гідролізовану) дріжджову масу піддавати обробці спиртом з екстракцією водорозчинних вітамінів групи В.
3. Для вилучення стеринів і фосфоліпідів висушений дріжджовий шріт варто піддавати додатковій обробці спиртом або іншим розчинником.
4. Отриманий після екстракції стеринів та видалення спирту білково-дріжджовий шріт можна використовувати для харчових або кормових цілей, а також для отримання нуклеїнових кислот [2].
2.2 Накопичення біомаси дріжджів
Приготування поживного середовища.
Середовище повинне відповідати певним вимогам:
а) забезпечувати максимальний приріст біомаси;
б) складатися з відносно дешевих компонентів;
в) мати гарну фільтруючу здатність;
Найбільш оптимальним субстратом для накопичення біомаси дріжджів у процесі отримання ергостерину є н
-алкани, оскільки вони сприяють максимальному виходу цільового продукту. Під час використання н
-алканів проблема складу поживного середовища являється особливо актуальною, оскільки макро- та мікроелементи відсутні у вихідній сировині. За загальноприйнятою схемою в розчин макроелементів вносять наступні солі: сульфат амонію (~1,5 кг/м3
) та хлорид натрію (~0,9 кг/м3
). Розчин мікроелементів готується окремо і складається із ряду сульфатів, що являються джерелом таких речовин, як магній, манган, залізо, цинк, мідь, натрій. Крім того, для підтримки постійного значення рН, що знижується в процесі культивування, в ферментер подається аміачна вода, яка одночасно є додатковим джерелом азоту [3].
Компоненти поживних середовищ для основного процесу біосинтезу та приготування посівного матеріалу подаються у змішувачі 3 та 3а відповідно (див. схему у додатку 1). Підготовлена вода подається у змішувачі із резервуарів 1 та 1а через мірники 2 та 2а відповідно. Приготовані поживні середовища подаються для подальшого проведення стерилізації.
Стерилізація поживного середовища.
Використовується безперервний метод стерилізації. Приготовлені середовища із буферних збірників 4 та 4а подаються в стерилізаційну колону 5 та 5а, через які пропускають гостру пару (тиск пари близько 505 Па). Пару подають зверху по внутрішній трубі, що має щілиноподібні прорізи, завдяки чому він надходить у середовище, швидко її нагріваючи. Середовище в колону подається знизу й рухається по спіралі навколо внутрішньої труби.
Безперервний метод стерилізації має ряд переваг: можливість автоматичного регулювання процесу, швидке й рівномірне нагрівання середовища, забезпечення більш повної стерильності середовища й ін.
Середовища, нагріті в колоні до необхідної для стерилізації температури (~130 °С), надходять у витримувачі 6 та 6а, де вони витримуються певний час за температури 125 – 130 °С. Час витримки залежить від складу середовища й триває 5 – 10 хвилин. Звідси стерильні середовища надходять спочатку у теплообмінники-рекуператори 7 та 7а, а потім у пластинчасті теплообмінники 8 та 8а, де вони охолоджуються до 30 – 35 °С (на виході) [4].
Поживне середовище для отримання посівного матеріалу подається в інокулятор 10.
Процес біосинтезу.
Умови культивування дріжджів загалом визначені наявністю в даному випадку чотирьохфазної системи: газ – вода – рідкі парафіни – клітини дріжджів.
Вивчення розподілу клітин в рідких фазах показало, що 1/3 клітин росте у водній фазі, споживаючи краплі алкану діаметром менше 3 мкм, а близько 2/3 клітин адсорбується на поверхні парафінових крапель діаметром від 3 до 5 мкм. Адсорбція на краплях великого діаметру незначна, тому диспергування н
-алканів у водній фазі має велике значення в процесі культивуванні. Окрім перемішування на ефективність диспергування впливає поверхневий натяг, тому внесення ПАР знижує поверхневий натяг і підвищує питому швидкість росту дріжджів.
Оскільки вуглеводневі субстрати можуть засвоюватись дріжджами тільки в аеробних умовах, то на ефективність процесу великою мірою впливає аерація середовища. На окиснення н
-алканів потребується в 2,6 – 2,8 разів більше кисню, ніж на окиснення вуглеводів, що викликає значне тепловиділення під час процесу культивування. Таким чином, одночасно з проблемою підвищення інтенсивності аерації виникає проблема забезпечення ефективного тепловідведення.
Великий вплив на ріст і розвиток клітин дріжджів чинить рН середовища, значення якого в процесі ферментації підтримується на рівні 4,0 – 4,5. Проте, в якості проміжного продукту окиснення н
-алканів виділяються жирні кислоти, що знижує рН водної фази в процесі вирощування і потребує постійної подачі лужного агенту в ферментер. Одним із головних параметрів процесу культивування являється температура середовища, яка не повинна підтримуватись на рівні 32 – 34 °С. Вирощування за більш високих температур (більше 36 °С) призводить до зменшення швидкості росту клітин, зменшення активності біосинтезу білкових речовин, і як результат, зниження економічного коефіцієнта засвоювання субстрату. Перебіг процесу за занадто низьких температур (нижче 28 °С) також призводить до зниження швидкості росту, тобто відбувається зниження продуктивності ферментеру.
Всі секції послідовно сполучені між собою. Для подачі повітря і перемішування культуральної рідини в кожній секції змонтований ежектор, який обертається від двигуна, встановленого на кришці ферментера. Ежектор є двоярусною конструкцією з нижньою та верхньою горловинами для входу та виходу рідини. Повітря засмоктується ежектором і рухається по трубі, яка герметично сполучена з турбіною. Для створення необхідної циркуляції культуральної рідини в кожній секції змонтовані дифузори, перегородки, конічні вставки. На дифузорах змонтовані змійовики-теплообмінники для охолодження культуральної рідини [5].
Суспензія клітин послідовно проходить через усі секції апарату. Із останньої секції виходить суспензія із мінімальним вмістом н-парафінів і максимальною концентрацією біомаси. Це досягається наявністю в ферментері Б-50 двохступеневого культивування: в секціях 1 – 9 – відбувається активний ріст і розвиток дріжджових клітин, а в 10 – 12 – процес «дозрівання» біомаси. Цей процес передбачає витримування біомаси в секціях, до яких припинена подача субстрату (н
-алканів) та поживних речовин. Життєдіяльність клітин відбувається за рахунок ендогенних джерел живлення із одночасним використанням залишкових вуглеводнів. Стадія «дозрівання» (голодування) дріжджів спрощує їх виділення та виключає необхідність екстрактивної очистки біомаси від залишкових вуглеводнів.
Встановлений в кожній секції ежекційний щілинний перемішувально-аеруючий пристрій забезпечує достатнє перемішування та інтенсивне змішування повітря із рідиною, насичуючи її киснем.
Для відведення тепловиділень із ферментера, що складає близько 34 – 40 МДж/кг, в кожну секцію вбудовані змійовики з теплообмінною поверхнею 3000 м2
.
Час вирощування дріжджів складає 8 – 8,5 год. Витрата повітря на одиницю робочого об’єму 113 м3
/м3
∙год.
Засів проводять великою кількістю інокуляту. Культивування проводять за високої температури і сильної аерації середовища, що містить великий надлишок джерел вуглецю по відношенню до джерела азоту [3].
Отримання біомаси дріжджів.
Культуральну рідину із ферментера 9 перекачують у двохступеневий флотатор 11, де відбувається початкове концентрування дріжджових клітин у їх суспензії. Із флотатора суспензія перекачується через деаератор 12, де відбувається відділення розчинених у суспензії газів. Підготовлена таким чином суспензія проходить дві стадії сепарування на сепараторах 13. Концентрована суспензія дріжджових клітин подається у буферний резервуар 14, з якого вона вже безпосередньо потрапляє на розпилювальну сушарку 15. Далі суха біомаса дріжджових клітин надходить на стадії отримання ергостерину (провітаміну D2
)[2].
2.3 Виробництво концентрату вітамінів групи В
Біомаса дріжджів подрібнюють в шнековому пресі 1 (додаток 2) і подають в автоклав 2, де вона піддається кислотному гідролізу за температури 110 °С протягом 20 – 30 хв. До 100 кг обробленої біомаси додають 20 л води і 10 мл соляної кислоти (густина 1190 кг/м3
). Кислотний гідроліз підвищує вихід ергостерину за рахунок вивільнення його частки, що перебуває у зв’язаному із білками стані. Із автоклаву дріжджі потрапляють у коагулятор 3 із зворотнім холодильником 4, куди на кожні 100 кг обробленої біомаси вводять 85 л етилового спирту із доведенням його об’ємної частки в розчині до 50 %. В коагуляторі масу витримують 40 – 50 хв за інтенсивного перемішування та температури 75 – 78 °С. Відбувається коагуляція білкових речовин, що полегшує подальший процес фільтрування та екстрагування водорозчинних вітамінів. Після цього оброблену масу подають в охолоджувач 5, де її охолоджують до 10 °С і потім фільтрують через барабанний вакуум-фільтр 6. Фільтрат подається в збірник 7, а із нього – у вакуум-випарювальний апарат 8 з поверхневим конденсатором 9. Конденсат спирту подають в збірник спирту-дистиляту 10, після чого подається на ректифікацію. Після відгонки спирту і частини води у вакуум-прегонному апараті залишається рідкий концентрат вітамінів групи В із вмістом сухих речовин 50 %. Концентрат подають у збірник 11, після чого, в залежності від призначення кінцевого препарату вітамінів, його подають на стадії додаткової очистки та розфасовки.
Білковий залишок з вакуум-фільтра 6 подається в дистиляційний апарат 12 для відгонки спирту.
Після відгонки спирту білковий залишок подають в бункер 13, а із нього у вальцьову сушарку 14, де вміст вологи в ньому зменшують до 2 %.
Отримані висушені пластівці подрібнюють в дробарці 15 до борошна, яке подається на подальшу обробку [2].
2.4 Виробництво концентрату провітаміна
D
2
та технологія його трансформації у вітамін
D
2
Отримання ергостерину-сирця.
В екстракторі 16 із зворотнім холодильником дріжджове борошно обробляють трикратним об’ємом спирту-ректифікату за температури 78 °С. Потім дріжджі відфільтровують в друк-фільтрі 17. Перший екстракт подають в збірник 18, а осад – у другий екстрактор 19, де його знову обробляють трикратним об’ємом спирту. Білковий осад відфільтровують на друк-фільтрі 20. Другий екстракт також подають у збірник 18. Із збірника 18 отриманий екстракт подається у вакуум-перегонний апарат 21, де спирт випаровують під вакуумом. Спирт після відгонки потрапляє в збірник 22, звідки він потім подається на ректифікацію. Фільтрат після відгонки спирту випарюють до вмісту сухих речовин 70 %. Із 100 кг сухих дріжджів отримується 20 – 25 кг ліпідного концентрату.
У вакуум-апарат вносять 45 %-й розчин гідроксиду натрію з розрахунку 6 кг на 100 кг отриманого борошна. Відбувається омилення жирів. Омилений розчин ергостерину кристалізують за температури 0 °С в кристалізаторі 23. Отримані кристали ергостерину-сирця відфільтровують на нутч-фільтрі 24 і подають на перекристалізацію. Міжкристальний спиртовий розчин подають у вакуум-перегонний апарат 25 для відгонки спирту. Лужний розчин, що залишається в перегонному апараті подається на утилізацію. Спирт-конденсат із конденсатора 26 подають в збірник 27, звідки потім потрапляє на ректифікацію. Білкову масу із друк-фільтра 20 направляють в дистилятор 28, де її розводять водою, кількість якої дорівнює подвійній масі осаду. Випарений з дистилятора 28 спирт направляють в збірник 29. Білкова маса із дистилятора після нейтралізації подається в бункер 30, а потім у вальцьову сушарку 31, дробарку 32 і далі на розфасовку.
Перекристалізація ергостерину-сирця.
Ергостерин-сирець піддають афінації в апараті 33 спиртом (65 % мас.) в кількості 10 кг на 1 кг сирця. Афіновану масу подають на друк-фільтр 34. Відфільтровані афіновані кристали ергостерину направляють на перекристалізацію в апарат 35 в спирто-бензольному розчині (4:1). Маточний розчин, отриманий під час афінації, подається в збірник 36, а потім в перегонний апарат 37 для відгонки спирту. Із перегонного апарату спирт потрапляє в збірник 38, із якого подається в збірник 63а і ректифікаційну колону 62. Розчин ергостерину в спирто-бензольній суміші із апарату 35 подають в кристалізатор 39 і нутч-фільтр 40. Кристали ергостерину І після промивання висушуються у вакуум-сушильному апараті 41 (температура плавлення кристалів не повинна бути менше ніж 160 °С). Маточник кристалізації І потрапляє в збірник 42, а потім у вакуум-перегонний апарат 43 для відгонки спирто-бензольного розчинника, який збирають в приймачі 44.
Згущений маточник піддають кристалізації в апараті 45. Кристали ергостерину ІІ відділяють від маточника ІІ на нутч-фільтрі 46.
Ергостерин ІІ подають на афінацію разом із ергостерином-сирцем. Маточник ІІ із нутч-фільтра 46 подається в збірник 47, а із нього у вакуум-випарний апарат 48. Випарений спирт подають в збірник 49, звідки він через збірник 44а подається на повторне використання.
Опромінення ергостерину і отримання концентрату вітаміну
D
2
.
Рядом досліджень було доведено, що в ефірному розчині процес активації ергостерину протікає не так інтенсивно, як в спирті. Якщо в спирті максимум активації досягається через 30 – 40 хв, то в середовищі ефіра через 150 – 200 хв, що дозволяє уникнути переопромінення розчину. Це особливо важливо у разі виробництва кристалічного ергокальциферолу, оскільки продукти переопромінення ергостерину утримують ергокальциферол в розчині, перешкоджаючи його кристалізації. Сухий ергостерин І опромінюють в розчині сірчаного ефіру. Вважають, що ергостерин спочатку ізомерується в прекальциферол, який під час нагрівання переходить в ергокальциферол і частково в люмістерин. В реактор 50 із сушарки подають ергостерина, а із збірника 51 – ефір. Отриманий розчин фільтрують через друк-фільтр 52, фільтрат через мірник 53 подають в збірник 54, звідки він безперервно протікає через опромінювач 55. Опромінений розчин потрапляє в збірник 56, потім в перегонний апарат 57 для відгонки ефіру і виділення непрореагованого ергостерину, який відфільтровують в друк-фільтрі 58 і подають на перекристалізацію. Фільтрат стандартизується по активності олією в змішувачі 59, фільтрується через нутч-фільтр 60 в збірник 61, звідки подається на розфасовку [2].
3.
Розрахунок Ферментера марки Б-50
3.1 Розрахунок габаритних розмірів ферментера
Продуктивність ферментера:
P
= 200
кг/год (за вологою біомасою);
Коефіцієнт заповнення ферментера:
0,7
;
Концентрація дріжджових клітин в культуральній рідині:
C
= 20
кг/м3
(за вологою біомасою);
Тривалість одного циклу культивування: Т = 12 год;
Кількість секцій ферментера:
n
= 12.
Культивування проводиться в два етапи: ріст біомаси та витримка біомаси, т.з. «голодування», протягом якого дріжджами споживаються залишки вуглеводнів, отриманих із поживного середовища. Етап росту триває 9 год, витримка біомаси триває 3 год. Процес розраховано таким чином, що в секціях 1 – 9 відбувається ріст біомаси, а в секціях 10 – 12 – дозрівання. Тож тривалість одного циклу в кожній секції становить 1 год.
Запуск ферментера відбувається із заповненням секції 1 поживним середовищем та внесенням посівного матеріалу. По завершенню 1-го циклу (1 год) вміст секції 1 перекачується в секцію 2, а секція 1 знову заповнюється поживним середовищем та інокулятом. Через 12 год після початку процесу, із секції 12 відбирається культуральна рідина і направляється на виділення біомаси, а в звільнену секцію 1 знову подається свіже поживне середовище та вноситься культура продуцента. Тож продуктивність ферментера кількісно дорівнюватиме об’єму культуральної рідини, яка зливається із секції 12. Оскільки продуктивність ферментера нам відома – P = 200 кг/год (за вологою біомасою), а концентрація дріжджових клітин в культуральній рідині становить C = 20 кг/м3
, то об’єм культуральної рідини в одній секції становитиме:
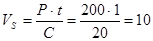 м3
,
м3
,
деt – тривалість одного циклу культивування, год.
Об’єм однієї секції, враховуючи коефіцієнт заповнення 0,7, становитиме:
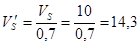 м3
,
м3
,
Габаритні розміри ферментера являють собою висоту секцій (h), зовнішній (R) та внутрішній (r) радіуси тороїда . Висота, при чому, дорівнює різниці зовнішнього та внутрішнього радіусів (R – r).
Об’єм однієї секції становитиме:
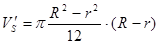 . .
Значення зовнішнього та внутрішнього радіусів R та rвизначаються із розрахункової таблиці (див. додаток 3) за відомим значенням об’єму ферментера V`S
.
У додатку 3 найближче значення об’єму секції ферментера (14,33 м3
) лежить на перетині значень зовнішнього та внутрішнього радіусів 4,4 м та 1,3 м, відповідно. Тож для конструювання даного ферментера приймемо наступні габаритні розміри:
3.2 Розрахунок товщини стінки ферментера
Розрахунок проводять за формулою:
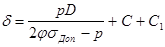
де δ – товщина стінки, м;
р – розрахунковий тиск, МПа (стерилізація ведеться за тиску гострої пари 0,3039 МПа);
D – діаметр апарата, м. В даному випадку, коли розрахунок товщини стінки ведеться для однієї секції, неможливо виділити значення діаметру, оскільки основа секції має форму трапеції. Тому, замість діаметру використаємо розрахунковий розмір А, що чисельно дорівнює середньому арифметичному висоти та середньої лінії трапеції. За габаритних розмірів тороїда (див. п. 3.1.) R = 4,4 м та r = 1,3 м, значення А становитиме:
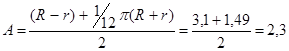 , м. , м.
φ – коефіцієнт міцності зварних швів, який становить 0,7 – 1 (приймемо 0,85);
σДод.
– напруга, що допускається, МПа (для неіржавіючої сталі за температури 132,9 °С, тобто за температури насиченої пари і стерилізації, σДод.
=140 МПа);
С – додаток до розрахункової товщини для компенсації корозії, м; С = Пτа
, де П – корозійна проникність, м/рік (приймемо, що П = 10-4
м/рік), τа
– амортизаційний термін служби апарата, років (приймемо 10 років);
С1
– додаток на заокруглення до цілого числа міліметрів.
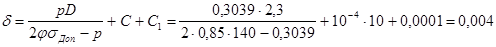 м. м.
- товщина стінки ферментера δ = 4 мм
[5].
Висновки
1. Процес накопичення біомаси, збагаченої провітаміном D2
(ергостерином), найкраще проводити на поживному середовищі, що містить н
-алкани, як найоптимальніший субстрат для отримання цільового продукту.
2. Для отримання біомаси продуцента на середовищі, збагаченому н
-алканами, рекомендовано використовувати 12-секційний тороїдний біореактор марки Б-50. Даний ферментер забезпечує специфічні особливості кінетики накопичення біомаси та умови перемішування і аерації.
3. Для вилучення вітамінів, міцно зв’язаних із білково-ферментними комплексами, і збільшення таким чином їхнього виходу, необхідно піддавати дріжджі гідролізу і автолізу.
4. Для забезпечення належної консистенції необхідно автолізовану (гідролізовану) дріжджову масу піддавати обробці спиртом з екстракцією водорозчинних вітамінів групи В.
5. Для вилучення стеринів і фосфоліпідів висушений дріжджовий шріт варто піддавати додатковій обробці спиртом або іншим розчинником.
6. Отриманий після екстракції стеринів та видалення спирту білково-дріжджовий шріт можна використовувати для харчових або кормових цілей, а також для отримання нуклеїнових кислот.
7. Для конструювання ферментера марки Б-50 продуктивністю 200 кг вологої біомаси на годину приймемо наступні габаритні розміри:
- зовнішній радіус тороїда R
= 4,4 м
;
- внутрішній радіус тороїда r
= 1,3 м
;
- висота секції h
=
R
–
r
= 4,4 – 1,3 = 3,1
м
.
- товщина стінки ферментера δ = 4 мм
.
Список використаної літератури
ергостерин вітамін ферментер синтез
1. Под ред. Н.С. Егорова. Промышленная микробиология: Учеб. пособие для вузов по спец. «Микробиология» и «Биология». – М.: Высш. шк., 1989. – 688 с.: ил.
2. Шнайдман Л.О. Производство витаминов. – М.: издательство «Пищевая промышленность», 1973. – 444 с.
3. Под ред. Егорова Н.С. Биотехнология т. 5: Производство белковых веществ – М.: Высш. шк., 1986. – 142 с.: ил.
4. Тимощенко Л.В., Чубик М.В. Основы микробиологии и биотехнологии: Учебное пособие . – Томск: Изд-во Томского политехнического университета, 2009. – 194 с.
5. Сидоров Ю.І., Влязло Р.Й., Новіков В.П.. Процеси і апарати мікробіологічної промисловості. Технологічні розрахунки. Приклади і задачі. Основи проектування виробництв. У 3 ч. – Ч. І. Ферментація. – Львів: Видавництво Національного університету «Львівська політехніка», 2004. – 240 с.
6. Бекер М.Е. Введение в биотехнологию. Пер. с латышского. – М.: издательство «Пищевая промышленность», 1978. – 228 с.
7. Касаткин А.Г. Основные процессы и аппараты химической технологии. Издание седьмое. – М.: Государственное научно-технологическое издательство химической литературы, 1961. – 831 с.
8. Под ред. К.А. Калунянца. Оборудование микробиологических производств. – М.: Агропромиздат, 1987. – 389 с.
|
