Введение
Реки занимают исключительное место в жизни человека. Они играют большую роль в обеспечении водой населения, сельского хозяйства и промышленности. На ряде рек проводится разработка рассыпных месторождений полезных ископаемых. Многие из них являются источниками строительных материалов, используются для сплава леса и перевозки грузов. Велика их рекреационная ценность – как традиционных мест отдыха, любительского рыболовства и водного туризма. На многих реках оборудованы туристические маршруты, действуют дома отдыха и здравницы. Ряд рек являются местами обитания промысловых видов рыб.
В Пермской области насчитывается 29 тыс. рек, из них две крупных, сорок средних, около 29 тыс. малых; общая протяжённость водотоков превышает 90 тыс. км (Комлев, Черных, 1984).
В настоящее время реки Прикамья изучены недостаточно, но они имеют большое практическое значение. Поэтому следует обратить особое внимание на проблему их охраны, в решении которой необходимым этапом является формирование представлений о естественном состоянии речных экосистем. Это представление может быть положено в основу регионального гидроэкомониторинга в качестве эталона экологической нормы.
Значительного обилия в реках достигает зообентос, который служит пищей для многих беспозвоночных и позвоночных животных.
Зообентос реки Сылвы изучен достаточно хорошо (Громов, 1959; Алексевнина, Буйдов, 1997; Паньков, 2004). В результате проведённых работ составлены видовые списки основных групп бентосных животных. Изучено пространственное распределение донных беспозвоночных. Выявлены закономерности сезонной динамики их сообществ. Установлены особенности межгодовой динамики, раскрыта связь между количественными показателями развития зообентоса и изменениями уровня воды в реке, показана связь устойчивости донной фауны к паводкам с гранулометрическим составом грунта. Исследовано питание ряда видов беспозвоночных, изучена трофическая структура речного бентоса, выявлены трофические зоны и установлены закономерности их распределения в речном континууме.
В то же время дрифт р. Сылвы изучен не достаточно, хотя это явление – одно из важнейших аспектов жизни зообентоса.
Реклама

Термином «дрифт» в современной гидробиологии обозначают как совокупность организмов, дрейфующих по речной струе по воле течения, так и явление такового дрейфа.
Явление дрифта впервые привлекло внимание гидробиологов в 30-е – 40-е годы XX века, когда ученые обнаружили массовый снос бентосных животных при прохождении полых и паводковых вод. С начала 60-х годов как у нас в стране, так и за рубежом начинаются интенсивные работы по всестороннему изучению этого явления.
Дрифт беспозвоночных – очень важное явление в жизни текучеводных экосистем. Миграции донных беспозвоночных в толще речной струи играют большую роль в динамике отдельных зообентоценозов и общем распределении донной фауны в русле водотоков. Дрифт является механизмом, обеспечивающим более или менее оптимальное размещение животных в бентали в соответствии с ресурсами среды и потребностями отдельных видов. Благодаря миграциям с вышележащих участков, происходит восстановление разрушенных сообществ и реколонизация опустошенных биотопов после воздействия экстремальных факторов гидрологического режима.
Наконец, дрейфующие беспозвоночные используются в пищу многими видами рыб; их роль особенно велика в горных и предгорных реках, где слабо развит зоопланктон, а животные, скрывающиеся в скважинах грунта, малодоступны (Шубина, 1986). В частности, преимущественно организмами дрифта питается молодь лосося (Шустов, 1977).
Несмотря на многолетние усилия специалистов, исследование дрифта далеко от своего завершения и хранит еще много тайн, а имеющиеся сведения нередко противоречивы и ждут своего объяснения. Таким образом, продолжение работ в этой области представляется актуальным.
Целью моей работы является изучение дрифта донных беспозвоночных р. Сылвы и его суточной динамики в районе заказника «Предуралье» летом 2005–2006 гг.
При этом решались следующие конкретные задачи:
1. Определение таксономического состава дрифта;
2. Изучение вертикального распределения организмов дрифта;
3. Изучение суточной динамики интенсивности дрифта.
1. Обзор литературы
Термином «дрифт» (от англ. drift
– пассивный снос течением) в современной гидробиологии обозначают как совокупность организмов, дрейфующих в речной струе по воле течения, так и явление такового дрейфа. В обоих случаях термин имеет широкую и узкую трактовки. Под дрифтом в «широком смысле слова» подразумеваются как организмы суши, случайно упавшие в воду так и гидробионты; указанное понимание термина свойственно многим специалистам, выделяющим в составе дрифта соответственно «воздушную»
и «водную»
фракции (Шустов, 1977), или аллохтонный
и автохтонный
компоненты (Чебанова, 1992). Дрифтом в «узком смысле слова» обычно называют только животных бентоса, в силу тех или иных причин оказавшихся в толще речной струи (Шубина, 1986; Богатов, 1989); синонимом такого понимания дрифта является более старый термин «бентосток».
Реклама

Оставив в стороне вопрос о «воздушной» фракции, в дальнейшем мы ограничимся рассмотрением дрифта бентосных животных, т.е. будем придерживаться последнего, более строгого определения термина.
Существует несколько подходов к классификации дрифта.
Так, среди дрейфующих по течению организмов, в зависимости от причин, по которым они оказались в толще речной струи, специалисты различают пассивный
(постоянный) и активный
(поведенческий) компоненты. Или «эврисиртон»
– организмы бентоса, которые оказались в струе в результате недостаточного сопротивления течению и «эконосиртон»
, к которому относятся организмы, оказавшиеся в толще воды в связи с особенностями жизненного цикла (Константинов, 1969, Богатов, 1983).
В. Богатовым (1988) предложена иная классификация, в которой за основу взяты не причины, вызвавшие дрифт, а принцип, который учитывает возрастание или убывание биомассы организмов в результате дрифта изучаемых групп гидробионтов на определенном участке реки за сутки.
Использование данного принципа позволило выделить пять типов дрифта бентосных животных.
Нейтральный тип дрифта, в результате которого биомасса гидробионтов на грунте увеличивается только за счет их продукции либо практически не изменяется.
Отрицательный или экстремальный тип дрифта, в результате которого биомасса животных на грунте снижается в результате вертикальных миграций. Такой тип дрифта наблюдается при воздействии экстремальных факторов, которые, однако, не ведут к разрушению сообщества. Например, при попадании в водоток малых концентраций ядохимикатов или в период прохождения паводковых вод.
Положительный тип дрифта, в результате которого биомасса организмов на грунте увеличивается. Положительный тип дрифта наблюдается, как правило, при восстановлении численности на биотопах, подвергшихся ранее воздействию экстремальных факторов.
Катастрофический тип дрифта – это крайняя форма отрицательного дрифта, которая ведет к разрушению сообщества. Такой дрифт может иметь место при значительном антропогенном загрязнении и во время природных катастроф.
Предимагинальный дрифт связан с массовым подъемом в толщу воды взрослых нимф и куколок амфибиотических насекомых для линьки на имаго.
Традиционно большое внимание специалисты уделяют вопросам о причинах дрифта и его биологическом смысле. Многочисленными работами выяснено, что дрифт – это естественная особенность всех обитаемых водотоков мира.
В самой общей форме, дрифт обусловлен двумя процессами: активным подъемом донных беспозвоночных в толщу воды, продиктованным необходимостью в перемене биотопа (в том числе водной среды на воздушную при имагинальном типе дрифта) и пассивным смывом с грунта тех животных, которые не могут противостоять течению (Задорина, 1987б
).
В первое время считалось, что все животные, дрейфующие в толще речной струи, покинули бенталь не по своей воле, т.е. дрифт донных беспозвоночных имеет всецело пассивный характер. Такую его особенность, как приуроченность, большей частью, к темному времени суток, сторонники указанной точки зрения объясняли ночной активностью бентонтов, покидающих убежища и выползающих на поверхность грунта с наступлением сумерек. Позднее было показано, что ночной дрифт не является полностью пассивным процессом; весьма убедительно это продемонстрировали В.Я. Леванидов и И.М. Леванидова (1981, с. 24): «Если допустить, что турбулентные силы в реке, подобной Хору, столь велики, что осуществляют за короткое время (полчаса-час) подъем личинок с глубины до 7 м к поверхности, то столь же резкое опускание их на дно среди ночи или с наступлением дня не может быть объяснено гидродинамическими силами, а объясняется только их целенаправленными движениями. Логично предположить, что если бы личинкам не было целесообразно сплывать в толще речной струи, то подхваченные течением, они активно возвращались бы на дно в самом начале этого процесса, при этом влеклись бы какое-то время в нижнем горизонте. В действительности же численность личинок в придонном слое почти всегда в 4 – 5 раз ниже, чем у поверхности».
Авторы цитированной работы делают вывод, что в дрифте одновременно могут находиться животные, попавшие в толщу речной струи по разным причинам: одну группу составляют беспозвоночные, покинувшие бенталь по своей воле, другая представлена бентонтами, сорванными с грунта течением.
Как различать пассивный и активный компоненты на практике, в настоящее время не вполне ясно, поскольку предложенные специалистами признаки того и другого уязвимы для критики. Принято считать, что пассивный дрифт, в отличие от активного, характеризуется постоянством параметров в течение суток (Чебанова, 1992).
Было бы также неправильным считать, что дневной дрифт является всецело пассивным процессом, поскольку активность отдельных групп донных животных в светлое время суток может сопровождаться их целенаправленными миграциями в толще воды. Последние могут быть спровоцированы, например, столкновениями конкурирующих особей или хищников и их жертв. Кроме того, хорошо известно, что молодь амфибиотических насекомых, в частности, некоторых видов хирономид и ручейников, ведет временный планктонный образ жизни (Константинов, 1958; Лепнева, 1964), причем личинки в это время демонстрируют положительный фототаксис, не свойственный особям старших возрастов, обитающих в бентали. Наконец, к светлому времени суток бывает приурочен вылет ряда групп амфибиотических насекомых, которому предшествует активный предимагинальный дрифт (Леванидов, Леванидова, 1981).
Заслуживает более детального обсуждения вопрос о биологическом смысле поведенческого дрифта (пассивный, по определению, такого смысла не имеет). В литературе высказано немало соображений на этот счет. Одним из первых было предположение, что бентосные животные активно поднимаются в толщу речной струи, избегая напряженной конкуренции за пищу и пространство (Леванидова, Леванидов, 1965), т.е. внешней причиной вертикальных миграций организмов является перенаселенность бентали. Последняя может возникнуть из-за сокращения площади речного дна при понижении уровня воды, увеличения численности донных беспозвоночных в результате размножения и резкого возрастания биомассы зообентоценозов вследствие параболического роста животных на последних стадиях развития (Леванидова, 1968).
Распространенным доводом в пользу указанной точки зрения служило наблюдение, что количественные характеристики дрифта находятся в прямой зависимости от плотности организмов на грунте (Шустов, 1978; Шубина, 1986; и др.).
Однако, В.В. Богатовым (1983) путем несложных математических выкладок было показано, что данный аргумент не вполне убедителен, поскольку принятые в то время показатели дрифта, не учитывающие скорости течения, не давали возможности оценить величину сноса животных с единицы площади речного дна и, следовательно, не были сопоставимы с показателями развития донной фауны.
Поскольку конкуренцию за место и пищу, как оказалось, нет оснований считать одной из причин целенаправленных вертикальных миграций бентосных животных, традиционное объяснение поведенческого дрифта утратило свою привлекательность. Ему на смену пришло иное понимание этого явления.
В.В. Богатовым (1983) было показано, что интенсивность дрифта (Nd
– количество животных, снесенных через единицу площади живою сечения потока за единицу времени) определяется двумя основными параметрами: миграционной активностью животных (М – количество организмов, снесенных за единицу времени с единицы площади бентали) и средней дистанцией их дрифта (L). В свою очередь, миграционная активность определяется относительной миграционной активностью (М» – доля животных бентоса, принявшая участие в вертикальных миграциях за определенный отрезок времени) и плотностью животных в бентали, а средняя дистанция дрифта является функцией от времени нахождения животных в толще воды (Т) и скорости течения (V).
Осуществленный анализ позволил перейти к более детальному осмыслению природы активного дрифта. Из вышеперечисленного следует, что интенсивность дрифта, среди прочих, зависит и от двух поведенчески обусловленных параметров: удельной миграционной активности животных и временем их нахождения в толще воды (Богатов, 1983).
Установлено, что эти параметры зависят как от некоторых факторов среды, так и особенностей биологии мигрирующих организмов (прежде всего, их видовой принадлежности и стадии онтогенеза). Так, оказалось, что наивысшие показатели М» и Т характерны для самых молодых особей, что хорошо согласуется с эмпирически регистрируемым преобладанием в составе дрифта животных ранних стадий развития. Считается, что биологический смысл этого явления состоит в обеспечении рассредоточения молоди донных животных после выхода из кладок (Лепнева, 1964; Леванидов, Леванидова, 1981).
В некоторых случаях повышенная склонность к вертикальным миграциям обнаруживается у животных, испытывающих потребность в перемене биотопа, связанную с завершением той или иной стадии индивидуального развития. Характерные примеры этого приводят В.Я. Леванидов и И.М. Леванидова (Леванидова, 1968; Леванидов, Леванидова, 1981): личинки ручейников Arctopsyche
,
Macronema
,
Amphipsyche
и Hydropsyche
живут в быстротекучих водах, но перед окукливанием поднимаются к поверхности и массами сплывают вниз по течению, оседая на спокойных участках реки; молодые личинки поденок Epeorus
(
Iron
)
latericius
Sinitsh. обитают по всему руслу горных речек, взрослые же нимфы – только на быстрых, мелких перекатах.
В условиях р. Сылвы на протяжении личиночной стадии развития меняют биотопы поденки Potamanthidae и Ephemeridae; их молодые личинки встречаются по всему руслу, зрелые же нимфы – только в рипали (Паньков, 2002).
Здесь же следует упомянуть и об имагинальном дрифте, представляющем яркий пример радикальной смены биотопа по завершении крупного этапа онтогенеза.
Таким образом, в нормальных условиях обитания динамика дрифта обусловлена не только численностью донной фауны, но и характером жизненных циклов беспозвоночных определяющих, так сказать, «фонд мигрантов» в зообентосе (Леванидова, 1968).
Весьма мощное воздействие на миграционное поведение гидробионтов оказывает ухудшение условий обитания; избегая воздействия неблагоприятных факторов среды, организмы поднимаются в толщу воды и сплывают вниз по течению. Известно, что интенсивность дрифта резко возрастает при нагрузке потока взвешенными и влекомыми веществами, ухудшении респираторных условий (уменьшении расходов и скорости течения воды, ее сильном прогреве, снижении концентрации кислорода), изменении рН, токсическом загрязнении, нарушении стабильности донных отложений (Ключарева, 1963). Миграции в толще воды завершаются оседанием животных на более пригодных для жизни участках реки, при этом бентонты демонстрируют высокую избирательность к субстратам (Леванидова, 1968).
Таким образом, к настоящему времени явление дрифта изучено весьма обстоятельно, что позволяет говорить о вполне сложившемся в рамках речной экологии самостоятельном направлении научных исследований. О зрелости этого направления свидетельствует, в частности, весьма дифференцированный подход к предмету изучения, обнаруживающий себя, например, в различных схемах классификации дрифта.
2. Описание района исследования
2.1 Физико-географическая характеристика района исследования
Физико-географические условия на территории Пермского края в настоящее время достаточно хорошо изучены (Коробков, 1959; Шкляев, Балков, 1963; Проблемы физической…, 1966; Комлев, Черных, 1984).
Территория Прикамья охватывает части трех крупных тектонических элементов: Главного Уральского мегаантиклинория, Предуральского краевого прогиба и Русской (Восточно-Европейской) платформы.
Восточную часть края занимает западный склон Северного и Среднего Урала, представляющий собой увалы высотой 500 – 700 м над уровнем моря. Среди них отдельные каменистые возвышенности достигают высоты более 1000 м. В седловинах между увалами часто встречаются заболоченные участки.
Западнее расположено предгорье Урала, которое постепенно переходит в Русскую равнину. Предгорье с севера на юг пересекается долиной Камы. В северной части края имеются плосковершинные возвышенности высотой до 200 м, покрытые лесом, – «пармы». По западу края тянутся Вятские увалы. Центр края между предгорьем Урала и Вятскими увалами носит характер слегка волнистой равнины с отметками над уровнем моря до 200 – 250 м. Южную часть занимает северное окончание Уфимского плато, представляющее собой возвышенную волнистую равнину, в значительной степени расчлененную глубоко врезанными долинами.
Наличие горной и равнинной частей обуславливает значительные климатические различия на территории края.
Основной воздушной массой, формирующей климат края, служит континентальный воздух, что связано с особенностями географического положения Уральских гор. Будучи вытянутыми в меридиональном направлении, горные цепи Урала находятся на пути движения западных влагонесущих масс; в то же время они не препятствуют вторжению арктических масс воздуха, проникающих на юг.
Это находит свое подтверждение в значительных годовых и суточных колебаниях почти всех метеорологических элементов.
Величина суммарной солнечной радиации увеличивается с севера на юг с 78,3 до 91,3 ккал/см2
в год. Средняя годовая температура воздуха в равнинной части изменяется с севера на юг от –2 до +20
С. В горных районах с увеличением высоты местности над уровнем моря наблюдается понижение температуры на 0,5 – 0,70
С на каждые 100 м.
Распределение годовой суммы осадков по территории Прикамья чрезвычайно неравномерно. Их наибольшее количество выпадает на хребтах и склонах гор.
Преобладающее количество осадков – до 60 – 70% выпадает в виде дождя в летнее – осенний период. Осадков холодного сезона гораздо меньше, чем летних, несмотря на продолжительную зиму. В горной части территории пик сдвигается на октябрь – ноябрь. Число дней с осадками уменьшается с севера на юг с 200 до 140.
Высота снежного покрова в пределах края не отличается особой пестротой, хотя характерно убывание ее с северо-востока на юго-запад. Снег лежит в среднем 161 – 192 дня.
Важным климатообразующим фактором является температурный режим воздуха. От температуры воздуха зависят интенсивность таяния снега, замерзание рек и многие другие явления. На территории края среднегодовые температуры, в среднем близкие к нулю градусов, понижаются с юго-запада на северо-восток. Самый теплый месяц – июль, самый холодный – январь.
Ветры в Пермской края определяются общими воздушными течениями, характерными для всего востока Европы. Преобладающее направление юго-западное. Ветра вызываются местными воздушными течениями, образующимися вследствие изменения температуры воздуха с высотой.
Расположение речной сети определяется рельефом местности.
Пермский край имеет густую речную сеть – до 545 малых рек длиной около 30 тыс. км.
Особенно густа речная сеть на северо-востоке края, где на 1 км2
территории приходится 21 – 23 км рек. В центральной части и на крайнем юго-западе края густота речной сети составляет 17 – 19 км на 1 км2
площади.
Основной рекой края является Кама, протекающая в его пределах на протяжении 1000 км. Река берет начало от Верхнекамской возвышенности с высотами до 380 м, расположенной на западе Прикамья южнее Веслянской низменности.
Всю речную сеть Пермского края можно разделить на бассейны Северного Прикамья, реки Вишеры, Среднего Прикамья, реки Чусовой, Южного Прикамья и реки Белой.
Река Сылва предгорного типа, берет начало в районе крайней западной гряды среднего Урала и протекает, большей частью, по территории Приуральской равнины.
Река Сылва в прошлом являлась самым крупным притоком Чусовой; ныне, после завершения гидростроительства, она впадает в Сылвенский залив Камского водохранилища.
Длина реки, достигавшая ранее 587 км (Справочник…, 1936; Коробков, 1959), с заполнением водохранилища значительно сократилась и составляет ныне 493 км, площадь водосбора соответственно уменьшилась с 22140 км2
до 19700 км2
(Коробков, 1959; Ресурсы поверхностных…, 1967). Средняя высота водосбора составляет 227 м, при среднем уклоне русла 0,3%, мутность вод до 150 г./м3.
Река протекает в умеренно-континентальном климате, наибольшая температура воды достигает +25º C.
В бассейне р. Сылвы широкое распространение получили карстовые явления. Густота речной сети близка к средней по Пермской края: 0,5–0,6 км/км2
. Модуль годового стока равен 7,5 л/с с 1 км2
.
По классификации М.И. Львовича, Сылва относится к рекам преимущественно снегового питания, кроме этого, значительную часть в питании рек составляют дождевые и грунтовые воды. Высокое продолжительное половодье наблюдается около ста дней в году. Характерны устойчивая летне-осенняя межень и низкая зимняя. Летне-осенняя межень часто прерывается дождевыми паводками. Залесенность (57%) и закарстованность (23%) бассейна оказывают регулирующую роль во внутригодовом распределении стока, в связи с этим р. Сылва не пересыхает в засушливые периоды и не перемерзает даже в особо суровые зимы. Амплитуда многолетних колебаний уровня воды на реке у с. Подкаменное составляет 9,2 м.
Средняя продолжительность ледостава 157 дней, ледовые образования появляются около 1 ноября, ледостав наступает в первой декаде ноября. Весенний ледоход начинается в середине апреля, к концу апреля река, как правило, полностью очищается ото льда. Средняя толщина льда в конце зимы составляет 60 см.
Комплексный заказник «Предуралье» расположен в среднем течении реки Сылвы. Его территория вытянута вдоль реки Сылвы. Лишь в отдельных местах она достигает ширины 1 – 3 км, в основном же ограничена склонами каньонообразной речной долины (Наумкин, Севастьянов, Лавров, 2004). Русло реки в районе заказника асимметрично. Асимметричность проявляется в наличии больших глубин у левого берега (более 3,5 м) и отмелей у правого. Береговая линия выражена хорошо у обоих берегов. В период половодья пойма ежегодно заливается водой, ширина реки в этот период достигает 120 м. В отдельные многоводные годы вода может заливать и часть первой надпойменной террасы, а в годы катастрофического половодья (1979) практически всю террасу. Средняя ширина реки Сылвы в межень 70–90 м при средней глубине 1,4 м. В пределах территории заказника р. Сылва не принимает ни одного притока (Воронов, Циберкин и др., 2000).
Среднее течение выделяется на основании общих для него признаков: сходных на всем его протяжении термического режима, характера грунтов, морфологии русла (наличие плесов с выраженной рипалью и перекатов). Для среднего течения характерно обильное развитие высшей водной растительности, преимущественно рдестов (Паньков, 2004).
По особенностям термического режима Средняя Сылва, согласно классификации В.Я. Леванидова (1981), принадлежит к группе умеренно тепловодных водотоков. Преобладающие грунты на перекатах и медиали плесов – валунные и галечные, в рипали – заиленные, со значительной примесью песка. На основании этих признаков среднее течение может быть отнесено к одной зоне – гипоритрали с признаками метапотамали. Вместе с тем на столь значительном протяжении среднее течение реки не остается единообразным: в его пределах выделяется четыре участка.
Комплексный заказник «Предуралье» расположен на Кишертско – Кунгурском участке (пос. Кишерть – г. Кунгур), который имеет длину около 20 км. На этом участке Сылва пересекает Сылвенский кряж (Уфимский вал), что сказывается на ее характере. Река течет в узкой долине среди высоких крутых берегов, изобилующих обнажениями коренных пород. Интенсивность руслового процесса понижена; в силу этого система пойменных водоемов практически отсутствует. Ширина реки местами уменьшается до 70 м; глубины снижаются до 2,0 – 2,5 м; скорость течения возрастает. Сылва на этом участке имеет облик полугорного водотока (Паньков, 2004).
2.2Характеристика зообентоценозов среднего течения р. Сылвы
дрифт таксономический интенсивность динамика
Зообентос среднего течения реки Сылвы изучался сотрудниками кафедры зоологии беспозвоночных и водной экологии с 1992 г.
С самого начала исследований на р. Сылве специальное внимание уделялось проблеме установления таксономического состава. С этой целью, помимо таксономической обработки гидробиологических проб, осуществлялись фаунистические сборы, в том числе крылатых фаз амфибиотических насекомых. В составе зообентоса р. Сылвы, ее притоков и водоемов поймы было зарегистрировано свыше четырехсот видов, из них более половины оказались новыми для области, 27 были отмечены впервые для Урала (Паньков, 1997 а, б).
По данным Н.Н. Панькова (2004), в составе зообентоса р. Сылвы, ее притоков и водоемов поймы к настоящему времени выявлено 428 видов и форм. Основу таксономического разнообразия донной фауны составляют членистоногие, главным образом, амфибиотические насекомые. Наиболее разнообразно представлены двукрылые (не менее 174 видов), богаты видами ручейниками(61), поденки (37), веснянки (18) и стрекозы (15).
Из прочих беспозвоночных разнообразны моллюски (67) и малощетинковые черви (27).
В составе зообентоса Средней Сылвы зарегистрировано 312 таксонов. Наиболее богаты видами двукрылые – 132 вида, существенный вклад в разнообразие донной фауны вносят также ручейники – 43 вида, моллюски – 49 видов, поденки – 29 видов и малощетинковые черви – 20 видов (Паньков, 2004).
Фауна в экологическом отношении очень разнородна. Наряду с избегающими заиления литофильными реобионтными видами хирономид подсемейства Orthocladiinae, ручейников семейств HydropsychidaePsychomyiidae, Goeridae, поденок Heptageniidae, клопов Aphelocheiridae, веснянок Leuctridae, Capniidae, Perlodidae, моллюсков Planorbidae (Ancylus
fluviatilis
Mueller, 1774) и олигохет Naididae богато представлены элементы пелофильных факультативно – речного и равнинно – речного комплексов: хирономиды трибы Chironomini, ручейники семейства Leptoceridae, поденки семейств Caenidae, Ephemeridae, Potamanthidae, Polymitarcyidae, и Baetidae (подсемейство Cloeoninae), моллюски Bithyniidae, Valvatidae, Pisidiidae, Euglesidae, Sphaeriidae, олигохеты Tubificidae.
Несмотря на значительную протяженность среднего течения, в его пределах формируется ограниченное число зообентоценозов, не обнаруживающих каких либо заметных тенденций в изменении структуры и количественных показателей развития по продольному профилю водотока. Лишь на пограничных с соседними зонами участках присутствуют немногие элементы, в целом для Средней Сылвы не характерные. В частности, на пограничном с Нижней Сылвой Кишертско-Кунгурском участке отмечен ряд видов, приуроченных, главным образом, к нижнему течению. Таковы моллюски Cyrenastrum
solidum
(Normand, 1844), Rivicoliana
rivicola
(Lamark, 1818), Viviparus
viviparus
(Linnaeus, 1758) и амфиподы Corophium
curvispinum
Sars, 1895.
Животные, преимущественное распространение которых связано с Верхней или Нижней Сылвой, лишь в малой степени влияют на типичную для среднего течения структуру зообентоценозов, ни в коей мере (за исключением C
.
curvispinum
) не затрагивая комплекса руководящих форм.
Зообентоценозы среднего течения довольно близки по структуре. Их сходство обусловлено широким распространением в пределах Средней Сылвы большого числа видов, из которых массовыми являются личинки ручейников Hydropsyche
contubernalis
McLachlan, 1865, Cheumatopsyche
lepida
(Pictet, 1834),Psychomyia
pusilla
(Fabricius, 1781), поденокEphemerella
ignita
(Poda, 1761) иEphemera
lineata
Eaton, 1870, хирономид Microtendipes
pedellus
(DeGeer, 1776), моллюски A
.
fluviatilis
, Cyrenastrum
scaldianum
(
Normand, 1844), Pisidium
amnicum
(Mueller, 1774).
В пределах Средней Сылвы наибольшее распространение получает зообентоценоз H
.
contubernalis
, приуроченный к валунным, галечным и гравийно-галечным грунтам перекатов и медиали плесов; на его долю приходится не менее 80% площади речного ложа. Численность сообщества составляет 2,9–4,9 тыс. экз/м2
при биомассе 8,8–10,9 г/м2
. Более половины биомассы складывается за счет членов доминантного комплекса, из которых наибольшее значение имеют ручейники H
.
contubernalis
, Ch
.
lepida
и P
.
pusilla
, клопы Aphelocheirus
aestivalis
(Fabricius, 1794) моллюски A
.
fluviatilis
. Наряду с руководящими видами часто встречаются жесткокрылые семейства Elmmiidae, в отдельные годы заметную роль играют хирономидыM
.
pedellus
и Tanytarsus
gr
.
gregarius
Kieffer, 1909, ручейники Hydropsyche
pellucidula
(Curtis, 1834) и двустворчатые моллюскиHenslowiana
suecica
(ClessininWesterlund, 1873).
Около 15% площади речного русла занимает зообентоценозE
.
lineata
+
P
.
luteus
, приуроченный к песчано-гравийно-галечным умеренно заиленным грунтам рипали плесов. Его численность составляет 3,2 тыс. экз./м2
при биомассе 11,2 г/м2
. В состав лидеров сообщества, помимо указанных видов, входят ручейники H
.
contubernalis
, стрекозы Gomphus
vulgatissimus
(Linne, 1758), моллюскиP
. a
mnicum
и хирономиды M
.
pedellus
. Наряду с ними обычны ручейники Lepidostoma
hirtum
(Fabricius, 1775), моллюскиCincinna
piscinalis
(Mueller, 1774),
Lymnaea
auricularia
(Linnaeus, 1758) иBithynia
tentaculata
(Linnaeus, 1758), поденки Caenis
macrura
Stephens, 1835. В отдельные годы наблюдалось массовое развитие ручейников Goera
pilosa
(Fabricius, 1775)(1994), поденок Ephoron
virgo
(Oliver, 1791) (с 1997) и E
.
ignita
(1994), входивших здесь в число видов-доминантов.
В пределах Кишертско-Кунгурского участка среднего течения получает распространение зообентоценозC
.
curvispinum
, приуроченный к заиленным валунным грунтам рипали. Численность зообентоценоза составляет 6,94 тыс. экз/м2
, биомасса 13,3 г/м2
. До 60% биомассы формируется за счет видов доминантного комплекса, к которым, помимо корофиума, относятся моллюски Lymnaea
tumida
(Heeld, 1836),
B
.
tentaculata
,
C
.
piscinalis
,
P
. a
mnicum
и поденкиPotamanthus
luteus
(Linnaeus, 1767).
3. М
атериал и методы его обработки
Отбор проб дрифта осуществлялся на участке среднего течения р. Сылвы в районе заказника «Предуралье» в июне – июле 2005 г. и 2006 г. Пробы отбирались на одной станции, расположенной на перекате (прилож. 35).
Для улавливания дрейфующих беспозвоночных применялась ловушка – коническая горизонтальная сетка из мельничного газа №29 с площадью входного отверстия 706,5 см2
и глубиной 0,93 м.
Организмы дрифта помещались в стеклянные емкости (пенициллинки), этикетировались, фиксировались 4%-ным формалином. На этикетке проставлялся номер пробы, место и дата взятия пробы.
Для учета дрифта применялся метод дробного лова: в каждой точке подряд отбиралось две пробы (с экспозицией ловушки по 5 минут), которые затем соединялись в одну интегральную пробу.
Это связано с тем, что при более длительной экспозиции часть гидробионтов покидает ловушку, двигаясь по ее стенкам навстречу течению, а мельничный газ забивается сносимыми водой неорганическими взвесями и частицами органического происхождения: чехликами личинок амфибиотических насекомых, кусочками водных растений, фитопланктоном и т.д., что существенно снижает фильтрационные способности ловушки и приводит к вымыванию части организмов из нее (Задорина, 1987 а).
Одновременно отбиралось по две интегральные пробы дрифта – в придонном иповерхностном горизонтах. При облове поверхностного горизонта ловушка устанавливалась с таким расчетом, чтобы она захватывала и поверхностную пленку воды.
Для изучения суточной динамики бентостока пробы отбирались каждые два часа в течение суток (один суточный учет в июне 2005 г., три суточных учета в июле 2005 г., одна – в июне и три – в июле 2006 г.).
Камеральная обработка собранного материала осуществлялась в лаборатории гидробиологии на кафедре зоологии беспозвоночных и водной экологии ПермГУ. Организмы дрифта извлекались из пробы при помощи пинцета, с использованием бинокулярного микроскопа МБС-1, подсчитывались и определялись, по возможности до вида, с применением «Определителя…» (1977) и «Определителя насекомых…» (Плавильщиков, 1994).
Личинки комаров-звонцов Сhironomidae, некоторых Trichoptera, Ephemeroptera и прочих организмов определены только до группы, поскольку их основную массу в пробе составили очень молодые формы, которые не подлежат более точной идентификации.
При дальнейшей обработке подсчитанного материала была рассчитана интенсивность дрифта.
Интенсивность дрифта – это количество организмов, снесенных через единицу площади живого сечения потока за единицу времени (экз/м2
в час).
Расчет интенсивности дрифта производился по формуле:
Nd
=n/(s*t),
где Nd
– интенсивность дрифта
n – численность (экз.) организмов в пробе
s – площадь рамки (м2
)
t – время экспозиции (час).
Всего проведено 8 суточных учетов дрифта и обработано 192 количественные пробы.
Пользуясь случаем выражаю благодарность своему научному руководителю Н.Н. Панькову, А.С. Козлову за консультации по определению организмов дрифта.
4. Результаты исследований и их обсуждение
4.1 Таксономическая структура дрифта
В результате наших исследований в составе водной фракции дрифта р. Сылвы зарегистрированы 23 таксономические группы животных, в составе воздушной фракции дрифта – 13 таксономических групп животных.
Разнообразнее всего в составе водной фракции дрифта представлены амфибиотические насекомые: Ephemeroptera (16 видов), Trichoptera (12 видов), Diptera (5 семейств) (табл. 1).
В пробах встречаются кишечнополостные Coelenterata (Hydra
sp
.), малощетинковые черви Oligochaeta, Turbellaria
, пиявки Hirudinea (Erpobdella octoculata
Linne, 1758, Hellobdella stagnalis
Linne, 1758, Piscicola geometra
(Linne, 1761)), моллюски (9 видов), ракообразные Corophium
curvispinum
Sars, 1895, Ostracoda, водные клещи Hydracarina, клопы Micronecta
sp
.
и Aphelocheirus
aestivalis
(Fabricius, 1794), личинки и имаго жуков Elmidae, Dytiscidae, личинки веснянок Perlodidae(табл. 1).
В составе водной фракции дрифта наиболее богаты видами поденки Ephemeroptera: Cloeon luteolum
(Muller, 1776), Baetis fuscatus
(Linnaeus, 1761), B. inexpectatus
(Tshernova, 1982), B. muticus
(Linnaeus, 1758), B. niger
(Linnaeus, 1761), B. scambus
(Eaton, 1870), B. vernus
(Curtis, 1830), Heptagenia fuscogrisea
(Retzius, 1783), Heptagenia sulphurea
(Muller, 1776), Ecdyonurus aurantiacus
(Burmeister, 1839), Ephemera lineata
(Eaton, 1870), Potamanthus luteus
(Linnaeus, 1767), Caenis horaria
(Linnaeus, 1758), Caenis macrura
(Stephens, 1835), Habrophlebia lauta
(McLachlan, 1884), Ephemerella ignita
(Poda, 1761) (табл. 1).
Среди ручейников встречаются следующие формы: Cheumatopsyche lepida
(Pictet, 1834), Hydropsyche contubernalis
(McLachlan, 1865), Hydropsyche pellucidula
(Curtis, 1834), Agraylea sp., Hydroptila sp., Ithytrichia lamellaris
(Eaton, 1873), Orthotrichia costalis
(Curtis, 1834),Ttricholeiochiton
fagesii
(Guinard, 1879), Triaenodes
bicolor
(Curtis, 1834), Brachycentrus
subnubilus
(Curtis, 1834), Psychomyia
pusilla
(Fabricius, 1781), Polycentropus
flavomaculatus
(Pictet, 1834) (табл. 1).
Таблица 1. Таксономический состав и встречаемость организмов дрифта
| ВОДНАЯ ФРАКЦИЯ
|
| Таксон
|
V*
|
| Hydra
sp
.
|
+++
|
| Oligochaeta |
+++
|
| Turbellaria
|
+
|
| Erpobdella octoculata
Linne, 1758 |
+
|
| Hellobdella stagnalis
Linne, 1758 |
+
|
| Piscicola geometra
(Linne, 1761) |
++
|
| сем. Spheariidae indet. |
++
|
| Sphaerium corneum
(Linnaeus, 1758) |
+
|
| сем. Euglesidae indet. |
+
|
| сем. Valvatidae indet. |
+
|
| Valvata sp.
|
+
|
| Bithynia tentaculata
(Linnaeus, 1758) |
++
|
| Opisthorchophorus hispanicus
(Servain, 1880) |
+
|
| сем. Lymnaeidae indet. |
+
|
| Lymnaea sp.
|
+
|
| сем. Planorbidae indet. |
++
|
| Ancylus fluviatilis
Mueller, 1774 |
+
|
| Armiger crista
(Linnaeus, 1758) |
+
|
| Bradybaena sp.
|
+
|
| Acrolocsus sp.
|
+
|
| Corophium curvispinum
Sars, 1895 |
+++
|
| Ostracoda
sp.
|
++
|
| Hydracarina |
+++
|
| Micronecta sp.
|
+++
|
| Aphelocheirus aestivalis
(Fabricius, 1794) |
++
|
| сем. Dixidae indet. |
+
|
| сем. Elmidae indet. |
++
|
| Riolus sp.
|
+
|
| сем. Dytiscidae indet. |
+
|
| отр. Trichoptera indet. |
++
|
| сем. Hydropsychidae indet. |
++
|
| Cheumatopsyche lepida
(Pictet, 1834) |
+
|
| Hydropsyche contubernalis
McLachlan, 1865 |
++
|
| Hydropsyche pellucidula
(Curtis, 1834) |
+
|
| Сем. Hydroptilidae indet. |
+++
|
| Agraylea sp.
|
+
|
| Hydroptila sp.
|
++
|
| Ithytrichia lamellaris
Eaton, 1873 |
++
|
| Orthotrichia costalis
(Curtis, 1834) |
+
|
| Продолжение табл. 1 |
| Таксон
|
V
*
|
| Tricholeiochiton fagesii
(Guinard, 1879) |
++
|
| Сем. Leptoceridae indet. |
+++
|
| Triaenodes bicolor
(Curtis, 1834) |
+
|
| Brachycentrus subnubilus
(Curtis, 1834) |
+ |
| Psychomyia pusilla
(Fabrricius, 1781) |
+ |
| Polycentropus flavomaculatus
Pictet, 1834 |
+ |
| отр. Ephemeroptera indet. |
+
|
| сем. Baetidae indet. |
+++
|
| Cloeon luteolum
(Muller, 1776) |
+++ |
| Baetis fuscatus
(Linnaeus, 1761) |
+ |
| B. inexpectatus
(Tshernova, 1982) |
+ |
| B. muticus
(Linnaeus, 1758) |
++ |
| B. niger
(Linnaeus, 1761) |
+ |
| B. scambus
Eaton, 1870 |
+++ |
| B. vernus
Curtis, 1830 |
+ |
| Heptagenia fuscogrisea
(Retzius, 1783) |
+ |
| Heptagenia sulphurea
(Muller, 1776) |
+ |
| Ecdyonurus aurantiacus
Burmeister, 1839 |
+ |
| Ephemera lineata
Eaton, 1870 |
+ |
| Potamanthus luteus
(Linnaeus, 1767) |
+ |
| Caenis horaria
(Linnaeus, 1758) |
+ |
| Caenis macrura
Stephens, 1835 |
++ |
| Habrophlebia lauta
McLachlan, 1884 |
+
|
| Ephemerella ignita
(Poda, 1761) |
+
|
| сем. Perlodidae indet. |
+
|
| отр. Diptera indet. |
++
|
| Atherix ibis
(Fabricius, 1798),
личинки |
+
|
| сем. Сhironomidae indet. |
+++
|
| сем. Simuliidae indet. |
+++
|
| сем. Culicidae indet. |
+
|
| сем. Limoniidae indet. |
+
|
| сем. Ceratopogonidae indet. |
++
|
| ВОЗДУШНАЯ ФРАКЦИЯ
|
| отр. Araneae indet. |
+ |
| сем. Trombidiidae indet. |
+ |
| сем. Leniphiidae indet. |
+ |
| сем. Clubionidae indet. |
+ |
| H
y
r
a
cantium sp.
|
+ |
| класс Insecta indet. |
+ |
| отр. Trichoptera indet. |
++ |
| Hydropsyche sp.
|
+ |
| Cheumatopsyche lepida
(Pictet, 1834) |
+ |
| сем. Hydroptilidae indet. |
++ |
| Hydroptila sp.
|
+ |
| Ithytrichia lamellaris
(Eaton, 1873) |
+ |
| сем. Leptoceridae indet. |
+ |
| Окончание табл. 1 |
| Таксон
|
V
*
|
| Psychomyia pusilla
(Fabricius, 1781) |
+ |
| отр. Ephemeroptera indet. |
+ |
| Сем. Baetidae indet. |
+++ |
| Baetis sp.
|
++ |
| B. vernus
(Curtis, 1830) |
+ |
| Caenis horaria
(Linnaeus, 1758) |
+ |
| Caenis macrura
Stephens, 1835 |
+ |
| Collembola indet. |
+++ |
| сем. Sminturidae indet. |
+ |
| Podura sp.
|
+ |
| отр. Psocoptera indet. |
+ |
| Tardigrada sp.
|
+ |
| отр. Homoptera indet. |
++ |
| отр. Coleoptera indet. |
+ |
| сем. Staphilinidae indet. |
+ |
| сем. Curculionidae indet. |
+ |
| сем. Ptilidae indet. |
+ |
| сем. Elmidae indet. |
++ |
| сем. Lumiidae indet. |
+ |
| отр. Neuroptera indet. |
+ |
| отр. Lepidoptera indet., личинки |
++ |
| отр. Hymenoptera indet. |
+++ |
| сем. Formacidae indet. |
+ |
| отр. Thysanoptera indet. |
+++ |
| отр. Diptera indet. |
+++ |
| сем. Tipulidae indet. |
+ |
| сем. Limoniidae indet. |
+ |
| сем. Сhironomidae indet. |
+++ |
| сем. Simuliidae indet. |
+++ |
*V – встречаемость таксона: +++ – массовый, ++ – обычный, + – редкий.
Среди дрейфующих организмов наиболее многочисленны кишечнополостные Hydra
sp
.
, малощетинковые черви Oligochaeta, ракообразные Corophium curvispinum
Sars, 1895, клопы Micronecta
sp
., водные клещи Hydracarina, личинки ручейников (Hydroptilidae, Leptoceridae), личинки поденок Ephemeroptera (Baetidae), личинки и куколки комаров-звонцов Сhironomidae, Simuliidae (рис. 1).
Значительно реже в пробы попадают личинки и имаго жуков Elmidae, личинки Ceratopogonidae, личинки мух Atherix ibis
(Fabricius, 1798), веснянок Perlodidae, ручейников Brachycentridae, Psychomyiidae, Polycentropodidae, поденок Baetis fuscatus
(Linnaeus, 1761), B. inexpectatus
(Tshernova, 1982), B. vernus
Curtis, 1830, Heptagenia fuscogrisea
(Retzius, 1783), Ecdyonurus aurantiacus
(Burmeister, 1839), Ephemera lineata
Eaton, 1870, Potamanthus luteus
(Linnaeus, 1767), Caenis horaria
(Linnaeus, 1758), Habrophlebia lauta
(McLachlan, 1884) и Ephemerella ignita
(Poda, 1761), пиявки и моллюски.
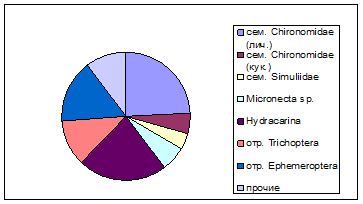
Рис. 1. Таксономическая структураорганизмов водной фракции дрифта
Таким образом, можно сказать, что структура дрифта не является точным отражением структуры зообентоса, возможно, это связано с различиями в миграционной активности животных, а так же с их разной способностью удержаться на грунте в условиях сильного течения.
Здесь же следует рассмотреть и воздушную фракцию в составе дрифта, или аллохтонный компонент дрифта (Чебанова, 1992). К данной группе относятся животные суши, случайно оказавшиеся в воде.
Постоянным компонентом в пробах являются имагинальные стадии Diptera, Hymenoptera, представители Thysanoptera (рис. 2).
Реже, в основном, в вечернее время, в пробах обнаруживаются взрослые представители амфибиотических насекомых Trichoptera и Ephemeroptera. Из ручейников чаще всего встречаются имаго Hydroptilidae, из поденок – Baetidae (Baetis sp.
). Единичными экземплярами представлены имаго Caenis macrura
(Stephens, 1835), Caenis horaria
(Linnaeus, 1758), Psychomyia pusilla
(Fabricius, 1781), Ithytrichia lamellaris
(Eaton, 1873), Cheumatopsyche lepida
(Pictet, 1834).
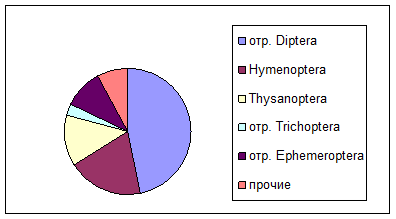
Рис. 2. Таксономическая структураорганизмов воздушной фракции дрифта
Малую долю сносимых в воду животных составляют личинки Lepidoptera, имаго Neuroptera, Coleoptera, Homoptera, Collembola, Araneae. Вероятно, данные группы животных сносятся ветром, либо смываются с берегов во время дождей.
4.2 Вертикальное распределение организмов дрифта
Как видно из таблицы 2, вертикальное распределение организмов дрифта не вполне однородно.
Таблица 2. Вертикальная структура дрифта
| Таксон
|
Поверхностный
горизонт
|
Придонный
горизонт
|
| днем
|
ночью
|
днем
|
ночью
|
| Hydra
sp.
|
6,7 |
6,7 |
2,1 |
1,5 |
| Oligochaeta indet. |
4,8 |
5,0 |
4,4 |
4,7 |
| Erpobdella octoculata
Linne, 1758 |
0,0 |
0,1 |
0,1 |
0,0 |
| Hellobdella stagnalis
Linne, 1758 |
0,0 |
0,3 |
0,0 |
0,0 |
| Piscicola geometra
(Linne, 1761) |
0,1 |
0,3 |
0,0 |
0,2 |
| сем.
Spheariidae indet. |
0,1 |
0,3 |
0,0 |
0,1 |
| Sphaerium corneum
(Linnaeus, 1758) |
0,1 |
0,0 |
0,0 |
0,0 |
| Sphaerium nitidum
|
0,2 |
0,1 |
0,0 |
0,0 |
| Valvata sp.
|
0,0 |
0,0 |
0,0 |
0,11 |
| Bithynia tentaculata
(Linnaeus, 1758) |
0,3 |
0,5 |
0,0 |
0,0 |
| сем.
Lymnaeidae indet. |
0,0 |
0,1 |
0,0 |
0,0 |
| Lymnaea sp.
|
0,1 |
0,2 |
0,1 |
0,0 |
| сем. Planorbidae indet. |
0,0 |
0,1 |
0,3 |
0,6 |
| Corophium curvispinum
Sars, 1895 |
7,0 |
8,4 |
0,6 |
1,2 |
| Ostracoda
|
1,8 |
1,0 |
1,0 |
1,2 |
| Hydracarina |
51,0 |
95,3 |
15,0 |
23,3 |
| Micronecta sp.
|
20,3 |
24,7 |
1,0 |
20,9 |
| Aphelocheirus a
estivalis
(Fabricius,1794) |
0,0 |
0,1 |
0,0 |
0,3 |
| сем. Elmidae indet. |
1,1 |
1,1 |
1,3 |
1,6 |
| Отр. Trichoptera indet. |
4,4 |
17,3 |
2,6 |
13,0 |
| Сем. Hydropsychidae indet. |
0,5 |
1,5 |
0,0 |
1,8 |
| Cheumatopsyche lepida
(Pictet, 1834) |
0,0 |
0,1 |
0,0 |
0,3 |
| Hydropsyche contubernalis
(McLachlan, 1865) |
0,5 |
3,0 |
0,3 |
1,3 |
| Сем. Hydroptilidae indet. |
1,4 |
2,7 |
0,2 |
0,9 |
| Hydroptila sp.
|
0,5 |
1,0 |
0,1 |
0,7 |
| Ithytrichia lamellaris
(Eaton, 1873) |
0,4 |
1,7 |
0,2 |
1,4 |
| Orthotrichia costalis
(Curtis, 1834) |
0,0 |
0,2 |
0,0 |
0,1 |
| Tricholeiochiton fagesii
(Guinard, 1879) |
0,3 |
2,3 |
0,2 |
1,5 |
| Сем. Leptoceridae indet. |
0,8 |
21,2 |
0,2 |
9,0 |
| Brachycentrus subnubilus
(Curtis, 1834) |
0,1 |
0,2 |
0,0 |
0,0 |
| Psychomyia pusilla
(Fabrricius, 1781) |
0,1 |
0,1 |
0,0 |
0,2 |
| Отр. Ephemeroptera indet. |
0,1 |
0,1 |
0,0 |
0,0 |
| Сем. Baetidae indet. |
1,3 |
16,9 |
0,9 |
8,1 |
| Cloeon luteolum
(Muller, 1776) |
0,3 |
51,4 |
0,5 |
17,7 |
| Baetis fuscatus
(Linnaeus, 1761) |
0,0 |
0,1 |
0,0 |
0,0 |
| B. inexpectatus
(Tshernova, 1982) |
0,0 |
0,3 |
0,0 |
0,2 |
| B. muticus
(Linnaeus, 1758) |
0,2 |
0,6 |
0,1 |
0,4 |
| B. niger
(Linnaeus, 1761) |
0,0 |
0,3 |
0,0 |
0,0 |
| B. scambus
(Eaton, 1870) |
0,5 |
15,5 |
0,2 |
1,4 |
| B. vernus
(Curtis, 1830) |
0,1 |
0,1 |
0,0 |
0,3 |
| Baetis sp.
|
1,3 |
1,0 |
0,8 |
0,4 |
| Heptagenia sulphurea
(Muller, 1776) |
0,1 |
0,7 |
0,1 |
0,4 |
| Potamanthus luteus
(Linnaeus, 1767) |
0,0 |
0,2 |
0,2 |
0,2 |
| Caenis macrura
(Stephens, 1835) |
0,2 |
1,7 |
0,3 |
2,5 |
| Ephemerella ignita
(Poda, 1761) |
0,0 |
0,3 |
0,1 |
0,0 |
| сем. Perlodidae indet. |
0,1 |
0,0 |
0,0 |
0,1 |
| отр. Diptera indet., куколки |
0,0 |
0,3 |
0,0 |
0,1 |
| Atherix ibis,
личинки |
0,1 |
0,1 |
0,0 |
0,01 |
| сем. Сhironomidae, личинки |
37,3 |
88,4 |
21,2 |
50,71 |
| сем. Сhironomidae, куколки |
14,7 |
20,7 |
2,4 |
4,6 |
| сем. Simuliidae, личинки |
3,3 |
15,7 |
2,3 |
10,4 |
| сем. Simuliidae, куколки |
0,4 |
0,1 |
0,2 |
0,0 |
| сем. Culicidae |
1,1 |
1,1 |
0,0 |
0,0 |
| сем. Ceratopogonidae |
0,0 |
2,2 |
0,1 |
0,0 |
| Прочие |
0,2 |
9,3 |
0,5 |
14,4 |
| ВСЕГО
|
163,7
|
422,2
|
59,8
|
197,6
|
Большинство организмов сносятся в поверхностном горизонте воды (табл. 2), что еще раз доказывает тот факт, что дрифт не является полностью пассивным процессом (Леванидов, Леванидова, 1981).
Более высокая плотность животных в верхних слоях воды объясняется их целенаправленными миграциями, которые могут быть спровоцированы столкновениями конкурирующих особей. Это может быть связано с вылетом ряда групп амфибиотических насекомых, которому предшествует активный предимагинальный дрифт (Леванидов, Леванидова, 1981).
Пассивное всплытие организмов происходит вместе с фрагментами перифитонных матов (сем. Naididae) или увлечением их с турбулентными токами воды (личинки и взрослые жуки Elmidae) (Паньков, 2007).
Литературные данные о вертикальном распределении организмов дрифта довольно противоречивы. Так в мощном предгорном потоке (р. Хор) плотность всплывающих организмов в верхнем горизонте была в 4 – 5 раз выше, чем в придонном (Леванидов, Леванидова, 1981).
В небольшой горной реке Щугор, напротив, наибольшая численность дрейфующих организмов приурочена к придонному горизонту (Шубина, 1986). Мигрирующие беспозвоночные преобладают в придонном горизонте и в р. Волге (Аристовская, 1945).
Таким образом, вертикальное распределение организмов дрифта еще не достаточно изучено и нуждается в дальнейшем исследовании.
4.3 Суточная динамика интенсивности дрифта
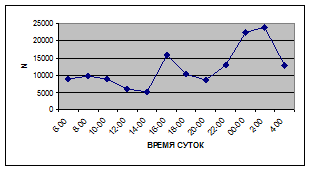
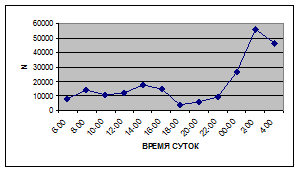
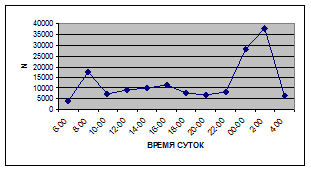
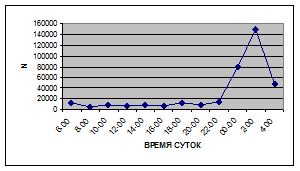
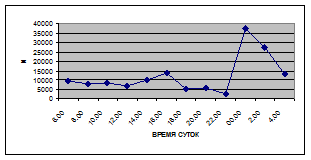
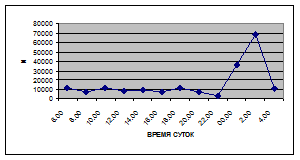
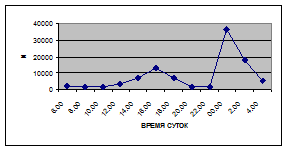
Суточная динамика интенсивности дрифта донных беспозвоночных р. Сылвы описывается кривой довольно сложной конфигурации, на которой могут быть выделены максимумы и минимумы первого и второго порядков (рис. 3).
Рассмотренная кривая на уровне элементов первого порядка имеет одновершинную форму с наивысшими показателями с двенадцати до двух часов ночи и низкими показателями в дневные часы.
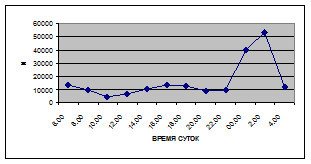 
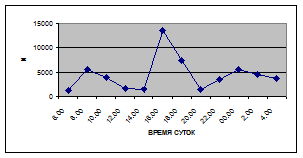
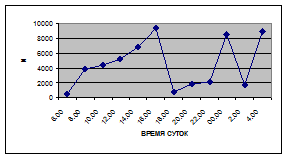
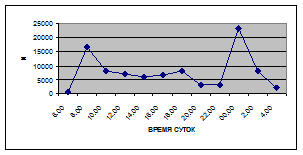
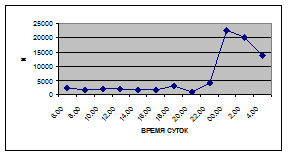
а) 29 июня 2005 г. д) 19 июня 2006 г.
б) 06 июля 2005 г. е) 11 июля 2006 г.
в) 16 июля 2005 г. ж) 18 июля 2006 г.
г) 18 июля 2005 г. з) 27 июля 2006 г.
Рис. 3. Суточная динамика интенсивности дрифта (N, экз/ м2
в час, водная фракция)
Важной характеристикой дрифта является амплитуда колебаний его параметров в течение суток. В р. Сылве максимальная интенсивность дрифта (Nmax) превосходит минимальную (Nmin) в 4,5 – 23,4 раз.
Минимальные показатели интенсивности дрифта приурочены к дневному времени суток, в основном, с четырех часов утра до восьми часов вечера. Они изменяются в пределах от 7,2 до 21,4 тыс. экз/ м² в час.
В этот период мигрируют животные с разной склонностью к дрифту. Это могут быть и животные, сравнительно легко расстающиеся с бенталью и животные, в целом не расположенные к вертикальным миграциям.
К ним относятся водные клещи Hydracarina и клопы Micronecta sp.
, ракообразные Corophium curvispinum
Sars, 1895 и Ostracoda
sp
.,
личинки амфибиотических насекомых Trichoptera, Ephemeroptera, Diptera и другие (прилож. 1, 5, 9, 13, 17, 21, 25, 29).
Далее, с десяти до двенадцати часов ночи, наблюдается увеличение интенсивности дрифта до 40,5 тыс. экз/ м² в час. Это происходит за счет увеличения миграционной активности амфибиотических насекомых: личинок и куколок Сhironomidae, Leptoceridae, Baetidae (Cloeon luteolum
(Muller, 1776)).
К двум часам ночи интенсивность дрифта возрастает до максимальных значений в суточном цикле, в среднем до 63,8 тыс. экз/ м² в час. Эти показатели обусловлены подъемом в толщу воды массовых групп животных. К ним относятся личинки и куколки Сhironomidae, Trichoptera, Cloeon luteolum
, Baetis
scambus
и другие таксоны (прилож. 1, 5, 9, 13, 17, 21, 25, 29).
Приуроченность наивысших показателей интенсивности дрифта к первой половине ночи отмечена и в литературе (Леванидов, Леванидова, 1981; Богатов, 1983; Шубина, 1986; Задорина, 1987б
).
Именно в эти часы дрифт достигает не только максимальной интенсивности, но и максимального разнообразия – в пробах отмечены все зарегистрированные таксономические группы (прилож. 1, 5, 9, 13, 17, 21, 25, 29).
Пик интенсивности дрифта обусловлен возрастанием миграционной активности всех участвующих в дрифте групп. Наибольшая интенсивность дрифта в эти часы зарегистрирована у личинок Ephemeroptera – 20,1 тыс. экз/ м², Trichoptera – 9,8 тыс. экз/ м² и у личинок Сhironomidae– 15,1 тыс. экз/ м² в час (рис. 4).
Иногда максимальная интенсивность дрифта в течение суток достигает показателей в 148,7 тыс. экз/ м² в час (18 июля 2006 г.), а в другие дни она может быть ниже в несколько раз (6 июля 2005 г. – 23,8 тыс. экз/ м² в час).
Вероятно, различия показателей интенсивности дрифта напрямую связаны с погодными и гидрологическими условиями. Так, 6 июля 2005 г. температура воздуха была +13…+150
С, а 18 июля 2006 г. – +34…+360
С, при этом уровень воды в начале июля 2005 г. был выше уровня воды в этот же период 2006 г.

Рис. 4. Суточная динамика интенсивности массовых таксонов водной фракции дрифта (N, экз/ м² в час)
С наступлением утренних сумерек интенсивность дрифта резко уменьшается и достигает минимальных показателей – 6,5 тыс. экз/ м² в час. Возможно, к этому времени большинство организмов удовлетворили свои потребности в расселении.
На фоне дневного минимума можно наблюдать пики второго порядка. Такие пики были отмечены 29 июня 2005 г. в шесть часов утра и в четыре часа дня, 6 июля 2005 г. – в восемь и четыре часа, 16 июля 2005 г. – в восемь и четыре часа, 18 июля 2005 г. – в четыре часа дня и 11 июля 2006 г. – в восемь и четыре часа. 19 июня, 18 и 27 июля 2006 г. пиков второго порядка вообще не наблюдалось (рис. 3).
Большую долю в максимальных пиках второго порядка составляют водные клещи Hydracarina, клопы Micronecta
sp
.,
ракообразные Corophium
curvispinum
Sars, 1895, личинки и куколки Сhironomidae.
Миграционная активность бентосных животных находится в противофазе с пищевой активностью хариуса – одного из наиболее многочисленных видов рыб на Сылве (Паньков, 2002). Низкие показатели дрифта утром можно объяснить наибольшей активностью хариуса в эти часы.
В целом показатели интенсивности дрифта р. Сылвы имеют довольно высокие значения и сильно отличаются от литературных данных по разным водотокам. Так для р. Лувеньги (Кольский полуостров) интенсивность дневного дрифта изменялась от 45 до 125 экз/ м² в час в конце июня – начале июля (Задорина, 1987б
); для р. Лижмы (бассейн Онежского озера) интенсивность дрифта составила 554–1545 экз/ м² в час (Шустов, Широков, 1980); в р. Порье (бассейн Белого моря) интенсивность дрифта в июле составила 665–1194 экз/ м² в час (Задорина, 1987а
).
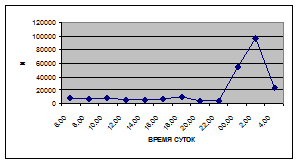
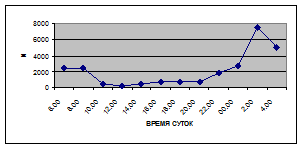
Суточная динамика интенсивности воздушной фракции дрифта не обнаруживает четкой закономерности (рис. 5).
На кривых, описывающих суточный ритм интенсивности воздушной фракции дрифта выделяется от одного до трех пиков (от 1,5 до 17,6 тыс. экз/ м² в час), которые приходятся как на светлое, так и на темное время суток.
Вероятно, это объясняется тем, что в воздушной фракции дрифта присутствуют организмы с разной биологией и с различной суточной активностью.
а) 29 июня 2005 г. д) 19 июня 2006 г.
б) 06 июля 2005 г. е) 11 июля 2006 г.
в) 16 июля 2005 г. ж) 18 июля 2006 г.
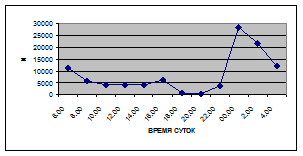 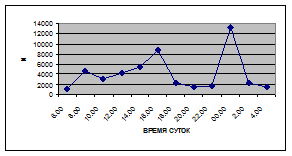
г) 18 июля 2005 г. з) 27 июля 2006 г.
Рис. 5. Суточная динамика интенсивности дрифта (N, экз/ м² в час, воздушная фракция)
Тем не менее, имеются некоторые общие черты суточной динамики водной и воздушной фракции дрифта.
В большинстве случаев максимальные пики интенсивности воздушной фракции дрифта приходятся на ночное время суток. Исключение составляют показатели интенсивности дрифта 6 июля 2005 г. и 11 июля 2006 г. Здесь максимальные пики интенсивности дрифта отмечены в дневное время. Это связано с увеличением суточной активности взрослых особей Сhironomidae, Hymenoptera и Thysanoptera.
Минимальные показатели интенсивности дрифта воздушной фракции приурочены к дневному времени суток и изменяются в пределах от 1,5 до 7,5 тыс. экз/ м² в час.
Днем самая высокая интенсивность дрифта воздушной фракции отмечена 6 июля 2005 г. (13,5 тыс. экз/ м² в час) в четыре часа, 16 июля 2005 г. в восемь часов утра (16,6 тыс. экз/ м² в час) и 19 июня 2006 г. (13,2 тыс. экз/ м² в час) в четыре часа за счет суточной активности имаго Diptera и Thysanoptera (рис. 5, 6; прилож. 6, 10, 18).

Рис. 6. Суточная динамика интенсивности массовых таксонов воздушной фракции дрифта (
N, экз/ м² в час)
Максимальные показатели интенсивности дрифта воздушной фракции приходятся на двенадцать часов ночи и изменяются в пределах 2,7 – 36,5 тыс. экз/ м² в час.
Самая высокая интенсивность дрифта воздушной фракции была отмечена 19 июня 2006 г. (36,5 тыс. экз/ м² в час) в двенадцать часов ночи, когда наблюдался вылет поденок Baetis
sp
.
и двукрылых (рис. 6; прилож. 18).
В
ыводы
1. В составе водной фракции дрифта беспозвоночных р. Сылвы летом 2005–2006 гг. зарегистрировано 23 таксономические групп, в воздушной фракции – 13.
В водной фракции дрифта наиболее разнообразно представлены амфибиотические насекомые: Ephemeroptera (16 видов), Trichoptera (12 видов), Diptera (5 семейств).
Достаточно богаты видами моллюски (Sphaerium corneum
(Linnaeus, 1758), Valvata sp.
, Bithynia tentaculata
(Linnaeus, 1758),
Opisthorchophorus hispanicus
(Servain, 1880), Lymnaea sp.
, Ancylus fluviatilis
Mueller, 1774, Armiger crista
(Linnaeus, 1758), Bradybaena sp.
, Acrolocsus sp.
), пиявки (Erpobdella octoculata
Linne, 1758, Hellobdella stagnalis
Linne, 1758, Piscicola geometra
(Linne, 1761)).
По численности в водной фракции дрифта преобладают личинки и куколки комаров-звонцов Сhironomidae, личинки мошек Simuliidae, личинки поденок Baetis
scambus
(Eaton, 1870), Cloeon
luteolum
(Muller, 1776), ручейников Leptoceridae, Hydroptilidae, водные клещи Hydracarina, клопы Micronecta
sp
.
Существенную роль в составе дрифта играют личинки ручейников Hydroptilidae, Leptoceridae и поденок Baetidae, ракообразные C
.
curvispinum
Sars, 1895, клопы Micronecta
sp
.
В составе воздушной фракции дрифта обычным компонентом являются имагинальные стадии Diptera (Сhironomidae, Simuliidae), Hymenoptera и Thysanoptera. В вечернее время можно наблюдать взрослых представителей амфибиотических насекомых Trichoptera (Hydroptilidae) и Ephemeroptera (Baetidae).
Малую долю сносимых в воду животных составляют личинки Lepidoptera, имаго Neuroptera, Coleoptera, Homoptera, Collembola, Araneae.
2. Вертикальное распределение организмов неоднородно: плотность дрейфующих организмов в поверхностном горизонте в 2,0 – 2,7 раза выше, чем в придонном. Это объясняется активным подъемом организмов в толщу воды.
3. Суточная динамика интенсивности дрифта описывается кривой сложной конфигурации, на которой выделяются максимумы и минимумы первого и второго порядков.
Наивысшие показатели интенсивности водной фракции дрифта (23,8 – 148,7 тыс. экз/ м² в час) приурочены к самому темному времени суток (2 часа), показатели интенсивности воздушной фракции дрифта (2,7 – 36,5 тыс. экз/ м² в час) – к двенадцати часам ночи. Минимальные показатели интенсивности водной фракции дрифта (3,9 – 18,0 тыс. экз/ м² в час) и воздушной фракции дрифта (0,3 – 1,7 тыс. экз/ м² в час), как правило, наблюдаются в утренние и дневные часы.
В определении суточной динамики водной фракции дрифта наибольшую роль играют личинки Сhironomidae, личинки поденок Ephemeroptera, Trichoptera, водные клещи Hydracarina, воздушной фракции дрифта – имагинальные стадии Сhironomidae, Hymenoptera и Thysanoptera.
Литература
1. Алексевнина М.С., Буйдов А.С. Фауна хирономид (Diptera, Chironomidae) р. Сылвы (бассейн Камы) // Вестн. Перм. ун-та, 1997. С. 148 – 152.
2. Аристовская Г.В. О значении сноса донных организмов р. Волги // Труды Общества естествоиспытателей при Казанском ун-те. Казань, 1945. Т.57. Вып. 1 – 2. С. 3 – 10.
3. Богатов В.В. Донные беспозвоночные и их дрифт в некоторых реках Дальнего Востока. Автореф. дис… канд. биол. наук. Ленинград, 1983. – 18 с.
4. Богатов В.В. Классификация дрифта речного бентоса // Гидробиол. журн., 1988. Т. 24. №1. С. 29 – 33.
5. Богатов В.В. Некоторые особенности динамики бентостока в условиях дождевого паводка // Систематика и экология речных организмов. Владивосток: ДВО АН СССР, 1989. С. 112 – 119.
6. Воронов Г.А., Циберкин Н.Г. и др. Учебно-научная база «Предуралье» Пермского университета // Вестн. Перм. ун-та, Вып. 3 (Заказник «Предуралье»). Пермь, 2000 С. 20 – 56.
7. Громов В.В. Донная фауна нижнего течения р. Сылвы // Изв. ЕНИ при Перм. ун-те, 1959. Т. 14. Вып. 3. С. 67 – 83.
8. Задорина В.М. Выбор экспозиции ловушки при сборе проб дрифта // Гидробиол. журн., 1987 а
. Т. 23. №2. С. 79 – 83.
9. Задорина В.М. Суточная динамика дрифта водных беспозвоночных в реках Кольского полуострова // Биология внутренних вод: инфор. бюлл. Л. Наука, 1987 б
. №73. С. 22 – 25.
10. Ключарева О.И. О скате и суточных вертикальных миграциях донных беспозвоночных Амура // Зоол. журн., 1963. Т. 42. Вып. 11. С. 1601–1612.
11. Комлев А.М., Черных Е.А. Реки Пермской области. Пермь, 1984. – 58 с.
12. Константинов А.С. Биология хирономид и их разведение // Труды Саратовского отделения Всесоюз. НИИ озерного и рыбного хозяйства, 1958. Т. 5. С. 1 – 362.
13. Константинов А.С. Сиртон и бентосток Волги близ Саратова в 1966 г. // Зоол. журн., 1969. Вып. 48. №1. С. 20 – 29.
14. Коробков И.М. Малые реки Пермской области и использование их в народном хозяйстве. Пермское книжное издательство, 1959. – 58 с.
15. Леванидов В.Я., Леванидова И.М. Дрифт личинок насекомых в крупной предгорной реке на примере реки Хор (бассейн Уссури) // Беспозвоночные животные в экосистемах Дальнего Востока. Владивосток: ДВНЦ АН СССР, 1981.С. 22 – 37.
16. Леванидова И.М. Бентос притоков Амура (эколого-фаунистический обзор) // Изв. Тихоокеанского НИИ мор. рыб. хозяйства и океанографии. Магадан, 1968. Т. 64. С. 181 – 289.
17. Леванидова И.М., Леванидов В.Я. Суточные миграции донных личинок поденок в реке Хор // Зоол. журн., 1965. Т. 44. Вып. 3. С. 373 – 389.
18. Лепнева С.Г. Ручейники. М., Л., 1964. – 562 с. (Фауна СССР; Новая Серия; №88. Т 2. Вып. 1).
19. Наумкин Д.В., Севастьянов В.М., Лавров И.А. Кунгурский заповедный край. – Пермь: ООО «Раритет – Пермь», 2004. – 120 с.
20. Определитель пресноводных беспозвоночных Европейской части СССР, Л., 1977. – 512 с.
21. Паньков Н.Н. Структурные и функциональные характеристики сообществ зообентоса реки Сылвы (бассейн Камы) / Дис. … канд. биол. наук. Пермь, 1997 а. – 230 с.
22. Паньков Н.Н. К познанию биологического разнообразия водоемов Западного Урала. Фауна поденок (Insecta: Ephemeroptera) р. Сылвы и их роль в донных биоценозах // Проблемы биологического разнообразия водных организмов Поволжья. Тольятти, 1997 б. С. 134 – 139.
23. Паньков Н.Н. Дрифт донных беспозвоночных в предгорной реке Среднего Урала (на примере реки Сылвы, заказник «Предуралье») // Биология внутренних вод: проблемы экологии и биоразнообразия. Тезисы докл. 12 Междунар. конф. молодых ученых (Борок, 23–26 сентября 2002 года). Борок, 2002. С. 86 – 87.
24. Паньков Н.Н. Структурные и функциональные характеристики зообентоценозов р. Сылвы (бассейн Камы). Пермь, 2004. – 161 с.
25. Паньков Н.Н. Основные итоги изучения дрифта реки Сылвы (заказник «Предуралье», 1997–2004 гг.) // Вестн. Перм. ун-та. 2007. Биология. В печати.
26. Плавильщиков Н.Н. Определитель насекомых: краткий определитель наиболее распространенных насекомых европейской части России. М.: Топикал. 1994. – 544 с.
27. Проблемы физической географии Урала. М., 1966. – 295 с.
28. Ресурсы поверхностных вод СССР. Т. 11 Средний Урал и Приуралье. Вып. 1. Кама. Л., 1967. – 536 с.
29. Справочник по водным ресурсам СССр. Т. ХII «Урал и Приуралье». М., Л., 1936. – 664 с.
30. Чебанова В.В. Динамика дрифта беспозвоночных в лососевых реках разного типа (юго-восток Камчатки) // Гидробиол. журн., 1992. Т. 28. №4. С 31 – 39.
31. Шкляев А.С., Балков В.А. Климат Пермской области. Пермь, 1963. – 191 с.
32. Шубина В.Н. Гидробиология лососевой реки Северного Урала. Л.: Наука, 1986. – 158 с.
33. Шустов Ю.А. Дрифт донных беспозвоночных в лососевых реках бассейна Онежского озера // Гидробиол. журн., 1977. Т. 13. Вып. 3. С. 32 – 37.
34. Шустов Ю.А. Дрифт беспозвоночных в притоках Онежского озера // Лососевые нерестовые реки Онежского озера. Л., 1978. С 50 – 53.
35. Шустов Ю.А., Широков В.А. Методика изучения дрифта беспозвоночных в реках // Гидробиол. журн., 1980. Т. 16. №3. С. 100 – 102.
|
