Мариинский Лесотехнический техникум
РЕФЕРАТ
по биологии
Генетика человека
студентка группы ОКД-11
Дочевская Анастасия;
преподаватель химии и биологии:
Мартынова Л. П.
09.03.2011
Цель работы
1. Узнать закономерности наследственности.
2. Раскрыть закономерности изменчивости.
3. Разобраться в генных мутациях.
Содержание:
1.Закономерности наследственности
2.Закономерности изменчивости
a) Типы изменчивости
b) Факторы приводящие к мутациям
c) Мутагены
d) Типы мутаций
3.Генные мутации
a) Наследственные болезни
b) Статистика рождения детей с наследственными болезнями
Литература
1.
Закономерности наследственности
наследственность мутация генная болезнь
Насле́дственность
— способность организмов передавать свои признаки и особенности развития потомству. Благодаря этой способности все живые существа (растения, грибы, или бактерии) сохраняют в своих потомках характерные черты вида. Такая преемственность наследственных свойств обеспечивается передачей их генетической информации. Носителями наследственной информации у организмов являются гены.
Методы изучения наследственности человека
· Генеалогический метод
— составление родословного дерева многих поколений и изучение типа наследования (доминантный или рецессивный, сцепленный с полом или аутосомный), частоты и интенсивности проявления наследственных свойств. Результатом изучения обычно является определение типа наследования, а также риска проявления наследственных нарушений у потомков;
· Цитогенетический метод
— изучение хромосомных наборов здоровых и больных людей. Результат изучения — определение количества, формы, строения хромосом, особенности хромосомных наборов обоих полов, а также хромосомных нарушений;
· Биохимический метод
— изучение изменений в биологических параметрах организма, связанных с изменением генотипа. Результат изучения — определение нарушений в составе крови, в околоплодной жидкости и т. д.;
· Близнецовый метод
— изучение генотипических и фенотипических особенностей однояйцевых и разнояйцевых близнецов. Результат изучения — определение относительного значения наследственности и окружающей среды в формировании и развитии человеческого организма;
· Популяционный метод
— изучение частоты встречаемости аллелей и хромосомных нарушений в популяциях человека. Результат изучения — определение распространения мутаций и естественного отбора в популяциях человека.
Реклама

- Моногибридное скрещивание
М
оногибридным
называется скрещивание, при котором родители различаются по проявлению лишь одного из признаков. В одном из опытов Г. Мендель в качестве родителей выбрал особей чистых линий (то есть особей, которые при скрещивании друг с другом на протяжении многих поколений давали потомство с набором тех же самых признаков). Он исследовал наследование окраски семян гороха — она может быть желтой или зеленой. Г. Мендель ставил опыт таким образом, что в одном эксперименте материнские растения имели желтые семена, а отцовские — зеленые, а в другом — наоборот. Такая система из двух скрещиваний носит название реципрокного
скрещивания. При этом одно из скрещиваний (любое) называется ПРЯМЫМ, а другое — ОБРАТНЫМ. (в данном случае результаты прямого и обратного скрещивания были одинаковыми.) Из гибридов первого поколения Г. Мендель путем самоопыления получал гибриды второго поколения и т. д. В нашем случае схема скрещивания будет выглядеть так.
 зеленые семена х зеленые семена х 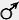 желтые семена желтые семена
| F1 |
4/4 желтые |
| F2 |
1/4 зеленые |
2/4 желтые |
1/4 желтые |
| F3 |
зеленые |
1/4 зеленые |
3/4 желтые |
желтые |
| F4 |
зеленые |
зеленые |
зеленые желтые |
желтые |
Из схемы видно, что у всех особей F1 проявился признак только одного родителя, а именно — желтая окраска семян. Проявляющийся в первом поколении гибридов признак Г. Мендель назвал доминантным
(а само явление — доминированием), а исчезающий — редессивным
. Описанная закономерность известна под названием закона-(или правила) единообразия первого поколения. Иногда ее также называют первым законом Менделя, что не совсем верно. Сам ученый формулировал лишь «закон комбинации различающихся признаков», включающий в себя, по сути, правило расщепления и правило независимого наследования. Кроме того, важно заметить, что правило единообразия гибридов первого поколения отражает не закономерности наследования признаков, а особенности их реализации в организме. При размножении гибридов F1 во втором поколении, наряду с доминантным, у части особей проявился отсутствовавший в фенотипе гибридов" первого поколения рецессивный признак. Г. Мендель обнаружил, что особей с доминантным признаком примерно втрое больше, чем с рецессивным (то есть произошло расщепление в соотношении 3 : 1). Эти результаты легли в основу закона расщепления. Дальнейшее размножение гибридов F2 показало, что особи с рецессивным признаком давали в ряду поколений только особей, у которых- также проявлялся лишь рецессивный признак; а группа с доминантным признаком оказалась разнородной. Одна ее часть в ряду поколений давала только особей с проявлением доминантного признака, а другая при размножении расщеплялась в соотношении по фенотипу 3:1. Рассматриваемые организмы диплоидны, то есть состоят из клеток, содержащих двойной набор хромосом. Гомологичные хромосомы имеют идентичные участки — гены, в которых содержится информация о том или ином признаке, например, цвете семян. Однако признак этот может проявляться в фенотипе различным образом — семена могут быть зелеными, а могут быть и желтыми. Собственно цвет (желтый или зеленый) определяется тем или иным состоянием гена (последовательностью нуклеотидов в цепи ДНК). В рассматриваемом случае ген окраски семян имеет две альтернативные формы (аллеля). Аллели
— формы (их может быть не только две, но и больше — явление множественного аллелизма) одного и того же гена, располагающиеся в одинаковых участках (локусах) гомологичных хромосом. Таким образом, соматические клетки содержат два аллеля одного гена. При этом, несмотря на то, что аллели могут быть разными (гетерозиготное состояние), в фенотипе проявляется только один из них — он называется доминантным. Рецессивный же аллель влияет на фенотип только в том случае, если он, находится в обоих гомологичных хромосомах (гомозиготное состояние). Образующиеся в результате мейоза гаплоидные гаметы содержат всего лишь один аллель того или иного гена. На схеме доминантные аллели обозначаются латинской заглавной буквой, а рецессивные — прописной (буква при этом используется одна и та же, что подчеркивает, что оба аллеля ответственны за проявление одного и того же признака). Схема нашего скрещивания с учетом сказанного будет выглядеть так:
Реклама

| Р
|
зеленые семена |
X |
желтые семена |
| фенотип: генотип: гаметы: |
аа a, a |
AA A, A |
| F1 фенотип: генотип: гаметы: |
желтые семена Aa A, a |
| F2 фенотип: генотип: |
3 желтые AA Aa Aa |
:
|
1 зеленые aa |
В скрещивании участвуют и особи чистых линий. Это означает, что они гомозиготны по выбранному признаку. При оплодотворении материнская и отцовская гаметы сливаются. Поскольку доминантный аллель подавляет работу рецессивного, а все гибриды F1 имеют одинаковый гетерозиготный генотип Аа, у них проявляется желтая окраска семян. Гибриды F1 способны образовывать 2 типа гамет: А и а, каждая из которых с равной вероятностью может слиться с любой другой. В результате в F2 образуются следующие генотипы: АА, аа, Аа и Аа (или: АА, 2Аа, аа). Как видно, генотипов с двумя рецессивными генами втрое меньше. Этим объясняется расщепление по фенотипу 3:1. Расщепление по генотипу составляет 1 : 2 : 1, то есть 1АА : 2Аа : 1аа. Гомозиготы АА и аа могут образовывать гаметы только одного типа, поэтому при самоопылении у их потомков расщепления не происходит. Гетерозиготы же Аа размножаются аналогично гибридам F1.
- Дигибридное скрещивание
Г.
Мендель продолжил свои исследования, но для экспериментов выбрал растения, отличающиеся друг от друга двумя признаками, то есть по двум парам аллелей. Скрещивание таких организмов называется дигибридным
. В одном из экспериментов семена гороха отличались не только окраской, но и формой (часть из них была гладкой, а часть — морщинистой):
Р
желтые гладкие семена х зеленые морщинистые семена
Все потомки первого поколения имели гладкие семена желтого цвета. Во втором поколении гибридов проявилось уже четыре фенотипа: желтые гладкие, зеленые гладкие, желтые морщинистые и зеленые морщинистые семена. Причем расщепление по фенотипу каждого признака в отдельности было таким же, как и при моногибридном скрещивании — количество желтых семян было втрое больше, чем зеленых, а количество гладких — втрое больше, чем морщинистых. На основании этого был сформулирован еще один принцип, который известен под названием закон независимого наследования (распределения) признаков, суть которого состоит в том, что альтернативные проявления одного признака могут сочетаться с любыми альтернативными проявлениями другого признака. Попробуем объяснить этот закон на основании хромосомной теории наследственности. Согласно этой теории аллели локализуются в гомологических хромосомах. В опытах Г. Менделя гены, кодирующие цвет и форму семян, располагались в разных хромосомах (обозначим ген окраски как А — желтая и а — зеленая, а ген формы как В — гладкая и b — морщинистая). В протекании процесса мейоза, приводящего к образованию гамет, есть одно непреложное правило: гомологичные хромосомы должны разойтись к разным полюсам и «уйти» в разные гаметы. А вот какая именно из гомологических хромосом (с доминантным или рецессивным геном — это, разумеется, относится только к гете-розиготам) отойдет к какому полюсу, дело случая. В нашем примере:
Р
|
желтые гладкие семена |
X |
зеленые морщинистые семена |
| фенотип: |
желтые гладкие семена |
зеленые морщинистые семена |
| генотип: |
AABB |
aabb |
| гаметы: |
AB |
ab |
| F1 |
| фенотип: |
желтые гладкие |
генотип:
Гаметы:
|
AaBb
AB, aB, Ab, ab
|
Различные варианты генотипов (и соответствующих им фенотипов), образующиеся при слиянии гамет в результате скрещивания, удобно рассчитывать по решетке Пеннета, располагая их в ячейках, на которые она поделена.
- Сцепление генов
К
ак выяснилось закон независимого распределения генов справедлив лигаь для генов, расположенных в разных хромосомах. На самом деле в любом организме число генов очень велико (десятки тысяч), а число их носителей — хромосом — ограничено: так, у человека 23 пары хромосом, у кукурузы — 10, а у дрозофилы — всего 4. Соответственно, в каждой хромосоме должно быть по несколько сотен или тысяч генов. Из того факта, что при образовании гамет к полюсам клетки в мейозе отходят хромосомы, а не гены, следует, что гены, локализованные в одной хромосоме, должны наследоваться вместе. Это подтверждают опыты Томаса Ханта Моргана, проведенные на плодовой мушке дрозофиле. Он исследовал дигибридное скрещивание для двух признаков: цвета тела (серое и черное) и длины крыла (длинные и зачаточные).
· Р
серое тело, длинные крылья (GGLL) Х черное тело, зачаточеные крылья (ggll)
· гаметы : GL gl
· F1 серое тело, длинные крылья GgLl
· Поскольку оба гена лежат в одной хромосоме, образуется только 2 типа гамет: GL и gl
F1 гаметы: 2GL, 2gl
F2
|
| фенотип: 3 серое тело, длинные крылья генотипы: GGLL, GgLl, GgLlили 1GGLL : 2GgLl : 1ggll |
1 черное тело, зачаточные крылья ggll |
Таким образом, в F2 наблюдается расщепление по фенотипу 3 : 1 вместо ожидаемого в соответствии с генетикой Менделя 9:3:3:1. Закономерность, суть которой сводится к тому, что гены, локализованные в одной хромосоме, наследуются преимущественно вместе, известна под названием закона Моргана. Слово преимущественно не случайно, ибо сам Морган обнаружил и объяснил отклонения от этого правила. Так как гены, лежащие в одной хромосоме, наследуются вместе, их называют сцепленными. Все гены одной хромосомы образуют ГРУППУ сцепления. Введем еще одно понятие. Анализирующим называется скрещивание изучаемого организма с формой, имеющей рецессивный гомозиготный генотип и соответственно образующей только один тип гамет с рецессивными аллелями. При анализирующем скрещивании (в данном случае оно является также и возвратным) серой длиннокрылой гетерозиготы из F1 с черной короткокрылой гомозиготой из родительского поколения Р у Т. X. Моргана помимо форм с ожидаемыми фенотипами — серое тело, длинные крылья и черное тело, зачаточные крылья — в соотношении 1 : 1 появились особи со смешанными признаками:
Р
серое тело, длинные крылья (GgLl) Х черное тело, зачаточеные крылья (ggll)
Fа(анализир.) 41,5 % серое тело, длинные крылья 41,5 % черное тело, зачаточные крылья 8,5 % серое тело, зачаточные крылья 8,5 % черное тело, длинные крылья
Т. X. Морган, объясняя полученные результаты, предположил, что гомологичные хромосомы, образующие на первой стадии мейоза хиазмы (перекресты), способны обмениваться отдельными участками в результате возникающих разрывов и последующих рекомбинаций. Это явление было названо кроссинговером. Оно приводит к тому, что аллели из гомологичных хромосом меняются друг с другом местами. Таким образом, в данном случае, кроме «нормальных» гамет GL и gl, образуются (в гораздо меньшем количестве) гаметы GI и gL. Именно они и определяют появление «неожиданных» особей. Процесс обмена участками между гомологичными хромосомами приводит к генетической рекомбинации. Особей, образующихся из гамет с новым сочетанием аллелей, называют рекомбинантными. Чем дальше друг от друга на хромосоме расположены гены, тем чаще между ними происходит кроссинговер и тем выше процент появляющихся рекомбинантных особей. На этом явлении основано построение генетических карт — определение последовательности расположения генов в хромосоме и примерного расстояния между ними.
- Взаимодействие генов
Более поздними исследованиями было показано, что, кроме сцепления, отклонения от менделевского наследования вызываются еще рядом причин, одной из которых являются эффекты, связанные с взаимодействием генов. Оказалось, что как аллельные, так и неаллельные гены способны взаимодействовать друг с другом, вызывая появление новых признаков. Взаимодействие аллельных генов Неполное доминирование — явление, при котором доминантный ген не полностью подавляет работу рецессивного, в результате развивается промежуточный признак. Примером может служить окраска цветка у растения ночная красавица с расщеплением по фенотипу в F2 1:2:1. Р
красный цветок (АА) Х белый цветок (аа)
· F1 фенотип: розовый цветок генотип: Аа гаметы: А А Х а а
· F2 фенотип: 1/4 красный цветок 2/4 розовый цветок 1/4 белый цветок генотип: АА Аа аа
Множественный аллелизм
— явление существования более двух альтернативных аллельных генов, имеющих различные проявления в фенотипе. Например, четыре группы крови у человека определяются сочетанием в генотипе аллелей А, В и О одного и того же гена I. Взаимодействие неаллельных геновКомплементарное взаимодействие
— (взаимодополнительное действие генов) — явление, когда признак развивается только при взаимном действии двух доминантных неаллельных
генов, каждый из которых в отдельности не вызывает развитие признака. Комбинативное взаимодействие
— явление, когда два неаллельных гена, взаимодействуя между собой обусловливают развитие нового признака, при этом каждый ген имеет собственное фенотипическое проявление. Эпистаз
— тип взаимодействия генов, при котором один ген подавляет действие другого (неаллельного) гена. Полимерия
— явление, когда несколько неаллель-ных генов отвечают за сходное воздействие на развитие одного и того же признака. Чем больше таких генов присутствует в генотипе, тем ярче проявляется признак. Часто явление полимерии наблюдается при наследовании количественных признаков — удойность коров, яй-ценосность, вес тела и т. д. Плейотролия
— множественное действие гена. В этом случае один ген отвечает за развитие нескольких признаков.
2.
Закономерности изменчивости
К наследственной изменчивости относят изменения признаков организма, которые определяются генотипом и сохраняются в ряду поколений. Иногда это крупные изменения, например коротконогость, отсутствие рогов у домашнего скота, отсутствие пигмента (альбинизм) или оперения. В результате таких изменений возникли также карликовый рост душистого горошка, красная береза, растения, дающие махровые цветки, но чаше это мелкие, едва заметные уклонения от нормы. Наследственные изменения называют мутациями.
Дарвин называл наследственную изменчивость неопределенной, индивидуальной изменчивостью, подчеркивая тем самым ее случайный, ненаправленный характер и относительную редкость. Мутации возникают вследствие изменения структуры гена (т. е. последовательности нуклеотидов в ДНК) или хромосом и служат единственным источником генетического разнообразия внутри вида. Благодаря мутационному процессу появляются различные варианты генов, составляя резерв наследственной изменчивости. Однако бесконечное разнообразие генотипов живых организмов, уникальность каждого генотипа обусловлены комбинативной изменчивостью — перекомбинацией хромосом в процессе полового размножения и участков хромосом в процессе крос-синговера. При этом типе изменчивости сами гены не изменяются, изменяются их сочетания и характер взаимодействия в генотипе.
Мутации бывают доминантные и рецессивные.
Большинство мутаций рецессивны и не проявляются у гетерозигот. Это очень важно для существования вида. Мутации в данных условиях оказываются, как правило, вредными, так как вносят нарушения в тонко сбалансированную систему биохимических реакций. При изменении условий внешней среды некоторые мутации могут оказываться полезными и носители таких мутаций получают преимущество в процессе естественного отбора.
В гомозиготном состоянии мутации нередко понижают жизнеспособность или плодовитость особи. Мутации, резко снижающие жизнеспособность, частично или полностью останавливающие развитие, называются полулетальными или летальными, У человека к таким мутациям относятся ген гемофилии и ген серповидно-клеточной анемии, определяющий синтез аномального гемоглобина.
Если мутация возникает в половых клетках, то она обнаруживается только в следующем поколении. Такие мутации называют генеративными. Мутации могут происходить и в соматических клетках, проявляясь лишь у данного организма. Но при бесполом размножении они могут передаваться потомству.
Мутации, образующиеся в результате замены одного или нескольких нуклеотидов в пределах одного гена и приводящие к изменению строения белков, называются генными. Один ген может мутировать неоднократно. Так возникает серия аллельных генов.
Изменения структуры хромосом служат причиной хромосомных мутаций. Они возникают вследствие утраты части хромосомы. Если оторвавшийся участок может присоединиться к негомологичной хромосоме, то образуется новая комбинация генов. Довольно широко распространено удвоение участка хромосомы.
К мутациям относится также изменение числа хромосом. Вследствие нерасхождения какой-либо пары гомологичных хромосом в мейозе одна из образовавшихся гамет содержит на одну хромосому меньше, а другая на одну хромосому больше, чем в нормальном гаплоидном наборе. При слиянии с другой гаметой возникает зигота с меньшим или большим числом хромосом по сравнению с диплоидным набором, характерным для вида.
У простейших и у растений часто наблюдается кратное увеличение числа хромосом. Такое изменение хромосомного набора носит название полиплоидии. С увеличением числа хромосомных наборов в кариотипе возрастает надежность генетической системы, уменьшается вероятность снижения жизнеспособности в случае мутаций. Полиплоидия нередко влечет за собой повышение жизнеспособности, плодовитости и других жизненных свойств. В растениеводстве это свойство используют, искусственно получая полиплоидные сорта культурных растений, отличающиеся высокой продуктивностью. У высших животных полиплоидия не встречается.
Типы изменчивости
Различают два вида изменчивости: фенотипическую (модификационная) и генотипическую. Фенотипическая изменчивость – это изменение организмов под действием факторов среды и эти изменения не наследуются. Эта изменчивость не затрагивает гены организма, наследственный материал не изменяется. Модификационная изменчивость признака может быть очень велика, но она всегда контролируется генотипом организма. Границы фенотипической изменчивости, контролируемые генотипом организма, называют нормой реакции. Широкая норма реакции приводит к повышению выживаемости. Интенсивность модификационной изменчивости можно регулировать. Модификационная изменчивость направлена. К статистическим закономерностям модификационной изменчивости относятся вариационный ряд изменчивости признака и вариационная кривая. Вариационный ряд представляет ряд вариант, (есть значений признака) расположенных в порядке убывания или возрастания (например: если собрать листья с одного и того же дерева и расположить их по мере увеличения длины листовой пластинки, то получается вариационный ряд изменчивости данного признака). Вариационная кривая – это графическое изображение зависимости между размахом изменчивости признака и частотой встречаемости отдельных вариант данного признака. Наиболее типичный показатель признака – это его средняя величина, то есть среднее арифметическое вариационного ряда. Различают следующие виды фенотипической изменчивости: модификации, морфозы и фенокопии. Модификации – это ненаследственные изменения генотипа, которые возникают под действием фактора среды, носят адаптивный характер и чаще всего обратимы (например: увеличение эритроцитов в крови при недостатке кислорода). Морфозы – это ненаследственные изменения фенотипа, которые возникают под действием экстремальных факторов среды, не носят адаптивный характер и необратимы (например: ожоги, шрамы). Фенокопии – это ненаследственное изменение генотипа, которое напоминает наследственные заболевания (увеличение щитовидной железы на территории, где в воде или земле не хватает йода). Генотипическая изменчивость - при генотипической изменчивости происходит изменение наследственного материала и, обычно, эти изменения наследуются. Это основа разнообразия живых организмов. Различают два вида генотипической изменчивости: мутационная и комбинативная. Комбинативная изменчивость основывается на возникновении новых комбинаций генов родителей. При комбинативной изменчивости в результате слияния родительских гамет возникают новые комбинации генов, однако сами гены и хромосомы остаются неизменными (пример: каждый новый организм является новый комбинацией генов родителей). Механизмы комбинативной изменчивости: 1) независимое расхождение хромосом в анафазу І мейоза. 2) Кроссенговер 3) Случайное слияние гамет 4) Случайный подбор родительских пар Мутационная изменчивость в основе этой изменчивости лежит изменение структуры гена, хромосомы или изменения числа хромосом. Мутация – это спонтанное изменение генетического материала. Мутации возникают под действием мутагенных факторов: А) физических (радиация, температура, электромагнитное излучение); Б) химических (вещества, которые вызывают отравление организма: алкоголь, никотин, колхицин, формалин); В) биологических (вирусы, бактерии). Различают несколько классификаций мутаций. Классификация 1.
Мутации бывают полезные, вредные и нейтральные. Полезные мутации: мутации, которые приводят к повышенной устойчивости организма (устойчивость тараканов к ядохимикатам). Вредные мутации: глухота, дальтонизм. Нейтральные мутации: мутации никак не отражаются на жизнеспособности организма (цвет глаз, группа крови). Классификация 2.
Мутации бывают соматические и генеративные. Соматические (чаще всего они не наследуются) возникают в соматических клетках и затрагивают лишь часть тела. Они будут наследоваться следующим поколениям при вегетативном размножении. Генеративные (они наследуются, т.к. происходят в половых клетках): эти мутации происходят в половых клетках. Генеративные мутации делятся на ядерные и внеядерные (или митохондриальные). Классификация 3.
По характеру изменений в генотипе мутации подразделяются на генные, хромосомные, геномные. Генные мутации (точковые) не видны в микроскоп, связаны с изменением структуры гена (генные мутации изменяют последовательность нуклеотидов в молекуле ДНК и ген перестаёт работать). Эти мутации происходят в результате потери нуклеотида, вставки нуклеотида, замены одного нуклеотида другим. Эти мутации могут приводить к генным болезням: дальтонизм, гемофилия. Таким образом, генные мутации приводят к появлению новых признаков. Хромосомные мутации связаны с изменением структуры хромосом. Может произойти делеция – потеря участка хромосомы, дупликация – удвоение участка хромосомы, инверсия – поворот участка хромосомы на 1800, транслокация – это перенос части или целой хромосомы на другую хромосому. Причиной этого может быть разрыв хроматид и их восстановление в новых сочетаниях. Геномные мутации приводят к изменению числа хромосом. Различают анеуплоидию и полиплоидию. Анеуплоидия связана с изменением числа хромосом на несколько хромосом (1, 2, 3): А) моносомия общая формула 2n-1 (45, Х0), болезнь – синдром Шерешевского-Тернера. Б) трисомия общая формула 2n+1 (47, ХХХ или 47, ХХУ) болезнь – синдром Клайнфельтра. В) полисомия Полиплоидия – это изменение числа хромосом, кратное гаплоидному набору (например: 3n 69). Организмы могут быть автоплоидными (одинаковые хромосомы) и аллоплоидными (разные наборы хромосом).
Сходство между комбинативной и мутационной изменчивостью заключается в том, что в обоих случаях потомство получает набор генов каждого из родителей. Мутационная изменчивость является одним из главных факторов эволюционного процесса. В результате мутаций могут возникать полезные признаки, которые под действием естественного отбора дадут начало новым видам и подвидам.
Факторы приводящие к мутации
В естественных условиях мутация появляется под влиянием факторов внешней и внутренней среды и обозначается термином «естественные (или спонтанные) мутации».
Причиной генных, или так называемых точечных, мутаций является замена одного азотистого основания в молекуле Д.Н.К. на другое, потеря, вставка, или перестановка азотистых оснований в молекуле Д.Н.К. Отсюда следует - ген мутирующий у человека могут развиться патологические состояния, патогенез которого различен.
На факторы вызывающие мутации на генном уровне оказало соответствующее влияние окружающей среды (подагру, некоторые формы сахарного диабета). Подобные заболевания чаще проявляются при постоянном воздействии неблагоприятных или вредных факторов окружающей среды (нарушение режима питания и др.). Мутация гена может повлечь за собой нарушение синтеза белков, выполняющих пластические функции. Вероятная причина таких заболеваний синдром Элерса - Данлоса.
В стадии изучения находится заболевания, в основе которых лежит недостаточность механизмов восстановления измененной молекулы Д.Н.К.
Генная мутация может привести к развитию иммунодефецит-ных болезней (аплазия вилочковой железы в сочетании агаммагло-булинемией). Причиной аномальной структуры гемоглобина являе-тся замена в молекуле остатка глутаминовой кислоты на остаток валина.
Известен ряд мутаций генов, контролирующих синтез факто-ров свертывания крови.
Генные мутации могут быть причиной нарушения транспорта различных соединений через клеточные мембраны. Они связаны с нарушением функций мембранных механизмов и с дефектами в некоторых системах.
Если мутация на генном уровне возникает при действии различных физических., химических, биологических факторов, то это называют мутагенезом .
Основой мутации являются первичные повреждения в молекуле Д.Н.К.
Мутагены
Мутагены
(от греч. γεννάω — рождаю) — химические и физические факторы, вызывающие наследственные изменения — мутации. Впервые искусственные мутации получены в 1925 году Г. А. Надсеном и Г. С. Филипповым у дрожжей действием радиоактивного излучения радия; в 1927 году Г. Мёллер получил мутации у дрозофилы действием рентгеновских лучей. Способность химических веществ вызывать мутации (действием иода на дрозофилы) открыта И. А. Рапопортом. У особей мух, развившихся из этих личинок, частота мутаций оказалась в несколько раз выше, чем у контрольных насекомых.
Классификация
Мутагенами могут быть различные факторы, вызывающие изменения в структуре генов, структуре и количестве хромосом. По происхождению мутагены классифицируют на эндогенные
, образующиеся в процессе жизнедеятельности организма и экзогенные
— все прочие факторы, в том числе и условия окружающей среды.
По природе возникновения мутагены классифицируют на физические, химические и биологические:
1.Физические мутагены
-ионизирующее излучение;
-радиоактивный распад;
-ультрафиолетовое излучение;
-моделированное радиоизлучение и электромагнитные поля;
-чрезмерно высокая или низкая температура.
2.Химические мутагены
-окислители и восстановители (нитраты, нитриты, активные формы кислорода);
-алкилирующие агенты (например, иодацетамид);
-пестициды (например гербициды, фунгициды);
-некоторые пищевые добавки (например, ароматические углеводороды, цикламаты);
-продукты переработки нефти;
-органические растворители;
-лекарственные препараты (например, цитостатики, препараты ртути, иммунодепрессанты).
-К химическим мутагенам условно можно отнести и ряд вирусов (мутагенным фактором вирусов являются их нуклеиновые кислоты — ДНК или РНК)
3.Биологические мутагены
-специфические последовательности ДНК — транспозоны;
-некоторые вирусы (вирус кори, краснухи, гриппа);
-продукты обмена веществ (продукты окисления липидов);
-антигены некоторых микроорганизмов.
Типы мутаций
Мутации, помимо качественных свойств, характеризует и способ возникновения. Спонтанные (случайные) - мутации, возникающие при нормальных условиях жизни. Спонтанный процесс зависит от внешних и внутренних факторов (биологические, химические, физические). Спонтанные мутации возникают у человека в соматических и генеративных тканях. Метод определения спонтанных мутаций основан на том, что у детей появляется доминантный признак, хотя у его родителей он отсутствует. Проведенное в Дании исследование показали, что примерно одна из 24000 гамет несет в себе доминантную мутацию. Ученый же Холдейн рассчитал среднюю вероятность появления спонтанных мутаций, которая оказалась равна 5*10-5 за поколение. Другой ученый Курт Браун предложил прямой метод оценки таких мутаций, а именно: число мутаций разделить на удвоенное количество обследованных индивидов.
Генные (точковые) мутации
затрагивают, как правило, один или несколько нуклеотидов, при этом один нуклеотид может превратиться в другой, может выпасть (делеция), продублироваться, а группа нуклеотидов может развернутся на 180 градусов. Например, широко известен ген человека, ответственный за серповидно - клеточную анемию, который может привести к летальному исходу. Соответствующий нормальный ген кодирует одну из полипептидныз цепей гемоглобина. У мутантного гена нарушен всего один нуклеотид (ГАА на ГУА). В результате в цепи гемоглобина одна аминокислота заменена на другую (вместо глутамина - валин). Казалось бы ничтожное изменение, но оно влечет за собой роковые последствия: эритроцит деформируется, приобретая серповидно - клеточную форму, и уже не способен транспортировать кислород, что и приводит к гибели организма. Генные мутации приводят к изменению аминокислотной последовательности белка. Наиболее вероятное мутация генов происходит при спаривании тесно связанных организмов, которые унаследовали мутантный ген у общего предка. По этой причине вероятность возникновения мутации повышается у детей, чьи родители являются родственниками. Генные мутации приводят к таким заболеваниям, как амавротическая идиотия, альбинизм, дальтонизм и др.
Интересно, что значимость нуклеотидных мутаций внутри кодона неравнозначна: замена первого и второго нуклеотида всегда приводит к изменению аминокислоты, третий же обычно не приводит к замене белка. К примеру, "Молчащая мутация"- изменение нуклеотидной последовательности, которая приводит к образованию схожего кодона, в результате аминокилотная последовательность белка не меняется.
Геномные мутации
Главная отличительная черта геномных мутаций связана с нарушением числа хромосом в кариотипе. Эти мутации так же подразделяются на два вида: полиплоидные анеуплоидные.
Полиплоидные мутации ведут к изменению хромосом в кариотипе, которое кратно гаплоидному набору хромосом. Этот синдром впервые был лишь обнаружен в 60-ых годах. Вообще полиплодия характерна в основном для человека, а среди животных встречается крайне редко. При полиплоидии число хромосом в клетке насчитывается по 69 (триплодие), а иногда и по 92 (тетраплодие) хромосомы. Такое изменение ведет практически к 100 % смерти зародыша. Триплодие имеет не только многочисленные пороки, но и приводит к потере жизнеспособности. Тетраплодие встречается еще реже, но так же зачастую приводит к летальному исходу.
Анеуплоидные же мутации приводят к изменению числа хромосом в кариотипе, некратное гаплоидному набору. В результате такой мутации возникают особи с аномальным чипом хромосом. Как и триплодия, анеуплодия часто приводит к смерти еще на ранних этапах развития зародыша. Причиной же таких последствий является утрата целой группы сцепления генов в кариотипе.
В целом же, механизм возникновения геномных мутаций связан с патологией нарушения нормального расхождения хромосом в мейозе, в результате чего образуются аномальные гаметы, что и ведет к мутации. Изменения в организме связаны с присутствием генетически разнородных клеток. Такой процесс называется мозаицизм.
Геномные мутации одни из самых страшных. Они ведут к таким заболеваниям, как синдром Дауна (трисомия, возникает с частотой 1 больной на 600 новорожденных), синдром Клайнфельтера и др.
Хромосомные мутации
Хромосомные мутации приводят к изменению числа, размеров и организации хромосом, поэтому их иногда называют хромосомными перестройками. Хромосомные перестройки делятся на внутри- и межхромосомные. К внутрехромосмным относятся:
Дубликация - один из участков хромосомы представлен более одного раза.
Делеция - утрачивается внутренний участок хромосомы.
Инверсия - повороты участка хромосомы на 180 градусов.
Межхромосомные перестройки (их еще называют транслокации) делятся на:
Реципрокные - обмен участками негомологичных хромосом.
Нереципрокные - изменение положения участка хромосомы.
Дицентрические - слияние фрагментов негомологичных хромосом.
Центрические - слияние центромер негомологичных хромосом.
Хромосомные мутации проявляются у 1% новорожденных. Однако интересно, исследования показали, что нестабильность соматических клеток здоровых доноров не исключение, а норма. В связи с этим была высказана гипотеза о том, что нестабильность соматических клеток следует рассматривать не только как патологическое состояние, но и как адаптивную реакцию организма на измененные условия внутренней среды. Хромосомные мутации могут обладать фенотипическими явлениями. Наиболее распостраненный пример - синдром "Кошачьего крика" (плачь ребенка напоминает мяукание кошки). Обычно носители такой делеции погибают в младенчестве. Хромосомные мутации часто приводят к паталогическим нарушениям в организме, но в то же время хромосомные перестройки сыграли одну из ведущих ролей в эволюции. Так, у человека 23 пары хромосом, а у обезьяны - 24. Таким образом различие составляет всего одна хромосома. Ученые предполагают, что в процессе эволюции произошла хотя бы одна перестройка. Подтверждением этого может служить и тот факт, что 17 хромосома человека отличается от такой же хромосомы шимпанзе лишь одной перецентрической инверсией. Такие рассуждения во многом подтверждают теорию Дарвина.
3.
Генные мутации
Генные (точковые) мутации - это изменения числа и/или последовательности нуклеотидов в структуре ДНК (вставки, выпадения, перемещения, замещения нуклеотидов) в пределах отдельных генов, приводящие к изменению количества или качества соответствующих белковых продуктов.
Замены оснований приводят к появлению трех типов мутантных кодонов: с измененным смыслом (миссенс-мутации), с неизмененным смыслом (нейтральные мутации) и бессмысленных, или терминирующих кодонов (нонсенс-мутации).
В результате миссенс-мутании в кодируемом данным геном полипептиде одна аминокислота замещается на другую, поэтому фенотипическое проявление мутации зависит от функциональной значимости затронутого домена. Так замены аминокислот в активных центрах белков могут сопровождаться полной потерей их функциональной активности. К примеру, миссенс-мутация в 553-м кодоне гена FAC, приводящая к замене лейцина на пролин, делает продукт этого гена неспособным комплементировать функциональный дефект в клетках больных анемией Фанкони.
Не всякая замена аминокислоты отразится на функциональной активности белка, вследствие чего происшедшая мутация может остаться не вьшвленной. Этим объясняется факт отмечаемого несовпадения частоты мутаций в определенном гене и встречаемости мутантов по нему. Кроме того, в силу вырожденности генетического кода, не всякая замена основания приведет к миссенс-мутации, возможно, она окажется нейтральной.
В результате нонсенс мутации кодон, определяющий какую-либо аминокислоту, превращается в один из стоп-кодонов, не транслирующихся на рибосомах (UAA UAG, UGA). Появление такого кодона не в конце структурного гена, а внутри него, приводит к преждевременной терминации трансляции и обрыву полипептидной цепи. Нонсенс-мутации обладают наибольшим повреждающим действием, так как образующиеся при преждевременной терминации трансляции белки не способны к модификации, часто не защищены от действия протеолитических ферментов и быстро деградируют.
Вставки, перемещения или выпадения отдельных оснований или их коротких последовательностей в пределах гена вызывают сдвиг рамки считывания. Природа таких мутаций была изучена при анализе аминокислотной последовательности белков фага Т4, кодируемьгх геном дикого типа е+ и тремя разными мутантными генами е, содержащими взаимно супрессирующие фреймшифт (сдвигающие рамку считывания)-мутации. Оказалось, что некоторые единичные мутации являются следствием одновременных изменений нескольких соседних нуклеотидов. И, скорее всего, единичная мутация со сдвигом рамки возникает в результате вставки двух соседних нуклеотидов, а не одного. При возникновении мутаций со сдвигом рамки считывания меняются все триплеты ниже сайта дупликации или делеиии по ходу считывания, при этом повышается вероятность возникновения стоп-кодонов и, соответственно, терминации трансляции.
С точки зрения структурно-функциональной организации генов, происходящие внутри них замены, вставки, выпадения, перемещения нуклеотидов можно объединить в следующие группы:
1) мутации в регуляторных областях генов
• мутации в промоторной части (например, регуляторном элементе с последовательностью PuCPuCCC и внутри ТАТА-бокса у гена р-глобина) снижают уровень синтеза белкового продукта; • мутации в сайте полиаденилирования снижают уровень транскрипции (характерно для афроамериканцев, страдающих талассемией; подробно о гемоглобинопатиях см. часть П Медицинская генетика).
Таким образом, мутации в регуляторных 5' и 3'-нетранслируемых областях генов вызывают количественные изменения соответствующих продуктов и проявляются фенотипически (клинически) в зависимости от порогового уровня белков, при котором их функция еще сохраняется;
2) мутации в кодирующих областях генов
• мутации в экзонах могут приводить к преждевременному окончанию белкового синтеза. Именно это происходит, к примеру, в случае 6-талассемии: в результате мутаций внутри экзона гена гемоглобина белковая цепь оказывается укороченной и не обладает активностью; • мутации в интронах способны генерировать новые сайты сплайсинга, которые, конкурируя с нормальными (исходными), в итоге, заменяют их. Возникновение замен в гене гемоглобина, замедляющих сплайсинг, известно и для В0-, и для В+-талассемии. • мутации в сайтах сплайсинга (на стыках экзонов и нитронов), нарушают процессинг первичного РНК-транскрипта и приводят к трансляции бессмысленных белков: удлиненного при неправильном вырезании интронов либо укороченного при вырезании экзонов. Так, в результате одиночных замен в донорском участке сплайсинга гена гемоглобина процессинг нарушается, что приводит к развитию В0- или р+-талассемии. А мутация сдвига рамки считывания в акцепторном участке сплайсинга гена ХРА приводит к полной инактивации белка и, как следствие, к развитию тяжелой формы пигментной ксеродермы.
Замены нуклеотидов в кодирующих областях генов, не сопровождающиеся заменами аминокислот в силу вырожденности генетического кода, приводят к нейтральным мутациям, не оказывающим заметного влияния ни на функцию соответствующего белка, ни на его структуру.
Наследственные болезни
Наследственные болезни, патологические состояния организма, обусловленные изменениями генетического материала — мутациями. В широком смысле термин «наследственные болезни» включает не только хромосомные и генные болезни, вызываемые соответствующими мутациями, но и мультифакториальные болезни, развитие которых связано с взаимодействием нормальных полиморфных генов, формирующих предрасположенность к заболеванию, с факторами внешней среды. Условно к наследственным болезням можно также отнести болезни, связанные с присутствием в организме некоторых редких вариантов белков (обычно ферментов) и развивающиеся в ответ на более или менее специфические внешнесредовые воздействия, например, на прием некоторых лекарственных средств. В узком смысле под термином «наследственные болезни» понимают моногенные заболевания, т. е. обусловленные мутациями отдельных генов. Многие наследственные болезни проявляются как врожденные состояния. Для наследственных болезней характерно также семейное накопление (заболевание встречается у нескольких членов семьи). Термины «наследственные болезни», «семейные болезни» и «врожденные болезни» (с которыми индивид рождается на свет) не являются синонимами. зрелом возрасте.
Классификация
Наследственные болезни многочисленны (известно свыше 6000) и разнообразны по проявлениям. Тип наследования лежит в основе классификации моногенных наследственных болезней. Для значительной части наследственных болезней тип наследования установлен — патологические признаки, также как и нормальные, могут наследоваться аутосомно-доминантно, аутосомно-рецессивно и сцепленно с полом (Х-сцепленный доминантный, Х-сцепленный рецессивный и Y-сцепленный типы наследования). Термин «аутосомный» указывает на то, что мутантный ген локализован в аутосоме, «Х-сцепленный» — в половой Х-хромосоме, а «Y-сцепленный» — в половой Y-хромосоме. Выделение доминантного и рецессивного типов наследования существенно с медицинской точки зрения, так как при доминантном типе наследования клиническое проявление болезни обнаруживается у гомо- и гетерозигот, а при рецессивном — только у гомозигот, т. е. значительно реже. Основные методы, с помощью которых устанавливается тот или иной тип наследования, — клинико-генеалогический, базирующийся на анализе родословных, и более точный сегрегационный анализ, объектом которого, как правило, являются так называемые ядерные семьи (т. е. родители и дети).
К наиболее часто встречающимся (частота 0,5-0,1 на 1000 человек) наследственным болезням с аутосомно-доминантным типом наследования относятся нейрофиброматоз, поликистоз почек (взрослый тип), несовершенный дентиногенез, хорея Гентингтона, сфероцитоз, одна из форм детской тугоухости и некоторые другие. При анализе родословных, в которых прослеживается этот тип наследования, выявляется, что один из родителей, как правило, болен; вероятность развития болезни у детей 50%; оба пола поражаются одинаково часто; наблюдается «вертикальная» передача наследственной болезни в ряду поколений. Примером довольно распространенных (по крайней мере в европейских популяциях) наследственных болезней с аутосомно-рецессивным типом наследования являются муковисцидоз, фенилкетонурия, адреногенитальный синдром, врожденная нейросенсорная тугоухость. При этом типе наследования родители в абсолютном большинстве случаев здоровы, но являются гетерозиготными носителями мутантного гена; вероятность рождения больного ребенка составляет 25%; оба пола поражаются с одинаковой частотой; родители больных детей часто оказываются кровными родственниками. Рецессивные наследственные болезни, сцепленные с X-хромосомой (например, миопатия Дюшенна, гемофилия А и В, ихтиоз), как правило, проявляются у мальчиков. Если мать является гетерозиготной носительницей мутантного гена, то вероятность рождения больного мальчика у нее составляет 50%. Девочки практически здоровы, но половина из них являются носителями мутантного гена. Больной отец не передает болезнь сыновьям, но все его дочери — гетерозиготные носительницы мутантного гeнa. Наследственные болезни с доминантным типом наследования, сцепленного с X-хромосомой, весьма редки (например, витамин D-резистентный рахит). Проявление заболевания не зависит от пола, однако более тяжело протекает у мальчиков. Среди детей больного мужчины все сыновья здоровы, все дочери — больны. Больные женщины передают мутантный ген половине сыновей и дочерей.
Для многих наследственных болезней характерна генетическая гетерогенность, т. е. к сходным фенотипическим проявлениям могут приводить различные мутационные изменения генетического материала. Такие изменения могут затрагивать либо разные гены, контролирующие обычно один метаболический путь (как, например, для ряда аутосомно-рецессивных форм мукополисахаридозов, ихтиоза, врожденной катаракты и других наследственных болезней, для которых существуют формы с разными типами наследования), либо разные сайты одного и того же гена (некоторые формы -талассемии, миопатии Дюшенна и Беккера, муковисцидоз и многие другие наследственные болезни). В результате реализации международной программы «Геном человека» тысячи генов наследственных болезней локализованы в определенных участках хромосом, а многие сотни генов выделены и идентифицированы — установлена их точная структура, а также природа мутационных изменений. Исследования молекулярно-генетических механизмов, лежащих в основе возникновения моногенных наследственных болезней, позволили обнаружить различные типы мутаций, ведущих к нарушению работы генов при целом ряде наследственных болезней.
Так как для многих наследственных болезней первичный биохимический дефект еще не известен, их классифицируют по клиническому принципу. Выделяют наследственные болезни нервной, эндокринной, сердечно-сосудистой, пищеварительной систем, наследственные болезни кожи и ее придатков, глаз, скелета, системы крови и т. п. Такая классификация в значительной степени условна, так как при большинстве наследственных болезней в патологический процесс вовлекаются органы, входящие в разные системы организма.
Большинство наследственных болезней проявляется уже в детском возрасте или даже при рождении. Проявление наследственных болезней, как правило, варьирует. Присутствие или отсутствие признака при наличии в генотипе гена, детерминирующего его, а также степень проявления признака определяются пенетрантностью и экпрессивностью
гена, которые зависят, с одной стороны, от взаимодействия с другими генами, с другой — от взаимодействия с факторами окружающей среды. Примером, показывающим насколько может варьировать проявление наследственной патологии человека, определяемой моногенно, является успешное лечение специальной диетой больных фенилкетонурией, которое позволяет практически полностью исправить проявление мутантного гена.
Основные причины искажений в картине наследования (отклонения от моногенного наследования) наследственных болезней — неполная пенетрантность и спорадические случаи заболевания, которые включают вновь возникающие мутации в половых клетках, фенокопии (внешне сходные по клинической картине заболевания, обусловленные факторами внешней среды) и другие. Наследование по правилам Менделя не прослеживается также при наследственных заболеваниях, обусловленных мутациями митохондриальных генов. Известно около 30 наследственных митохондриальных заболеваний.
Статистика рождения детей с наследственными болезнями
Примерно 5-6 детей из 100 рождаются с какими-нибудь генетически обусловленными заболеваниями. В большинстве — это заболевания с генетическими предрасположенностями. Это могут быть пороки развития, нарушения в интеллектуальном развитии ребенка. В эти 5-6 процентов входят наследственные заболевания, возникшие впервые или унаследованные от одного из родителей.
Литература
Интернет-ресурсы.
Учебники биологии и химии за 10-11 класс.
Медицинские энциклопедии.
Беседа с преподавателями биологии и химии.
|
