РЕФЕРАТ
по дисциплине: Ботаника
ТЕМА: ”Деление и онтогенез клетки”
2009
План
1. Строение ядра.
2. Деление клетки.
2.1 Митоз
2.2 Мейоз
3. Онтогенез растительной клетки.
Список литературы
1. Строение ядра
Ядро – обязательная и существенная часть живой клетки всех эукариотических организмов. Ядро растительной клетки открыл Роберт Броун. Классические труды по выяснению роли ядра в жизни клетки принадлежат русскому ботанику Герасимову (1890-1905). Ядро контролирует жизнедеятельность клетки, сохраняет генетическую информацию и передает ее дочерним клеткам в процессе клеточного деления. Клетки с удаленным ядром быстро гибнут. Единственным типом клеток, которые остаются живыми и во взрослом состоянии не имеют ядра, являются ситовидные трубки, но живут они недолго: обычно один вегетационный период.
Живая клетка содержит одно ядро. Однако существуют многоядерные клетки некоторых водорослей и низших грибов. Двухъядерные бывают клетки выстилающего слоя пыльников.
В световой микроскоп ядро плохо видно, потому что преломляет свет не намного больше, чем окружающая цитоплазма. Поэтому без специальной окраски ядро тяжело обнаружить.
Ядро – крупный клеточный органоид. Размеры ядра зависят от вида растения, типа, возраста и состояния клетки. В вегетативных клетках высших растений размеры ядра от 5 до 25 мк. У однодольных ядра крупнее, чем у двудольных. У голосеменных крупнее, чем у покрытосеменных. Самые крупные ядра (до 500 мк) в половых клетках голосеменных.
Форма ядра чаще шаровидная, в прозенхимных клетках – вытянутая. В молодых клетках занимает центральное положение, затем смещается к оболочке, оттесняемое вакуолью.
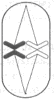
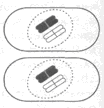
Ядро эукариотической клетки окружено двумя мембранами, которые образуют покровы ядра (рис. 1). Между ними находится перинуклеарное пространство. Внутренняя мембрана агранулярного типа. Наружная мембрана гранулярная: к ней прикрепляются рибосомы. В некоторых местах эта мембрана объединяется с ЭПР. Покровы ядра можно рассматривать как специализированную, локально дифференцированную часть ЭПР.
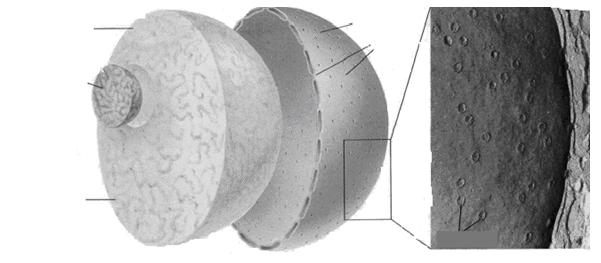 Рис. 1. Строение ядра:
Рис. 1. Строение ядра:1- ядрышко; 2- нуклеоплазма; 3- хроматин; 4- мембраны ядра; 5- ядерные поры.
Покровы ядра имеют поры, которые размещены у некоторых растений в правильном порядке. Мембраны смыкаются вокруг каждой поры и образуют края поры. Ядерная пора не является простым отверстием, а имеет сложную структуру. Кольцо состоит из глобулярных телец, связанных фибриллами. Это кольцо окружает пору. Имеется центральная гранула и система фибрилл между гранулой и кольцом. Через ядерные поры осуществляется обмен веществ между ядром и цитоплазмой, например, выход в цитоплазму матричной РНК и рибосомных субчастиц или поступление в ядро рибосомных белков, нуклеотидов и молекул, которые регулируют активность ДНК.
Реклама

Содержимое ядра представляет собой гелеобразный матрикс, называемый нуклеоплазмой. В состав ее входят разные химические вещества (белки, нуклеотиды) в виде истинного коллоидного раствора. В нуклеоплазме находятся хроматин и ядрышки. Хроматин состоит из отрицательно заряженной ДНК и положительно заряженных специальных белков – гистонов и липидов. Гистоны и ДНК объединены в структуры, которые по виду напоминают бусы. Их называют нуклеосомами. В составе нуклеосом ДНК плотно упакована и защищена от воздействия ферментов. Между нуклеосомами расположены нити ДНК с прикрепленными к ним негистоновыми белками.
В интерфазе хроматин переходит в более диспергированное состояние. Часть его остается плотно спирализованной и по-прежнему хорошо окрашивается. Эта часть называется гетерохроматином. Гетерохроматин имеет вид темных пятен, расположенных обычно ближе к покровам ядра. Остальной, более рыхло спирализованный хроматин, локализуется ближе к центру ядра. Это эухроматин.
Во время деления ядра хроматин конденсируется и образует более туго спирализованные нити или палочковидные тельца – хромосомы, названные так Вальдейером в 1888 году. Количество их для клеток каждого вида постоянно. В любой клетке гороха, например, находятся 14 хромосом – по семь от каждого родителя. В подсолнечнике – 34. Это приведены хромосомные числа в соматических клетках – диплоидный набор 2n. Половые клетки, или гаметы, содержат только половину количества хромосом, характерную для соматических клеток организма. Количество хромосом в гаметах называют гаплоидным набором хромосом. Гаплоидные половые клетки находятся в зрелых пыльцевых зернах и в зародышевом мешке семязачатка.
Клетки, имеющие более двух наборов хромосом, – полиплоидные. Приставки три-, тетра- и так далее показывают, во сколько раз увеличено количество хромосом, то есть: степень плоидности: 3n – триплоид, 4n – тетраплоид, 5n –пентаплоид и так далее. У растений полиплоидия встречается гораздо чаще, чем у животных. Большинство растений способно к вегетативному размножению и поэтому эффективно воспроизводятся в полиплоидном состоянии. Растения-полиплоиды чаще характеризуются крупными размерами, повышенным содержанием ряда веществ, устойчивостью к неблагоприятным факторам окружающей среды и другими хозяйственно полезными качествами. Они представляют собой важный источник изменчивости и могут быть использованы как исходный материал для селекции и создания высокоурожайных сортов растений. Среди наиболее важных сельскохозяйственных культур полиплоиды – пшеница, хлопчатник, сахарный тростник, банан, картофель, подсолнечник. Красивые садовые цветы (хризантемы, георгины) – также полиплоидные.
Реклама

Искусственно полиплоидные растения получают при помощи колхицина – алкалоид, который угнетает образование митотического веретена в результате нарушения образований микротрубочек. Встречаются полиплоиды и в природных популяциях.
Одна из первых попыток построения схемы строения хромосом принадлежит Кольцову (1934).
Хромосома состоит из двух хроматид, основой которых является молекула ДНК. Неконденсированный участок ДНК формирует первичную перетяжку, скрепляющую хроматиды – центромеру. На ней расположены дисковидные тельца, на которых фиксируются нити веретена деления. У некоторых хромосом есть вторичная перетяжка, которая не имеет белковых телец. Она отделяет от остальной части хромосомы спутник. Вокруг вторичной перетяжки в ядре формируется ядрышко.
Ядрышко – сферическая структура внутри ядра. Шлейден в 1842 г. впервые указал на существование ядрышек. Граница между ядрышком и нуклеоплазмой не выражена, потому что ядрышко не окружено мембраной и находится в непосредственном контакте с другими компонентами ядра. Ядрышко имеет плотную структуру, состоящую из элементов двух типов – гранулярных и фибриллярных.
Некоторые из гранул содержат РНК, их можно сравнить по размерам с цитоплазматическими рибосомами. Мелкие гранулы представляют собой белок. Фибриллярный компонент содержит ДНК.
В центральной части ядрышка начинается скручивание рибосомной РНК и идет сборка рибосом, которая завершается в цитоплазме. Таким образом, в ядрышках синтезируются рибосомальные РНК. Деление и образование новых эукариотических клеток связано с делением ядра. Новые ядра всегда возникают в результате деления уже имеющихся.
2. Деление клетки
2.1 Митоз
Процесс деления у эукариот можно разделить на две стадии: митоз и цитокинез.
Митоз (от греч. «митос» – нить) – это образование из одного ядра двух дочерних ядер, морфологически и генетически эквивалентных друг другу. Цитокинез включает деление цитоплазматической части клетки с образованием дочерних клеток.
Биологическая роль митоза состоит в одинаковом распределении редуплицированных хромосом между дочерними клетками, что обеспечивает образование генетически равнозначных клеток.
Интерфазу, период между двумя последовательными митотическими делениями, можно разделить в свою очередь на три периода. Первый начинается после митоза. В это время увеличивается количество цитоплазмы, включающей разные органеллы; синтезируются вещества, стимулирующие или ингибирующие остальную часть цикла. Во второй период происходит удвоение генетического материала (ДНК). В третий период формируются структуры, непосредственно принимающие участие в митозе, например, компоненты нитей веретена.
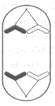
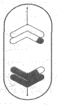
В процессе митоза выделяют несколько стадий: профазу, метафазу, анафазу и телофазу (рис. 2). В профазе хромосомы укорачиваются и утолщаются и распределяются более упорядоченно. В конце профазы покровы ядра и ядрышко исчезают.
В метафазе хромосомы окончательно обособляются и собираются в экваториальной плоскости посредине между полюсами бывшего ядра. Хроматиды начинают отделяться друг от друга, но остаются связанными в области центромеры. В это время микротрубочки образуют ряд нитей, расположенных между полюсами ядра – митотическое веретено.
Рис. 2. Сравнительная схема митоза и мейоза (на примере одной пары гомологичных хромосом). Обозначения: n- плоидность; с- количество хроматина.
В анафазе делятся центромеры. Каждая хромосома разделяется на две самостоятельные хроматиды, которые становятся дочерними хромосомами.
Нити веретена, прикрепленные к хромосомам, укорачиваются и способствуют расхождению и движению дочерних хромосом в противоположные стороны. При этом центромеры двигаются впереди, а плечи хромосом тянутся сзади. Микротрубочки постоянно формируются на одном конце нити веретена и разрушаются на другом.
К моменту телофазы дочерние хромосомы достигают полюсов клетки, веретено исчезает, хромосомы удлиняются и постепенно становятся вновь неразличимыми в форме хроматиновых нитей. Одновременно появляются ядрышки и покровы ядра вокруг двух новых ядер.
Митоз завершается, когда хромосомы становятся невидимы и два дочерних ядра вступают в интерфазу. Эти ядра генетически эквивалентны друг другу и материнскому ядру.
Продолжительность митоза зависит от организма, типа ткани. Однако профаза всегда самая продолжительная, анафаза – самая короткая. В апексе корня продолжительность профазы 1-2 часа; метафазы – 5-15 мин.; анафазы – 2-10 мин.; телофазы – 10-30 мин.; интерфазы – 12-30 часов.
После митоза происходит процесс деления цитоплазмы – цитокинез. Образуется бочковидная система волокон – фрагмопласт.
В зависимости от того, где будет перегородка между дочерними ядрами, выделяют типы деления клетки:
1) тангенциальный: перегородка закладывается параллельно поверхности и обуславливает рост организма в толщину;
2) антиклинальный: перегородка закладывается перпендикулярно поверхности. Организм нарастает своей поверхностью;
3) периклинальный: перегородка закладывается под углом к поверхности. Деление в объем.
Митоз осуществляется по мере роста растений, поэтому его обозначают термином соматическое деление ( от греч. «сома» – тело).
2.2 Мейоз
Мейоз ( от греч. «мейезис» – уменьшение ) – способ деления клеток, при котором в отличие от митоза происходит редукция (уменьшение) количества хромосом и переход клеток из диплоидного состояния в гаплоидное.
У растений мейоз – основа образования гамет и гаплоидных спор. Мейоз состоит из двух последовательных делений ядра, в процессе которых удвоение ДНК происходит один раз. Отличительной особенностью первого деления мейоза является сложная и растянутая во времени профаза. Хромосомы принимают вид длинных тонких нитей. Гомологичные хромосомы спариваются (конъюгируют) друг с другом. Каждый гомолог происходит от одного из родителей и состоит из двух идентичных хроматид. Таким образом, гомологичные пары состоят из четырех хроматид. Конъюгация гомологичных хромосом является обязательной частью мейоза. В гаплоидных клетках конъюгация не встречается, так как в них отсутствуют гомологи.
Объединенные пары гомологичных хромосом называются бивалентами. В это время четыре прижатые друг к другу хроматиды могут обмениваться отдельными участками (кроссинговер). В любой данной точке кроссинговер может быть у двух хроматид. В результате кроссинговера хроматиды имеют другой состав генов. В конце профазы I фрагментируются покровы ядра и образуется веретено деления. В метафазе I биваленты размещаются по экватору веретена, образуя экваториальную пластинку. Центромеры гомологичных хромосом расположены по противоположным сторонам от экваториальной плоскости. В метафазе митоза наоборот: центромеры отдельных хромосом находятся в экваториальной плоскости. В анафазе I начинается движение гомологичных хромосом к противоположным полюсам клетки. В анафазе митоза центромеры делятся и расходятся идентичные хроматиды. В анафазе I мейоза центромеры не делятся, хроматиды остаются вместе, а разъединяются гомологичные хромосомы. Однако из-за обмена фрагментами в результате кроссинговера хроматиды не идентичны, как в начале мейоза.
Завершается первое деление телофазой I, когда спирализация хромосом ослабляется, они удлиняются и вновь становятся невидимыми.
Телофаза переходит в интерфазу. Но у многих организмов интерфаза отсутствует. Затем в каждой из дочерних клеток происходит второе деление митоза. Во втором делении митоза также выделяют стадии: профаза II, метафаза II, анафаза II, телофаза II. По сущности, это митоз в клетке с гаплоидным набором хромосом. При этом к противоположным полюсам расходятся сестринские хроматиды. Несмотря на то, что в процессе мейоза генетический материал удваивается только один раз, происходит два деления, в результате чего образуется четыре клетки, каждая с уменьшенным вдвое (гаплоидным) количеством хромосом, а благодаря кроссинговеру, и с новыми, отличными от родительских, типами хромосом. При митозе наборы хромосом дочерних ядер и материнского были идентичными.
Эндомитоз – внутреннее деление ядра, которое отличается от обычного кариокинеза тем, что ядрышко и покровы ядра сохраняются, а хромосомы разделяются в продольном направлении, и все остаются в составе одного ядра. Встречается у видов родов Lycopersicum, Taraxacum. Распространенность и механизм эндомитоза пока не известны.
3. Онтогенез растительной клетки
С момента образования и до отмирания клетки проходят определенный путь развития, в котором выделяют три основные фазы: эмбриональную, растяжения и дифференциации.
Эмбриональная фаза развития – процесс новообразования клеток путем их деления. Образовавшиеся в результате деления клетки мелкие, расположены плотно, оболочка тонкая, богатая пектином, мало клетчатки. В цитоплазме развита ЭПР, много рибосом.
Фаза растяжения. Клетки растут, увеличиваются в объеме. Протопласт вакуолизируется. Мелкие многочисленные вакуоли сливаются, размер их увеличивается, количество сокращается. Цитоплазма становится более жидкой.
В фазе дифференциации клетки приобретают индивидуальные особенности. Специализация клеток происходит в соответствии с их расположением в растении. Обязательный момент дифференциации – образование вторичной оболочки. Характерные морфологические особенности приобретает протопласт.
Выполнившие свои физиологические функции клетки отмирают. Толстостенные клетки и после отмирания продолжают функционировать как проводящие или механические ткани. Тонкостенные после отмирания протопласта сплющиваются под давлением соседних клеток и постепенно рассасываются. Этот процесс называется облитерация (лат. obliteratus – прекращение).
Известно, что соседние клетки как бы сцементированы срединной пластинкой. Поэтому, когда она разрушается при помощи некоторых веществ, то оболочки соседних клеток отклеиваются друг от друга, и клетки разъединяются. Процесс разъединения клеток в результате разрушения срединной пластинки называют мацерацией. Впервые метод мацерации для изучения строения растительных клеток применил немецкий ботаник Мольденгауэр (1812). Естественная мацерация происходит, когда пектиновые вещества срединной пластинки при помощи пектиназы переходят в растворимое состояние и потом вымываются водой. Наблюдается естественная мацерация в переспевших плодах груши, дыни, персика. Естественная мацерация происходит в черешках листьев перед листопадом, в плодоножках и лепестках перед их опаданием. Мацерация наглядно показывает, что каждая клетка имеет свою собственную оболочку.
При переходе клеток из эмбрионального во взрослое состояние наблюдается частичная мацерация клеток, когда срединная пластинка разрушается по углам клеток. В результате тургорного давления соседние клетки в этих местах округляются, образуются межклетники. Межклетники такого типа называют схизогенными, образующиеся путем расщепления. Некоторые межклетники возникают путем растворения, распада целых клеток. Это лизигенные межклетники.
Межклетники обоих типов могут служить вместилищем разнообразных продуктов секреции.
Список литературы
1. Бавтуто Г.А., Еремин В.М. Ботаника: Морфология и анатомия растений. Мн., 1997. – 375 с.
2. Васильев А.Е. и др. Ботаника: Морфология и анатомия растений. М., 1988. – 480 с.
3. Грин Н., Стаут У., Тейлор Д. Биология. В 3-х т. М., 1990.
4. Лемеза Н.А., Лисов Н.Д. Клетка – основа жизни. Мн., 1997. – 168 с.
5. Хржановский В.Г. Курс общей ботаники. М., 1976. – 272 с.
6. Яковлев Г.П., Челомбитько В.А. Ботаника. М., 1990. – 367 с.
|