На правах рукописи
Автореферат
диссертации на соискание ученой степени
доктора биологических наук
Романова Алексея Анатольевича
Специальность 03.02.08 – Биология (Биологические науки)
Закономерности формирования и динамики авифауны гор Азиатской Субарктики
Москва - 2010
Работа выполнена на кафедре зоологии и экологии биолого-химического факультета Московского педагогического государственного университета, в ФГУ «Государственный природный заповедник «Путоранский»
Научный консультант:
доктор биологических наук, профессор
Константинов Владимир Михайлович
Официальные оппоненты:
доктор биологических наук, профессор
Остапенко Владимир Алексеевич
доктор биологических наук, академик РАЕН
Равкин Евгений Соломонович
доктор биологических наук, профессор
Рогачева Энергия Васильевна
Ведущая организация:
Научно-исследовательский
Зоологический музей
Московского государственного
Университета им. М.В. Ломоносова
Защита состоится «____» ноября 2010 г. в ______ часов на заседании диссертационного совета Д 212.154.20 при Московском педагогическом государственном университете по адресу: 129164, г. Москва, ул. Кибальчича, д. 6, корп. 5, ауд. 304.
С диссертацией можно ознакомиться в библиотеке Московского педагогического государственного университета по адресу:
119992, г. Москва, ул. Малая Пироговская, д. 1.
Автореферат разослан «_____» ______________ 2010 г.
Ученый секретарь диссертационного совета,
кандидат биологических наук С.П. Шаталова
Введение
Актуальность
проблемы. Познание механизмов ландшафтно-биотопического размещения, адаптивных особенностей, закономерностей географического распространения и динамики ареалов птиц обширных горных областей имеет исключительно важное значение для решения таких фундаментальных теоретических проблем орнитологии, как палеогеографические и экологические аспекты формирования фаунистических комплексов современной суши (Кищинский, 1974, 1977; Исаченко, Шляпников, 1989; Назаренко, 1990, 1992; Жуков, 2004; Чернов, 2008). Исходя из этого, мы предприняли попытку аналитического обобщения орнитофаунистических исследований в горах Азиатской Субарктики (в дальнейшем сокращенно называемых ГАС). Предполагая естественные пути развития экосистем горных областей, их можно рассматривать в качестве модели для решения таких фундаментальных проблем биологической науки как познание исторических закономерностей формирования, динамики и развития фаун, последующего мониторинга трансформации субарктических горных экосистем.
Реклама

Горы Азиатской Субарктики до сих пор еще слабо изучены. При этом, развитие исследований в этих регионах и объективно существующая географо-экологическая общность цепи ГАС придает исключительную актуальность выявлению общих закономерностей формирования и динамики авифауны на всем ее протяжении с учетом региональной неоднородности и высотно-поясной дифференциации. Особую актуальность приобретают познание закономерностей, определяющих суть, причины и механизмы, интенсивность и направленность пространственных изменений видового состава, соотношения таксономических и географо-генетических групп птиц, присутствия или отсутствия представителей тех или иных типов фаун. Фундаментальных обобщений по авифаунам столь обширных горных областей Севера Азии до сих пор не проводилось. Глобальная по своей сути работа Ю.И. Чернова (1978), посвященная структуре животного населения Субарктики, не имела основной целью выявление специфики горно-субарктических экосистем. Орнитологические исследования А.А. Кищинского (1988) на северо-востоке Азии охватывали хотя и значительную, но все же часть Северной Азии, и могут рассматриваться как базисные для развития теоретических обобщений намного более обширной и разнообразной в природном отношении всей цепи гор Азиатского Севера.
Основная цель работы – выявление общих закономерностей формирования и динамики авифауны гор Азиатской Субарктики с учетом высотно-ландшафтной, региональной и провинциальной неоднородности; комплексный анализ авифауны гор Азиатской Субарктики в сопоставлении с прилежащими равнинами и горами юга Палеарктики.
Достижение этой цели подразумевает решение следующих задач:
1. Выполнение полной инвентаризации видового состава всей авифауны гор Азиатской Субарктики, включающей целенаправленный сбор, систематизацию и критический анализ всех материалов по авифауне.
2. Выявление и уточнение характера пребывания, границ ареалов, особенностей распространения, экологии птиц в экстремальных условиях гор Азиатской Субарктики.
3. Выявление и анализ пространственно-временных трендов изменения горизонтальных и вертикальных границ распространения видов в специфических условиях горных систем.
Реклама

4. Выявление специфических для гор Азиатской Субарктики абиотических факторов среды, определяющих закономерности формирования и динамики горно-субарктической авифауны.
5. Проведение всестороннего, детального сравнительного анализа фауны и населения птиц ключевых горных систем Азиатской Субарктики; выявление общих тенденций и региональных отличий в распределении птиц в пределах исследуемой области.
6. Выявление уровня общности происхождения и формирования авифауны гор и равнин Азиатской Субарктики.
7. Выявление высотно-поясной, широтной и меридиональной дифференциации авифауны гор Азиатской Субарктики.
8. Анализ высотно-поясной изменчивости летнего населения птиц по основным показателям (лидирующие по обилию виды, плотность, видовое богатство) и выявление основных пространственных и временных трендов.
Основные положения, выносимые на защиту:
1. Закономерности формирования и динамики фауны и населения птиц плато Путорана рассматриваются в качестве модели этих процессов, характерных для всех горных систем Азиатской Субарктики. В пределах ГАС, образующих единую обширную область с однородным типом физико-географической среды, действуют сходные факторы пространственной дифференциации фауны и населения птиц.
2. Фаунистическая, географо-генетическая, и экологическая гетерогенность авифауны горных систем Азиатской Субарктики отражает закономерности разнонаправленного поэтапного ее формирования в условиях ландшафтов, имеющих перигляциальный (или аналогичный) генезис.
3. Авифауна гор Азиатской Субарктики характеризуется специфическим типом высотно-поясной дифференциации, в соответствии с которым, с достаточно высокой степенью автономности, сформировались четко идентифицируемые самостоятельные авифауны гольцового, подгольцового и лесного поясов. На фоне доминирующей высотно-поясной изменчивости авифауны выражены также ее межрегиональные и внутрирегиональные отличия.
4. Наряду с гидротермическим режимом, континентальностью климата, и определяемым ими господствующим типом растительности, - в горах Азиатской Субарктики в качестве ведущего фактора пространственной динамики авифауны выступает литогенная основа.
5. Более однородная авифауна формируется в однородном ландшафте, сохраняющем непрерывность на обширном трансконтинентальном пространстве. В противоположность этому, наиболее регионально-специфичные авифауны формируются в дизъюнктивных «островных» областях однородного ландшафта.
6. Основу авифауны гор Азиатской Субарктики формируют равнинные виды не имеющие специальных адаптаций к жизни в горах. Формирование авифауны Субарктики связано с расширением гнездовых ареалов птиц не только на равнинах, но и в обширных горных ее областях. В последних процесс идет не только в горизонтальной, но и в вертикальной плоскости, образующих отчетливо выраженное трехмерное пространство ареала.
Научная новизна работы. Впервые детально и в сравнительном аспекте проанализирована фауна и население птиц ключевых горных систем всей Азиатской Субарктики. В работе впервые проведен анализ специфических абиотических факторов среды перигляциального генезиса, определяющих закономерности формирования горно-субарктической авифауны. Впервые составлены полные видовые списки гнездовой авифауны гольцового, подгольцового и лесного поясов всей горной области Азиатской Субарктики. На обширном пространстве от Корякского нагорья до Приполярного Урала включительно выявлены пространственные изменения авифауны в вертикальной и горизонтальной плоскости. Исследованы закономерности динамики территориального распределения видов и специфических орнитокомплексов в основных высотно-ландшафтных поясах гор Севера Азии. Выявлены факторы пространственной дифференциации, а также степень их воздействия на размещение видов, пространственно-временную и количественную характеристики населения птиц. Впервые крупнейший горный массив Заполярья - плато Путорана используется как модельный регион для познания общих закономерностей формирования и динамики фауны и населения птиц горно-субарктических экосистем.
Теоретическое значение. Проведенные исследования вносят определенный вклад в развитие экологии животных и современных направлений зоогеографии, основанных на ландшафтных методах изучения фауны и населения животных. Важное теоретическое значение имеют выявленные закономерности географического распространения и биотопического размещения птиц, особенности их экологии, выводы о тесной взаимосвязи авифаун равнин и гор Азиатской Субарктики, высотно-ландшафтной основе формирования авифауны этой обширной области суши. Выдвинутые предположения позволяют по новому интерпретировать механизмы, последовательность формирования и пути развития современного биологического разнообразия перигляциальных ландшафтов обширных горных областей Северной Евразии. Итоги представленной работы могут служить основой дальнейших исследований, «стартовой индикационной площадкой» долговременного мониторинга изменений горно-субарктической авифауны.
Практическое значение. Результаты исследований послужили обоснованием межевания земель и корректировки существующих границ ГПЗ «Путоранский», расширения его территории за счет кластерных участков на Таймыре, включения данного ООПТ в Список Всемирного Природного Наследия ЮНЕСКО. Разработаны и предложены к реализации - комплексная программа многолетнего мониторинга за состоянием биологического разнообразия в ГАС, программа мониторинга популяций орлана-белохвоста и пискульки на севере Красноярского края. Данные мониторинга за состоянием популяций редких и исчезающих видов птиц ежегодно используются в процессе ведения Красной книги России и Красной книги Красноярского края. В рамках реализации региональной экологической программы Таймырского района Красноярского края были подготовлены и опубликованы научно-популярные фотокниги «Плато Путорана – жемчужина таймырского заполярья» (2006, 2008), «Плато Путорана – горная страна полярных стихий» (2008).
Апробация работы и публикации. Основные положения и результаты исследований были представлены на научных форумах и научно-практических семинарах различного ранга: орнитологической конференции «Взаимодействие организмов в тундровых экосистемах» (Сыктывкар, 1989), конференции «Орнитологические проблемы Сибири» (Барнаул, 1991), Х Всесоюзной орнитологической конференции (Витебск, 1991), Международном Арктическом Симпозиуме им. Виллема Баренца (Москва, 1998); XI и XII Международных орнитологических конференциях Северной Евразии (Казань, 2001; Ставрополь, 2006), Международной орнитологической конференции «Waterbirdsaroundtheworld» (Эдинбург, Великобритания, 2004), Международных симпозиумах «Современное состояние популяций, управление ресурсами и охрана гусеобразных птиц Северной Евразии» (Олонец, 2003; Санкт-Петербург, 2005), Международном зоологическом конгрессе (Пекин, Китай, 2004), Сибирской зоологической конференции (Новосибирск, 2004), Международных семинарах «Основные направления изучения и охраны пискульки» (Хельсинки, Финляндия, 2005; Москва, 2007), Научно-практической конференции «Сохранение разнообразия животных и охотничье хозяйство России» (Москва, 2005), Международном научно-практическом семинаре «Экологические проекты крупнейших промышленных корпораций России» (Всемирный Банк Реконструкции и Развития, Московский офис, 2005), орнитологической конференции «Экология врановых в естественных и антропогенных ландшафтах» (Ставрополь, 2007), IV Международной орнитологической конференции «Современные проблемы орнитологии Сибири и Центральной Азии» (Улан-Удэ, 2009), Международной научно-практической конференции «Комплексное управление экосистемами в Российской Арктике: проблемы и перспективы» (Москва, 2009), научно-практическом семинаре РАН «Спутниковая система определения местоположения и сбора данных системы ARGOS ее применение для решения зоологических и экологических задач» (Москва, 2009).
Основные положения и материалы исследований по теме диссертации опубликованы в 59 работах общим объемом 89 печатных листов. В число указанных работ входят 2 персональные монографии объемом 27,7 печатных листов.
Структура и объем диссертации. Диссертация состоит из введения, 6 глав, выводов и приложения. Основной текст занимает 401 страницу машинописного текста. Список использованной литературы включает 524 наименования на русском и иностранных языках. Работа иллюстрирована 13 рисунками, содержит 99 таблиц, 18 из которых вынесены в приложение.
Глава 1 Районы и методы исследований, объем выполненных работ
1.1 Районы и сроки полевых исследований
Объекты наших исследований - авифауны горных систем Северной Азии, лежащие в пределах Субарктики: Корякского и Колымского нагорьев, гор Якутии (хребты Верхоянский, Черского, Кулар, Полуосный), Анабарского плато, плато Путорана, Приполярного и Полярного Урала. Понятие Субарктики мы принимаем в трактовке широко распространенной у зоологов, ботаников и физико-географов (Юрцев, 1966; Чернов, 1978; Кищинский, 1988; Голубчиков, 1996; Куваев, 2006), и определяемой как тип физико-географической среды, территориально соответствующий подзоне южных тундр, лесотундре и северным окраинам северотаежной подзоны.
В основу большинства разделов настоящей работы положены итоги 20-летних исследований, проведенных нами на плато Путорана в период 1988–2008 гг. и изложенных в персональных монографиях (Романов, 1996, 2003). В пределах плато Путорана выражены все основные особенности физико-географической среды, характерные для всех гор Азиатской Субарктики, что позволяет рассматривать его как модельный регион для выявления и познания общих закономерностей формирования и динамики авифауны в горно-субарктических условиях. Более объективному анализу закономерностей обсуждаемых в нашей работе также способствовали собственные многочисленные орнитологические исследования на Полярном Урале и других регионах Арктики и Субарктики от Кольского полуострова до Чукотки.
1.2.Методы исследований, объем и количественные показатели выполненных работ
Территориально арена наших исследований на плато Путорана составила 284000 км2
; непосредственно обследовано около 26000 км2
.
Основой для характеристики состояния и пространственно-временной динамики горно-субарктической авифауны и населения птиц на современном этапе их формирования стали количественные учеты птиц в модельном регионе – плато Путорана.
В гнездовой период и период послегнездовых кочевок обследование велось преимущественно маршрутным методом. В процессе линейного перемещения по территории через день-два делались остановки (на 2–7 суток) для стационарного обследования местности радиальными пешими маршрутами. Одноразовые переходы между остановками составляли 15-65 км, радиус обследования вокруг каждой многодневной стоянки – 15-50 км. Во время весенних и осенних миграций предпочтение отдавалось стационарным исследованиям. В период сезонных миграций проведено более 800 часов круглосуточных наблюдений за птицами. Большая часть сухопутных учетов (73% суммарной длины) повторялась 2-5 раз, меньшая (27%) проведена однократно. Учеты птиц и вычисление показателей обилия (ос/км²) для каждого вида выполнялись по методике Ю.С. Равкина (1967). Регистрировали всех птиц визуально и по голосу. На маршрутах учитывали всех встреченных птиц независимо от статуса их пребывания. За пару принимались: встреченные вместе территориальные самец и самка, территориальный поющий самец, птица с кормом, птица с отвлекающими демонстрациями, жилое гнездо, семейная группа слетков, не распавшийся выводок. Численность водных и околоводных видов определялась прямыми подсчетами птиц на акваториях озер и руслах рек, а также на береговой полосе, с последующим пересчетом количества особей на 1 км береговой линии.
Суммарная протяженность учетных маршрутов на плато Путорана составила 8857 км, из них 4063 км – в лесных ландшафтах лесного пояса, 2553 км – в береговой полосе рек и озер лесного пояса, 238 км – в горных редколесьях подгольцового пояса, 587 км в горных тундрах гольцового пояса. Общая протяженность лодочных маршрутов составила 1616 км. Максимальная высота на которой проводились исследования – 1450 м н.у.м.. Высоту местности определяли по приборам глобального позиционирования (GPS), а длину пройденных маршрутов – по крупномасштабным картам и показаниям шагомера. Суммарная протяженность учетных маршрутов в других регионах Евразийской Арктики и Субарктики составила 760 км.
В наших аналитических обобщениях по населению птиц мы использовали однолетние данные, собранные в разные годы, поскольку невозможно обследовать обширные труднодоступные территории за один сезон. Корректность такого подхода для зоогеографических исследований была фактически подтверждена целым рядом работ, в том числе проведенных в таких обширных регионах как, например, Алтай, Восточно-Европейская и Западно-Сибирская равнины (Равкин, 1984; Равкин, Равкин, 2005; Цыбулин, 2009).
Базовый информационный блок, послуживший отправной точкой для дальнейших обобщений, был сформирован как на основании наших данных, так и с привлечением большого числа опубликованных материалов других исследователей. Наиболее значимые работы из числа привлеченных: по Корякскому и Колымскому нагорьям (Кищинский, 1968, 1974, 1977, 1980, 1988), по хребтам Верхоянскому и Черского (Воробьев, 1963; Кищинский, 1988; Борисов и др., 1996, 2007), хребтам Кулар и Полуосный Сыроечковский-мл. и др., 1996), по Анабарскому плато (Бабенко, 2007; Поспелов, 2007), по Приполярному и Полярному Уралу (Естафьев, 1977, 1981; Морозов, 1989; Рябицев, 2001; Селиванова 2002, 2008; Селиванова, Естафьев, 2004; Головатин, Пасхальный, 2002, 2003, 2004, 2005). Для репрезентативного и унифицированного обобщения, анализа и интерпретации были отобраны только сопоставимые данные по гнездовой авифауне тех ГАС или их обширных областей, в пределах которых четко выражены гольцовый, подгольцовый и лесной высотно-ландшафтные пояса.
Достоверность гнездования признавалась в соответствии с критериями, рекомендованными Комитетом Европейского Орнитологического Атласа – EOAC (TheEBCCAtlas…, 1997). Гнездование считалось доказанным при его подтверждении фактическими материалами (находки гнезд, яиц, выводков, встречи птиц с кормом для птенцов), вероятным – при достаточно высокой численности птиц, демонстрирующих элементы гнездового поведения (токование, спаривание, беспокойство у гнезд), возможным – при летнем пребывании птиц на постоянных участках в подходящих для гнездования условиях.
Фауна гнездящихся птиц характеризуется по типам фаун (Штегман, 1938) и в свете современных представлений о географо-генетических группах птиц (Чернов, 1976, 1980; Кищинский, 1977, 1988).
Статистическую обработку материалов осуществляли с применением методов вариационной статистики (Лакин, 1973, 1990), в том числе - программ С.П. Харитонова (1999) part.exe и part_fix.exe «Cравнение выбора долей по методу критерия Стьюдента для выборочных долей».
Сходство авифаун сравниваемых горных систем определялось по коэффициенту фаунистической общности (КФО), рассчитывавшемуся по формулам:
Серенсена
КФО = 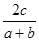 100% 100%
и Жаккара
КФО = 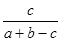 100% 100%
(Песенко, 1982; Чернов, 1975, 2008). В обеих формулах - а
и в
- число видов в каждой из двух фаун, с
- количество видов, общих для двух фаун.
Для выявления провинциальных отличий в населении птиц разных участков модельного горного региона (плато Путорана) был использован коэффициент сходства населения (КСН), рассчитывавшийся по формуле:
КСН = 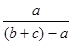 100% (Наумов, 1964), 100% (Наумов, 1964),
где а
— сумма наименьших (из двух) показателей обилия видов, общих для обоих сравниваемых районов, в
и с
— общее обилие птиц первого и второго районов.
В номенклатуре и при составлении списков птиц мы следовали Л.С. Степаняну (1990, 2003). Названия некоторых видов приняты по Списку птиц Российской Федерации (Коблик и др., 2006).
1.3 Основные понятия и термины
В работе дано определение используемых специфических или допускающих неоднозначную трактовку терминов и понятий.
Глава 2 Краткая история исследований авифауны гор Азиатской Субарктики
Авифаунистическая изученность различных горных систем Азиатской Субарктики остается весьма различной: от скрупулезно и систематически обследовавшихся (плато Путорана, Приполярный и Полярный Урал) до почти не изученных (хребет Черского, внутренние области Верхоянского хребта и Колымского нагорья). Тем не менее, к настоящему времени мы располагаем достаточным объемом знаний для составления общего целостного представления об авифауне гор Азиатской Субарктики и теоретического осмысления процессов и закономерностей, формирующих ее.
Глава 3 Особенности среды обитания и специфика формирования пространственной дифференциации фауны и населения птиц гор Азиатской Субарктики
3.1 Общие физико-географические особенности гор Азиатской Субарктики как среды обитания птиц
ГАС расположены на севере Евразии приблизительно в одном широтном диапазоне (60º00'–70º00' с.ш.). Занимаемая ими суммарная площадь около 1190000 км2
, что составляет 7 % от площади России. ГАС имеют сходные природные условия (Пармузин, 1979; Атлас СССР, 1983; Гвоздецкий, Голубчиков, 1987; Голубчиков, 1996). Следствие этого - достаточно однородная и весьма суровая среда обитания птиц: господство низко- и среднегорных форм рельефа (1000-3000 м н.у.м.), континентальный или резко континентальный типы субарктического (реже умеренного) климата, суровые морозные и долгие зимы, обильная и разветвленная гидросеть, образование наледей на реках, почти повсеместное распространение многолетнемерзлых грунтов. Все рассматриваемые регионы или их части в плейстоцене были центрами оледенения, и в силу горного рельефа освободились от ледников позднее сопредельных равнин (Кинд, 1965, 1969, 1976; Котляков, 1968; Авенариус и др., 1978; Голубчиков, 1996; Соломина, 1999). Поэтому в облике современных ландшафтов ГАС прослеживается перигляциальный генезис (Вангейм, 1976; Матюшкин, 1976; Верещагин, Громов, 1977; Верещагин, Барышников, 1985), а авифауна (особенно гольцовая) вероятно моложе, чем авифауна равнинной Субарктики, и становление ее продолжается. В горах Азиатской Субарктики сменяются лесной (горно-северотаежный), подгольцовый (горные редколесья и кустарники) и гольцовый (горно-тундровый) высотно-ландшафтные пояса (Норин, 1995; Голубчиков, 1996; Куваев, 2006).
3.2 Формирование авифауны в ледниковых ландшафтах гольцов
Реконструкция развития ледников в конце плейстоцена – начале голоцена (Cоломина, 1999), и выявленные нами современные экологические связи и пространственное распределение птиц позволили выдвинуть гипотезу о том, что ледниковые геоморфологические структуры, образованные поздне-плейстоценовыми ледниками, создавая экологические предпосылки обогащения гольцовой авифауны «негорным» комплексом видов, вероятно оказывают непосредственное влияние на ее формирование и пространственную неоднородность в течение всего постледникового периода (Романов, 2009). Это подтверждает статистически достоверная пространственная сегрегация сообществ птиц, формирующихся на «гольцовых плакорах» с одной стороны, и в ледниковых долинах – с другой. Пространственная дифференциация арктоальпийских (альпийских) видов и равнинно-тундровых вселенцев, обусловленная литогенной неоднородностью поверхности, согласуется с историей их формирования и различиями в стратегии освоения гольцов. Арктоальпийские (тундряная куропатка, хрустан, рогатый жаворонок, пуночка) и альпийские (американский конек) виды, а также - обыкновенная каменка, осваивали горные вершины благодаря уже выработавшимся у них специальным адаптациям к экстремальным условиям внешней среды еще в период оледенения (Кищинский, 1988), и вероятно представляют более древний элемент гольцовой авифауны. Позднее, когда ледники начали разрушаться и вскоре исчезли, все виды птиц гольцового комплекса полностью или почти полностью сохранили преимущественно горно-вершинный характер своего распространения. Активного их перемещения в освободившиеся ледниковые долины, расположенные на меньших высотах, не произошло. Подтверждением этому служит тот факт, что в настоящее время почти все территориальные пары этих видов мы находим в верхних или внедолинных частях гольцов без явных признаков последнего оледенения (Романов, 2009). Равнинно-тундровые вселенцы, не имея столь глубоких адаптаций, при проникновении в горы использовали традиционные для себя экологические условия, сформировавшиеся на локальных участках перигляциальных ландшафтов, связанных с деятельностью ледников. Они расширили видовое богатство этой авифауны позднее, проникая на вершины гор по участкам былого расположения ледников по мере их интенсивного таяния.
В ледниковых долинах после разрушения ледников экологические условия оказались мягче, разнообразнее, и в целом ближе к зональной тундре, чем к суровым горно-субарктическим условиям «гольцовых плакоров» (Норин, Китсинг, 1982). В современную эпоху с поздне-плейстоценовыми ледниковыми долинами, занимающими, например, на плато Путорана не более 25-30% площади горных вершин, связано распространение более 2/3 всего видового состава авифауны этих районов (в т.ч. почти 100% равнинно-тундровых вселенцев), тогда как с более обширными пространствами гольцовых водоразделов (70-75% площади горных вершин) связано не более 1/4 всех видов (в т.ч. почти 100% арктоальпийских и альпийских видов). Суммарные показатели обилия птиц в ледниковых долинах всегда в 2-4 раза выше, чем на водоразделах (Романов, 2009). Возможно, это связано также с тем, что ледники могли быть источником повышенной продуктивности сопредельных экосистем. На это указывают некоторые современные исследования, например на Аляске (Hoodetal., 2009).
Равнинные водные и околоводные виды заселяют неглубокие запрудные ледниковые озера, равнинно-тундровые - хорошо дренированные склоны блюдцеобразных озерных котловин, склоны и поверхности боковых и конечных морен, лугово-кустарниковые – влажные долинки речек, сток которых зарегулирован конечной мореной. На ледниковых озерах гнездятся чернозобая гагара, чирок-свистунок, морянка, большой и средний крохали, серебристая и сизая чайки, полярная крачка. На пляжах из мелкой гальки и на песчано-илистых отмелях по берегам ледниковых озер встречаются галстучник, белохвостый песочник, кулик-воробей и сибирский пепельный улит. В тундрах, покрывающих склоны и днища ледниковых долин, распространены золотистая ржанка, краснозобый конек, весничка, варакушка, полярная овсянка, лапландский подорожник. Связь обитателей равнин с ледниковыми формами рельефа особенно четко прослеживается при выборе мест устройства гнезд. Белохвостый песочник, лапландский подорожник, сибирский пепельный улит связаны с каменистыми грядами конечных морен. На склонах боковых морен, имеющих вид сухих щебнистых бугров, гнездятся морянки и золотистые ржанки, а на их вершинах – полярные крачки.
На конусообразных каменных буграх, камах, успешно гнездятся морянки, золотистые ржанки, сизая чайка. Гнездовые местообитания краснозобого конька, веснички, варакушки и полярной овсянки в пределах ледниковых долин тесно связаны с заболоченными участками широких долинок многочисленных речек с мохово-осоковым кочкарником и зарослями низкорослого ивнякас осоковой кочкарней и зарослями низкорослого ивняка. Лапландские подорожники в ледниковых долинах местами заселяют кочкарные тундры по подножию пологих склонов сильно разрушенных боковых морен. Из равнинных птиц за пределы ледниковых долин очень редко выходят лишь адаптированные к условиям плакоров зональной тундры золотистая ржанка, краснозобый конек и лапландский подорожник.
Наибольшую роль в формировании неоднородности фауны и населения птиц горных вершин Азиатской Субарктики, в прошлом подвергавшимся оледенению, играют геоморфологические структуры ледникового генезиса, механический состав грунтов и гидрологические условия (в частности – дренаж поверхности). Учитывая, что эти факторы в совокупности определяют многие особенности структуры растительного покрова, а мезоклиматические различия в пределах гольцового пояса обусловлены особенностями рельефа, в качестве главного фактора пространственной дифференциации авифауны гольцов следует признать литогенную основу, с которой связаны и гидрологические условия. Литологические условия определяют также основные закономерности пространственной дифференциации фауны и населения птиц в других регионах с экстремальной внешней средой, например – на арктическом о.Врангеля (Стишов, 1985, 1989). Таким образом, можно предполагать ведущее значение литогенной основы в формировании авифауны и дифференциации населения птиц однородных в зональном (Чернов, 1971) или высотно-поясном (Романов, 2009) отношении территорий, находящихся в экстремальных условиях. В пределах этих территорий средообразующая роль растительности снижается и усиливается воздействие на органический мир абиогенных факторов.
3.3 Формирование сообществ птиц в районах образования долинных наледей
Одна из важнейших особенностей ледового режима рек гор Азиатской Субарктики – широкое распространение долинных наледей (Колосов, 1938; Котляков, 1968; Голубчиков, 1996). Процесс формирования сообществ птиц на наледных полях, в определенной мере, моделирует общие закономерности экологического пути освоения птицами перигляциальных пространств – участков суши, освобождающихся от ледовых образований или находящихся в непосредственной близости от их границ. В районах образования крупных наледей происходит существенная трансформация ландшафта (Фотиев, 1962), оказывающая существенное влияние на формирование специфических сообществ птиц. Параметры многолетней наледи не постоянны во времени (Толстихин, 1965, 1966). На каком то этапе, несмотря на ежегодное образование зимой, она начинает фенологически раньше разрушаться, а затем полностью стаивать за лето, активизируются процессы зарастания и смены сукцессий на освобождающихся ото льда грунтах (Работнов, 1937). Сокращение наледей и интенсивное зарастание наледных полей стало особенно заметно в условиях потепления климата (Материалы отчетов Красноярского краевого и Норильского городского туристических клубов, 1959-2009). У различных наледей эти процессы не синхронны, что позволяет выявить закономерности формирования авифауны наледных полей на более ранних и более поздних стадиях их существования. На плато Путорана нами выявлено 3 этапа формирования сообществ птиц, связанных с многолетним сокращением летних размеров наледей и хронологически соответствующих 3 этапам постепенного зарастания наледных полей (Работнов, 1937).
Пути и скорость их зарастания определяют темп и характер формирования сообществ птиц (Романов, 2009). Наледные поля на пионерном этапе зарастания осваиваются только ржанкообразными (100%), на втором этапе – преимущественно ржанкообразными (75%), и лишь на тертьем этапе – преимущественно воробьинообразными (80%). Видовое разнообразие в процессе формирования авифауны наледных полей возрастает в геометрической прогрессии. Среди первых вселенцев (n=2) - только гемиаркты, связанные в своем современном распространении с экологически экстремальными высокоширотными областями. На 2-м этапе зарастания, когда сомкнутость растительного покрова приближается к 90%, вселенцы (n=4) представлены гипоарктическими и бореально-гипоарктическими видами, связанными с экологически менее экстремальными ландшафтами. На 3-м этапе зарастания вслед за более существенным увеличением разнообразия экологических условий, сообщества птиц наледных полей пополняются 10 видами из числа гипоарктических, бореально-гипоарктических, арктоальпийских и широкораспространенных видов (Романов, 2009). На 3-м этапе зарастания, авифауна наледных полей насчитывает 16 видов (галстучник, сибирский пепельный улит, мородунка, белохвостый песочник, фифи, средний кроншнеп, полярная крачка, краснозобый конек, желтоголовая и белая трясогузка, весничка, варакушка, черноголовый чекан, обыкновенная каменка, полярная овсянка, овсянка-крошка). Поливидовые сообщества птиц тундроподобных ландшафтов наледных полей формируются видами из числа характерных представителей региональной авифауны. В нашем примере – авифауны плато Путорана. Экологические особенности тех или иных биотопов наледных полей определяет весьма своеобразное сочетание вобранных видов птиц (Романов, 2009). В основном, оно представлено околоводными северными видами, и широкораспространенными видами, связанными с азональными зарослями кустарников и берегами водоемов. Структура сообществ птиц наледных полей имеет определенное сходство со структурой сообществ тундровой зоны. В их состав входят гемиарктические виды — 2 (12%), гипоарктические — 5 (31%), бореально-гипоарктические — 5 (31%), широкораспространенные - 3 (20%), арктоальпийские — 1 (6%) виды. Плотность гнездового населения птиц тундроподобных местообитаний в районах зимнего образования мощных наледей невелико – 69 ос/км2
(Романов, 2009).
3.4 Гидрологические факторы, определяющие формирование авифауны гор Азиатской Субарктики
Уровень воды в ранне-весенний период,во многом зависящий от степени континентальности климата, в немалой степени определяет географию миграционных путей птиц в пределах обширных горных стран Азиатской Субарктики. Так, основной пролет водных и околоводных птиц на плато Путорана идет вдоль наиболее полноводных рек запада региона (Романов, 2005, 2006). В его центральной части, где реки ранней весной менее полноводны, пролет не столь интенсивен (Романов, 1996). Русла рек востока Путорана, за зиму полностью иссушенные, заполняются водой лишь после начала интенсивного таяния снега в горах. Поздний и очень бурный подъем уровня воды делает подобные водотоки абсолютно недоступными для птиц, из-за чего пролет птиц на востоке Путорана практически не наблюдается.
Динамика уровня воды в гнездовой период определяет в ГАС характер устройства гнезд, начало откладки яиц и успех размножения многих околоводных видов птиц (Морозов, 1984; Романов, 1996, Романов и др., 2007). Например, у галстучника откладка яиц начинается после прекращения подъема уровня воды в озерах, в период его стабилизации и дальнейшего плавного понижения. Чем раньше пара начинает гнездование, тем выше место для гнезда она выбирает. Откладка яиц может начаться и раньше окончания весеннего половодья, если пара занимает гнездовой участок на вершине очень высокого берега, который никогда не затапливается (Кречмар, 1966; Романов, 1996, 2006, Романов и др., 2007). В ГАС обычны высокие летние паводки (Гвоздецкий, Михайлов, 1987; Голубчиков, 1996). Перекрывая доступ водным и околоводным птицам к и без того малочисленным гнездовым биотопам, они крайне негативно влияют на успех размножения, и соответственно лимитируют их численность (Романов, 2004).
Влияние охлаждающего эффекта водоемов на формирование природно-территориальных комплексов известно в отношении влияния морей северной части Тихого океана, имеющих «полярный» гидрологический режим: ареалы многих видов птиц, зонально ориентированные в Азиатской Субарктике по широте, в горных районах ее северо-восточной части характерной ступенью «опускаются» на юг в меридиональном направлении. По узкой полосе североохотской прибрежной лесотундры на юг до 59º с.ш., до Тауйской губы и залива Бабушкина, проникают виды арктической фауны: белоклювая гагара, морянка, зимняк, чернозобик, белохвостый песочник, круглоносый плавунчик, лапландский подорожник, краснозобый конек (Кищинский, 1968). Таким образом, формируются «инверсионные» гипоарктические сообщества птиц в пределах бореальной зоны умеренных широт. Этот феномен – характерная зоогеографическая особенность северо-востока Азии (Кищинский, 1988; Лобков, 2003) и Колымского нагорья, в частности (Кищинский, 1968, 1988).
В некоторых ГАС авифауна формируется в условиях охлаждающего эффекта крупных внутренних водоемов. Например, озера плато Путорана занимают первое место по объему водной массы в пределах Субарктики, и образуют второй по объему поверхностный резервуар пресной воды в России после Байкала (Пармузин, 1964, 1975). Сильный охлаждающий эффект путоранских озер и отсутствие на прибрежных пространствах развитой древесно-кустарниковой растительности (Водопьянова, 1976) обуславливает низкое разнообразие биоты приозерий, в частности бедность видового состава гнездовой авифауны (n=27) и низкие показатели плотности населения птиц (50-170 ос/км²). Сообщества птиц, формирующиеся в специфичных экологических условиях приозерий, в целом имеют хорошо выраженный гипоарктический облик (Романов, 1996). «Очаги» гипоарктической авифауны формируются значительно южнее областей ее господства - в пределах северной тайги. К специфическим видам авифауны приозерий, очень редко встречающихся в других местообитаниях лесного пояса, следует отнести гипоарктов – щеголя, белохвостого песочника, среднего кроншнепа, краснозобого конька, полярную овсянку, гемиарктов – морянку и малого веретенника, бореало-гипоарктов – золотистую ржанку, мородунку, турухтана, гаршнепа, желтоголовую трясогузку (Романов, 1996).
Глава 4
Общие закономерности высотно-ландшафтной дифференциации авифауны гор Азиатской Субарктики
В соответствии с господствующим типом высотной поясности, в ГАС дифференцированы сменяющие друг друга с высотой авифауны лесного, подгольцового и гольцового поясов. Закономерности формирования авифауны в пределах каждого из поясов имеют либо общий характер и сходное проявление на всем протяжении от Урала до Корякского нагорья, либо демонстрируют определенный вектор их пространственных изменений в пределах Азиатского континента.
Равнинные элементы формируют основу авифауны каждого пояса, качественно сближают ее с авифаунами смежных высотных поясов и ландшафтов равнинных предгорий. Авифауна лесного пояса гор Азиатской Субарктики, в отличие от гор юга Палеарктики, достаточно однородна. Она состоит из видов широко распространенных на севере бореальной зоны и одновременно участвующих в формировании лесной авифауны не менее 4 из 6 горных стран Азиатской Субарктики. Из 185 видов лесной авифауны ГАС, 96 (52%) таких видов более или менее повсеместно формируют авифауну лесного пояса субарктических гор на всем протяжении от Урала до Корякского нагорья.
Доминирующая закономерность формирования подгольцовой авифаунывсей цепи ГАС связана со встречным взаимовлиянием северо-таежной авифауны нижней части горных склонов и гольцовой авифауны горных вершин. В основном - за счет проникновения многих обитателей лесного пояса выше границы распространения древесной растительности. Благодаря этому особенности подгольцовой авифауны каждой горной страны в значительной степени определяются особенностями местной лесной авифауны. Из 103 видов подгольцовой авифауны ГАС, лишь 26 из них (25%) формируют ее более или менее повсеместно. Большая их часть (75%) входит в состав подгольцовых авифаун 1-2, реже 3 горных систем.
Авифауна гольцового, подгольцового и лесного поясов гор Азиатской Субарктики, на современном этапе их формирования, имеют вполне определенные специфичные черты за счет ряда видов, экологически тесно связанных с горными ландшафтами.
Ядро гольцовой авифауны почти всех ГАС формируют арктоальпийские (тундряная куропатка, хрустан, рогатый жаворонок, пуночка) и альпийские (американский конек) виды, а также - обыкновенная каменка. Горный характер авифауны гольцов ГАС (за исключением Урала) определяют также горные формы - монгольский зуек, горный дупель, альпийская завирушка, сибирский вьюрок, и виды, связанные в своем генезисе с Азиатской Субарктикой (длиннопалый песочник (Леонович, 1973; Томкович, 1980), в частности горными ее районами (сибирский пепельный улит (Воробьев, 1963; Андреев, 1980; Кищинский, 1988; Романов, 2008), большой песочник (Tomkovich, 1997; Томкович, 2001). Все эти виды формируют своеобразный горный комплекс, представленный в наиболее полной форме в восточной половине цепи ГАС от Корякского нагорья до Верхоянского хребта. Западнее, на плато Путорана, обнаружить его в более или менее «целостном виде» уже не удается. Здесь повсеместно обычен лишь сибирский пепельный улит. Альпийские виды приобретают роль одной из ведущих географо-генетических групп в гольцовых авифаунах восточных регионов Азиатской Субарктики (от Корякского нагорья до Верхоянского хребта), где они представлены максимально, как по числу видов (n=5-6), так и по доле участия (11-32%).
Горную специфику подгольцовой авифауне ГАС придают сибирский пепельный улит, кроншнеп-малютка, кедровка (N
.
c
.
kamtschatkensis
Barr.-Ham.), щур (P
.
e
.
kamtschatkensis
(Dyb.), авифауне лесного пояса - каменушка, длинноносый и большой крохали, сибирский пепельный улит, кроншнеп-малютка, горная трясогузка, оляпка и бурая оляпка. К горному комплексу близки синехвостка и сибирская чечевица, на что указывают некоторые особенности их экологии, например, преимущественно горно-лесное распространение на значительной части гнездового ареала. Сокращение числа горных видов в региональных авифаунах с востока на запад следует признать объективной закономерностью пространственного формирования авифауны в пределах всей цепи гор Азиатской Субарктики.
Глава 5 Закономерности формирования и динамики пространственно-временной структуры населения птиц гор Азиатской Субарктики
5.1 Общие закономерности формирования структуры гнездового населения птиц в условиях высотной поясности гор Азиатской Субарктики
В ГАС с высотой поступательно сокращаются видовое богатство, плотность населения птиц, обилие абсолютного большинства видов (табл. 1).
Таблица 1. Плотность населения (ос/км²) птиц в различных высотных поясах гор Азиатской Субарктики
| № |
Высотно-ландшафтный пояс |
Приполярный и Полярный Урал |
Плато Путорана |
Анабарское плато |
Горы Якутии |
Колымское нагорье |
Корякское нагорье |
Всего в ГАС |
| 1 |
Гольцовый |
149 |
47 |
42 |
− |
14 |
− |
63 |
| 2 |
Подгольцовый |
194 |
164 |
147 |
− |
55 |
− |
140 |
| 3 |
Лесной |
420 |
405 |
185 |
− |
145 |
− |
289 |
Примечание: для составления таблицы использованы данные по Уралу (Естафьев, 1977, 1981; Селиванова 2002, 2008), плато Путорана (Романов, 1996, 2003, 2006; Романов и др., 2007), Анабарскому плато (А.Е. Волков, личное сообщение; Бабенко, 2007; Поспелов, 2007), Колымскому нагорью (Кищинский, 1968, 1988).
Значительные отличия между населением птиц 3 высотных поясов ГАС, выявленные с применением коэффициента сходства населения - КСН (Наумов, 1964), указывают на высокую степень автономности процесса его формирования в пределах каждого из поясов. Например, на плато Путорана уровень сходства населения лесного и подгольцового поясов не превышает 29%, подгольцового и гольцового – 18%, а лесного и гольцового – составляет всего 2%. С подъемом на каждые 100 м плотность населения птиц сокращается в среднем (n=4) – на 28 ос/км². В большинстве горных регионов основное сокращение плотности населения птиц происходит при переходе из подгольцового пояса в гольцовый, и чуть менее существенное - при переходе из лесного пояса в подгольцовый. Имеются и отличия в выявленной общей закономерности. Например, на Анабаре повышенная общая суровость климата и очень пологие склоны невысоких поднятий (более напоминающих увалы и низкие сопки, нежели настоящие горы) существенно нивелируют, а местами даже сводят на нет различия экологических условий лесного и подгольцового поясов. Поэтому в силу очень плавного и постепенного перехода между ними смена сообществ и интенсивность сокращения плотности населения птиц на Анабарском плато весьма незначительна.
На плато Путорана в рамках снижения от подножий к вершинам общей плотности населения птиц, выявлены 4 группы видов, имеющие различные векторы вертикальной динамики обилия. 1 – поступательно сокращающие обилие с высотой (дербник, белая куропатка, горная и белая трясогузки, таловка, бурый дрозд, обыкновенная чечетка, овсянка-крошка). 2 – поступательно увеличивающие обилие с высотой (обыкновенная каменка). 3 – сохраняющие стабильное обилие в лесном и подгольцовом поясах и сокращающие его лишь в гольцах (азиатский бекас, желтоголовая трясогузка, весничка). 4 – имеющие максимальное обилие в подгольцах и сокращающие его в сторону вершин и подножий склонов (сибирский пепельный улит, обыкновенная кукушка, болотная сова, краснозобый конек, варакушка, сибирская чечевица, полярная овсянка). В силу региональной специфики экологических условий в разных горах Азиатской Субарктики направления пространственного изменения обилия одних и тех же видов в них могут не совпадать. Например, в Колымском нагорье таловка и обыкновенная чечетка достигают максимального обилия не в лесном поясе (как на плато Путорана), а в подгольцовом (Кищинский, 1968). Наиболее крайние формы таких тенденций могут проявляться в том, что некоторые виды, обитающие на большей части ГАС преимущественно (или в том числе) в лесном поясе, на востоке «покидают» пределы лесного пояса и населяют только вышележащий подгольцовй пояс. Таковы, например, азиатский бекас в Корякском нагорье или варакушка в Колымском нагорье (Кищинский, 1988).
В пределах цепи гор Азиатской Субарктики плотность населения птиц на уровне всех высотно-ландшафтных поясов сокращается в направлении с запада на восток. Это согласуется с сокращением в этом же направлении общей продуктивности растительного покрова (Национальная стратегия и план действий по сохранению биоразнообразия России; http://www.sci.aha.ru/biodiv/npd/ind1.htm). Корреляция сокращения плотности населения птиц с долготой близка к достоверности для показателей плотности населения в пределах гольцового, подгольцового и лесного поясов (табл. 2).
Таблица 2. Достоверность корреляции между долготой и плотностью населения птиц
| Высотно-ландшафтный пояс |
n
|
R
(коэффициент корреляции между долготой и плотностью населения птиц) |
P
(достоверность коэффициента корреляции) |
| Гольцовый |
4 |
-0.89 |
0.10 |
| Подгольцовый |
4 |
-0.92 |
0.07 |
| Лесной |
4 |
-0.94 |
0.06 |
В 10 раз в восточном направлении сокращается плотность населения птиц гольцового пояса, и не столь существенно – подгольцового (в 3,5 раза) и лесного (в 3 раза) поясов.
Выявлен сходный состав лидирующих видов во многих горах Азиатской Субарктики, что свидетельствует в пользу не только общности ядра их авифаун, но и определенного единства экологических и зоогеографических закономерностей формирования основы населения птиц этих регионов. Пространственная преемственность населения птиц поддерживается в горизонтальной плоскости видами лидирующими по обилию одновременно в нескольких регионах ГАС, а в вертикальной – одновременно лидирующими в двух, обычно смежных высотно-ландшафтных поясах. В лесном поясе большей части цепи ГАС лидируют 7 видов, из которых 4 (таловка, вьюрок, обыкновенная чечетка, овсянка-крошка) являются общими. В населении птиц подгольцового пояса лидируют таловка и обыкновенная чечетка. В гольцах ГАС почти повсеместно лидирует обыкновенная каменка в различном сочетании еще с 5 видами, обычно – представителями рода Anthus
. В западной части цепи ГАС, на Урале это – луговой и краснозобый коньки (Естафьев, 1977, 1981; Головатин, Пасхальный, 2005), а восточнее – американский и краснозобый коньки (Кищинский, 1980; Романов, 1996, 2004). Широкое распространение и массовый характер пребывания разных видов коньков в верхних поясах гор севера Евразии, видимо, в значительной мере объясняется, с одной стороны, их экологической пластичностью, а с другой стороны, достаточно глубокими экологическими адаптациями некоторых видов этой группы к экстремальным условиям горных вершин (Ирисов, 1997; Ирисова, 2002).
5.2 Внутрирегиональная изменчивость плотности и структуры гнездового населения птиц в пределах отдельно взятой горной страны
Огромные размеры и сложная орография горных стран Азиатской Субарктики предопределяют внутрирегиональные (провинциальные) особенности населения птиц. В качестве модельного региона, где c помощью коэффициента сходства населения – КСН (Наумов, 1964) выявлены особенности внутрирегиональных различий в плотности и структуре населения птиц, выбрано плато Путорана.
Наиболее существенные внутрирегиональные отличия выявлены в населении птиц подгольцового пояса. В подгольцах самые низкие показатели минимального (3%) и максимального (49%) сходства населения. Общий уровень абсолютных значений КСН также заметно ниже, чем в гольцовом и лесном поясах: более чем в половине сравниваемых пар районов сходство населения птиц не превышает 20%; всего в 10 из 55 сравниваемых пар районов сходство населения несколько превышает 30%. Ярко выраженная провинциальность подгольцовых сообществ птиц по сравнению с гольцовыми или, тем более, лесными обусловлена неоднородными и менее стабильными экологическими условиями, а также почти полным отсутствием видов адаптированных к ним.
В пределах гольцового пояса провинциальные отличия населения птиц не столь контрастны, как в подгольцовом поясе. В гольцах, как и в подгольцах довольно значительна амплитуда между минимальным (8%) и максимальным (67%) показателем коэффициента сходства населения (КСН). Однако абсолютные их значения, равно как и общий уровень абсолютных значений КСН, выше, чем в подгольцовом: более чем в половине сравниваемых пар районов сходство населения птиц превышает 30%; всего в 9 из 67 сравниваемых пар районов сходство населения ниже 20%. Экологические условия в гольцовом поясе более экстремальны и одновременно более стабильны, чем в подгольцовом. В населении птиц гольцов значительна доля участия альпийских и арктоальпийских видов хорошо адаптированных к условиям субарктических горных вершин. Все это обеспечивает большую общую пространственно-временную устойчивость населения птиц гольцов.
В лесном поясе провинциальные отличия населения птиц выражены намного меньше, чем в гольцовом и подгольцовом поясах. Меньше амплитуда между минимальным (36%) и максимальным (67%) показателем коэффициента сходства населения (КСН). Абсолютные значения КСН, равно как и общий уровень абсолютных значений КСН, выше, чем в гольцовом и тем более в подгольцовом поясах. Более чем в половине сравниваемых пар районов сходство населения птиц превышает 50%. Лишь в 4 из 45 сравниваемых пар районов сходство населения ниже 40%. В целом, высокие абсолютные значения коэффициента сходства населения (КСН) характерны для всех сравниваемых пар районов. Это свидетельствует о достаточно стабильном и равноценном взаимовлиянии сообществ птиц даже взаимоудаленных районов на уровне лесного пояса. В гольцовом и подгольцовом поясах сообщества птиц этих же районов, при их попарном сравнении, дают гораздо больший разброс значений КСН.
Выявленные векторы изменения плотности населения и обилия большинства фоновых видов указывают на то, что их пространственная динамика в ГАС имеет концентрически центробежный характер. Эти показатели имеет тенденцию увеличения от наиболее высоких внутренних районов горных стран, где их величины минимальны, в сторону периферии (Воробьев, 1963; Кищинский, 1968, 1980; Естафьев, 1977; Селиванова, 2002, 2008; Романов, 1996, 2003, 2004; Головатин, Пасхальный, 2005; Борисов и др., 1996, 2007; Бабенко, 2007; Поспелов, 2007). Особенно четко эта закономерность проявляется в населении птиц самой нижней и самой верхней части высотного профиля: в лесном и гольцовом поясах.
Провинциальные отличия в пределах всех высотно-ландшафтных поясов характерны как в общих показателях населения птиц, так и в специфике пространственных изменений обилия отдельных видов, что подробно было рассмотрено на примере плато Путорана. Оказалось, что в лесном, подгольцовом и гольцовом поясах закономерности провинциальных отличий в плотности населения птиц весьма специфичны. В лесном поясе провинциальные отличия плотности населения определяются различным составом основных лесообразующих пород и бонитетом лесов в зависимости от географической широты местности и континентальности климата. Этим объясняется наличие двух равноценных трендов сокращения этого показателя – с юга (566-673 ос/км²) на север (295-309 ос/км²) и с запада (364-488 ос/км²) на восток (251 ос/км²). В подгольцовом поясе основным фактором, определяющим провинциальные отличия плотности населения в пределах отдельной горной системы, следует признать пространственную неравномерность развития полосы кустарников и стлаников, связанную, прежде всего, с уровнем континентальности климата. В подгольцах Путорана это подтверждается наличием лишь одного достоверно выявленного тренда сокращения плотности населения – с запада (196-388 ос/км²) на восток (71 ос/км²). Зависимости плотности населения от широты местности в пределах подгольцов Путорана не выявлено. В гольцовом поясе Путорана провинциальные отличия плотности населения определяются характером господствующих горно-тундровых типов растительности, связанных, в основном, с абсолютной высотой местности, чем объясняется наличие трендов увеличения этого показателя от наиболее высокой средней части плато (30 ос/км²) к более низким южным (43-46 ос/км²) и северным (102-115 ос/км²) окраинам. Повышенная плотность населения птиц в гольцах самых северных частях гор Азиатской Субарктики обусловлена не столько собственно географической широтой, сколько их непосредственным контактом с зональной тундрой и лесотундрой, достаточно плотно заселенными птицами, и в качестве «донора» поддерживающими достаточно высокий уровень обилия ряда видов в гольцах.
Сообщества птиц лесного пояса, по сравнению с гольцовыми и подгольцовыми, более разнообразны, стабильны и однородны в пространстве и времени. На это, кроме небольших провинциальных отличий выявленных по коэффициенту сходства населения (КСН), указывает минимальное превышение показателей видового богатства и плотности населения в различных районах лесного пояса, равномерное распределение по территории региона более 50% видового состава, несущественные провинциальные отличия в составе лидеров, основная часть которых встречается в этом качестве в лесном поясе почти повсеместно.
Провинциальные отличия населения, диагносцируемые с помощью КСН, неодинаковы на уровне каждого из рассматриваемых высотно-ландшафтных поясов. Характерна также высотно-поясная специфичность провинциальных различий основных параметров населения (видового состава, плотности, состава лидирующих и фоновых видов), а в ряде случаев и закономерностей определяющих их пространственные тренды. Это свидетельствует в пользу достаточно высокого уровня автономности формирования населения птиц горной Субарктики на разных высотах.
5.3 Закономерности пространственной изменчивости гнездового населения птиц гор Азиатской Субарктики в пределах однородного ландшафта
Во всех ГАС общие закономерности формирования пространственной изменчивости населения птиц в пределах однородного ландшафта сходны. В значительной мере они зависят от абиогенных факторов, в частности - литогенной основы (абсолютная высота местности, угол наклона поверхности, экспозиция склона, механический состав грунтов). Плотность населения птиц заметно возрастает в устьевых и пойменных местообитаниях - 808-1324 ос/км², на склонах южной экспозиции - 570-780 ос/км², ледниковых геоморфологических структурах - 79-200 ос/км²), и сокращается - на плоских участках межгорных долин - 236-260 ос/км², склонах северной экспозиции - 89 ос/км², горных водоразделах без следов деятельности поздне-плейстоценовых ледников – 8-30 ос/км² (Романов, 1996, 2003, 2009).
Локальное размещение многих видов птиц в горах Азиатской Субарктики поддерживается горно-котловиннным характером местности и усиливается склонностью целого ряда неколониальных видов образовывать «микро-ассоциации». Поливидовые ассоциации образуют 1-2 территориальные пары 3-5 видов (воробьинообразных) в одном небольшом участке, вокруг которого на значительном расстоянии (0,3-20 км) в пределах абсолютно сходных условий этих видов нет. Такие ассоциации образуют мелкие виды воробьинообразных, ни один из которых не выступает в роли покровителя по отношению к остальным. В случае опасности все участники оказываются одинаково уязвимы. Их объединяет компактное расположение наиболее подходящих для гнездования и кормодобывания местообитаний. Как показали наши исследования плато Путорана, подобного рода поливидовые ассоциации (n=447) встречаются более чем в 3 раза чаще, чем моновидовые (n=131). Разница в пользу поливидовых ассоциаций высоко достоверна (P=0.0001). В лесном поясе поливидовые «микро-ассоциации» обычно составляют сибирская завирушка, бурый дрозд, таловка, зарничка и вьюрок, в подгольцовом и гольцовом поясе – весничка, варакушка и полярная овсянка (Романов, 1996, 2003, 2004, 2006). Можно предположить, что причина преимущественного формирования поливидовых ассоциацией заключается в существовании определенной самоорганизации птичьих сообществ – тенденции поддерживать экологически, а затем, видимо, эволюционно закрепленную плотность населения, определенное сочетание и интенсивность взаимодействия между разными видами даже при минимальном количестве особей в условиях «недонаселенности» гор Азиатской Субарктики. Учитывая расширение признания гипотезы о самоорганизации животных сообществ вообще (Лапкин, Извеков, 1995; Князева, Курдюмов, 2004), данное предположение представляется еще более убедительным. Не исключен еще один аспект выявленной на плато Путорана закономерности. В условиях крайне низкой населенности, далекой от потенциально возможной, и, следовательно, почти при отсутствии межвидовой конкуренции, горно-субарктические ландшафты, вероятно, более активно осваиваются целыми сообществами птиц, нежели отдельными видами. И хотя механизмы выявленной закономерности пока еще не исследованы, все это вполне согласуется с взглядами на сообщества как целостные самоорганизующиеся живые системы, способные эволюционировать (Шварц, 1973; Уитеккер, 1980; Разумовский, 1981; Чернов, 1984; Северцов, 1990). К сожалению, теория эволюции сообществ, к построению которой ближе всех подошел В.В. Жерихин (1987, 1994, 1997, 2003), еще далека от полноценной разработки (Воронцов, 2004).
5.4 Закономерности вертикальной изменчивости гнездового населения птиц гор Азиатской Субарктики в пределах однородного ландшафта
В ГАС отличия в общей плотности населения птиц и обилии многих видов обнаруживаются при относительно небольшом смещении по вертикали в пределах однородного ландшафта любого из высотно-ландшафтных поясов (Кищинский, 1988; Романов, 1996). Размещение птиц по высотному профилю в пределах лесного пояса ГАС подчиняется важной закономерности: концентрации их в нижних частях пояса. В пределах подгольцового пояса гор Азиатской Субарктики вертикальная дифференциация размещения видов связана с экологической спецификой двух основных предпочитаемых местообитаний. Верхние подгольцы Путорана, отличающиеся максимально разреженными древостоем и кустарниковой растительностью, населяют виды открытых тундровых пространств гольцов: тундряная куропатка, хрустан, золотистая ржанка, американский и краснозобый коньки, обыкновенная каменка. Виды, в большинстве своем связанные с зарослями кустарников и обитающие в основном в лесных ландшафтах Путорана, тяготеют в пределах подгольцов к нижней их полосе, непосредственно примыкающей к расположенному ниже лесному поясу. Среди них кукша, варакушка, таловка, бурый дрозд, обыкновенная чечетка, полярная овсянка, овсянка-крошка (Романов, 1996, 2003, 2006). В основе вертикальной изменчивости населения птиц в пределах гольцового пояса гор Азиатской Субарктики лежит поступательная трансформация растительного покрова с высотой. Например, на плато Путорана более богатый видовой состав и максимальная плотность населения птиц выявлена в нижней полосе гольцового пояса, составляющей лишь 18% их жизненного пространства в вертикальной плоскости. Здесь представлены 80% (n=16) всего видового состава птиц гольцов, а плотность населения (79 ос/км²) более чем в 2 раза выше средней (35 ос/км²) по данному высотному поясу. С высотой происходит резкое падение обоих показателей (8 видов; 22,5 ос/км²) и минимальных значений они достигают в самой обширной верхней части гольцового пояса (5 видов; 8 ос/км²), составляющей почти 60% жизненного пространства птиц в вертикальной плоскости. Из-за особой экстремальности экологических условий горных вершин сюда проникают лишь 20% видов птиц, формирующих авифауну гольцов, а плотность населения более чем в 4 раза ниже средней по данному высотному поясу (Романов, и др., 2007).
5.5 Общие закономерности формирования гнездового населения птиц озерно-речной системы гор Азиатской Субарктики
В сухопутных и водных местообитаниях векторы изменения плотности населения птиц, связанные с увеличением абсолютной высоты, различны. На плато Путорана плотность населения птиц водно-околоводных местообитаний постепенно возрастает от лесного пояса (6 особей на 1 км береговой линии) к подгольцовому, где достигает максимальной величины (10,3 особи на 1 км береговой линии). Далее, по мере увеличения абсолютных высот местности, она неуклонно снижается в гольцовом поясе, но достигает здесь значений (5,4 особи на 1 км береговой линии) лишь не намного уступающих соответствующим показателям лесного пояса. Сокращение обилия ряда фоновых (или лидирующих) видов от подгольцового пояса в сторону лесного и гольцового позволяет сделать предположение о том, что оптимальные местообитания в вертикальной составляющей ареала этих видов расположены в подгольцовом поясе, а субоптимальные – в лесном и гольцовом поясах. Например, на плато Путорана таковы морянка, синьга, обыкновенный турпан, сибирский пепельный улит, полярная крачка. Для сибирского пепельного улита подобная ситуация указывает на его тесные экологические связи со специфическими условиями горных вершин Азиатской Субарктики. Морянка, синьга, обыкновенный турпан и полярная крачка, будучи экологически тесно связанными с зональными гипоарктическими ландшафтами (Данилов, 1966; Успенский, 1969; Кищинский, 1988; Рябицев, 2001; Кречмар, Кондратьев, 2006), и в условиях ГАС осваивают преимущественно горные аналоги этих ландшафтов, господствующие в подгольцовом поясе.
В населении птиц водно-околоводных местообитаний лесного пояса лидируют – большой крохаль, сибирский пепельный улит, малая и серебристая чайки, полярная крачка, подгольцового – морянка, синьга, обыкновенный турпан, фифи, сибирский пепельный улит, полярная крачка, гольцового – галстучник, сибирский пепельный улит, перевозчик, кулик-воробей, белохвостый песочник, полярная крачка. Таким образом, в водно-околоводных местообитаниях всех поясов лидируют 13 видов, из которых лишь 2 являются общими, а каждый из 11 остальных – специфичным только для одного высотного пояса.
Максимальная плотность населения птиц водно-околоводных местообитаний превышает минимальную лишь в 1,9 раза, тогда как в сухопутных местообитаниях – в 8,6 раза.В водно-околоводных местообитаниях сходство населения лесного и подгольцового поясов, определенное c помощью коэффициента сходства населения – КСН (Наумов, 1964), составляет 26%, подгольцового и гольцового – 25%, а лесного и гольцового – 40%. Амплитуда между минимальным (25%) и максимальным (40%) показателем КСН в водно-околоводных местообитаниях намного меньше, чем в сухопутных (соответственно 2% и 29%), а абсолютные значения КСН - выше. Все это указывает на то, что в пределах ГАС сообщества птиц интразональных (внепоясных) водно-околоводных местообитаний более устойчивы, стабильны и однородны в пространстве, чем сухопутных.
5.6 Закономерности сезонной динамики населения птиц в горах Азиатской Субарктики
Существуют два экологически сходных пространственных вектора послегнездовых кочевок в Субарктике: в широтно-зональной плоскости в северном направлении из крайней северной тайги в лесотундру, и в вертикальной высотно-поясной плоскости вверх из лесного пояса в подгольцы и гольцы (Михеев, 1964; Данилов, 1966; Кречмар, 1966; Успенский, 1969; Исаев, 1994, 1996; Рыжановский, 1997; Романов, 1988, 1989, 1996, 2001, 2003, 2004, 2006; Селиванова, Естафьев, 2001; Романов, Карпов, 2002; Селиванова 2002, 2008).
По нашим наблюдениям на плато Путорана, динамика населения птиц гольцового, подгольцового и лесного поясов на рубеже гнездового и послегнездового периодов в подавляющем большинстве случаев имеет иммиграционный тип. Устойчивое увеличение плотности населения в послегнездовой период происходит на фоне одновременного сокращения видового состава (табл. 3).
Таблица 3. Изменения в населении птиц в послегнездвой период
Высотный пояс
|
Увеличение плотности населения на рубеже гнездового и послегнездового периодов |
Сокращение видового состава на рубеже гнездового и послегнездового периодов |
| Гольцовый пояс |
в 2,6-5 раз |
в 1,5-1,9 раза |
| Подгольцовый пояс |
в 1,5-22 раза |
в 1,1-10 раз |
| Лесной пояс |
в 1,1-1,7 раза |
в 1,2-1,9 раза |
Выявленные ежегодные перемещения большого количества особей многих равнинных видов в верхние части высотного профиля, вероятно, можно рассматривать как составной компонент сезонных аспектов формирования и динамики авифауны гор Азиатской Субарктики, а также - глобального процесса освоения равнинными видами горных ландшафтов Азиатской Субарктики.
В послегнездовой период с точки зрения стратегии экономии энергетических затрат в северотаежной Субарктике для многих птиц более оптимальны горные ландшафты нежели равнинные. Массовый подъем кочующих выводков из горно-лесного пояса в вышележащие высотные пояса позволяет многим молодым птицам воробьинообразных быстрее оказаться в кормных местообитаниях горных вершин. Кроме этого, у молодых птиц в процессе послегнездовых кочевок происходит «запечатление и образование связи» с территорией будущего гнездования (Lohrl, 1959; Catchpole, 1972; Brewer, Harrison, 1975; Ralph, Mewaldt, 1975; Berndt, Winkel, 1979; Рыжановский, 1997). Поэтому правомерно предположить, что молодые птицы, оказавшиеся выше верхней границы леса, могут успешно «запечатлеть» инородный ландшафт горного редколесья или тундры как образ потенциально возможной территории гнездования. Вероятно, в том числе с этим связана весьма высокая доля особей в популяциях некоторых видов, ежегодно успешно гнездящихся не только в нижней части лесного пояса ГАС, но и у верхних пределов распространения лесной растительности (Кищинский, 1988; Романов, 1996, 2006; Романов и др., 2007). Возможно, что наряду с некоторыми экологическими факторами внешней среды, привлекающих птиц в гнездовые биотопы подгольцового и верхней части лесного поясов, важное значение имеет опыт «запечатления» ими этих горных ландшафтов в период послегнездовых кочевок первого года жизни.
Глава 6
Современное состояние авифауны гор Азиатской Субарктики как отражение закономерностей ее формирования и динамики
6.1 Таксономическая структура авифауны гор Азиатской Субарктики; региональные фаунистические различия
Из 657 видов птиц гнездящихся в России (Степанян, 2003; Коблик и др., 2006) в ГАС гнездится 197 видов (30% авифауны РФ). Таксономическая структура гнездовой авифауны и пропорции соотношений удельного веса различных отрядов (в %) в общем сохраняются на всем протяжении цепи ГАС от Приполярного Урала до Корякского нагорья. Такая закономерность, с учетом того, что показатель представленности общей гнездовой авифауны гор Азиатской Субарктики в большинстве ее частей достигает 65-69%, свидетельствует в пользу значительного сходства авифаун отдельных регионов между собой и общей однородности авифауны рассматриваемой части материковой суши.
Таксономическая структура гнездовой авифауны ГАС соответствует зональным и ландшафтным особенностям рассматриваемой части Азии с доминированием 4 отрядов, характерных для бореального и гипоарктического поясов Палеарктики: воробьинообразных – 91 вид (46,4 % авифауны ГАС), ржанкообразных - 41 вид (20,8%), гусеобразных -
27 видов (13,7 %), соколообразных - 15 видов (7,6 %).
В обширных горных областях Азии выявлена тенденция уменьшения таксономического разнообразия на уровне отрядов с продвижением в северном направлении, что является проявлением общезональной закономерности (табл. 4).
Таблица 4. Изменение таксономического разнообразия на уровне отрядов с широтой
Регионы в пределах
90-100º в.д.
|
Число отрядов |
Регионы в пределах
132-135º в.д.
|
Число отрядов |
| Таймыр |
7 |
Хребты Кулар и Полуосный |
11 |
| Плато Путорана |
13 |
Хребты Верхоянский и Черского |
12 |
| Алтай-Саянский регион |
19 |
Хингано-Буреинское нагорье |
18 |
Суммарно на долю гусеобразных, соколообразных, ржанкообразных, воробьинообразных, как на плато Путорана, так и в хребтах Верхоянского и Черского, приходится 85% отмеченных видов. Севернее эта доля еще более возрастает (до 89-90%), а южнее несколько сокращается (до 79-81%). Аналогичная закономерность увеличения к северу от гор Азиатской Субарктики и сокращения – к югу от них прослеживается в доле участия гусеобразных и ржанкообразных. Напротив, вектор изменения доли участия воробьинообразных и соколообразных имеет обратное направление.
6.2 Широтная и меридиональная дифференциация авифауны гор Азиатской Субарктики
Проведенный нами анализ видового состава авифаун отдельных регионов Евразии, выявил, что в цепи ГАС, в отличие от равнин, центр относительного видового разнообразия смещен восточнее Урала и расположен на плато Путорана. К западу, в сторону Приполярного и Полярного Урала, видовое разнообразие авифауны снижается на 10% (гнездовой – на 6%), к востоку, в сторону Верхоянья, хребта Черского и Колымского нагорья - на 15% (гнездовой – на 3-5%), и далее, в Корякском нагорье – на 26% (гнездовой – на 22%). Центры видового разнообразия авифауны как в горах Азиатской Субарктики (Путорана), так и в горах Южной Сибири (Алтай-Саянский регион) проецируются на меридиональный внутриконтинентальный трансект по 90-100º в.д. (Рогачева, 1988; Романов, 1996; Бисеров, 2006; Баранов, 2007). Выявленное повышенное видовое разнообразие авифауны Путорана соответствует особенностям его положения в пределах Енисейской зоогеографической границы, сформировавшейся по линии ледникового разрыва авифаунистических комплексов и ареалов видов (Рогачева, 1988). Пограничное положение между темнохвойной западно-сибирской тайгой и светлохвойной тайгой Средней и Восточной Сибири обусловливает переходный характер путоранской авифауны и ее повышенное видовое богатство. В составе этой авифауны находятся виды и подвиды - типичные представители различных орнитокомплексов (таежных и тундровых), господствующих западнее или восточнее Енисейской границы и не распространяющихся далее от нее. Золотистая ржанка, серая ворона, камышевка-барсучок и серая мухоловка находят на плато Путорана восточный предел распространения. Такие виды, как клоктун, горбоносый турпан, азиатская бурокрылая ржанка, сибирский пепельный улит, кроншнеп-малютка, белопоясничный стриж, американский конек, сибирский жулан, соловей-свистун, черная ворона, дрозды сибирский и Науманна, бурый дрозд, сибирская чечевица не проникают западнее Енисейской зоогеографической границы, а территория плато Путорана составляет часть западной периферии их ареала. Последних видов в путоранской авифауне несколько больше, что свидетельствует в пользу преобладающего влияния на фауну Путорана орнитокомплексов, распространенных восточнее Енисейской зоогеографической границы и самого плато, за счет которых в основном и поддерживается повышенное видовое богатство авифауны обсуждаемого региона (Романов, 1996). У большого числа видов – и западных, и восточных – в районе плато Путорана проходит граница подвидов, имеющих западное и восточное распространение (Степанян, 2003). Таковы Delichon
urbica
urbica
(L.) и Delichon
urbica
lagopoda
(Pall.), Motacilla
alba
dukhunensis
Sykes и Motacilla
alba
ocularis
Swinh., Phylloscopus
collybita
fulvescens
Sever. иPhylloscopus
collybita
tristis
Blyth., Phylloscopus
trochiloides
viridanus
Blyth. и Phylloscopus
trochiloides
plumbeitarsus
Swinh., Pinicola
enucleator
enucleator
(L.)иPinicola
enucleator
kamtschatkensis
(Dub.). Случаев одновременного распространения соответствующих пар подвидов на территории Путорана пока выявлено немного (Романов, 1996, 2004, 2006; Романов, Голубев, 2007). Это доказано лишь в отношении Motacilla
alba
dukhunensis
и Motacilla
alba
ocularis
. Для воронка и теньковки зарегистрированы только восточные подвиды (Delichon
urbica
lagopoda
(Pall.) и Phylloscopus
collybita
tristis
Blyth.), для щура – только западный подвид (Pinicola
enucleator
enucleator
(L.).
На повышение видового разнообразия авифауны Путорана возможно начинает оказывать процесс расселения видов, имевших до оледенения ареалы значительно шире и сокративших их в период ледникового времени, не образовав разрывов, а в настоящее время быстро расселяющихся на свои исконные территории (Матюшкин, 1976). На юге Палеарктики так ведут себя, например камышницаи зяблик (Баранов, 2003; 2007). Предполагаем аналогичный процесс для ряда видов, имеющих в пределах среднего звена цепи ГАС (плато Путорана, горы Якутии) хорошо заметный «южный прогиб» северной границы ареалов (широконоска, луток, чеглок, обыкновенная пустельга, озерная чайка, большой и малый пестрый дятлы, пятнистый конек, зеленая пеночка, обыкновенный снегирь, овсянка-ремез, дубровник). Мы также связываем данную деформацию ареалов с распространением в прошлом ледников и перигляциальных ландшафтов, оттеснивших эти виды к югу. Недостаточный объем наблюдений не позволяет сделать репрезентативные выводы о положительных трендах динамики северных границ их распространения и восстановления доледниковых ареалов. Однако, в пользу этого предположения говорят, например, регистрации в 2006-2007 гг. первых фактов гнездования на территории Путорана обыкновенной пустельги, зеленой пеночки, овсянки-ремеза (Романов, 2006; Романов, Голубев, 2007, Романов, 2008), а также исследования, подтверждающие поступательное смещение в северном направлении границ природных зон (Харук и др., 2004; Loarieetal., 2009).
Для авифаун гор Азиатской Субарктики нами выявлен высокий в целом уровень их взаимного сходства. Это подтверждено высокими значениями коэффициента фаунистической общности - КФО, полученными по формулам Серенсена и Жаккара (Песенко, 1982; Чернов, 1975, 2008). Максимальное сходство авифауна каждого из горных регионов Азиатской Субарктики проявляет с авифаунами ближайших в широтном направлении соседних регионов. Одинаковые (n=85%) максимально высокие уровни сходства проявляет авифауна гор Якутии с авифаунами ближайших к западу (плато Путорана) и ближайших к востоку (Колымское нагорье) горных систем. Это позволяет предполагать формирование наиболее однородной авифауна в этой области горной Субарктики. Вероятно, это связано с тем, что перестройки экосистем большинства горных систем Якутии в плейстоцене были менее катастрофичны, чем в экосистемах других регионов цепи ГАС (Караваев, 1955; Нейштадт, 1957; Хотинский, 1977; Савина, 1986; Андреев, 1989; Томская, 1989). Это обусловило в целом относительно более стабильное развитие местной авифауны, которая (видимо главным образом в качестве донора) в свою очередь обеспечивала возможность видообмена с ближайшими регионами, и среди прочего способствовало формированию повышенного видового разнообразия авифауны плато Путорана. Несомненно, однородность авифауны указанных горных областей поддерживается также сходством современных экологических условий.
Авифауна гор Азиатской Субарктики имеет четко оформленное общее фаунистическое ядро. Приблизительно половину гнездовой авифауны всей цепи ГАС формируют виды, широко распространенные в северной тайге, лесотундре, частично в южной тундре и одновременно входящие в состав авифаун почти всех его регионов от Урала до Корякского нагорья (Воробьев, 1963; Кищинский, 1968, 1980; Естафьев, 1977, 1981; Сыроечковский-мл. и др., 1996; Романов, 1996, 2003, 2004; Рябицев, 2001; Селиванова, 2002, 2008; Головатин, Пасхальный, 2005; Борисов и др., 1996, 2007; Бабенко, 2007; Поспелов, 2007).
Формирование и пространственно-временная динамика авифауны гор Азиатской Субарктики определяется также закономерностями широтно-зонального характера. Например, при движении в северном направлении общее видовое разнообразие гнездовой авифауны сокращается со 129 видов на Приполярном Урале (Селиванова, 2008) до 94 – на Полярном Урале (Головатин, Пасхальный, 2005). Показательно сокращение (с 19 до 2 видов) в северном направлении видового разнообразия гнездящихся гусеобразных плато Путорана (Романов, 2004). Закономерность, согласно которой видовое богатство авифауны уменьшается в более высоких широтах, прослеживается и в горах Якутии. В средней части Верхоянского хребта (65º30'–66º30' с.ш.) установлено пребывание 94 видов (Борисов и др., 2007). Севернее, на Полуосном кряже и хребте Кулар (69º00'–70º00' с.ш.) разнообразие авифауны сокращается до 77 видов (Сыроечковский-мл. и др., 1996). Авифаунистические отличия между северными и южными частями Колымского нагорья отражены в зоогеографическом районировании региона (Кищинский, 1968). Среди 6 зоогеографических участков, дифференциация 4 связана в значительной мере с широтой местности. Отличия между авифауной северных и южных частей выявлены и в Корякском нагорье (Кищинский, 1980). Общий гипоарктический облик авифауны этой горной страны более ярко выражен в ее южной и центральной частях, в зоне лесотундры. На крайнем северо-востоке, в пределах тундровой зоны, возрастает роль тундровых (гемиарктических) видов.
Существует еще один аспект широтного распространения некоторых птиц. По гольцовым вершинам гор Азиатской Субарктики равнинно-тундровые виды (морянка, горбоносый турпан, лапландский подорожник) и арктоальпийский вид (пуночка) проникают намного южнее границ своего зонального ареала. На крайнем северо-востоке Азии подобные случаи были выявлены достаточно давно (Кищинский, 1988). Исследованиями последних двух десятилетий аналогичный характер проникновения на юг лапландского подорожника и пуночки (на расстояние до 500 км) выявлен и в более континентальных ГАС – плато Путорана, хребтах Северо-Восточной Якутии (Сыроечковский-мл. и др., 1996; Романов, 2006; Поспелов, 2007).
В горах Азиатской Субарктики имеет место и обратное явление: некоторые виды птиц проникают по нижней (лесной) части долин крупных горных рек намного севернее, чем по долинам равнинных рек и тем более – по равнинным водоразделам. В долинах рек ГАС основная часть ареалов таких видов приобретает вид своеобразных максимально выступающих в северном направлении «языков». Они проецируются на интразональные элементы ландшафта – долины горных рек, в которых градиенты климатических факторов сглаживаются особенно сильно. Наиболее отчетливо это выражено на севере Средней Сибири, где древесная растительность проникает до 72º с.ш. (Чернов, 1975; Абаимов и др., 1997). Самые продуктивные и высокобонитетные леса в этой части Азиатской Субарктики приурочены к горным долинам плато Путорана, где они заходят почти на 2º севернее, чем на сопредельных равнинах или Анабарском плато (Гвоздецкий, Голубчиков, 1987; Абаимов и др., 1997). Поэтому, например, повсеместно обычные или даже многочисленные в лесах Путорана свиристель, сибирская завирушка, зарничка, вьюрок, белокрылый клест, в более угнетенных лесах Анабара, лежащих на широте путоранских лесов, становятся крайне редкими и встречаются спорадично (Бабенко, 2007; Поспелов, 2007). А такие виды как тетеревятник, перепелятник, речная крачка, мохноногий сыч, вертишейка, желна, малая мухоловка, синехвостка, щур находят в лесах речных долин Путорана северный предел своего распространения (Морозов, 1984; Романов, 1996, 2006, 2009).
Таким образом, обладая значительной вертикальной дифференциацией ландшафтов, горы Азиатской Субарктики служат своеобразными миграционными коридорами. В процессе формирования авифаун, горы обеспечивают высотно эшелонированную систему разнонаправленных путей пространственного распространения для многих видов птиц, благодаря чему по подходящим ценозам разных высотных поясов происходит взаимопроникновение северных форм на юг, а южных на север. Все это, несомненно, влияет на повышение уровня видового богатства в процессе формирования авифауны горных стран.
6.3 Особенности формирования вертикальной неоднородности авифауны гор Азиатской Субарктики
От подножия к вершинам, в соответствии с последовательной сменой трех основных высотно-ландшафтных поясов, на соответствующих высотах формируются горно-северотаежная, подгольцовая и гольцовая авифауны.
Соотношение таксономических групп, слагающих основу авифауны гольцов, подгольцов и лесного пояса, в целом сохраняется по всему высотному профилю. Наиболее значимы (по мере убывания) воробьинообразные (43-57%), ржанкообразные (18-28%), гусеобразные (12-14%), соколообразные (8%). От подножий к вершинам ГАС возрастает удельный вес ржанкообразных (с 18 до 28%), в частности куликов (с 14 до 23%), что сближает структуру авифаун горных и равнинных тундр. Удельный вес воробьинообразных оказывается максимальным (57%) в подгольцах. Это связано с экологическими особенностями данного пояса, где доминируют кустарники, редины и стланики, т. е. местообитания, к освоению которых более других групп адаптированы воробьинообразные.
Закономерности вертикального изменения доли видов различных географо-генетических групп следующие. Широкораспространенные виды наиболее значимы по доле участия на всех уровнях высотного профиля, несмотря на то, что этот показатель сокращается от подножий к вершинам (с 41 до 29%). Аналогичная тенденция выявлена и у бореальных видов (с 25 до 11%). Закономерное увеличение удельного веса в авифауне от подножий к вершинам в ГАС происходит у большего числа географо-генетических групп (n=5): эоарктов (с 0,5 до 2,4%), гемиарктов (с 7,6 до 11%), гипоарктов (с 10 до 15%), арктоальпийских (с 1,1 до 6,1%) и альпийских (с 0,5 до 7,3%) видов. За счет этого авифауна постепенно с высотой приобретает специфические горные черты, а также – особенности, которые сближают ее с авифауной равнинной тундры.
В пределах лесного пояса формируется наиболее богатая и разнообразная авифауна, 185 видов которой составляют 94% авифауны гор Азиатской Субарктики (табл. 5). Выше, в экстремальных условиях горных вершин, приближающим их по параметрам внешней среды к арктическим тундрам, формируются значительно более бедные видами подгольцовая и гольцовая авифауны. Подгольцовую авифауну формируют 103 вида, гольцовую – 82 вида, что соответственно составляет 52,3% и 41,6% всей авифауны гор Азиатской Субарктики.
Таблица 5. Изменение количества видов в горах Азиатской Субарктики с высотой
| № |
Высотно-ландшафтный пояс |
Приполярный
и Полярный Урал
|
Плато Путорана |
Анабарское плато |
Горы Якутии |
Колымское нагорье |
Корякское нагорье |
Всего в ГАС |
| 1 |
Гольцовый |
52 |
40 |
18 |
43 |
19 |
21 |
82 |
| 2 |
Подгольцовый |
61 |
52 |
14 |
48 |
34 |
52 |
103 |
| 3 |
Лесной |
119 |
129 |
68 |
119 |
120 |
90 |
185 |
| 4 |
Всего видов |
129 |
137 |
77 |
130 |
133 |
107 |
197 |
Видовой состав авифаун при переходе от одного к другому высотно-ландшафтному поясу меняется постепенно. При наличии в авифауне каждого вертикального пояса своих специфических видов, авифауны двух соседних поясов имеют в своем составе много общих видов. Из 185 видов птиц гнездящихся в лесном поясе ГАС и 103 видах - в подгольцовом поясе, 95 видов являются общими для авифаун обоих поясов. Из 103 видов птиц гнездящихся в подгольцовом поясе и 82 видах - в гольцовом поясе, 63 вида являются общими для авифаун этих поясов. Более того, из-за пониженного расположения «снеговой линии» жизненное пространство здесь значительно уже, чем в горах более южных широт, например - горах юга Палеарктики. Благодаря этому птицы различных фаунистических и географо-генетических комплексов, имеющие специфические адаптации к жизни в тайге, тундре и горах, оказываются в горах Азиатской Субарктики в непосредственной близости друг от друга. При наличии соответствующих экологических предпосылок они имеют возможность не только достаточно быстро перемещаться из пояса в пояс, но образовывать совместные «смешанные группировки».Это позволяет сохранять непрерывность и максимальную эффективность формирования и трансформации авифауны в соответствии с направлением пространственно-временных изменений условий внешней среды в горах. Широкое вертикальное распространение многих видов птиц в горах Азиатской Субарктики предопределяет большое общее биоразнообразие даже в высотных поясах с экстремальными условиями, и как следствие – сохраняет высокую потенциальную возможность успешного эволюционного развития горных сообществ и формирования горной авифауны в целом.
6.4 Автономность формирования орнитокомплексов различных высотных поясов гор Азиатской Субарктики
Анализ степени общности гольцовых, подгольцовых и горно-лесных авифаун ГАС, проведенный с использованием формул Серенсена и Жаккара (Песенко, 1982; Чернов, 1975, 2008), выявил следующие закономерности.
В пределах цепи ГАС большее единообразие и однородность основной части видового состава характерно для горно-лесной авифауны, и значительно меньшее для подгольцовой и гольцовой авифауны. Горно-лесная авифауна ГАС в современном ее виде сформировалась в пределах единой области Северной Азии с повсеместным господством северотаежных лесов и гипоарктических редколесий. В условиях непрерывности лесных ландшафтов и повсеместной сопряженности горных и равнинных типов лесов подавляющее большинство видов осваивает их повсеместно, так как не имеет непреодолимых преград, препятствующих расселению. Раздробленность, взаимная удаленность и изоляция участков подгольцовых и гольцовых поясов в совокупности с более разнообразными и нестабильными экологическими условиями обусловила меньшее взаимное сходство региональных авифаун на уровне каждого из этих поясов.
Авифауна каждого высотно-ландшафтного пояса всех гор Азиатской Субарктики проявляет максимальное сходство с авифауной соответствующего пояса ближайших в широтном направлении соседних регионов.
Для горно-лесной, подгольцовой и гольцовой авифаун выявлены области с максимально высоким уровнем взаимной общности. В цепи ГАС положение этих областей различно. Наиболее качественно однородная (83-84%) горно-лесная авифауна сформировалась в горах Якутии и ближайших к ним горных системах: с запада - плато Путорана, с востока - Колымском нагорье. Это согласуется с положением о единстве таежной сибирской фауны, в становлении которой особенно велика роль весьма специфического восточносибирского (Ангарского) фаунистического комплекса (Чернов, 1975). Сходство авифаун регионов, одновременно охватывающих горно-предгорные и равнинные территории Северной Азии (55-58,3%) (Чернов, 1975), меньше, чем горно-таежных авифаун ГАС (83-84%). Это свидетельствует о более высокой однородности авифауны горно-таежных районов и дает основание рассматривать формирование авифауны ГАС, как оригинальный процесс, протекающий в едином, относительно, самостоятельном экологическом и зоогеографическом пространстве. Повышенное сходство горно-таежных авифаун ГАС можно также предположительно объяснить и тем, что их основу одновременно формируют виды, адаптированные к освоению горно-гипоарктических или горно-бореальных ландшафтов. Другие виды, экологически связанные со специфичными местообитаниями отдельных равнинных регионов, не проникают в горы и понижают взаимное сходство авифаун этих регионов.
Взаимно наиболее схожие (61-65%) подгольцовые авифауны формируются в Корякском нагорье и в соседних с ним горах Якутии и Колымском нагорье. Вероятно, это тесно связано со становлением, так называемой, берингийской лесотундры и ее горных дериватов, очень характерных для указанной территории и особенно восточной ее части (Кищинский, 1988).
Наиболее однородная (63%) гольцовая авифауны формируется в северной области цепи ГАС: на плато Путорана и ближайших с востока горах Якутии и с запада - Приполярном и Полярном Урале. Эта область занимает срединную, переходную с точки зрения взаимопроникновения видов, часть Евразийской Субарктики, авифауна которой в известной мере объединяет элементы европейских и азиатских северных биомов (Pleske, 1928; Успенский, 1969; Rogachevaetal., 1995). Многие из этих элементов (обитателей равнинных тундр) проникают в горы Субарктики на всем отрезке от Урала до Верхоянского хребта, что и поддерживает повышенное сходство гольцовых авифаун указанных горных стран.
Выявленные различия уровня общности соответствующих авифаун указывают на определенную автономность формирования авифауны на уровне каждого высотно-ландшафтного пояса ГАС. В пользу этого свидетельствует также и то, что максимально сходные гольцовые, подгольцовые и горно-таежные авифауны формируются на разных участках горной цепи Азиатской Субарктики и под влиянием различных факторов.
6.5 Общность происхождения и формирования авифауны гор и равнин Севера Азии
Проведенный нами сравнительный анализ современных авифаун равнин и гор Азиатской Субарктики подтвердил то, что подавляющее большинство видов (70-81%) из сопредельных равнин проникают в соответствующие горные регионы, активно осваивают их, и формируют основу региональной гнездовой авифауны. Это в значительной мере определяет высокий уровень сходства авифаун крупных соседних равнинных и горных регионов. Авифаунам последних это придает в целом также преимущественно более «экологически равнинный» облик. Ареалы большинства равнинных видов Северной Палеарактики захватывают ГАС. Свободный обмен видами между равнинными и горными регионами Азиатской Субарткики обуславливает более однородную авифауну на всем ее протяжении независимо от региональных особенностей рельефа.
Некоторые виды (галстучник, золотистая ржанка, варакушка, краснозобый конек, лапландский подорожник) заселяют в горных условиях участки аналогичные тем, которые они осваивают на равнинах. Ряд видов (обыкновенная кукушка, сибирская завирушка, синехвостка, бурый дрозд, таловка, вьюрок), обладающих большей экологической пластичностью или адаптационными возможностями, могут заселять местообитания в различной степени отличающиеся от исходных – тех, которых они более или менее строго придерживаются на большей части равнинной составляющей своего ареала. Но независимо от уровня экологической пластичности и отличий в стратегии освоения чужеродного ландшафта, большинство равнинных видов, проникающих в верхние части горно-лесного пояса, в подгольцы и еще выше в гольцы, обладают общими особенностями. Все они относительно легко адаптируются к новым условиям обитания и местам гнездования, многие из них не имеют узкой кормовой специализации. Например, в авифауне Полярного Урала оказываются те же виды, которые осваивают и сопредельные пространства равнинной лесотундры (Морозов, 1989; Романов, 1989; Бутьев, Костин, 1997; Рябицев, 2001; Головатин, Пасхальный, 2005). Подавляющее большинство из отмеченных в лесотундре птиц – 110 из 130 – были обнаружены на Полярном Урале (Пасхальный, Головатин, 2005).
Число равнинных видов проникающих в горы, а следовательно, и интенсивность влияния равнинной авифауны на авифауну сопредельного горного региона уменьшается от подножья к вершинам.
Как один из элементов продолжающегося процесса формирования авифауны гор Азиатской Субарктики мы рассматриваем закономерность, в соответствии с которой многие равнинные виды обычные на большей части Северной Палеарктики при движении на восток «поднимаются» в горы, достоверно расширяя вверх вертикальную составляющую своих ареалов. На фоне обеднения равнинной авифауны Азиатской Субарктики с запада на восток, доля равнинных видов проникающих в горные регионы последовательно возрастает в том же направлении (с 70 до 81%). Вероятно, это связано с увеличением в авифауне более восточных районов Азиатской Субарктики удельного веса видов лучше адаптированных к суровым условиям внешней среды этой обширной части континента. Это в свою очередь определяет то, что большая часть видов местной авифауны может успешно осваивать не только равнинные территории, но и беспрепятственно проникать в горные регионы.
В «макромасштабе» авифауны гор Азиатской Субарктики по числу видов беднее авифаун сопредельных с ними крупнейших равнин Евразии. Но эта особенность не всегда проявляется при сравнении авифаун гор Азиатской Субарктики и примыкающих вплотную к ним меньших по площади равнинных предгорий. Например, авифауны Верхоянского хребта и плато Путорана демонстрируют различные векторы изменения видового богатства при движении от центра горной страны к их предгорьям. При движении вглубь Верхоянской горной страны наблюдается качественное и количественное обеднение авифауны, а на плато Путорана – обратная закономерность. Это связано с региональной спецификой литогенной основы. В хребты с большими абсолютными высотами и треугольным профилем в поперечном сечении проникновение многих видов птиц из предгорий сдерживают повышенные углы наклона речных долин, сокращение их длины и, следовательно, - распространения пойменных биотопов, по которым в основном идет проникновение равнинных видов в горы. Монолитные горные массивы, имеющие в поперечном сечении профиль трапеции, повсеместно расчлененной глубокими сквозными тектоническими долинами, абсолютная высота дна которых лишь ненамного превышает высоту окружающих предгорий, позволяют проникать вглубь гор большему числу видов птиц. Эти региональные особенности литогенной основы, способствуя проникновению равнинных видов вглубь горных массивов по лесным местообитаниям в горизонтальной плоскости, не менее эффективно затрудняют этот же процесс в вертикальном направлении.
Продолжение формирования авифауны гор Азиатской Субарктики за счет равнинных видов логично ожидать и в современную эпоху. Этому объективно способствуют соответствующие изменения внешней среды: потепление климата (Kaufmanetal., 2009; Loarieetal., 2009; Postetal., 2009), таяние ледников (Svenssonetal., 2004; Velicogna, Wahr, 2005; Komori, 2008), продвижение кустарниковой растительности вверх по горным склонам (Ваганов и др., 1996; Naurzbaev, etal., 2004; Головатин, Пасхальный, 2005).
Проведенное нами сравнение равнинных и горных авифаун Азиатской Субарктики с использованием формул Серенсена и Жаккара (Песенко, 1982; Чернов, 1975, 2008), выявило высокое сходство (60-80%) северо-таежных авифаун гор и равнин этой области суши. Сходство авифаун равнинных и горных тундр значительно меньше (18-32%). Это обусловлено тем, что лесные ландшафты занимают преимущественно непрерывное единое пространство, а тундровые – хотя и обширные, но в значительной мере изолированные участки. Повсеместный в пределах Азиатской Субарктики непосредственный контакт равнинных и горных северо-таежных лесов обеспечивает возможность широкого обмена видами между их авифаунами (и прежде всего проникновение равнинных птиц в горы), а следовательно и большую однородность. Тундровые авифауны гор и равнин Азиатской Субарктики более обособлены, имеют меньше сходства и потому не столь однородны как лесные в силу большей взаимной пространственной удаленности, как в горизонтальной, так и в вертикальной плоскости. Пространственные дизъюнкции тундровых ландшафтов затрудняют свободный обмен видами между авифаунами их горных и равнинных аналогов, заметно снижают интенсивность проникновения равнинных видов из равнинных тундр в горные тундры гольцов. Лишь в наиболее северных районах Азиатской Субарктики, где на обширных пространствах равнинные тундры непосредственно смыкаются с горными, обмен видами между их авифаунами значительно активизируется, в силу чего существенно возрастает и уровень их взаимного сходства.
В составе авифауны лесных ландшафтов гор Азиатской Субарктики нет специфических горных подвидов и качественно она идентична авифауне сопредельных равнин. В подгольцовой авифауне наиболее восточных ГАС уже появляются подвиды кедровки (N
.
c
.
kamchatkensis
Barr.-Ham.) и щура (P
.
e
.
kamtschatkensis
(Dyb.), связанные с горными зарослями кедрового стланика. Но и они сформировались в процессе адаптаций к условиям гор равнинных исходных форм (Кищинский, 1988). В гольцах гор Азиатской Субарктики широко распространены арктоальпийские виды (тундряная куропатка, хрустан, рогатый жаворонок, пуночка), а также - обыкновенной каменка. В рамках рассматриваемой области суши они являются характерными обитателями высокогорий. Но в более широком пространственном диапазоне, охватывающем в целом Арктику и Субарктику, арктоальпийцы имеют «двойственное» распространение: как в верхних поясах гор, так и в зональных тундрах на равнинах. Конфигурация ареалов и экологические связи арктоальпийцев свидетельствует о том, что окончательное становление арктоальпийских таксонов и их ареалов происходило, по-видимому, в процессе формирования мезофильно-тундровых типов экосистем как в горах, так и на равнинах (Кищинский, 1988). Полагаем, что все это свидетельствует о генетической связи происхождения и формирования авифаун горных и равнинных аналогов северо-таежных, лесотундровых и тундровых ландшафтов.
На фоне активного и повсеместного проникновения равнинных птиц в горы Азиатской Субарктики обратный процесс выхода горных птиц в предгорья и тем более сопредельные равнины практически отсутствует. Авифауна равнин по отношению к авифауне ГАС выступает как «донор», а горная по отношению к равнинной - лишь как «реципиент». Практически однонаправленный видообмен между горными и равнинными авифаунами - проявление глобальной закономерности. Суть ее в том, что экстремальные ландшафты осваивают, прежде всего, менее специализированные, экологически пластичные, как правило, широко распространенные виды из соседних ландшафтов с более мягкими экологическими условиями. На равнинах Субарктики вектор этого видообмена направлен почти исключительно горизонтально на север (Чернов, 1975, 1980; Стишов и др., 1989), в ГАС – почти исключительно вертикально вверх. Виды имеющие глубокие адаптации к наиболее экстремальным условиям существования, как в горах, так и на равнинах, достаточно строго приурочены в своем распространении к областям с господством этих условий. Это ограничивает возможность их проникновения, а тем более освоения, любых иных местообитаний, даже – с более мягкими характеристиками внешней среды. Поэтому типично горные виды птиц практически не спускаются на гнездовье в более низкие высотные пояса, а эоаркты и гемиаркты не встречаются к югу от границ соответственно арктической и типичной тундровой подзон (Чернов, 1975, 1980; Стишов и др., 1989; Головатин, Пасхальный, 2005).
6.6 Фаунистическая и географо-генетическая структура авифауны гор Азиатской Субарктики; территориальная удаленность как фактор формирования региональных авифаун
Горы Азиатской Субарктики лежат в пределах обширной полосы перехода от таежных к южно-тундровым ландшафтам. Поэтому авифауна гор Азиатской Субарктики гетерогенна по происхождению, сформирована представителями различных типов фаун и географо-генетических групп.
В формировании авифауны гор Азиатской Субарктики принимают участие виды 6 типов фаун (Штегман, 1938). Наиболее значимы элементы сибирского (71 вид - 36%) и арктического (30 вид - 15%) типов авифаун. Элементы европейского (19 видов – 9,6%), китайского (13 видов – 6,6%), тибетского (4 вида – 2,0%) и неарктического (1 вид – 0,5%) типов фаун, центры становления которых лежат далеко от ГАС (или как минимум за их пределами), играют второстепенную роль, так как число их невелико, и в силу этого они не имеют сколько-нибудь существенного значения в формировании современных сообществ птиц в ГАС. При этом, они превносят специфику в формирование качественного разнообразия видового состава птиц, отражающую историко-региональный аспект взаимодействия авифаун в процессе их формирования и динамики. Немало в авифауне гор Азиатской Субарктики широкораспространенных видов (59 видов - 30,1%). Их происхождение пока недостаточно выяснено, что в определенной мере ограничивает развитие представлений о генезисе авифауны ГАС.
Влияние элементов того или иного типа фауны в структуре авифауны горного региона достоверно сокращается с пространственной удаленностью конкретной горной системы от ареала исторического формирования того или иного типа фауны. Доля видов арктического и сибирского типов фауны имеет устойчивую тенденцию к сокращению в южном направлении, а китайского и европейского типов фауны – в северном. Благодаря этому арктико-сибирская авифауна субарктических хребтов Верхоянского и Черского четко локализована в ряду пространственно дифференцированных авифаун горных стран, лежащих между 128-150 меридианами. Авифауна расположенного южнее Алданского нагорья преимущественно сибирская с небольшим участием видов китайской фауны. Авифауна еще более «южного» Сихотэ-Алиня является уже по существу китайской, хотя и с большой долей сибирских видов (Бисеров, 2006). Промежуточное положение между авифаунами Алдана и Сихотэ-Алиня занимает авифауна Хингано-Буреинского нагорья, которая почти в равных долях представлена видами китайской и сибирской фаун, а также широкораспространенными видами (Бисеров, 2006).
Выявлена региональная дифференциация авифауны гор Азиатской Субарктики в широтном направлении. Сибирские виды, почти везде составляют около 40% видового состава. Но к западу от плато Путорана их доля понижается и уже в крайне западной части пояса ГАС, на Приполярном и Полярном Урале, их доля не превышает – 32,6%. Одновременно с этим, Урал, будучи единственным регионом цепи ГАС непосредственно приближенным к историческому центру формирования европейского типа фауны, имеет в составе своей авифауны достаточно большую долю европейских видов (около 14%). В более восточные горы Азиатской Субарктики европейские элементы почти вообще не проникают. Их доля весьма незначительна на плато Путорана (около 3%), а еще восточнее, в горах Якутии и Колымском нагорье, - ничтожно мала (не более 1 %). На Анабарском плато и Корякском нагорье их нет. Еще одна закономерность формирования авифауны ГАС связана с изменениями в широтном направлении особенностей распространения видов китайского типа фауны. На фоне небольшой в целом доли участия китайских видов в авифауне ГАС, максимальное влияние они оказывают на авифауну Колымского нагорья, что проявляется и в большем числе зарегистрированных видов (n=11), и в их повышенном участии в местной авифауне (8,3%). Вероятно, это объясняется проникновением ряда китайских видов из южных районов Дальнего Востока по тайге вдоль охотского побережья, или по долинам рек с пойменными смешанными лесами неморального типа и разнообразными кустарниково-луговыми местообитаниями (Кищинский, 1968). К западу и востоку от Колымского нагорья доля китайских видов в региональных авифаунах поступательно уменьшается, наиболее существенно в западном направлении. Только в авифауне восточной половины цепи ГАС представлены виды сибирско-американского (n=2), американского (n=1) и тибетского (n=4) типа фауны. Особенностям распространения последних способствовал горный характер территорий, непосредственно связывающих историческую область формирования тибетской фауны и восточные горы Азиатской Субарктики. По ним, не встречая глобальных экологических препятствий, ряд тибетских видов и проник далеко на север: в горы Якутии, Колымское и Корякское нагорье.
Горные системы, в том числе и горы Азиатской Субарктики, носят интразональный характер в общей схеме природной зональности Палеарктики (Гвоздецкий, Голубчиков, 1987; Исаченко, Шляпников, 1989). Вместе с тем, в ГАС развиваются экологические условия, соответствующие всем основным географическим зонам Палеарктики, которые эти горы пересекают. В горных условиях сталкиваются два различных ландшафта - гипоарктический и бореальный, с образованием, широкого спектра переходных ландшафтов, иногда весьма специфичных (берингийская лесотундра). Экотонный характер авифауны наиболее выражен в северных частях ГАС и обусловлен смешением элементов упомянутых ландшафтов в следствии островного ленточного или горно-долинного взаимопроникновения тундровых и лесных участков. Мозаика размещения соответствующих ландшафтов в ГАС всецело зависит от рельефа местности: по вершинам горных массивов тундра простирается далеко на юг, между тем как по межгорным долинам северотаежные и пойменные леса проникает к северу. Все это обуславливает неоднородность авифауны ГАС по сочетанию формирующих ее представителей 8 географо-генетических групп: эоарктических - 3 (1,5%), гемиарктических - 15 (7,6%), гипоарктических - 18 (9,1%), бореально-гипоарктических - 28 (14,2%), бореальных - 47 (23,8%), широкораспространенных - 75 (38,3%), арктоальпийских - 5 (2,5%), альпийских - 6 (3,0%). Значительный вклад в авифауну ГАС бореальных, бореально-гипоарктических и гипоарктических видов птиц, суммарно составляющих 47% гнездовой авифауны, полностью согласуется с доминированием соответствующих зональных ландшафтов северной тайги, лесотундры и южной тундры в рамках рассматриваемой нами части Евразии. Гемиарктические и эоарктические виды играют второстепенную роль, так как число их невелико, и в силу этого они не имеют сколько-нибудь существенного значения в формировании современных сообществ птиц в ГАС. Гемиаркты и эоаркты связаны в своем распространении, главным образом, с подзонами типичной и арктической тундры, которые значительно удалены от северных границ цепи ГАС. Немногие из этих видов смогли, достаточно широко расселиться в этих горах, войти в состав гнездовой авифауны и почти повсеместно занять в местных сообществах устойчивые позиции (краснозобая гагара, морянка, зимняк, галстучник, полярная крачка). Некоторым гемиарктам и эоарктам удалось проникнуть в ГАС лишь локально – преимущественно по горно-тундровым ландшафтам гольцового пояса в областях максимального сближения или, что реже, - непосредственного контакта зональных тундр и северных отрогов горных массивов (тулес, азиатская бурокрылая ржанка, плосконосый плавунчик, кулик-воробей, краснозобик, малый веретенник, длиннохвостый поморник, лапландский подорожник). Наименее значима в гнездовой авифауне ГАС доля альпийских и арктоальпийских видов. Однако, несмотря на низкую долю участия, именно они определяют в процессе формирования качественного разнообразия авифауны ГАС ее горную специфику.
Важным индикатором региональной дифференциации авифауны цепи гор Азиатской Субарктики в широтном направлении является соотношение видов преимущественно «западного» и «восточного» распространения. Подобная индикация позволяет выявить значение горных массивов в качестве меридиональных экологических рубежей, ограничивающих участие определенных видов птиц в формировании авифауны более широких областей горной Субарктики.
Наиболее значительно участие «восточных» видов (n=30) в формировании авифауны восточной половины цепи гор Азиатской Субарктики. Их количество максимально в Колымском нагорье (n=28), и несколько меньше в горах Якутии (n=22), расположенных от него к западу, и Корякском нагорье (n=20), расположенном к востоку от него. Для видов рассматриваемой категории горы Якутии и Корякское нагорье в пределах ГАС представляют наиболее четко выраженные зоогеографические рубежи. Некоторые виды, при этом, оказываются еще западнее - на плато Путорана (Романов, 1996, 2003), или восточнее – в северной тайге Северной Америки (Snyder, 1957; Gabrielson, Lincoln, 1959; Godfrey, 1966; Портенко, 1972, 1973). На плато Путорана западный предел распространения находят 8 «восточных» видов. В гнездовой авифауне наиболее западных частей ГАС, на Приполярном и Полярном Урале, «восточных» видов нет.
Из общего списка видов, имеющих преимущественно «западное» распространение в пределах ГАС (n=25), большинство (n=18) не проникают восточнее Приполярного и Полярного Урала. Очевидно, что лишь в этой, крайне западной, области цепи ГАС роль «западных» видов в формировании авифауны существенна. Для них Урал в цепи ГАС представляет наиболее четко выраженный зоогеографический рубеж. Намного меньше «западных» видов (n=7) оказываются еще восточнее - на плато Путорана. На Анабарском плато восточный предел распространения находят лишь единичные «западные» виды – синьга, золотистая ржанка, желтоголовая трясогузка, рябинник. В гнездовой авифауне восточной части цепи ГАС, от Верхоянского хребта до Корякского нагорья, «западных» видов нет.
6.7 Расширение ареалов как закономерный компонент формирования и динамики авифауны гор Азиатской Субарктики
Для ряда видов птиц в ГАС выявлена динамика северных границ ареалов, пространственный и количественный рост популяционных группировок (Морозов, 1987; Кищинский, 1988; Сыроечковский-мл. и др., 1996; Романов, 1996, 2003, 2006, 2009; Головатин, Пасхальный, 2005; Рогачева и др., 2008). Формирование авифауны гор Азиатской Субарктики продолжается, в основном, за счет иммигрантов, расселяющихся из более южных областей. Благодаря этому за несколько прошедших десятилетий видовое разнообразие, например, авифаун плато Путорана и субарктических гор Якутии увеличилось на 6%. Расширение ареалов в широтном направлении – не столь распространенный процесс, хотя известны примеры расселения с востока на запад полярной овсянки (Морозов, 1987) и американского бекасовидного веретенника (Томкович, 1984; Рогачева, 1988; Лаппо, 1996).
Нами выявлено, что за 20 лет (1988-2008 гг.) гнездовой ареал синехвостки в Средней Сибири расширился к северу на 600 км, и к настоящему времени полностью охватывает горные районы плато Путорана, простираясь на север вплоть до 69º50' с.ш. (Романов, 2009). Ареал расширялся по днищам долин в горизонтальном направлении и по склонам - в вертикальном. В новообразованной части ареала прослеживается широтное изменение его неоднородности. Так, его северная половина сформирована небольшими изолированными участками с низким обилием птиц, а южная - обширными сплошными территориями, заселенными птицами с сравнительно высоким обилием, позволяющим отнести синехвостку к обычным фоновым видам. В пределах плато Путорана в 1988-2008 гг. гнездовые ареалы расширили к северу малая чайка (на 300 км), речная крачка (300 км), мохноногий сыч (600 км). В частности, в последние десятилетия ряд находок малой чайки в Центральной Сибири показали, что произошла полная или частичная ликвидация отмечавшегося ранее ледникового разрыва в ареале (Рогачева и др., 2008). Более того, восстановление единства ареала по линии бывшего разрыва сопровождается явной экспансией малой чайки в северном направлении на плато Путорана. Расширение северных границ гнездовых ареалов на плато Путорана возможно также у полевого луня, перепелятника, чеглока, обыкновенной пустельги, проникающих из более южных районов (Романов, 2009). Первые случаи гнездования отдельных пар перепелятника и обыкновенной пустельги уже зарегистрированы. Подтверждение гнездования полевого луня и чеглока, по нашему мнению, является только вопросом времени. Выявленное на плато Путорана расширение северной границы у малой чайки, речной крачки, мохноногого сыча, полевого луня имело характер резкого пульсационного выселения за пределы ареала. Это идентифицировалось по резкому (2-5 лет) появлению гнездящихся особей там, где их раньше достоверно не было, заметным увеличением частоты встречаемости кочующих особей, или тем и другим одновременно (Романов, 1996, 2006, 2009).
В районе Полуосного кряжа и хребта Кулар Е.Е.Сыроечковский-мл. с коллегами (1996) получил доказательства смещения к северу границ гнездового ареала в 1960-1996 гг. у речной крачки на 400 км, сибирского конька - 150 км, теньковки – 300 км, малой мухоловки - 200 км, черноголового чекана – 40 км, белобровика – 50 км.
Известные данные по расширению ареалов птиц на Полярном Урале и сопредельных к горам равнинах (Морозов, 1987, 1995, 2002, 2003; Головатин, Пасхальный, 2005). В частности В.В. Морозов (1987) констатирует, что широконоска, сибирская завирушка, теньковка и полярная овсянка появились в южных тундрах при своем естественном расселении в северном и западном направлении, а сизая чайка, серая ворона и сорока проникли в кустарниковые тундры явно в недавнее время благодаря освоению Крайнего Севера.
Приведенные факты указывают на то, что столь массовый и глобальный процесс как расширение северных границ ареалов птиц, придающий динамизм современному формированию авифауны Северной Евразии, охватил и горы Азиатской Субарктики.
6.8 Вертикальная составляющая ареала и изменение высотного распределения видов с широтой и долготой
В трехмерной структуре ареала в условиях гор особое значение приобретает его вертикальная составляющая (Куваев, 2006; Панфилов, 1960; Ирисов, 1997). Амплитуда абсолютных высот в горах Азиатской Субарктики достигает 1000-2500 м. Это предопределяет широкие возможности для освоения птицами пространства по вертикали: распространение в однородном ландшафте на разных абсолютных высотах, расселение из оптимальных местообитаний в субоптимальные при подъеме из ниже лежащих высотных поясов в выше лежащие. Более того, «сдвиг» в какой-то части гнездового ареала в вертикальном направлении позволяет некоторым видам далее расселяться вновь по горизонтальной поверхности, но уже в пределах более высоко эшелонированных ландшафтов. Например, золотистая ржанка, широко распространенная в тундровой зоне Таймыра (Рогачева, 1988; Nowak, Pavlov, 1995; Hötker, 1995), резко повышает вертикальный уровень своего ареала у северного склона плато Путорана, и на высоте около 1000 м распространяется по гольцам далее к югу в горизонтальной плоскости (Романов, 1996, 2006; Романов и др., 2007).
У большинства видов птиц в условиях гор Азиатской Субарктики за счет распространения в вертикальной плоскости ареал принимает объемный (трехмерный) характер, с достаточно большой амплитудой между нижней и верхней границей. Пределы вертикального распространения определяются биотопической избирательностью видов, структурой высотной поясности и абсолютными высотами конкретного горного региона.
В различных ГАС верхняя граница вертикального распространения всех видов проходит на различной высоте и поэтому не линейна. В группе видов с наиболее существенной разницей между максимальным и минимальным уровнем верхней границы - глухая кукушка (1030 м), краснозобый конек (1930 м), обыкновенная каменка (1700 м), варакушка (1600 м). Намного меньше эта разница у таловки (600 м), щура (650 м), полярной овсянки (630 м), лапландского подорожника (550 м). Среди модельных видов на максимальную высоту (до 2000 м н.у.м.) поднимаются краснозобый конек, обыкновенная каменка, варакушка, на минимальную (до 1000 м н.у.м.) – весничка, полярная овсянка, лапландский подорожник.
Последовательное изменение высотных пределов распространения многих видов в цепи гор Азиатской Субарктики прослеживается как в широтном так и в меридиональном направлениях. В более высоких широтах (на Полярном Урале, Анабарском плато) в связи с более низким общим положением границ высотно-ландшафтных поясов верхние границы вертикального распространения видов расположены ниже. Изменение вертикальных пределов распространения с запада на восток у многих видов происходит достаточно сходно: основное увеличение вертикальной составляющей ареалов большинства видов происходит в восточном направлении, что соответствует более высокому положению ландшафтных поясов в горах в условиях резко континентального климата. Нижняя граница вертикального распространения обычно совпадает с уровнем подножий горных склонов. Однако в ряде случаев она может проходить и значительно выше этого уровня, и тогда ареал на схеме сечения своего трехмерного пространства как бы «зависает» на определенной высоте. Такой приподнятый уровень нижней границы вертикального распространения у некоторых видов можно наблюдать почти на всем протяжении цепи гор Азиатской Субарктики, у других - лишь в отдельных ее регионах. Подъем нижней границы почти всегда ведет к сужению вертикального распространения. Исключением является лишь характер вертикального распространения щура в Колымском нагорье. В ряде случаев экологические условия региона резко ограничивают возможности вертикального распространения некоторых видов, в результате чего нижняя и верхняя граница занимаемого ими жизненного пространства почти совпадают, и тогда на схеме трехмерного сечения ареала можно наблюдать лишь уровень их пребывания в горах. Например, у лапландского подорожника на Приполярном Урале, Анабаре и хребте Кулар на севере Якутии.
Уровень нижней границы вертикального распространения лежит значительно выше уровня подножий: во-первых, у арктических видов (лапландский подорожник, пуночка), а во-вторых – у различных видов в восточной половине пояса ГАС (от Верхоянского хребта до Корякского нагорья). Это объясняется тем, что в рассматриваемых нами пределах Субарктики, пуночка повсеместно связана исключительно с горным ландшафтом, а лапландский подорожник в регионах с господством у подножий северной тайги образует изолированные, лежащие высоко в гольцах, горно-тундровые популяции. На востоке цепи ГАС (Колымское и Корякское нагорья) «отрыв» нижней границы распространения от горных подножий у ряда видов происходит в следствии того, что оказываются смещены вверх по высотному профилю наиболее оптимальные для них местообитания. Это относится, например, к обыкновенной каменке связанной с вертикально расчлененным горным рельефом, а также к зеленой пеночке, варакушке, обыкновенной чечевице, щуру связанными на Северо-Востоке Азии в основном с подгольцовыми зарослями кедрового и ольховникового стлаников. Выявленные тенденции можно проследить на примере и других видов, встречающихся почти по всему высотному профилю (в том числе и у подножия гор), но имеющих определенно повышенные показатели обилия в его средней или верхней части. Среди них сибирская завирушка, таловка, соловей-красношейка.
Выявленное у ряда видов на востоке Азиатской Субарктики расширение вертикальной составляющей ареала или «подъем» его нижней границы, вероятно, представляет собой один из промежуточных этапов продолжающегося формирования горного аспекта авифауны ГАС.
Выводы
1. Авифауна гор Азиатской Субарктики формируется в системе общих зонально-ландшафтных и высотно-поясных закономерностей. Сокращение видового разнообразия птиц происходит в северном направлении и с высотой - от подножий к вершинам.
2. Высока общность таксономической структуры и видового состава гнездовой авифауны гор Азиатской Субарктики, насчитывающей 197 видов. Половину авифауны рассматриваемых горных систем формируют виды, широко распространенные на пространстве от Урала до Корякского нагорья. Общность структуры населения птиц поддерживается в горизонтальной плоскости видами, лидирующими по обилию одновременно в нескольких горных регионах Азиатской Субарктики, а в вертикальной – одновременно лидирующими в двух, обычно смежных, высотно-ландшафтных поясах. Наиболее однородная авифауна формируется в горах Якутии и ближайших горных системах, лежащих к западу (плато Путорана) и востоку (Колымское нагорье) от них.
3. В горах Азиатской Субарктики большинство видов птиц населяет широкий диапазон высот, охватывающий, как правило, не менее двух высотных поясов. Ареалы видов принимают объемный (трехмерный) характер. Широкое вертикальное распространение многих видов птиц определяет большое общее биоразнообразие даже в высотных поясах с экстремальными условиями, и как следствие – сохраняет высокую потенциальную возможность успешного эволюционного развития горных сообществ и формирования горной авифауны в целом.
4. Установлено, что в ряду горных систем Азиатской Субарктики, в отличие от сопредельных равнин, центр видового разнообразия смещен восточнее Урала и находится на плато Путорана, что соответствует его расположению в пределах Енисейской зоогеографической границы, сформировавшейся по линии ледникового разрыва авифаунистических комплексов и ареалов видов. На повышение видового разнообразия авифауны Путорана оказывает процесс расселения видов, имевших до оледенения ареалы значительно шире.
5. Горы Азиатской Субарктики, обладая значительной вертикальной дифференциацией ландшафтов формируют высотно эшелонированную систему разнонаправленных путей пространственного распространения многих видов птиц. Благодаря этому по соответствующим ценозам разных высотных поясов происходит взаимопроникновение северных форм на юг, а южных на север, что влияет на повышение уровня видового богатства в процессе формирования авифауны горных стран.
6. В зоогеографическом отношении авифауна гор Азиатской Субарктики имеет специфичные черты за счет видов (n=24), экологически связанных с горными ландшафтами. Сокращение числа горных видов в региональных авифаунах с востока на запад следует признать объективной закономерностью пространственного формирования авифауны в пределах всей цепи гор Азиатской Субарктики.
7. Большинство (70-81%) видов проникающих из сопредельных равнин в горные регионы формируют основу региональной гнездовой авифауны. Равнинные элементы качественно сближают авифауны различных высотных поясов и ландшафтов предгорий. Это определяет высокий уровень сходства авифаун соседних равнинных и горных регионов, а авифаунам последних придает более равнинный экологический облик. Выход горных птиц в предгорья практически отсутствует. Авифауна равнин по отношению к авифауне гор Азиатской Субарктики выступает как «донор», а горная по отношению к равнинной - лишь как «реципиент».
8. На фоне обеднения равнинной авифауны Азиатской Субарктики с запада на восток, доля равнинных видов, проникающих в горные регионы, последовательно возрастает в том же направлении. Многие равнинные виды обычные на большей части Северной Палеарктики при движении на восток поднимаются в горы, расширяя вверх вертикальную составляющую своих ареалов, что можно рассматривать как один из промежуточных этапов продолжающегося формирования горного аспекта авифауны гор Азиатской Субарктики.
9. Разнонаправленные изменения видового богатства при движении от центра горной страны к предгорьям связаны с региональной спецификой литогенной основы. В хребты с большими абсолютными высотами и треугольным профилем в поперечном сечении проникновение многих видов птиц из предгорий сдерживают повышенные углы наклона речных долин. Горные массивы, имеющие в поперечном сечении профиль трапеции, расчлененной глубокими сквозными долинами, позволяют проникать вглубь гор большему числу видов птиц. Эти особенности литогенной основы, способствуя проникновению равнинных видов вглубь горных массивов по лесным местообитаниям в горизонтальной плоскости, не менее эффективно затрудняют этот же процесс в вертикальном направлении.
10. В горах Азиатской Субарктики большее сходство видового состава характерно для горно-лесной авифауны, и значительно меньшее - для подгольцовой и гольцовой. В условиях непрерывности лесных ландшафтов и повсеместной сопряженности горных и равнинных лесов подавляющее большинство видов осваивает их повсеместно. Раздробленность и изоляция участков подгольцовых и гольцовых поясов обусловила формирование более регионально-специфичных авифаун каждого из этих поясов.
11. В цепи гор Азиатской Субарктики положение областей с максимально высоким уровнем общности горно-лесной, подгольцовой и гольцовой авифаун различно. Пространственная обособленность этих областей, а также разные величины уровня общности соответствующих авифаун указывают на автономность формирования авифауны на уровне каждого высотного пояса. Наиболее качественно однородная горно-лесная авифауна сформировалась в горах Якутии и ближайших к ним горных системах - плато Путорана и Колымском нагорье. Взаимно наиболее схожие подгольцовые авифауны формируются в Корякском нагорье и соседних с ним горах Якутии и Колымском нагорье. Наиболее однородная (63%) гольцовая авифауны формируется в северной области цепи гор Азиатской Субарктики - на плато Путорана, Приполярном и Полярном Урале, в горах Якутии.
12. Нами выдвинута гипотеза о том, что ледниковые геоморфологические структуры, образованные поздне плейстоценовыми ледниками, создавая экологические предпосылки обогащения гольцовой авифауны не горными видами, оказывают непосредственное влияние на ее формирование и пространственную неоднородность. Выявлена статистически достоверная пространственная сегрегация сообществ птиц, формирующихся на «гольцовых плакорах» с одной стороны, и в ледниковых долинах – с другой. Арктоальпийские и альпийские виды осваивали горные вершины благодаря выработавшимся у них адаптациям к экстремальным условиям еще в период оледенения, и вероятно представляют более древний элемент гольцовой авифауны. Равнинно-тундровые вселенцы, не имея подобных адаптаций, при проникновении в горы использовали типичные для себя экологические условия, сформировавшиеся на локальных участках перигляциальных ландшафтов. Они обогатили гольцовую авифауну позднее, проникая на вершины гор по участкам былого расположения ледников по мере их интенсивного таяния.
13. В горах Азиатской Субарктики с высотой поступательно сокращаются плотность населения птиц и обилие абсолютного большинства видов. В большинстве горных регионов основное сокращение плотности населения птиц происходит при переходе из подгольцового пояса в гольцовый, и чуть менее существенное - при переходе из лесного пояса в подгольцовый.
14. Плотность населения птиц на уровне гольцового, подгольцового и лесного высотно-ландшафтных поясов сокращается в направлении с запада на восток. Это согласуется с сокращением в этом же направлении общей продуктивности растительного покрова. Корреляция сокращения плотности населения птиц с долготой близка к достоверности для показателей плотности населения в пределах всех высотных поясов.
15. Пространственная динамика плотности населения птиц и обилия большинства фоновых видов в горах Азиатской Субарктики имеет концентрически центробежный характер. Эти показатели имеют тенденцию увеличения от наиболее высоких внутренних районов горных стран, где их величины минимальны, в сторону периферии.
16. Сообщества птиц лесного пояса, по сравнению с гольцовыми и подгольцовыми, более разнообразны, стабильны и однородны в пространстве и времени. Помимо высоких значений коэффициента сходства населения, здесь выявлены минимальные амплитуды видового богатства и плотности населения птиц, относительно равномерное распределение по территории региона более половины видового состава, несущественные провинциальные отличия в составе лидеров.
17. Горы Азиатской Субарктики охвачены глобальным процессом расширения северных границ ареалов птиц, придающим динамизм современному формированию авифауны Северной Евразии. Для ряда видов птиц удалось впервые выявить отчетливо выраженную динамику границ ареалов (в горизонтальной и вертикальной плоскости), пространственный и количественный рост популяционных группировок.
Список опубликованных работ по теме диссертации
Монографии
1. Романов А.А. Птицы плато Путорана. М. 1996. 297 с.
2. Романов А.А. Орнитофауна озерных котловин запада плато Путорана. М. 2003. 144 с.
Статьи в изданиях, влюченных в Перечень ВАК РФ
3. Романов А.А. Сибирский пепельный улит (Heteroscelus brevipes) на плато Путорана // Бюл. МОИП, отделение биол. Т. 113. Вып. 3. М. 2008. С. 12-17.
4. Романов А.А. Динамика расширения ареала синехвостки (Tarsigercyanurus) на севере Средней Сибири // Бюл. МОИП, отделение биол. Т. 114. Вып. 1. М. 2009. С. 22 – 25.
5. Романов А.А. Экология и территориальные связи пискулек (Anser erythropus), гнездящихся на плато Путорана, Средняя Сибирь // Бюл. МОИП, отделение биол., Т. 114. Вып. 3. М. 2009. С. 3 – 10.
6. Романов А.А. Связи птиц с ледниковыми ландшафтами гольцов плато Путорана (Средняя Сибирь) // Вестник Московского государственного областного университета, серия «Естественные науки», № 1. 2010. С. 25-34.
7. Романов А.А. Речные наледи и авифауна горной Субарктики (плато Путорана, Средняя Сибирь) // Вестник Московского государственного областного университета, серия «Естественные науки», № 1. 2010. С. 35-45.
8. Романов А.А., Морозов В.В. К экологии овсянки крошки на плато Путорана. // Бюл. МОИП, отделение биол. Т. 98. Вып. 4. М. 1993. С. 28 – 33.
9. Романов А.А., Рупасов С.В. Орлан–белохвост (Haliaeetusalbicilla) на севере Средней Сибири // Бюл. МОИП, отделение биол. Т. 114. Вып. 4. М. 2009. С. 44 – 49.
10. Романов А.А., Поспелов И.Н. Внутриконтинентальные пространственные связи пискулек (Anser erythropus) горно-субарктических регионов Центральной Палеарктики // Экология, Екатеринбург, 2009. №5. С. 101-104.
Статьи, материалы выступлений и докладов.
11. Романов А.А. Роль речных долин в формировании авифауны тундры и лесотундры. // Сборник науч. трудов ВНИИ природа. М. 1988. С. 113 – 118.
12. Романов А.А. Население птиц юго-восточного побережья Чёшской губы // Сборник науч. трудов ВНИИ природа. М. 1989. С. 157 – 162.
13. Романов А.А. К распределению и численности некоторых видов гусеобразных на острове Вайгач // Растительный и животный мир заповедных островов. М. 1989. С. 78-82.
14. Романов А.А. Зональные и интразональные группировки птиц в тундре. // Взаимодействие организмов в тундровых экосистемах. Сыктывкар, 1989. С. 58
15. Романов А.А. К распространению некоторых видов птиц на плато Путорана. // Орнитология. Вып. 25. 1991. С. 174 – 175.
16. Романов А.А. О залетах некоторых видов птиц на остров Вайгач и Югорский полуостров. // Орнитология. Вып. 25. 1991. С. 174.
17. Романов А.А. Редкие и малоизученные птицы плато Путорана. // Орнитологические проблемы Сибири. Барнаул, 1991. С. 130 – 132.
18. Романов А.А. Анализ орнитофауны гольцового и подгольцового поясов плато Путорана. // Заповедное дело. Научно-методические записки по заповедному делу; вып. 2. М. 1997. С. 33 – 52.
19. Романов А.А. Редкие виды птиц плато Путорана // Вопросы сохранения ресурсов малоизученных редких животных Севера. Материалы к Красной книге. М. 1998. С. 67 – 71.
20. Романов А.А. Птицы плато Путорана (Южный Таймыр) // Природное и культурное наследие Арктики: изучение и сохранение. М. 1998. С. 78 – 79.
21. Романов А.А. Результаты орнитологических исследований на западе плато Путорана в 1999 г. // В кн.: Актуальные проблемы изучения и охраны птиц Восточной Европы и Северной Азии. Казань. 2001. С. 529 – 531.
22. Романов А.А. Особенности осеннего пролёта воробьинообразных в южных районах плато Путорана. // В кн.: Актуальные проблемы изучения и охраны птиц Восточной Европы и Северной Азии. Казань. 2001. С. 528 – 529.
23. Романов А.А. Пискулька на плато Путорана. // Казарка, № 7. 2001. С. 105 – 115.
24. Романов А.А. Орнитофауна котловин озёр Накомякен и Собачье (плато Путорана). // Изучение биологического разнообразия на Енисейском экологическом трансекте. Животный мир. М. 2002. С. 353 – 371.
25. Романов А.А. Лебедь-кликун (Cygnuscygnus) на плато Путорана. // Современное состояние популяций, управление ресурсами и охрана гусеобразных Северной Евразии. / Сб. тезисов докладов международного симпозиума. Петрозаводск. 2003. С. 131 – 132.
26. Романов А.А. Орнитофауна юго-западных районов Путоранского заповедника. // Научно-методические записки комиссии по биоразнообразию (секция заповедного дела) РАН; вып. 11. М. 2003. С. 5 – 33.
27. Романов А.А. Список птиц Путоранского заповедника // Корнеева Т.М., Стишов М.С. Птицы заповедников России / Современное состояние биологического разнообразия на заповедных территориях. М. 2003. С. 100-207.
28. Романов А.А. Новые находки мест гнездования пискульки на юго-западе плато Путорана. Казарка, № 9. 2004. С. 139-153.
29. Романов А.А. Физико-географическая характеристика плато Путорана // Фауна позвоночных животных плато Путорана. М. 2004. С. 18-28.
30. Романов А.А. История исследования орнитофауны плато Путорана // Фауна позвоночных животных плато Путорана. М. 2004. С. 92-95.
31. Романов А.А. Общие особенности орнитофауны плато Путорана // Фауна позвоночных животных плато Путорана. М. 2004. С. 96-112.
32. Романов А.А. Аннотированный список видов птиц плато Путорана // Фауна позвоночных животных плато Путорана. М. 2004. С. 113-299.
33. Романов А.А. Результаты и перспективы по изучению и сохранению пискульки на плато Путорана // тезисы докладов Сибирской зоологической конференции. Новосибирск. 2004. С. 178-179.
34. Романов А.А. Актуальные направления исследований, охраны и рационального использования некоторых видов животных плато Путорана. // Сохранение разнообразия животных и охотничье хозяйство России. (Материалы научно-практической конференции). М. 2005. С. 300-303.
35. Романов А.А. Гусеобразные (Anseriformess) горных районов северо-запада Эвенкии // Гусеобразные птицы Северной Евразии (Тезисы докладов третьего международного симпозиума 6-10 октября 2005 г.). Санкт-Петербург. 2005. С.223-225.
36. Романов А.А. Плато Путорана – важнейший регион сохранения разнообразия птиц Средней Сибири // Материалы Х11 орнитологической конференции. Ставрополь. 2006. С. 217-218.
37. Романов А.А. Видовой состав, численность и ландшафтно-биотопическое размещение птиц в бассейне р. Северной // Изучение и охрана животных сообществ плато Путорана. Сборник научных трудов. М. 2006. С. 9-70.
38. Романов А.А. Фауна и население птиц центральной части котловины оз. Кета // Изучение и охрана животных сообществ плато Путорана. Сборник научных трудов. М. 2006. С. 71-102.
39. Романов А.А. Наблюдения за весенним пролетом птиц в котловинах озер Агата Верхняя и Кета // Изучение и охрана животных сообществ плато Путорана. Сборник научных трудов. М. 2006. С. 103-121.
40. Романов А.А. Список видов птиц плато Путорана // Изучение и охрана животных сообществ плато Путорана. Сборник научных трудов. М. 2006. С. 256-263.
41. Романов А.А Фауна и население гусеобразных (Anseriformes) бассейна р.Северная // Сборник трудов НИИ Сельского Хозяйства Крайнего Севера. Новосибирск. 2007. С. 51-64.
42. Романов А.А. О встречах некоторых видов птиц в западных предгорьях плато Путорана // Биоразнообразие экосистем плато Путорана и сопредельных территорий. Сборник научных трудов. М. 2007. С. 297.
43. Романов А.А. Особенности гнездования и миграций пискулек (Anser erythropus) образующих путоранскую территориальную группировку // Вопросы природопользования на Крайнем Севере: Сборник научных трудов / ГУАП. СПб. 2008. С. 61-73.
44. Романов А.А. Биологическое разнообразие и ландшафты Таймыра. М.: Изд-во «Искра». 2008. 98 с.
45. Романов А.А. Плато Путорана. М.: Изд-во «Искра». 2008. 64 с.
46. Романов А.А. Распространение и динамика численности соколообразных плато Путорана (Средняя Сибирь) // Алтайский зоологический журнал. Барнаул. 2009. Вып. 4. С. 52-57.
47. Романов А.А. Экологические связи птиц с ледниковыми ландшафтами гольцов плато Путорана // Тезисы докл. IV Международной орнитологической конференции «Современные проблемы орнитологии Сибири и Центральной Азии». Улан-Удэ. 2009. С. 178-181.
48. Романов А.А. Речные наледи как фактор формирования авифауны горной Субарктики // Тезисы докл. IV Международной орнитологической конференции «Современные проблемы орнитологии Сибири и Центральной Азии». Улан-Удэ. 2009. С. 181-184.
49. Романов А.А., Карпов В.Н. К характеристике послегнездового периода жизни воробьинообразных на плато Путорана // Изучение биологического разнообразия на Енисейском экологическом трансекте. Животный мир. М. 2002. С. 372 – 379.
50. Романов А.А., Голубев С.В. Орнитологические находки в бассейне р. Котуй // биоразнообразие экосистем плато Путорана и сопредельных территорий. Сборник научных трудов. М. 2007. С. 298-299.
51. Романов А.А., Рупасов С.В. Врановые птицы западной части плато Путорана // Экология врановых в естественных и антропогенных ландшафтах. М. – Ставрополь. 2007. С. 86-87.
52. Романов А.А., Рупасов С.В., Журавлев Е.А., Голубев С.В. Птицы бассейна р. Курейки // Биоразнообразие экосистем плато Путорана и сопредельных территорий. Сборник научных трудов. М. 2007. С. 7-70.
53. Голубев С.В., Романов А.А. Некоторые данные по птицам города Норильска и его окрестностей // Биоразнообразие экосистем плато Путорана и сопредельных территорий. Сборник научных трудов. М. 2007. С. 71-90.
54. Забродина Е.Ф., Куваев В.Б., Ларин В.В., Романов А.А., Сыроечковский Е.Е. Государственный природный заповедник «Путоранский» // Заповедники Сибири, т. 1. М.: «Логата». 1999. С. 90 – 101.
55. Карпов В.Н., Романов А.А. Анализ орнитофауны плато Путорана // Тез. докл. Всесоюз. орнитологическойконф. Витебск. 1991. С. 265 – 266.
56. Romanov A.A. Birds of the Putorana plateau // Heritage of the Russian Arctic: Research, Conservation and International Co-operation. Moscow: Ecopros Publishers. 2000. p. 323 – 328.
57. Romanov A.A. Lesser White-fronted Goose (Anser Erythropus) on the Putorana Plateau (southern Taimyr) // Waterbirds around the world. Abstracts book. Edinburg, UK. 2004. P. 165-166.
58. Romanov A.A., Rupasov S.V. [Locality Report of Arctic Breeding Conditions] The River Severnaya basin, Putorana plateau, Russia // Arctic Birds: Newsletter of International breeding conditions survey. № 6. 2004. P. 13-14.
59. Romanov A.A., Golubev S.V. [Locality Report of Arctic Breeding Conditions] The River Kotuy basin, Putorana plateau, Russia // Arctic Birds: Newsletter of International breeding conditions survey. № 10. 2008. P. 17.
|
