ПЛАН
ВСТУП
РОЗДІЛ І. ДОСЛІДЖЕННЯ ГЕНОМУ ЛЮДИНИ
1.1 Вивчення геному людини в рамках міжнародної програми "Геном людини"
1.2 Будова геному людини
РОЗДІЛ ІІ. МЕТОДИ ДОСЛІДЖЕННЯ ГЕНОМУ ЛЮДИНИ
2.1Гібридизація клітин у культурі
2.2Відбір клітинних гібридів за допомогою методу НАТ
2.3Внутрішньохромосомне картування генів за допомогою хромосомних перебудов
2.4Мікроклітини й ізольовані хромосоми
2.5Картування генів за допомогою ДНК-зондів
РОЗДІЛ ІІІ МЕДИЧНІ ТА ЕТИЧНІ АСПЕКТИ ДОСЛІДЖЕННЯ ГЕНОМУ ЛЮДИНИ
3.1Пошкодження генів і спадкові хвороби
3.2Онкогени
3.3Клонування
ВИСНОВКИ
СПИСОК ВИКОРИСТАНОЇ ЛІТЕРАТУРИ
ВСТУП
XX століття стало століттям найбільших відкриттів у всіх областях природознавства, століттям науково-технічної революції, яка змінила і вигляд Землі, і вигляд її мешканців. Можливо, однієї з основних галузей знання, які будуть визначати вигляд нашого світу в наступному столітті, є генетика. З цією порівняно молодою наукою завжди було зв'язано чимало суперечок і протиріч, але, завдяки останнім досягненням генетики і генної інженерії, цілком може вважатися самостійною дисципліною у таких областях, як дослідження геному людини і клонування. Завдяки цим дослідженням було відкрито широкі перспективи розвитку біотехнологій і лікування різних захворювань.
Проект дослідження генома людини має колосальне значення для вивчення молекулярних основ спадкоємних хвороб, їх діагностики, профілактики і лікування. Варто звернути увагу на те, що за останні десятиліття в індустріально розвинутих країнах частка спадкоємних хвороб у загальному обсязі захворювань значно збільшилася.
Саме спадковістю обумовлена схильність до ракових і серцево-судинних захворювань. У значній мірі це зв'язано з екологічною ситуацією, із забрудненням навколишнього середовища, тому що багато відходів промисловості і сільського господарства є мутагенами, тобто змінюють людський генофонд. З огляду на сучасний рівень розвитку генетики можна припустити, що наукові відкриття майбутнього дозволять шляхом зміни геному адаптувати людини до несприятливих умов зовнішнього середовища. Що ж стосується боротьби зі спадкоємними захворюваннями, то їх лікування шляхом заміни хворих генів на здорові здається реальним уже зараз.
Реклама

Усе це означає, що людина одержить можливість не тільки змінювати живі організми, але і конструювати нові форми життя. У зв'язку з цим виникає цілий ряд серйозних питань.
Тому метою нашої роботи буде: вивчення даних про дослідження геному людини.
Задачі:
1. Зібрати дані щодо робіт, метою яких було вивчення геному людини, провести аналіз даних цих робіт.
2. Встановити методи визначення локалізації генів в ДНК людини.
3. Розглянути можливість використання даних про геном людини в медицині.
РОЗДІЛ І
.
Д
ОСЛІДЖЕННЯ ГЕНОМУ ЛЮДИНИ
1.1
Вивчення геному людини в рамках міжнародної програми "Геном людини"
Наприкінці XX століття генетика впритул підійшла до рішення одного з фундаментальних питань біологічної науки - питання про повну розшифровку спадкоємної інформації про людину.
У реалізації грандіозного проекту по розшифровці генетичного коду ДНК, який отримав назву HUGO (Human Genome Organization) взяли участь 220 учених з різних країн, у тому числі і п'ять радянських біологів. Була створена власна програма "Геном людини", керівником якої став академік Олександр Олександрович Баєв.
Вперше ідея організації подібної програми була висунута в 1986 році. Тоді ідея здалася неприйнятою: геном людини, тобто сукупність усіх його генів містить біля трьох мільярдів нуклеотидів, а в кінці 80-х років витрати на визначення одного нуклеотиду складали біля 5 доларів США. Крім того технології 80-х дозволяли одній людині визначати не більш 100 000 нуклеотидів у рік. Проте, вже в 1988 року Конгрес США схвалив створення американського проекту досліджень у цій області, керівник програми Дж. Уотсон.
Створено апарати, здатні секвенувати до 35 млн. послідовностей нуклеотидів у рік. Одним з важливих досягнень стало відкриття так називаної полімеразной ланцюгової реакції, яка дозволяє з мікроскопічних кількостей ДНК за кілька годин одержати обсяг ДНК, достатній для генетичного аналізу.
Програма приносила корисні результати. Суть робіт полягала в наступному: спочатку проводилось картування геному (визначення положення гена в хромосомі), локалізація деяких генів, а після цього секвенування (визначення точної послідовності нуклеотидів у молекулі ДНК). До 1990 року число ідентифікованих генів досягло 5000, з них картовано 1825, секвеновано - 460. Вдалося локалізувати гени, зв'язані з найтяжкими спадкоємними хворобами, такими, як хорея Гентінгтона, хвороба Альцгеймера, м'язова дистрофія Дюшена, кистозный фіброз і ін.
Реклама

Коли з'явилася програма HUGO, виникли так називані геномні компанії, які зайнялися самостійно зайнялися розшифровкою геному. Як приклад можна привести американську організацію за назвою Institute of Genomic Research (TIGR) або компанію Human Genome Sciences Inc. (HGS). Так у жовтні 1994 Крек, Вентер, глава вищезгаданої компанії TIGR, повідомив, що в розпорядженні його корпорації знаходиться бібліотека з 35000 фрагментів ДНК, синтезованих за допомогою РНК на генах, отриманих лабораторним шляхом. Ці фрагменти порівняли з 32 відомими генами спадкоємних захворювань. Виявилося, що 8 з них цілком ідентичні, а 19 гомологічні. TIGR виявився власником найціннішої наукової інформації, але його керівники заявили, що хімічна будова усіх послідовностей з цієї бібліотеки засекречене і буде зроблене надбанням гласності тільки в тому випадку, якщо за компанією буде визнане право власності на всі 35000 фрагментів.
1.2 Будова геному людини
Гаплоїдний геном людини містить 3∙109
п.н. Повторювані послідовності ДНК складають близько 30%. Кількість копій цих послідовностей у геномі людини варіює від одиниць до декількох тисяч. Інші 70%, тобто приблизно 2∙109
п.н., являють собою "унікальні" послідовності, які присутні у вигляді однієї або одиничних копій. Близько 90% РНК, які транскрибуються з унікальної ДНК (мРНК), не залишає ядро клітини. Тільки 10%, що відповідає в хромосомі 2∙ 108
п.н., транспортується в цитоплазму, де відбувається трансляція. Виходячи з того, що процесована мРНК, яка кодує білок, складається в середньому з 1500 нуклеотидів, можна підрахувати, що людський геном містить інформацію для кодування близько 130000 білків (2∙108
: 1500 = 130000). Часто структурні гени, які кодують ті або інші поліпептиди, утримуються в геномі людини у вигляді декількох копій. Немає точного способу визначення частки таких генів або ступеня їхньої повторюваності. Проте є підстави думати, що число різних поліпептидів, які кодуються геномом людини, знаходиться в діапазоні від 30000 до 100000.
Для оцінки числа структурних генів у геномі людини можна запропонувати наступний підхід. Відома нуклеотидна послідовність фрагмента ДНК величиною 60000 п.н., яка містить гени β-глобінової родини (див. Рис. 1.1).
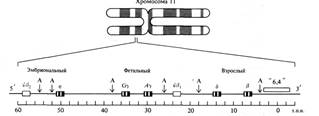
Рис1.1. Родина β -глобинових генів людини розташована в хромосомі 11. Величина представленого фрагмента хромосоми близько 60 т.п.н. Він містить п'ять функціонально активних генів (ε, Gγ
, Gγ,
δ и β) і два псевдогена (Ψβ2 и Ψβ2). Буквою А позначені положення Аlu-повторів-послідовностей, повторених у геномі людини близько 300 000 разів.
Цей фрагмент ДНК входить до складу одинадцятої хромосоми і містить п'ять функціональних структурних генів β, δ, A
γ, G
γ, і ?, які кодують чотири різних поліпептиди (два гени в кодують ідентичні білки). Таким чином, на кожен білок приходиться по 15000 (60000 : 4) пар нуклеотидів. Подібні дані отримані при вивченні генів β-глобінової родини. Фрагмент ДНК величиною 30000 п.н., розташований у шістнадцятій хромосомі, містить три функціональних гена: αl, α2 і ζ,
які кодують два білки, α і ζ,.
І в цьому випадку виходить те ж співвідношення-15000 п.н. на індивідуальний білок. Якщо прийняти цю цифру за середню кількість ДНК людського геному, яка приходиться на один білок, то можна зробити висновок, що гаплоїдний геном кодує 2∙109
: 1,5∙104
= 130000 індивідуальних поліпептидів. Число різних структурних генів може бути трохи менше в тому випадку, якщо середній рівень повторюваності по геному в цілому виявиться вище, ніж у генів глобінової родини.
Картування десятків тисяч генів являє собою надзвичайно важку задачу, хоча її і полегшує те, що деякі гени зібрані в групи, так називані кластери. Кластеризоване розташування мають гени глобінів, білків головного комплексу гістосумісності, імуноглобулінів. Труднощі вивчення генетики людини обумовлені тим, що аналізоване потомство нечисленне, покоління змінюються повільно, а підбор пар, природно, не піддається плануванню. Задача картування людського геному виявилася істотно полегшеною завдяки освоєнню методів роботи із соматическими клітинами.
Ген, відповідальний за колірну сліпоту (дальтонізм), був локалізований у Х-хромосомі в 1911 році. Особливості спадкування генів, зчеплених з Х-хромосомою, дозволили віднести до цієї групи зчеплення більш ніж 100 локусів. Хромосомна локалізація аутосомних генів була вперше проведена в 1968 році. Визначено розташування локусу, який кодує антигени груп крові Даффі, що, подібно антигенам групи АВО й іншим антигенам крові, знаходяться на поверхні еритроцитів. Порівняння спадкування досліджуваного гена з розподілом аберрантної хромосоми показало, що він локалізований у цій хромосомі. З тих пір на підставі аналізу родоводів визначені групи зчеплення для 70 генів людини. Картування багатьох з цих генів стало можливим після того, як було показано їхнє зчеплення з іншими генами, локалізацію яких удалося установити методами генетики соматичних кліток. Прикладом цього служить картування гена резус-фактора, уперше відкритого в 1939 році. У 1971 р. вивчення родоводів показало, що ген Rh
сегрегує счеплено з геном РЕРС,
якийкодує пептидазу С. Роком пізніше при вивченні соматичних клітин ген РЕРС
був локалізований у хромосомі 1. Таким чином, стала відомою група зчеплення і для гена Rh, яка
кодує резус-фактор. Картовано близько 500 аутосомних генів. Переважна більшість цих генів локалізовано методами генетики соматичних кліток.
За останніми даними, Міжнародний консорціум учених заявив про закінчення робіт з розшифровки генома людини. За словами глави Національного інституту по дослідженню геному людини д-ра Фрэнсиса Коллінса, дослідження завершено достроково. Програма по дослідженню послідовності генетичного коду людини, що стартувала в 1990 р., була розрахована на 15 років. У програмі по дослідженню геному людини брали участь учені, зокрема, зі США, Франції, Німеччини, Великобританії, Німеччини, Японії і Китаю.
Згідно отриманим даним, геном людини складається з 3,12 млрд пар нуклеотидів, які формують собою від 35 до 40 тис. генів. Геном може бути розшифрований тільки на 99,99%, оскільки що залишилися 0,01% є індивідуальною складовою кожної людини і не вкладаються в стандартні схеми. Остання доповідь про хід робіт з дослідження геному людини була зроблена д-ром Ф.Колінсом у 2000 р. На той момент вченим удалося розшифрувати геном людини на 97%. Керівник інституту також заявив, що незабаром результати робіт будуть представлені міжнародної наукової громадськості.
РОЗДІЛ ІІ
.
МЕТОДИ ДОСЛІДЖЕННЯ ГЕНОМУ ЛЮДИНИ
2.1 Гібридизація клітин у культурі
У I960 р. було показано, що при спільному культивуванні клітин двох різних ліній вони можуть зливатися, утворювати гібриди, які містять геноми обох батьківських форм. Перші гібридні клітини були отримані при злитті клітин різних ліній мишей, які культивувались Крім внутрішньовидових отримані і міжвидові гібриди, наприклад, між клітинами людини і миші, миші і хом'ячка і навіть миші і курчати.
Утворення гібридних клітин відбувається набагато частіше, якщо в культуру додані деякі речовини, наприклад поліетиленгліколь або інактивовані віруси. Для цієї мети часто використовують вірус Сендай. У вірусів звичайно є один і більш специфічних ділянок, завдяки яким вони можуть зв'язуватися з рецепторами клітини-хазяїна. На поверхні вірусу Сендай таких ділянок небагато. Таким чином, одна вірусна частка здатна утворити місток, з'єднавшись відразу з двома клітинами. Внаслідок дуже малого розміру вірусної частки клітини виявляться надзвичайно тісно зближені. При цьому може відбутися злиття плазматичних мембран клітин і утворитися дикаріон -
клітина з двома ядрами. Потім ядра також можуть злитися з утворенням синкаріону,
якиймістить хромосоми обох батьків. По незрозумілим причинах протягом декількох перших розподілів гібридної клітини відбувається швидка втрата хромосом одного з поєднуваних видів. У клітинних гібридів миша-хом'ячок відбувається втрата хромосом миші. У гібридів кліток миші і людини втрачаються хромосоми людини. Звичайно через 30 поколінь гібридна клітинна лінія миша-людина містить повний хромосомний набір миші і тільки сім людських хромосом. Ця цифра є середньою, деякі клітини містять тільки одну-дві пари людських хромосом, тоді як інші 20. Одна клітинна лінія, хромосоми якої втрачаються після злиття, називається донорною,
а інша - реципієнтною.
Варіабельність втрати хромосом людини в клітинних гібридів мишей-людин полегшує картування людських генів. Для картування генів миші використовують клітинні гібриди миша-хом'ячок. Якщо присутність продукту досліджуваного гена корелює з наявністю якої-небудь однієї хромосоми в гібриді, то цей ген, швидше за все, локалізований у цій хромосомі. Повинні дотримуватися дві умови. По-перше, досліджувана ознака, яка кодується хромосомами людини, повинна чітко (на клітинному рівні) відрізнятися від аналогічної ознаки миші. Наприклад, досліджувана лінія клітин людини містить мутантну лактатдегідрогеназу A (LDH-A). Цей фермент відрізняється від білка, який кодується відповідним мишачим геном. Ці дві форми легко розділяються при гель-електрофорезі. Друга умова, необхідна для картування - можливість ідентифікації даної людської хромосоми, яка присутня у досліджуваній клітинній лінії.
Зчеплення генів у соматичних клітках запропоновано називати синтенією,
від грецького "син" - спільно, "тіні"-підтримувати. Цей термін уведений для того, щоб відрізняти дані про хромосомну локалізацію, отримані в дослідах із соматичними клітинами, від результатів по зчепленню генів, отриманих при аналізі родоводів. Якщо два гени присутні або обоє відсутні в гібридних клітинних лініях, то вони називаються синтеничними.
2.2 Відбір клітинних гібридів за допомогою методу НАТ
Втрата хромосом у гібридів між клітинами миші і людин - це випадковий процес. Те, яка хромосома збережеться і стабілізується в гібридній лінії, випадковий процес. Однак із усієї популяції гібридних клітин можна відібрати стабільні лінії, які містять бажані гени і хромосоми, використовуючи селекційні методи. Ці процедури аналогічні тим, що використовуються для добору певних класів рекомбінантів, які утворяться при схрещуваннях бактерій або дріжджів.
Відсутність у батьківської лінії мишачих клітин якої-небудь істотної функції, наприклад здатності синтезувати необхідний метаболіт, може бути супресовано при внесенні елементів геному людини. З такої гібридної лінії можуть поступово втратитися всі людські хромосоми, крім тієї, котра містить ген, відповідальний за незамінну функцію. Таким чином, можна відібрати гібридні клони, які містять конкретні людські хромосоми. Такий попередній добір полегшує картування генів, розташованих у цій хромосомі.
Один дуже широко використовуваний метод добору гібридних кліток називається НАТ-селекцією. Це назва-абревіатура англійських назв речовин: гіпоксантина, аміноптерина і тимідина, які присутні у селективному середовищі. Аміноптерин блокує синтез пуринів і піримідинів. Для того щоб синтезувати ДНК, клітини повинні реутилізувати пурини і піримідини, які утворяться при деградації полінуклеотидів. В обміні нуклеотидів бере участь велика кількість ферментів. Один з цих ферментів - тимідинкіназа (ТК), яка здійснює перетворення тимідину в тимідинмонофосфат. Якщо у мутантній лінії клітин миші, яка використовується відсутній цей фермент, то на селективному середовищі HAT, в якому відсутній тимідинмонофосфат, можуть рости клітини, які одержали хоча б одну з гомологічних хромосом людини, які містять ген ТК. Цитологічні дослідження показали, що всі клітинні лінії, які відбираються, містять 17 хромосому людини. За допомогою таких клітинних ліній можуть бути картовані й інші гени людини (крім гена ТК),
локалізовані на сімнадцятій хромосомі. Задача зводиться до ідентифікації продуктів генів людини, що напрацьовуються у відібраних гібридних лініях.
Селективне середовище HAT може бути використана для добору клонів, які містять і інші хромосоми. Наприклад, фермент фосфорибозил-гіпоксантин-трансфераза (HPRT) бере участь у синтезі пуринів. Якщо лінія кліток миші втрачає здатність синтезувати HPRT, то цей дефект може бути відшкодований присутністю відповідного гена людини. Показано, що всі гібридні клони, відібрані за цією ознакою, містять людську Х-хромосому, у складі якої знаходиться ген HPRT.
Розроблено інші селективні схеми, що дозволяють проводити картування генів, локалізованих і на інших хромосомах, крім X і 17.
2.3 Внутрішньохромосомне картування генів за допомогою хромосомних перебудов
Ідентифікація індивідуальної хромосоми, у якій знаходиться досліджуваний ген - це тільки перший етап картування. Основною задачею є встановлення порядку генів і їхня точна локалізація. У деяких випадках метод аналізу родоводів дозволяє розташувати на генетичній карті хромосоми три і більш маркерів. Використання більш ефективних методів генетики соматичних кліток може дати більш точну інформацію. Істотна допомога в таких дослідженнях належить хромосомним перебудовам.
Ген, який кодує кислу фосфатазу 1 (ACPI) еритроцитів, розташований на другій хромосомі. При цитологічному вивченні каріотипів двох дітей (із двох різних родин), у яких були множинні уроджені аномалії, далося показати наявність делецій термінальної області короткого плеча другої хромосоми. В одного з дітей делеція хромосоми починалася з термінального району смуги 2р23. Культура клітин, отриманих від цієї дитини, мала ACPI-активність. Ці клітини несли алель А
в одній з гомологічних хромосом і аллель В
в іншій. У другої дитини делеція захоплювала і проксимальний район смуги 2рЗЗ. Клітини, які містять таку хромосому, були позбавлені активності цього ферменту. Таким чином, можна зробити висновок, що ген ACPI
локалізований у смузі 2р23.
У клітинах, які культивуються часто виявляються хромосомні перебудови, відсутні в індивідуумів, які служили донорами вихідних клітин. В інших випадках перебудови, наприклад транслокації, уже присутні в мутантних організмах.
Зміна рівня нагромадження ферменту може допомогти картувати відповідний дуплікований ген. Ген GOT1,
якийкодує розчинну глутамат-оксалацетат—трансаміназу, розташований у 10 хромосомі. Були ідентифіковані дві клітинні лінії, у яких фрагмент хромосоми 10 був транслокований на хромосому 17 або 21. Крім того, у клітин, які досліджувались була збережена і вихідна хромосома 10. Лінія з транслокацією 10/21 характеризується рівнем активності GOT, у порівнянні з контрольним, який має дві нормальні хромосоми і не має дублікованих сегментів. Лінія з транслокацією 10/17 характеризується рівнем активності GOT, приблизно на 50% більшим, ніж в інших ліній. От чому припускають, що лінія з транслокацією 10/17 містить три копії гена GOT. Цей ген повинен бути локалізований на сегменті хромосоми 10, який присутній у транслокації 10/17, але не 10/21.
2.4
Мікроклітини й ізольовані хромосоми
Селекція за типом HAT і інші способи дозволяють одержувати стабільні клітинні лінії, які несуть індивідуальні людські хромосоми. Існують більш прямі методи, які дозволяють одержувати гібридні клітинні лінії, які містять певні компоненти геному людини. Це гібридизація кліток миші з мікроклітинами людини, які несуть неповний геном, і ендоцитоз мишачими клітинами ізольованих хромосом людини.
Мікроклітини
являють собою хромосоми, оточені ділянками ядерної і плазматичної мембран. При рості клітин у присутності колцеміду через кілька днів ядро розпадається на кілька мікронуклеусів, кожний з яких містить тільки одну або кілька хромосом. Ці клітини обробляють цитохалазином і центрифугують. Таким чином, відокремлюється фракція мікроклітин, які являють собою мікронуклеуси, оточені тонким шаром цитоплазми, укладеної між ядерною і плазматичною мембранами. Цитологічні методи дозволяють визначити, які саме хромосоми містять отримані мікроклітини. Плазматична мембрана зберігає рецептори для вірусу Сендай, що дозволяє проводити гібридизацію кліток миші з отриманими мікроклітинами, використовуючи цей вірус. Лінії кліток, які утворяться в результаті гібридизації, несуть визначені людські хромосоми. Гени людини, які експресуються в отриманих гібридних лініях, можуть бути, таким чином, віднесені до визначених хромосом.
Препарат хромосом людини, який не містить домішки клітин, може бути отриманий при відповідній обробці, яка розкриває плазматичну і ядерну мембрани клітини. У сімдесятих роках були розроблені методи фракціонування і проточної мікрофлуориметрії, які дозволяють здійснювати сортування хромосом на фракції, які містять індивідуальні хромосоми людини високої чистоти. Коли вільні хромосоми додають до клітин миші, які культивуються, то ці клітини можуть захоплювати цілі хромосоми в процесі ендоцитозу. Всередині реципієнтної клітини захоплені хромосоми звичайно деградують, розпадаючись на фрагменти. Якщо такі фрагменти містять центромерні області, то вони можуть підтримуватися як єдине ціле. Фрагменти, позбавлені центромери, можуть транслокуватися на мишачі хромосоми. У тому і іншому випадку визначені людські гени будуть експресуваться в гібридних клітинах. Вбудовування фрагментів у хромосому миші відбувається з дуже низькими частотами, від 10-4
до 10-7
на клітину. Фрагменти, які вбудовуються, можуть бути як дуже дрібними, не видимими у світловий мікроскоп, так і досить великими, включаючи плечі хромосом і навіть цілі хромосоми. Такий перенос генетичного матеріалу від донора, який супроводжується вбудовуванням у хромосоми реципієнта, називається трансформацією
(як у бактерій) або трансфекцією.
Картування генів у визначених областях хромосом може бути зроблене в тому випадку, якщо експресію досліджуваного гена можна тестувати у трансформованій клітині, а убудований фрагмент хромосоми ідентифікується цитологічно. При аналізі трансфікованих фрагментів даної хромосоми, які розрізняються за величиною, можна визначити синтенію та порядок генів. Природно, це можливо тоді, коли гени, відсутні в малих фрагментах, виявляються при вбудовуванні фрагментів більшого розміру .
Крім такого прямого підходу, порядок генів і їхнє відносне зчеплення можна вивчати за частотами котрансформації
цих генів. Такі експерименти зводяться до виміру частот спільного вбудовування в геном однієї клітини двох різних генів.
2.5 Картування генів за допомогою ДНК-зондів
Методи картування генів, які обговорювалися в попередніх розділах, були засновані на експресії досліджуваних генів у культурах клітин. Гени, які кодують ферменти клітинного метаболізму і гени, які кодують поверхневі антигени, такі, як білки головного комплексу гістосумісності або антигени групи крові, задовольняють цій умові. Гени, які не володіють подібним фенотипічним проявом, не можуть бути картовані лише з використанням описаних методів. Це обмеження вдається перебороти за допомогою методів генної інженерії. При цьому стає можливим картування будь-яких послідовностей ДНК, для яких можна одержати відповідний ДНК-зонд. Ці методи внесли воістину революційний переворот у генетичний аналіз гібридів соматичних кліток.
Перше застосування методів роботи з рекомбінантними ДНК для картування генів людини включало ДНК-гібридизацію в розчині. Таким чином, вдалося картувати гени α- і β-глобінів. Зонди, які представляють собою комплементарні ДНК (кднк) цих генів, були отримані за допомогою зворотної транскрипції очищених α- і β-глобінових матриць. Ці зонди у певних умовах не взаємодіють один з одним або з глобіновими генами миші, але утворять стабільні дуплекси при обжизі з відповідними послідовностями ДНК людини. Глобінові кДНК-зонди використовували для тестування в розчині препаратів ДНК, виділених з різних гібридних клонів. Утворення стабільного дуплекса між кДНК-зондом і ДНК досліджуваного препарату свідчило про присутність людських глобінових генів у геномі відповідної гібридної лінії. Каріологічний аналіз виявлених у такий спосіб гібридних клонів дозволив зробити висновки, що гени β-глобінової родини розташовані в хромосомі 11, а α-глобінові в хромосомі 16.
Для проведення гібридизаційного аналізу в розчині потрібно велика кількість ДНК. Додаткові труднощі пов'язані також з можливістю перехресної гібридизації між гомологічними генами людини і миші. Більшість цих труднощів можуть бути подолані за допомогою блот-гібридизації за Саузерном. На першому етапі відповідний ДНК-зонд мітять радіоактивними ізотопами (такими, як 32
Р або 3
Н) за допомогою нік-трансляції. Потім з гібридних ліній клітин, які несуть визначені хромосоми людини, виділяють препарати тотальної ДНК. Препарати ДНК обробляють рестрикуючими ендонуклеазами, а отримані фрагменти ДНК розділяють гель-електрофорезом, денатурують і переносять на нітроцелюлозні мембранні фільтри. На фільтрах здійснюється гібридизація з тим або іншими міченими ДНК-зондами. Для більшої ефективності можна розділяти ампліфіковані фрагменти хромосомної ДНК. Для цього використовують їхнє попереднє клонування на фагових векторах.
Після гібридизації фільтри накладають на рентгенівську плівку, на якій після прояву виявляються смуги, які відповідають рестрикційним фрагментам, які містять послідовності ДНК, гомологічні даному зондові . Гени співвідносять з певними хромосомами людини на підставі кореляції між присутністю даної хромосоми в гібридній клітинній лінії і наявністю фрагмента людської ДНК, який гібридизується на радіоавтограмі. Даний метод більш чуттєвий, ніж гібридизація ДНК у розчині і, головне, більш специфічний.
Метод гібридизації по Саузерну, поєднаний з підходами, розробленими для генетичного аналізу соматичних клітин, з успіхом застосовувався не тільки для картування визначених генів, але і для локалізації послідовностей ДНК із невідомими функціям, знайденими в бібліотеках генів людини.
РОЗДІЛ ІІІ
.
МЕДИЧНІ ТА ЕТИЧНІ АСПЕКТИ ДОСЛІДЖЕННЯ ГЕНОМУ ЛЮДИНИ
3.1 Пошкодження генів і спадкові хвороби
Варто також звернути увагу на питання, зв'язані з діагностикою, профілактикою і лікуванням спадкоємних захворювань, де аналіз генома стає усе більш діючим і надійним способом визначення хвороби.
Якщо існує тест, що з достатнім ступенем точності дає можливість виявити те або інше захворювання, то чи буде цей тест обов'язковим для всіх, або тестування буде залежати від бажання пацієнта?
З 10 тис. відомих захворювань людини близько 3 тис. спадкові хвороби. Вони необов'язково успадковуються (передаються нащадкам). Просто викликані вони порушеннями спадкоємного апарата, тобто генів (у тому числі в соматичних клітинах, а не тільки в статевих). Виявлення молекулярних причин "поломки" генів найважливіший результат проекту.
Спадковість безпосередньо зв'язана з продовженням роду, але і тут виникає питання про бажання і готовність чоловіка і жінки пройти обстеження перед тим, як завести дитини. Якщо, скажемо, обоє дружина є носіями гена спадкоємного захворювання, то з 25% імовірністю патологічний ген проявиться в дитини. У такому випадку проведення пренетальної діагностики, зв'язаної з виділенням кліток плоду з крові матері, просто необхідно, але тоді, у випадку виявлення патології плоду в свою чергу встає питання про можливості і навіть обов'язковості аборту.
Число вивчених хвороботворних генів швидко росте, і через 34 роки ми пізнаємо всі 3 тис. генів, відповідальних за ті або інші патології. Це допоможе розібратися в генетичних програмах розвитку і функціонування людського організму, зокрема, зрозуміти причини раку і старіння. Знання молекулярних основ захворювань допоможе їхній ранній діагностиці, а виходить, і більш успішному лікуванню. Адресне постачання ліками уражених кліток, заміна хворих генів здоровими, керування обміном речовин і багато інших мрій фантастів на наших очах перетворюються в реальні методи сучасної медицини. Одним з таких є генотерапія.
Основна частина робіт з генотерапії спадкових і інших захворювань орієнтована на одержання коригуючих послідовностей і векторів, їхній перенос і вбудовування в клітини-реципієнти. Для цієї мети випробуються плазмідні і вірусні вектори, балістичні мікроінфузії, трансплантація клітин, оскільки за наявним даним відомо, що для терапевтичного ефекту досить всього 5 - 10% нормально функціонуючих клітин. Серед можливих векторів для доставки коригуючих ДНК до клітин і тканин-мішеней при гемотерапії муковісцидозу розглядаються вірусні, плазмідні, ліпосомні і пептидні конструкції. Однак до клінічних іспитів потрібно ще вирішити непрості питання взаємодії генетичних препаратів з клітинами, стійкості ефекту.
Перспективним методом генотерапії вважається технологія ex vivo, коли клітинний матеріал, взятий від пацієнта, реплантується йому після екстракорпоральної генокорекції. Уже є приклад успішного лікування сімейної форми гіперхолестеринемії з використанням такого підходу.
3.2
Онкогени
Трансформація культур клітин за допомогою ДНК недавно привела до відкриття генів, які беруть участь у канцерогенезі. Ці гени називають онкогенами.
Деякі форми ракових захворювань мають явно виражену генетичну природу. Характер спадкування вказує на те, що захворювання може визначатися одним геном. Як приклад подібного захворювання можна привести ретинобластому, яка наслідується по домінантному типі. Це захворювання виявляється в дитинстві як ракова поразка одного або двох очей, що швидко переходить на мозок, що при відсутності лікування приводить до ранньої смерті хворого. Хоча відомі багато видів ракових захворювань, спадкування яких підкоряється законам менделевського розщеплення ознак, усі вони досить рідкі й охоплюють лише незначну частку людей хворих раком. Більш розповсюджені форми раку не мають такої чітко з'ясованої генетичної природи, хоча схильність до них, імовірно, успадковується.
На клітинному рівні рак, однак, є виразно генетичним захворюванням. Ракова клітка передає свої неопластичні властивості дочірнім кліткам. Цим можна пояснити високу проліферативную активність ракових тканин. Таким чином, перетворення нормальної клітини в ракову зв'язано з якимись генетичними змінами. Це підтверджується експериментами, у яких ракові клітки передавали свої властивості нормальним кліткам у культурі при трансформації останніх хромосомної ДНК із ракових клітин. Іншим підтвердженням генетичної природи рака більш загального порядку служить той факт, що онкогенні віруси, які викликають рак, при перетворенні клітки в ракову виявляються вбудованими в її геном. Таке перетворення клітини або тканини називається раковою трансформацією.
Цей термін не слід плутати з терміном "генетична трансформація", що позначає включення ДНК у геном після улучення вільної ДНК у клітку.
Ретровіруси, які викликають ракову трансформацію, це віруси, які містять РНК. За допомогою ферменту ревертази вони здатні синтезувати ДНК-копії в ході зворотної транскрипції.
Ці ДНК-копії здатні вмонтуватися в геном клітини. Інтегрована копія називається провірусом. Деякі ретровіруси, відомі як активно трансформуючі віруси, високо онкогенні. Вони викликають неопластичні захворювання в заражених ними тварин. У культурі клітин ці віруси викликають трансформацію клітин, яка протікає з високою ефективністю. Близько 20 таких вірусів було виділено з пацюків, мишей, мавп, кішок, курчат і індиків (наприклад, віруси саркоми Харвея і саркоми Малони виділені з пацюків і мишей). Крім генетичної інформації, необхідної для своєї власної реплікації, ці віруси несуть специфічні гени, які називаються онкогенами, які відповідальні за їх здатність викликати ракову трансформацію. Зараз відомо близько 15 генів one,
включаючи ген src
вірусу саркоми Рауса, який уражає курей, ген mos
вірусу саркоми мишей і ген ras
вірусу саркоми пацюків.
Недавні дослідження показали, що онкогени гомологічні по деяких послідовностях ДНК нормальних кліток ссавців, не інфікованих ретровірусами. Це відкриття лежить в основі двох важливих гіпотез, що відносяться до виникнення раку на клітинному рівні. За однією з цих гіпотез онкогени пішли від гомологічних генів, які є у нормальних клітинах. У рамках іншої гіпотези ракова трансформація є наслідком аномальної експресії нормального гена. Остання може спостерігатися не тільки у випадку трансформації, викликаної вірусами, але і при інших формах раку.
Додатковим підтвердженням гомології між онкогенами і послідовностями нормальних кліток служить і той факт, що вони, як правило, кодують аналогічні білки. Наприклад, онкоген вірусу саркоми Рауса кодує тирозиню-специфічну кіназу. Цей білок являє собою фосфопротеїн молекулярної маси 60000. Такий же білок виділяється і з нормальних клітин курчати, хоча в цих клітинах його кількість у 100 разів менше, ніж у ракових. Використання клонуваннях фрагментів онкогенів в експериментах по ДНК-гібридизації виявило гомологію між геном ras
вірусу саркоми Харві і геном із клітин карциноми сечового міхура людини. Аналогічні результати отримані для гена rasк
вірусу саркоми Кірштейна і відповідного гена з клітин карциноми легені. Зараз вважається загальновизнаним, що онкогени утворились в результаті вбудовування мряк, деяких клітинних генів у геном ретровірусів.
3.3 Клонування
Клонування – "одержання ідентичних нащадків за допомогою безстатевого розмноження" По-іншому визначення клонування звучить так "Клонування - це процес виготовлення генетично ідентичних копій окремої клітини або організму". Тобто ці організми схожі не тільки зовні, але і генетичний код, закладений у них, однаковий.
Мабуть, одним з найбільш яскравих досягнень генетики за останній час є експеримент по клонуванню вівці, успішно завершений в лютому 1997 року вченими Рослінського університету в Шотландії під керівництвом Яна Вілмута. Подібні експерименти по клонуванню тварин проводилися і раніш: ще в 70-і роки професорові Гердону з Оксфордского університету вдалося здійснити пересадження ядра й у такий спосіб клонувати жаб, у 1995 році були клоновані пацюки, проводилися експерименти з іншими ссавцями з тією відмінністю, що замість клітин молочної залози використовувалися клітини ембріона. Колін Стюарт, відомий генетик, який працює в Лабораторії дослідження ракових захворювань у Меріленді США, вважає, що успіх Вілмута багато в чому обумовлений тим, що йому удалося виріши проблему відторгнення ядра донорською клітиною, створивши для ядра необхідну живильну оболонку.
Успіх клонування ссавців не залишає сумнівів у тім, що подолання технічних труднощів, зв'язаних із клонуванням людини, - лише справа часу. У зв'язку з цим виникає безліч питань етико-правового характеру, які варто розглянути докладніше.
Думки вчених по питанню дозволу клонування людини розділилися. Сам Ян Вілмут вважає, що клонування людини абсолютно неприпустимо, його французький колега Жан-Француа Матії переконаний у необхідності того, "щоб ООН розробила спеціальні міжнародні обов'язкові норми по біоетиці, враховуючі останні досягнення науки, аж до внесення доповнень у Декларацію прав людини". Саймон Фішел, науковий директор клініки в Ноттінгемі, навпроти, вважає, що "у багатьох відносинах клонування може привести до величезних переваг". З цією ідеєю згодний, наприклад, і російський академік Струнніков.
У Європі вже є законодавча основа для заборони клонування людини - недавно Рада Європи схвалила Конвенцію по правах людини і біомедицині, у яку потрібно буде внести лише деякі доповнення. Цей документ накладає строгі обмеження на можливі зловживання досягненнями медичної і біологічної науки, у квітні 1997 року був відкритий до підписання 40 країнами - членами Ради Європи. У Великобританії прийнятий у 1990 році закон "Про запліднення й ембріологію" забороняє клонування людини з використанням клітин ембріона.
Суперечки з приводу заборони клонування ледве було не привели до згортання проекту Вілмута, але вченим удалося відстояти результати своєї роботи і продовжити дослідження.
Зараз проводяться куди більш людяні експерименти по вирощуванню ссавців, органи яких надалі можна буде пересаджувати людині. Так технологія трансплантації ядра збільшить шанси на успіх при пересадженню людині свинячого серця. Велике значення нових методів для сільського господарства. Доктор Рон Джеймс, науковий співробітник фірми "ППЛ Терапевтікс", яка придбала права на результати роботи Вілмута, вважає цілком реальним клонування елітних порід великої рогатої худоби й інших сільськогосподарських тварин. Клонування може бути застосоване і для порятунку тварин, занесених у Червону книгу, і відновлення лісів, так необхідних для збереження балансу в атмосфері. Нова технологія пересадження ядра спростить створення трансгенних рослин і тварин, тобто організмів, у геном яких внесений який або сторонній ген, що обумовлює ті або інші властивості, наприклад холодостійкість і велика продуктивність, або
вироблення визначених речовин, зокрема рідких лік.
Досить цікава точка зору, яку дотримує Аксель Кан, директор Лабораторії досліджень в області генетики і молекулярної патології при Паризькому інституті молекулярної генетики. У своїй статті, присвяченій можливості клонування людини, вона у першу чергу розглядає соціальні наслідки експериментів у цієї області. Вона вважає, що якщо раніш було можливим лікування спадкоємних хвороб шляхом заміни генів, то нові технології, застосовувані для клонування, відкривають куди більш широкі перспективи. Кан відзначає, що в сучасному суспільстві усе більше людей хоче мати гарантію того, що всі їхній спадкоємні ознаки в точності будуть передані наступному поколінню.
Можливо, що це зв'язано з усе більшою глобалізацією культури і втратою окремими країнами і культурами своєї самобутності. Тим часом проблема, зв'язана з нездатністю мати дітей унаслідок захворювань, визначеного стилю життя або інших причин, у розвинутих суспільствах здобуває усе більше значення. Саме тому технологія штучного запліднення ICSI (intracytoplasmic sperm injection), яка дозволяє парам, не здатним до відтворення, мати дітей, одержала в суспільстві широку підтримку. Що ж стосується технологій, застосовуваних при клонуванні, то вони дають можливість обходитися генофондом тільки одного з батьків, що робить цілком реальним народження дітей навіть у гомосексуальних шлюбах. З цього випливає, що за певних умов суспільна думка може схилитися на користь дозволу клонування людини.
Клонування органів і тканин - це задача номер один в області трансплантології, травматології й в інших областях медицини і біології. При пересадженні клонованого органа не треба думати про придушення реакції відторгнення і можливих наслідків у вигляді раку, який розвився на тлі імунодефіциту. Клонування органів стануть порятунком для людей, які потрапили в автомобільні аварії або які-небудь інші катастрофи, або для людей, яким потрібна радикальна допомога через захворювання літнього віку (зношене серце, хвора печінка і т.д.). Клонування допоможе людям, які страждають важкими генетичними хворобами. Якщо гени, які визначають яку-небудь подібну хворобу, утримуються в хромосомах батька, то в яйцеклітину матері пересаджується ядро її власної соматичної клітки, - і тоді з'явиться дитина, позбавлена небезпечних генів, точна копія матері. Якщо ці гени утримуються в хромосомах матері, то в її яйцеклітину буде переміщене ядро соматичної клітини батька, - з'явиться здорова дитина, копія батька.
ВИСНОВКИ
1. Встановили, що найбільш фундаментальні дослідження геному людини проводились в рамках міжнародної програми "Геном людини".
2. Ознайомились з результатами діяльності цієї програми, основним з яких на даний момент є встановлення фізичної карти геному людини.
3. Детально ознайомились з методами, які застосовуються для вивчення функцій генів, їх розміщенні в хромосомах. Особливу увагу звернули на гібридизацію клітин культур, внутрішньо хромосомне картування та картування за допомогою ДНК-зондів.
4. Розглянуто можливості використання знань про тонку структуру геному людини в медицині.
СПИСОК ВИКОРИСТАНОЇ ЛІТЕРАТУРИ
1. Айала Ф., Дж. Кайгер. Современная генетика. - М.:Мир, 1988. – 368 с.
2. Баев А.А. "Геном человека": некоторые этико-правовые проблемы настоящего и будущего // Человек, 1995, №2.
3. Баев А.А. Ключи от шифра. О программе "Геном человека" // Человек, 1990, №1.
4. Стрельчук С.І. Генетика с основами селекції. – К.: Фітоцентр, 2000. – 288с.
5. Пехов Л.П. Биология с основами экологии. – Спб, 2000. – 679с.
|
