РЕФЕРАТ
АНТИТЕЛА И КЛЕТОЧНЫЕ РЕЦЕПТОРЫ ДЛЯ НИХ
2008
Основная функция специфического иммунного ответа — это специфическое распознавание чужеродных антигенов. В распознавании участвуют молекулы двух разных типов — иммуноглобулины и Т-клеточные рецепторы антигенов. Структурное разнообразие этих молекул, благодаря которому они способны распознавать множество самых разных антигенов, возникает в результате многочисленных генных рекомбинаций.
Иммуноглобулины представляют собой группу гликопротеинов, которые содержатся в плазме крови и в тканевой жидкости у всех млекопитающих. Некоторые иммуноглобулиновые молекулы структурно связаны с плазматической мембраной В-клеток и функционируют как антигенспецифичные рецепторы. Другие присутствуют в плазме или в лимфе как свободные молекулы. Синтез антител осуществляют В-клетки, но для этого необходим контакт с антигеном и вызванное им созревание В-клеток в антителообразующие клетки. К АОК относятся, в частности, секретирующие значительные количества антител плазматические клетки. Мембраносвязанные иммуноглобулины незрелых В-клеток имеют ту же самую антигенсвязывающую специфичность, что и антитела, образуемые зрелыми АОК.
ИММУНОГЛОБУЛИНЫ - ОСОБОЕ СЕМЕЙСТВО БЕЛКОВ
У большинства высших млекопитающих обнаружено пять классов иммуноглобулинов — IgG, IgA, IgM, IgD и IgE, которые различаются по размерам молекул, заряду, аминокислотному составу и содержанию углеводов.
Помимо различий между классами, существует и весьма значительная гетерогенность в пределах каждого класса. Так, по электрофоретическим свойствам иммуноглобулины настолько разнообразны, что встречаются во всех фракциях нормальной сыворотки, от б до у.
Иммуноглобулины - бифункциональные молекулы
Каждый иммуноглобулин выполняет две функции. Одна область его молекулы предназначена для связывания с антигеном, другая осуществляет так называемые эффекторные функции. К ним относится связывание иммуноглобулина с тканями организма, различными клетками иммунной" системы, определенными фагоцитарными клетками и первым компонентом комплемента при активации этой системы по классическому пути.
Принадлежность иммуноглобулина к определенному классу и подклассу определяется структурой тяжелой цепи/
Реклама

Основная структурная единица иммуноглобулина любого класса состоит из двух одинаковых легких и двух одинаковых тяжелых полипептидных цепей, удерживаемых вместе дисульфидными связями. От типа тяжелых цепей зависит принадлежность молекулы иммуноглобулина к тому или иному классу и подклассу. Так, у человека четыре подкласса igGимеют тяжелые цеп» соответственно г1,г2, г3 и г4;все они выявляются иммунохимически как г-цепи, но незначительно отличаются друг от друга.
К 1, 2, 3 и 4 подклассам IgG относится соответственно около 66%, 23%, 7% и 4% общего числа молекул этого класса. Известны также два подкласса IgA, но подклассов IgM, IgD и IgE человека пока не обнаружено. Разнообразие классов и подклассов иммуноглобулинов обусловлено изотипической изменчивостью их молекул.
В процессе эволюции подклассы иммуноглобулинов возникли, по-видимому, позже классов. Поэтому подклассы IgG человека очень сильно отличаются от четырех подклассов IgG, идентифицированных у мыши.
У каждого класса иммуноглобулинов свой набор функций.
Все иммуноглобулины — это гликопротенны; содержание углеводов в них варьирует от 2—3% у IgG до 12—14% у IgM, IgD и IgE.
IgG
. Это главный изотип Ig нормальной сыворотки человека; на его долю приходится 70—75% общего количества сывороточных иммуноглобулинов.Молекула IgG представляет собой четырехцепочечный мономер с коэффициентом седиментации 7S и мол. массой 146 кДа. При этом белки igG3 несколько крупнее белков других подклассов из-за слегка большей по размерам г3-цепи. Иммуноглобулины класса G равномерно распределены между внутри- и внесосудистым пулами и составляют большинство антител вторичного иммунного ответа, а также основную часть антитоксинов. Кроме того, именно IgG обеспечивают невосприимчивость ребенка к инфекциям в первые несколько месяцев жизни. У человека антитела всех подклассов IgG проникают через плаценту в организм плода, создавая напряженный пассивный иммунитет на весь неонатальный период. У млекопитающих тех видов, для которых характерна передача материнского иммуноглобулина потомству только после рождения, например у свиньи, lgG, поступающий с молоком, избирательно проникает из желудочно-кишечного тракта в кровоток новорожденного.
IgM
.
К этому классу относится примерно 10% общего пула иммуноглобулинов сыворотки. Молекула IgM представляет собой пентамер основной четырехцепочечной единицы. Отдельная тяжелая цепь имеет мол. массу -65 кДа, а вся молекула — 970 кДа. Антитела этого класса содержатся преимущественно во внутрисосудистом пуле иммуноглобулинов и доминируют в качестве «ранних» антител, чаще всего при иммунном ответе на сложные по антигенному составу патогенные микроорганизмы.
Реклама

IgA
. Белки этого класса составляют 15—20% общего количества иммуноглобулинов в сыворотке человека, где они более чем на 80% представлены в виде мономера — четырехцепочечной единицы. Однако в сыворотке большинства других млекопитающих IgA присутствует большей частью в полимерной форме, чаше всего как димер четырехцепочечной единицы. IgA — это главный класс иммуноглобулинов серозно-слизистых секретов, таких как слюна, молозиво и молоко, а также отделяемого слизистой оболочки дыхательных и мочеполовых путей.
Секреторные IgAотносятся к подклассу IgA I или IgA2 и представлены в основном димерной формой с коэффициентом седиментации 11S и мол. массой 385 кДа. Они присутствуют в большом количестве в серозно-слизистых секретах, где связаны с другим белком, называемым секреторным компонентом.
IgD
. Этот класс составляет менее 1% всех иммуноглобулинов плазмы, но обильно представлен на мембране многих В-клеток. Биологическая роль данного класса иммуноглобулинов до конца не известна. Предположительно он участвует в антиген-зависимой дифференцировке лимфоцитов.
IgE
. Концентрация этого класса иммуноглобулинов в сыворотке исчезаюше мала, но он выявляется на поверхностной мембране базофилов и тучных клеток у любого человека. Кроме того, IgE сенсибилизированы клетки слизистых оболочек, в частности носовой полости, бронхов и конъюнктивы. Возможно, IgE имеют существенное значение в ангигельминтозном иммунитете, однако в развитых странах с ними чаще всего связан патогенез аллергических заболеваний, например бронхиальной астмы и сенной лихорадки.
СТРОЕНИЕ АНТИТЕЛ
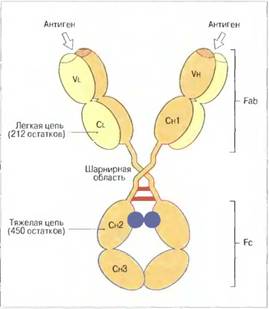
Основная четырехцепочечная структурная единица иммуноглобулиновых молекул образована полипептидными цепями двух разных типов. Меньшие по размерам цепи имеют мол. массу 25 кДа и одинаковы у всех классов, тогда как более крупные цепи, мол. массой 50—77 кДа, структурно различны у разных классов и подклассов иммуноглобулинов. Полипептидные цепи удерживаются вместе ковалентными и нековалентными связями.
Каждая цепь содержит вариабельную и константную области.
У большинства позвоночных легкие цепи существуют в двух различных изотопических формах, обозначенных каппа и лямбда. В молекуле иммуноглобулина могут объединяться пары легких и тяжелых цепей любого типа, но обе цепи в паре относятся к одному типу.
Как установили Хильшманн, Крейг и др., легкие цепи состоят из двух различных областей. С-концевая половина цепи одинакова у легких цепей всех типов; она названа константной, или CL-областью. В то же время Н-концевая половина этой цепи имеет множество вариантов аминокислотной последовательности, из-за чего названа вариабельной, или VL-областью.
Молекулы lgG имеют «типичную» для антител структуру.
В качестве «типичного» антитела можно рассматривать молекулу IgG. В ней имеется две внутрииепочечные днсульфндные связи в каждой легкой цепи — по одной в вариабельной и константной областях — и четыре таких связи в каждой тяжелойцепи, которая вдвое длиннее легкой. Каждая дисульфидная связь замыкает пептидную петлю из 60—70 аминокислотных остатков; при сравнении аминокислотных последовательностей этих петель выявляется удивительно высокая степень их гомологии. В основном поэтому каждая полипептидная цепь иммуноглобулина образует несколько глобулярных доменов с весьма сходной вторичной и третичной структурой.
Пептидная петля, замкнутая дисульфидной связью, — это центральная часть «домена», в котором всего насчитывается примерно 110 аминокислотных остатков. Как в легких, так и в тяжелых цепях первые от Н-конца домены образованы соответственно вариабельными областями Vlи Vh. Тяжелые цепи lgG, IgA н IgD имеют еще три домена — Сн1, Сн2 и Сн8, составляющих константную область. В цепяхнепосредственно за Сн1 следует один дополнительный домен, поэтому С-концевые домены тяжелых цепей IgM и IgE гомологичны СнЗ-домену IgG.
Модель молекулы IgGl с изображением глобулярных доменов тяжелой и легкой цепей. Обратите внимание на взаимное сближение доменов СнЗ и разделение доменов Сн2, между которыми расположены углеводные компоненты. На этом рисунке дисульфидные связи между Н- и L-цепями не показаны.
По данным рентгеноструктурного анализа удалось реконструировать б-углеродный скелет и построить компьютерные модели целых молекул IgG. Модельные IgG имеют вид Y- и Т-образных структур, и аналогичные формы IgG выявлены с помощью электронной микроскопии.
В Fab-области молекулы иммуноглобулина гомологичные домены легких и тяжелых цепей располагаются парами; СуЗ-домены двух тяжелых цепей также образуют пару, но Су2-домены разделены углеводными компонентами.
Несмотря на структурное сходство гомологичных доменов, междоменные взаимодействия в разных парах существенно различаются. Например, вариабельные домены контактируют друг с другом слоями, состоящими из трех сегментов цепи, а константные — слоями из четырех сегментов. Модель молекулы IgGl в общем адекватно отражает структуру элементарных единиц в составе иммуноглобулинов всех изотипов, однако каждый класс и подкласс имеет свои характерные отличия в деталях строения.
IgG
. Четыре подкласса IgG человека лишь слегка различаются по аминокислотной последовательности тяжелых цепей. Этими различиями, относящимися в основном к шарнирной области, обусловлены изотипические вариации расположения и числа межцепочечных дисульфидных связей. Из четырех подклассов наиболее выраженной структурной особенностью — удлиненной шарнирной областью — обладает lgG3, чем объясняется его более высокая мол. масса и, отчасти, повышенная биологическая активность.
IgM
. У человека IgM обычно обнаруживается в виде пентамера основной четырехцепочечной структурной единицы. Отличие его м-цепи от г-цепей IgG состоит в иной аминокислотной последовательности и наличии дополнительного константного домена с С-концевым пептидом из 18 аминокислотных остатков. Субединицы пентамера соединены дисульфидными связями между С^З-доменами и, вероятно, между С-концевыми пептидами. По данным электронной микроскопии молекула IgM имеет плотно сложенный центр, от которого расходятся пять ветвей.
На микрофотографиях антитела IgM, связавшиеся с бактериальным жгутиком, видны в «крабовидной» конфигурации. Такая форма молекулы IgM свидетельствует о том, что тяжелые цепи в области между Cμ2 и CμЗ могут легко изгибаться, хотя по структуре эта область не гомологична шарнирной области IgG. Взаимное расположение разных частей молекулы IgM, принявшей «крабовидную» конфигурацию, по-видимому, связано с активацией ею комплемента.
Молекулу IgM характеризуют еще два свойства: многочисленные присоединенные к м-цепи олигосахариды и добавочная пептидная J-цепь, которая предположительно принимает участие в полимеризации мономерных единиц, предшествующей выходу IgM из синтезирующей его клетки. J-цепь представляет собой полипептид из 137 аминокислотных остатков, образующий домен иммуноглобулинового типа. Каждая молекула IgM содержит только одну J-цепь. Она соединена дисульфидными связями с С-концевыми. состоящими из 18 аминокислотных остатков пептидами тяжелых цепей отдельных мономеров. Имеется наблюдение, что в клетках, секретирующих IgM преимущественно в форме гексамера, отсутствуют свободные J-цепи.
IgA
. Состоящая из 472 аминокислотных остатков б-цепь свертывается с образованием четырех доменов: Vh, Ccd, Сос2 и СосЗ. Аналогично IgM тяжелая цепь IgA содержит дополнительный С-концевой пептид из 18 аминокислотных остатков с остатком цнстеина в предпоследней позиции. Этот остаток способен ковалентно взаимодействовать с J-цепью, соединяющей две молекулы с образованием димера. На электронных микрофотографиях димеры IgA выглядят как двойные Х-формы, что свидетельствует о соединении двух мономерных субъединиц конец-в-конец и об участии в этом соединении С-концевых областей СаЗ.
Секреторный IgAпредставлен главным образом димерной формой с коэффициентом седиментации 11S. Полностью собранная молекула состоит из двух мономеров IgA. одного секреторного компонента и одной J-цепи. Как все эти пептидные цепи связаны между собой, до конца не ясно. В противоположность J-цепи секреторный компонент синтезируется не в плазматических, а в эпителиальных клетках. Молекулы IgA, удерживаемые в димерной конфигурации J-цепью и секретируемые субэпителиальными плазматическими клетками слизистых оболочек, при прохождении через эпителиальный покров активно связывают секреторный компонент. Он способствует доставке антител slgA в выделения организма, а также защищает эти антитела от протеолиза.
Преобладающий подкласс IgA, как в сыворотке, так и в выделениях организма, — это IgAl. Однако в просвете толстой кишки около 60% IgA составляет подкласс lgA2. Многие бактерии в микрофлоре верхних дыхательных путей, приспособленные к условиям обитания, выделяют протеазы, расщепляющие IgAl.
IgD
. К IgD относится меньше 1% иммуноглобулинов сыворотки. Этот белок гораздо более чувствителен к протеолизу, чем IgGI, IgG2, IgA или IgM и, кроме того, проявляет тенденцию к спонтайному протеолнзу. По-видимому, его д-цепи удерживаются вместе всего одной дисульфидной связью и соединены с большим количеством углеводых цепей.
IgE
. Молекула IgEсостоит из более крупных е-цепей, содержащих большее число аминокислотных остатков и образующих пять доменов.
МЕЖДУ МОЛЕКУЛАМИ АНТИТЕЛ ВОЗМОЖНЫ ИЗОТИПИЧЕСКИЕ, АЛЛОТИПИЧЕСКИЕ И ИДИОТИПИЧЕСКИЕ РАЗЛИЧИЯ В АМИНОКИСЛОТНОЙ ПОСЛЕДОВАТЕЛЬНОСТИ
Изотпические различия. Гены изотипических вариантов антител в норме присутствуют в геноме всех представителей данного вида. Например, в геноме каждого человека имеются одновременно гены гА-, г2-, г3-, г4-, м-, бъ-, б2-, д-, е-, к- и л-цепей, представляющих собой поэтому изотипы.
Аллотипические различия. Они отражают генетические различия между особями одного вида, в том числе по аллелям данного локуса. Например, у человека вариант IgG3 с остатком фенилаланина в 436 позиции уЗ-цепи, названный G3m, встречается не у всех индивидов, т. е. представляет собой аллотип. Чаще всего аллотипы различаются константной областью тяжелых цепей.
Идиотипические различия. Существование разных идиотипов антител обусловлено различиями в аминокислотной последовательности вариабельных доменов, и особенно их гипервариабельных участков. Эти же области определяют специфичность антнгенсвязывающего центра антител. Индивидуальные идиотипы антител обычно характерны для разных клонов В-клеток. Однако иногда у различных В-клеточных клонов идиотипы антител могут быть общими.
ЭФФЕКТОРНЫЕ ФУНКЦИИ АНТИТЕЛ
Основная, первичная функция антител — связывание с антигеном. В некоторых случаях оно непосредственно ведет к достижению эффекта, например, обеспечивая нейтрализацию бактериального токсина или предотвращая проникновение вируса в клетки. Однако чаще взаимодействие антител с антигеном остается безрезультатным, пока они не осуществят свои вторичные, «эффекторные» функции.
Один из наиболее важных эффекторных механизмов действия IgGl и lgG3 состоит в активации системы комплемента — группы особых сывороточных белков, принимающих участие в воспалительных реакциях. Связываясь с антигеном. IgM, IgGl и lgG3 способны активировать каскад протеолитических реакций, осуществляемых системой комплемента. Менее эффективен в этом отношении lgG2; антитела же lgG4, IgA, IgD и IgE не активируют комплемент.
К зффекторным функциям иммуноглобулинов относится также их избирательное взаимодействие с различными типами клеток при участии специальных рецепторов клеточной поверхности.
КЛЕТОЧНЫЕ РЕЦЕПТОРЫ ДЛЯ АНТИТЕЛ
Существует три типа рецепторов клеточной поверхности для IgG
Клеточные рецепторы для IgG опосредуют ряд эффекторных функций антител. Перекрестная сшивка антигеном антител Ig, связанных с рецепторами, инициирует ту или иную биологическую активность клетки, причем разные рецепторы могут индуцировать одни и те же активности, среди которых главные — фагоцитоз, зависимая от антител клеточная цитотоксичность, высвобождение медиаторов и презентация антигена.
На поверхности клеток выявлены к настоящему времени три группы рецепторов IgG человека: FcyRI, FcyRIIи FcyRIII. Все они имеют внеклеточные домены, в значительной степени гомологичные V-областям иммуноглобулинов, т. е. относятся к иммуноглобулиновому суперсемейству молекул, как и специфичный для IgA рецептор FcaR.
Свойства и распределение рецепторов для IgG
Рецептор FcyRIна клетках человека связывает с высоким сродствоммономерный IgG и имеет более ограниченное распространение, чем другие рецепторы.
Рецептор FcyRIIэкспрессируют очень многие клетки, часто как единственный рецептор Ig. Он связывает с низким сродством IgG, причем только в составе иммунных комплексов или агрегатов.
Рецептор FcyRIIlобильно гликозилирован и экспрессируется в формах с мол. массой от 50 до 80 кДа. В форме FcyRI Па его экспрессируют макрофаги, НК-клетки и некоторые Т-клетки; эта форма связывает мономерный, а также включенный в иммунный комплекс IgG с аффинностью 3 ч ЙП7 М"1. В форме FcyRIIIb, связанной с клеточной мембраной посредством GPI, этот рецептор экспрессируют только гранулоциты; FcyRIIIb связывает IgG с низким сродством.
Дополнительное разнообразие клеточных рецепторов для IgG
Три типа Fcy-рецепторов встречаются в 12 различных изоформах, и, кроме того, описан генетический полиморфизм для FcyRII и FcyRIIl. Наряду с этой внутренней гетерогенностью существует, как установлено, и другая, обусловленная экспрессией данных рецепторов на клеточной поверхности в виде комплексов с полипептидными цепями иного происхождения. Два типа таких цепей идентифицированы. Они соединяются с рецепторами разных типов:
• FcyRI, аналогично FceRI, образует комплекс с димером изу-цепей, соединенных дисульфидной связью,
• FcyRIIla может ассоциировать с теми же димерами г-цепи, либо с димерами ж-цепи, либо с гетеродимерами из у- и ж-цепей.
Ассоциированные цепи, вероятно, обеспечивают экспрессию рецепторов на плазматической мембране и, кроме того, важны, по-видимому, для передачи сигнала внутрь клетки. В случае Fcylllb, заякоренного в мембране при помощи GPI, вероятно нет необходимости ни в г-, ни в ж-цепях.
IgE связывается с двумя различными клеточными Fce-рецепторами.
В настоящее время известны два различных клеточных рецептора для IgE. На тучных клетках и базофилах обнаружен высокоаффинный «классический» рецептор FceRI, относящийся к иммуноглобулиновому суперсемейству молекул. Существенно отличный от него низкоаффинный Fc-рецептор для IgE выявлен на лейкоцитах, в том числе и на лимфоцитах. Низкоаффинный рецептор не принадлежит к иммуноглобулиновому суперсемейству, но в значительной степени гомологичен некоторым лектинам животных, например лектину, связывающему маннозу.
FceR/ - высокоаффинный клеточный рецептор к IgE
Молекула FceRI образована 4 полипептидными цепями. Входящая в ее состав гликозилированная ос-цепь экспонирована над поверхностью клетки. Антитела к б-цепи способны блокировать связывание IgE с этим рецептором и самостоятельно индуцировать высвобождение гистамина из лейкозных базофилов крысы. Углеводные компоненты б-цепи, вероятно, защищают ее от действия сывороточных протеаз, но не имеют значения для связывания IgE и обусловленного IgE высвобождения гистамина.
Одна в-цепь и две соединенные дисульфидной связью г-цепи — это важные компоненты ару2-рецепторной единицы. Они необходимы для экспрессии рецептора на поверхности клетки и, вероятно, для трансмембранной передачи сигнала.
Рецептор FceRI взаимодействует с С-концевой частью тяжелых цепей IgE, а именно с Се2-и/или СеЗ-доменами. Это связывание высокоспецифично и характеризуется очень высокой константой. Однако ни взаимодействие рецептора с одновалентным IgE, ни связывание специфического лиганда одной молекулой IgE, по-видимому, не активирует тучные клетки или базофилы, поскольку при этом не происходит высвобождения гистамина. Для начала дегрануляции необходимо, чтобы несколько соединенных с клеточной поверхностью молекул IgEбыли перекрестно связаны антигеном или другими лигандами, вызывающими дегрануляцию.
Углеводные компоненты молекулы IgE, вероятно, не участвуют во взаимодействии с FceRI; они важны для секреции IgE В-клетками.
Раньше считалось, что высокоаффинные рецепторы для IgE имеются только на тучных клетках и базофилах, но недавно установлено, что они могут присутствовать также на клетках Лангерганса и на стволовых клетках.
FcfR// - низкоаффинный клеточный рецептор для IgE
Лимфоцитарный FceRII человека, или антиген CD23, содержит типичный для мембраносвязанной молекулы трансмембранный домен, но расположен в мембране необычно — «вверх ногами», т.е. С-концом снаружи клетки. В отличие от других Fc-рецепторов он относится не к иммуноглобулиновому суперсемейству молекул, а к филогенетически более древнему суперсемейству животных лектинов.
К настоящему времени идентифицированы, клонированы и секвенированы две формы FceRIIчеловека, одинаковые по структуре внеклеточных доменов, но различные в Н-концевой, цитоплазматической области. Рецептор FceRlla постоянно присутствует в норме на В-клетках, тогда как экспрессия FceRIIb индуцируется цитокином ИЛ-4 на Т-клетках, В-клетках, моноцитах и эозинофилах. Часто эта экспрессия бывает повышенной на В-клетках и моноцитах у больных экземой и на лимфоцитах при сенной лихорадке.
АНАЛИЗ СТРУКТУРЫ И ФУНКЦИИ АНТИТЕЛ
Получение протеолитических фрагментов для анализа структуры и функции антител
Растительная протеаза папаин расщепляет молекулу IgG в шарнирной области между Cyl - и Су2-доменами на два идентичных антигенсвязывающих фрагмента Fabи один способный кристаллизоваться фрагмент, Fc. Изучение этих фрагментов существенно помогло установлению структуры и функций антител, поскольку дало возможность отделить область Fab, взаимодействующую с антигеном, от области Fc, ответственной за эффекторные функции, например за фиксацию комплемента, связывание с моноцитами или проникновение через плаценту.
При продолжительном папаиновом гидролизе от Fc-фрагмента отщепляется фрагмент Fc', состоящий из двух неполных СуЗ-доменов. Другой фермент, часто применяемый в структурно-функциональных исследованиях, — пепсин. Он расщепляет молекулу IgG на два крупных фрагмента — F2, включающий в себя целиком обе Fab-части, соединенные в шарнирных областях тяжелых цепей, и pFc', который соответствует двум СуЗ-доменам этой молекулы.
Молекулу IgG расщепляют и многие другие ферменты. Так, кратковременное воздействие трипсином на обработанные кислотой Fc-фрагменты позволяет выделить Су2-домен для последующего структурно-функционального сопоставления его с другими субфрагментами молекулы IgG, в частности с pFc'.
Связывание антителами разнообразных антигенов обеспечивают гипервариабельные последовательности антигенраспознающих центров.
Аминокислотная последовательность некоторых коротких участков вариабельной области тяжелых и легких цепей чрезвычайно изменчива. В легких цепях эти сегменты, названные гипервариабельными, находятся вблизи 30, 50 и 95 позиций. Их называют также участками, определяющими комплементарность, поскольку именно они образуют антигенсвязывающие центры молекулы антитела. Последовательности между гипервариабельными сегментами названы каркасными. В V-области каждой легкой и тяжелой цепи имеется по три CDRи по четыре FR.
Вариабельные области легких и тяжелых цепей пространственно свернуты так, что гипервариабельные участки находятся близко друг к другу и образуют на поверхности молекулы структуру, связывающую антиген. Такие сегменты располагаются чаше всего на изгибах пептидной цепи.
В структуре антител идентифицировано много участков, отвечающих за эффекторные функции.
Если локализация антигенсвязывающих центров была установлена очень быстро, то последовательности, ответственные за большинство эффекторных функций, долгое время не удавалось точно локализовать. Некоторые предварительные данные были получены в опытах по ингибированию функциональной активности антител их субфрагментами, образующимися при ферментативном расщеплении, однако дело продвигалось медленно, пока не стало возможным применение метода сайт-направленного мутагенеза, который позволяет вызывать избирательно замену различных аминокислотных остатков в известной пептидной последовательности и таким образом определять значение различных остатков для осуществления функции.
Прежде всего, этот метод был использован для изучения механизма активации комплемента антителами IgG. К тому времени уже было известно, что Clqвзаимодействует с Су2-доменом IgG. С помощью сайт-направленного мутагенеза удалось выявить участок связывания Clq — боковые цепи трех аминокислотных остатков в Су2-домене, Glu-318, Lys-320 и Lys-322. Эта последовательность, по-видимому, типична для участков взаимодействия молекул IgG с Clq.
В случае IgM механизм активации комплемента, вероятно, иной. Свободный циркулирующий IgM в звездообразной конфигурации не способен, очевидно, активировать комплемент, но приобретает эту способность после связывания с антигеном. По предположению Файнстайна и др., при связывании с полимерным или перекрестносвязанным антигеном Р2_единицы IgM отклоняются от плоскости своего исходного положения так, что пентамер приобретает «крабовидную» конфигурацию, вполне различимую при электронной микроскопии. Эти конформационные изменения, вероятно, обнажают кольцо сайтов для связывания Clq, скрытых при звездообразной конфигурации пентамерной молекулы IgM из-за тесного сближения соседних мономеров. Участок связывания Clqнаходится в CμЗ-дoмeнe, причем его структурная локализация аналогична локализации возможного сайта той же специфичности в Су2-домене.
Молекулы IgG взаимодействуют с разнообразными клеточными Fc-рецепторами. По данным исследований с применением сайт-направленного мутагенеза, высокоаффинный рецептор FcyRI на моноцитах связывается со структурным мотивом г-цепи, расположенным вокруг остатка лейцина в позиции 235, между Су2-доменом и шарнирной областью.
Не так давно раскрыт механизм взаимодействия IgG материнского молока с FcRn, экспрессированным на кишечном эпителии новорожденного крысенка; предполагается, что он близок к механизму связывания материнского IgG человека с hFcRn — плацентарным аналогом FcRn. Сайт в составе Fc, связывающий
FcRn, находится на стыке доменов Сн2 и СнЗ. перекрывая участок взаимодействия со стафилококковым белком А. Вероятно, основное функциональное значение в этом участке имеют три или четыре остатка гистидина: по-видимому, от них зависит связывание IgG с FcRn при рН 6,5 и его высвобождение при рН 7,5.
Для идентификации сайтов молекулы IgE, связывающихся с FceRI тучных клеток или с FceRII В-клеток, были применены методы генетической инженерии. Синтез рекомбинантных пептидов, соответствующих тем или иным отрезкам последовательности е-цепи, и сопоставление их ингибируюшего влияния на взаимодействие IgE с клеточными рецепторами позволили установить, что связывание IgE с FceRI, по-видимому, опосредовано пептидом из 76 остатков между Се2 и СгЗ. тогда как FceRlI, вероятно, распознает структуру, состоящую из аминокислотных остатков СгЗ-доменов обеих е-цепей.
Получены также некоторые данные о топографии взаимодействий между Fc-частью молекулы lgG и белком AStaphylococcusaureus. Сайт связывания находится, предположительно, в области соединения доменов Су2 и СуЗ IgG.
|
