Міністерство освіти і науки України
Житомирський державний університет імені Івана Франка
Кафедра ботаніки
Реферат
з Біології
ТЕМА: “Локалізація та розподіл ферментних систем у рослинній клітині”
Виконала:
студентка ІІІ курсу 32 групи
природничого факультету
спеціальність «Біологія. Хімія»
Чернишенко Лілія Юріївна
Перевірила:
доцент кафедри ботаніки, к.б.н.
Перепелиця Л.О.
Житомир – 2010
План
Вступ
1 Історія дослідження ферментів
2 Структура і механізм дії ферментів
3 Функції ферментів
4 Класифікація ферментів
5 Локалізація ферментів у клітині
6 Створення нових ферментів
Висновки
Список літератури
Вступ
Актуальність теми:
тема вивчається для глибокого розуміння фізіологічних процесів, що протікають в рослинній клітині. Знання теми необхідні для кращого сприйняття інших тем, наприклад, «Фотосинтез», «Біосинтез білку». Адже ферменти беруть участь у всіх хімічних реакціях, що відбуваються в живому організмі.
Мета:
викласти матеріал, що стосується ферментних систем рослинної клітини; показати роль ферментів в процесах життєдіяльності; з’ясувати їх функції та будову.
1 Історія дослідження ферментів
Вивчення ферментів почалось з дослідження головним чином двох біологічних процесів – перетравлення їжі тваринами та бродіння цукру з утворенням спирту під дією дріжджів. Можна вважати, що вперше дію ферментів спостерігав у 1833 році А. Пайєн та Ж. Персо, які встановили, що спирт осаджує з солоду якусь речовину, що здатна перетворювати крохмаль в цукор. Пайєн і Персо назвали це активне начало діастазою (від грецького «роз’єднання») і показали, що воно руйнується при нагріванні. Введений термін «діастаза» застосовували протягом деякого часу в якості загальної назви для усіх ферментів. Але на перших етапах цього дослідження було встановлено, що бродіння під дією дріжджів представляє собою каталітичний процес. Звідси і виникло поняття фермент (від латинського fermentum – бродіння, закваска).
В кінці 19 століття між хіміком М. Бертло та батьком мікробіології Луї Пастером виникла суперечка про природу бродіння. Бертло вважав, що цей процес обумовлюється утворенням каталізаторів (ферментів) живими клітинами дріжджів. Пастер вважав, що здатність викликати бродіння притаманна тільки живим клітинам дріжджів, що діляться, та ніяк не відокремлена від них.
Реклама

У 1878 році німецький фізіолог В. Кюне запропонував назву «ензим» (від грецького – в «заквасці») не тільки для активного начала дріжджів, а й для інших ферментів. У 1897 році Е. Бухнер встановив дуже цікавий факт. Він показав, що якщо зруйнувати клітини дріжджів, розтираючи їх з піском, та віджати з них сік, цей сік може швидко зброджувати цукор. Це відкриття спричинило кінець дискусії між послідовниками Пастера та Бертло. Було розпочато роботу по виділенню та очищенню ферментів.
В наш час кількість ідентифікованих ферментів становить більше 800. Для того, щоб було відомо про який фермент йде мова, була введена формальна система класифікації та нумерації всіх відомих ферментів. Але ферменти мають ще й «тривіальні» назви, що використовуються в розмові, а також в науковій літературі. Правило для тривіальних назв було запропоновано Е. Дюкло у 1898 році. Згідно з цим правилом назва ферменту утворюється шляхом додавання закінчення –аза до слова (чи його частини), що позначає речовину, на яку діє цей фермент. (Речовина, на яку діє фермент, називається субстратом.) Наприклад, фермент, що діє на цукор, повинен називатися сахаразою; фермент, що діє на жири – ліпазою і т.д.
З часом список ферментів збільшувався і ставало зрозумілішим, що на один і той же субстрат може діяти більш ніж один фермент, але кожен з них діє по своєму. Тому було змінено номенклатуру ферментів так, що кожна назва відображала тип хімічної зміни, що викликається даним ферментом. Таким чином, пропозиція Дюкло зумовила створення простого методу присвоєння ферментам назв, в яких міститься відома інформація про них.
2 Структура і механізм дії ферментів
Активність ферментів визначається їхньою тривимірною структурою. В ранніх дослідженнях було отримано багато доказів того, що ферменти – це білки. Нездатність проходити крізь напівпроникну мембрану, особливості їх розчинності, денатурація при нагріванні і в сильному лужному чи кислотному середовищі – всі ці властивості ферментів співпадають зі звичайними характеристиками білків. В наш час багато ферментів отримано в чистому вигляді та проаналізовано їх будову. У всіх випадках доведено, що вони являють собою білки. Як і всі білки, ферменти синтезуються у вигляді лінійного ланцюжка амінокислот, який згортається певним чином. Кожна послідовність амінокислот згортається особливим чином, і молекула (білкова глобула), що виходить, володіє унікальними властивостями. Кілька білкових ланцюжків можуть об'єднуватися у білковий комплекс.(Рис.1) Найбільші рівні структури білків — третинна та четвертинна структури — руйнуються при нагріванні або під дією деяких хімічних речовин.
Реклама

Рис.1. Просторова структура тріозфосфат-ізомерази
Щоб каталізувати реакцію, фермент повинен зв'язатися з одним або кількома субстратами. Білковий ланцюжок ферменту згортається таким чином, що на поверхні глобули утворюється щілина або западина, до якої приєднуються молукули субстрату. Ця область називається ділянкою (сайтом) зв'язування субстрата. Зазвичай вона співпадає з активним центром ферменту або знаходиться поблизу від нього. Деякі ферменти містять також ділянки зв'язування кофакторів або іонів металів.
У деяких ферментів присутні також ділянки зв'язування малих молекул, що не беруть безпосередньої участі в реакції і часто, але не обов'язково, є субстратами або продуктами метаболічного шляху, в який входить фермент. Вони зменшують або збільшують активність ферменту, що створює можливість для зворотного зв'язку або регуляції роботи ферменту.Для активних центрів деяких ферментів характерне явище кооперативності.
Специфічність
Одна з найбільш характерних властивостей ферментів – це їх специфічність, в міру якої кожен фермент діє тільки на одну речовину або дуже невелику кількість споріднених речовин. Специфічність дії ферментів – найважливіше біологічне явище, без якого неможливий впорядкований обмін у живій природі, відповідно і саме життя. Концентрація каталізатора – розмір його активної поверхні – визначає швидкість змін, що відбуваються під його впливом. Високо специфічні каталізатори визначають які саме речовини підлягають перетворенню. Специфічність каталізаторів завдяки своїй структурі визначає серед багатьох шляхів той єдиний шлях, за яким повинна йти реакція. Він має здатність направляти реакцію по одному шляху. Цю напрямленість надають процесам обміну внутрішньоклітинні ферменти. Якщо б ферменти на мали специфічності, їх дія призводила б до швидкого розпаду клітинного матеріалу і при цьому не відбувалося б біосинтезу.
Ступінь специфічності у різних ферментів варіює. В більшості випадків фермент діє тільки на одну речовину і каталізує лише одну реакцію. В інших випадках фермент може діяти на ряд близькоспоріднених речовин, каталізуючи кожен раз одну й ту ж саму реакцію. Ферменти демонструють високий рівень стереоспецифічності (просторової специфічності), регіоселектівності (специфічності орієнтації) і хемоселектівності (специфічності до хімічних груп).
Модель «ключ-замок»
У 1890 році Еміль Фішер припустив, що специфічність ферментів визначається точною відповідністю форми ферменту і субстрата. Таке припущення називається моделлю «ключ-замок». Фермент з'єднується з субстратом з утворенням короткоживучого фермент-субстратного комплексу. Проте, хоча ця модель пояснює високу специфічність ферментів, вона не пояснює явища стабілізації перехідного стану, який спостерігається на практиці.
Кінетика ферментативної реакції
Крива насичення хімічної реакції (рівняння Міхаеліса-Ментен), що ілюструє співвідношення між концентрацією субстрата [S] і швидкістю реакції V.(Рис.2) Найпростішим і найпоширенішим описом кінетики односубстратних ферментатівних реакцій є рівняння Міхаеліса-Ментен. На сьогоднішній момент описано і кілька складніших типів кінетики ферментів. Наприклад, якщо реакція вимагає кількох молекул субстрата або різних субстратів, часто реакція протікає через утворення третинного комплексу. Для дії багатьох ферментів також типове утвореня перехідних комплексів (станів), що описується «механізмом пінг-понг».
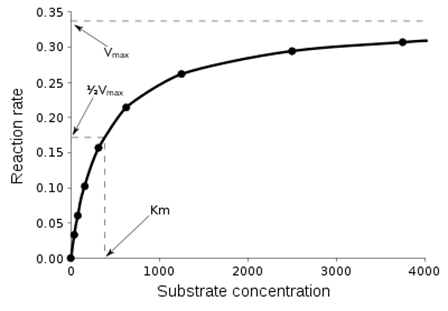
Рис.2. Крива насичення хімічної реакції
3 Функції ферментів
Ферменти є біологічними каталізаторами, вони присутні у всіх живих клітинах і сприяють перетворенню одних речовин (субстратів) на інші (продукти). Ферменти виступають в ролі каталізаторів практично у всіх біохімічних реакціях, що відбуваються в живих організмах — ними каталізується біля 4000 хімічно окремих біореакцій. Ферменти грають найважливішу роль у всіх процесах життєдіяльності, скеровуючи та регулюючи обмін речовин організму. Подібно до всіх каталізаторів, ферменти прискорюють як пряму, так і зворотну реакцію, знижуючи енергію активації процесу. Хімічна рівновага при цьому не зміщується ні в прямий, ні у зворотний бік. Відмінність ферментів від небілкових каталізаторів полягає у їхній високій специфічності — константа дисоціації деяких субстратів з білком-ферментом може досягати менш ніж 10−10 моль/л. Ферменти широко використовуються і в народному господарстві — харчовій, текстильній промисловості, у фармакології.
4 Класифікація ферментів
Комісія по ферментам у 1961 році розробила засновану на класифікації систему присвоєння кодових чисел (шифрів) індивідуальним ферментам. Шифр кожного ферменту складається з чотирьох чисел і створюється за наступним принципом.
- Перше число вказує до якого з шести головних класів належить даний фермент:
1. КФ 1: Оксидоредуктази — ферменти, що каталізують окислення або відновлення. Приклад: каталаза, алкогольдегідрогеназа
2. КФ 2: Трансферази — ферменти, що каталізують перенесення хімічних груп з однієї молекули субстрата на іншу. Серед трансфераз особливо виділяють кінази, що переносять фосфатну групу, як правило, з молекули АТФ.
3. КФ 3: Гідролази — ферменти, що каталізують гідроліз хімічних зв'язків. Приклад: естерази, пепсин, трипсин, амілаза, ліпопротеїнліпаза
4. КФ 4: Ліази — ферменти, що каталізують розрив хімічних зв'язків без гідролізу з утворенням подвійного зв'язку в одному з продуктів.
5. КФ 5: Ізомерази — ферменти, що каталізують структурні або геометричні зміни в молекулі субстрата.
6. КФ 6: Лігази — ферменти, що каталізують утворення хімічних зв'язків між субстратами за рахунок гідролізу АТФ. Приклад: ДНК-полімераза
- Друге число означає підклас. У оксидоредуктаз воно вказує природу тієї групи в молекулі донора, яка піддається окисленню; у трансфераз – природу групи, що транспортується; у гідролаз – тип зв’язків, що гідролізуються; у ліаз – тип зв’язків, що підлягають розриву; у ізомераз – тип реакції ізомеризації, що каталізується; у лігаз – тип утвореного зв’язку.
- Третє число означає підпідклас.
- Четверте число вказує порядковий номер ферменту в даному підпідкласі.
Зазвичай ферменти іменують за типом реакції, яку він каталізує, додаючи суфікс -аза до назви субстрату (наприклад лактаза — фермент, що бере участь в перетворенні лактози). Таким чином, у різних ферментів, що виконують одну функцію, буде однакова назва. Такі ферменти розрізняють по інших властивостях, наприклад, по оптимальному pH (лужна фосфатаза) або локалізації в клітині (мембранна АТФ-аза).
Оксидоредуктази
— клас ферментів, що каталізують реакції окислення, тобто перенесення електронів з однієї молекули (відновника, донора електронів) на іншу (окислювач, акцептор електронів).
Реакції, що каталізуються оксидоредуктазами, виглядають так:
A- + B → A + B-
Тут A
— відновник (донор електронів), а B
— окислювач (акцептор електронів)
У біохімічних перетвореннях окислительно-відновні реакції іноді виглядають складнішими. Наприклад, одна з реакцій гліколізу:
Pн + гліцеральдегид-3-фосфат + НАД+ → НАД · H + H+ + 1,3-діфосфогліцерат
Тут окислювачем виступає НАД+, а гліцеральдегид-3-фосфат є відновником.
Систематичні назви ферментів класу утворюються по схемі «донор:акцептор + оксидоредуктаза». Проте широко використовуються й інші схеми іменування. Коли можливо, ферменти називають у вигляді «донор + дегідрогеназа», наприклад гліцеральдегид-3-фосфатдегідрогеназа, для другої реакції вище. Іноді назва записується як «акцептор + редуктаза», наприклад НАД+-редуктаза. У окремому випадку, коли окислювачем є кисень, назва може бути у вигляді «донор + оксидаза». Згідно з міжнародною класифікацією ферментів, оксидоредуктази відносяться до 1 класу
ферментів, в межах якого виділяють 22 підкласи:
· КФ 1.1 — ферменти, що взаємодіють з CH—OH—групою донорів;
· КФ 1.2 — ферменти, що взаємодіють з альдегідною або оксо-групою донорів;
· КФ 1.3 — ферменти, що взаємодіють з CH—CH—групою донорів;
· КФ 1.4 — ферменти, що взаємодіють з CH—NH2 групою донорів;
· КФ 1.5 — ферменти, що взаємодіють з CH—NH групою донорів;
· КФ 1.6 — ферменти, що взаємодіють з НАД · H або НАДФ · H;
· КФ 1.7 — ферменти, що взаємодіють з іншими азотовмісними сполуками в якості донорів;
· КФ 1.8 — ферменти, що взаємодіють з сірковмісною групою донорів;
· КФ 1.9 — ферменти, що взаємодіють з гемовою групою донорів;
· КФ 1.10 — ферменти, що взаємодіють з дифенолами та спорідненими сполуками в якості донорів;
· КФ 1.11 — ферменти, що взаємодіють з пероксидом в якості акцептора (пероксидази);
· КФ 1.12 — ферменти, що взаємодіють з воднем в якості донорів;
· КФ 1.13 — ферменти, що взаємодіють з одиночними донорами із вбудуванням молекулярного кисню (оксигенази);
· КФ 1.14 — ферменти, що взаємодіють з парними донорами з вбудовуванням молекулярного кисню;
· КФ 1.15 — ферменти, що взаємодіють з супероксид-радікалами в якосі акцепторів;
· КФ 1.16 — ферменти, що окилюють іони металів;
· КФ 1.17 — ферменти, що взаємодіють з CH чи CH2 групами;
· КФ 1.18 — ферменти, що взаємодіють з залізосірчаними білками в якості донорів;
· КФ 1.19 — ферменти, що взаємодіють з відновленим флаводоксином в якості донора;
· КФ 1.20 — ферменти, що взаємодіють з фосфором чи миш'яком в якості донора;
· КФ 1.21 — ферменти, що взаємодіють з молекулами виду X—H и Y—H з утворенням зв'язку X—Y;
· КФ 1.97 — решта оксидоредуктаз.
Гідролази
(КФ 3) — клас ферментів, що каталізують гідроліз ковалентного зв'язку. Загальний вигляд реакції, що каталізується гідролазами, виглядає наступним чином:
A–B + H2O → A–OH + B–HСистематична назва гідролаз включає назву субстрату, що розщеплюється, з додаванням після неї слова -гідролаза. Однак, як правило, в тривіальній назві слово гідролаза скорочується до суфіксу «-аза».
Згідно з міжнародною класифікацією ферментів гідролази віднесені до класу (КФ 3). Клас, в свою чергу, підрозділяється на 13 підкласів в залежності від типу зв'язку, що гідролізується.
Лігази (від лат. ligāre — «зшивати», «зв'язувати») — клас ферментів (КФ 6), здатних каталізувати з'єднання двох молекул з утворенням нового хімічного зв'язку (лігування). При цьому зазвичай відбувається відщеплення (гідроліз) невеликої хімічної групи від однієї з молекул. Зазвичай реякція має вигляд:
Ab + C → A—C + b
де малі букви позначають невеликі хімічні групи, що відщеплюються лігазою.
Зазвичай назви лігаз включаються в себе слово «лігаза» (наприклад, ДНК-лігаза) або слово «синтетаза» (наприклад, аміноацил-тРНК-синтетаза). Через те, що деякі лігази додають вуглекислоту до молекули, вони мють назва карбоксилаз. Відмітьте, не слід плутати назви «синтетаза» і «синтаза», остання каталізує синтез молекул без відщеплення малої групи і згідно класифікації ферментів групується разом з ліазами.
В класифікації міжнародної комісії по ферментам, лігази класифікуються як КФ 6 та поділяються на 6 підгруп:
· КФ 6.1 включає лігази, що формують зв'язки вуглець-кисень
· КФ 6.2 включає лігази, що формують зв'язки вуглець-сірка
· КФ 6.3 включає лігази, що формують зв'язки вуглець-азот (включаючи аргінінсукцинат-синтетазу)
· КФ 6.4 включає лігази, що формують зв'язки вуглець-вуглець
· КФ 6.5 включає лігази, що формують фосфодиефірні зв'язки
· КФ 6.6 включає лігази, що формують зв'язки азот-метал
5 Локалізація ферментів у клітині
В кожній рослинній клітині синтезуються білкові молекули, які діють як каталізатори. Сотні різних ферментів, які діють в кожній клітині, прискорюють численні ферментативні реакції, послідовність яких становить так звані метаболічні шляхи. Ферментативні реакції поділяють на реакції синтезу (анаболічні) та реакції розпаду (катаболічні). Ферменти характеризуються специфічністю, з’єднуючись із субстратом, вони утворюють короткоживучий фермент – субстратний комплекс, який по закінченні реакції розпадається на продукти реакції та фермент. Фермент в реакції не змінюється. Концентрації ферменту і субстрату при певних умовах значно впливають на швидкість ферментативної реакції. Концентрація ферменту помітно позначається на швидкості ферментативної реакції тоді, коли спостерігається значний надлишок субстрату. Як правило, згадану залежність на графіку можна показати прямою лінією. Лише в разі порушення нормального перебігу реакції (під впливом інгібіторів або активаторів, при затримці пересування молекул субстрату до молекул ферменту) спостерігається відхилення від загальної закономірності.
Ферменти резистентного дихання
Мітохондрії рослин відрізняються від мітохондрій тварин тим, що в них можуть функціонувати два різних шляхи перенесення електронів від субстрату типу НАД•Н та сукцинату до О2. Якщо один з цих шляхів пригнічується з допомогою KCN-ціанідів, що блокують цитохромоксидазу, то інший шлях не блокується ціанідом, а тому його називають ціанід резистентним ланцюгом перенесення електронів. Аналогічно сам окислювальний процес, що призводить до утворення такого термінального ланцюга, має назву ціанідрезистентного дихання, або дихання, стійкого до ціаніду. Слід підкреслити, що ці два ланцюги не зовсім незалежні. Справа в тому, що в обох випадках введення в ланцюг електронних пар здійснюється внутрішньою та зовнішньою НАДН-дегідрогеназою та сукцинатдегідрогеназою і далі вони передають на убіхінон. Тільки потім, після убіхінону, спостерігається розділення їхніх дальших шляхів. В нормальному ланцюгу електрони поступово переносяться через ФП, цитохроми b- і с- типу та цитохромоксидазу до О2, тоді як в ціанід резистентному ланцюгу електрони переносяться від убіхінону на ФП, а потім через ціанідрезистентну термінальну оксидазу теж на О2. Природу термінальної оксидази поки що не вивчено. Вважають, що це залізо сірчаний білок, що не є гемопротеїном. Фізіологічне значення ціанідрезистентного дихання не зрозуміле. Вважають, що воно відповідає за клімактерій у плодів, тобто посилення дихання в процесі перед дозріванням плодів. Є дані, що клімактерій індукується етиленом, тому вважають, що етилен стимулятор ціанідрезистентного дихання. Доведено, що у деяких проростків цей тип дихання функціонує на ранніх етапах (бубнявіння насіння).
Допоміжні ферменти
До ферментних систем, які беруть участь у диханні, слід віднести також карбоксилази. Вони каталізують відщеплення СО2 від різних органічних кислот, в результаті чого утворюються сполуки з укороченим вуглецевим ланцюгом. Наприклад, таким чином при декарбоксилуванні пірувату утворюється оцтовий альдегід і т.д. Слід також згадати ще одну групу ферментних систем, які не беруть безпосередньої участі в окисленні субстратів та перенесенні електронів, але беруть активну участь в підготовці субстратів дихання. Нагадаємо, що здатність ферментів дегідрогеназ здійснювати окислення поширюється лише на сполуки з невеликою, досить простої структури молекулою (органічні кислоти, спирти, цукри). Однак при диханні використовуються досить складні органічні сполуки типу поліцукрів, білків, жирів .Саме тому використанню їх передує перетворення їхньої структури з допомогою ферментів гідролітичного комплексу. Потім в дію вступають ферменти, діяльність яких направлена на активування молекул мономерів, шляхом утворення їхніх фосфорних ефірів. Дальше активування досягається перетворенням хімічної структури молекули за допомогою ізомераз. Слід назвати також групу трансфераз, які каталізують відщеплення та перенесення окремих фрагментів вуглецевого ланцюга (трансальдолази та транскетолази).
Ізоферменти
Ізоферменти – це ферменти з однаковою каталітичною активністю, але з різною структурою. Зазвичай вони складаються з певних субодиниць. Різноманітне поєднання субодиниць дає ферменти, які мають одну й ту саму активність, але при цьому можуть розділятися на різні фракції за допомогою сучасних методів хімії білка. Основна роль ізоферментів – це регуляцію метаболічних процесів в клітині. Різні ізоферменти знаходяться в різних органелах рослинної клітини. Чим пояснити такий розподіл, до кінця ще не зрозуміло.
6 Створення нових ферментів
Команді учених із США і Ізраїлю вдалося створити ферменти, що прискорюють реакцію, для якої в природі не існує природного каталізатора. Щоб досягти максимальної ефективності нового ферменту, дослідники імітували in vitro процес його "еволюції". Робота опублікована в одному з найавторитетніших наукових журналів - Nature.
У своїй роботі учені задалися метою створити фермент, який каталізував би реакцію відщеплення протона, пов'язаного з вуглецем. У природі відповідного фермента не існує. На першій стадії експерименту дослідники визначили структуру активного центру майбутнього фермента. Потім вони визначили, якою повинна бути повна послідовність амінокислот, щоб вони змогли "згорнутися" і утворити активний центр заданої структури. За допомогою нової методики професор Девід Бейкер з Університету Вашигтона в Сієтлі проаналізував десятки тисяч можливих варіантів і виділив близько 60 "кандидатів". Учені синтезували їх і перевірили на наявність біологічної активності. Цей тест пройшли вісім "претендентів", їх яких дослідники відібрали три найбільш перспективні варіанти. Фахівці з відділу структурної біології Інституту Вайсмана визначили тривимірну структуру одного з трьох ферментів. Вона виявилася практично ідентична структурі, передбаченій за допомогою комп'ютерного аналізу. Проте ефективність цих ферментів була істотно нижча, ніж у їх природних "родичів", яких природа відбирала мільйони років.
Дослідники з Ізраїлю змоделювали еволюцію ферменту in vitro. Вони випадковим чином вносили в послідовності ферментів зміни, після чого аналізували активність одержаних мутантів. Після семи циклів внесення мутацій активність ферменту збільшилася в 200 разів в порівнянні з початковим варіантом, розробленим на основі комп'ютерної моделі. Швидкість реакції у присутності нового фермента зростала на шість порядків. Внесені мутації трохи змінювали структуру активного центру ферментів, що оптимізувало протікання безпосередньо реакції відщеплення, або змінювали гнучкість всього ланцюга, що сприяло прискоренню звільнення продукту реакції. Розроблений ученими алгоритм дозволяє створювати ферменти, що каталізують необхідні реакції, і збільшувати їх активність до рівня природних аналогів. Крім того, отримані результати допоможуть краще зрозуміти принципи роботи ферментів.
Висновки
Дослідження ферментів почалось у 19 столітті. Пізніше було введено термін «ензим» вченим В.Кюне. Ферменти за хімічною природою є білками. Ферменти є каталізаторами усіх хімічних реакцій в рослинній клітині. Ферменти широко використовуються і в народному господарстві — харчовій, текстильній промисловості, у фармакології. Одна з найбільш характерних властивостей ферментів – це їх специфічність, в міру якої кожен фермент діє тільки на одну речовину або дуже невелику кількість споріднених речовин. Специфічність дії ферментів – найважливіше біологічне явище, без якого неможливий впорядкований обмін у живій природі, відповідно і саме життя.
Сьогодні наука стрімко розвивається. Вчені виявляють нові ферменти, які досі були невідомі. Для того, щоб не розгубитися у великій кількості назв ферментів, потрібно знати принципи їх класифікації. У рефераті подано сучасну систему класифікації ферментів. Усі ферменти поділяють на шість головних класів.
Рослинна клітина багата на ферменти. Вони локалізовані в органелах рослинної клітини і каталізують життєво важливі процеси. Проте вчені пішли далі і створили штучні ферменти, які каталізують реакції, для яких в природі не існує каталізаторів.
Список літератури
1. Гудвин Т., Мерсер Э. Введение в биохимию растений, т.1. – М.: Мир, 1986. – С. 387
2. Диксон М., Уэбб Э. Ферменты, т.1. – М.: Мир, 1982. – С. 331-333, 292-294
3. Мосс Ф. Ферменты. – М.: Мир, 1970. – С. 10-13, 57-62.
4. Мусієнко М.М. Фізіологія рослин. – К.: Вища школа, 1995. – С. 33-34, 186-187
5. http://sciencereview.org
6. http://uk.wikipedia.org
|
