Реферат
На тему
Критические периоды развития зрительной и слуховой систем
Зрительная система новорожденных котят и обезьян
Последствия правильного и неправильного установления связей изучались в зрительной системе новорожденных котят и обезьян. Рецептивные поля нейронов сетчатки, латерального коленчатого тела и зрительной коры уже к рождению имели строение, аналогичное взрослым животным, кроме слоя 4 зрительной коры. К моменту рождения кортикальные клетки в слое 4 управляются обоими глазами, и в течение первых 6 недель жизни они формируют связи подобные таковым у взрослых, когда каждая клетка слоя 4отвечает только на сигналы от одного глаза, в то время как клетки, локализующиеся в других слоях, продолжают получать бинокулярную информацию. Если в течение первых трех месяцев жизни один из глаз остается закрытым, это приводит к слепоте на этот глаз и потере его способности управлять кортикальными клетками. Кортикальные колонки, получающие сигналы от депривированного глаза, уменьшаются в размерах, в то время как колонки, получающие сигналы от противоположного, недепривированного, — увеличиваются.
Закрытие глаза у взрослого животного не влияет на колоночную архитектуру зрительной коры или на ответы корковых клеток. Во время критических периодов можно добиться обратного развития изменений, вызванных сенсорной депривацией, если открыть ранее закрытый глаз и закрыть недепривированный. Дополнительным доказательством наличия конкуренции между двумя глазами служит эксперимент, в котором оба глаза остаются закрытыми в раннем возрасте. Когда ни один из глаз не имеет преимуществ, продолжает развиваться нормальная колоночная архитектура, однако каждая клетка в коре при этом управляется только одним глазом. Подобным же образом при подрезании мышц глазного яблока у детенышей обезьян (что приводило к косоглазию (страбизму)), только небольшое количество нейронов получало сигналы от обоих глаз, хотя каждый глаз получал нормальное количество света. Роль функциональной активности в доминировании того или иного глаза также может быть показана в экспериментах, в которых блокада импульсов в обоих зрительных нервах при помощи тетродотоксина предотвращала разделение коры на колонки глазного доминирования. Спонтанная импульсная активность и нейротрофины (neurotrophins) могут также влиять на правильное формирование связей коры.
Реклама

У незрелых совят развитие слуховой системы также значительно выражено в критические периоды. Призмы, помещенные перед глазами, приводят к смещенному расположению рецептивных полей. Таким образом, возникает диспропорция между картами окружающего пространства в четверохолмии (tectum), соответствующими зрительным (верхние холмики) и слуховым (нижние холмики) ощущениям. В течение первых месяцев жизни это несоответствие корректируется в результате изменения карты окружающего пространства в слуховой коре. По прошествии критического периода подобного рода сдвиги становятся уже невозможны. У сов, выращиваемых в специальных условиях, когда они получают большое количество сенсорного опыта, критический период, в течение которого возможны изменения в связях коры, удлиняется. Эксперименты с сенсорной депривацией являются важными для понимания развития высших функций мозга.
Мы несколько раз подчеркивали, что для правильной работы нервной системы требуется высокая специфичность ее связей. Также понятно, что развитие связей между нейронами продолжается и после рождении на протяжении различного периода времени у разных животных. Например, котята рождаются слепыми на оба глаза. Если мы поднимем веко, чтобы свет попал в глаз, это приведет к сужению зрачка, хотя раньше этот котенок никогда не видел света и является полностью слепым. К 10-му дню жизни котенка появляются первые признаки зрения, и, таким образом, он начинает распознавать объекты и паттерны. Когда котята вместо нормальных условий выращиваются в темноте, у них сохраняется зрачковый рефлекс, однако они остаются слепыми. Как полагают, имеется иерархия уровней восприятия, с «постоянными» и «переменными» связями, образующимися в разных частях мозга.
Изменения в функционировании зрительной системы во время развития порождают ряд вопросов. Каковы удельные доли генетических факторов и приобретаемого опыта в данном процессе? До какой степени те нейронные сети, которые необходимы для зрения, уже сформировались и готовы к работе к моменту рождения? Какое значение в их развитии имеет свет, падающий на сетчатку? В чем причина того, что котенок, выращенный в темноте, слепой: связано ли это с тем, что не сформировались какие-то связи в коре, или произошло разрушение каких-то ранее сформированных связей? Все эти концепции можно охарактеризовать одной фразой: «природа и воспитание». С накоплением знаний это выражение трансформировалось в «природа или воспитание» или даже более строго: «природа против воспитания». В этой главе будет показано, что оба эти фактора (как и следовало ожидать) являются необходимыми.
Реклама

Зрительная система является удобным объектом для прямого изучения вопросов, связанных с развитием, потому что пункты переключения сигнала в коре (relay stations) легко достижимы и количество света, а также естественные стимулы можно легко варьировать. В пределах зрительной системы мы опять отдельно выделяем эксперименты, выполненные на обезьянах и кошках. Для достижения цели более удобно начать эту главу с подробного рассмотрения работ, результаты которых логически следуют из материалов. Работы, выполненные на незрелой зрительной системе, дают основу для анализа других сенсорных систем, в частности слуховой системы амбарной совы (barn owl).
Они также дают возможность предполагать, какие механизмы играют роль в модификации корковых связей в начальный период жизни.
Имеется много данных об организации корковых связей, которые лежат в основе восприятия зрительной информации у взрослых кошек и обезьян. Простая клетка в коре избирательно «узнает» один строго определенный тип зрительных стимулов, такой как движение узкой вертикально ориентированной полоски света, расположенной в определенном месте зрительного поля обоих глаз. Подобного рода ответы возможны благодаря точным и определенным образом организованным связям между клетками коры, сетчатки и латерального коленчатого тела. Естественным является любопытство: присутствуют ли эти клетки и связи уже в мозгу новорожденного животного, или они развиваются как результат приобретенного зрительного опыта, когда зрительные образы в раннем периоде жизни каким-то образом переориентируют случайный набор существующих связей, модифицируя его под определенную задачу.
Для изучения обезьян, не имеющих зрительного опыта, новорожденных обезьян отбирали сразу после рождения (или кесарева сечения), после чего за ними ухаживали таким образом, чтобы исключить зрительные стимулы. Для предотвращения формирования зрения до того периода, когда обезьяны станут достаточно взрослыми для изучения, проводили сшивание век или на роговицу накладывали полупрозрачный материал, который пропускал свет, однако изображение воспринималось очень размыто. Подобным же образом в течение первых недель жизни исследовались не имеющие зрительного опыта котята и хорьки.
Рецептивные поля и свойства кортикальных клеток новорожденных животных
Новорожденные обезьяны сразу проявляют выраженную зрительную активность и способны к фиксации взгляда. Ответы корковых нейронов во многом сходны с ответами у взрослых животных. Например, запись сигнала от отдельных клеток первичной зрительной коры (V1
) показывает, что клетки не отвечают на диффузное освещение. Как и у взрослых животных, они лучше всего отвечают на полоски света или тени определенной ориентации, проецируемые на определенные области сетчатки2)
. В опытах с животными, не имевшими предыдущего зрительного опыта, диапазон ориентации ничем не отличается от такового у взрослых животных. Рецептивные поля также организованы в антагонистические "on"- и "off"-зоны, управляемые обоими глазами. Более того, при проникновении в толщу коры в косом направлении при движении электрода наблюдается смена предпочтительных ориентации в строго определенной последовательности(рис. 1). На рис. 1 также показаны карты предпочтительных ориентации с характерными «вертушками». Таким образом, очевидно, что все ориентации представлены в равной степени уже к моменту рождения.
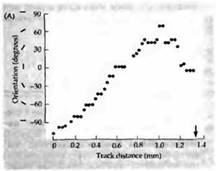 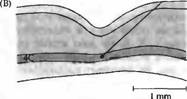
Рис. 1. Колонки ориентации в отсутствие зрительного опыта. (А) Ось ориентации рецептивного поля клеток при прохождении злектрода под косым углом к коре 17-дневного детеныша обезьяны, глаза которого были закрыты при помощи шва на второй день после рождения. Ориентация зрительных полей изменяется постепенно по мере пересечения колонок, что говорит о том, что нормальные колонки ориентации присутствуют в коре животного, не имеющего зрительного опыта. Серые точки — ипсилатеральный глаз; черные точки — контралатеральный глаз. (В) Черной точкой отмечено повреждение в конце проникновения электрода в слой 4. (С) Колонки ориентации, выявленные при помощи оптических методов у 14-дневного котенка, глаза которого были закрыты швом с рождения. Цветные полоски (справа) показывают ориентацию стимула. Стоит отметить, что уже имеются «вертушки».
Глазодоминантные колонки у новорожденных обезьян и котят
При рождении большинство клеток во всех слоях первичной зрительной коры уже получают управление от обоих глаз — некоторые в основном от правого, другие — от левого, и некоторые — в одинаковой степени от обоих. На рис. 2 показано распределение ответов нейронов всех слоев коры по отношению к глазному предпочтению у взрослых и ювенильных обезьян.
Степень доминирования может быть более наглядно выражена в виде гистограммы, где нейроны сгруппированы в семь категорий согласно частоте разрядов, с которой они отвечают на стимуляцию правого или левого глаза. Клетки в группе 1 и 7 на рис. 2 управляются только зрительными стимулами, предъявляемыми на один из глаз, в то время как группы со 2 по 6 отвечают в разной степени на сигналы от обоих глаз.
Гистограммы на рис. 2А и В выглядят похоже в пределах диапазона глазного предпочтения. Однако они не отражают важной и поразительной разницы между новорожденными и взрослыми обезьянами, что касается свойств клеток слоя 4. В пределах этого слоя у новорожденных обезьян клетки управляются от обоих глаз, а после 6 недель развития — только от одного. Вне слоя 4 кортикальные клетки новорожденных обезьян демонстрируют ответы, аналогичные клеткам взрослых, за исключением того, что сигналы от некоторых клеток могут быть менее интенсивными или временно отсутствовать.
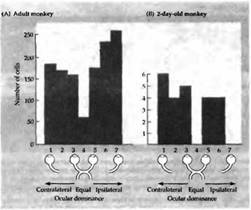
Рис. .2. Распределение глазного доминирования в зрительной коре новорожденной обезьяны. Клетки в группах 1 и 7 гистограммы управляются только одним глазом (ипсилатеральным или контралатераль ным). Все другие клетки получают сигналы от обоих глаз. В группах 2, 3, 5 и 6 один из глаз доминирует. В группе 4 оба глаза оказывают одинаковое влияние. (А) Нормальная взрослая обезьяна, (В) Нормальная 2-дневная обезьяна.
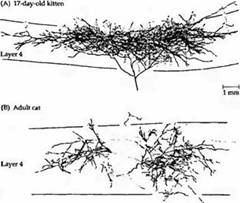
Рис. 3. Возрастная зависимость паттернов ветвления аксонов нейронов ЛКТ, заканчивающихся в слое 4, выявленных при введении пероксидазы хрена. (А) Аксон 17-дневного котенка. Аксон разветвляется на большой непрерывной территории слоя 4 зрительной коры. (В) У взрослой кошки аксон нейрона ЛКТ заканчивается на двух различных лучках, между которыми находятся неокрасившиеся волокна, идущие от другого глаза.
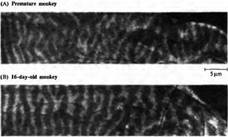
Рис.4. Глазодоминантные колонки в слое 4 стриарной коры зрительно «наивной» обезьяны и более зрелого животного, которое имело зрительный опыт. (А) Обезьяна была рождена на 8 дней раньше срока, незрелой, при помощи кесарева сечения. Для проведения сечения использовались инфракрасные очки и операция проходила в полной темноте. На второй день жизни в правый глаз был введен радиоактивный пролин, после чего животное держали в темноте еще 7 дней. Авторадиография показывает слой 4 ипсилатеральной первичной зрительной коры. Уже различимы колонки глазного доминирования. (В) Подобное сечение через слой 4 16-дневного детеныша обезьяны, рожденного в срок и содержащегося в нормальных световых условиях. За 9 дней до опыта в правый глаз ввели радиоактивный пролин. Границы колонок глазного доминирования хорошо различимы.
Формирование глазодоминантных колонок
В слое 4 Ле Вей, Визель и Хьюбелъ (а также Ракич) нашли важное и поразительное анатомическое различие между взрослыми и новорожденными. Причиной того, что клетки в пределах слоя 4 у новорожденных управляются обоими глазами заключается в том, что ветвления нервных окончаний волокон из ЛКТ значительно перекрывают друг друга (рис. 3 и 4). Отдельные аксоны, достигающие слоя 4 из ЛКТ, у новорожденных простираются на более широкий участок, чем у взрослых. В результате зоны, получающие информацию от каждого из глаз, перекрываются. Это отличается от паттерна строения этого слоя у взрослых, когда глазодоминантные колонки, отвечающие на сигналы от правого или левого глаза, имеют четко определенные границы. На рис. 4 показано, что глазодоминантные колонки могут быть обнаружены у обезьяны еще при рождении, хотя их развитие к этому моменту еще не закончено. Границы колонок у обезьян, не имеющих зрительного опыта, выглядят более смазанными и менее четко определены.
В течение первых 6 месяцев жизни животного аксоны из ЛКТ, направляющиеся в слой 4, подвергаются ретракции, что приводит к уменьшению ветвистости, напоминая «подрезанное» дерево (рис. 3). Таким образом, в пределах слоя 4 устанавливаются отдельные корковые области, каждая из которых управляется только одним глазом (рис. 5). Сравнимые изменения во время развития происходят и на предыдущем этапе зрительного пути, в латеральном коленчатом теле. Волокна зрительного нерва от обоих глаз врастают в ЛКТ, области их ветвления значительно перекрываются, до тех пор пока они не разделятся на отдельные слои. Постнатальное развитие колонок глазного доминирования и слоев ЛКТ происходит и у животных, выращенных в полной темноте. Эти морфологические наблюдения могут объяснить изменения в глазном доминировании. «Пятна» в области первичной зрительной коры, выявленные при окрашивании на цитохромоксидазу, определяются анатомически уже к моменту рождения, что связано с нормальным развитием глазодоминантных колонок. Полоски в зрительной области 2 (зона 18) четко различимы у новорожденных обезьян, не имеющих зрительного опыта.
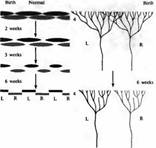
Рис. 5. Ретракция окончаний аксонов ЛКТ в пределах слоя 4 коры в течение первых 6 недель жизни. На рисунке показано перекрытие сигналов, исходящих из правого (R) и левого (L) глаза, имеющееся с рождения, и последующее разделение в различные кластеры, связанные с соответствующими колонками глазного доминирования. Перекрытие при рождении больше у котят, чем у детенышей обезьян.
Литература
1. Sanes, J. Marshall, L. M McMahan, U. J. 1978. Reinnervation of muscle fiber basal lamina. J. Cell .
78: 176-198.
2. Saunders, N. R Kitchener, P., Knott, G. Nicholls, J. Potter, A. and Smith, T.J. 1998. Development of walking, swimming and neuronal connections after complete spinal cord in the neonatal opossum Monoelphis J. Neurosci.
18: 339-355.
3. Schwab, M. E and Cani, P. 1988. Olgodends and CNS melin are nonpessive substrates for neute growth andbroblast spreading in vitro. J. Neurosci.
8: 238-2393.
|
