Реферат
"Ионный канал. Проводимость и проницаемость"
Проводимость
и
проницаемость
ионного канала
Проводимость ионного канала зависит от двух факторов: во-первых, от той легкости, с которой ионы проходят через открытый канал. Это внутреннее свойство канала известно как проницаемость канала. Во-вторых, проводимость зависит от концентрации ионов около устьев канала. Ясно, что если ионы калия отсутствуют как внутри, так и снаружи клетки, то не может быть и тока. При этом становится не важно, какова проницаемость канала или насколько велик мембранный потенциал. Если присутствует всего несколько ионов калия, то, при одной и той же величине проницаемости и стандартном трансмембранном потенциале, ток через ионный канал будет гораздо меньше, чем при избыточной концентрации ионов калия. Эти взаимоотношения могут быть представлены следующим образом:
Открытый канал –> проницаемость.
Проницаемость + ионы –> проводимость.
В уже рассмотренных примерах ионного тока концентрации ионов калия были одинаковыми по обе стороны мембраны. Что произойдет, если мы сделаем концентрации этого иона в пипетке и в ванночке различными? Представим, что после получения конфигурации outside-out, концентрация калия в ванночке составляет 3 ммоль (подобно нормальной внеклеточной концентрации этого иона), а внутри электрода эта концентрация равняется 90 ммоль (подобно цитоплазматической концентрации). Если калиевый канал в мембране будет открыт, то ионы калия начнут двигаться из пипетки в ванночку. Такой ток будет происходить даже при отсутствии потенциала на пипетке, так как движущей силой для ионов калия является градиент концентраций. Если же мы зарядим пипетку положительно, градиент потенциала на мембране будет еще более увеличивать движение положительно заряженных ионов калия наружу (к минусу). В результате ток ионов через калиевые каналы будет возрастать. И наоборот, если придать пипетке отрицательный заряд, движение ионов калия из электрода наружу через мембрану замедлится и канальный ток уменьшится. Важно отметить, что при достаточно большом отрицательном заряде ионы калия начнут движение внутрь,
то есть против градиента концентрации. Ток ионов калия через канал зависит от электрического потенциала мембраны и градиента концентрации калия. Сочетание этих двух факторов формирует электрохимический градиент для калия. В отличие от начального результата, полученного при одинаковых концентрациях ионов калия по обе стороны мембраны, в данном примере ионный ток равен нулю при потенциале на пипетке -85 мВ. При этом потенциале стремление ионов калия выйти через канал наружу по градиенту концентраций полностью уравновешено трансмембранной разницей электрического потенциала, которая направляет движение ионов в противоположном направлении. Этот трансмембранный потенциал называется калиевым равновесным потенциалом (Е
K
)
Равновесный потенциал зависит только от концентрации ионов по обе стороны мембраны, но не от свойств ионного канала или механизма проникновения ионов через канал.
Реклама

Какой именно потенциал необходим для того, чтобы уравновесить эффект реальной разницы концентраций ионов калия? Предположение о том, что Е
к
просто пропорционален разнице между внутриклеточной [К]i
и внешней [К]0
концентрацией ионов калия, не совсем верно. Точнее, равновесный потенциал зависит от разницы между логарифмами
концентраций:
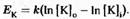
Константа k
определяется из формулы RT
/(
zF
),
где R
–
газовая постоянная, Т
– абсолютная температура, z – валентность иона (в данном случае +1) и F
– число Фарадея (число кулонов электричества в 1 молярном растворе иона). Таким образом, 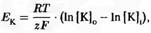 , что можно представить как , что можно представить как 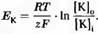 . .
Это уравнение называется уравнением Нернста для ионов калия. Аналогично, можно построить уравнение Нернста и для других основных ионов. Отношение RT/(zF)
измеряется в вольтах и равно примерно 25 мВ при температуре 20° С. Иногда более удобно пользоваться десятичным (log), а не натуральным логарифмом (ln). Тогда значениемRT
/(
zF
)
должно быть умножено на In 10, или 2,31, что даст в результате 58 мВ. То есть
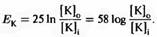 . .
При температуре тела млекопитающих (37°С) вместо 58 мВ следует использовать 61 мВ. Для клетки, показанной на рис. 2.7, значение £к
= -85 мВ соответствует отношению концентраций 1/30.
Необходимо отметить, что диффузия иона по градиенту концентрации не строго зависит от его концентрации. Во всех растворах, кроме очень слабых, ионы взаимодействуют друг с другом, что проявляется в электростатическом притяжении или отталкивании заряженных частиц. Результатом таких взаимодействий является снижение эффективной концентрации ионов. Эффективная концентрация иона называется активностью. Поэтому более точным теоретическим параметром для уравнения Нернста является соотношение активностей, а не соотношение концентраций. Однако поскольку суммарные концентрации ионов внутри и вне клетки близки, соотношение активности ионов не будет существенно отличаться от соотношения концентраций.
Реклама

Без приложения потенциала ток составляет около 4 рА, тогда как с потенциалом на пипетке -85 мВ ионный ток равен нулю. Следовательно, ионный ток определяется не абсолютным значением мембранного потенциала (V
m
),
a разницей между мембранным потенциалом и равновесным потенциалом для данного иона, в данном случае для иона калия (Ек
).
Эта разница Vm
– Ек
является движущей силой для прохождения ионов через канал. Вновь обратимся к рис. 2.7F: при мембранном потенциале, равном нулю, движущая сила составляет +85 мВ.
Вторым характерным свойством зависимости «ток–напряжение», является ее нелинейность. При сдвиге от равновесного потенциала -85 м В в сторону деполяризации (то есть к нулю) ток меняется более быстро, чем при переходе в сторону гиперполяризации. Это происходит из-за того, что проводимость канала является функцией концентрации иона. В нашем примере концентрация ионов калия внутри пипетки гораздо выше концентрации этого иона во внешнем растворе. Это приводит к тому, что током, идущим наружу, переносится больше ионов, чем током, направленным внутрь. По мере сдвига от равновесного потенциала в сторону деполяризации, этот эффект становится все более заметным. Поэтому зависимость «ток–напряжение» имеет направленный вверх изгиб, несмотря на то, что проницаемость этого типа канала практически не зависит от потенциала.
Нелинейные отношения «ток–напряжение» наблюдаются также в ионных каналах, обладающих выпрямляющими свойствами. В таких каналах проницаемость зависит от потенциала, поэтому при определенном потенциале ионы двигаются в одном направлении гораздо легче, чем в обратном. Одним из таких примеров является потенциалзависимый калиевый канал, называемый каналом внутреннего выпрямления (inwardrectifier). Такой тип канала позволяет ионам калия двигаться внутрь клетки при потенциале, более негативном, чем равновесный калиевый потенциал. Однако при потенциалах, менее негативных по отношению к равновесному калиевому потенциалу, выходящий ток или очень мал, или полностью отсутствует.
Каким же образом ионы проходят через каналы? Одним из возможных механизмов является диффузия через водную среду, заполняющую пору. Представление о диффузии лежало в основе ранних гипотез о процессе ионной проницаемости. Однако для большинства каналов простая диффузия описывает ионную проницаемость недостаточно адекватно. Главная причина в том, что проникающие ионы вступают во взаимодействие с белками ионного канала. Так, в растворе, благодаря наличию заряда, ионы всегда покрыты гидратной оболочкой. В случае катионов молекулы воды ориентированы таким образом, что отрицательно заряженный кислоро обращен в сторону иона. Если пора ионного канала достаточно узкая, необходимо некоторое количество энергии, чтобы освободить ион от ассоциированных молекул воды и позволить ему проникнуть через этот участок. В канале ион может быть объектом притяжения или отталкивания электростатическими зарядами стенки канала. Взаимодействие иона с комплементарными центрами ионного канала может приводить к тому, что процесс проникновения будет представлять собой своеобразный «перескок» иона с одного центра связывания на другой. Такие взаимодействия иона со стенкой канала могут влиять как на ионную избирательность, так и на скорость потока ионов. Модели, описывающие ионную проницаемость по этому механизму, называются моделями Эйринга. В целом, такие модели описывают ионную избирательность и проводимость гораздо более адекватно, чем модели простой диффузии.
Функционирование ионных каналов дает возможность нейронам реагировать на сигналы из внешней среды или от других нейронов, передавать импульсы на большие расстояния к исполнительным органам или к другим нейронам. Таким образом, вся сложная система восприятия и анализа сигналов, так же как генерация двигательной команды, определяется, в конечном счете, активностью ионных каналов.
Важно понимать, что все ионные токи, лежащие в основе нейрональной сигнализации, обусловлены пассивным движением ионов через открытые ионные каналы по градиенту концентрации и в зависимости от заряда клеточной мембраны. Другими словами, нейроны используют электрохимические градиенты для генерации потока ионов и, как следствие, для формирования электрических сигналов. Потенциально ионные токи могли бы нарушать эти градиенты, однако в действительности этого не происходит, так как клетки используют энергию, образуемую в ходе метаболизма, для поддержания ионного состава цитоплазмы. Специализированные механизмы, лежащие в основе активного транспорта ионов.
Каналы совершают переходы между открытым и закрытым состояниями. Каждый канал имеет присущее ему время открытого состояния. Когда каналы активированы, вероятность их открытия возрастает. Деактивация снижает частоту открытия. Каналы также могут быть инактивированы или блокированы. Каналы могут быть классифицированы по типу их активации: механочувствительные, потенциал-активируемые, лиганд-активируемые. Ионы движутся через каналы пассивно в соответствии с градиентом концентрации или электрическим градиентом на мембране.
Результирующий поток ионов через канал по градиенту концентрации может быть снижен противоположно направленным электрическим градиентом. Электрический потенциал, снижающий результирующий поток какого-либо иона до нуля, называется равновесным потенциалом данного иона. Отношение между равновесным потенциалом и градиентом концентрации описывается уравнением Нернста. Движущая сила для движения ионов через мембрану есть разница между равновесным и мембранным потенциалами. Ионный ток, протекающий через одиночный канал, зависит от движущей силы для данного иона и проводимости канала для этого иона. В свою очередь, проводимость зависит от проницаемости данного канала и внешней и внутренней концентраций ионов.
Электрические сигналы в нервной системе генерируются движением ионов через мембрану нервной клетки. Эти ионные токи протекают через водные поры трансмембранных белков, известных как ионные каналы.
Каналы различаются по своей избирательности: некоторые катионные каналы пропускают только натрий, калий или кальций, другие являются менее избирательными. Анионные каналы сравнительно не избирательны для малых анионов, но они пропускают в основном ионы хлора, так как хлор является самым распространенным анионом внеклеточной и внутриклеточной жидкостей.
Литература
Терлецкий Н.А. О пользе и вреде излучения для жизни. (Relata Refero.)
Эбелинг В., Энгель А., Файстель Р. Физика процессов эволюции.
Поппер К.Р. Объективное знание. Эволюционный подход.
Малинецкий Г.Г. Хаос. Структуры. Вычислительный эксперимент.
|
