Ионные механизмы потенциала покоя
Содержание
Введение
Идеальная клетка
Ионное равновесие
Электрическая нейтральность
Влияние внеклеточного калия и хлора на мембранный потенциал
Литература
В состоянии покоя на мембране нейронов поддерживается постоянный электрический потенциал, при этом внутриклеточная среда заряжена отрицательно по отношению к внеклеточной. Поскольку внутри клетки больше ионов калия и меньше ионов натрия и хлора, чем снаружи, то каждый из этих ионов стремится проникнуть сквозь мембрану в направлении своего электрохимического градиента, то есть ионы калия стремятся наружу, а ионы натрия и хлора - внутрь клетки. Этому перемещению препятствует мембранный потенциал. Если представить себе упрощенную модель клетки, мембрана которой проницаема лишь для калия и хлора, то в ней возможен такой баланс между концентрационными градиентами и мембранным потенциалом, при котором суммарный ток каждого из этих ионов равен нулю. Тогда мембранный потенциал будет в точности равен равновесному потенциалу для этих двух ионов.
Если в такой модели изменить внеклеточную концентрацию калия, то изменится равновесный потенциал для калия, а следовательно и мембранный потенциал. Изменение внеклеточной концентрации хлора, напротив, приведет к соответствующему изменению его внутриклеточного уровня. При этом ни равновесный потенциал для хлора, ни мембранный потенциал не изменятся.
Мембрана реальной клетки проницаема также для натрия. В состоянии покоя ионы натрия непрерывно перемещаются внутрь клетки, снижая таким образом отрицательный заряд на мембране. Поскольку при этом калий уже не находится в равновесии, он начинает вытекать из клетки. Если бы не механизмы компенсации, то такое перемещение натрия и калия привело бы к изменению их внутриклеточных концентраций. Этого не происходит благодаря работе натрий-калиевого насоса-обменника, переносящего ионы натрия наружу и ионы калия внутрь клетки в пропорции 3:2. Мембранный потенциал покоя зависит от калиевого и натриевого равновесного потенциала, относительной проницаемости клеточной мембраны для этих ионов, а также от стехиометрии натрий-калиевого насоса. В состоянии покоя скорость переноса натрия и калия насосом в точности компенсирует пассивную утечку этих ионов. Натрий-калиевый обменник вносит также непосредственный вклад в мембранный потенциал (в размере нескольких милливольт), поскольку он перемещает больше положительных зарядов наружу, чем внутрь клетки.
Реклама

В зависимости от механизмов транспорта, равновесный потенциал для хлора может быть как более, так и менее отрицательным по отношению к потенциалу покоя. Наличие значительной хлорной проницаемости играет важную роль в обеспечении электрической стабильности многих клеток, несмотря на то, что распределение ионов хлора не столь важно для формирования потенциала покоя.
Возникновение электрических сигналов в нервной клетке происходит в основном за счет изменения проницаемости мембраны для таких ионов, как натрий и калий. Увеличение проницаемости мембраны позволяет ионам перемещаться внутрь или наружу клетки в направлении своих электрохимических градиентов. Изменения проницаемости зависят от активации ионных каналов. Перемещение ионов через каналы в мембране приводит к изменению заряда на ней и, следовательно, мембранного потенциала. Чтобы понять принцип возникновения электрического сигнала, необходимо уяснить сущность ионных градиентов на мембране, а также то, как они влияют на потенциал покоя.
Начнем с рассмотрения модели идеальной клетки, показанной на рис.1.1. В этой клетке содержатся ионы калия, натрия и хлора, а также крупные анионы. Клетка помещена в раствор, содержащий ионы натрия и калия. Другие ионы, присутствующие в реальных клетках, не играют важной роли в формировании потенциала покоя и потому не рассматриваются в данной модели. Концентрации ионов внутри и снаружи клетки приблизительно соответствуют средним величинам, полученным на клетках лягушки. В клетках птиц и млекопитающих эти значения несколько выше, а у таких морских беспозвоночных, как кальмар - значительно выше, чем у лягушки (см. табл. 1.1). В настоящей модели рассматривается клеточная мембрана, проницаемая для калия и хлора, но не для натрия и внутриклеточных анионов. Для того чтобы клетка пребывала в стабильном состоянии, необходимо выполнение трех условий:

Рис.1.1 Распределение ионов в идеальной клетке.
Мембрана проницаема для К и Cl, но непроницаема для Na и внутриклеточных анионов (А). Концентрационный градиент ионов калия способствует выходу этих ионов из клетки (черная стрелка); градиент потенциала стремится перенести ионы калия внутрь клетки (серая стрелка). В состоянии покоя эти две силы уравновешивают друг друга. Концентрационный и электрический градиенты для хлора имеют противоположные направления. Ионные концентрации выражены в миллимолях (ммоль).
Реклама

1. Как внутриклеточная, так и внеклеточная среды должны быть электрически нейтральными. Так, например, раствор не может состоять из одних лишь ионов хлора. Отрицательный заряд этих ионов должен компенсироваться такими положительными ионами, как натрий или калий (иначе взаимное отталкивание одинаково заряженных частиц приведет к разрушению раствора).
2. Клетка должна находиться в осмотическом равновесии. В противном случае вода будет проникать в клетку (или вытекать из нее) до тех пор, пока равновесие не будет достигнуто. Осмотическое равновесие наступает при условии равенства суммарной концентрации растворенных частиц внутри и вне клетки.
3. Суммарный заряд каждого отдельно взятого иона, переносимый через мембраны клетки, должен равняться нулю.
Как создаются и поддерживаются ионные градиенты и соответствующий электрический потенциал? На рис.1.1 показано, что ионы находятся в положении обратной пропорциональности: ионы калия более концентрированы внутри клетки, а ионы хлора снаружи. Представим себе, что мембрана клетки проницаема только для ионов калия. Возникает вопрос, почему эти ионы не диффундируют из клетки наружу до тех пор, пока концентрации внутри и снаружи клетки не сравняются. Причина этого заключается в том, что если ионы калия покидают клетку, то снаружи накапливается положительный заряд
, а внутри клетки образуется избыток заряда отрицательного. Возникший таким образом электрический потенциал снижает скорость перемещения ионов калия, а при достижении определенного уровня приводит к полному его прекращению. Это равновесный потенциал для калия (Ек
). При потенциале Ек
электрический градиент полностью уравновешивает градиент химический, в результате чего движение ионов прекращается. Отдельные ионы калия по прежнему втекают в клетку и вытекают из нее, но суммарный ток равен нулю. Ионы калия находятся в равновесии.
Условия нахождения ионов калия в равновесии те же самые, что описаны и при рассмотрении нулевого суммарного тока через одиночный канал в небольшом участке (patch) мембраны.
В этом случае концентрационный градиент уравновешивался потенциалом, приложенным к patch-электроду.
Важное отличие описываемой здесь ситуации в том, что перемещение ионов само по себе производит электрический потенциал, уравновешивающий и останавливающий это перемещение.
Другими словами, равновесие в данной модели достигается автоматически и является неизбежным. Напомним, что в главе 2 равновесный потенциал для калия получался из уравнения Нернста:
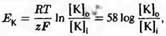
где [К] 1
и [К] 0
- внутриклеточная и внеклеточная концентрации калия, соответственно. Для клетки, изображенной на рис.1.1, получим ЕK
= - 85 мВ.
Допустим теперь, что в мембране, кроме калиевых, присутствуют еще и хлорные каналы. Поскольку для анионов z = - 1, получим равновесный потенциал для хлора:

или, пользуясь свойствами логарифма,

Для модели идеальной клетки получим соотношение концентраций хлора, которое тоже равно 1: 30, и хлорный равновесный потенциал, равный - 85 мВ. Как и в случае с калием, мембранный потенциал величиной в - 85 мВ в точности уравновешивает стремление ионов хлора двигаться в направлении их концентрационного градиента, т.е. внутрь клетки.
Обобщая вышесказанное, можно заключить, что мембранный потенциал препятствует перемещению как хлора внутрь клетки, так и калия из клетки наружу. Равновесные потенциалы для двух ионов равны благодаря тому, что соотношения их внеклеточных и внутриклеточных концентраций одинаковы (1: 30). Поскольку калий и хлор единственные ионы в нашей модели, способные проникать через мембрану клетки, и при - 85 мВ оба иона находятся в равновесии, то клетка может находиться сколь угодно долго в состоянии покоя, при котором суммарное перемещение ионов в клетку и из клетки будет равно нулю.
Перемещение ионов калия наружу и ионов хлора внутрь клетки приводит к накоплению отрицательного заряда в клетке и положительного - во внеклеточном пространстве. На первый взгляд, такая ситуация противоречит принципу электрической нейтральности, однако это не так. Ионы калия, покидая клетку, накапливаются в непосредственной близости от ее мембраны, в то время как их отрицательно заряженные спутники остаются внутри клетки вблизи от внутренней поверхности той же мембраны. И те и другие фактически оказываются вне основной части раствора, как вне-, так и внутриклеточного. Также и ионы хлора, входя в цитоплазму, остаются вблизи от мембраны, а их спутники, оставленные перед проходом через мембрану, скапливаются поблизости от нее. Образуются два слоя ионов - катионов снаружи и анионов внутри клетки, которые удерживаются у мембраны благодаря взаимному притяжению. Таким образом, мембрана играет роль электрической емкости, разделяющей и запасающей заряд.
Вышесказанное не означает, что ионы калия и хлора прикованы к поверхностям мембраны. Отдельные ионы свободно обмениваются с ионами внутри - или внеклеточного раствора. Тем не менее, заряд, накопленный на мембране, остается неизменным, а растворы - нейтральными.
Интересным представляется вопрос, какую долю от общего количества ионов в клетке составляют ионы, накапливающиеся на мембране. Доля их весьма незначительна. Если предположить, что диаметр клетки составляет 25 мкм, то при концентрации 120 ммоль общее количество катионов (а следовательно, и анионов) получится 4 1012
. При мембранном потенциале - 85 мВ величина заряда, разделенного мембраной, составляет приблизительно 5 1011
одновалентных ионов на см2
. При площади поверхности клетки 8 10-5
см2
получается, что на внутренней поверхности мембраны накапливается около 4 107
отрицательных ионов, или одна стотысячная часть общего числа ионов во внутриклеточном растворе. Следовательно, перемещение ионов калия и хлора, достаточное для создания мембранного потенциала, никак не влияет на концентрации ионов в клетке.
На мембранный потенциал нейронов, а также многих других клеток, влияют изменения внеклеточной концентрации калия, но не хлора. Обратимся к модели идеальной клетки. Сделаем допущение (для рассмотрения данной ситуации), что объем внеклеточной жидкости бесконечно велик, и что перемещение ионов не влечет за собой значительного изменения концентрации ионов вне клетки. На рис.1.2А показаны изменения внутриклеточного ионного состава и мембранного потенциала, вызванные повышением внеклеточного уровня калия с 3 до 6 ммоль. Для сохранения первоначальной осмолярности, одновременно с добавлением 3 ммоль калия из раствора удалили 3 ммоль натрия, в результате чего общая концентрация растворенных ионов осталась на уровне 240 ммоль. Увеличение внеклеточной концентрации калия приводит к снижению его трансмембранного градиента, движущего ионы из клетки наружу. При этом на начальном этапе мембранный потенциал не меняется. В результате происходит перенос суммарного положительного заряда внутрь клетки. Вследствие накопления положительного заряда на внутренней поверхности мембраны она деполяризуется. Ионы натрия благодаря деполяризации выходят из состояния равновесия и начинают двигаться внутрь клетки. Перемещение ионов калия и хлора продолжается до тех пор, пока не будет достигнуто новое равновесное состояние, основанное на новом соотношении концентраций и новом уровне мембранного потенциала, в данном примере - 68 мВ.
Вход калия и хлора сопровождается током в клетку некоторого количества воды, приводящим к небольшому увеличению объема клетки. В процессе достижения нового состояния равновесия концентрация калия возрастает с 90 до 91 ммоль, хлора - с 4 до 7,9 ммоль, а объем клетки увеличивается на 3,5%. На первый взгляд кажется, что количество вошедшего в клетку хлора превышает количество калия, но представим себе, каковы были бы эти концентрации, если бы объем клетки не увеличился: концентрации обоих ионов были бы выше на 3,5%. Следовательно, вместо 7,9 ммоль концентрация хлора была бы 8,2 ммоль, а концентрация калия была бы на уровне 94,2 ммоль, то есть увеличение обеих концентраций составило бы 4,2%. Таким образом, сначала калий и хлор входят в клетку в приблизительно равных количествах (за исключением незначительного количества, необходимого для изменения заряда на мембране), а уже затем вода втекает в клетку, снижая концентрации обоих ионов до окончательного уровня.

Рис.1.2 Влияние изменений внеклеточных концентраций ионов на внутриклеточные концентрации и на мембранный потенциал. (А) Внеклеточный уровень калия удвоен, а концентрация натрия снижена с целью сохранения осмолярности на прежнем уровне. (В) Половина ионов внеклеточного хлора заменена на анионы, не проникающие через мембрану (А). Ионные концентрации выражены в миллимолях (ммоль), объем внеклеточного пространства рассматривается как бесконечно большой в сравнении с объемом клетки, поэтому вход и выход ионов через мембрану не изменяет их внеклеточных концентраций.
Сходные рассуждения применимы и в случае увеличения внеклеточной концентрации хлора, с той лишь разницей, что при этом мембранный потенциал не меняется. На рис. 1.2В показан результат 50-процентного снижения концентрации хлора во внеклеточном растворе путем замены 60 ммоль хлора на другой анион, не проникающий через мембрану. Ионы хлора вытекают из клетки, тем самым деполяризуя мембрану до нового хлорного равновесного уровня (-68 мВ). Калий, выведенный из равновесия, также начинает вытекать из клетки. Как и в предыдущем примере, оба иона покидают клетку в равных количествах, увлекая за собой воду. Снижение внутриклеточного уровня калия невелико в процентном отношении, поскольку начальная его концентрация в клетке высока. Напротив, снижение внутриклеточного уровня хлора весьма существенно, и оно влечет за собой значительное изменение равновесного потенциала для хлора. По мере выхода хлора из клетки значение равновесного хлорного потенциала вновь приближается к первоначальному. Процесс продолжается до тех пор, пока равновесные потенциалы для хлора и калия не сравняются и мембранный потенциал не восстановит свое прежнее значение
1. Шлегель Г.Г. История микробиологии.
2. Лотова Л.И. Морфология и анатомия высших растений.
3. Алексеев В.И., Каминский В.А. Прикладная молекулярная биология.
4. Галимов Э.М. Феномен жизни. Происхождение и принципы эволюции.
|
