МИНИСТЕРСТВО ОБРАЗОВАНИЯ РОССИЙСКОЙ ФЕДЕРАЦИИ
Государственное образовательное учреждение
высшего профессионального образования
«Мичуринский государственный педагогический институт»
Кафедра зоологии и экологии
Евстратов Алексей Анатольевич
Экологическая структура населения жужелиц (
Caleoptera
,
Carabidae
) плодовых садов
Курсовая работа
Научный руководитель –
доцент, к.б.н.
Касандрова Любовь Ивановна
МИЧУРИНСК
2008
Содержание
Введение
Глава I. Материал и методы работы
1.1. Природно-климатическая характеристика района исследования
1.2. Методы учёта и сбора жужелиц
1.3. Методы анализа структуры населения жужелиц
Глава II. Видовой состав населения жужелиц плодовых садов
Глава III.Экологическая структура населения жужелиц плодовых садов
3.1. Экологические группы жужелиц по биотопическому преферендуму
3.2. Экологические группы жужелиц по степени увлажнения предпочитаемых биотопов
Заключение
Библиографический список
Введение
В борьбе с вредителями сельскохозяйственных культур получило широкое распространение сочетание химического метода с биологическим. В связи с этим большое практическое значение приобретает изучение хищных насекомых, среди которых заслуживают внимания жуки из семейства жужелиц (Carabidae).
Жужелицы являются хорошими индикаторами почвенных и растительных условий. На них проведены исследования по зоогеографии, географической изменчивости, жизненным формам.
Семейство жужелиц – важнейший компонент почвенного населения беспозвоночных животных. Жужелицы встречаются практически во всех ландшафтах суши и тонко реагируют на изменения почвенно-растительных и микроклиматических условий, поэтому они используются как биоиндикаторы. (Мазохин - Поршняков, 2001).
Семейство жужелиц очень крупное. Оно насчитывается около 30000 видов жужелиц, из них около 2300 видов в России (Крыжановский, 2002). Размеры жуков колеблются в большом диапазоне: от мелких (1,2 мм) до крупных (90 мм). У некоторых усики нитевидные, у части жуков ноги длинные, бегательные. Надкрылья иногда срастаются по шву, крылья бывают недоразвитые. Окраска чаще чёрная, бурая, с металлическим отливом, реже пёстрая. У многих развиты анальные железы, выделяющие защитную жидкость. Личинки камподеовидные.
Реклама

Жуки являются важной составной частью естественных биоценозов и агроценозов. Семейство Carabidae обладает широкой экологической пластичностью (Шарова, 1981). Они различаются по способам питания, занимаемым ярусам, местообитаниям, сезонной и суточной активностью. Жужелицы обитают в почве и на её поверхности, реже на деревьях, в древесине, муравейниках, термитниках, пещерах. Большое разнообразие наблюдается в суточной и сезонной активности жужелиц. Условно можно разделить жужелиц на три группы: виды с дневной, ночной активностью и безразличных в этом отношении (Крыжановский, 1983). Активность в сезоне обычно связана с особенностями биологии вида, с оптимальным сочетанием температуры и влажности.
Жужелицы преимущественно мезо- и гигрофилы, поэтому они активны в умеренно теплые и влажные сезоны года.
По способам питания жужелиц можно условно разделить на три основные группы: зоофаги – хищники, фитофаги – растительноядные, миксофитофаги – жужелицы, обладающие смешанным питанием (Крыжановский, 1983; Шарова, 1981).
Жужелицы играют заметную роль в ограничении численности многих беспозвоночных животных, а сами входят как компонент в рацион позвоночных. Личинки, склонные к сапрофагии, принимают активное участие в почвообразовательных процессах. Большинство жужелиц – многоядные хищники, питающиеся почвенными беспозвоночными. Так, красотелы поедают гусениц, настоящие жужелицы (Carabus) истребляют многие виды насекомых и моллюсков. Растительноядные формы могут повреждать культурные растения, например, хлебная жужелица (Zabrustenebrioides) опасна для полей пшеницы. Личинки ряда жужелиц, например, бомбардиров, – эктопаразиты куколок других жуков.
Общеизвестна полезная роль жужелиц-хищников в уничтожении ряда вредителей полевых культур и лесов. Наибольшее хозяйственное значение имеют крупные хищные жужелицы. Например, за 20 дней виды рода Carabus, Pterostichus на 1 гектаре картофеля уничтожают 1,6-2млн. личинок колорадского жука (Scherney, 1962, цит. по Касандровой, 1970). Жужелицы средних и мелких размеров при высокой их численности тоже являются эффективными энтомофагами. Например, Pseudoophonuspubesceus (=Harpalusrufipes), AmaraaulicaPz. и др. уничтожают личинок различных видов щелкунов (Иняева, 1965). Неоднократно отмечалась роль жужелиц в снижении численности хрущей (Иняева, 1965; Пацановский, 1957).
Роль личинок жужелиц в сельском хозяйстве значительна. Личинки жужелиц способны уменьшить численность проволочников настолько, что всякая борьба с ними становится излишней (Григорьева, 1950; Иняева, 1965; Масайтис, 1929; Черепанов, 1957; Шуровенков, 1962 и др.).
Реклама

Личинки жужелиц ещё больше, чем взрослые жуки, предпочитают животную пищу. Лишь при недостатке воды или отсутствии подходящей животной пищи личинки жужелиц питаются тканями растений. Растительноядными преимущественно бывают личинки родов Zabrus, Harpalus и Amara.
В лесном хозяйстве, кроме видов рода Calosoma и рода Carabus, к числу полезных относят жужелиц родов Dromius, Tachyta, Agonum (Померанцев, 1949).
Отмечена полезная роль жужелиц и в плодовых садах, в которых Carabidae представлены хищниками и миксофитофагами. В число их жертв входит ряд серьёзных вредителей: яблонная плодожорка, яблонная моль, восточная плодожорка, гусеницы кольчатого шелкопряда. В умеренных широтах личинок гусениц яблонной моли и яблонной плодожорки поедают жуки Ophonusrufipes (=Harpalusrufipes), Anisodactylussignatus, Amaraingenua, Pterostichusversicolor (=Poecilusversicolor) и др. (Касандрова, 2005). В садах Крыма Broscuscephalotes, Pterostichusmelanarius. Harpalusrufipes, Anisodactylussignatus, Licinusdepressus питаются куколками и гусеницами старших возрастов яблонной плодожорки, совок, пядениц, листовёрток, личинками жуков долгоносиков, паутинными клещами (Эйдельберг, 1985).
По обилию видов и численности, а также по биологической роли на сельскохозяйственных землях жужелицы имеют особое значение. Личинок жужелиц по характеру питания можно объединить в несколько экологических групп (Тишлер, 1971): хищники, эктопаразиты, сапрофаги и фитофаги.
Путём обследования содержимого желудка и кишечника 12 наиболее обычных видов жужелиц на клеверных полях Чехии были получены точные данные о составе их пищи (Skyhravy, 1958, цит. по Тишлеру, 1971). У Carabus и Brachynus в кишечнике не найдено частиц пищи, потому что у видов этих родов пищеварение внекишечное. У Calathusfuscipes и Pterostichuslepidus пищевые остатки состояли из тлей, муравьев и гусениц бабочек. О видах Amara и Zabrus уже давно известно, что они фитофаги (Кряжева, 1965). Многие Amara питаются семенами злаковых трав и сорняков (Кочетова, 1936). В ходе развития важнейшие виды родов Harpalus, Zabrus и Amara от исходного хищничества перешли через всеядность к преимущественно растительноядному образу жизни. Это относится как к личинкам, так и к взрослым особям.
У жужелиц выявлены враги и паразиты (Иняева, 1965; Тишлер, 1971). Врагами жужелиц являются грачи (Corvus), скворцы (Sturnus) и многие другие птицы, а также кроты (Talpa) и мыши. Из мелких животных крупные коротконадкрылые жуки, или стафилиниды, могут парализовать и убивать жужелиц своей токсично действующей слюной. Характерным паразитом жужелиц является муха Vivianiacinerea. Оплодотворённые самки нематод из рода Heterotylenchus внедряются в куколок мелких жужелиц (Heterotylenchusbovieni в виды Bembidion, Heterotylenchusstammeri в Clivinafossor), в полости тела которых они выращивают своих многочисленных личинок.
Шесть видов жужелиц занесены в Красную книгу России, а двадцать девять видов – в Красную книгу Тамбовской области (Белевитин, Ганжа, Касандрова, 2000; Тихомиров, 2002).
Изучение жужелиц представляет интерес с двух сторон:
1) использование их в биологическом методе борьбы с вредителями сельскохозяйственных культур, с учётом того, что большинство видов этого семейства являются многоядными хищниками и полифагами;
2) использование в качестве индикаторов почвенных и растительных условий, так как хищники и полифаги не строго лимитируются в своём распределении одним лишь фактором, в первую очередь наличием определённых видов растений, как это бывает у фитофагов (Гиляров, 1965).
Обе стороны изучаемого объекта тесно связаны между собой. Выявление видового состава жужелиц их распределение в различных биотопах представляет интерес и с точки зрения использования жужелиц для биологической защиты растений (Н.В. Яценко, Л.Ф. Скрылёва, Л.И. Касандрова и др., 2005).
По изучению состава и структуры населения жужелиц в различных ландшафтах имеется большая и разносторонняя литература (Гиляров, 1978; Душенков, Черняховская, 1982; Булохова, 1995а, 1995б; Шарова и др., 1997). Особое внимание в последнее время уделялось изучению жужелиц агроценозов (Иняева, 1965; Шарова. 1969; Касандрова, 1977; Карпова, 1984а, 1984б; Будилов, 1989, 1990а, 1990б; Шарова, Попова, 1990; Романкина, 1994).
Особенно детально изучены жужелицы полевых агроценозов (Аверин, 1939; Кряжева, 1963 - 1965; Мордкович, 1964). Меньше опубликовано работ го изучению жужелиц плодовых садов, но интерес к ним возрастает и в России, и за рубежом: в Канаде (Hagley, 1975), в ФРГ (Steiner, 1974; Beijer, 1981), в Польше (Gzechowski, 1982), в Венгрии (Kadar, Szel, 1989; Kadar, Szentkiralyi, 1991), в Прикарпатье и Закарпатье (Егоров, 1976), в Узбекистане (Дадамирзаев, Давлетшина, 1982), в предгорье Копетдага (Сабирова, 1984), в Молдавии (Мацюк, Верлан, 1987), в Крыму (Эйдельберг, 1989). В нашей стране: в Среднем Поволжье (Утробина, 1964); в Алтайском заповеднике (Хмельков, Ковригин, 1985); в Дагестане (Сайпулаева, 1986) и др. (цит. по Романкиной, 2001).
В Тамбовской области изучение населения жужелиц садов было начато в 1965 году Л.И. Касандровой и в 1970 году защищена кандидатская диссертация, опубликован ряд работ (Касандрова, 1970, 1977, 1978, 1981, 1982, 1989, 1997). К этим исследованиям в дальнейшим подключилась М.Ю. Романкина, защитившая кандидатскую диссертацию в 1996 году, тоже давшая ряд публикаций (Романкина, 1994, 2000, 2001). Изучены видовой состав, распределение, экологическая структура, трофические связи. В последние годы была составлена обобщающая сводка по хищным жукам из семейства жужелиц и насекомоядных птиц промышленных садов Мичуринского района Тамбовской области (В.Н. Яценко, Л.Ф. Скрылёва, Л И. Касандрова и др., 2005), и видовой состав жужелиц (Coleoptera, Carabidae), в состав которой вошли сведения о видовом составе жужелиц садов Тамбовской области.
В Тамбовской области изучена карабидофауна плодовых садов на плакоре и в меньшей мере пойменных садов (Касандрова, Родюкова, 1987; Касандрова, Тимохова, 2000).
В основе данной работы положены материалы 1965-1969 гг. В это время экологическая характеристика жужелиц была недостаточно ясна. Разные авторы один и тот же вид жужелиц нередко относили к разным экологическим группам. Впервые Шарова (1978) в сводке о жужелицах Московской области уточнила экологические характеристики более 200 видов жужелиц. Эта характеристика в дальнейшем стала общепринятой. В связи с этим выбрана тема данной работы.
Целью настоящей работы было выяснение эколого-фаунистической структуры населения жужелиц в плодовых садах бывшей ЦГЛ и ВНИИС им. И.В. Мичурина.
Задачи:
1. Уточнение видовой структуры населения жужелиц в плодовых садах.
2. Выяснение экологической структуры населения жужелиц садов по биотопическому преферендуму.
3. Выяснение экологической структуры населения жужелиц садов по степени увлажнения биотопов.
4. Выяснение влияния на экологическую структуру полива садов в условиях лесостепной зоны.
Объём работы: работа изложена на 35 страницах печатного текста, состоит из введения, 3-х глав и заключения. В работе 8 таблиц и 2 рисунка. Библиографический список содержит 30 названий.
Глава
I
. Материал и методы работы
1.1.
Природно-климатическая характеристика района исследования
Весь материал для анализа видовой и экологической структуры населения жужелиц был предоставлен кафедрой зоологии и экологии МГПИ.
Основные исследования были проведены в течение трёх вегетационных периодов (1965, 1967, 1968 гг.) в садах Центральной генетической лаборатории (ЦГЛ) им. И.В. Мичурина и Всесоюзного научно-исследовательского института (ВНИИС) им. И.В. Мичурина Мичуринского района Тамбовской области (Касандрова, 1970).
Тамбовская область расположена в лесостепной зоне, занимая часть Окско-Донской низменности. Рельеф равнинный, преобладающий ландшафт – степной. Наибольшие лесные массивы имеются на западе области на территории Мичуринского района. Почвы, характерные для этого района – выщелоченные чернозёмы и серые лесные.
Климат Тамбовской области умеренно-континентальный с устойчивой зимой и преобладанием тёплой, нередко полузасушливого характера погодой в летний период. Вегетационный период начинается с переходом средней суточной температуры воздуха через –5°С (15-17 апреля) и заканчивается с падением её ниже +5°С осенью (14-18 октября). В ноябре среднесуточная температура опускается ниже 0°С, промерзает почва, выпадает снег. Количество атмосферных осадков в отдельные годы очень неустойчиво. В городе Мичуринске, по многолетним данным, за год выпадает 475-500 мм атмосферных осадков.
За период исследования погода заметно различалась по годам (табл. 1). 1965 год – полузасушливый. Он отличался низкой суммой положительных температур и неравномерным их распределением в течение вегетационного периода: тёплая погода сменялась резким похолоданием во второй декаде июня, первой и второй декадах июля и второй декаде августа (табл. 2, рис.2). 1967 год характеризовался наиболее благоприятными погодными условиями с устойчивыми среднесуточными температурами с мая по сентябрь и постепенным и понижением в конце вегетационного периода. Похолодание было только в первой декаде июня. Обильные осадки с конца мая выпадали через одну декаду. 1968 год – год избыточного увлажнения (за вегетационный период выпало осадков около годовой нормы) и неблагоприятного сочетания метеоэлементов: похолодания сопровождались обильными осадками в первой декаде мая и ливневыми дождями в июле. Август был засушливым, особенно третья декада (0,1 мм осадков при среднедекадной температуре –21,6°С). Почва после переувлажнения покрылась растрескивающейся коркой.
Рис. 1. Карта Тамбовской области.
Условные обозначения:
 – граница подзон северной и типичной лесостепи – граница подзон северной и типичной лесостепи
Таблица 1
Колебание сумм температур воздуха и количества осадков в окрестностях города Мичуринска
| Годы |
Сумма за вегетационный период |
| температур воздуха (в град.) |
осадков (в мм)
|
дней с осадками более 1 мм |
| 1964 |
2456,2 |
358,9 |
49 |
| 1965 |
2265,0 |
317,3 |
48 |
| 1966 |
2711,1 |
255,5 |
45 |
| 1967 |
2631,7 |
380,0 |
45 |
| 1968 |
2567,3 |
448,4 |
58 |
| Средние многолетние |
2719,0 |
303,0 |
Обследованные сады расположены на правом берегу реки Лесной Воронеж в 4 км к западу от г. Мичуринска, занимая площадь около 100 га. Со всех сторон сады окружены ветроломными лесополосами, состоящими из тополей и кустарников, в основном представленных жёлтой акацией. С юго-восточной стороны они отделены небольшим прогоном (200-300 м) от широколиственной рощи, с западной – выходят к реке. На территории садов имеются два небольших водоёма для стока воды в период сильных дождей и таяния снега (рис. 2). Почвы в исследованных садах – серые лесные, их механический состав – лёгкие и средние суглинки.
В качестве опытных были взяты два яблоневых сада, возраст которых свыше тридцати лет. В зависимости от содержания почвы, сады получили названия:
«Пар I» – междурядья в саду в течение ряда лет находились под чёрным паром. Их обработка (культивация, боронование, дискование) в течение тёплого периода года была тщательной и прекращалась только во время уборки урожая. Осенью проводили зяблевую вспашку. До 1967 года в этом саду был искусственный полив летом.
«Пар II» – междурядья тоже в течение ряда лет находились под чёрным паром, но сад не подвергался искусственному поливу, поэтому накопление влаги в почве было ниже, чем в саду «пар I». Вследствие некоторого снижения требований к агротехнике почва в этом саду частично в 1967 году, особенно в 1968 году стала покрываться разнотравьем; местами встречались дернинки. Этому процессу способствовали обильные осадки в 1968 году.
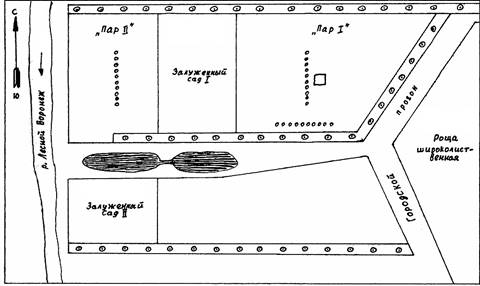
Рис. 2. Схема расположения опытных садов в окрестностях г. Мичуринска и ловушек в них
Условные обозначения:
 – водоёмы; – водоёмы;
 – ловушки. – ловушки.
1.2.
Методы учёта и сбора жужелиц
Жужелицы были собраны с помощью почвенных ловушек, в качестве которых были использованы стеклянные банки ёмкостью 0,5 л. С диаметром входного отверстия 7,5 см. Ловушки закапывали в землю так, чтобы край входного отверстия находился на уровне поверхности земли. Почву вокруг ловушки слегка утрамбовывали, чтобы после оседания её входное отверстие ловушки не оказалось выше уровня земли (в каждом саду находилось по 10 ловушек). На дно ловушек насыпали слой почвы примерно на ¼ часть высоты ловушки. Жуки, попавшие в ловушки, закапывались в почву на её дне, почти не делая попыток к бегству.
Проверку ловушек и подсчёт имаго жужелиц в 1965 году проводили через 3-5 дней, в 1967 и 1968 годах – строго через 5 суток в течение всего вегетационного периода. Одновременно с подсчётами жужелиц, подсохшую землю заменяли свежей, протирали внутренние стекла ловушек.
Ловушки расставляли в приствольных кругах плодовых деревьев на расстоянии не более 0,5 м от ствола дерева во избежание их потерь, а особенно во время частых обработок почвы в садах. Приствольные круги обрабатывались значительно реже. Кроме того, кроны деревьев, создавая тень, предохраняли в жаркие дни почву в ловушках от сильного иссушения, а в дождливую погоду от наполнения их водой, обеспечивая относительно благоприятные условия для временного пребывания жужелиц в них. В каждом саду находилось по 10 ловушек, которые располагались в центре сада, в одну линию в рядах деревьев. Одна ловушка от другой стояла на расстоянии 10 м. В саду «пар I», кроме ловушек в центре сада, стояла вторая группа ловушек на окраине, в первом ряду от лесополосы и широколиственной рощи. По расположению эти группы ловушек были взаимно перпендикулярны друг другу.
1.3. Методы анализа структуры населения жужелиц
Для определения жужелиц использовали бинокулярный микроскоп МБС, ручную лупу, пинцет. Идентификацию видов жужелиц провели по определителю насекомых Европейской части СССР (ред. Бей-Биенко, 1965. Т.2).
Участие того или иного вида жужелиц выражено в индексах доминирования по шкале Ренконена (Renkonen, 1932, цит. по Шишовой, 1992). К доминирующими видам были отнесены те виды, обилие которых составляло 5% и более от общего числа жужелиц, отловленных в данном биотопе, к субдоминантам – с обилием от 2 до 5%, к редким – с обилием менее 2%, к супердоминантам – более 50%.
Динамическая плотность жужелиц выражена в числе экземпляров на 10 ловушко-суток.
Степень сходства населения жужелиц рассчитали по формуле Жаккара (Чернов, 1972):
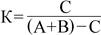 , ,
где
С – число видов для двух сравниваемых биотопов,
А – число видов в первом биотопе,
В – число видов во втором биотопе.
Экологическая характеристика видов жужелиц по биотопическому преферендуму дана по Шаровой (1981).
Глава
II
. Видовая структура населения жужелиц плодовых садов
Сады представляют собой агробиоценозы, возникшие в результате деятельности человека. Они характеризуются высокой однородностью растительного покрова. Относительная устойчивость органического состава агробиоценозов характерна и для их фауны. В целом она характеризуется обеднением видового состава населения по сравнению с первичными биоценозами, меньшим числом доминантов и высокой их численностью.
В отличие от полевых агробиоценозов для садов характерна более высокая стабильность экологических условий, так как плодовые культуры являются многолетними. Фауна жужелиц обследованных садов по видовому разнообразию, мало уступает фауне жужелиц полей. Так, в агроценозах кормоовощного севооборота А.А. Попова (1986) сообщает о наличии 71-ого вида жужелиц из 27-ми родов. Видовой состав жужелиц в садах также разнообразен. Касандрова (1970) зарегистрировала 65 видов жужелиц из 22 родов в 6 садах. Романкина (1996) сообщает о 52-х видах из 7-ми садов.
Таблица 2.
Состав доминантов в садах в окрестностях города Мичуринска
| Доминирующие виды жужелиц |
1965 |
1967 |
1968 |
| 1. Amara communis (Panzer, 1797) |
+ |
+ |
+ |
| 2. Amara ingenue (Duftschmid, 1812) |
+ |
+ |
+ |
| 3. Anisodactylussignatus (Panzer, 1797) |
+ |
+ |
+ |
| 4. Bembidionlampros (Herbst, 1784) |
– |
– |
+ |
| 5. Broscus cephalotes (Linnaeus, 1758) |
– |
– |
+ |
| 6. Harpalusdistinguendus (Duftschmid, 1812) |
+ |
+ |
– |
| 7. Microlestes minutulus (Goeze, 1777) |
– |
+ |
– |
| 8. Clivina fossor (Linnaeus, 1758) |
+ |
– |
+ |
| 9. Harpalusrufipes (De Geer, 1774) |
+ |
+ |
+ |
| 10. Poecilus cupreus (Linnaeus, 1758) |
+ |
+ |
+ |
| 11. Poecilus versicolor (Sturm, 1824) |
+ |
+ |
+ |
Число доминантов
Численность в %
|
8
91,4
|
8
95,2
|
9
91,3
|
«+» – доминировал «–» – нет
В обследованных садах в течение трёх лет зарегистрировано высокое число видов (46) жужелиц.
Таблица 3.
Видовой состав и биотопическая характеристика фауны жужелиц в садах ЦГЛ
Виды
|
Биотопическая характеристика |
Присутствие или отсутствие жуков в ловушках |
| 1965 |
1967 |
1968 |
| 1. Calosoma investigator (Illiger, 1798) |
лс |
+ |
+ |
+ |
| 2. Cylindera germanica (Linnaeus, 1758) |
лг |
– |
– |
+ |
| 3. Clivina fossor (Linnaeus, 1758) |
лг, бл |
+ |
+ |
+ |
| 4. Broscus cephalotes (Linnaeus, 1758) |
п |
+ |
+ |
+ |
| 5. Calathus halensis (Schaller, 1783) |
лс |
+ |
+ |
– |
| 6. Calathus ambiguus (Paykull, 1790) |
лг, п |
+ |
+ |
+ |
| 7. Calathus erratus (C.R. Sahlberg, 1827) |
лг, п |
– |
+ |
+ |
| 8. Calathus melanocephalus (Linnaeus, 1758) |
лс |
+ |
+ |
+ |
| 9. Synuchus vivalis (Illiger, 1797) |
лс, бл |
+ |
+ |
+ |
| 10. Bembidion biguttatum (Fabricius, 1779) |
бл |
– |
+ |
– |
| 11. Bembidion bipunctatum (Linnaeus, 1761) |
лг, п |
– |
+ |
– |
| 12. Bembidion lampros (Herbst, 1784) |
лг, п |
+ |
+ |
+ |
| 13. Bembidion quadrimaculatum (Linnaeus, 1761) |
лг, п |
+ |
+ |
+ |
| 14. Agonum muelleri (Herbst, 1784) |
лг, п |
– |
+ |
– |
| 15. Stomis pumicatum (Panzer, 1796) |
лс, бл |
– |
+ |
– |
| 16. Poecilus cupreus (Linnaeus, 1758) |
лг, п |
+ |
+ |
+ |
| 17. Pterostichus anthracinus (Illiger, 1798) |
лг, п |
+ |
– |
+ |
| 18. Poecilus lepidus (Leske, 1785) |
лг, п |
+ |
+ |
+ |
| 19. Pterostichus melanarius (Illiger, 1798) |
лс |
+ |
+ |
+ |
| 20. Pterostichus nigrita (Paykull, 1790) |
лс, бл |
+ |
+ |
– |
| 21. Pterostichus oblongopunctatus (Fabricius,1787) |
лс |
+ |
– |
+ |
| 22. Poecilus punctulatus (Schaller, 1783) |
п |
– |
+ |
+ |
| 23. Poecilus versicolor (Sturm, 1824) |
лг, п |
+ |
+ |
+ |
| 24. Microlestes minutulus (Goeze, 1777) |
лг, п |
+ |
+ |
+ |
| 25. Amara bifrons (Gyllenhal, 1810) |
лг, п |
– |
+ |
– |
| 26. Amara aenea (De Geer, 1774) |
лг, п |
+ |
+ |
+ |
| 27. Amara apricaria (Paykull, 1790) |
лг, п |
+ |
+ |
+ |
| 28. Amara communis (Panzer, 1797) |
лг, п |
+ |
+ |
– |
| 29. Amara familiaris (Duftschmid, 1812) |
лг, п |
+ |
– |
+ |
| 30. Amara fulva (O. Müller, 1776) |
п |
+ |
+ |
– |
| 31. Amara ingenue (Duftschmid, 1812) |
п |
+ |
+ |
+ |
| 32. Amara similata (Gyllenhal, 1810) |
лг, п |
+ |
+ |
– |
| 33. Amara ovata (Fabricius, 1792) |
лг |
– |
– |
+ |
| 34. Harpalus rufipes (De Geer, 1774) |
п |
+ |
+ |
+ |
| 35. Harpalus griseus (Panzer, 1797) |
п |
+ |
+ |
+ |
| 36. Harpalus affinis (Schrank, 1781) |
лг, п |
– |
+ |
– |
| 37. Harpalus calceatus (Duftschmid, 1812) |
ст |
– |
+ |
– |
| 38. Harpalus distinguendus (Duftschmid, 1812) |
лг, п |
+ |
+ |
+ |
| 39. Harpalus froelichi (Sturm, 1818) |
ст |
– |
+ |
– |
| 40. Harpalus latus (Linnaeus, 1758) |
лс |
– |
+ |
– |
| 41. Harpalus luteicornis (Duftschmid, 1812) |
лг |
– |
+ |
– |
| 42. Harpalus tardus (Panzer, 1797) |
лг, п |
– |
– |
+ |
| 43. Harpalus smaragdinus (Duftschmid, 1812) |
лг, п |
+ |
+ |
– |
| 44. Harpalus zabroides (Dejean, 1829) |
ст |
+ |
+ |
+ |
| 45. Anisodactylus signatus (Panzer, 1797) |
п |
+ |
+ |
+ |
| 46. Anisodactylus binotatus (Fabricius, 1787) |
п |
+ |
– |
+ |
| Итого |
31 |
39 |
30 |
| Общих видов: 21 (45,7%) |
| Специфичных видов: 14 (30,4%) |
1 |
10 |
3 |
Условные обозначения:
Биотопическая характеристика: лс – лесная; бл – болотная; лс, бл – лесо-болотная; лг, бл – лугово-болотная; лг – луговая; п – полевая; лг, п – лугово-полевая; ст – степная.
Таблица 4.
Родовой состав жужелиц изучаемых садов
| Род |
Число видов |
% |
Экз. |
% |
| Agonum Bonelli, 1810 |
1 |
2,2 |
1 |
0,01 |
| Amara Bonelli, 1810 |
9 |
19,6 |
2965 |
23,28 |
| Anisodactylus Dejеan, 1829 |
2 |
4,3 |
1553 |
12,20 |
| Bembidion Latreille, 1802 |
4 |
8,7 |
157 |
1,23 |
| Broscus Panzer, 1813 |
1 |
2,2 |
47 |
0,37 |
| Calathus Bonelli, 1810 |
4 |
8,7 |
35 |
0,28 |
| Calosoma F. Weber, 1801 |
1 |
2,2 |
9 |
0,07 |
| Clivina Latreille, 1802 |
1 |
2,2 |
200 |
1,57 |
| Cylindera Westwood, 1831 |
1 |
2,2 |
1 |
0,01 |
| Harpalus Latreille, 1802 |
11 |
23,9 |
7150 |
56,14 |
| Microlestes Schmidt – Gobel, 1846 |
1 |
2,2 |
14 |
0,11 |
| Poecilus Bonelli, 1810 |
4 |
8,7 |
571 |
4,48 |
| Pterostichus Bonelli, 1810 |
4 |
8,7 |
19 |
0,15 |
| Stomis Clairville, 1806 |
1 |
2,2 |
1 |
0,01 |
| Synuchus Gyllenhal, 1810 |
1 |
2,2 |
12 |
0,09 |
| Всего |
46 |
100 |
12735 |
100 |
Таблица 5.
Видовой состав жужелиц сада «Пар I» (окраина)
| Виды |
Количество экземпляров |
%
|
| 1967 |
1968 |
Всего |
| 1. Calosoma investigator |
2 |
– |
2 |
0,10 |
| 2. Clivina fossor |
5 |
21 |
26 |
0,33 |
| 3. Broscus cephalotes |
1 |
– |
1 |
0,05 |
| 4. Calathus erratus |
1 |
– |
1 |
0,05 |
| 5. Calathusmelanocephalus |
1 |
– |
1 |
0,05 |
| 6. Synuchus vivalis |
5 |
– |
5 |
0,26 |
| 7. Bembidionlampros |
– |
2 |
2 |
0,10 |
| 8. Bembidionquadrimaculatum |
– |
1 |
1 |
0,05 |
| 9. Poecilus cupreus |
12 |
– |
12 |
0,61 |
| 10. Pterostichus anthracinus |
– |
1 |
1 |
0,05 |
| 11. Poeciluslepidus |
1 |
2 |
3 |
0,15 |
| 12. Pterostichus nigrita |
1 |
– |
1 |
0,05 |
| 13. Pterostichus oblongopunctatus |
– |
3 |
3 |
0,15 |
| 14. Poecilus versicolor |
5 |
16 |
21 |
1,07 |
| 15. Amara bifrons |
1 |
– |
1 |
0,05 |
| 16. Amara aenea |
1 |
– |
1 |
0,05 |
| 17. Amara apricaria |
– |
1 |
1 |
0,05 |
| 18. Amara ingenue |
633 |
52 |
685 |
35,04 |
| 19. Harpalusrufipes |
589 |
217 |
806 |
41,23 |
| 20. Harpalusgriseus |
– |
1 |
1 |
0,05 |
| 21. Harpalus affinis |
2 |
– |
2 |
0,10 |
| 22. Harpalusdistinguendus |
41 |
– |
41 |
2,10 |
| 23. Anisodactylussignatus |
254 |
82 |
336 |
17,19 |
| Всего экземпляров |
1555 |
400 |
1955 |
100 |
| Всего видов |
17 |
13 |
23 |
Таблица 6.
Видовой состав жужелиц сада «Пар I» (центр)
Виды
|
Количество экземпляров |
%
|
| 1965 |
1967 |
1968 |
Всего |
| 1. Calosoma investigator |
– |
– |
2 |
2 |
0,03 |
| 2. Cylindera germanica |
– |
– |
1 |
1 |
0,01 |
| 3. Clivina fossor |
62 |
17 |
43 |
122 |
1,72 |
| 4. Broscus cephalotes |
1 |
2 |
– |
3 |
0,04 |
| 5. Calathus halensis |
2 |
1 |
– |
3 |
0,04 |
| 6. Calathus ambiguus |
4 |
2 |
5 |
11 |
0,16 |
| 7. Calathus erratus |
– |
4 |
6 |
10 |
0,14 |
| 8. Calathus melanocephalus |
2 |
– |
– |
2 |
0,03 |
| 9. Synuchus vivalis |
3 |
1 |
– |
4 |
0,06 |
| 10. Bembidionlampros |
– |
7 |
2 |
9 |
0,13 |
| 11. Bembidionquadrimaculatum |
4 |
1 |
3 |
8 |
0,11 |
| 12. Stomis pumicatum |
– |
1 |
– |
1 |
0,01 |
| 13. Poecilus cupreus |
37 |
30 |
2 |
69 |
0,97 |
| 14. Poeciluslepidus |
10 |
3 |
3 |
16 |
0,23 |
| 15. Pterostichus melanarius |
2 |
1 |
– |
3 |
0,04 |
| 16. Pterostichus oblongopunctatus |
1 |
– |
1 |
2 |
0,03 |
| 17. Poecilus versicolor |
9 |
13 |
12 |
34 |
0,48 |
| 18. Microlestes minutulus |
– |
2 |
– |
2 |
0,03 |
| 19. Amara bifrons |
– |
1 |
– |
1 |
0,01 |
| 20. Amara aenea |
2 |
1 |
– |
3 |
0,04 |
| 21. Amara apricaria |
7 |
4 |
8 |
19 |
0,27 |
| 22. Amara familiaris |
– |
– |
5 |
5 |
0,07 |
| 23. Amara fulva |
1 |
1 |
– |
2 |
0,03 |
| 24. Amara ingenue |
51 |
1696 |
89 |
1836 |
25,94 |
| 25. Harpalusrufipes |
1350 |
1916 |
309 |
3575 |
50,50 |
| 26. Harpalusgriseus |
10 |
1 |
1 |
12 |
0,17 |
| 27. Harpalus affinis |
– |
1 |
– |
1 |
0,01 |
| 28. Harpaluscalceatus |
– |
1 |
– |
1 |
0,01 |
| 29. Harpalusdistinguendus |
6 |
461 |
5 |
472 |
6,67 |
| 30. Harpalusfroelichi |
– |
14 |
– |
14 |
0,20 |
| 31. Harpalustardus |
– |
– |
2 |
2 |
0,03 |
| 32. Harpalus zabroides |
1 |
1 |
3 |
5 |
0,07 |
| 33. Anisodactylussignatus |
97 |
560 |
166 |
823 |
11,63 |
| 34. Anisodactylus binotatus |
1 |
– |
1 |
2 |
0,03 |
| Всего экземпляров |
1665 |
4731 |
683 |
7079 |
100 |
| Всего видов |
24 |
27 |
22 |
34 |
Таблица 7.
Видовой состав жужелиц сада «Пар II»
| Виды |
Количество экземпляров |
%
|
| 1965 |
1967 |
1968 |
Всего |
| 1. Calosoma investigator |
– |
– |
2 |
2 |
0,05 |
| 2. Clivina fossor |
5 |
16 |
31 |
52 |
1,40 |
| 3. Broscus cephalotes |
17 |
10 |
16 |
43 |
1,16 |
| 4. Calathus halensis |
– |
2 |
– |
2 |
0,05 |
| 5. Calathus ambiguus |
– |
1 |
– |
1 |
0,03 |
| 6. Calathus erratus |
– |
2 |
1 |
3 |
0,08 |
| 7. Calathusmelanocephalus |
– |
– |
1 |
1 |
0,03 |
| 8. Synuchus vivalis |
– |
– |
3 |
3 |
0,08 |
| 9. Bembidion biguttatum |
– |
1 |
– |
1 |
0,03 |
| 10. Bembidionbipunctatum |
– |
1 |
– |
1 |
0,03 |
| 11. Bembidionlampros |
10 |
4 |
92 |
106 |
2,86 |
| 12. Bembidionquadrimaculatum |
15 |
4 |
10 |
29 |
0,78 |
| 13. Agonum muelleri |
– |
1 |
– |
1 |
0,03 |
| 14. Poecilus cupreus |
41 |
37 |
36 |
114 |
3,08 |
| 15. Pterostichus anthracinus |
1 |
– |
– |
1 |
0,03 |
| 16. Poeciluslepidus |
17 |
9 |
11 |
37 |
1,00 |
| 17. Pterostichus melanarius |
– |
– |
2 |
2 |
0,05 |
| 18. Pterostichus nigrita |
1 |
– |
– |
1 |
0,03 |
| 19. Pterostichus oblongopunctatus |
4 |
– |
1 |
5 |
0,14 |
| 20. Poecilus punctulatus |
– |
1 |
1 |
2 |
0,05 |
| 21. Poecilus versicolor |
48 |
40 |
175 |
263 |
7,10 |
| 22. Microlestes minutulus |
1 |
10 |
1 |
12 |
0,32 |
| 23. Amara bifrons |
– |
2 |
– |
2 |
0,05 |
| 24. Amara aenea |
2 |
– |
1 |
3 |
0,08 |
| 25. Amara apricaria |
1 |
4 |
– |
5 |
0,14 |
| 26. Amara communis |
2 |
1 |
– |
3 |
0,08 |
| 27. Amara familiaris |
6 |
– |
9 |
15 |
0,41 |
| 28. Amara fulva |
1 |
– |
– |
1 |
0,03 |
| 29. Amara ingenue |
115 |
233 |
30 |
378 |
10,21 |
| 30. Amara similata |
1 |
1 |
– |
2 |
0,05 |
| 31. Amara ovata |
– |
– |
2 |
2 |
0,05 |
| 32. Harpalusrufipes |
108 |
1061 |
912 |
2081 |
56,21 |
| 33. Harpalusgriseus |
1 |
2 |
5 |
8 |
0,22 |
| 34. Harpalus affinis |
– |
3 |
– |
3 |
0,08 |
| 35. Harpalusdistinguendus |
15 |
93 |
10 |
118 |
3,19 |
| 36. Harpalusfroelichi |
– |
2 |
– |
2 |
0,05 |
| 37. Harpalus latus |
– |
2 |
– |
2 |
0,05 |
| 38. Harpalusluteicornis |
– |
1 |
– |
1 |
0,03 |
| 39. Harpalussmaragdinus |
2 |
1 |
– |
3 |
0,08 |
| 40. Anisodactylussignatus |
60 |
125 |
206 |
391 |
10,56 |
| Всего экземпляров |
474 |
1670 |
1558 |
3702 |
100 |
| Всего видов |
23 |
29 |
23 |
40 |
Таблица 8.
Массовые виды жужелиц
| Виды |
Пар I |
Пар II |
| окраина |
центр |
| 1 . Harpalusrufipes |
41,2
—————
|
50,5
—————
|
56,2
—————>
|
| 2. Amaraingenua |
35
<————
|
25,9
—————
|
10,2
—————
|
| 3. Anisodactylussignatus |
17,1
<————
|
11,6
—————
|
10,6
—————
|
| 4. Harpalusdistinguendus |
2
—————
|
6,7
——> <——
|
3,2
—————
|
| 5. Poecilusversicolor |
+ |
+ |
7,2 |
| 6. Poeciluscupreus |
+ |
+ |
3 |
| 7. Bembidionlampros |
+ |
+ |
2,9 |
| Всего |
4 |
4 |
7 |
Соотношение доминантов и субдоминантов
|
3
–
1
|
4
–
–
|
4
–
3
|
В первом саду («пар I») 36 видов, причём в центре 34 вида, а на окраине 23 вида, во втором («пар II») 40 видов. Всего в трёх садах было собрано 12 735 экземпляра жужелиц. Численность жужелиц оказалась самой высокой в саду «пар I» центр (7079), а в этом же саду, но на окраине – 1955 экземпляра.
Число видов жужелиц в разные годы не было постоянным: в 1965 году был зафиксирован 31 вид, в 1967 году – 39, а в 1968 – 30. Общих видов за три года исследований оказалось 21, специфичных – 14.
В исследуемых садах наибольшим числом видов представлены роды Amara (9 видов), Pterostichus (8 видов), Harpalus (11 видов), Bembidion (4 вида) и Calathus (4 вида).
Из 46 видов жужелиц массовых выделено всего 7, которые составляют от 93,2% в саду «пар II» до 95,3 % в саду «пар I» на окраине (Табл. 4.).
Ежегодно в садах доминировали 8-9 видов. Общий список доминантов (численность свыше 15%) и субдоминантов (от 5 до 15%), отмеченных за 3 года, включают 11 видов (Табл. 4.). Они составили 92,6% от общего числа отловленных жуков.
В течение всех трёх лет в качестве доминантов или субдоминантов в садах встречались Harpalusrufipes (50%), Amaraingenua (24,5%), Anisodactylussignatus (9,1%), Poecilusversicolor (7,6%), а также Poeciluscupreus, Amaracommunis, доминирующие в отдельных садах (Табл. 4.).
Массовые виды играют основную роль в комплексе жужелиц. Но редкие и единичные виды не менее важны, так как они являются резервами биоценоза и вступают в действие, а иногда становятся массовыми, при резкой смене условий.
Глава
III
.Экологическая структура населения жужелиц плодовых садов
Сочетание в плодовых садах некоторых условий, характерных для полей, с одной стороны, и лесов, с другой, оказывает влияние на формирование фауны жужелиц в них. Фауна жужелиц опытного сада представляет сложный комплекс видов, различных по экологической характеристике.
Экологическая группа – это группа организмов, включающая разные виды, сходно реагирующие на какой-либо фактор среды, имеющий важное морфофизиологическое значение (температура, свет, влажность и т. д.). К сходным условиям организмы могут приспосабливаться по-разному, вырабатывая разную «стратегию» использования ресурсов среды. Поэтому в пределах многих экологических групп можно найти организмы, резко отличающиеся друг от друга по внешнему облику и внутреннему строению.
В карабидологии существуют два подхода к выделению экологических групп: по биотопическому преферендуму (Шарова, 1982) и степени увлажнения биотопов (Булохова, 1985).
3.1. Экологические группы жужелиц по биотопическому преферендуму
Кроме основополагающей работы Шаровой (1982) мы использовали работы и других авторов (Касандрова, 1971; Попова, 1985; Шишова, 1994; Булохова, 1995; Романкина, 1996; Касандрова, Попова, Романкина, 2007).
Комплекс жужелиц опытного сада включает представителей 10 экологических групп по биотопическому преферендуму от лугово-полевой до лугово-болотной. Такое разнообразие экологической структуры отражает разнообразие условий, сложившихся в саду и возможных источниках формирования населения жужелиц. Сочетание в плодовом саду некоторых условий, характерных для леса и поля, создало благоприятную среду для лугово-полевых, луговых, полевых и лесных видов, а также так же гигрофилов. Сходное число экологических групп в плакорных садах отмечает Романкина (1996): 6-9.
В целом в садах ЦГЛ по количеству видов доминирует лугово-полевая группа (43,5%). А по количеству экземпляров – полевая группа (86,3%). Значительна доля видов лесной группы: 13,0% видового обилия. Значение луговой группы резко снижено по видовому обилию 6,5% и особенно по численному – 0,3%.
Ниже приводится характеристика экологических групп жужелиц изученных садов.
Лугово-полевая группа
Жужелицы лугово-полевой группы, составляют основу видов открытых пространств в садах, преобладают по видовому обилию в садах ЦГЛ 43,5%, что характерно для большинства агроценозов. А по численному обилию уступают полевой группе (11,5%)
По данным Романкиной (1996), проводившей исследование в промышленных садах Мичуринского района, жужелицы этой группы преобладали как по видовому, так и по численному (62,5%) обилию в залуженном саду.
По нашим данным эту группу в садах ЦГЛ составляют 20 видов, из них 4 вида – массовые. Они составляют ядро лугово-полевой группы по численному обилию.
Полевая группа
В изучаемых садах по численному обилию первое место занимает полевая группа, включающая 8 видов жужелиц (86,3% численного обилия). Высокой численностью полевая группа обязана доминанту Harpalusrufipes, составившему 59% от численности этой группы.
В садах ЦГЛ за время наблюдений отмечено разное число видов по зонам и изменение их численной доли. Виды полевой группы более концентрировались в центральной зоне сада «пар I», что в 2 раза больше, чем в саду «пар II», где было отмечено по 7 вида (таблица 6). Численное обилие этой группы преобладало в центральной зоне «пар I» (57% от всего количества полевой группы), а наименьшее количество отмечено на окраине сада «пар I» (16,6 % от общего количества экземпляров полевой группы).
Лесная группа
Лесная группа в садах ЦГЛ играет заметную роль. В этой группе встречается достаточно много видов (13,0%), а по численному обилию очень малочисленная (0,3%).
В исследуемых садах большее число видов отмечено в саду «пар II» (6), а наименьшее – 3 – на окраине сада «пар I». По численному составу лидерство держит центральная часть сада «пар I» (42,9% от общего количества экземпляров данной группы). Виды лесной группы не являются в садах массовыми.
Луговая группа
Луговая группа в комплексе жужелиц сада не играет очень заметной роли, как представители лугово-полевой группы. Она сильно уступает другим группам по видовому (6,5%) и по численному обилиям (0,03%).
В садах ЦГЛ за время наблюдений отмечено разное число видов по зонам и изменение их численной доли. Виды луговой группы более концентрировались в саде «пар II». А на окраине сада «пар I» экземпляров совсем не найдено. Численное обилие этой группы преобладало в саду «пар II» (75% от всего количества полевой группы).
В луговой группе не отмечено ни одного массового вида.
Степная группа
Степная группа в садах ЦГЛ представлена тремя видами: Harpaluscalceatus, Harpalusfroelichi, Harpaluszabroides. Количество экземпляров 22. По видовому разнообразию у этой группы (как и у луговой) 6,5% от общего количества видов, встречающихся в изучаемых садах. А по количеству экземпляров – 0,17% от общего числа. Массовых видов в степной группе нет. 91% экземпляров пойманы в центральной части сада «пар I», а вот на окраине этого же сада в ловушки не попал не один жук этой группы.
Лесо-болотная группа
Лесо-болотная группа включает 3 вида (Pterostichusnigrita, Stomispumicatum и Synuchusvivalis). Видовое и численное обилие этой группы составило 6,5% и 0,12%, соответственно. Виды этой группы были отмечены во всех садах и относятся к редким (5-6 экземпляров).
Болотная группа
Болотная группа представлена только одним видом – Bembidionbiguttatum и одним экземпляром, попавшем в ловушку в саду «пар II». Эта группа самая малочисленная по числу видов и экземпляров: 2,2% и 0,008% соответственно.
Лугово-болотная группа
Как и болотная группа, лугово-болотная представлена только одним видом – Clivinafossor. Этот вид многочисленный – 200 экземпляров (1,6%). Встречается во всех садах, но большее число жуков поймано в центральной части сада «пар I» (61% от общего числа жуков этой группы). Присутствие в ловушках жужелиц этой группы обусловлено нахождением на территории ЦГЛ небольших водоёмов и р. Лесной Воронеж.
3.2. Экологические группы жужелиц по степени увлажнения предпочитаемых биотопов
Дефицит влаги – одна из наиболее существенных особенностей наземно-воздушной среды жизни. Протекание всех биохимических процессов в клетках и в организме в целом возможны только при достаточном обеспечении его водой. Вся эволюция наземных организмов шла под знаком приспособления к добыванию и сохранению влаги. Водный обмен со средой у гигрофильных видов более интенсивен, чем у мезофильных и тем более ксерофильных видов. Режимы влажности среды на суше очень разнообразны. Велика суточная и сезонная изменчивость содержания водяных паров в атмосфере. Обеспечение водой наземных организмов зависит также от режима выпадения осадков, наличия водоёмов, запаса почвенной влаги, близости грунтовых вод.
Влажность является ограничивающим фактором среды, определяющим географический ареал вида и лимитирующим его численность. В соответствии с этим, И.Х. Шарова, Н.А. Булохова (1995) предложили на примере карабидокомплексов лугов Брянской области сгруппировать виды жужелиц с разным биотопическим преферендумом в 4 экологические группы по степени увлажнения предпочитаемых биотопов:
1) I группа – гигрофилы (береговые и болотные виды);
2) II группа – мезогигрофилы (лугово-болотные и лесо-болотные виды);
3) III группа – мезофилы закрытых ландшафтов (лесные виды);
4) IV группа – мезофилы открытых пространств (лугово-полевые, полевые, луговые виды).
Так как, потребности во влаге (воздуха или почвы) у представителей различных групп далеко не одинакова, разделение насекомых на эти группы носит более или менее относительный характер.
Доминируют в комплексе жужелиц изучаемых садов мезофиллы открытых пространств по числу видов (32), видовому (69,7%) и численному (97,8%) обилию. По обилию они составили намного больше половины от числа зафиксированных видов и собранных жуков. Заметна роль мезофиллов закрытых биотопов, представленных лесной группой. Эти две экологические группы были зафиксированы во всех сада. Виды остальных групп локализовались в двух зонах сада или одной. К первым относятся мезоксерофилы и мезогигрофилы. Особенностью их распределения была приуроченность к центральной зоне сада, в которой отмечены обе группы. Виды гигрофильной группы были обнаружены только в центральной зоне.
Ниже приводится характеристика экологических групп по степени увлажнения предпочитаемых биотопов в саду.
Мезофилы открытых пространств
Преобладающей группой как по видовому (69,7%), так и по численному (97,8%) обилию являются мезофилы открытых пространств. Эта тенденция прослеживается во всех зонах садов. Число видов варьировало от 3 до 20, видовое обилие от 6,5% до 43,5%, а численное от 0,03 до 86,3%. Наибольшее видовое и численное обилие встречалось в центральной части сада «пар I».
Доминирование видов открытых пространств характерно для агроценозов.
Мезофилы закрытых ландшафтов
Мезофилы закрытых пространств занимают второе место по видовому (13,0%) и одно из последних мест по численному (0,27%) обилию. Видовое обилие этой группы варьировало по садам от 6,5% до 13,0%. Так как в эту экологическую группу входит только одна группа по биотолическому преферендуму – лесная, то она и определяет динамику мезофиллов закрытых биотопов садов – концентрация видов данной группы наблюдается в саду «пар II» (40% численного обилия) и центральной части сада «пар I» (43%) и резкое снижение численного обилия в окраинной части сада «пар I» (17%). Доминирующей по видовому обилию является сад «пар II» (100%), а по численному – центральна часть сада «пар I» (43%).
Мезогигрофилы
Группа включает четыре немногочисленных вида: Clivinafossor, Pterostichusnigrita, Stomispumicatum, Synuchusvivalis. Видовое обилие составило 8,7%, а численное – 1,7%. Эта группа является общей для двух садов и видовое обилие одинаково – по 3 вида в каждой зоне. А численное обилие их не одинаково – 59% от численности мезогигрофилов сосредоточено в центральной части сада «пар I», а на окраине этого же сада только 15%.
Гигрофилы
Группа немногочисленна и включает только один вид: Bembidionbiguttatum. Единственный экземпляр был пойман в саду «пар II». Видовое обилие гигрофилов составило 2,2%, численное обилие – 0,008%. Эта группа специфична и была зарегистрирована только в одном из садов.
Заключение
В результате трёхлетнего (1965, 1967, 1968 гг.) изучения населения жужелиц и его динамики в садах ЦГЛ было выявлено богатое биоразнообразие: 46 видов из 15 родов, 8 экологических групп по биотопическому преферендуму и 4 по степени увлажнения предпочитаемых биотопов. Доминируют мезофилы открытых пространств, что характерно для агроценозов. Особенностью агроценоза садов является значительная доля мезофиллов закрытых биотопов и немногочисленность гигрофилов. Эти характеристики свидетельствуют о большой устойчивости агроценоза садов.
Особенностью родового состава является большое видовое разнообразие родов: Harpalus – 11, Amara – 9, что характерно для агроценозов лесостепи. Доминирующим по видовому и численному обилию является род Harpalus 23,9% и 56,14%, соответственно. Второе место по видовому и численному обилию занимает род Amara (19,6 % и 23,28% соответственно).
Массовых видов жужелиц выявлено 7, что свидетельствует о полидоминантности. Они составляют видовое ядро населения жужелиц садов. На их долю приходится более 90% численного обилия. Harpalusrufipes является доминирующим во всех зонах сада, имеет наибольший индекс доминирования и относится к полевой группе.
Видов немногочисленных и редких – 9. Численное обилие этих видов колеблется от 0,01% до 0,1%. Несмотря на низкую численность, эти виды являются резервом биоценоза, который используется при смене экологических условий.
Зоны сада различаются по числу видов и составу специфичных видов. Наибольшее число видов в центральной зоне сада «пар I» (24). Все сады имеют общие виды. В число специфичных видов центральной части сада «пар I» вошёл вид, который занесён в Красную книгу Тамбовской области (Cilinderagermanica).
Видовой состав и массовые виды жужелиц динамичны во времени. Для массовых видов характерна относительная или динамичная стабильность. Структура доминирования в целом имеет тенденцию к полидоминантности.
По биотопическому преферендуму выделено 8 экологических групп. Среди них отмечены группы с противоположными экологическими требованиями (полевая и лесная, степная, болотная и степно-полевая). Из 8 групп наиболее многочисленными являются три группы (лугово-полевая, полевая и лесная). Лугово-полевая группа доминирует по видовому обилию 43,5%, полевая – по численному (86,3%). Второе место по видовому обилию (13,0%) занимает лесная группа. Распределение экологических групп по зонам сада находится в соответствии с микроклиматическими условиями и экологическими требованиями видов жужелиц. В центральной зоне наиболее типичные и благоприятные условия для большинства видов и всех экологических групп по биотопическому преферендуму.
По степени увлажнения предпочитаемых биотопов выделено 4 экологических группы. В экологическом спектре доминируют мезофилы открытых пространств, к которым относятся лугово-полевые, полевые виды, что характерно для агроценозов лесостепной зоны. Особенностью изучаемых садов является то, что в экологический спектр входят группы, объединяющие гигрофильные виды: мезогигрофилы и гигрофилы.
Экологическая структура динамична во времени. Анализ погодичной динамики экологической структуры показал, что видовое и численное обилие зависило от погодных условий.
Таким образом, разнообразие условий, сложившихся в садах ЦГЛ, обусловило разнообразие видовой и экологической структуры населения жужелиц, имеются условия для поддержания устойчивости агроэкосистемы сада.
Библиографический список
1. Богач Я. Экологические факторы формирования почвенной мезофауны агроценозов в зонах широколиственных лесов и лесостепи: Автореф. дисс. канд. биол. наук. - М, 1983. - С.25 – 30.
2. Большой энциклопедический словарь / под. ред. К.Л. Мазохина-Поршнякова - М., 2001. - С. 294.
3. Видовой состав жужелиц (Coleoptera, Carabidae) Тамбовской области: препринт / Л.И.Касандрова, А.А. Попова, М.Ю.Романкина, Т.В.Шаламова, М.И.Шишова. - Мичуринск: МГПИ, 2007. – 44 с.
4. География Тамбовской области: учеб.пособие / под. ред. М.К. Снытко - Воронеж: Центрально-Чернозёмн. кн. изд-во; 1985. - 93 с.
5. Душенков В.М. Фауна и экология жужелиц (Coleoptera, Carabidae) пахотных земель Центрального Нечерноземья: Автореф. дис. к.б.н. М.: 1983 - С. 14.
6. Касандрова Л.И. Распределение и динамика численности жужелиц в плодовых садах: Автореф. дис. канд. биол. наук. - М., 1970. - 21 с.
7. Касандрова Л.И. К изучению комплекса жужелиц (Coleoptera, Carabidae) лесостепи Тамбовской области // VII Междунар. симп. по энтомофауне Ср. Европы. - Л., 1977. - С. 45-46.
8. Касандрова Л.И. К изучению комплекса жужелиц плодового питомника // Проблемы почвенной зоологии, - Минск, 1978. - с. 109-110.
9. Касандрова Л.И. Жужелицы (Coleoptera, Carabidae) плодового питомника // Экология животных и фаунистика. - Тюмень, 1981. № 81. - С. 52-61.
10. Касандрова Л.И., Хмырова Т. Жужелицы на полях пропашных культур. Формирование животного и микробного населения агробиоценозов. Тез. докл. Всес. совещания, - М., 1982. - С. 50-51.
11. Касандрова Л.И. Жужелицы в агроценозе плодового сада лесостепи // Проблемы интенсификации садоводства: тезисы докладов областной научной конференции. - Тамбов, 1989. - С. 155-156.
12. Касандрова Л.И. Особенности структуры комплекса жужелиц в агроценозе клевера // Экология животных. - Мичуринск. 1997. С. 23-33.
13. Касандрова Л.И. Видовой состав жужелиц (Caleoptera, Carabidae) Тамбовской области: препринт / Л.И. Касандрова, А.А. Попова, М.Ю. Романкина, Т.В. Шаламова, М.И. Шишова. – Мичуринск: МГПИ, 2007. – 44 с.
14. Крыжановский О.Л. Семейство Carabidae - жужелицы // Определитель насекомых Европейской части СССР. - Л., 1965. - 4.2. - С. 29-77.
15. Попова А.А. Сезонная динамика численности доминантных видов жужелиц на полях овощного севооборота ЦЧхоза «Роща» Мичуринского района Тамб. обл. - В. кн.: Новые проблемы зоологической науки и их отражение в вузовском преподавании: Ч. 1. - Ставрополь, 1979, С. 140-141.
16. Попова А.А. Сезонная динамика численности доминантных видов жужелиц на полях овощного севооборота. // В.кн.: Проблемы почвенной зоологии.: Тез. докл. VII Всесоюз. совещ. - Киев, 1981. С. 168-169.
17. Попова А.А. Жужелицы в овощном севообороте. // В.кн. Антропогенное воздействие на фауну почв. - М., 1982 а, С. 96-102.
18. Попова А.А. Доминантные виды жужелиц в овощном севообороте. // В. кн. Формирование животного и микробного агроценозов: Тез. докл. Всесоюз. совещ., М., 1982 б, С. 52-53.
19. Романкина М.Ю. Особенности фауны жужелиц молодых и многолетних яблоневых садов Тамбовской области в условиях лесостепи // Фундаментальная и методическая подготовка специалиста по экологии и охране природы. - Орёл, 1994. - ч. 1 - С. 155.
20. Романкина М.Ю. Сезонная динамика активности жужелиц в яблоневых садах // Эколого-фаунистические исследования в Центральном
Черноземье и сопредельных территориях. - Липецк, 2000. - С. 92-93.
21. Романкина М.Ю. Население жужелиц (Coleoptera, Carabidae) в яблоневых садах и прилегающих агроландшафтов северной лесостепи России:
монография. - М.: Мичуринск, 2001. - 162 с. - Соавт. И.Х. Шарова.
22. Шарова И.Х. Характеристика основных комплексов жужелиц (Carabidae) Московской области // Проблемы почв. зоол. - Казань, 1969. - С. 195-196.
23. Шарова И.Х. Жизненные формы жужелиц (Coleoptera, Carabidae ).- М., 1981 а.- С. 360.
24. Шарова И.Х., Булохова Н.А. Динамика экологической структуры населения жужелиц (Coleoptera, Carabidae) в луговых сообществах под влиянием антропогенных факторов // Фауна и экология жужелиц лугов на юго-западе России. - Брянск, 1995. - С. 38-44.
25. Шарова И.Х., Попова А.А. Зависимость сезонной динамики активности жужелиц (Coleoptera, Carabidae) в агроценозах от смены культур и почвенных условий в лесостепной зоне // Структура и динамика популяций почвенных и наземных беспозвоночных животных. - М., 1990. - 4.1. - С. 12-20.
26. Шарова И.Х., Романкина М.Ю. Видовой состав и структура населения жужелиц в агроценозах Центрального Черноземья // Экология животных. — Мичуринск, 1997. - С. 4-19.
27. Шарова И.Х., Романкина М.Ю. Население жужелиц (Coleoptera, Carabidae) в яблоневых садах и прилегающих агроландшафтах северной лесостепи России. - М. -Мичуринск, 2001. - 161 с.
28. Шишова М.И. Динамика структуры населения и популяции массовых видов жужелиц (Coleoptera, Carabidae) в лесонасождениях северной лесостепи России: Автореф. дис. канд. наук. - М., 1994. - 16 с.
29. Яценко В.Н. Биотехнические мероприятия по защите садов от вредителей как одно из направлений экологизации садов./Яценко, Скрылёва,
Касандрова и др. - 2005. - 25 с.
30. Kryzhanovskij O.L. et altera A Checklist of the Ground - Beetles of Russia and Adjacent Lands (Insecta, Coleoptera, Carabidae) Pensoft Publishers, sofm Moscow, 1995.-C. 5-270.
|
