Содержание
Транспорт субстратов и продуктов
Механизмы клеточной проницаемости
Организация транспортных систем
Способы сопряжения транспорта с энергией метаболизма
Регуляция транспортных процессов
Транспорт веществ из клетки в среду: секреция и экскреция
С клеточной мембраной связан целый ряд важнейших метаболических процессов. Вот главные из них:
репликация ДНК;
биосинтез белков, липидов, компонентов клеточной стенки;
дыхание, фотосинтез;
клеточное деление;
мембранный транспорт, который и будет предметом рассмотрения в данной главе.
Мембранным транспортом будем называть транслокацию веществ через биологические мембраны с обязательным участием молекул-посредников: "подвижных переносчиков" или "каналообразующих" компонентов.
Следует различать пассивное проникновение веществ через мембрану без участия посредников и активное проникновение веществ через мембрану с участием посредников - собственно транспорт.
Пассивная проницаемость мембраны - это проникновение через нее веществ за счет теплового движения молекул. Конечным итогом такого процесса является уравнивание внеклеточной и внутриклеточной концентраций вещества. Начальная скорость физической диффузии зависит от внешней концентрации вещества, а изменение температуры мало влияет на скорость процесса.
Для большинства гидрофильных природных субстратов коэффициент диффузии через двойной липидный слой мембраны имеет очень низкую величину, поэтому скорость их диффузии недостаточна для обеспечения нормального протекания метаболических процессов.
За счет физической диффузии осуществляется проникновение в клетки молекул воды, некоторых газов, а также углеводородов и гидрофобных ксенобиотиков.
В некоторых случаях истинного транспорта так же, как и при физической диффузии, происходит лишь уравнивание внешней и внутренней концентраций вещества. Такие процессы носят название облегченной диффузии. Они осуществляются с участием белков, и скорость их достаточно велика. Типичным примером является проникновение веществ через наружную мембрану грамотрицательных бактерий с участием беяков-поринов.
Значительно чаще транспорт приводит к заметному концентрированию транспортируемых веществ в клетке, так что >> So
. Такой транспорт представляет собой термодинамическую работу и требует затраты энергии. Его называют концентрирующим или активным транспортом.
Реклама

Особенностью транспортных процессов, в отличие от диффузии, является также их стереоспецифичность, в результате которой близкие по химической структуре вещества конкурируют при транспорте за общий переносчик. Ограниченное количество молекул переносчика в мембране приводит к тому, что зависимость начальной скорости транспорта от концентрации субстрата описывается уравнением гиперболы, формально сходным с уравнением Михаэлиса-Ментен, описывающим ферментативную кинетику с аналогичными параметрами:
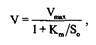
где V - начальная скорость транспорта; V - максимальная скорость при насыщающей концентрации субстрата; K. m
- концентрация субстрата, при которой скорость транспорта равна половине максимальной. В этом случае говорят, что транспортный процесс подчиняется "кинетике насыщения".
Параметры К. П1
и называют параметрами транспортной системы: К. т
характеризует сродство транспортного посредника к субстрату, aVmex
пропорциональна количеству посредника в мембране и скорости его функционирования. Для вычисления этих параметров, как и в ферментативной кинетике, используют графики линейных аппроксимат уравнения Михаэлиса-Ментен.
Одной из первых моделей транслокации субстратов через биологические мембраны была модель "подвижного" переносчика, в которой предполагалось присутствие интегрального мембранного компонента, способного к образованию гидрофобного комплекса с гидрофильным субстратом, экранирующего последний от гидрофобной внутримембранной среды. Предполагалось, что образованный комплекс диффундирует на внутреннюю поверхность мембраны и там освобождает субстрат во внутриклеточную среду. По этому типу действительно осуществляется перенос ионов некоторыми ионофорами. Однако подобный механизм, как правило, не обеспечивает концентрирование субстрата в клетке. Вторая модель предполагает наличие в мембране гидрофильного канала, через который могут проникать субстраты. В отличие от малоспецифичных каналов, образуемых поринами, он должен обладать высокой специфичностью за счет "эстафетной" передачи субстрата от одного центра связывания к другому. Такой канал может стать асимметричным и обеспечить концентрирование субстрата в клетке.
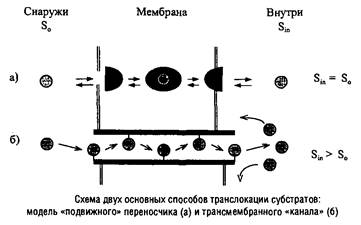
Реальные транспортные системы часто включают более одного белкового компонента, а интегральные мембранные белки-"переносчики" многократно пересекают мембрану, образуя в ней сложную гидрофильную структуру. Молекулярные механизмы транслокации субстрата через такие структуры остаются до конца не расшифрованными.
Реклама

По типу молекулярной организации транспортные системы можно разделить на два больших класса.
Транспортные системы, включающие периплазматические связывающие белки, которые обеспечивают "узнавание" и "доставку" субстрата к мембранному переносчику. Такие системы чувствительны к осмотическому шоку и зависят от энергии АТР. К ним относятся системы транспорта некоторых аминокислот, Сахаров, неорганических катионов.
Транспортные системы, включающие только интегральные мембранные компоненты. Такие системы, как правило, осуществляют одновременный перенос субстрата и одновалентных неорганических катионов и зависят от энергии ТЭП. К ним относятся системы транспорта большинства аминокислот, Сахаров, органических кислот и др.
Для концентрирования веществ внутри клеток необходимо превращение равновесного процесса "облегченной" диффузии в одновекторный процесс "активного" транспорта. Для этого необходима затрата энергии, т.е. создание своего рода "энергетического привода" для транспорта.
Сопряжение транслокации субстрата с энергией метаболизма осуществляется двумя основными путями.
Энергия может затрачиваться на такую химическую модификацию субстрата, которая делает его неспособным взаимодействовать с переносчиком на внутренней поверхности мембраны, а также проникать через мембрану чисто диффузионным путем, что предотвращает его "утечку" из клетки.
Энергия может затрачиваться на такую модификацию переносчика, которая делает его неспособным взаимодействовать с субстратом на внутренней поверхности мембраны, что также предотвращает "утечку" субстрата из клетки.
Системы первого типа фактически осуществляют первый этапов метаболизма этих субстратов и поэтому называются системами векторного метаболизма или реакциями переноса радикалов. К ним, например, относится фосфотрансфе-разная система транспорта Сахаров и сахароспиртов, называемая также системой векторного фосфорилирования, и некоторые другие системы. Фосфотрансферазная система опосредует следующую цепь реакций.

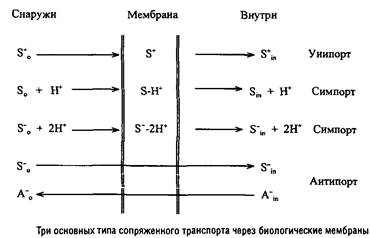
Системы второго типа, в свою очередь, подразделяются на системы "первичного" активного транспорта, генерирующие ТЭП и системы "вторичного" активного транспорта, использующие ТЭП для транспорта органических и неорганических субстратов. В некоторых случаях, например в системах со "связывающими" белками, энергия АТР непосредственно используется в транспорте субстратов. Системы "вторичного" активного транспорта распространены более широко и могут функционировать в соответствии с тремя основными механизмами.
Катионы транслоцируются в клетку по градиенту электрического потенциала путем своеобразного электрофореза.
Незаряженные соединения транслоцируются в клетку совместно с катионами Н+
или Na+
.
Анионы также могут транслоцироваться в клетку путем сим-порта, присоединяя такое количество катионов, которого достаточно для перевода комплекса субстрата с переносчиком в положительно заряженную форму. Кроме того, анионы внешней среды могут обмениваться на внутриклеточные анионы. По механизму антипорта могут транслоцироваться и катионы, например у прокариот широко распространена система антипорта Н+
и Na+
, а у эукариот - система антипорта К+
и Na+
.
Как и регуляция процессов внутриклеточного метаболизма, она осуществляется на двух уровнях: на уровне биосинтеза белковых посредников и на уровне функционирования готовых посредников.
Основными механизмами регуляции биосинтеза переносчиков транспортных систем являются индукция, репрессия и катаболитная репрессия.
Как и в случае ферментов, по типу индукции и катаболитной репрессии регулируется биосинтез компонентов тех транспортных систем, субстраты которых участвуют в процессах катаболизма. По типу репрессии избытком субстрата регулируется главным образом биосинтез аминокислотных транспортных систем.
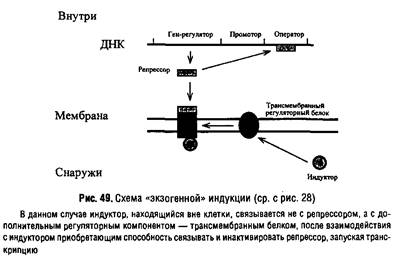
Особенность регуляции некоторых транспортных процессов состоит в том, что индукция осуществляется не внутриклеточным субстратом, а внеклеточным субстратом.
Такая индукция называется экзогенной и требует наличия промежуточного регуляторного интегрального мембранного белка, передающего сигнал индуктора на репрессор.
Подобный тип индукции характерен, например, для транспортной системы гексозофосфатов, фосфоглицерата, некоторых трикарбоновых кислот, а также компонентов фосфотрансферазной системы.
Картина регуляции осложняется тем, что у многих организмов для одного и того же субстрата часто используется несколько транспортных систем, отличающихся по специфичности и величине кинетических параметров. Существуют системы с узкой специфичностью, предназначенные только для одного или небольшого числа сходных субстратов, и с широкой специфичностью. Например, у Escherichiacoli существуют четыре системы для транспорта ароматических аминокислот: три из них специфичны только для одной из этих аминокислот, а четвертая является общей для всех.
Регуляция активности белковых посредников транспортных систем может осуществляться способом обратимой ковалентной модификации или путем нековалентного взаимодействия с эффекторами. В последнем случае, если эффектор взаимодействует с транспортной системой, находясь на той же стороне мембраны, что и субстрат, говорят о цис-регуляции. Например, отрицательная цис-кооперативность обнаруживается при транспорте пролина у Escherichiacoli: избыток субстрата тормозит свой собственный транспорт из среды. У галобактерий, наряду с обычными четырьмя транспортными системами для ароматических аминокислот, существует высокоспецифичная - для тирозина, обладающая очень высоким сродством к субстрату, активность которой подавляется избытком тирозина по бесконкурентному типу.
Если эффектор взаимодействует с транспортной системой, находясь по разные стороны мембраны относительно субстрата, говорят о транс-регуляции транспорта. Например, некоторые аминокислоты, в частности ароматические, находясь внутри клетки, тормозят свой собственный транспорт из среды.
События, связанные с регуляцией транспортных процессов, иногда оказывают существенное влияние на процессы метаболизма в целом. Ярким примером является участие фосфотрансферазной системы в регуляции биосинтеза белков по типу катаболитной репрессии. Оказалось, что уровень сАМР у Escherichiacoli облигатно зависит от функционирования фосфотрансферазной системы, причем главную роль в этой связи играет специфический для глюкозы компонент Е 111.
В отсутствие глюкозы все компоненты системы, в том числе и Е I 11, находятся в фосфорилированном состоянии за счет резерва PEP. Фосфорилированный Е 111, взаимодействуя с аденилатциклазой, переводит ее в активное состояние, в результате чего внутриклеточный уровень сАМР повышается и активируется транскрипция "слабых" оперонов, в том числе систем транспорта и метаболизма других Сахаров.
Напротив, в присутствии глюкозы степень фосфорилирования Е I 11 снижается в связи с переносом фосфорильного остатка на глюкозу в процессе ее транспорта. В результате уменьшается активность аденилатциклазы, снижается уровень сАМР и блокируется транскрипция ряда "сахарных" оперонов.
Следует добавить, что нефосфорилированная форма Е II I, по-видимому, может инактивировать транспортные системы других Сахаров, предотвращая поступление последних в клетку, что еще более усиливает катаболитную репрессию.
Каков же механизм катаболитной реперессии в случае, когда подавляется синтез ферментов, ответственных за катаболизм самой глюкозы, а в качестве более выгодных в энергетическом смысле субстратов выступают, например, органические кислоты или водород?
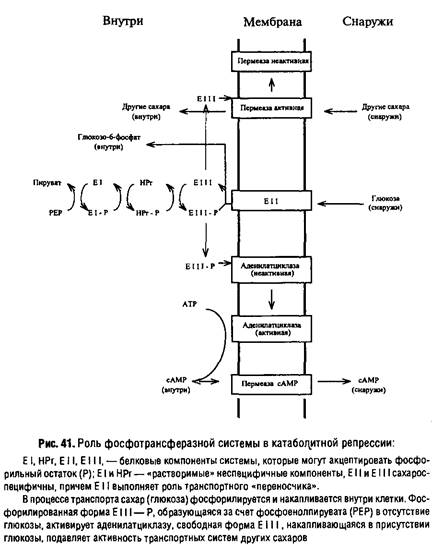
Ведь тогда участие фосфотрансферазной системы невозможно. Чтобы понять механизм явления, необходимо обратить внимание на нижнюю часть рис.41, где изображена система экскреции сАМР. Значение этих систем для регуляции метаболизма мы рассмотрим подробнее в следующем параграфе, а здесь отметим только, что одним из способов снижения уровня сАМР может служить активирование его выброса из клетки, например наложением на мембрану ТЭП, т.е. путем "энергизации" мембраны, степень которой, естественно, будет выше всегда, когда используется более выгодный в энергетическом отношении субстрат. Таким образом, если субстрат обеспечивает энергетические потребности клетки и создает необходимую степень "энергизации" мембраны, он может вызывать подавление использования других субстратов, от которых зависит уровень с AMP в клетке.
Рассмотрим сначала процессы секреции, т.е. выделение из клетки белков.
Секретируемые белки синтезируются в виде более длинных предшественников, которые подвергаются процессингу, как правило, на этапе транслокации через мембрану. Они содержат на NHj-конце так называемый сигнальный пептид из 15-30 аминокислот, которые удаляются специальной сигнальной пептидазой, локализованной в мембране. Транслокация белка через мембрану обычно протекает одновременно с трансляцией, хотя известны случаи посттрансляционной транслокации. После образования сигнального пептида трансляция временно прекращается в результате присоединения к рибосоме нуклеопротеидного ингибитора и полисомный комплекс перемещается к мембране, где локализован аппарат секреции, включающий рецепторы рибосомы и сигнального пептида. Происходит формирование трансмембранной "поры". Сигнальный пептид закрепляется на своем рецепторе, и трансляция возобновляется, причем растущая пептидная цепь "проталкивается" через мембрану. Сигнальный пептид отщепляется сигнальной пептидазой, и "зрелая" молекула фермента отделяется от рибосомы. После завершения трансляции рибосомный комплекс покидает мембрану и диссоциирует на субчастицы для подготовки нового цикла трансляции.
Сходный механизм используется при образовании белков наружной мембраны грамотрицательных бактерий. Такой белок содержит гидрофобную "якорную" последовательность, которая позволяет ему закрепиться в мембране, процесс его секреции часто называют экспортом.
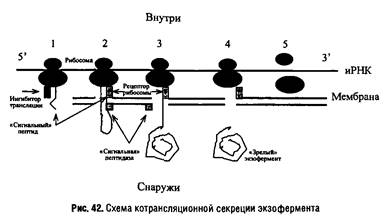
У эукариот решающее значение в процессе секреции ферментов имеют эндоплазматический ретикулум и аппарат Гольджи.
Для описания процесса выделения из клеток низкомолекулярных веществ используют термин экскреция. Она может выполнять три основные физиологические функции:
а) удаление ингибитора - токсичного для клетки вещества, продуцируемого эндогенно или поступающего из окружающей среды;
б) удаление эффектора, избыток которого нарушает нормальные физиологические процессы в клетке;
в) удаление конечного продукта метаболизма с целью "сброса" окислительно-восстановительных эквивалентов, создания в среде резерва источников питания или запасания энергии в виде ТЭП.
Дополнительные функции секреции могут состоять в выделении химических "сигналов", регулирующих физиологическое состояние популяции.
Хорошо изученными примерами экскреции первого типа являются транспортные системы, кодируемые плазмидами и опосредующие энергозависимое удаление антибиотике в-тетрациклинов, а также анионов арсената и катионов Cd2+
из клеток устойчивых к этим ингибиторам микроорганизмов.
Пример экскреции второго типа - так называемый выброс индуктора в системе метаболизма лактозы под действием глюкозы, а также экскреция сАМР в результате повышения ТЭП. Оба эти эффекта представляют собой дополнительные проявления регуляции метаболизма по механизму катаболитной репрессии.
Наконец, примером экскреции третьего типа может служить выделение лактата в процессе молочнокислого брожения, в результате чего происходит "сброс" восстановительных эквивалентов и формирование ТЭП.
Важное физиологическое значение регуляции внутриклеточного уровня низко молекулярных веществ путем их экскреции из клетки позволяет рассматривать эти механизмы как особый уровень регуляции метаболизма, который мы предлагаем называть мембранной регуляцией.
|
