Содержание ДНК в нервных клетках. Проблема "избыточной" ДНК
Содержание
Введение
1. Репликативный синтез ДНК и пролиферация нервных клеток
2. Репарация ДНК в мозге животных
3. Особенности организации хроматина в нервных клетках
4. Рибонуклеиновые кислоты мозга
5. Мозгоспецифическая экспрессия генов
6. Характеристическая последовательность нуклеотидов в РНК мозга
7. Альтернативный процессинг пре-мРНК в мозге
8. Экспрессия генома и онтогенез мозга животных
9. Экспрессия генов в ЦНС беспозвоночных
10. Экспрессия генов в развивающейся нервной системе позвоночных
Выводы
В большинстве областей головного мозга клетки являются диплоидными. У человека, в частности, на обеих парах хромосом каждой клетки содержится около 6 пг ДНК, т.е. около 4-1012 Д или 6 109 пар нуклеотидов - Общая длина молекул ДНК диплоидного набора хромосом клетки человека близка к 1,5 м. В целой клетке ДНК больше, но ненамного, - за счет митохондриальной ДНК. Это превышение достигает нескольких десятков процентов - для средних и больших по размеру нейронов.
В конце 1960-х - начале 1970-х годов в результате многочисленных исследований методом цитофотометрии ДНК по Фельгену получило широкое распространение представление о существовании избыточной по сравнению с диплоидным набором ДНК в клетках мозга млекопитающих и птиц. Особенно популярной была точка зрения о тетраплоидности крупных нейронов - таких, как клетки Пуркинье мозжечка и пирамидные нейроны гиппокампа.
Применение более совершенных методов в последующие годы показало, что большинство "подозреваемых" нейронов в действительности содержит диплоидное количество ДНК, хотя в ограниченных популяциях нейронов определенных типов содержание ДНК может быть более высоким. Наиболее достоверно гипердиплоидизация обнаружена в клетках Пуркинье мозжечка, в небольшой части которых выявлено избирательное умножение генов рибосомальной РНК. Общепринятое почти до недавнего времени представление о накоплении избыточной ДНК в ядрах нейронов неокортекса млекопитающих в первые недели постнатального онтогенеза не подтвердились при исследовании более совершенными методами.
В нейронах некоторых беспозвоночных явление полиплоидизации распространено очень широко: в ЦНС брюхоногих моллюсков, например, полиплоидными являются практически все крупные нейроны.
Реклама

Выше мы обращали внимание на отсутствие качественных отличий ДНК клеток мозга от ДНК других клеток организма, имея в виду одинаковый набор генов, но в то же время глубокие различия в наборе работающих и неработающих генов. В течение последних четырех лет появились данные об особой роли в функциях мозга ряда монотонно повторяющихся тринуклеотидных последовательностей в ДНК. Например, тринуклеотид CAG содержится в различных частях генома группами по 8-33 копий. Увеличение числа копий в 3-10 раз ассоциируется с рядом тяжелых нервных болезней. Аналогичная ситуация установлена для последовательностей CGG, GCC и CTG. Это служит яркой иллюстрацией того, как велико значение точной организации всех элементов ДНК нейронов, даже относительно простых и монотонных.
Формирование нейронольных популяций в головном мозге крыс и мышей в основном завершается к моменту рождения. Исключение составляют популяции зернистых нейронов обонятельных луковиц, зубчатой фасции гиппокампа и коры мозжечка, формирование которых наиболее активно протекает в первые три недели после рождения, а в небольших масштабах, по-видимому, происходит и у взрослых животных. Попытки доказать возможность митотического деления полностью дифференцированных нейронов большинством цитологов признаны неубедительными. Лишь при создании определенных весьма специфических условий invitro возникает репликация ДНК и митозы в мотонейронах из спинного мозга цыплят.
Источником образования нейронов в период активного нейрогенеза служат камбиальные клетки специальных герминативных зон. При этом миграция нейробластов в соответствующие зоны созревающего мозга весьма упорядочена. Так, для неокортекса млекопитающих, имеющего хорошо выраженную микроколончатую организацию, топография микроколонок определяется топографией пролиферативных единиц вентрикулярного слоя. Последние представляют собой морфологически обособленные группы камбиальных клеток, которые, претерпевая ряд клеточных делений, дают начало соответствующей обособленной группе нейронов в определенной зоне неокортекса, т.е. микроколонке.
В мозге взрослых теплокровных животных нейрогенез чрезвычайно ограничен. У приматов нейрогенез, по-видимому, полностью заканчивается в первые месяцы после рождения, т.е. задолго до полного функционального созревания мозга.
У беспозвоночных животных формирование нейроналъных популяций ЦНС может продолжаться, по-видимому, на протяжении всей жизни. Так, у аплизии нейрональные популяции всех центральных ганглиев многократно умножаются в первые месяцы постметаморфного онтогенеза, причем выделяется ограниченная во времени стадия позднего ювенильного развития, на которой происходит бурное увеличение численности нейронов одновременно во всех ганглиях. Еще более стремительно при этом возрастает объем нейропиля. Одновременно с этим у агошзии появляется способность к наиболее сложной для нее форме обучения - сенситизации.
Реклама

Формирование популяций клеток глии в головном мозге млекопитающих наиболее активно протекает в первые недели постнаталъного онтогенеза и в ограниченных масштабах продолжается в течение всей жизни. Основным источником глиальных клеток, как и нейронов, служат плюрипотентные клетки герминативных зон. Однако в отличие от нейронов, которые утрачивают способность к митотическому делению до миграции из герминативной зоны, глиальные клетки сохраняют ее и в местах своей будущей конечной дифференцировки.
Действие систем "ремонта", репарации ДНК в любой клетке является необходимым условием нормального функционирования ее генетического аппарата. Этот процесс особенно важен для клеток долгоживущих, медленно обновляющихся популяций, классическим примером которых являются именно клетки нервной системы. Замечено, что прогрессивные нарушения функционирования нервных клеток в стареющем мозге человека и животных коррелируют с постепенным накоплением повреждений в их ДНК. При действии умеренных доз гамма-облучения в нейронах и глиальных клетках наблюдается стимуляция репаративного синтеза ДНК. Вместе с тем в нейронах мозжечка гамма-облучение invivo сопровождается накоплением каких-то нерепарируемых повреждений ДНК, которые в конце концов приводят к деградации ДНК и гибели нейронов. Очевидно, существует определенная пороговая доза повреждений, после которой системы репарации ДНК в клетках уже не способны справиться с их дезорганизующим действием.
Большая часть синтеза ДНК в мозге интактных взрослых крыс обусловлена именно процессами репарации. Скорость репарации многих экспериментально индуцированных повреждений ДНК мозга очень невелика. Относительно быстрое удаление части таких повреждений в первые часы после их индукции обычно сменяется фазой гораздо более медленной репарации. В первую очередь репарируются повреждения транскрипционно активных, важных для выживания и полноценного функционирования клеток генов, тогда как в репрессированных областях хроматина повреждения могут накапливаться. В основе такой избирательности может лежать более высокая доступность транскрибируемых участков хроматина ферментам репарации и совместное расположение транскрипционных и репаративных ферментов в определенных участках ядерного матрикса.
В мозге млекопитающих обнаружены практически все ферменты, необходимые для эффективной репарации повреждений ДНК. Так, в мозге крыс и кроликов имеются основные ферменты, необходимые для синтеза дезоксинуклеозидтрифосфатов, - рибонуклеотидредуктаза и тимидинкиназа. Активность этих ферментов особенно высока в мозге эмбрионов и новорожденных животных. У взрослых животных она сохраняется на довольно низком уровне, соответствующем невысоким потребностям в пополнении фондов дезоксинуклеозидтрифосфатов для репаративного синтеза ДНК. Содержание различных ДНК-полимераз в мозге млекопитающих также зависит от возраста. Функциональное значение различных ДНК-полимераз в клетках эукариот довольно хорошо исследовано.
Главной релликативной ДНК-полимеразой является поли-мераза а. Ведущую роль в репликации ядерной ДНК наряду с а-полимеразой выполняет ДНК-полимераза 5: предполагается, что 5-полимераза осуществляет непрерывный синтез лидирующей цепи ДНК, а а-полимераза - синтез фрагментов Оказаки "запаздывающей" цепи. ДНК-полимераза; небольшой вклад в общую активность вносит ДНК-полимераза у. ДНК-полимераза р осуществляет стимулируемый ультрафиолетовым облучением репаративный синтез ДНК в изолированных ядрах нейронов и является, таким образом, главной, если не единственной, репаративной полимеразой в зрелых нейронах.
ДНК-полимераза р является конститутивным ферментом, синтезируемым в клетках всех типов на более или менее одинаковом уровне, не зависимом от их пролиферативной активности. Это самая часто "ошибающаяся" из всех эукариотических ДНК-полимераз: при копировании ДНК invitro она делает одну ошибку на каждые 103
- 104
нуклеотидов. По-видимому, существуют какие-то дополнительные факторы, повышающие точность ее работы invivo.
В мозге млекопитающих обнаружены также и другие ферменты, принимающие участие в репликативном и репаративном синтезе ДНК. Наиболее подходящей по своим каталитическим свойствам на роль репаративной экзонуклеазы является ДНКаза ВШ, которая, по-видимому, относится к мозгоспецифическим ферментам. В ядрах нейронов и глии мозга взрослых морских свинок выявлена ДНК-лигаза, участвующая в завершающих этапах репаративного синтеза. Непонятной остается функция обнаруженной в мозге человека терминальной дезоксинуклеотидилтрансферазы, способной к нематричному синтезу ДНК. Интересно, что этот широко распространенный у млекопитающих фермент обнаруживается только в клетках тимуса и нервной системы. Высказываются предположения, что его роль связана с уникальной способностью этих клеток запасать и хранить ненаследуемую информацию.
ДНК-топоизомеразы I и II, участвующие в различных матричных процессах регуляции топологической структуры ДНК, обнаружены в ядрах нейронов и глиальных клеток.
В заключение остановимся на гипотезе о так называемой метаболической ДНК.
Предполагается, что, помимо стабильной геномной ДНК, в дифференцированных клетках существует специфическая фракция относительно быстро обменивающейся ДНК, которая, по-видимому, представлена экстракопиями наиболее активных генов. В целом эта концепция до сих пор не получила достаточного обоснования, хотя сообщения о существовании быстро обменивающихся фракций ДНК в тех или иных клетках время от времени появляются в научной литературе. Наиболее убедительны данные о существовании таких ДНК в эмбриональных фибробластах цыпленка, где ей приписывается роль межклеточного переносчика информации.
Вопрос о существовании метаболически лабильной ДНК в клетках мозга животных и ее природе остается спорным. Быстрое включение радиоактивных предшественников в ДНК мозга и последующее быстрое исчезновение их из этой ДНК наблюдали многие исследователи.
Однако имеются и сообщения о высокой стабильности радиоактивной метки, включившейся в ДНК мозга.
В литературе приводятся убедительные аргументы в пользу того, что быстрый обмен вновь синтезированной ДНК является артефактом, связанным с ее радиоактивной деградацией вследствие включения меченых предшественников.
Полагают также, что метаболическая нестабильность этой ДНК обусловлена быстрой гибелью значительной части активно делящихся клеток. Тем не менее некоторые исследователи считают, что метаболически лабильная ДНК в клетках мозга действительно существует и может выполнять какие-то особые, хотя пока и не выясненные функции.
Недавно стабильность ДНК в клетках коры мозжечка человека исследовали по соотношению в ней природных изотопов углерода. Оказалось, что основная масса ДНК в этих клетках чрезвычайно стабильна.
Ясно, что если метаболическая ДНК в мозге человека и существует, она составляет ничтожно малую от суммарной ДНК долю.
Хроматин всех эукариот организован в виде серии повторяющихся нуклеопротеидных частиц - нуклеосом. Каждая нуклеосома состоит из так называемого кора, содержащего октамер гистонов Н2А, Н2В, НЗ, Н4 и обернутый вокруг него участок ДНК - 146 н. п., а также линкерной области, ассоциированной с гистоном HI, играющим особую роль в соединении нуклеосом друг с другом и в образовании наднуклеосомньгх уровней организации хроматина.
Нуклеосомная и наднуклеосомная организации, несомненно, играют важную, хотя и во многом еще не выясненную роль в функциональной активности хроматина. Укладка двуспиральной ДНК в нуклеосоме сопровождается сильными искажениями ее вторичной структуры и кардинально изменяет условия ее взаимодействия с различными регуляторными белками. Показано, что наличие нуклеосом в промоторной области генов препятствует инициации транскрипции РНК-голимеразой II. В то же время наличие нуклеосом в кодирующей области не препятствует элонгации уже инициированных цепей мРНК. Поэтому начальные реакции транскрипции генов invivo обеспечиваются специальными механизмами, исключающими образование нуклеосом в критических участках промоторной области. В кодирующей же области активно транскрибируемых генов отмечается лишь частичное нарушение нуклеосомной структуры, объясняемое временным вытеснением гистонов движущейся РНК-полимеразой.
Образование локальных наднуклеосомных структур с участием гистона HI, по-видимому, является универсальным самоподдерживающимся механизмом выключения генов. Исследования последних лет показали также, что тканеспецифическая транскрипция генов обеспечивается сложными взаимодействиями различных негативных и позитивных регуляторов, узнающих специфические последовательности в промоторах индивидуальных генов. Характерными особенностями структуры активных участков хроматина являются: частичное или полное нарушение нуклеосомной и наднуклеосомной организации, присутствие особых вариантов гистонов или их постсинтетическая модификация, существование локальных торзионных напряжений в доменах хроматина и избирательная ассоциация с ядерным матриксом.
Преобладающая часть ядерной ДНК мозга организована в типичную нуклеосомную структуру. Однако по сравнению с хроматином печени хроматин мозга более гетерогенен по длине линкерных участков и обладает более разнообразным набором негистоновых белков. Длина нуклеосомных единиц в большинстве эукариотических клеток близка к 200 н. п. Некоторые вариации этой величины связаны с изменчивостью в длине линкерных участков, тогда как с кором нуклеосом всегда ассоциирован фрагмент ДНК длиной 146 н. п. В дифференцированных нейронах неокортекса нуклеосомная ДНК необычно короткая и составляет - 160-162 н. п., а клетки неастропитарной глии неокортекса и нейроны мозжечка имеют обычные по длине нуклеосомы. Переход в нейронах неокортекса к атипично коротким нуклеосомам коррелирует с окончательной дифференцировкой этих нейронов: у кроликов, мышей и крыс он проходит в первую неделю постнатального онтогенеза, а у более зрелорождающихся морских свинок - между 32-м ч 44-м днями эмбриогенеза. Аналогично, в пренатально дифференцирующихся нейронах гипоталамуса крыс короткие нуклеосомы присутствуют уже за два дня до рождения.
Укорочение нуклеосом не всегда сопровождает процесс морфофункционального созревания нейронов. Например, укорочения не наблюдается при созревании постмитотических нейронов неокортекса, изолированных из мозга 16-дневных эмбрионов крыс и культивируемых на селективной среде до возраста, соответствующего второй неделе постнаталъного онтогенеза. В мозжечке созревание зернистых нейронов в первый месяц постнатального онтогенеза сопровождается не уменьшением, а даже увеличением длины нуклеосом. Вообще усредненные данные о длине нуклеосом не следует автоматически переносить на всю сложную популяцию нервных клеток. Так, в мозге крыс процесс укорочения нуклеосом характерен только для нейронов глубоких слоев неокортекса. Существенная разница в длине нуклеосом существует и у различных глиальных клеток.
Функциональное значение описанного перехода к коротким нуклеосомам при созревании нейронов неокортекса до настоящего времени остается невыясненным. Предположение о том, что укороченные нуклеосомы обеспечивают укладку полинуклеосомной цепи в более открытую, способствующую транскрипции наднуклеосомную структуру, является упрощенным.
Исследования этого вопроса на более широком круге объектов подтверждают, что однозначной связи между транскрипционной активностью генов и длиной нуклеосом не существует. Транскрипционная активность хроматина определяется, как уже отмечено выше, большим числом факторов. Возможно, существование коротких нуклеосом в хроматине нейронов лишь каким-то образом облегчает действие этих факторов. В частности, могут изменяться условия взаимодействия линкерных участков хроматина с гистоном HI и негистоновыми белками. В хроматине нейронов неокортекса содержание гистона HI составляет - 0,5 молекулы на 1 нуклеосому, что примерно в два раза ниже, чем его содержание в хроматине глиальных клеток и клеток соматических тканей. Это хорошо согласуется с данными о более высокой доле активного хроматина в мозге по сравнению с соматическими тканями и в нейронах по сравнению с глиальными клетками.
Фракция активного хроматина нейронов практически не содержит гистона HI, а аналогичная фракция хроматина из клеток неастроцитарной глии содержит его в очень малом количестве. Напротив, фракции неактивного хроматина нейронов и глии содержат сравнительно высокие количества HI. Снижение доли активного хроматина в нейронах и глиальных клетках мозга человека при болезни Альцгеймера также коррелирует с повышением содержания гистона HI.
Таким образом, роль гистона HI в образовании наднуклеосомных уровней организации хроматина, а также его избирательное вытеснение из активных участков хроматина в настоящее время уже не вызывают сомнений. Менее изучена роль различных вариантов гистона HI. Известно, что из шести обнаруженных в клетках млекопитающих вариантов гистона HI, HI а и Н1в в значительных количествах обнаруживаются лишь в активно делящихся клетках, Hlc, - d, - е - в делящихся и неделящихся, а гистон Н1° характерен для терминально дифференцированных неделящихся клеток. Прекращение пролиферативной активности нейронов коррелирует с накоплением гистона Hie, а терминальная дифференцировка нейронов - с накоплением HI°.
Коровые гистоны в хроматине нейронов неокортекса представлены большим числом вариантов, для многих из которых обнаружены формы, конъюгированные с убиквитином. Одна из форм гистона Н2В, по-видимому, является мозгоспецифической. Высокое содержание соединенных с убиквитином форм гистонов в хроматине обычно связывают с высокой матричной активностью.
Хроматин нервных клеток является динамичным, активным образованием, состояние которого изменяется в ходе нормального развития и при действии различных внешних и внутренних стимулов. В первые недели постнатального онтогенеза в хроматине нейронов неокортекса крыс происходит заметное уменьшение числа участков с повышенной чувствительностью к ДНКазе в мозге стареющих животных эти изменения еще более выражены, что свидетельствует об уменьшении матричной активности хроматина в ходе старения.
Одним из механизмов регуляции матричной активности хроматина является обратимое ацетилирование гистонов. В ядрах нейронов ацетилирование гистонов более выражено по сравнению с глиальными клетками. Это коррелирует с более высокой активностью эндогенных РНК-полимераз нейронов и с большим числом активных участков инициации транскрипции. С другой стороны, в первые дни постнатального онтогенеза степень ацетилирования гистонов в неокортексе крыс быстро уменьшается.
Обнаружено, что конечная дифференцировка нейронов неокортекса и мозжечка крыс в первые недели постнатального онтогенеза сопровождается изменениями в наборах негистоновых белков хроматина, наиболее примечательным из которых можно считать появление белков - J5 и - 38 кД, имеющих специфическое сродство к однонитчатой ДНК. Свое влияние на матричные процессы они оказывают, способствуя локальному расплетению двойной спирали ДНК.
В ядрах нейронов неокортекса обнаружены также белки, имеющие сродство к специфической левоспиральной форме ДНК - Z-ДНК. В последние годы такие Z-ДНК-связывающие белки привлекают большое внимание как возможные регуляторы транскрипции и других генетических процессов.
При терминальной дифферениировке нейронов происходят также изменения в наборах особой фракции негистоновых белков, прочно связанных с ДНК, некоторые из которых являются мозгоспецифическими.
Важную роль в регуляции транскрипции генов могут играть, наконец, процессы фосфорилирования, метилирования и, возможно, другие посттрансляционные модификации негистоновых белков хроматина. Заметные изменения в наборах фосфорилируемых и метилируемых негистоновых белков наблюдаются в ядрах нейронов и глиальных клеток неокортекса крыс в первые недели постнатального онтогенеза. При этом главное различие между ядрами нейронов и глиальных клеток состоит в большем мешлировании группы специфических для нейронов негистоновых белков - 100 кД. В ядрах нейронов и глии мозга мышей обнаружены протеинкиназная и метилазная активности, значительная часть которых прочно ассоциирована с хроматином.
Протеинкиназа осуществляет цАМФ-независимое фосфорилирование белков хроматина. Ее активность в ядрах нейронов значительно выше, чем в ядрах глиальных клеток. При действии ряда нейромедиаторов на нейроны мозга крыс наблюдается фосфорилирование ядерных белков и стимуляция синтеза РНК. Фосфорилирование части негистоновых белков индуцируется в клетках верхнего шейного ганглия при действии фактора роста нервов. В хромаффинных клетках надпочечников фосфорилирование негистоновых белков хроматина пАМФ-зависимой протеинкиназой является центральным звеном в транссинаптической регуляции синтеза тирозин-3-мо-нооксигеназы ацетилхолином. Показано, что фосфорилирование негистоновых белков хроматина повышается при выработке оборонительных условных рефлексов.
Общее содержание РНК в большинстве нервных клеток очень велико. Среднее отношение РНК: ДНК достигает 50 и сравнительно редко бывает ниже 3. Это превышает отношение, характерное для особенно интенсивно метаболирующих клеток секреторных тканей, где оно составляет 2-4,5. В мотонейронах головного мозга и в спинальных ганглиях количество РНК в одной клетке достигает 500-2500 пг. Такое обилие РНК обусловлено главным образом наличием мощного рибосомального белоксинтезирующего аппарата в цитоплазме нейрона. Быстро обменивающаяся мессенджер-РНК, тоже относительно широко представленная в нейронах, занимает в количественном отношении скромное место по сравнению с рибосомальной РНК.
Рибосомальная РНК мозга и аппарат трансляции не имеют принципиальных отличий от других тканей и органов. Поэтому, отметив особую мощность последнего, сосредоточим далее внимание на особенностях синтеза и многообразии мРНК мозга, обусловливающих качественное своеобразие белков и многих других компонентов нервных клеток.
Так же как и во всех дифференцированных клетках и тканях организма, в нейронах и глиальных клетках мозга "работает" лишь часть генов. Это, с одной стороны, гены, ответственные за продукцию белков, необходимых для обеспечения метаболических процессов, более или менее сходных в разных клетках и тканях, а с другой стороны, гены, участвующие в синтезе белков, регулирующих специфические функции данной ткани. Активность остальных генов, не нужных для функций данных клеток, подавлена. Доля активных генов в каждой данной ткани обычно невелика - менее 10% - и неодинакова в разных клетках и тканях. В мозге эта доля выше, чем в других органах, что отражает особую сложность его функций. К сожалению, точные значения этих параметров пока не установлены.
В последние годы проводится систематическое исследование характера экспрессии большого числа индивидуальных генов в мозге млекопитающих. Для этого случайно выбранные из коллекции кДНК клонов мозга крысы последовательности гибридизуются с препаратами поли+РНК из разных тканей в так называемых Нозерн блоттах. По результатам гибридизации можно судить о присутствии соответствующих данному клону последовательностей РНК в исследуемых тканях, их количестве, гетерогенности и т.д. Анализ таким методом 191 случайно выбранного клона позволил авторам разбить все экспрессируемые в мозге поли+РНК на четыре класса: I - "нерегулируемые", т.е. одинаково экспрессируемые во всех исследованных тканях и, скорее всего, кодирующие так называемые белки "домашнего хозяйства", необходимые для жизнедеятельности любой клетки; II - регулируемые, т.е. экспрессируемые в клетках всех трех тканей, но в разной степени; III - мозгоспецифические, т.е. экспрессируемые только в клетках мозга; IV - редкие, т.е. присутствующие в мозге в количестве, недостаточном для строго воспроизводимого обнаружения использованным методом. Если считать мозгоспецифическими РНК III и IV классов, в эту категорию попадает более половины синтезируемых в мозге мРНК, что хорошо согласуется с оценками, полученными с помощью методов суммарной ДНК-РНК-гибридизации.
Важно отметить, что определение нуклеотидной последовательности клонированных мозгоспецифических мРНК позволяет воспроизвести аминокислотную последовательность кодируемого белка.
Такой анализ был осуществлен Сатклиффом, например, для одного из клонов, кодирующих РНК III класса. Этот клон гибридизуется с моз-госпецифической мРНК, присутствующей в разных отделах мозга крысы, но в разном количестве. Кодируемый этой РНК полипептид не содержит участков гомологии с ранее изученными белками.
С помощью антител к синтетическим фрагментам этого полипептида изучена его локализация в мозге. Он обнаружен в аксонах, иннервирующих клетки Пуркинье в мозжечке, пирамидные нейроны поля САЗ гиппокампа; в радиальных волокнах глубоких слоев цингулярной и соматосенсорной коры; в группах волокон свода, стриатума, латерального обонятельного тракта и других структур. Источником этих волокон являются нейроны медиального ядра трапециевидного тела, центральной покрышки моста, вентромедиального и базальноаркуатного ядер гипоталамуса, а также нейроны, разбросанные по другим структурам.
Эту работу Сатклиффа и соавторов можно считать образцом технически выполнимой и универсальной методологии поиска новых мозгоспецифических мРНК и исследования первичной структуры, мест синтеза и функциональной роли кодируемых ими мозгоспецифических белков.
Особый интерес в этом отношении представляют мРНК III и IV классов, имеющие ограниченную локализацию внутри мозга и его отдельных структур. Судя по содержанию наиболее редких мозгоспецифических мРНК, они должны транскрибироваться в ограниченных популяциях нервных клеток и, возможно, связаны с узкой функциональной специализацией этих популяций.
Для избирательного клонирования таких РНК в последние годы используют процедуру "вычитания", суть которой заключается в удалении из суммарных препаратов кДНК последовательностей, общих для разных отделов мозга, и последующем клонировании такого "вычтенного" препарата кДНК. Так, уже клонированы мРНК, имеющие ограниченное распространение в неокортексе обезьян, но их изучение еще только начинается.
В лаборатории J.С. Venter начато составление всеобъемлющего каталога генов, экспрессирующихся в мозге человека. Для этого авторы, осуществляют крупномасштабное сиквенирование клонов из библиотек кДНК мозга человека и его частей. К настоящему времени они получили частичные последовательности для примерно 3000 клонов, большинство из которых являются ранее не известными. Ближайшей задачей является картирование соответствующих генов в хромосомах человека и использование полученных данных для идентификации генов, ответственных за различные наследственные заболевания.
Несколько мозгоспецифических клонов из коллекции Сатк-лиффа и соавторов проявляли необычный характер гибридизации с препаратами поли+РНК, узнавая в них общую короткую мозгоспецифическую РНК. Эта малая РНК найдена во всех отделах мозга крыс, хотя и в неодинаковых количествах. Анализ нуклеотидной последовательности нескольких клонов показал, что единственным общим для них элементом является консервативная 82-нуклеотидная последовательность, которая и ответственна за гибридизацию с ВС1 РНК. Инвариантными в этой последовательности являются 51 нуклеотид] еще 24 нуклеотида одинаковы в 90% проанализированных клонов. Справа эта последовательность соседствует с олигонуклеотидами, содержащими в основном остатки аденина. Остальные примыкающие или близкие к ней последовательности в разных клонах совершенно различны. Присутствие одной и той же или сходной нуклеотидной последовательности в разных мозгоспецифических РНК как бы маркирует их, что и послужило основанием для ее обозначения как характеристической seguence"). Эта последовательность присутствует в некодирующих областях многих генов и соответствующих пре-мРНК и в большинстве случаев удаляется при процессинге последних. Ее функция до сих пор неизвестна, хотя есть основания полагать, что она служит неспецифическим стимулятором транскрипции содержащих ее генов. В любом случае ID-последовательность не является нейроспецифическим регулятором транскрипции, поскольку она присутствует и в многих пре-мРНК соматических тканей.

В препаратах цитоплазматической РНК мозга также обнаружены короткие молекулы РНК, содержащие ID-последовательность: ВС1, ВС2 и ТЗ. Синтез этих малых РНК осуществляется РНК-полимеразой III, узнающей консервативные участки в ID-последовательности, гомологичные внутреннему промотору генов РНК-полимеразы III. Обнаруженные в клетках эукариот малые РНК, как показано в последние годы, играют важнейшую роль в таких процессах, как сплайсинг и З'-процессинг пре-мРНК, трансляция и трансмембранный транспорт секретируемых белков и т.п.
Альтернативный процессинг заключается в образовании различных зрелых мРНК из одного первичного транскрипта в результате соединения различных комбинаций экзонов и/или использования различных сигналов полиаденилирования. Одним из наиболее хорошо изученных случаев использования альтернативного сплайсинга для образования различных продуктов в разных клетках является система синтеза регуляторных пептидов, кодируемых геном кальцитонина. Этот ген кодирует небольшое семейство пептидов, два из которых (кальцитонин и катакальцин) преимущественно образуются в клетках щитовидной железы, а третий - в клетках нервной системы.
Исследование структуры гена кальцитонина и зрелых мРНК, кодирующих эти пептиды у крысы и человека, показало, что I-III экзоны являются общими для этих мРНК. IV экзон, кодирующий кальцитонин и катакальцин, присутствует только в мРНК кальцитонина и содержит специфический для нее сигнал полиаденилирования, а V и VI экзоны, содержащие CGRP-кодирующую и 3'-некодирующую последовательности, включая специфический для CGRP-мРНК сигнал полиаденилирования, присутствуют только в CGRP-мРНК. Механизмы выбора различных путей сплайсинга первичного транскрипта CT/CGRP-гена в клетках щитовидной железы и нейрона пока еще остаются неясными и сейчас интенсивно изучаются.
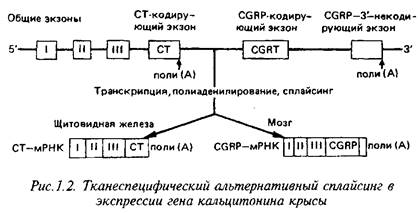
Альтернативный сплайсинг первичных транскриптов обеспечивает также разнообразие белков миелина в ЦНС млекопитающих. В частности, показано, что различные основные белки миелина у мыши кодируются одним геном shi-локуса: при этом белок 21,5 кД кодируется мРНК, содержащей последовательность всех 7 экзонов гена, белок 18,5 кД - всех, кроме II экзона, белок 17 кД - всех, кроме VI экзона, а белок 14 кД - всех, кроме II и VI экзонов. При этом образующиеся белки идентичны по аминокислотным последовательностям, кодируемым общими экзонами.
Еще одним примером служит так называемый РРТ ген крысы, который кодирует препротахикинины а - и р-типов - предшественники целого семейства нейропептидов - тахикининов. Первый из них содержит последовательность вещества Р, а второй - вещества Р и вещества К. Анализ первичной структуры показал, что эти РНК образуются в результате альтернативного сплайсинга по экзонам, кодирующим вещество К.
Разнообразие потенциал-зависимых и лиганд-зависимых ионных каналов в мембранах нервных клеток обеспечивается существованием кодирующих такие каналы мультигенных семейств и опять-таки альтернативным сплайсингом - Один и тот же ген у дрозофилы кодирует четыре полипептидные цепи, участвующие в формировании функционально активного К-канала. Эти полипептиды имеют одинаковые центральные домены, содержащие характерные для потенциал-зависимых каналов элементы.
Как и в описанных выше системах, разные полипептидные цепи К-канала возникают в результате альтернативного сплайсинга одного первичного транскрипта. Для формирования функционально активного К-канала необходима ассоциация четырех одинаковых или разных полипептидов: очевидно, что комбинирование различных полипептидов может обеспечить широкое разнообразие различающихся по физиологически значимым параметрам каналов.
Еще один яркий пример использования альтернативного сплайсинга - образование семейства синаптических рецепторов глутаминовой кислоты. Каждый из четырех рецепторов этого семейства существует в двух вариантах, различающихся лишь коротким сегментом, предшествующим четвертому трансмембранному домену. Согласно существующей топологической модели рецептора этот сегмент имеет цитоплазматическую локализацию. В генах каждого из рецепторов альтернативные варианты 38-ами-нокислотного сегмента кодируются двумя соседними экзонами, а сами варианты рецепторов образуются в результате альтернативного сплайсинга пре-мРНК по этим экзонам. Добавим, что существование альтернативных вариантов для каждого из рецепторов функционально значимо: они имеют различные фармакологические и кинетические свойства и по-разному распределены в отделах ЦНС.
Наконец, роль альтернативного сплайсинга показана при образовании четырех форм тирозингидроксилазы у человека, трех форм ацетилхолинэстеразы в электрическом органе ската, трех форм периферию у мыши, полипептидов у аплизии, специфических для нейрона R15, и в ряде других случаев.
Очевидно, альтернативный сплайсинг является эволюционно древним и широко распространенным в клетках нервной системы способом увеличения качественного разнообразия синтезируемых в них полипептидов.
Выше уже отмечено, что общее число транскрибируемых в мозге генов в 1,5-2 раза выше, чем во всех остальных тканях, и составляет, по-видимому, несколько десятков тысяч. Многообразие экспрессируемых в целом мозге генов объясняется двумя причинами:
1) разнообразием таковых в каждом индивидуальном нейроне и 2) различиями в наборах генов, экспрессируемых в разных нейронах. Именно налагаясь друг на друга, эти два фактора являются причиной исключительного разнообразия образующихся мРНК и соответствующих белков. Следует подчеркнуть, что разнообразие синтезируемых в любой ткани последовательностей РНК связано преимущественно с относительно редко встречающимися молекулами, которые составляют небольшую долю от общей массы РНК. К мозгу это приложимо в большей мере, чем к любому другому органу или ткани. Поэтому измерения суммарного синтеза РНК и ее общего количества практически не позволяют судить о качественных характеристиках транскрипции генома.
Транскрибируемость уникальных последовательностей ДНК в мозге млекопитающих прогрессивно возрастает в позднем эмбриогенезе и раннем постнатальном онтогенезе, достигая максимума к моменту функционального созревания. Обнаружено, что транскрибируемость генома в различных отделах мозга человека неодинакова: в гностических областях коры больших полушарий она выше, чем в проекционных, в лобной коре левого полушария значительно выше, чем правого, в мозжечке и стволовых отделах мозга уровень транскрипции - промежуточный. Эти данные позволяют сделать вывод о том, что развитие сложных гностических функций в мозге человека связано с прогрессивным увеличением генетического разнообразия составляющих его клеточных элементов.
В последние годы появились работы, позволяющие сделать прямые оценки степени генетической специализации клеток мозга. Они основаны на исследовании локализации различных мозгоспецифических антигенов с помощью моноклональных антител и локализации индивидуальных мРНК с помощью комплементарных клонированных последовательностей. Главным недостатком этих методов является их низкая представительность: в каждой работе исследуется ничтожная доля от всех синтезируемых в клетках мозга мРНК и белков. Добавим также, что в сфере этих исследований в большинстве случаев оказываются белки и мРНК, присутствующие в мозге в относительно высокой концентрации и уже в силу этого обстоятельства экспрессируемые в обширных популяциях нервных клеток. Так, обнаружены антигены, специфические для основных типов нервных клеток, а также для нейронов определенных отделов мозга. В рамках соответствующих отделов антигены могут иметь еще более выраженную клеточную специфичность. Известны антигены, которые экспрессируются в перекрывающихся популяциях нейронов в одних отделах ЦНС и неперекрывающихся популяциях - в других. Аналогичные, хотя пока и не столь многочисленные данные получены при исследовании локализации мозгоспецифических мРНК гибридизацией insitu. Судя по морфологическим, нейрофизиологическим, нейрохимическим и другим критериям, в мозге млекопитающих минимальными единицами такой специализации являются группы из десятков-сотен клеток, число которых в мозге крысы - 7104, а в мозге человека - 5-107, Это заведомо превышает общее число экспрессируемых в них генов. По-видимому, морфофункциональная специфичность клеток в мозге определяется уникальностью всей комбинации экспрессируемых в них генов и положением этих клеток в специфических нейронных ансамблях. В то же время тот факт, что такие группы насчитывают десятки-сотни клеток с одинаково экспрессируемым геномом, может объясняться необходимостью повышения надежности работы всей системы в целом. Однако в ЦНС некоторых беспозвоночных такая специализация распространяется на индивидуальные нейроны.
С помощью ДНК-РНК-гибридизации показано, что в ЦНС моллюска-кальмара экспрессируется 46% уникальных последовательностей генома, что достаточно для кодирования нескольких десятков тысяч различных мРНК. Следовательно, генетическая сложность клеточных элементов в ЦНС высокоорганизованных беспозвоночных сопоставима с таковой в ЦНС млекопитающих. В отличие от млекопитающих, однако, у кальмара не обнаружено специфической для ЦНС популяции молекул поли+РНК.
Уникальную возможность для исследования экспрессии генов в индивидуальных нейронах представляет ЦНС брюхоногих моллюсков, состоящая из небольшого числа крупных, во многих случаях легко идентифицируемых нейронов, сосредоточенных в нескольких ганглиях. У аплизии некоторые нейроны достигают размеров 0,5 мм и содержат - 0,25 мкг ДНК и - 5 нг поли+РНК, что вполне достаточно для анализа методами молекулярного клонирования.
С помощью процедуры дифференциальной гибридизации клонированы гены и мРНК, специфически зкспрессирующиеся в отдельных нейронах ЦНС аплизии. В их число входят гены, продукты которых хорошо идентифицированы; например ген, кодирующий гормон откладки яиц, и четыре родственных ему гена. Будучи активными в разных нейронах, они кодируют синтез нескольких физиологически активных пептидов, секреция которых индуцирует стереотипный поведенческий репертуар откладки яиц. При этом один и тот же пептид может действовать как нейрогормон на клетки соматических тканей и нейромедиатор на определенные нейроны. Экспрессия других генов этого семейства в клетках атриальных желез приводит к синтезу нейроактивных пептидов, вызывающих активацию сумчатых клеток, а также секретируемых во внешнюю среду пептидов, обладающих активностью половых феромонов. Таким образом, кодируемые этим семейством генов пептиды регулируют различные компоненты одного сложного поведенческого репертуара.
Очень интересен также наплизии, участвующий в регуляции водно-соленого баланса. Он кодирует две мРНК, образующиеся в результате альтернативного сплайсинга. Одна из них является преобладающим продуктом экспрессии гена в одном из нейронов, тогда как укорочения форма мРНК преимущественно синтезируется в некоторых других нейронах. Следовательно, у аплизии существуют механизмы выбора различных путей сплайсинга в разных нейронах.
Изучение экспрессии генов в индивидуальных нейронах этого моллюска позволяет сделать некоторые принципиальные выводы, которые могут быть применимыми и к ЦНС позвоночных животных. Главный из этих выводов состоит в том, что присутствие множества "редких" молекул РНК в суммарных препаратах поли*РНК, изолированных из целой ЦНС или ее крупных отделов, является следствием активной экспрессии этих РНК в небольших популяциях нервных клеток, а не их экспрессии на одинаково низком уровне в обширных популяциях клеток.
Сравнение популяций мРНК, синтезируемых в нейронах, использующих один и тот же классический медиатор, но функционально различных, показывает, что такие нейроны обычно различаются экспрессией нескольких мРНК, каждая из которых специфична лишь для одного из сравниваемых нейронов. Очевидно, общее разнообразие синтезируемых в ЦНС мРНК складывается из перекрывающихся, но не одинаковых популяций мРНК, образуемых в индивидуальных нейронах. Остается добавить, что рассмотренные исследования охватывают лишь относительно часто встречающиеся молекулы мРНК. Не исключено, что в действительности различия в популяциях синтезируемых мРНК между индивидуальными нейронами носят более сложный характер.
К популярным объектам нейрогенетики из числа беспозвоночных относится и одна из свободно живущих нематод - Caenorhabditiselegans. Одним из важных результатов ее исследования является обнаружение так называемых селекторных генов. Последние играют ключевую роль в онтогенезе нейронов нематоды: их продукты индуцируют включение серии "генов-реализаторов", формирующих фенотип нейронов. Примером селекторного гена может служить ген INS-4, детерминирующий специфичность синаптических контактов идентифицируемого мотонейрона VA. При мутациях этого гена мотонейрон VA образует синаптические контакты, в норме ему не свойственные. Никаких других заметных нарушений в развитии ЦНС при этом не обнаруживается. Продуктом гена INS-4 является гомеобокс-содержащий ДНК-связывающий белок. Очевидно, он регулирует транскрипцию группы генов-реализаторов, непосредственно участвующих в установлении специфических для данного мотонейрона синаптических контактов.
Наконец, классический объект генетических исследований - дрозофила - позволил получить обширную информацию о сложных процессах онтогенеза центральной нервной системы. Эту информацию можно обобщенно представить в виде модели многоступенчатой генетической детерминации нейрогенеза, основные положения которой, по-видимому, сохраняют силу и для позвоночных животных. В соответствии с этой моделью существует четыре класса генов, регулирующих последовательные этапы нейрогенеза. Первый, наименее изученный класс, включает гены, ответственные за преобразование части клеток недифференцированной вентральной эктодермы в предшественники нервных клеток. Ко второму классу относятся гены, детерминирующие свойства клеток-предшественников в соответствии с их положением в той или иной части ЦНС. Среди этих генов - многие из так называемых гомейотических или сегментарных генов, которые детерминируют также и формирование общего плана строений организма в целом. При этом одни и те же гены обычно детерминируют и формирование определенного сегмента организма, и специфические свойства локализованного в нем сегмента ЦНС. Заметим, однако, что сегментарная и нейроспецифическая экспрессия гомейотических генов контролируется разными механизмами. Гены третьего класса детерминируют свойства индивидуальных нейробластов в зависимости от их локализации внутри данного сегмента ЦНС. Представителем этого класса, по-видимому, является ген prospero, экспрессирующийся только в определенных нейробластах каждого сегмента ЦНС, но не в образующихся из них нейронах. Мутации этого гена приводят к выпадению из развивающейся нервной системы нескольких специфических клеточных линий. Наконец, гены четвертого класса детерминируют индивидуальные свойства каждого из нейронов, образующегося при делении одного нейробласта. Представителями этого класса являются гены fushitarazu и even-skipped. Интересно, что один и тот же ген может участвовать в контроле разных этапов нейрогенеза. Так, ген ftz является типичным гомейотическим геном, участвующим в формировании общего сегментарного плана строения ЦНС и организма в целом, т.е. входит во второй класс генов. На более поздних этапах он участвует в детерминации свойств индивидуальных нейронов: в каждом сегменте ЦНС он экспрессируется в группе из 30 идентифицируемых нейронов, являющихся потомками - 8 нейробластов. При этом ни в одном из нейробластов ген ftz не экспрессируется, а процент экспрессирующих его клеток в потомстве индивидуальных нейробластов варьирует от 0 до 100%. Заметим, однако, что сегментарная и нейроспецифическая экспрессия гена ftz контролируется разными механизмами. Еще один фундаментальный вывод описанных исследований состоит в том, что индивидуальная специфичность элементов нервной системы определяется не уникальными генами, количество которых заведомо меньше числа самих элементов, а уникальными комбинациями взаимодействующих генов, надежность которых обеспечивается чертами функциональной избыточности.
Гены, ответственные за формирование специфического фенотипа функционально зрелых нейронов, изучены главным образом для относительно просто устроенных нейронных структур, например сетчатки глаза дрозофилы. Каждый элемент сетчатки - омматидий - состоит из 8 четко различимых по морфологии и локализации фоточувствительных нейронов и 12 ненейрональных клеток. Дифференцировка нейронов в развивающихся омматидиях протекает в строго фиксированной последовательности. Недифференцированные клетки в зачатках омматидиев являются эквипотенциальными, а судьба дифференцирующихся клеток определяется не их происхождением, а индуцирующими стимулами со стороны соседствующих с ними ранее дифференцированных клеток.
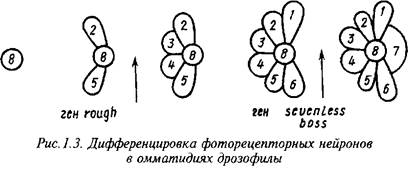
Некоторые из генов, ответственных за генерацию и рецепцию этих стимулов, хорошо изучены: ген seveniess кодирует мембранный рецептор, необходимый для индукции дифференцировки нейрона R7, а ген brideofsevenless - за образование в нейроне R8 лиганда, действующего на этот рецептор. Ген seveninabsentia кодирует ядерный белок, интерпретирующий сигнал, воспринимаемый рецептором sevenless. Ген rough кодирует транскрипционный фактор, необходимый для возникновения в нейронах R2 и R6 сигнала, индуцирующего дифференцировку соседних клеток в нейроны R3 и R4. Другой транскрипционный фактор, необходимый для дифференцировки нейронов Rl, R3, R4 и R6, кодируется геном seven-up. Ключевым элементом в реализации генетических программ функционального созревания фоторецепторных нейронов является ДНК-связывающий белок, кодируемый геном glass, а специфическими для индивидуалъньгх фоторецепторных нейронов компонентами этих программ - гены, кодирующие разные формы фоторецепторных белков.
Важную роль в окончательной морфологической дифферен-цировке нейронов играют гены, кодирующие молекулы избирательной межклеточной адгезии. Одной из разновидностей таких молекул у насекомых являются фасциклины, обеспечивающие взаимное узнавание и ассоциацию растущих аксонов в развивающейся ЦНС и, в конечном итоге, формирование стереотипной системы аксонных пучков. В эмбриональной ЦНС разные фасциклины экспрессируются в разных популяциях нейронов и их отростков: каждый фасцикяин экспрессируется в аксонах - 5 из - 30 аксонных пучков передних и задних комиссур каждого ганглия и - 15 нейронах каждого полусегмента. В эмбриогенезе экспрессия фасциклинов наблюдается уже в отдельных клетках морфологически недифференцированной вентральной нейроэктодермы, сохраняясь в образующихся из этих клеток нейробластах, ганглионарных материнских клетках и нейронах. По-видимому, фасциклины входят в число наиболее ранних молекулярных маркеров индивидуальных нейрональньгх линий. Обнаруженные закономерности при исследовании относительно просто устроенной ЦНС беспозвоночных не всегда находят прямые аналогии в более сложно организованных ЦНС высших позвоночных животных. Тем не менее с известной осторожностью их можно использовать при интерпретации данных по экспрессии генов в мозге позвоночных.
Возникновение ЦНС из недифференцированных клеток эктодермы до настоящего времени остается одним из самых загадочных процессов в эмбриогенезе позвоночных. Считается, что исходно клетки эктодермы не имеют самостоятельного нейрогенного потенциала, а их дифференцировка в нервные клетки индуцируется какими-то сигналами, поступающими из нижележащих клеток эмбриональной мезодермы. Молекулярная природа этих сигналов и их первичных эффектов в клетках эктодермы до последнего времени была совершенно неизученной. Совсем недавно обнаружено, что, например, компонентом такого первичного эффекта в эмбрионах шпорцевых лягушек является индукция гена Х1НЬохб, содержащего характерную для многих генов, регулирующих онтогенез, эволюционно консервативную последовательность, так называемый гомеобокс. Он тесно связан с определением локализации групп клеток и тканей, с "географией" клеток и функций в организме.
Экспрессия этого гена характерна для наиболее ранних стадий нейрогенеза. Ген ХШЬохб имеет четко выраженный сегментарный характер экспрессии в эмбриональной ЦНС: его транскрипты обнаруживаются только в задних двух третях нервной трубки. Подобно многим гомейотическим генам дрозофилы ген ХШЬохб перестает экспрессироваться на поздних стадиях эмбриогенеза.
Ген XlHboxl экспрессируется в спинном мозге, причем передняя граница экспрессирующей его области - очень резкая и в точности совпадает с границей спинного и головного мозга. При инактивации кодируемого этим геном белка специфическими антителами нарушается развитие передних отделов спинного мозга: они трансформируются в структуру, морфологически идентичную продолговатому мозгу.
Очевидно, кодируемые этими генами белки являются специфическими транскрипционными факторами, регулирующими генетическую детерминацию клеточных линий в определенных отделах эмбриональной ЦНС. Они обеспечивают реализацию позиционной информации в общем процессе нейрогенеза.
В геноме мыши обнаружено четыре комплекса тесно сцепленных гомеобокс-содержащих генов, Нох-1, - 2, - 3, и - 4. Индивидуальные гены каждого комплекса аналогично гомейотическим генам дрозофилы экспрессируются в различных относительно передне-задней оси тела областях ЦНС. Морфологически различимые сегменты в эмбриональной ЦНС позвоночных обнаруживаются лишь на ранних стадиях развития заднего мозга в виде так называемых ромбомеров, тогда как на более поздних стадиях и в мозге взрослых животных рудиментом сегментарного строения остается лишь распределение ядер черепно-мозговых нервов.
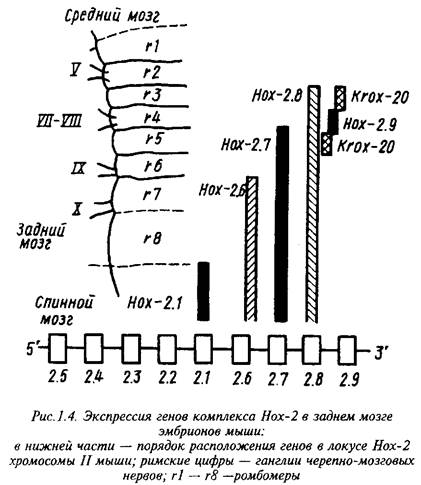
Индивидуальные гены Нох-2 комплекса имеют общую область экспрессии в каудальных отделах эмбриональной нервной трубки, тогда как передние границы экспрессирующей области у них различны: чем ниже данный ген расположен в Нох-2 локусе хромосомы, тем выше расположена эта граница.
Сейчас интенсивно накапливаются и обобщаются новые данные в этой области. Эволюционная консервативность регуляторных генов, экспрессируемых в сегментарных отделах эмбриональной ЦНС позвоночных, и явные аналогии в характере их экспрессии с гомейотическими генами дрозофилы делают весьма привлекательной гипотезу об их роли в детерминации общего плана строения и идентичности отдельных клеточных линий в развивающейся ЦНС.
Ни один из исследованных генов Нох комплексов не экс-прессируется в передних отделах головного мозга позвоночных в эмбриогенезе. Эти отделы не проявляют явных признаков сегментарного строения. Являясь относительно эволюционно новыми приобретениями, они, по-видимому, имеют специфичные механизмы генетической детерминации. Показано, в частности, что в детерминации отделов переднего и промежуточного мозга существенную роль играет ген Dlx, гомологичный гомейотическому гену D11 дрозофилы. Аналогично, в детерминации среднего мозга и, возможно, передних отделов мозжечка участвует протоонкоген Wnt-1 и гомеобокс-содержащие гены Еп-1 и - 2.
Имеются данные, заставляющие по-новому взглянуть на вопрос о функциях гомеобокс-содержащих генов у высших животных. Клонирование генов, кодирующих транскрипционные факторы Oct-1, Oct-2 и Pit-1, показало, что они являются членами особого подсемейства гомеобокс-содержащих генов, в которое входит также ген Unc-86 нематод. Общим признаком, объединяющим эти гены, является длинная консервативная последовательность, POU-домен, частью которой является гомеобокс. Ген Unc-86 участвует в детерминации нейронных линий у нематод. Несколько генов POU-подсемейства специфически экспрессируются в отделах мозга млекопитающих: Вгп-1, - 2, - 3 и Tst-I. Все четыре гена экспрессируются в эмбриональной нервной системе, включая клетки герминативных зон.
Динамика экспрессии индивидуальных генов в развивающемся мозге очень различна и по мере развития отделов мозга приобретает черты, отражающие их паттерн экспрессии в мозге взрослых животных. Экспрессия этих генов в клетках герминативных зон может означать, что судьба индивидуальных клеточных линий в нервной системе предопределена уже до начала их миграции из герминативной зоны.
По-видимому, кодируемые гомеобокс-содержащими генами транскрипционные факторы могут регулировать экспрессию генов не только в формирующихся клеточных линиях в раннем эмбриогенезе млекопитающих, но и в дифференцированных клетках взрослого организма, причем на очень разных уровнях клеточной специфичности.
Наконец, особого рассмотрения заслуживает тот факт, что в мозге млекопитающих есть ряд генов, особенно быстро реагирующих на разнообразные экстремальные воздействия. Их включение в ряде ситуаций, в частности, сопровождающихся повышением уровня внутриклеточного Са2+, служит как бы первым относительно неспецифичным звеном реакции генома нейрона. Далее следует включение более специфичных участков генома. В число таких генов "первоочередного реагирования" входят гены, открытые ранее в связи с исследованием протоонкогенов. В частности, гены c-fos и c-jun кодируют факторы - регуляторы транскрипции других генов, - белки Fos и Jun. Последние входят, в свою очередь, в состав белка АР-1, связывающегося с определенными локусами ДНК, - например возле 5'-конца гена, кодирующего фактор трофики нейронов - NGF. В экстремальных ситуациях - при эпилептических судорогах, требующих срочного усиления трофики нейронов, - этот механизм включает синтез NGF. Аналогичные механизмы с участием генов c-fos и c-jun срабатывают под влиянием болевых импульсов и других экстремальных воздействий на ЦНС, включая целые каскады транскрипции генов, кодирующих, например, такие противоболевые факторы, как опиодные пептиды и другие регуляторы. В последнее время c-fos и другие гены раннего реагирования нередко называют "третьим мессенджером". Наконец, появляется все больше данных об участии генов этого типа в механизмах запоминания.
1. Большинство нейронов ЦНС позвоночных являются диплоидными; небольшая доля нейронов в некоторых отделах ЦНС может содержать избыточное по сравнению с диплоидным количество ДНК.
2. Репликативный синтез ДНК в дифференцированных нейронах отсутствует; в мозге взрослых млекопитающих репликация ДНК связана главным образом с ограниченными процессами размножения глиальных клеток.
3. В клетках мозга млекопитающих имеются активно функционирующие системы репарации ДНК, поддерживающие целостность и эффективность генетического аппарата.
4. Хроматин нервных клеток имеет типичную для эукариотических клеток нуклеосомную организацию. Особенностями хроматина нейронов неокортекса млекопитающих являются необычно короткие нуклеосомные единицы, присутствие редких вариантов гистонов, высокое разнообразие негистоновых белков и высокая матричная активность.
5. В мозге млекопитающих экспрессируется несколько десятков тысяч уникальных генов, из которых не менее половины имеют мозгоспецифический характер экспрессии.
6. Огромное разнообразие экспрессируемых в мозге генов складывается из перекрывающихся, но не одинаковых популяций генов, экспрессируемых в отдельных нервных клетках.
7. Наряду с разнообразнейшими мозгоспецифическими мессенджер-РНК в центральной нервной системе синтезируется ограниченное число особых малых РНК с последовательностями нуклеотидов, общими для всех отделов мозга.
8. Разнообразие белков и регуляторных пептидов, синтезируемых в мозге, определяется не только большим набором экспрессируемых генов, но и системой альтернативного сплайсинга пре-мРНК.
|
