Федеральное агентство по образованию
Южно – Уральский Государственный Университет
Факультет физической культуры и спорта
Кафедра теории и методики физической культуры и спорта
Курсовая работа
По дисциплине: концепции современного естествознания
Современные концепции возникновения жизни
Выполнил студент
группы ФКиС – 237
Стрижов Артём Николаевич
Проверил: Полтинкина И.В.
Челябинск - 2005
Содержание
Введение
Глава
I
. Обзор литературы
§1.1 Гипотеза Крика – Оргела
§1.2 О возможностях случайного посева
§1.3 Теория Ю. А. Колясникова
§1.4
Теория У. Мартина и М. Рассела
§1.5 Гипотеза Сванте Аррениуса
§1.6
Гипотеза А.И Опарина
Глава
II
. Опытно – экспериментальная часть
§2.1 Абиотический синтез биомономеров
§2.2 Абиотический синтез полимеров
Заключение
Список литературы
Введение
Гипотезы — это леса, которые возводят
перед зданием и сносят, когда здание готово.
И. В. Гете
Жизнь — это самое прекрасное и в конечном итоге самое важное из всего того, что существует на нашей планете. Научное раскрытие тайн живой природы имеет исключительное, решающее значение как в теоретическом, мировоззренческом отношении, так и во всех тех областях практической деятельности человека, где он имеет дело с живыми объектами,—в области медицины, сельского хозяйства и ряда отраслей промышленности.
Проблема сущности жизни не может быть рационально разрешена без понимания ее возникновения и развития. Господствовавший еще недавно в естествознании механистический материализм, игнорируя это положение, тщетно пытался познать живой организм как какой-то сложный механизм, разбирая его, как часы, на отдельные винтики и колесики и на основании изучения этих деталей стремясь познать живое существо как целое.
Конечно, детальный анализ веществ и явлений, свойственных живым телам, чрезвычайно важен, совершенно необходим для правильного понимания жизни. Блестящие успехи современной биохимии и биофизики являются наглядным доказательством этого. Но как бы глубоко и полно мы не познали вещества, структуры и процессы, лежащие в основе организации современных живых тел, мы никогда, исходя только из одного этого, не сможем ответить на вопрос, почему эта организация является именно такой, какой она есть, в частности такой «целесообразной», т. е. организацией, при которой строение всех составляющих организм частей (молекул, органоидов и органов) так совершенно приспособлено к выполняемым ими функциям, а весь организм как целое приспособлен к существованию в данных условиях внешней среды.
Реклама

Все религиозные учения видят причину этой целесообразности в творческой воле создателя, который породил жизнь согласно определенному плану и направил
ее прогрессивное развитие к единой финальной цели, к конечному совершенству. Такой точки зрения придерживаются, однако, не только теологи, но и представители других идеалистических учений (холисты, финалисты, органицисты).
Только на основе диалектико-материалистических представлений, рассматривающих жизнь как особую качественно новую форму организации и движения материи, возникшую когда-то в процессе эволюционного развития материи, можно рационально подойти к решению проблемы происхождения жизни.
Успехи, достигнутые в настоящее время в различных областях современного естествознания, позволяют нам на указанной основе наполнить конкретным содержанием и первую часть нашей формулы, раскрыть те пути, которыми происходило возникновение жизни из безжизненной материи.
В разных пунктах Вселенной, на различных объектах нашего звездного мира эволюционное развитие материи осуществлялось и осуществляется различными путями и в различных темпах. Поэтому в схеме мы не должны его себе представлять как какую-то прямую линию, а скорее как целый пучок различных путей, отдельные ответвления которых могут приводить к очень сложным, совершенным, но глубоко различным между собой формам организации и движения материи.
Изучение зарождения жизни на Земле представляет собой исследование лишь одного примера события, которое должно было происходить в мире бессчетное число раз. Поэтому выяснение вопроса о том, как появилась жизнь на Земле, безусловно, должно служить убедительным доводом в пользу теории о существовании жизни и на других объектах Вселенной.
Глава
I
. Обзор литературы
§1.1 Гипотеза Крика – Оргела
Позже идею панспермии в очень решительной форме – как доставку жизни к мёртвой Земле на космическом корабле – выдвинули Ф. Крик и Л. Оргел.
И в этом случае выступили учёные, мнение которых трудно игнорировать. Например, Френсис Харри Комптон Крик (р. 1916) – английский биофизик и генетик, лауреат Нобелевской премии. В 1953 г. совместно с Дж. Уотсоном он создал модель структуры ДНК (двойную спираль), что положило начало молекулярной генетике. Является автором трудов по расшифровке генетического кода, по нейрофизиологии и др Взгляды Крика и Оргела поддерживаются рядом других учёных.
Реклама

Однако, коммунистическая наука не считала такие взгляды материалистическими и на корню отвергала их, подобно идее божественного творения. Большинство советских учёных полагало, что идея панспермии – это лишь уход от решения проблемы происхождения жизни.
В памяти ещё не стёрлась история генетики и кибернетики, долго отвергавшихся коммунистической наукой, а теперь определяющих вершины научного прогресса. Не уготован ли такой же путь и науке о происхождении жизни? Вероятно, здесь не обойтись без борьбы мнений, но разве правильное понимание биологической истории менее существенно, чем иные фундаментальные проблемы?
Для земной науки оказалась непосильной загадка 100%-ной зеркальной асимметрии биологических молекул. К ней примкнули загадки единства аминокислотного набора, генетического алфавита и кода, «энергетической валюты» в виде АТФ. Комплекс фундаментальных загадок земной жизни довершился отсутствием ожидаемой предбиологической эволюции. Но фундаментальные загадки разом исчезают, если признать целенаправленное занесение первых клеток на Землю извне. Тогда обнаруживается точное соответствие между теоретическими ожиданиями и реальными фактами.
Такая же ситуация со взглядами А.И. Опарина. Все перечисленные особенностиранних этапов эволюции (однотипная хиральность биомолекул, одинаковый набор аминокислот, единые генетический алфавит, генетический код и АТФ, отсутствие предбиологической эволюции) противоречат ожиданиям, только если стоять на почве гипотезы А.И. Опарина о зарождении жизни на Земле.Достаточно перейти к гипотезе панспермии, как эти противоречия разом отпадают.
Если раньше гипотеза А.И. Опарина вступала в конфликт, главным образом, с фактами биохимии, то после успехов микропалеонтологии архея
стало ясно, что ей противоречит и этот раздел науки. Беда не в том, что обнаружилось непропорционально долгое формирование клеточного ядра. Важно, что в палеонтологической летописи вообще не оказалось места для длительной предбиологической эволюции, неминуемой в случае естественного возникновения жизни на Земле. При этом последующие палеонтологические открытия не способны изменить ситуацию.
Например, если бы выяснилось, что одноклеточные эукариоты
появились ранее, чем мы думаем сегодня (скажем, 3,3–3,5 млрд. лет назад), то малому времени предбиологической эволюции стало бы противоречить слишком долгое формирование многоклеточных организмов на основе одноклеточных эукариот (в таком случае оказалось бы, что многоклеточные формировались около 2 млрд. лет).
Палеонтологические данные о сроках возникновения жизни поставили дилемму:
– либо жизнь возникла на Земле сама собой, но это произошло с немыслимо высокой скоростью, из-за чего процесс не оставил следов в палеонтологической летописи,
– либо жизнь занесена на Землю извне.
На решение этой дилеммы существенно влияет парадокс молекулярной асимметрии земной жизни. Даже если бы предбиологическая эволюция, вопреки всему, длилась, скажем, 100–200 тысяч лет, что могло бы не оставить палеонтологических следов, то и тогда предшественниками биохимических процессов могли явиться лишь обычные химические реакции, не проявляющие предпочтения к молекулам определённой хиральности. Иначе говоря, независимо от длительности предбиологического этапа, на Земле обязательно остались бы следы жизни с разными типами хиральности молекул.
И наоборот – если предбиологическая эволюция протекала на иной планете, то нет и проблем хиральности, бесполезно искать на Земле наследие чисто химических процессов в виде равного (или неравного) участия в живой материи молекул левой и правой симметрии. Если эти следы и сохранились, то находятся вне Земли.
Известно, что земные организмы используют два разных генетических кода. Первый из них –собственно код генов на ДНК, а второй – это структурогенный код пространственного расположения генов в ядре. Первый код используется во всех организмах, второй – только у эукариот. Первый код явно возник не на Земле, а был занесен сюда, второй – так же неоспоримо является продуктом местной, земной эволюции.
Сравнение этих двух случаев особенно подчёркивает привнесенность первого кода и автономное происхождение второго. Тогда как палеонтология не обнаруживает периода формирования первого кода, на возникновение второго затрачены миллиарды лет от возникновения жизни до появления первых эукариот. И тогда, как биология не видит ступеней формирования кода ДНК, детально прослеживаются этапы становления генетической системы эукариот.
При поверхностном взгляде может показаться, что исходная позиция учёного – согласие или несогласие с возможностью занесения жизни из космоса – не способна повлиять на итог исследований. Мол, всё равно, в конце концов, нужно узнать, как возникло живое из неживого. Но нет! Исходная позиция, исходные ограничения имеют огромное, принципиальное значение!
Вообразим, что после изучения горных кряжей обжитых континентов учёные стали исследовать погребённые подо льдом горы Элсуорта в Антарктиде. И вдруг в горном массиве Винсон обнаружили верхушку горы, сложенную из полированных гранитных плит! Одни учёные стали думать – как такие плиты могли туда попасть? А другие, поскольку в Антарктиде не известно цивилизаций, сразу отмели возможность их искусственного происхождения и занялись поиском условий, ведущих к естественному формированию горной вершины из гладких гранитных блоков. Легко представить себе, в какие дебри завели бы такие поиски!
Аналогичная ситуация сложилась с попытками объяснить естественное возникновение жизни на Земле. Исследователи натолкнулись на неоспоримые факты, противоречащие гипотезе А.И. Опарина, но хорошо объяснимые идеей панспермии. И только идеей панспермии. Как же реагировать на это?
§1.2 О возможностях случайного посева
Для понимания первых этапов жизни на Земле и её дальнейшего развития важно знать не только, имела ли место панспермия, но если она произошла, то в каком варианте? Был ли это случайный перенос мельчайших клеток через космос, либо перенос их метеоритами, как представляли себе многие сторонники идеи панспермии, или же это была целенаправленная доставка на космическом корабле специально подобранных микроорганизмов в соответствии со сценарием Ф. Крика и Л. Оргела?
Литературные данные позволяют уточнить ситуацию. Одно из направлений исследований было связано с изучением возможности случайного переноса жизнеспособных клеток через космос.
После того, как П.Н. Лебедев экспериментально доказал, что свет оказывает давление, С. Аррениус рассчитал, что под давлением солнечных лучей споры с поперечником 1,5–2 мкм могут за 14 месяцев покинуть пределы нашей планеты. Позже американский учёный К. Саган указывал, что частицы размером 0,2–0,6 мкм могут быть вынесены с Земли световым давлением и за несколько недель могут достигнуть орбиты Марса. Орбиты Нептуна они могут достигнуть через несколько лет, а до ближайших звёзд долететь за несколько десятков лет. При этом он исходил из предположения, что споры движутся прямолинейно с достаточно высокой скоростью.
Однако подобные предположения отпадают, если вспомнить, что на орбите Земли ультрафиолетовая радиация Солнца имеет плотность 1,4*106
эрг на 1 см2
/с. Даже если допустить нереально высокую скорость перемещения спор в космическом пространстве, равную четверти скорости света, то полёт к Марсу займёт около 1000 секунд. Тогда доза облучения составит 1,4*109
эрг/см2
, что во много раз выше смертельной дозы 102
–106
эрг/см2
[Рубенчик, 1983].
Поскольку выяснилось, что ультрафиолетовая радиация Космоса гарантированно убивает незащищённые клетки (большинство видов клеток не может переносить и воздействие вакуума), особое внимание привлекли метеориты, способные стать для микроорганизмов как бы транспортными контейнерами.
Каждые сутки Земля принимает значительную массу этого космического вещества. За год выпадает от 10 тыс. до 1 млн. метеоритов и много метеоритной пыли. Вес метеоритов колеблется от миллиграммов до десятков тонн. Главное, что отличает метеориты от земных горных пород, это содержание в метеоритном веществе железа с примесью никеля. По количеству Fe метеориты подразделяются на железные, железокаменные и каменные. Больше всего на Землю попадает каменных метеоритов (около 90%), тогда как железных – около 5–6%.
В железных метеоритах металл подвергался воздействию температур выше 1500°С, а затем, как полагают некоторые исследователи, в течение десятков миллионов лет расплавленная масса охлаждалась и кристаллизовалась.
Для объяснения уникальных особенностей структуры метеоритов приходится заключить, что вещество, прошедшее глубокое плавление, составляет в них лишь некоторую часть. Как правило, плавлению при высокой температуре подвергалась лишь малая доля вещества, находившаяся при соударении исходных частиц непосредственно в зоне контакта. Главным формирующим процессом для них являлось медленное, длившееся миллионы лет, спекание мелких частиц в вакууме под действием сил адгезии и космических излучений.
В этом смысле микроструктура метеоритов близка не к монолиту чугунной отливки, а к микроструктуре твёрдосплавных инструментов из «победита», изготавливаемых путём прессования мельчайших гранул при температуре, намного ниже их точки плавления. И твёрдосплавные пластинки, и метеориты, на микроуровне представляют собой губчатую структуру, пронизанную густой сетью капилляров. Это очень важно с точки зрения возможного переноса Жизни через Космос.
Особый интерес представляют углеродистые хондриты, относящиеся к каменным метеоритам. Кроме железа, они содержат серу, связанную воду и до 5% углерода в виде различных органических соединений. Это битумообразные вещества, содержащие углеводороды, ароматические и жирные кислоты, серо- и хлорсодержащие органические соединения, углеводы и др.
В хондритах найдены аминокислоты, присутствующие в земных организмах (глицин, аланин, глутаминовая кислота и др.), а также аминокислоты, не свойственные им. Например, в метеорите Мурчисон выявлены не используемые земными организмами аминокислоты 2-метилаланин и саркозин. (Интересно, что 2-метилаланин и саркозин также оказались синтезированными в опытах, при которых моделировались условия первобытной Земли).
Микроорганизмы искали в метеоритах ещё в прошлом столетии. В 1864 г. вблизи французского селения Оргейль упал крупный метеорит. Луи Пастер сконструировал специальный зонд для стерильного извлечения проб из внутренних частей метеорита. Однако посевы на питательные среды дали отрицательный результат – жизнеспособные микроорганизмы не были обнаружены. В 1921 г. В Галипп и Ж. Суфлянд обследовали 21 метеорит различных типов и обнаружили в них как подвижные, так и неподвижные микроорганизмы.
В 1932 г. Ц. Липман провёл исследования внутренних зон метеоритов, тщательно стерелизовав перед этим их поверхности, и выявил в глубине значительное количество бактерий. Были обнаружены бактерии и в железистом хондрите Оргейль, в котором их безуспешно искал Пастер.
В 1961 г. Ф. Сислер выделил из метеорита Муррей несколько бактерий, которые росли на питательных средах, но не развивались при введении их в организмы крыс, мышей и кур. Однако при проверке в 1962 г. М.Н. Бриггсом эти данные не подтвердились.
В 1962 г. Ч. Байриев и С. Мамедов исследовали один из кусков Сихоте-Алинского железного метеорита через 15 лет после его падения и обнаружили бактериальную палочку, сохраняющую жизнеспособность при повышенной температуре.
Таким образом, в метеоритах были найдены жизнеспособные микроорганизмы. Но действительно ли эти микроорганизмы принесены из далёкого космоса? Например, Дж. Бернал предположил, что при вулканических извержениях земная пыль, вместе с содержащимися на ней микробными клетками, может уноситься в межпланетное пространство, встречаться там с метеоритами, прилипать к ним и с ними возвращаться на Землю.
Наконец, не исключено, что микроорганизмы могут проникать внутрь метеоритов уже после их падения. Чтобы проверить эту возможность, в 1970 г. А.А. Имшенецкий и С.С. Абызов провели эксперименты над образцами горных пород – туфа, пемзы, талька, магнезита, гранита, каменного угля – размером 10´10 см.
Образцы стерилизовались 8–12 часов в муфельной печи при 400–500°С, а затем в автоклаве при избыточном давлении 1 атм. Часть стерильных образцов помещалась в ящик с влажной почвой, а часть (для контроля) хранилась в лабораторных условиях. Через разные сроки (от 8 дней до 6 месяцев) поверхность каждого образца тщательно стерилизовалась и специальным сверлом отбиралась проба из его центральной части. Посевы проб на питательные среды обнаружили рост микроорганизмов даже тогда, когда образцами являлись такие плотные породы, как туф, гранит и др. В контрольных образцах микробы ни разу не обнаружились.
Таким образом, почвенные микроорганизмы находили способ проникать внутрь исследуемых пород.
Другая серия экспериментов проводилась с образцами метеоритов – Сихотэ-Алинского (железный октаэдрит), Чинге (железный атаксит), Кунашак и Саратов (каменные хондриты). В образцах были просверлены вертикальные каналы диаметром до 10 мм, затем образцы подверглись стерилизации и в каналы была внесена почва с чистой культурой бактерий Serratiamarcescens. В таком виде образцы были частично погружены в питательную среду. Через 2–15 суток во всех питательных средах, кроме омывающей метеорит Чинге, было зарегистрировано массовое размножение микроорганизмов.
Это показало, что для микроорганизмов проницаемы не только различные горные породы, но и большинство исследованных метеоритов. Следовательно, метеориты, приземляющиеся на влажную почву, способны загрязняться почвенноймикрофлорой,
из-за чего обнаруженные в них жизнеспособные клетки не могут служить доводом в пользу метеоритного переноса Жизни.
Если нельзя получить данные о переносе метеоритами жизнеспособных клеток, то не помогут ли они, по крайней мере, узнать, существует ли жизнь в мире, из которого они прилетают? Иначе говоря, не содержат ли метеориты следов мёртвых, ископаемых (фоссилизированных) организмов? Такие исследования были проведены.
В 1961 г. Ж. Клаус и Б. Надь обнаружили в метеорите Оргейль так называемые «организованные элементы», которые, по мнению исследователей, могли являться ископаемыми (фоссилизированными) остатками каких-то биологических образований. Эти элементы люминесцировали в ультрафиолетовом свете, окрашивались, как и бактерии, оснóвными анилиновыми красителями, давали положительную реакцию по Фёльгену, характерную для ДНК. Исследователи предполагали, что это остатки микроорганизмов внеземного происхождения.
Впоследствии такие исследования были продолжены. В метеоритах, найденных в Советском Союзе (Мигеи, Старое Борискино, Саратов) и за рубежом (Ивуна, Але, Беккевельд, Топк) были обнаружены сходные образования размером от 4 до 30 мкм. В метеоритах Оргейль и Ивуна количество таких включений достигало 1700 мг. По поводу происхождения этих образований высказывались серьёзные сомнения. В 1970 г. Г.П. Вдовыкин показал, что некоторые «организованные элементы» представляют собой минеральные образования.
Таким образом, результаты исследований метеоритов не подтвердили и не опровергли возможности существования Жизни вне Земли. Они также не противоречат и не дают весомого подтверждения предположению о переносе метеоритами Жизни из иных миров на Землю.
Другая группа фактов связана с единством биохимии земных организмов.
Предположим, что какая-то частичка космической пыли или метеорит принесли в земной Океан клетку прокариотической водоросли. Могло бы это дать старт устойчивой биологической эволюции? Возможно. Но сомнительно. Вероятнее, что за первым этапом быстрого размножения последовало бы исчерпание питательных веществ, отравление среды продуктами собственной жизнедеятельности – что мы часто наблюдаем при «цветении воды» в изолированных водоёмах – и гибель клеток.
Дело даже не в том, что обязательно погибли бы все до одной клетки. На периферии очага жизни, вероятно, сохранились бы немногочисленные клетки, позволяющие жизни «теплиться» далее. Но не смогло бы образоваться такое количество продуктов фотосинтеза, чтобы его можно было найти почти через четыре миллиарда лет, как они найдены, например, в разрезе Исуа (Западная Гренландия) в виде отложений нефтеподобного вещества.
Чтобы на Земле возникли значительные массы продуктов фотосинтеза, оставившие весомые отложения нефтеподобного вещества, должна была сформироваться устойчивая экосистема с замкнутым круговоротом наиболее критичных веществ (фосфора, азота, серы и др.). К этому времени (3,69–3,83 млрд. лет назад) в Океане уже должны были существовать, вероятно, не менее 3-4-х типов клеток, по характеру биохимических процессов напоминающих фотосинтезирующие водоросли, хемотрофов и животных.
Если бы в Океан был занесен только один тип клеток, от которого все остальные типы (родоначальники будущих крупных таксонов) произошли значительно позже, то, соответственно, и обнаруженное нефтеподобное вещество датировалось бы более поздней отметкой, а не появилось бы практически сразу за возникновением на Земле необходимых для жизни условий.
Но этого мало! Если предположить, что на Землю могли быть случайнозанесены метеоритом несколько типов клеток, создавших устойчивый замкнутый круговорот веществ, то метеорит должен был содержать не 3-4 вида клеток, а хотя бы втрое больше, чтобы в этойслучайной выборке обязательно присутствовали все необходимые типы. При этом все виды должны были бы иметь одинаковую биохимию, у них должны были бы совпадать и хиральности молекул, и наборы аминокислот, и т.д.
Если клетки случайно попали к нам из мира, где существовали разные ветви жизни (разные варианты биохимии), то невозможно, чтобы десяток принесенных метеоритами видов клеток случайно имел одинаковую биохимию. В случайной выборке обязательно должны были бы присутствовать клетки, относящиеся к иным ветвям Жизни, с иной биохимией.
Да, отдельные типы клеток с иной биохимией не могли идеально включиться в круговорот веществ. Но это не значит, что они должны были полностью погибнуть. Часть рассеянных по Океану клеток имела возможность продолжить существование. В таком случае они, вероятно, были бы обнаружены, как в метеорите Мурчисон выявлены не используемые земными организмами аминокислоты 2-метилаланин и саркозин.
То, что клетки с иной биохимией до сих пор не найдены, косвенно свидетельствует против случайного занесения Жизни на Землю с помощью метеоритов. Во всяком случае, против занесения её с планеты, имеющей формы Жизни с разной биохимией.
Пытаясь разобраться в филогении
прокариот, палеоботаники отмечают, что у них свободно сочетаются многие признаки. Поэтому система прокариот, по выражению Г.А. Заварзина, приобретает характермногомернойрешётки. Узлы решётки отвечают таксонам
, каждый из которых характеризуется определённым сочетанием признаков. Из-за того, что некоторые сочетания запрещены, в решётке образуются пробелы. Но остающиеся участки решётки не образуют структуру, которую можно было бы интерпретировать как филогенетическое дерево. Построение системы прокариот на филогенетической
основе Г.А. Заварзин считает принципиально невозможным». Такая ситуация заставляет задуматься – не потому ли стало невозможным построение филогенетического
древа прокариот, что на Земле в начале биологической эволюции одновременно появились не единицы, а десятки (или сотни?) видов прокариотических клеток с одинаковой биохимией?! Не потому ли мы не видим их филогенетического родства, что эволюционные пути прокариот разошлись задолго до их прибытия на Землю? Если такое предположение справедливо, то разговоры о случайномзанесении Жизни на Землю можно полностью прекратить – слишком уж невероятно случайное падение на Землю с метеоритами десятков видов живых клеток с одинаковой биохимией!
В случае если Жизнь переносится в наиболее примитивных формах, эволюция получает максимальный простор. Появляется возможность наиболее разнообразного, оригинального и безудержного развития. Единственное сохраняющееся ограничение для всей эволюции – единый, общий вариант биохимии. Если инопланетяне, действительно, занесли клетки с одним типом биохимии, то они тем самым избавили Землю от хорошо знакомых им, надо думать, проблем, связанных с несовместимостью разных ветвей жизни. Скорее всего, они выбрали для переноса клетки именно с такой биохимией, какой обладали в то время сами.
§1.3 Теория Ю. А. Колясникова.
На сегодня очевидно, что рождение Жизни могло произойти только в жидкой воде. Однако ни один исследователь не обратился к этому универсальному растворителю как к возможному, и, быть может, основному участнику кодирования белков в ДНК, а тем более как к главному дирижеру буквально всех процессов в нашем организме. Скорее всего, гипнотизирующая простота химического состава воды не давала даже повода к такой мысли. Но, исключив воду из рассмотрения возможных претендентов на матрицу синтеза предбиологической органики, исследователи за сорок лет безуспешных поисков были вынуждены обратиться к очень популярным ныне среди космобиохимиков идеям внеземного происхождения жизни и направленной панспермии.
Однако при таком простом составе структура аномальной по всем свойствам воды настолько сложна, что не поддается всем самым современным методам исследования.
В 1990 году Ю. А. Колясников предложил оригинальную политетрамерную модель структуры воды, в которой роль молекул играют не Н2
О, а сверхсжатые водные тетрамеры Н8
О4
, соразмерные кремнекислородным тетраэдрам – элементарным “кирпичикам” литосферы.
Важным элементом модели является обнаруженная графически рацемичность воды, то есть наличие в ней зеркально-симметричных, по распределению внутренних Н- связей, тетрамеров. А как полагали еще П. Кюри и В. И. Вернадский, правизна - левизна живого вещества есть следствие диссиметрии среды, в данном случае – воды. Кроме того, в живом веществе, вода находится в связанном состоянии, когда тетрамеры «развязываются», образуя, правые и левые спиральные цепочки, в которых в связи с дипольностью молекул может быть записана некая информация.
Попробуем представить, что происходило на Земле 4 млрд. лет назад.
Сейчас наука возвращается к взглядам Р. Декарта об изначально горячей Земле. Именно с таких позиций пытались представить возникновение Жизни, или биопоэз, Дж. Холдейн, А. И. Опарин, Дж. Бернал и другие, но без конкретного сценария образования самого первородного океана.
В своей теории (или сценарии), автор пытается восполнить этот пробел.
По мере остывания планеты ее плотный облачный покров, подобный венерианскому, опускался все ниже и ниже. Из облаков проливались горячие ливни, но поначалу они выкипали на лету. Наконец дождевые капли стали достигать поверхности, но они падали и тут же вскипали. Это продолжалось достаточно долго, вплоть до того момента, когда одна из них не выкипела целиком, упав, следовательно, на максимально охлажденную точку горячей протокоры, с которой и начался стремительный рост гидросферы. В составе тонкой пленки первородного бульона имелись все нужные для синтеза предбиологической органики компоненты, хотя сама вода была тогда в дефиците.
В той первой капле в подкипящем состоянии (250 – 200 о
С и 50 атм.) первая тетрамерная спиральная цепочка связанной с жестким силикатным субстратом воды, совершенно случайно оказалось левовращающей (с той же вероятностью она могла быть и правовращающей). На ней была синтезирована первая аминокислота, которая уже не случайно получилась левовращающей. К первой тетрамерной цепочке присоединилась вторая, тоже левая, как и синтезированная на ней аминокислота, то есть все последующие аминокислоты становились левовращающими. И так последовательно заработал водно-матричный механизм синтеза сразу хирально-чистой аминокислотной органики.
Однако одновременно с левовращающими «разворачивались» и правовращающие тетрамерные цепочки связанной воды, на которых позднее начался синтез менее термостойких сахаров – основы нуклеиновых кислот. Такой синхронный синтез полипептидов и полинуклеотидов неумолимо вел к образованию сложных нуклеопротеидных комплексов с записью в их примитивной РНК однозначного генетического кода.
Итак, главным фактором хирального очищения органики и возникновения генетического кода живого вещества Земли могло быть лишь одно из необычайных свойств воды, а именно ее собственная рацемичность. Следует отметить также, что в принятых сейчас сценариях биопоэза синтез рацемичной предбиологической органики происходил в уже существовавшем океане, в котором позднее произошла хиральная катастрофа.
В интерпретации же Ю. А. Колясникова все началось с самой первой капли, и далее шла стремительная экспансия бульонной пленки с синтезом в ней сразу хирально-чистой органики, что исключает маловероятную хиральную катастрофу. В результате на поверхности планеты образовался первобытный Солярис, огромная сеть бульонной пленки состоящей из предбиологического органического вещества.
С появлением в первом вулканическом катаклизме 4 млрд. лет назад на поверхности планеты свободной воды, уцелевшие фрагменты первобытного Соляриса дезинтегрировались в плазмиды, прионы и наноразмерные нуклеопротеидные комплексы. Избегая прямого контакта с чуждой им объемной политетрамерной водой, последние сворачивались, формируя белковые капсулы с РНК внутри них. Следовательно, вторым этапом биопоэза можно считать образование в первичной гидросфере бесчисленного множества разнообразных протовирусов, вироидов и прочих.
Позднее появились мембраны разного состава, а на их основе возникли первые клетки как вполне автономные истинно живые системы. Но и те, и другие продолжали использовать в своей жизнедеятельности первичную водную матрицу, обеспечивающую ускоренный синтез их биополимеров.
§1.4
Теория У. Мартина и М. Рассела
Уильям Мартин из Университета Генриха Гейне в Дюссельдорфе, Германия, и Майкл Рассел из Центра изучения окружающей среды, Университет Шотландии, Глазго, Великобритания утверждают, что первые живые организмы на Земле могли появиться внутри камней, выстилающих дно океана.
Более 4 миллиардов лет назад крошечные полости внутри минералов могли выступить в роли клеток.
Ключевой момент в этой теории – отложения сульфида железа (FeS). В горячих источниках на морском дне это соединение образует «соты» с ячейками шириной в несколько сотых миллиметра. Как считают Мартин и Рассел, эти ячейки – идеальное место для возникновения жизни По сравнению с другими гипотезами возникновения жизни на Земле, теория Мартина и Рассела уникальна тем, что она предполагают, что возникновение клетки предшествовало возникновению белков и самореплицирующихся молекул. С притоком горячей воды в ячейки попадают ионы аммония (NH4
+
) и монооксид углерода (CO), и сульфид железа выступает в роли одного из катализаторов синтеза органических веществ из неорганики. Простые соединения концентрировались в «камерах» из сульфида железа, что могло привести к возникновению сложных молекул – белков и нуклеиновых кислот.
Форд Дулитл из канадского университета Далхаузи, Галифакс считает данную теорию красивой и практически всеобъемлющей. Другие ученые согласны, что ячейки сульфида железа вполне могут быть «инкубаторами» первичных жизненных форм, однако указывают на «недостающее звено» между простыми органическими соединениями и химией живых существ. Так, Пьер Луиджи Люизи из Федерального института технологий в Цюрихе, Швейцария считает, что без объяснения происхождения ферментов все вышесказанное останется голой теорией.
Мартин и Рассел предположили, что живые организмы покинули каменные ячейки, когда научились сами строить клеточную стенку. Поэтому они выдвинули довольно спорное предположение о том, что жизнь на Земле возникала дважды. Об этом, по их мнению, свидетельствует большая разница в строении клеточной стенки у двух основных царств примитивных прокариот – бактерий и архебактерий.
С этим согласны далеко не все. Например, Томас Кавалье-Смит из Оксфордского университета в Великобритании говорит, что у бактерий и архебактерий есть сотни гомологичных генов, а также множество сходных признаков, таких как, скажем, способ встраивания белков в мембрану.
Мартин в ответ утверждает, что из-за способности бактерий обмениваться ДНК сейчас нам трудно установить последовательность событий только на основе генетики. Он предполагает, что выход обоих царств из каменных ячеек произошел около 3,8 миллиардов лет назад, в то время как самые древние ископаемые образцы, бесспорно свидетельствующие о наличии бактерий на Земле, относятся к периоду около 2,5 миллиардов лет назад, хотя некоторые исследователи говорят о возникновения жизни еще 3,5 миллиарда лет назад.
§1.5 Гипотеза Сванте Аррениуса
Даже если считать, что путь от неживой материи к прокариотической
клетке не сложнее, чем последующее формирование клеточного ядра (что далеко не так), а скорость эволюции неизменна (в действительности она постоянно нарастает), то и тогда создание первой клетки должно было бы длиться хотя бы столько, сколько длилось формирование ядра (т.е. 2,0¸2,4 млрд. лет). Как же воспринять практическое отсутствие этого промежутка времени, отсутствиестоль принципиально важного предбиологического этапа эволюции,этапа подхода к появлению Жизни?
Противоречие в сроках, как и противоречия однотипной биохимии, касаются ранних этапов биологической эволюции, не оставивших ясных следов в палеонтологической летописи. Это требует более внимательного и критического рассмотрения проблемы возникновения жизни на Земле в целом.
В бывшем СССР официальной наукой признавалась только „материалистическая теория происхождения жизни” А.И. Опарина (1922 г.) и примыкающие к ней работы. Речь шла о развитии клеток из так называемых коацерватных капель в водах Первичного океана. За рубежом же ещё ранее была высказана гипотеза С.
Аррениуса
о случайном занесении на Землю жизни из космоса, получившая название гипотезы панспермии.
Обращаясь к истории представлений о панспермии, можно отметить, что идея о существовании внеземной жизни высказывалась не только в религиозных учениях, в священной книге индусов Веде, в Авесте персов, в учении Заратустры, но и в трудах философов древнего мира. По учению о панспермии древнегреческого мыслителя Анаксагора, зародыши жизни распространены повсюду. Философская школа Эпикура учила, что существует множество обитаемых миров, подобных нашей Земле.
Один из сторонников этой школы Митридор сказал, что «считать Землю единственным населённым миром в беспредельном пространстве было бы такой же вопиющей нелепостью, как утверждать, что на громадном засеянном поле мог бы вырасти только один пшеничный колос».
Римский философ Лукреций Кар в поэме «О природе вещей» писал: «Весь этот видимый мир вовсе не единственный в природе, и мы должны верить, что в других областях пространства имеются другие земли с другими людьми и другими животными». Аналогичную мысль выражал Джордано Бруно. Он писал: «Существуют бесчисленные солнца, бесчисленные земли, которые кружатся вокруг своих солнц, подобно тому, как наши 7 планет кружатся вокруг нашего Солнца... На этих мирах обитают живые существа».
Вопреки старинному латинскому звучанию фамилии, шведский учёный Сванте Август Аррениус (1859-1927) относился к наиболее передовым мыслителям рубежа столетий. Он стал одним из основателей физической химии, автором теории электролитической диссоциации и основного уравнения химической кинетики, трудов по астрономии, астрофизике и биологии, лауреатом Нобелевской премии, иностранным членом-корреспондентом Петербургской Академии Наук, а позже – иностранным почётным членом Академии Наук СССР.
Кроме Аррениуса, в XIX и первой четверти XX ст. сторонниками учения о панспермии были выдающиеся зарубежные учёные Ф. Кон, Ю. Либих, Г. Гельмгольц, В. Томсон. В более позднее время мысль о возможности переноса спор бактерий через космическое пространство поддерживал американский учёный К. Саган. На территории бывшего СССР сторонниками учения о панспермии были С.П. Костычев, П.П. Лазарев, Л.С. Берг.
§1.6
Гипотеза А.И Опарина.
Пик исследований А. И. Опарина и его соавторов приходился на 50-60-е годы, хотя его книга «Происхождение жизни» была опубликована еще в 1924 году.
Появление жизни он рассматривал как единый естественный процесс, который состоял из протекавшей в условиях ранней Земли первоначальной химической эволюции, перешедшей постепенно на качественно новый уровень - биохимическую эволюцию.
С самого начала этот процесс был связан с геологической эволюцией. В настоящее время принято считать, что возраст нашей планеты составляет примерно 4,3 млрд лет. В далеком прошлом Земля была очень горячей (4000-8000 °С). По мере остывания образовывалась земная кора, а из воды, аммиака, двуокиси углерода и метана - атмосфера. Такая атмосфера называется «восстановительной», поскольку не содержит свободного кислорода. При падении температуры на поверхности Земли ниже 1000
C образовались первичные водоемы. Под действием электрических разрядов, тепловой энергии, ультрафиолетовых лучей на газовые смеси происходил синтез органических веществ-мономеров, которые локально накапливались и соединялись друг с другом, образуя полимеры. Можно допустить, что тогда же одновременно, с полимеризацией шло образование надмолекулярных комплексов-мембран.
По однотипным правилам синтезировались в «первичном бульоне» гидросферы Земли полимеры всех типов: аминокислоты, полисахариды, жирные кислоты, нуклеиновые кислоты, смолы, эфирные масла и др. Это предположение было проверено экспериментально в 1953 году на установке Стэнли Миллера, которому удалось получить многие вещества, имеющие важное биологическое значение, в том числе ряд аминокислот, аденин и простые сахара. Позднее в сходном эксперименте были синтезированы нуклеотидные цепи длиной в шесть мономерных единиц (простые нуклеиновые кислоты).
Органические вещества скапливались в сравнительно неглубоких водоемах, прогреваемых Солнцем. Солнечное излучение доносило до поверхности Земли ультрафиолетовые лучи, которые в наше время сдерживаются озоновым слоем атмосферы. Так энергией обеспечивалось протекание химических реакций между органическими соединениями и синтез полимеров.
Первичные клетки предположительно возникли при помощи молекул жиров (липидов).
Молекулы воды, смачивая только гидрофильные концы молекул жиров, ставили их как бы «на голову», гидрофобными концами вверх. Таким способом создавался комплекс упорядоченных молекул жиров, которые за счет прибавления к ним новых молекул постепенно отграничивали от всей окружающей среды некоторое пространство, которое и стало первичной клеткой, или коацерватом — пространственно обособившейся целостной системой. Коацерваты оказались способными поглощать из внешней среды различные органические вещества, что обеспечивало возможность первичного обмена веществ со средой.
Таким образом, первичная клеточная структура, по Опарину, представляла собой открытую химическую микроструктуру которая была наделена способностью к первичному обмену веществ, но еще не имела системы для передачи генетической информации на основе нуклеиновых кислот. Такие системы, черпающие из окружающей среды вещества и энергию, могут противостоять нарастанию энтропии и способствовать ее уменьшению в процессе своего роста и развития, что является характерным признаком всех живых систем.
Естественный отбор сохранял те системы, в которых были более совершенными функция обмена веществ и приспособленность организма в целом к существованию в данных условиях внешней среды.
В ходе естественного отбора выжили системы, имевшие особое строение белковых полимеров, что обусловило появление третьего качества живого - наследственности (специфичной формы передачи информации).
Концепция А. И. Опарина в научном мире весьма популярна. Сильной ее стороной является точное соответствие теории химической эволюции, согласно которой зарождение жизни - закономерный результат. Аргументом в пользу этой концепции служит возможность экспериментальной проверки ее основных положений в лабораторных условиях.
Слабой стороной концепции А. И. Опарина является допущение возможности самовоспроизведения коацерватных структур в отсутствие систем, обеспечивающих генетическое кодирование. В рамках концепции Опарина не решена главная проблема - о движущих силах саморазвития химических систем и перехода от химической эволюции к биологической, о причине таинственного скачка от неживой материи к живой.
Глава
II.
Опытно – экспериментальная часть
§2.1 Абиотический синтез биомономеров
Синтез аминокислот при действии электрических разрядов в газовой смеси, имитирующей возможный состав примитивной земной атмосферы, был осуществлен еще в 1953 г в широко известных в настоящее время опытах Миллера. Использованный для этой цели прибор Миллера изображён на рис 1. Он состоит из большого круглого сосуда, в котором находится исходная смесь газов и в котором производится электрический разряд, а также малой колбы с кипящей водой - в ней скапливаются получающиеся продукты.
При действии искрового или тихого разряда на смесь СН4
, NH3
, H2
и паров воды при постоянной (в течение недели) циркуляции смеси в малой колбе были обнаружены глицин, α-аланин, α -аминомасляная и α -аминоиз масляная кислоты, β-аланин, аспарагиновая и глутаминовая кислоты, саркозин и N-СН2
-аланин. Промежуточными продуктами этой реакции являлись альдегиды HCN.
Рис. 1. Прибор Миллера для синтеза органических соединений в восстановительной атмосфере под действием искровых разрядов
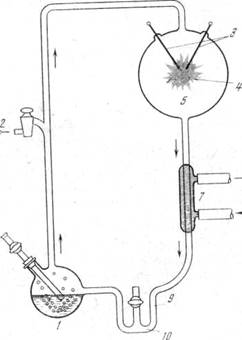
1 — колба с кипящей водой; 2 — к вакуумному насосу; 3 — электроды;
4 — искровой разряд; 5 — смесь газов (СН4,
NH3
, Н2
О, Н2
); 6 — выход воды из холодильника; 7 — холодильник; 8 — подача воды в холодильник; 9 — вода, содержащая
органические соединения; 10 — ловушка
Данные Миллера были подтверждены в работе Т. Е. Павловской и А. Г. Пасынского, в которой применялась несколько другая аппаратура, а избыточное количество водорода заменялось на СО. В последующих работах указанных авторов было обосновано положение, согласно которому любая реакция, приводящая к образованию альдегидов и HCN, обязательна должна была способствовать накоплению α -аминокислот в гидросфере первичной Земли.
Эбельсон, используя более сложную исходную смесь газов, включающую наряду с СН4
, NH3
и Н2
О также) СО, СО2
и N2
, получил аналогичную Миллеру смесь аминокислот.
Оро, используя С2
- и С3
-углеводороды и концентрированный NH4
OH, дополнил указанный список синтезированных в искровом разряде аминокислот лейцином изолейцином и валином.
Гроссенбахер, пропуская искровой разряд в специально сконструированном приборе в течение 100 - 200 час. через смесь NH3
, CH4
и Н2
, получил в водном ; растворе смесь аминокислот (соотношение в молях): аспарагиновая кислота — 2, треонин — 4, серии—14, глицин— 16, аланин—14, лизин — 4, лейцин — 2, изолейцин — 2 и глутаминовая кислота — 1. Кроме того, он обнаружил пептиды, состоящие из глицина и аланина (5:1), а также глицина и изолейцина. К концу опыта продукты полимеризации аминокислот выделялись из раствора в виде мелких многомолекулярных сфер или капель.
Менее многочисленны исследования по синтезу аминокислот при воздействии на смесь примитивных газов ионизирующими излучениями. Здесь можно назвать опыты Дозе и Раевского, обнаруживших образование кислых и нейтральных аминокислот при действии рентгеновых лучей на газовую смесь СН4
, NH3
, H2
, CO2
, N2
и Н2
О. При облучении β-лучами от линейного ускорителя (2 Мэв) растворов ацетата аммония Гассельстромом, Генри и Мурром были получены глицин и аспарагиновая кислота, а при Х-облучении 60
Со (5*108
р) твердого карбоната аммония Пашке, Чанг и Янг доказали образование глицина и аланина. Пальм и Кальвин, действуя на смесь метана, аммиака, водорода и паров воды пучком быстрых электронов (5 Мэв при дозе 1010
эрг), синтезировали глицин и аспарагиновую кислоту.
Образование аминокислот при простом нагревании исходных растворов было впервые обнаружено Фоксом, Джонсоном и Вегодским. Оро с сотрудниками показал, что ряд аминокислот (глицин, аланин, серии, аспарагиновая кислота, треонин) получается просто при нагревании в течение 40—60 час. водных смесей формальдегида и гидроксиламина при 80—100° С или даже при более низких температурах. Аналогичные результаты были получены Левом, Рисом и Маркхемом, которые нагревали 1,5 М водный раствор NH3
и HCN при 90° в течение 18 час. и обнаружили при этом значительное количество аминокислот, частично входящих в пептиды и освобождающихся после гидролиза продуктов реакции (в том числе, кроме глицина, аланина и аспарагиновой кислоты, они получили также лейцин, изолейцин, серии, треонин и глутаминовую кислоту).
Но, конечно, основное значение для первичного абиогенного синтеза органических веществ имела все же энергия коротковолновых ультрафиолетовых лучей, легко достигавшая земной поверхности вследствие отсутствия в доактуалистическую эпоху озонового экрана. Синтез аминокислот и других органических соединений при экспериментальной имитации такого рода условий особенно широко представлен сейчас в мировой литературе.
Т. Е. Павловская и А. Г. Пасынский при облучении водных растворов, содержащих 2,5% формальдегида и до 1,5% хлористого или азотнокислого аммония, ультрафиолетовыми лучами от лампы ПРК-2 в течение 20 час. установили образование следующих аминокислот: серина, глицина, глутаминовой кислоты, аланина, валина, фенилаланина и основных аминокислот — лизина, орнитина и аргинина.
В присутствии адсорбентов (бентонита, каолинита, лимонита и оптического кварца) были найдены те же аминокислоты (за исключением основных) и, кроме того, изолейцин. В последующей работе было показано образование треонина, а при замене формальдегида ацетальдегидом — аргинина и тирозина. Аналогичные результаты были получены другими авторами при облучении коротковолновым ультрафиолетовым светом смеси примитивных газов и их ближайших производных. Нужно отметить, что как при электрических разрядах, так и при освещении ультрафиолетовыми лучами в использованных для синтеза примитивных смесях образуются не только аминокислоты, но также органические кислоты и альдегиды, амины и амиды, в частности мочивина. Вместе с тем в этих условиях происходит полимеризция мономерных молекул, приводящая к образованию более сложных соединений.
§2.2 Абиотический синтез полимеров
Следующим за образованием мономеров шагом эволюции на пути к возникновению жизни являлось их объединение в полимеры, в частности в соединения, аналогичные белкам и нуклеиновым кислотам, играющим исключительно важную роль в организации живых существ.
Принципиальная возможность такой далеко идущей абиогенной полимеризации органических веществ подтверждается рядом опытов, где в качестве мономеров использовались в первую очередь аминокислоты и родственные им соединения.
Здесь прежде всего нужно упомянуть об исследованиях Акабюри, который еще в 1955 г. указывал на возможность в условиях примитивной Земли абиогенного синтеза протобелков (т. е. аминокислотных полимеров со случайным расположением аминокислотных остатков в полипептидной цепи). Вскоре он доказал эту возможность экспериментально, получая из формальдегида, аммиака и HCN аминоацетонитрил, поликонденсат которого адсорбировался на каолине с образованием полиглицина. Затем в полиглицин можно вводить боковые Цепи путем реакции с альдегидами или с ненасыщенными углеводородами. Таким путем Акабюри удалось получить полиглицин и его аналоги с молекулярным весом приблизительно равным
15 000.
Фокс и его сотрудники, имитируя вулканические условия, подвергали смесь 18 аминокислот нагреванию при 170° С на куске лавы в течение 6 час. Для того чтобы расплавить смесь до ее обугливания, бралось избыточное количество дикарбоновых аминокислот. В других случаях нагревание осуществлялось в присутствии полифосфатов, что позволяло снизить температуру нагревания до 100° С.
При таком температурном воздействии образуется янтарного цвета полимер с молекулярным весом в несколько тысяч (от 5 000 до 10 000), содержащий все взятые в опыт типичные для белка аминокислоты (потерявшие, однако, свою оптическую активность). Этот полимер был назван Фоксом протеиноидом за его сходство по ряду признаков с белками.
При дальнейших исследованиях были выявлены два очень интересных свойства протеиноидов: во-первых, то, что они обладают известной внутримолекулярной упорядоченностью, повторяющейся последовательностью аминокислотных остатков, и, во-вторых, имеют хотя и слабую, но вполне экспериментально обнаруживаемую ферментативную активность (катализ гидролиза, декарбоксилирования, аминирования и дезамннирования).
Из многочисленных работ по синтезу полипептидов в водной среде следует назвать следующие: поликонденсацию глицина при его нагревании в водном растворе аммония, при γ-облучении; ацетилглицин при термальной обработке растворов цианистого аммония, при нагревании смеси HCN, NH3
, при полимеризации аминоацетонитрила, цианамида и дицианамида.
Очень важным вопросом при синтезе полипептидов и белковоподобных соединений является та последовательность, с которой аминокислотные остатки связываются между собой в полипептидные цепи. При современном биологическом синтезе белков эта характерная для каждого индивидуального белка последовательность (первичная структура) определяется нуклеиновым кодом, но, как указывалось выше, уже при термальном синтезе Фокса, в отсутствие нуклеиновых кислот создался некоторый постоянный порядок соседствования аминокислот.
Штейнман присоединял первую в ряду аминокислоту к гранулам высокомолекулярного полимера (хлорметилированного полистирола) и таким образом получал смолу с присоединенной к ней одной аминокислотой. Затем он исследовал относительную эффективность присоединения к ней ряда других аминокислот с незащищенной аминогруппой.
Экспериментально определяя выходы различных дипептидов, Штейнман сопоставлял эти результаты с частотой аминокислотных пар, определенных по «Атласу белковых структур» в современных белках, и обнаружил значительное совпадение. Кальвин в своей книге «Химическая эволюция» дает модель системы воспроизведения полипептида без матрицы, на основании контроля со стороны растущего конца. Интересно сопоставить эти высказывания Кальвина с замечательным синтезом полипептида (грамицидина С), осуществленным Липманном.
В более сложных и довольно трудновоспроизводимых в природе условиях были синтезированы и полимеры нуклеотидов, образующие цепочки, аналогичные молекулам нуклеиновых кислот современных организмов.
Замечательные работы Корнберга и другие показали возможность синтеза нуклеиновых кислот invitro. Однако эти биосинтезы могут проходить только в присутствии соответствующего специфического фермента, и поэтому они не могут служить моделью для абиогенных процессов на примитивной Земле.
Некоторая вероятность такого синтеза была впервые намечена Шраммом в его работах с полифосфорным эфиром. В дальнейшем было показано, что цитидинфосфат может конденсироваться в олигонуклеотиды с помощью полифосфорной кислоты. В модельных опытах, более приближающихся к условиям первичной Земли, была показана способность карбодиимида стимулировать образование ди- и тринуклеотидов из смеси нуклеозидов и нуклеотидов в разведенных водных растворах (Оргель и сотр.).
Особую роль в процессах первичной полимеризации играют, по-видимому, фосфорные соединения. При температурах около 300° С и выше фосфорная кислота полностью конденсируется с образованием полифосфатов. Поэтому на поверхности примитивной Земли, с ее широко идущим локальным разогревом, полифосфаты и их органические соединения должны были образовываться в довольно широком масштабе.
В настоящее время неорганические полифосфаты обнаружены в значительных количествах у низших организмов: у бактерий, водорослей, грибов и некоторых простейших. Это линейные полимеры ортофосфорной кислоты с молекулярным весом 30 000—100 000. Из ряда объектов выделены также циклические тримета- и тетраметафосфаты.
Как было показано Н. С. Кулаевым и другими, неорганические полифосфаты играют существенную роль в энергетическом обмене низших организмов в противоположность высокоорганизованным живым существам, для которых они не являются характерными.
Все это позволяет думать, что на заре возникновения жизни на Земле неорганические полифосфаты вполне могли участвовать в образовании простейших механизмов сопряжения энергодающих и энергопотребляющих реакций, т. е. играть ту роль в предбиологическом и раннем биологическом обмене, которая сейчас осуществляется аденозинтрифосфорной кислотой.
Поннамперума, Саган и Маринер использовали этилметафосфат для модельных опытов по синтезу нуклеотидных компонентов. При этом необходимым источником энергии им служил ультрафиолетовый свет с длиной волны 2400—2900 А, который был вполне доступен примитивной земной поверхности. Синтезы велись в водных растворах аденина, аденозина, адениловой кислоты, рибозы и этилметафосфата. Смеси освещались ультрафиолетовым светом с длиной волны 2537 А при 40°С. При этом было установлено превращение аденина в аденозин, аденозина в аденозинмонофосфат (АМФ), АМФ в аденозиндифосфат (АДФ) и наконец АДФ в аденозинтрифосфат (АТФ). Таким образом, была доказана возможность абиотического синтеза в условиях доактуалистической земной поверхности аденозинтрифосфорной кислоты – этой основной «энергетической валюты» современных высших организмов.
Итак, широко развернувшиеся в настоящее время модельные опыты по абиотическому синтезу органических веществ в условиях, имитирующих примордиальные земные условия, позволяют нам с известной долей вероятности представить себе последовательный ход химической эволюции от простейших углеродистых соединений к высокополимерным веществам, из которых образовались те «организованные элементы» или примитивные организмы, остатки которых мы обнаруживаем в отложениях земной коры.
На этом этапе развития материи и произошел на нашей планете переход от химической эволюции к биологической, когда из гомогенного раствора органических веществ обособились индивидуальные целостные системы — пробионты, а затем и первичные живые существа.
Мы можем составить представление об этом переходе путем сопоставления современных данных сравнительной биохимии с лабораторными модельными опытами.
Заключение.
Зарождение жизни - точка отсчета для развития всего живого мира на Земле. Именно в этот момент начали функционировать фундаментальные законы существования живых организмов, которые с ходом поступательного развития жизни становились только более многоуровневыми и дифференцированными. Не поняв существа этих базисных законов, мы лишаем себя возможности осмыслить целый ряд важнейших аспектов в эволюционной биологии, цитологии, микробиологии, экологии, учении о биосфере и других науках, включая медицину и валеологию. Можно сказать, что понимание причины и механизма возникновения первичных живых форм является тем ключом, который облегчит проникновение в тайны существования растений, животных и человека, и поможет найти оптимальные подходы к их гармоничному, или органичному, сосуществованию. Именно поэтому «ориджинология» - наука о возникновении жизни, должна занять достойное место в ряду других наук.
Кроме познавательного аспекта, ориджинология имеет важнейшее практическое значение. В настоящее время на планете быстрыми темпами развиваются глобальные изменения, связанные, прежде всего с ухудшением состояния экосистем и здоровья населения планеты. Многие ученые сходятся во мнении, что главным резервом, на который человечество может опираться в своем дальнейшем существовании, является системный ресурс биосферы. Другим резервом, является системный ресурс самого человека, его огромные потенциальные возможности, которые могут развертываться в трудных ситуациях. В решении обоих этих проблем могут быть использованы знания, накопленные учеными в ходе исследований, посвященных возникновению жизни на Земле.
Сравнивая первые теории, выдвинутые несколько десятилетий назад Опариным, Холдейном и Берналом - первопроходцами в этой области исследований, - с положениями, достигнутыми к настоящему времени, мы видим, как из набора слабо обоснованных фактами гипотез, выросла современная теория, как неизмеримо увеличились наши знания. Мы знаем теперь слабые места исходных гипотетических построений. На многих примерах мы поняли, какие из этих построений надо отбросить, какие заслуживают сохранения. В результате отпали многие неясности, мы смогли точнее представить себе, как возникла жизнь.
Даже сейчас, не ожидая новых открытий, которые, несомненно, появятся в будущем, мы с высокой степенью вероятности можем утверждать, что жизнь развилась естественным путем из некоей преджизни, состоявшей из «органических» соединений, созданных в результате неорганических процессов.. эта теория хорошо согласуется с большим числом проверенных научных фактов и наблюдений, полученных специалистами в самых разных естественных науках. В этом ее преимущество перед более ранними гипотезами, в которых было немало противоречий и несуразиц, тем более если включать в их число постулаты о внеземном, а то и сверхъестественном происхождении жизни. Сейчас эти постулаты признаются не только недостоверными, но и просто излишними; наука их отбрасывает. Жизнь на Земле могла возникнуть естественным путем, и, судя по известным фактам, именно так она и возникла.
Список литературы
1. Камшилов М. М
. Эволюция биосферы. М.: «Наука», 1974. 57-69, 87, 90-91, 94-96, 117-120, 240с.
2. Колясников Ю. А.
К тайнам мироздания. Магадан: СВНЦ ДВО РАН, 1997. 225 с.
3. Компаниченко В. Н.
Возникновение жизни в глубинах гидротермальных систем. Хабаровск, 1996, 24-38, 98-100с.
4. Калвин В
. Химическая эволюция. М.: «Наука», 1971 – 10-14, 25-33, 45-49, 58-60 с.;
5. Опарин А
.И.
Материя – Жизнь – Интеллект. М.: «Наука», 1977, 5-6, 96-117 с.
6. Найдыш В.
М.
Концепции современного естествознания: Учеб. пособие. – М.: Гардарики, 2000. – 115-124с.
7. Мотылёва Л.
С., Скоробогатов В.А., Судариков А.М.
Концепции современного естествознания: Учебник для вузов – СПб.: Издательство Союз, 2000. – 228-237с
|
