и способы ее предсказания для трансмембранных белков
С высоким разрешением удалось установить структуру только одного класса мембранных белков – реакционного центра бактерий, однако и в этом случае положение белка относительно липидного бислоя не определено однозначно. Распространять принципы его организации на другие мембранные белки следует с осторожностью. Некоторую ясность может внести использование термодинамических принципов, а также учет того факта, что основная масса экспериментальных данных согласуется с предположением о высоком содержании в мембранных белках а-спиралей. Термодинамические факторы налагают определенные ограничения на то, какого типа белково-липидные структуры могут быть стабильными.
Мембранные белки – это амфифильные соединения
Любые мембранные белки, непосредственно контактирующие с гидрофобной сердцевиной липидного бислоя, должны быть амфифильными. Те участки полипептида, которые экспонированы в растворитель, скорее всего обогащены полярными и ионизируемыми аминокислотными остатками, а остатки, контактирующие с лилидными углеводородными цепями, должны быть в основном неполярными. Все это логически следует из энергетических принципов, рассмотренных в разд. 2.3.1. Заряженные или полярные аминокислоты вообще говоря могут находиться внутри бислоя, однако на это налагаются определенные ограничения.
Рассмотрим три уровня амфифильных структур в мембранных белках: первичную, вторичную и третичную амфифильность.
1. Первичные амфифильные структуры содержат протяженный участок из преимущественно неполярных аминокислотных остатков, длина которого достаточна для пересечения бислоя. Такие структуры выявлены как в реакционном центре, так и в бактериородопсине. У этих белков все пронизывающие мембрану элементы являются а-спиральными. а-Спиральная структура предпочтительна потому, что при этом образуются все водородные связи, в которых могут участвовать атомы водорода полипептидного каркаса. Альтернативная структура, у которой отсутствует одна из водородных связей, менее стабильна примерно на 5 ккал / моль. Все это позволяет высказать предположение о том, что поворот полипептидной цепи внутри мембраны маловероятен. В местах поворота от трех до пяти аминокислотных остатков не смогли бы образовать водородные связи, и это дестабилизировало бы структуру примерно на 15–20 ккал / моль. В глобулярных, водорастворимых белках области поворота располагаются преимущественно на поверхности белковой глобулы, где амидные группы могут образовывать водородные связи с водой; по-видимому, в молекулах мембранных белков повороты также будут происходить лишь в экспонированных в воду участках.
Реклама

Не исключено, что 3-слой тоже может образовывать трансмембранные элементы, имеющие, например, форму /J-цилиндров, как в случае порина. Требования, предъявляемые к образованию водородных связей атомами водорода полипептидного остова в подобных структурах, могут быть удовлетворены, но лишь при условии взаимодействия между отдельными ^-цепями. Как такая структура может встраиваться в мембрану, не совсем ясно, а ограничения, налагаемые механизмами сборки мембранных белков, вообще неизвестны.
2. Вторичные амфифильные структуры. В таких структурах гидрофобные остатки периодически встречаются вдоль цепи, и при укладке полипептида в определенную вторичную структуру они образуют сплошную поверхность. Периодичность некоторых элементов вторичной структуры указана в табл. 1. В качестве примера белков, в которых вторичные амфифильные структуры, по-видимому, играют важную роль, можно привести порины. В них полярные и неполярные аминокислотные остатки в каждой из /3-цепей чередуются. Все полярные остатки находятся на одной стороне складчатого слоя, выстилая наполненную водой пору. Заметим, что все сказанное о порине носит гипотетический характер.
Таблица 1. Параметры вторичной структуры
| Структура |
Периодичность или число остатков на виток |
Расстояние
между остатками, А
|
Радиус или ширина, А |
| Неизогнутая |
2,0 |
3,2–3,4 |
0,9–1,1 |
| (З-цепь |
| Изогнутая /З-цепь |
2,3 |
3,3 |
1,0 |
| Зю-Спираль |
3,0 |
2,0 |
1,9 |
| а-Спираль |
3,6 |
1,5 |
2,3 |
а-спираль, в которой гидрофобные остатки встречаются через каждую вторую или третью мономерную единицу, должна иметь гидрофобную и полярную поверхности. Подобные структуры часто представляют в виде спирального кольца с указанием боковых цепей – так, как это сделано на рис. Вторичные амфифильные структуры могут возникать в ситуациях, схематически показанных на рис. ЗЛО.
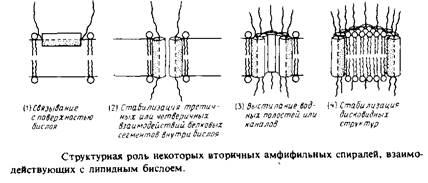
а. Поверхностно-активные сегменты белка; одна сторона спирали взаимодействует с гидрофобной областью липидного бислоя, а другая контактирует с водной фазой и полярной областью бислоя. Амфифильные а-спирали способны образовывать многие пептидные гормоны, а также разрушающие мембрану пептиды, например меллитин.
б. Трансмембранные элементы; неполярная поверхность спирали обращена к липидной фазе, а полярная выстилает водный канал, пронизывающий бислой. Это весьма распространенная модель, построенная главным образом исходя из результатов исследования никотинового ацетилхолинового рецептора, функционирующего как химически возбудимый канал. Однако основанные на экспериментальных данных выводы о том, что мембрану пронизывает именно амфифильная спираль, вызвали возражения. Такой наполненный водой канал, как в порине, может образовать и амфифильная 3-цепь.
Реклама

в. Трансмембранные элементы; неполярная часть поверхности контактирует с липидами, а полярные группы – с полярными группами других трансмембранных элементов. Именно этот принцип лежит в основе «вывернутых» структур, каким предположительно является бактериородопсин. Полярные взаимодействия между амфифильными спиралями в принципе могли бы стабилизировать взаимодействия между субъединицами в олигомерных белках.
3. Третичные амфифильные структуры. Об их существовании можно говорить только предположительно. Их гидрофобная поверхность должна формироваться на уровне третичной структуры остатков, расположенных в самых разных участках полипептидной цепи. Подобные структуры могут быть характерны для белков, связывающихся с бислоем, но не имеющих четко выраженных гидрофобных доменов, определяемых по любому из указанных выше критериев. Возможным примером такого рода является а-лактальбумин.
Ионизируемые аминокислотные остатки в трансмембранных сегментах
Многие модели мембранных белков предполагают, что в их трансмембранных сегментах находятся ионизируемые остатки. Эти остатки, вероятно, играют важную функциональную и / или структурную роль. В некоторых случаях эта роль однозначно установлена: 1) остатки лизина в бактериородопсине и родопсине образуют шиффовы основания с простетической группой ретиналя, что необходимо для светового возбуждения молекулы; 2) остатки гистидина в полипептидах реакционного центра бактерий участвуют в связывании с фотосинтетическими пигментами; 3) заряженные остатки в лактозопермеазе из Е. coliучаствуют в осуществлении этим белком транспортных функций; возможно, эти остатки образуют сеть водородных связей внутри молекулы белка.
Перенос заряженных групп из воды в среду с низкой диэлектрической проницаемостью внутри мембраны энергетически очень невыгоден, и эти группы необходимо каким-либо образом стабилизировать. Неоднократно предполагалось, что для стабилизации достаточно образования ионных пар, и этот принцип использовался при построении трехмерной модели бактериородопсина. Однако расчеты показали, что свободная энергия переноса ионной пары из воды в среду с низкой диэлектрической проницаемостью тоже весьма велика. Для дальнейшей стабилизации необходимы дополнительные полярные взаимодействия, возможно, с участием других полярных групп или с помощью водородных связей.
В принципе даже одиночная заряженная группа внутри мембраны может стабилизироваться через взаимодействия с полярными группами и при участии водородных связей, эффективно делокализующих заряд. Можно привести несколько примеров изолированных, де-сольватированных ионов, стабилизированных за счет взанмодействий в водорастворимых белках. Аналогичные принципы, по-видимому, действуют в случае заряженных остатков трансмембранных сегментов интегральных белков.
Однако представляется более вероятным, что ионизируемые аминокислоты нейтрализуются внутри мембраны за счет протонирования или депротонирования. Свободная энергия нейтрализации заряженных аминокислот, по оценкам, составляет примерно 10–17 ккал / моль. В отсутствие специфических условий для полярных взаимодействий, стабилизирующих заряженный остаток в трансмембранном сегменте, он скорее всего будет нейтрализован.
Заряженные аминокислоты в сегментах, экспонированных в водную среду
Как мы уже говорили, заряженные остатки распределены между двумя сторонами реакционного центра бактерий асимметрично. Такая асимметрия характерна и для некоторых других внутренних мембранных белков бактерий. Так, основные остатки Lys и Arg в четыре раза чаще встречаются в тех соединяющих трансмембранные элементы участках, которые расположены на внутренней стороне мембраны, а не на наружной. Для кислых остатков Asp и Glu подобная тенденция не выявляется. Возможно, эта асимметрия связана с механизмом сборки мембранного белка, но как именно – неясно. Более того, неизвестно, можно ли обобщить это наблюдение и имеет ли оно какую-либо предсказательную ценность.
В глобулярных, водорастворимых белках остатки пролина редко находятся в серединной части а-спирали. По данным исследований 58 белков, содержащих 331 а-спираль, выявлено 30 таких случаев. В половине из них пролин располагался в местах повреждения спирали, а в остальных случаях находился в области искривления или нерегулярности структуры.
В то же время у бактериородопсина пролиновые остатки расположены в средней части трех из семи трансмембранных спиралей, а у родопсина – в пяти из семи таких спиралей. Подобная тенденция выявлена и для других трансмембранных сегментов интегральных белков, особенно транспортных. Значение этого феномена неизвестно. Следует отметить, однако, что из-за наличия циклической боковой цепи пролин не образует водородных связей с остатками, находящимися на предыдущем витке а-спирали. Это может способствовать формированию структур, в которых водородная связь образуется за счет специфичного взаимодействия с остатком, расположенным в другом пронизывающем мембрану участке. Подобное полярное взаимодействие внутри бислоя могло бы стабилизировать трехмерную структуру мембранных белков.
Способы идентификации первичных амфифильных структур
Однозначная структурная информация о мембранных белках получена лишь в нескольких случаях, но зато в распоряжении исследователей имеются обширные данные об аминокислотной последовательности, основанные на результатах секвенирования ДНК. Для идентификации трансмембранных а-спиралей, предположительно имеющих длину – 20 остатков и состоящих преимущественно из гидрофобных аминокислот, разработано несколько методов анализа аминокислотной последовательности. В основе каждого из них лежит расположение аминокислот в ряд в соответствии с неким параметром, который отражает вероятность обнаружения этого остатка в трансмембранном сегменте.
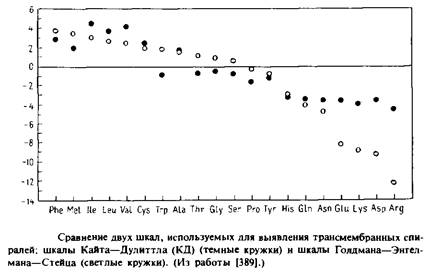
Существует два типа шкал. В одном случае аминокислоты классифицируют по их относительной полярности или «гидрофобности». Эти шкалы имеют термодинамическую природу и основаны на величине изменения свободной энергии при переносе аминокислоты из водного раствора в углеводородную среду. Однако число способов количественной оценки гидрофобности аминокислот весьма велико, и они не во всем согласуются между собой. Часто используются данные, относящиеся одновременно к более чем одной физической характеристике. Примером такого рода является шкала «гидропатии» Кайта и Дулиттла, основанная на данных о гидрофобности, измеряемой по потенциалу гидрации, а также о вероятности нахождения остатков внутри глобулы.
Шкала Голдмана, Энгелмана и Стейца основана на количественной оценке свободной энергии переноса а-спиралей из водной среды внутрь мембраны. На рис. сравнивается шкала Кайта – Дулиттла со шкалой Голдмана–Энгелмана–Стейца.
По оценка Энгелмана и др., изменение свободной энергии при внедрении в мембрану полианионной а-спирали длиной 20 остатков составляет 30 ккал / моль. Расчет основан на оценке площади поверхности спирали, экспонированной в растворитель. Вклад в энергию каждой боковой группы оценивали с учетом площади поверхности, экспонированной в водную среду внутри спирали. Учитывали также свободную энергию переноса в бислой полярных групп. Например, предполагали, что глутамин при переносе в бислой будет протонироваться и свободная энергия этого процесса составит 10,8 ккал / моль. Подобно этому, перенос гидроксилов будет «стоить» примерно 4,0 ккал / моль.
Все сказанное выше показывает, как выигрыш в энергии взаимодействий при переносе а-спирали внутрь бислоя может использоваться для «втягивания» в бислой полярных боковых групп. Например, один остаток аргинина может встроиться в бислой в составе неполярной трансмембранной спирали, если он депротонирован; для этого требуется 16,7 ккал / моль при рН 7,0. Суммарная свободная энергия переноса а-спирали по-прежнему останется отрицательной. Однако ситуация изменится, если в бислой понадобится встроить два аргининовых остатка или если аргинин будет положительно заряжен. Конечно, полярные остатки могут стабилизироваться внутри бислоя благодаря специфическим взаимодействиям, но реально учесть это при расчетах очень трудно. Например, боковые группы серина, цистеина и треонина могут образовывать водородные связи с полипептидным остовом, а кислые и основные остатки могут образовывать ионные пары; появление таких пар возможно, если эти остатки расположены через четыре или пять мономерных единиц друг от друга.
Второй тип шкал, который используется для классификации аминокислот, основан на данных о частоте, с которой аминокислоты действительно встречаются в пронизывающих мембрану сегментах.
При этом эмпирически учитывается гидрофобность, а также многие другие факторы, которые нельзя оценить количественно, как гидрофобность. Недостаток этого полуэмпирического подхода состоит в отсутствии точных данных о границах трансмембранных участков. Тем не менее подобные шкалы могут быть столь же полезны, как и шкалы, основанные на термодинамических параметрах. В качестве примера можно привести шкалу «склонности» к мембране Куна и Лейгха или шкалу «погруженности спирали в мембрану» Рао и Аргоса. Четыре наиболее гидрофобных остатка по шкале Голдмана–Энгелмана–Стейца являются также четырьмя остатками с наивысшим значением параметра по шкале Рао и Аргоса.
На рис. представлены профили трех разных мембранных белков, полученные с использованием различных шкал. При построении этих профилей учитываются средние значения чисел на шкалах, приписываемые каждой аминокислоте в пределах выбранного «окна»; это среднее откладывается относительно номера остатка в полипептиде. Например, если «окно» составляет 19 остатков, значение, приписанное положению 40, будет средним числом на шкале для всех аминокислот от 31 до 49 включительно. Значение, приписанное положению 41, будет средним для остатков с 32 по 50 и т.д. Пики на профиле соответствуют гидрофобным участкам или тем участкам, которые с большей вероятностью образуют трансмембранные спирали. Для построения профиля важен размер окна; большинство кривых на рис. были построены при размере окна в 19 остатков.
Попытаемся проинтерпретировать построенные профили. По шкале Голдмана–Энгелмана–Стейца пики при значениях, близких к нулю, соответствуют трансмембранным спиралям. Значение 1,25 по шкале Кайта–Дулиттла является наименьшим значением, отвечающим известной трансмембранной спирали в L-субъе-динице реакционного центра R. viridis. Во всех трех случаях, представленных на рис. 3.12, профили для субъединиц реакционного центра сходны.
На рис. приведены два профиля для цитохрома Р450 из микросом. Этот белок был выбран потому, что данные о его первичной структуре позволяют высказать предположение о наличии у него восьми трансмембранных спиралей. Однако имеющиеся экспериментальные данные указывают на существование только одного N-koh-цевого якоря в мембране. Как профиль Кайта–Дулиттла, так и профиль Голдмана–Энгелмана–Стейца выявляют N-конце-вой участок, но они указывают и на наличие одного или более дополнительных трансмембранных сегментов, что не соответствует действительности. Отметим, что многие из построенных моделей мембранных белков, которые основываются лишь на данных об аминокислотной последовательности, могут быть некорректными.
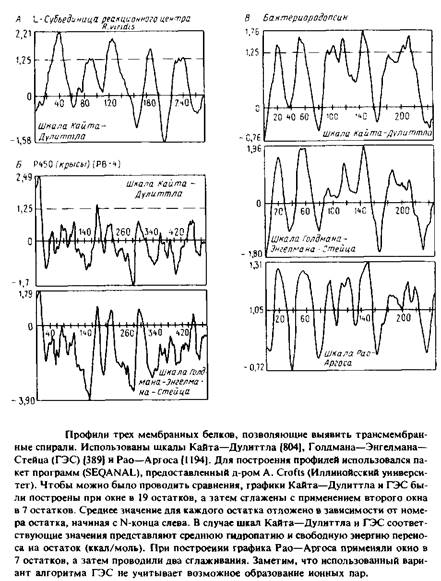
На рис. приведены три профиля для бактериородопсина. Несмотря на их сходство, видны различия в форме пиков, отвечающих семи трансмембранным сегментам. Алгоритм Голдмана–Энгелма-на–Стейца не учитывает стабилизирующего эффекта, связанного с образованием ионной пары из близко расположенных заряженных остатков в пределах одной спирали. С учетом этого фактора разделение между двумя последними спиралями становится более четким.
Одна из проблем, с которыми сталкивается применение всех описанных выше алгоритмов, состоит в том, чтобы исключить гидрофобные сегменты в известных глобулярных белках, не являющиеся трансмембранными, но располагающиеся внутри белка. Однако, когда мы ищем достаточно протяженные участки, эта проблема не возникает.
Отметим, что алгоритмы, используемые для выявления а-спиральных структур в растворимых глобулярных белках, например алгоритм Чоу–Фасмана, непригодны для обнаружения трансмембранных элементов. Эти алгоритмы неприменимы для описания структуры неглобулярных участков, какими являются сегменты, расположенные внутри бислоя.
Алгоритмы, предназначенные для идентификации трансмембранных участков, нельзя использовать в случае сегментов, являющихся вторичными амфифильными структурами или пересекающих мембрану в виде /3-слоя. В первом случае этот участок исключается из рассмотрения из-за наличия в нем полярных остатков, а во втором трансмембранный сегмент оказывается слишком коротким, поскольку для пересечения бислоя необходимо лишь 10–12 аминокислотных остатков в составе /3-структуры. Некоторые алгоритмы предназначались скорее для выявления ^-поворотов, а не самих трансмембранных элементов. Хотя это позволяет избежать некоторых проблем, связанных с выделением различных классов трансмембранных элементов, неясно, насколько приемлемыми они окажутся при более широком их применении.
Способы идентификации вторичных амфифильных структур
Разработано несколько подходов к выявлению вторичной амфифильности или асимметрии в распределении гидрофобных остатков в сегментах полипептидной цепи. Достаточно часто а-спирали и /3-слои в глобулярных белках характеризуются периодичностью в распределении гидрофобных остатков. Использование спирального кольца как качественного показателя не всегда оправданно, необходимы более количественные подходы. Основной из них – это определение периодичности в распределении гидрофобных остатков с помощью методов фурье-преобразования. В качестве примера можно привести гидрофобный момент.
1. Гидрофобный момент. Этот параметр был предложен Эйзенбергом и др. Он определяется как
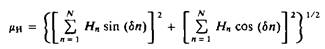
и представляет собой некую векторную сумму гидрофобности остатков в сегменте из N элементов. Гидрофобность каждого остатка представлена в виде вектора, который характеризуется углом, образуемым боковой цепью и осью полипептидного остова. Для а-спирали 6 = 100°. На рис. 3.9, Б «векторы» гидрофобности представлены в проекции на плоскость спирального кольца, и гидрофобный момент равен их векторной сумме. Гидрофильный остаток представляется вектором с отрицательной направленностью. Для случайной последовательности значение ци в силу случайного распределения гидрофобных остатков будет очень мало. В то же время в пептиде меллитине гидрофобные остатки расположены с одной стороны структуры, а полярные – с другой. Численное значение гидрофобного момента приписывается аминокислоте, находящейся в центре анализируемого сегмента. Следовательно, можно «просканировать» последовательность и приписать каждому положению среднюю гидрофобность, а также найти ^н-
Эйзенберг и др. проанализировали сегменты длиной 11 остатков из многих белков и пептидов, определив гидрофобный момент
и среднюю гидрофобность < // > для каждого из исследуемых сегментов. Для полипептидных сегментов глобулярных белков характерны низкие значения как <Я>, так и ци – Трансмембранные элементы гидрофобного характера имеют высокие значения <Я>, но низкие значения рн, являясь в основном неполярными. Пептиды и участки белков, относящиеся к «поверхностно-активным», имеют высокие значения цн из-за сильной асимметрии в распределении полярных и неполярных остатков. С помощью этого алгоритма были идентифицированы некоторые сегменты поверхностно-активных белков, например участки дифтерийного токсина и пируватоксидазы из Е. coli.
Гидрофобный момент служит количественной мерой периодичности в распределении гидрофобных остатков в разных участках полипептида. Важную роль при этом играет выбор 6. Гидрофобный момент является по существу одним из параметров фурье-преобразования функции гидрофобности. Более общие методы, описанные ниже, позволяют проанализировать все фурье-компоненты и выявить любую возможную периодичность.
2. Периодичность последовательности. Разработано много методов идентификации участков белковых молекул, для которых характерны периодические изменения гидрофобности вдоль цепи. Все они включают фурье-преобразование функции, зависящей от гидрофобности аминокислотных остатков вдоль полипептида. Наличие пика с периодом 3,6 указывает на то, что гидрофобный остаток в данном сегменте анализируемого полипептида встречается в среднем через каждые 3,6 остатка. Это означает, что сегмент является а-спиралью, на одной стороне которой находятся преимущественно гидрофобные остатки. Этот метод использовался для идентификации амфифильных участков в некоторых траспортных белках и белках, образующих каналы; в качестве примера можно привести ацетилхолиновый рецептор, натриевый канал, переносчик глюкозы, белок-разобщитель митохондрий и белок полосы 3 эритроцитов, являющийся анионным переносчиком. Однако четкие указания на то, что эти предполагаемые амфифильные спирали являются трансмембранными, отсутствуют.
Эти методы использовались также для анализа пептидов, взаимодействующих с мембранной поверхностью, и аполипопроте-инов.
Пептиды – модели мембранных белков
Пептиды стали использоваться для изучения белково-липидных взаимодействий много лет назад. В большинстве случаев это были природные мембраноактивные пептиды, в первую очередь грамицидин А, аламетицин и меллитин. В настоящее время в качестве модельных систем чаще применяют синтетические пептиды. При этом необходимо помнить о двух моментах: 1) при связывании пептида с мембраной существенны как первичная, так и вторичная амфифильности; 2) пептиды часто обладают полиморфизмом, т.е. способностью изменять конформацию в зависимости от окружения. Не; исключено, что в будущем с помощью синтетических пептидов удастся детально изучить белково-липидные взаимодействия, но пока мы еще очень далеки от этого.
|
