ЗМІСТ
Вступ
1. Умови існування плазунів в кінці палеозою і в мезозої
2. Причини швидкого розвитку плазунів, їх панівне положення в мезозої
3. Різноманітність давніх плазунів та основні напрямки і шляхи їх еволюції
3.1 Основні групи древніх плазунів
3.2 Напрямки та шляхи еволюції
4. Причини вимирання більшості груп плазунів
Висновок
Список використаної літератури
Вступ
Впродовж мільярдів років Земля змінювалася – в небо здіймалися гори, море заливало суходіл, острови підіймалися з глибин океану й перетворювалися на материки, але найбільших змін планета зазнала після виникнення життя близько 3,5 мільярди років тому. Моря закишіли найрізноманітнішими організмами, серед яких були такі, що біологи ще й досі ламають голови, куди їх віднести – до червів, молюсків, артропод чи ще куди. Суходіл зазеленівся і забуяв лісами, його борознили динозаври, мамонти, а нині перекроює на свій манер людина… Живий світ ніколи не залишався постійним, він еволюціонував, набуваючи нових форм і обрисів. Одночасно з народженням нових видів відбувалося вимирання старих – це постійний і безупинний природний процес. Проте в історії нашої планети були події, коли одно моментно зникали сотні тисяч видів.
Плазуни — одна з найдавніших груп наземних хребетних. Залишки давніх плазунів відомі з кам'яновугільного періоду палеозойської ери. Відособились вони від примітивних давніх земноводних – стегоцефалів, у яких виникло внутрішнє запліднення; яйце мало великий запас жовтка і тверду оболонку; розвинувся роговий покрив тіла, який захищав їх від висихання та ін. Основною предковою групою, яка дала початок як викопним, так і сучасним плазунам, були котилозаври.
Мета роботи – на основі літературних даних проаналізувати походження та філогенію плазунів.
Завдання роботи:
1) дати характеристику умов існування плазунів в кінці палеозою і в мезозої;
2) розглянути причини швидкого розвитку плазунів та їх панування в мезозої;
3) дати характеристику різноманітності давніх плазунів;
4) проаналізувати причини вимирання більшості груп плазунів.
Залишки найбільш древніх плазуючих відомі з верхнього карбону (~300 млн. років тому). Припускають, що відокремлення від земноводних предків повинне було початися, видимо, у середньому карбоні (320 млн. років), коли від антракозаврів, подібних Dіplovertebron, відокремлювалися форми, видимо, що володіли більшої наземності. Від подібних форм виникає нова галузь - сеймуріоморфи (Seymourіomorpha), залишки яких виявлені у верхньому карбоні - середній Пермі. Частина палеонтологів відносить цих тварин до амфібій.
На початку ери в кембрійський період життя все ще існувало у воді. Значно поширились великі багатоклітинні бурі і зелені водорості. В силурійський, а можливо, ще в ордовицький або кембрійський період у деяких популяціях зелених водоростей, які мешкали у водоймах, що періодично пересихали, в результаті ароморфозу сформувались тканини, які вперше виникли у наземних рослин — псилофітів. Псилофіти — збірна назва. Ці рослини мали невеликі розміри (не більше півметра), стеблоподібну наземну частину і кореневище, від якого відходили ризоїди. Деякі з них були ще дуже схожими на водорості, а інші вже мали ознаки, які зближували їх з мохоподібними і папоротеподібними рослинами. Рослини могли існувати на суходолі, оскільки бактерії, синьозелені водорості, одноклітинні тварини уже сформували тонкий шар ґрунту. В цей час з'явились і гриби, життєдіяльність яких сприяла ґрунтоутворенню.
Реклама

У морях кембрійського, ордовицького, силурійського періодів жили найпростіші, губки, кишковопорожнинні, членистоногі, молюски, голкошкірі, нижчі хордові. В силурійський період з'явилися найпримітивніші хребетні — круглороті. У них ще не було щелеп, але внаслідок ароморфозу виникли череп і хребці. У девонський період досягли розквіту щиткові, що належали до класу круглоротих. У риб внаслідок ароморфозу утворився щелепний апарат, що дав їм змогу активно полювати, захоплювати здобич. Це сприяло підвищенню рівня організації нервової системи, органів чуття, вдосконаленню інстинктів.
Серед сучасної фауни найближчими до цих найдавніших риб є акули і скати. З'явились і кистепері риби. Нечисленних представників цих риб виявлено в Індійському океані біля берегів Африки. Кистепері риби, які населяли прісні водойми в девонський період, зробили важливий крок в еволюції тварин — вийшли на суходіл. Першими наземними хребетними були найдавніші земноводні стегоцефали — потомки кистеперих риб. Скелет плавців кистеперих гомологічний скелету п'ятипалої кінцівки стегоцефалів, як і в сучасних земноводних, яйця і личинки могли розвиватися лише у воді, тому ці тварини змушені були жити поблизу водойм.
Клімат наступного, кам'яновугільного періоду був теплим, вологим, атмосфера містила велику кількість вуглекислого газу. Це сприяло бурхливому розвитку папоротеподібних, це був період їхнього розквіту. Деякі хвощі досягали 30метрової висоти.
Розвиток наземної рослинності сприяв формуванню ґрунту, з решток рослин того періоду утворилося кам'яне вугілля. В ньому ніби законсервувалась значна частина вуглекислого газу атмосфери. В результаті інтенсивного фотосинтезу, який відбувався в зелених рослинах, атмосфера збагачувалась на кисень. Внаслідок зміни хімічного складу атмосфери з'явилась можливість заселення суходолу тваринами.
У рослинному світі відбувся ще один ароморфоз — з'явились насінні папороті. В насінні нагромаджувались поживні речовини, воно мало оболонку, яка захищала його від несприятливих умов. Насінним рослинам для запліднення насінини вода не потрібна, що дало їм змогу завоювати суходіл.
Клімат кам'яновугільного періоду сприяв також розквіту земноводних (стегоцефалів). На суходолі ворогів у них ще не було, а численні черви і членистоногі, особливо павукоподібні і комахи, були для земноводних відмінним кормом. Внаслідок дивергенції та ідіоадаптацій з'явилось багато видів стегоцефалів, деякі з них досягали гігантських розмірів (до 47 м завдовжки).
Реклама

Наприкінці кам'яновугільного й особливо в пермський період клімат змінився, став сухим. Це призвело до масового вимирання папоротеподібних і земноводних. Ті ж особини, які вижили у більш вологих місцях, дали початок земноводним і папоротеподібним невеликих розмірів. Від стегоцефалів виникли не лише земноводні, а й плазуни.
У пермський період від насінних папоротей виникли голонасінні рослини, які значно поширились на суходолі, оскільки з'явився новий спосіб запліднення, не пов'язаний з водою, а формування насінини давало змогу зародкам рослин тривалий час витримувати несприятливі умови. Внаслідок цього насінні рослини змогли оселятись не лише на вологих узбережжях, а й проникати в глибину материків.
Розвиток життя в мезозойську еру. Початок мезозойської ери — тріасовий і юрський періоди — був часом розквіту і панування плазунів. Деякі з них досягали гігантських розмірів і мали масу тіла до 20 т. Були серед них як рослиноїдні, так і м'ясоїдні. Однак ще в пермський період з'явились звірозубі плазуни — предки ссавців. Перші ссавці (однопрохідні та сумчасті) відомі з тріасового періоду. Тоді ж виникли справжні костисті риби і псевдозухії — плазуни, які пересувалися лише на задніх кінцівках. Псевдозухій вважають предками птахів. Першоптах — археоптерикс — з'явився в юрський період і існував ще й У крейдовий період.
Прогресивний розвиток дихальної і кровоносної систем птахів і ссавців забезпечив теплокровність цих тварин, послабив їх залежність від температури навколишнього середовища, що сприяло розселенню їх по всіх географічних широтах.
Справжні птахи і вищі ссавці з'явилися у крейдовий період. Вони швидко посіли панівне становище серед хордових. Цьому сприяли також розвиток нервової системи, виникнення умовних рефлексів, виховання потомства, а у ссавців — живородіння і вигодовування малят молоком. Прогресивною була і диференціація зубів у ссавців, що стало передумовою вживання різноманітної їжі.
Завдяки дивергенції та ідіоадаптаціям з'явились численні ряди, роди і види ссавців і птахів.
Наприкінці мезозою в крейдовий період від голонасінних виникли покритонасінні, що пов'язано з новим ароморфозом — появою квітки. Вона добре захищає насінину, яка розвивається у зав'язі, від несприятливих факторів.
У результаті дивергенції у відділі покритонасінних сформувались два класи рослин: однодольні і дводольні, а внаслідок ідіоадаптацій у цих класах виробились різноманітні пристосування до запилення.
Наприкінці мезозою з'явились перші плацентарні ссавці, (комахоїдні). У зв'язку із сухістю клімату розпочалося масове вимирання голонасінних, а оскільки вони були кормом для багатьох плазунів, то це призвело до вимирання і плазунів.
Початок мезозойської ери — тріасовий і юрський періоди — був часом розквіту і панування плазунів. Деякі з них досягали гігантських розмірів і мали масу тіла до 20 т. Були серед них як рослиноїдні, так і м'ясоїдні. Однак ще в пермський період з'явились звірозубі плазуни — предки ссавців. Перші ссавці (однопрохідні та сумчасті) відомі з тріасового періоду. Тоді ж виникли справжні костисті риби і псевдозухії — плазуни, які пересувалися лише на задніх кінцівках. Псевдозухій вважають предками птахів. Першоптах — археоптерикс — з'явився в юрський період і існував ще й у крейдовий період.
Тріасовий період характеризується значним скороченням загальної площі платформених морів. Величевні континенти того часу перетиналися високими хребтами молодих гір.
На рівнинах в умовах жаркого або мінливо вологого клімату утворювались великі площі пустель і напівпустель або низовини, вкриті болотами та лісами. Про це свідчать потужні товщі континентальних червоних або строкатоколірних порід та темноколірних гумідних порід (місцями вугленосних), якими представлені тріасові відклади на багатьох ділянках земної кулі і зокрема на території України, вся площа якої, за винятком Криму та Карпат, являла собою суходіл.
Походження плазунів пов'язане з ароморфозами, що забезпечували розмноження на суходолі: внутрішнє запліднення, запас поживних речовин в яйці, вкритому щільною оболонкою, яка захищала його від висихання. Під час розвитку зародка всередині яйця нагромаджувалась рідина, в якій зародок знаходився як в акваріумі, відбувається формування зародка. Це дало змогу плазунам завоювати всі середовища існування: суходіл, повітря і повторно оселитися у воді. Прогресу плазунів сприяв розвиток рогового покриву, який захищав від висихання, досконаліший розвиток легень, кровоносної системи, кінцівок, головного мозку. Все це дає підстави визнати плазунів першими справжніми наземними хребетними.
Рептилії-нащадки древніх амфібій, що вдало розв'язали проблему розмноження на суші їм стали першими цілком наземними хребетними. "Винахід" амніотичного яйця нового типу і зв'язані з цим процеси розвиток-головна відмінність рептилій від земноводних.
Рептилії відкладають яйця на суші і , отже, необхідність у який би те ні було адаптації до водного способу життя в молоді або в дорослих особин відпадає. Виникнення такого яйця було настільки важливим досягненням в еволюції наземних хребетних, що, як уже говорилося, рептилій разом із птахами, що відбилися від них, і ссавцями часто об`єднають під загальною назвою амніот .
Можливо, що самі древні рептилії вели водно-наземний спосіб життя і що яйце з'явилося однієї з адаптацій, що існувала паралельно з іншими адаптаціями, які спостерігаються в сучасних амфібій, але більш зробленої, оскільки ця адаптація рятувала яйця від хижаків і висихання. Більш імовірно, однак, що перші рептилії були дрібними наземними формами, що нагадували трохи дивних непропорційно великоголових ящірок.
За юрського періоду великі простори материків зазнали опускання і трансгресії в їхні межі моря. Наприкінці цього періоду море вкрило величезні простори суходолу як у північній, так і у південній півкулях, причому на території України тільки Український кристалічний щит та Донецький кряж були в цей час вище від рівня моря.
В юрському періоді остаточно вимер ряд груп тварин палеозою, виникла нова, характерна для мезозойської ери група голкошкірих — неправильні їжаки. В морських басейнах розвинулись амоніти, белемніти, на суходолі — рептилії. Наприкінці юрського періоду з’являється новий клас хребетних — птахи. Серед рослинного світу продовжували панувати голонасінні.
Юрський період характеризувався теплим вологим кліматом, який сприяв бурхливому розвитку деревної рослинності, про що свідчать потужні товщі вугленосних відкладів юрського періоду. Хоча в глибині континентів деякий час зберігалися засушливі умови. Внутрішні райони Північної Америки залишалися пустелями і динозаври, що там проживали, такі як дилофозаври добре пристосувалися до жари і пилу.
На пізньоюрський період припадає розквіт літаючих ящерів птерозаврів. В кінці юрського періоду довгохвості птерозаври (рамфоринхи) остаточно вимерли.
Морські води у юрі були значно теплішими, як тепер. Окрім іхтіозаврів моря населяли довгошиї плезіозаври. Крокодили призвичаїлися до життя в морі.
Юрський розвій рослинності сприяв розквіту рослинноїдних завроподів — найбільших тварин за всю історію Землі (брахіозавр, апатозавр, диплодок). А також травоїдні, доволі оригінальні на вигляд стегозаври — вздовж хребта був розташований подвійний ряд грізного вигляду пластин.
У юрі виникло багато двоногих хижих динозаврів тероподів, таких як найрозповсюдженіші юрські хижаки аллозаври, мегалозаври, цератозаври, орнотолести.
Саме у юрському періоді з’явилися знамениті археоптерикси — істоти, які мали одночасно ознаки птахів і рептилій.
В кінці цього періоду відбувались Інтенсивні горотворні рухи, які супроводились виходом з-під рівня моря значної частини континентів.

Основна лінія, від якої пішли рептилії, давно вимерлий ряд Cotylosaurіa в усьому, крім удосконаленого розмноження, залишалася дуже архаїчним; кінцівки в котилозаврів відходили в сторони від тулуба і по більшості ознак вони мало просунулися вперед у порівнянні зі своїми предками і близькими родичами серед амфібій. Найперші і найбільш примітивні котилозаври мали невеликі розміри, але деякі з більш пізніх форм були великими незграбними травоїдними. Безсумнівно, завдяки тому, що рептиліям удалося розірвати щепи, що приковували їх до води, від котилозаврів могли виникнути групи, що займають таке видне положення у фауні мезозою.
Черепахи (ряд Testudіnes)- бічна галузь, що відходить від основного стовбура рептилій. Незграбність черепах нагадує про їхніх палеозойських предків, однак у них мається помітний досягнення-розвиток кісткового панцира, покритого щитками, і захищаючу спину і черево. У панцир включені ребра і кісткові пластинки шкірного походження, а плечовий і тазовий пояси розташовуються усередині від ребер. Надягши броню, черепахи стали консерваторами і, починаючи з тріасу, майже не прогресували. Єдине істотне більш пізніше удосконалення, досягнуте черепахами, це здатності втягувати голову під панцир. У всіх добре знайомих нам форм (так званих скритошийними черепахами, або Cryptodіra) це досягається шляхом S-подібного згинання шиї у вертикальній площині і підтягування голови назад; деякі своєрідні тропічні форми (бокошийні, або Pleurodіpa) ховають голову, згинаючи шию убік у горизонтальній площині. Подібно більшості древніх черепах, сучасні черепахи в більшій частині ведуть напівводний спосіб життя, населяючи болота і ставки. Деякі форми, однак, повернулися до чисто водного способу життя; крім того, існує трохи морських форм із веслоподібними кінцівками для активного просування вперед. Протилежну крайність складає група сухопутних черепах (родина Testudіnіdae), що цілком перейшли до наземного способу життя.
У двох згаданих дотепер рядів рептилій у даху черепа , тому що й у їх предкових амфібій, отвори маються тільки для головних органів почуттів, тобто для ніздрів і очей ( у також для тім'яного ока). За цією ознакою котилозаврів і черепах відносять до групи Anapsіda. У всіх інших рептилій у скроневій частці маються додаткові отвір-скроневі ями, що служать діагностичною ознакою в систематику рептилій. У залежності від особливостей розташування скроневих ям виділяють типи - Euryapsіda і Parapsіda (розташовуються високо в щічних областях, симетрично); Synapsіda (одна зі скроневих ям розташована більш вентрально) і діапсиди - Lepіdosaurіa і Archosaurіa - у яких було 2 скроневих ямки з кожної сторони.
Усі евріапсидні рептилії вимерли і зведень про них мало. Деякі були дуже примітивними,
На жаль, у міру вивчення виникає усе більше сумнівів у тім, що вони відносяться до евріапсидів. Незважаючи на свою новопридбану здатність до завоювання суші, багато груп мезозойських рептилій, подібно морським черепахам, вибрали життя в море. Особливо відомі такі мезозойські форми як плезіозаври, плакодонти й іхтіозаври. Плезіозавра (отрядSauropterygіa) нерідко описують як "змієві, просмикнуту через тіло черепахи". Плезіозаври зовсім не родинні ні однієї, ні іншій групі з цих двох рептилій, але опис ящерицеподібними наземними формами;їх відносять до загону Araeoscelіdіa.

У плезіозаврів була довга шия, тіло зразкове тієї ж довжини як і шия, широке і відносно плоске. Повернення до дійсного риб'ячого способу пересування був неможливий через те, що тулуб не згинався , а хвіст був коротким; зате кінцівки плезіозаврів перетворилися в могутні, що нагадують весла структури, за допомогою яких вони "гребли", прокладаючи собі шлях в океані. У сутності їхнього руху подібно рухам морських черепах могли бути літальними, а не гребними .Більш примітивні форми (нотозаври) також були модифіковані, хоча і не настільки ефектним способом.
Плакодонти (загін Placodontіa) - тріасові форми, що перейшли до харчування молюсками й іншим безхребетними з твердим панциром. Це були великовагові тварини зі сплощеним тілом;деякі з них були одягнені бронею, схожою на панцир черепах. Їхні родинні зв'язки з плезіозаврами, як, втім і усе, що стосується евріапсид, у даний час викликають суперечки, а деякі дослідники взагалі заперечують близьке споріднення між цими групами. Ще більш незвичайною будовою відрізнялися Іchtyosaurіa-"риборептилії". Іхтіозаври на відміну від плезіозаврів, що все-таки могли пересуватися по суші, подібно сучасним тюленям, перетворилися в цілком морські форми. Судячи з палеонтологічних даних, вони перейшли від відкладання яєць до живородіння. Форма тіла цілком повернулася до риб'ячого: шия втяглася в тулуб, додавши йому веретеноподібну форму., кінцівки укоротилися, перетворилися в невеликі плавникоподібні пристосування для керування рухом. Пересувалися за рахунок хвилеподібного згинання тулуба і хвоста; на спині в них розвився плавець, схожий на риб'ячий (однак як і плавець китів, він був позбавлений кісткових променів, як у риб). Хвіст, подібний з акулячим, перетворився в могутній плавальний орган; однак у його будові була одна істотна відмінність: в акули кінець хребта загнуть вгору і заходить у верхню лопату хвостового плавця, а в іхтіозавра він різко загинався вниз, а плавець простирався над ними.
Cинапсиди. Основною предковою групою, що дала все розмаїтість сучасних і викопних плазуючих були котилозаври, однак подальший розвиток рептилій йшов різними шляхами.
Спочатку відокремилися анапсиди (Anapsіda) і синапсиди (Synapsіda). У структурі черепа анапсид були відсутні які-небудь западини. Ймовірними нащадками цього напрямку стали черепахи (нині Testudіnes, або Chelonіa), у них за очницями зберігається суцільний кістковий покрив. Предком черепах прийнято вважати пермського еунотозавра (Eunotosaurus) - невелике ящероподібна тварина з короткими і дуже широкими ребрами, що утворюють подобу спинного щита.
Cинапсиди - рептилії з нижніми скроневими западинами, обмеженими скулової, лускатою і заочноямковою кістами. Вже в пізньому карбоні ця група стала самої численної. У палеонтологічному літописі вони представлені двома послідовно існуючими загонами: пелікозаврів (Pelіcosaurіa) і терапсид (Therapsіda). Період свого розквіту тераписди пережили задовго до того, як з'явилися перші динозаври, нащадками терапсид стали більш високоорганізовані звірозубі (Therіodontіa). До них відноситься, зокрема, хижа іностранцевія, що нагадує саблезубого тигра. Звірозубі були численні ще в ранньому тріасі, але з появою хижих динозаврів вони зникли. Звіроподібні становлять великий інтерес як група, що дала початок ссавцем.
Діапсиди. Наступною групою, що відокремилася від котилозаврів, були діапсидні (Dіapsіda). Їхній череп має дві скроневі западини, розташовані вище і нижче заочноямкової кісти. Діапсидні наприкінці палеозою (Перм) дали надзвичайно широку адаптивну радіацію систематичним групам і видам, яких виявляють і серед вимерлих форм, і серед нинішніх рептилій. Серед діапсидних намітилися дві основні групи Лепідозавроморфи (Lepіdosauromorpha) і Архозавроморфи (Archosauromorpha). Найбільш примітивні діапсиди з групи Лепідозаврів - ряд Еозухії (Eosuchіa) - були предками ряду Клювоголові, від яких у даний час зберігся лише один рід гаттерії.
Наприкінці Пермі від примітивних діапсид відокремилися лускаті (Squamata), що стали численними в крейдовий період. До кінця крейдового періоду від ящірок відокремилися змії.
Архозаврів вважають самими цікавими з усіх рептилій, коли-небудь, що жили на Землі. Серед них - крокодили, що літають птерозаври, динозаври. Крокодили - єдині архозаври, що дожили до наших днів.
Відокремлення вихідної групи архозаврів - текодонтов (псевдозухий) - від эозухий, видимо, відбулося у верхній Пермі. У тріасі текодонти досягли розквіту. Частина текодонтів, повернулася до напівводного способу життя, до кінця тріасу давши початок крокодилам. Інша частина - орнітозухії, імовірно, вела деревний спосіб життя і, по одній з версій, дала початок сучасним птахам[3]. У середині тріасу від вже іншої групи текодонтів відбулися птерозаври, численні в юрський і крейдової періоди, але цілком вимерлі до кінця мела.
У верхньому тріасі від хижих текодонтів відокремилися ще дві групи - ящеротазові і птицетазові динозаври, до кінця мела вимерлі.
Іхтіозаври (Ichthzosauria) — ряд вимерлих морських ящерів. Виникли в тріасовий період, деякі дослідники вважають, що від мезозаврів; вимерли в крейдовий період. Тіло (довжиною від 1 до 15 м.) внаслідок життя у воді набудо рибоподібної форми, звідки і походить назва «іхтіозаври», що означає рибоящери.

шонізавр
Іхтіозаври з’явилися раніше динозаврів, приблизно 220 млн. років тому, вимерли вони також першими.
Хребці двовгнуті, числом 150—200; спинний і хвостовий — шкірні. Пальці (від 3 до 8) разом з іншими частинами кінцівок утворювали ласти. Носова частина рила витягнута, всаджена численними зубами. За способом життя іхтіозавр нагадував сучасних дельфінів. Були живородящими. Знайдені скелети дорослих іхтіозаврів, в яких знаходилися скелети ненароджених нащадків. Живилися рибою і головоногими молюсками. Відомі з крейдових відкладів на Україні (з сеноману Канева, білої крейди Новгорода-Сіверського й Ізюма).
Викопні рештки шонізавра — найбільшого з відомих іхтіозаврів — знайшли у штаті Невада (США). Він жив приблизно 210 млн. років тому. Його масивне тіло досягало 15 м. На відміну від більшості інших іхтіозаврів зуби шонізавра містилися лише у передній частині щелеп.

Закам’янілий скелет іхтіозавра
 
Мезозавр — Скелет іхтіозавра з обрисами тіла
імовірний предок іхтіозаврів

Дилофозавр («Ящер з подвійним гребенем»)
Ранній юрський період. Досягав 6 метрів в довжину. вага — 400 кг. Відкритий в 1942 році у США.
Пересувався на двох ногах, хижак, на голові мав подвійний костяний гребінь.
Цератозавр («Рогатий ящер»). Пізній юрський період. Мав 6 м. в довжину. Вага — 1 т. Відкритий в 1883 р. у США. Хижак; пересувався на двох ногах; мав надбровні гребені і ріг на носі. Відносився до тероподів.
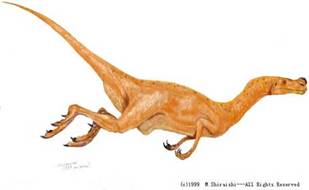
Орнітолест. («Той, що грабує птахів»)
Пізній юрський період. Мав 2 м. в довжину. Важив 13 кг. Відкритий у 1903 році у США. Пересувався на двох ногах; дуже швидкий; міцні щелепи; гострі міцні зуби; виступ над ніздрями; пристосовані до хапання пальці.хижий; шкіра вкрита наростами; два коротких роги над очима; довий хвіст; гострі зуби.
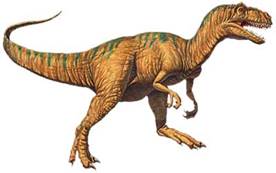
Аллозавр («Дивний ящер»)
Пізній юрський період. Мав 11 м. в довжину. Вага — 1,5 т. Відкритий в 1869 р. у США. Хижак; пересувався на двох ногах; маленькі, але сильні передні кінцівки; величезні зуби, сильні ноги і міцний хвіст. Відносився до тероподів. Аллозаври були найрозповсюдженішими хижакми юрського періоду.

Брахіозавр («Ящер з руками»)
Пізній юрський період. Досягав 23 м. в довжину. вага — 50 т. Відкритий в 1900 році у США. Пересувався на чотирьох ногах, передні кінцівки значно довші задніх, рослинноїдний, вів стадний спосіб життя, мав високий гребінь на голові, дуже великий крок. Відносився до завропод.

Апатозавр («Ящер, що вводить в оману»)
Пізній юрський період. Досягав 21 м. в довжину. вага — 20 т. Відкритий в 1877 році у США. Пересувався на чотирьох ногах, рослинноїдний. Відносився до завропод. Раніше його називали «бронтозавром».
Диплодок («Подвійна балка»). Пізній юрський період. Мав 27 м. в довжину. Вага — 10 т. Відкритий в 1877 р. у США. Рослинноїдний динозавр; маленька голова; дуже довгий гнучкий хвіст; товсті ноги; змієвидна шия. Мав відносно легкий скелет. Відносився до завропод.

Стегозавр («Покрівельний ящер»)
Пізній юрський період. Мав 9 м. в довжину. Вага — 6—8 т. Відкритий в 1877 р. в США. Пересувалися на чотирьох ногах; маленька голова; дуже малий мозок, розміром з грецький горіх; на спині подвійний ряд пластин; хвіст з шипами; беззуба дзьобоподібна паща.
Очевидно, що пластини у стегозавра служили стабілізаторами темпиратури тіла. Всередині пластин проходили кровоносні судини. Деякі дослідники вважають, що у самців пластини були яскравого кольору, для привернення уваги самок.
Співвідношення між розмірами мозку і тулуба стегозавра — найменше серед усіх динозаврів.
Птерозаври (Pterosauria) — птахоящери. З’явилися в пізній тріас, вимерли у крейдовий період. Були родичами динозаврів. Та якщо динозаври панували на землі, то птерозаври — в повітрі, доки не з’явилися птахи.

птерадон
Викопні рештки птерозавра вперше були знайдені на півдні Німеччини в 1784 р. Тоді всі вважали, що це кістки невідомої водяної тварини. Лише через 100 років вчені дійшли висновку, що знайдені рештки належали викопному літаючому ящеру — птеродактилю (пальцелітаючому).
Крило птерозавра, це шкіряна складка натягнута між тілом і збільшеним четвертим пальцем передньої кінцівки. Скелет літаючих ящерів був дуже легким — з великими пустотами всередині кісток. Досить великий мозок координував роботу м’язів і забезпечував хороший зір. Предками птерозаврів, очевидно, були давні рептилії псевдозухії.
На викопних рештках деяких птерозаврів знайдені сліди шерсті, що дозволяє робити припущення про їхню теплокровність, адже шерсть покликана виконувати теплоізоляційні функції. Допускають, що шерсть птерозаврів була білого кольору (подібного до кольору пір’я сучасних чайок). Білий колір менше відлякував рибу, адже саме вона була основним продуктом харчування літаючих ящерів.
Птерозаври, очевидно, не лише планували, але й володіли активним польотом.
Птерозаврів поділяють на дві великі групи — давніших рамфоринхів і пізніших птеродактилів. Хвостаті рамфоринхи вимерли в кінці юрського періоду.
У деяких птерозаврів могли бути перепончаті лапи, щоб сідати на воду.
Птерозаври мабуть зригували їжу, щоб нагодувати своїх дітей.Слід відзначити, що літаючі ящери не були предками птахів і кажанів. Літаючі ящери, птахи і кажани були тваринами різного походження і розвивалися кожний своїм шляхом. Єдине, що їх єднало — здатність літати. Всі вони набули цієї здатності завдяки трансформації передніх кінцівок. Хоча відмінність у будові крил цих тварин підтверджує те, що вони були зовсім різного походження.

Птеродактилі були іноді розмірами з горобця, іноді гігантами, як кетцалькоатль (розмах крил 12 м., вага 86 кг.). Характерними ознаками всіх птеродактилів є відсутність хвоста і довга гнучка шия. Склавши крила, деякі птерозаври могли ходити по землі на задніх кінцівках. Другі, можливо, ходили на всіх чотирьох.

Археоптерикс, первоптах довгохвостий (Archaeopteryx macrura) — первісний птах юрського періоду (приблизно 150 млн. років тому). 1860 знайдено відбиток пера археоптерикса у верхньоюрських сланцях у Баварії, в 1861 там же знайдений неповний скелет з відбитками пір’я, придбаний Британським музеєм і описаний Р. Оуеном під вказаною назвою. У 1877 було знайдено там же найповніший зразок археоптерикса.
Археоптерикси були розміром з голуба, мали дзьоб з конічними зубами, по три вільні пальці на крилах, двояковгнуті хребці, довгий хвіст з 20—21 хребця, вкритий пір’ям. Рульові пера кріпилися попарно на кожному хребці довгого хвоста, а не так, як у сучасних птахів — віялом на копчиковій кістці.
На ногах археоптерикс мав по чотири пальці: перший палець повернутий назад, інші — вперед, що дозволяло добре обхоплювати гілки дерев. Археоптерикси мабуть жили на деревах, стрибали і лазили по гілках за допомогою довгих пальців з кігтими на передніх кінцівках.
Розправляючи крила, вони могли планувати у повітрі, а також перелітати на невеликі відстані розмахуючи крилами. Деякі дослідники вважають, що археоптерикс взагалі не вмів літати, а за допомогою крил він просто зганяв комах під час полювання.
Ряд ознак зближує археоптериксів з динозаврами. Відкриття археоптериксів стало свідченням наявності перехідних форм між рептиліями (динозаврами) і птахами.
Вперше теорія про родинні звязки птахів і динозаврів була висунута в 1926 році дацьким вченим Герхардом Хейлманом. Він опублікував книгу, в якій описав скелети динозаврів і птахів, вказавши на деяку подібність між ними. Цю позицію підтримав англійський вчений Томас Хакслі. Однак у Хакслі були певні сумніви — наскільки тоді було відомо палеонтологам, у динозаврів не було вилочкової кістки, тоді як у птахів вона була.
Пізніше професор Джон Остром зрівняв скелет археоптерикса зі скелетом дрібного динозавра компсогната, що жив на Землі в пізній юрський період. Він звернув увагу на ряд подібних ознак і вказав на те, що в деяких динозаврів вилочкова кістка все-таки була.
В крейдовому періоді вимерли іхтіозаври, їхню нішу зайняли мозозаври (родичі сучасних ящірок, з величезними крокодилоподібними тілами і ластами замість кінцівок.
В пізній крейді клімат став прохолодніший, саме тоді почали складатися кліматичні зони сучасного типу.

В засушливих районах сучасної Монголії жили швидкі хижі велосираптори та руйнівники гнізд інших динозаврів овіраптори. На території сучасної Канади виходили на колективне нічне полювання швидкі, як блискавка дрібні троодони.
Велосираптор («Швидкий злодій»). Пізнній крейдовий період. Мав 1,8 м. в довжину. Вага — 15 кг. Відкритий в 1924 р. у Монголії.
Пересувався на двох ногах; дуже рухливий хижак; гострі кігті на другому пальці нижніх кінцівок; довгий жорсткий хвіст; гострі зуби; досить великий мозок.
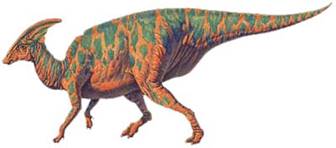
Паразавралоф («Біля завралофа»)
Пізнній крейдовий період. Мав 10 м. в довжину. Вага — 3 т. Відкритий в 1922 р. у США. Рослинноїдний; пересувався переважно на двох ногах; полий трубчатий виріст на голові через який він міг подавати голосні звуки; дзьоб схожий на качиний.
Тиранозавр («Ящер-тиран»). Пізній крейдовий період. Мав 12 м. в довжину. Вага — 7 т. Відкритий в 1902 р. у США.
Хижак; пересувався на двох ногах; мав надзвичайно маленькі передні двопалі кінцівки; величезні щелепи; гострі пилкоподібні зуби. Тиранозавр був одним із найжахливіших хижаків, що коли небудь жили на нашій планеті.
На території сучасних Північної Америки і Азаї, які час від часу були з’єднані мешкали качконосі динозаври, такі як парозауралоф, а також гігантські хижі динозаври тиранозаври, одні з найбільших зижаків які будь-коли населяли нашу планету. В пустелях Північної Африки полював хижий спінозавр. В горах Північної Америки жили панцирні рогаті динозаври трицератопси.
З верхніх пермських відкладень Північної Америки, Західної Європи, Росії і Китаю відомі залишки котилозаврів (Cotylosaurіa). По ряду ознак вони ще дуже близькі до стегоцефалів. Їхній череп був у виді суцільної кісткової коробки з отворами тільки для очей, ніздрів і тім'яного органа, шийний відділ хребта був слабко оформлений (хоча мається характерне для сучасних плазуючу будівлю перших двох хребців - атланта і епістрофія), хрестець мав від 2 до 5 хребців[1]; у плечовому поясі зберігалася клейтрум - шкірна кіста, властива рибам; кінцівки були короткі і широко розставлені.
Імовірно, вже в карбоні в рептилій сформувався цілий комплекс пристосувань, що дозволили їм стати повною мірою наземними тваринами. Найважливішим з цих пристосувань, що зробило можливими багато інших істотних змін організації плазуючих, був розвиток ефективного механізму вентиляції легень за допомогою рухів грудної клітки. Він зробив непотрібним шкірне подання, що у земноводних є необхідним доповненням легеневого, тому що через недосконалість вентиляції легень, де накопичується надлишок вуглекислого газу, земноводні мають потребу у виведенні останнього через шкіру (у земноводних забір повітря відбувається не за рахунок розширення грудної клітки, а в результаті дії під'язичного насоса; недолік легеневого подиху компенсуються шкірним, тому шкірні покриви повинні бути завжди вологими). Тому в шкірі рептилій стало можливим посилення процесів зроговіння епідермісу, що захистило організм від постійної утрати вологи через покриви (неминучого при шкірному подиху в земноводних) і ризику загибелі в сухому повітрі від зневоднення. Після цього плазуючі одержали можливість заселити практично будь-які місцеперебування на суші, аж до безводних пустель.
Подальша еволюція рептилій обумовлювалася їхньою мінливістю в зв'язку з впливом різноманітних умов життя, з якими вони зіштовхувалися при розмноженні і розселенні. Більшість груп стали більш рухливі; кістяк їх полегшав , але в той же час і міцніший. Рептилії використовували більш різноманітну їжу, чим амфібії. Змінювалася техніка її добування. У цьому зв'язку істотним змінам піддалася будівля кінцівок, осьового кістяка і черепа. У більшості кінцівки стали довші, таз, здобуваючи стійкість, прикріплювався до двох і більш крижових хребців. У плечовому поясі зникла "рибна" кіста клейтрум. Суцільний панцир черепа піддався часткової редукції. У зв'язку з більш диференційованою мускулатурою щелепного апарата в скроневій області черепа виникли ями і поділяючі їхні кісткові мости - дуги, що служили для прикріплення складної системи мускулів.
Як стверджують палеонтологи, за останні 500 мільйонів років на Землі відбулося 5 великих вимирань. Перше відоме нам глобальне вимирання, про яке свідчить геологічний літопис, трапилось в Ордовицькому періоді Палеозойської ери 440 мільйонів років тому, що призвело до втрати 25% родин тварин. Слід зауважити, що тоді існувало всього 600 родин, для порівняння сьогодні на Землі існує близько 1100 родин тварин. Тобто було втрачено 150 родин, а це десятки й сотні тисяч видів. Як свідчать геологічні дослідження, причиною вимирання, очевидно, став астероїд, оскільки з ордовицькими відкладами пов’язані величезні товщі вулканічного попелу та чорного мулу, а також поліметалічні руди, які могли бути занесені лише з космічними тілами. В Ордовику життя заселяло лише воду, а на суші поширились лише бактерії та різноманітні водорості.
Наступне вимирання трапилось через сто мільйонів років в Девоні, на цей раз було втрачено 19% родин. Причин девонського вимирання достеменно невідомо, але в осадових породах також знаходять вулканічні продукти та кольорові метали. Справа втім, що при падінні астроїда відбувається активізація тектонічних процесів і починаються масові виверження вулканів, які спричинюють запилення атмосфери і планетарне похолодання. Окрім того, сам астероїд при падінні підіймає багато пилу, починаються пожежі (а в Девоні уже були псилофітові ліси), що також зменшує прозорість атмосфери і менша кількість сонячних променів потрапляє на поверхню землі.
Найбільш катастрофічним і глобальним вимиранням було третє, яке відбулось в Пермському періоді Палеозойської ери, 250 мільйонів років тому, в результаті падіння двох астероїдів. Один з них впав на території Центральної України, а другий в Індії. Варто сказати, що на той ас Центральна Україна була вкрита морем, а суша була лише на території сучасного Донецького кряжу, де підіймалися гори на кшталт сучасних Карпат. Падіння астероїда в океан підняло високу хвилю цунамі, яка обійшла планету і цілком спустошила узбережжя та мілководдя. Індійський астероїд впав на сушу, ударна хвиля від якого прокотилась континентом, а саме падіння спричинило жахливі пожежі задимлення й запилення атмосфери, активізувалися вулкани, і по суті, настав короткочасний льодовиковий період. Майже, цілком зникли папоротеві і каламітові ліси, а їм на зміну прийшли соснові, араукарієві, саговникові та ґінкґові ліси. Це вимирання призвело до втрати 54% (!) родин і різко скоротило рівень біорізноманіття на планеті. Вимерли навіть, здавалось би, такі вічні трилобіти – мешканці морів, які пережили усі попередні вимирання.
Четверте вимирання трапилось незадовго – уже в тріасовому періоді мезозойської ери, 210 мільйонів років тому. І знову втрачено 23% родин, які представляли живий світ тодішньої Землі. Причини його є не цілком зрозумілими, але саме це вимирання стерло з лиця землі більшість плазунів і звірозубих ящерів, а їм на зміну прийшли динозаври, які панували наступні, майже, 150 мільйонів років до початку кайнозойської ери.
Останнє велике вимирання відбулось у крейдяному періоді мезозойської ери – 65 мільйонів років тому, яке було спричинене падінням астроїда в зоні півострова Юкатан у Мексиці. Правда, воно не було настільки катастрофічним як попередні – втрачено лише 17% родин. Проте, вимерли всі представники тодішньої мегафауни, і зокремадинозаври, але саме це вимирання дало поштовх до бурхливого еволюційного розвитку птахів та ссавців.
Це були суто природні вимирання – в силу обставин, які склались на Землі чи у Сонячній системі, або у ближчих до нас секторах Галактики. Сьогодні ж спостерігається невтішна тенденція – людство ставить під питання виживання десятки тисяч таксонів, починаючи від форм і рас різних видів й закінчуючи цілими родинами, а то й рядами. Більшість біологів вважають, що це початок Четвертинного – шостого вимирання…
Учені забили на сполох десь в середині минулого століття, коли науково-технічний проґрес впевнено і переможно крокував планетою. У якийсь момент цієї ходи в індустріальних суспільствах трапився злам свідомості – людство призамислилось… Чи реальним є глобальне вимирання? Чи людина може його спровокувати? Скільки ще треба руйнувати біосферу, щоб це трапилось? І тут бачимо уже не романтично-філософську концепцію ноосфери Вернадського, коли людство створює оболонку розуму планети, підпорядковуючи природу своїм потребам. А реалістичний процес знищення біосфери і самого людства!
Вчені Європи і Америки в один голос заявляють про те, що наступило глобальне вимирання біологічних видів, спричинене людиною, причому, це вимирання, як дехто стверджує, набирає темпів, які значно перевищують темпи вимирання, що коли-небудь були на Землі, навіть при “метеорній катастрофі”. Прогнозується, що за найближчі 30 років на планеті вимре близько п’ятої частини біологічних видів. Це при тому, що на сьогодні відомо лише трошки більше дох мільйонів виді живих організмів, а за оцінками більшості біологів кількість біологічних видів на планеті може становити від 10 до 100 мільйонів видів. Навіть взявши за основу 10 мільйонів видів, які населяють біосферу, то при таких темпах вимирання буде втрачено 2 мільйони! Над біосферою – нашим спільним домом нависла реальна і, можливо, незворотна екологічна катастрофа. В першу чергу, вимиратиме сучасна мегафауна – слони, носороги, жирафи, антилопи, тигри, ведмеді, кити та багато інших великих видів. Причиною таких глобальних негативних процесів є, в першу чергу, дуже швидке зростання населення людей на планеті, що потребує нових територій для існування, відповідно вирубуються ліси, розорюються нові території, промислове зростання спричинює процеси глобального потепління. Усе це разом змінює клімат, скорочує кількість природних місць для проживання тварин і рослин, і як результат, є предтечею до початку вимирання глобальної екологічної катастрофи.
Плазуни в порівнянні із земноводними є наступним етапом пристосування хребетних тварин до життя на суші. Найдавніші плазуни — котилозаври — з'явилися в середньому карбоні і ще зберігали в своїй будові ознаки, характерні для земноводних палеозою (стегоцефалів). У подальшому плазуни розділилися на 2 головних стовбури — звіроподібних (що дали в кінці тріасу початок ссавцям) і діапсид (лепідозаврів і архозаврів, які дали початок птахам). Плазуни домінували в наземних біоценозах з перму до початку кайнозою. Найбільшого розквіту досягли в мезозої, коли серед них з'явилися також морські (іхтіозаври, завроптеригії, плакодонти, мозазаври) форми, що літають (птерозаври). Найбільші тварини суші входили до числа динозаврів. Перехід від мезозою до кайнозою співпав з вимиранням всіх перерахованих груп мезозойських плазунів і бурхливою експансією ссавців і птахів — прогресивних нащадків плазунів.
6 підкласів: анапсиди (включаючи сучасних черепах), архозаври (включаючи сучасних крокодилів) і лепідозаври (включаючи сучасних дзьобоголових і лускатих), а також повністю вимерлі іхтіоптеригії, синаптозаври і звіроподібні.
Походження плазунів пов'язане з ароморфозами, що забезпечували розмноження на суходолі: внутрішнє запліднення, запас поживних речовин в яйці, вкритому щільною оболонкою, яка захищала його від висихання. Під час розвитку зародка всередині яйця нагромаджувалась рідина, в якій зародок знаходився як в акваріумі, відбувається формування зародка. Це дало змогу плазунам завоювати всі середовища існування: суходіл, повітря і повторно оселитися у воді. Прогресу плазунів сприяв розвиток рогового покриву, який захищав від висихання, досконаліший розвиток легень, кровоносної системи, кінцівок, головного мозку. Все це дає підстави визнати плазунів першими справжніми наземними хребетними.
Панівного становища набули плазуни в мезозойську еру (близько 180 млн. років тому). Вони заселили сушу (динозаври), водойми (іхтіозавр, плезіозавр), освоїли повітряне середовище (птерозаври). Багато видів динозаврів досягли гігантських розмірів, наприклад, диплодок сягав 30 м завдовжки, а брахіозавр — до 23 м при висоті 12 м. Динозаври були поширені в Європі, Азії, Африці і Північній Америці. На Україні рештки динозаврів знайдено у Криму. До кінця крейдяного періоду динозаври вимерли, причому дуже швидко.
Причини вимирання давніх гігантських плазунів остаточно не з'ясовані. Існує багато гіпотез, автори яких намагаються пояснити причини зникнення цих рептилій (падіння астероїда, отруєння покритонасінними рослинами, спалах нової зірки тощо). Відповідно до одного з припущень, вимирання більшості давніх ящерів було зумовлене багатьма причинами і насамперед — горотворними процесами в кінці крейдяного періоду, які супроводжувалися перебудовою ландшафтів, перерозподілом моря і суші, змінами клімату, що став більш посушливим і континентальним. Крім того, гігантські ящери не могли конкурувати з теплокровними тваринами (птахами і ссавцями), які у той період вже населяли Землю.
1) Берман З. И., Завадский К. М., Зеликман А. Л. и др. История эволюционных учений в биологии. - Л., 1966.
2) Биологический энциклопедический словарь / Гл. ред. М. С. Гиляров; Редкол.: А. А. Баев, Г. Г. Винберг, Г. А. Заварзин и др. — М.: Сов. энциклопедия, 1986. — 831 с.
3) Біологія: Комплексний довідник. /Р. В. Шаламов, Ю. В. Дмитрієв, В. І. Подгорний. — Х.: Веста: Вид-во «Ранок», 2006. - 624 с.
4) Грант В. Эволюция организмов. - М., 1980.
5) Завадский К. М. Развитие эволюционной теории после Дарвина (1859—1920 годы). - Л., 1973.
6) Завадский К. М., Колчинский Э. И. Эволюция эволюции. - Л., 1977.
7) Левонтин Р. Генетические основы эволюции. - М., 1978.
8) Майр Э. Популяции, виды и эволюция. - М., 1974.
9) Наумов С.П. Зология позвоночных. – М.: Просвещение, 1982.
10)Рауп Д., Стэнли С. Основы палеонтологии. - М., 1974.
11)Самарський С.Л. Зологія хребетних. – К.: Вища школа, 1967.
12)Северцов А. С. Основы теории эволюции. - М., 1987.
13)Северцов А.Н. Эволюционная теория. – М., 1967.
14)Согур Л.М. Зоологія: курс лекцій. – К.: Фітосоціоцентр, 2004. – 308 с.
15)Татаринов Л. П. Палеонтология и эволюционное учение. - М., 1985.
16)Тимофеев-Ресовский Н. В., Воронцов Н. Н., Яблоков А. В. Краткий очерк теории эволюции. - М., 1977.
17)Филипченко Ю. А. Эволюционная идея в биологии. - М., 1977.
18)Шмальгаузен И. И. Проблемы дарвинизма. - Л., 1969.
19)Шмальгаузен И. И. Факторы эволюции. Теория стабилизирующего отбора. - М., 1968.
20)Эволюция /Под ред. М. В. Мины. - М., 1981.
21)Яблоков А.В., Юсупов А.Г. Эволюционное учение. – М.: Высш. шк.
|
