Реферат
Понятие гиперцикла
2009
Рассмотрим последовательность реакций, в которой на каждом этапе продукты — с участием дополнительных реагентов или без них — подвергаются дальнейшим превращениям. Если в такой последовательности какой-либо из продуктов идентичен исходному реагенту какой-либо предшествующей стадии, то такая система подобна реакционному циклу, а цикл в целом — катализатору. В простейшем случае катализатор — это отдельная молекула, например фермент, превращающий субстрат в продукт;
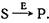
Механизм, который стоит за этой формальной схемой, требует по меньшей мере трехчленного цикла (рис. 1). Более сложные реакционные циклы, выполняющие фундаментальные каталитические функции, показаны на рис. 2 и 3. Цикл Бете — Вайцзекера (рис. 2) вносит существенный вклад в поддержание высокой скорости высвобождения энергии в "массивных звездах. Он, если можно так сказать, заставляет Солнце светить и, следовательно, является одной из наиболее важных внешних предпосылок существования жизни на Земле. Не меньшее значение, по-видимому, имеет цикл Кребса, или цикл лимонной кислоты [6], изображенный на рис. 3, хотя он касается внутреннего механизма живого. Этот цикл реакций опосредует и регулирует метаболизм углеводов и жирных кислот в живой клетке и выполняет также важные функции в анаболических.
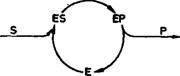
Рис. 1. Общий механизм ферментативного катализа по Михаэлису-Меитен.
В этом процессе участвуют по меньшей мере три интермедиата: свободный фермент (Е), комплекс фермент — субстрат (ES) и комплекс фермент — продукт (ЕР). Схема показывает, что каталитическое действие фермента эквивалентно циклическому воспроизведению интермедиатов при превращении субстрата (S) в продукт (Р). Впрочем, она дает лишь формальное представление истинного процесса, который может включать в себя многоступенчатую активацию субстрата и индуцированные конформационные изменения фермента.
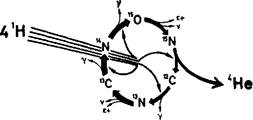
Рис. 2. Углеродный цикл, предложенный Бете и фон Вайцзекером, ответствен — по крайней мере частично — за высвобождение энергии в массивных звездах.
Его компоненты — 12
С, 13
N, 13
С, 14
N, 15
0 и l
5
N — постоянно воспроизводятся в ходе реакционного цикла. Циклическая схема в целом представляет собой катализатор, который превращает четыре атома 'Н в один атом 4
Не с высвобождением энергии в виде Y позитронов (е+) и нейтрино (v).
Реклама


Рис. 3. Цикл трикарбоновых кислот, или цикл лимонной кислоты, — это общий каталитический аппарат биологического окисления «топливных» молекул.
Полная схема цикла была предложена Кребсом, важный вклад в ее построение был внесен также Сеит-Дьёрди, Марциусом и Кнопом. Основные компоненты цикла? цитрат (С), чисаконитат (А), изоцитрат (I), а-кетоглутарат (К), сукцинил-СоА (S*), сукцинат (S), фумарат (F), 1-ма-лат (М) и оксалоацетат (О). Ацетат вступает в цикл в активированной форме в виде ацетил-СоА (этап 1) и реагирует с оксалоацетатом и Н2
0 с образованием цитрата (С) и СоА (+Н+). Во всех превращениях участвуют ферменты и такие кофакторы, как СоА (этапы 1, 5, 6), Fes
+
(этапы 2, 3), NAD+ (этапы 4,5,9), ТПФ, липоевая кислота (этап 5) и FAD (этап 7). На схеме не показаны дополнительные реагенты: Н2
0 (этапы 1, 3, 8), Pt
и QDP (этап 6) и продукты реакций — Н2
0 (этап 2), Н+ (этапы 1, 9) и QTP (этап 6). В результате всех реакций цикла происходит полное окисление двух ацетильных углеродов до CQ2
(и Н2
0). За одни оборот цикла генерируется 12 макроэргических фосфатных связей, причем одна из них образуется в самом цикле (QTP, этап 6), а 11—в результате окисления NADH и FADH2
[три пары электронов переносятся к NAD+ (этапы 4, 5,9) и одна пара — к FAD (этап 7)].
Однонаправленность циклического воспроизведения интермедиатов, конечно, предполагает, что система находится в состоянии, далеком от равновесия, и всегда связана с расходованием энергии, часть которой диссипирует в окружающую среду. С другой стороны, установление равновесия в замкнутой системе приведет к тому, что все стадии будут находиться в состоянии детального равновесия. Каталитическое действие в такой замкнутой системе будет микроскопически обратимо, т. е. оно будет происходить с равной эффективностью в обоих направлениях.
Теперь, пользуясь простой итерацией, построим иерархии реакционных циклов и зададим их частные свойства. Это означает, что на следующем этапе мы рассматриваем реакционный цикл, в котором по меньшей мере один, но, возможно, и все интермедиаты сами являются катализаторами. Отметим, что эти интермедиаты, будучи катализаторами, теперь остаются неизменными во время реакции. Каждый из них образуется из потока высокоэнергетического строительного материала с использованием каталитической поддержки от предшествующего интермедиата (рис. 4). При большом числе интермедиатов такая система становится очень сложной по составу и поэтому вряд ли может встретиться в природе. Самым известным примером является четырехчленный цикл, связанный с матричной репликацией молекулы РНК (рис. 5). Такого рода механизм исследовался invitro: использовали реакционную среду, содержащую нуклеозидтрифосфаты четырех типов в качестве высокоэнергетических строительных материалов, а также фаговую репликазу, которая играла роль постоянного фактора среды [7, 8] (более детальное описание дано Кюпперсом [9]). Каждая из двух цепей действует как матрица, инструктирующая синтез своей комплементарной копии,— как при фотографической репродукции.
Реклама

Простейшим представителем этой категории реакционных систем является отдельный автокатализатор или — в случае целого класса объектов I,-, несущих информацию,— самовоспроизводящаяся единица. Такой процесс формально может быть описан следующим образом:
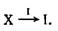
В этом реферате будут часто рассматриваться реакции такого типа; мы будем обозначать их символом 
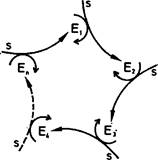
Рис. 4. Каталитический цикл представляет собой более высокий уровень организации в иерархии каталитических схем.
Компоненты цикла сами являются катализаторами, которые образуются из каких-либо высокоэнергетическнх субстратов (S), причем каждый интермедиат является катализатором для образования E+i. Каталитический цикл в целом эквивалентен автокатализатору, инструктирующему свое собственное воспроизведение. Для существования каталитического цикла достаточно, чтобы одни из интермедиатов был катализатором для одной из последующих реакций.
Такой истинно самовоспроизводящейся формой является двухцепочечная ДНК (в отличие от одноцепочечной РНК) — обе ее цепи одновременно копируются полимеразой (рис. 6). Эта же формальная схема применима и в случае прокариотической клетки, где все наследование осуществляется в основном независимо в каждой клеточной линии.
Кинетические уравнения как простых каталитических, так и автокаталитических систем при постоянной концентрации субстрата содержат член первого порядка по концентрации катализатора. Кривые роста, однако, четко дифференцируют две эти системы. Количество продукта для простого каталитического процесса в указанных условиях растет во времени линейно, тогда как для автокаталитических систем характерен экспоненциальный рост.
Строго говоря, автокаталитическую систему уже можно называть гиперциклической, ибо она представляет собой цикл катализаторов, которые сами являются циклами реакций. Мы, однако, ограничим использование этого термина такими системами, которые являются гиперциклическими по отношению к каталитической функции. В действительности это гиперциклы второй или более высокой степени, потому что они относятся к реакциям по меньшей мере второго порядка по концентрациям катализаторов.
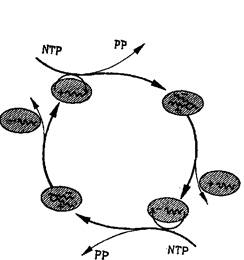
Рис. 5. Каталитический цикл, имеющий биологическое значение, — репликация одноцепочечной РНК.
Интермедиаты (плюс- и минус-цепи) участвуют в цикле как матрицы для своего взаимного воспроизведения. Матричная функция эквивалентна избирательному катализу. Нуклеозидтрифосфаты (NTP) представляют собой высокоэнергетический строительный материал, а пирофосфат (РР) является отходом реакций цикла. Комплементарное инструктирование, механизм которого будет обсуждаться в связи с рис. 11, является существенно автокаталитической функцией, т. е. функцией самовоспроизведения.
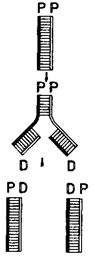
Рис. 6. Истинный процесс самовоспроизведения (одночленный каталитический цикл) — репликация ДНК.
Механизм этого чрезвычайно сложного процесса (см. рис. 12) обеспечивает связь каждой дочерней цепи (D) с соответствующей родительской цепью (Р).
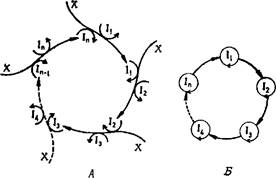
Рис. 7. Каталитический гиперцикл состоит из самоинструктируюшихся единиц Uс двойными каталитическими функциями.
В качестве автокатализатора или — в более общем случае—в качестве каталитического цикла ннтермедиат Uспособен инструктировать свое собственное воспроизведение, а кроме того, он оказывает каталитическое содействие воспроизведению следующего интермедиата (с использованием высокоэнергетичеекого строительного материала X). Упрощенная схема (Б) показывает иерархию циклов.
Каталитический гиперцикл — это система, связывающая автокаталитические, или самовоспроизводящиеся единицы друг с другом посредством циклической связи. Такая система изображена на рис. 7. как самовоспроизводящиеся единицы сами являются каталитическими циклами, например комбинациями плюс- и минус-цепей РНК, как показано на рис. 5. Однако процесс репликации как таковой должен прямо или косвенно стимулироваться дополнительными специфическими связями между различными репликативными единицами.
Реально такие связи могут обеспечиваться белками — продуктами трансляции предшествующих циклов воспроизведения РНК (рис. 8). Эти белки могут действовать как специфические репликазы или дерепрессоры или же как специфические защитные факторы, предохраняющие от деградации. Связи между самовоспроизводящимися единицами должны образовывать наложенный цикл — лишь тогда система в целом будет подобна гиперциклу. По сравнению с системами, показанными на рис. 4 и 5, гиперцикл является самовоспроизводящимся в более высокой степени.
Простейшим представителем систем этого класса снова является (квази)одношаговая система, т.е. усиленный автокатализатор. Мы сталкиваемся с такой системой в случае инфекции клетки РНК.
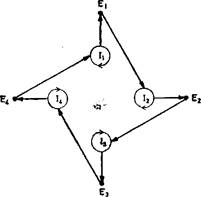
Рис. 8. Реалистическая модель гиперцикла второй степени, где носители информации «выдают» инструкции двух типов: во-первых, для собственного воспроизведении и, во-вторых, для трансляции с образованием интермедиатов второго типа, которые обладают оптимальными функциональными свойствами.
Каждый фермент Е; катализирует воспроизведение следующего носителя информации I;. Он может иметь и другие каталитические свойства, необходимые для осуществления процесса трансляции, метаболизма и т.д. В подобных случаях степень гиперциклической связи больше двух.
Если фаговая РНК (плюс-цепь) проникает в бактериальную клетку, то ее генотипическая информация транслируется аппаратом клетки-хозяина. Один из продуктов трансляции связывается с определенными факторами хозяина с образованием активного ферментного комплекса, который специфически реплицирует плюс- и минус-цепи фаговой РНК, действующие при воспроизведении как матрицы друг для друга [11]. Однако репликазный комплекс не размножает — в сколько-нибудь ощутимой степени — информационную РНК клетки-хозяина. В результате инфекции начинается гиперболический рост числа фаговых частиц, который в конце концов выходит на насыщение из-за ограниченности ресурсов клетки-хозяина.
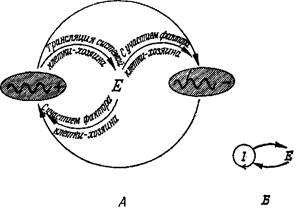
Рис. 9. Размножение РНК-содержащего фага в бактериальной клетке представляет собой простой гиперциклнческий процесс.
Инфекционная плюс-цепь, используя аппарат трансляции клетки-хозяина, сначала инструктирует синтез белковой субъединицы (Е), которая связывается с другими белками клетки-хозяииа с образованием фагоспецифичной РНК-репликазы. Этот репликазный комплекс узнает только определенные фенотипические особенности фаговой РНК, которые имеются как у плюс-, так и у минус-цепи благодаря симметрии определенных участков цепи РНК. В результате происходит вспышка синтеза фаговой РНК, которая — из-за гиперциклической природы — носит гиперболический характер (см. часть Б, рис. 17), пока не наступит насыщение по одному из интермедиатов или пока не истощатся запасы метаболитов клетки-хозяина. Схема Б показывает, что автокаталитическими илк самоинструктивиыми свойствами может обладать всего один из интермедиатов при условии, что другие «партнеры» связаны с ним с помощью циклической обратной связи.

Рис. 10. Иерархия циклических реакционных сетей очевидна из сопоставления этих схем (— химическое превращение, —* каталитическое действие).
Другой естественный гиперцикл может возникнуть в менделеевских популяциях в начальной фазе видообразования — пока численности популяций малы. Для репродукции генов необходимо взаимодействие между двумя аллелями (М и F), т. е. между гомологичными участками в мужской и женской хромосомах, которые затем появляются в потомстве в перестроенной комбинации. Тот факт, что кинетические уравнения популяционной генетики менделеевских популяций обычно не содержат гиперциклической нелинейности (которая ведет к гиперболическому, а не к экспоненциальному росту), объясняется насыщением, наступающим при сравнительно малых численностях популяций, когда скорость рождений (обычно) становится пропорциональной только численности самок.
Как видно из сравнения схем, показанных на рис. 10, гиперциклы представляют собой новый уровень организации. Этот факт проявляется в их уникальных свойствах. Не связанные друг с другом самовоспроизводящиеся единицы гарантируют сохранение ограниченного количества информации, которое может передаваться от поколения к поколению. Это оказывается одной из необходимых предпосылок дарвиновского поведения, т. е. отбора и эволюции. Подобно им, гиперциклы тоже селекционны, но, кроме того, они имеют интегрирующие свойства, которые делают возможной кооперацию между единицами, конкурирующими в других условиях. Более того, гиперциклы конкурируют даже еще более ожесточенно, чем дарвиновские виды, с любой самовоспроизводящейся единицей, не являющейся частью гиперцикла. Далее, они способны устанавливать глобальные формы организации благодаря свойству отбирать «раз и навсегда», которое не допускает сосуществования с другими гиперциклическими системами, если только они не стабилизированы связями более высокого порядка.
Простейший тип связи внутри гиперцикла — это прямая стимуляция или дерепрессия, которым в кинетических уравнениях соответствуют члены второго порядка для процессов образования. Члены для связей более высокого порядка могут войти тоже, и они, таким образом, определяют степень р гиперциклической организации.
Отдельные гиперциклы могут также связываться друг с другом с образованием иерархий. Это, однако, требует введения членов интерциклической связи, которые существенно зависят от степени организации. Чтобы установилось устойчивое сосуществование, для гиперциклов H1
и Н2
, имеющих степени внутренней организации р1
и р2
, необходимы члены интерциклической связи степени.
Цель этой работы состоит в том, чтобы разработать детальную теорию той категории сетей реакций, которые мы окрестили гиперциклами, и обсудить их значимость для биологической самоорганизации, особенно в связи с возникновением трансляции — события, которое можно считать решающим этапом в доклеточной эволюции.
|
