Реферат
Отдел голосемянные,
тип одноклеточных животных из группы эукариотов
Отдел Голосеменные
Что же такое семя, каково его биологическое и систематическое значение?
Общая характеристика.
Первые голосеменные появились в конце девонского периода около 350 млн. лет назад; вероятно, они произошли от древних папоротниковидных, вымерших в начале каменноугольного периода. В мезозойскую эру - эпоху горообразования, поднятия материков и иссушения климата - голосеменные достигли расцвета, но уже с середины мелового периода уступили свое господствующее положение покрытосеменным.
Отдел современных голосеменных насчитывает более 700 видов. Несмотря на относительно малую численность видов, голосеменные завоевали почти весь земной шар. В умеренных широтах Северного полушария они на огромных пространствах образуют хвойные леса, называемые тайгой.
Современные голосеменные представлены преимущественно деревьями, значительно реже - кустарниками и очень редко - лианами; травянистых растений среди них нет. Листья голосеменных значительно отличаются от других групп растений не только по форме и размерам, но и по морфологии и анатомии. У большинства видов они игловидные (хвоя) или чешуевидные; у отдельных представителей они крупные (например, у вельвичии удивительной их длина достигает 2-3 м), перисторассеченные, двулопастные и др. Листья располагаются поодиночке, по два или несколько в пучках.
Водопроводящая система (ксилема) голосеменных состоит преимущественно из трахеид, и лишь у некоторых групп имеются настоящие сосуды.
Подавляющее большинство голосеменных - вечнозеленые, одно - или двудомные растения с хорошо развитыми стеблем и корневой системой, образованной главным и боковым корнями. Расселяются они семенами, которые формируются из семязачатков. Семязачатки голые (отсюда название отдела), расположены на мегаспорофиллах или на семенных чешуях, собранных в женские шишки.
В цикле развития голосеменных наблюдается последовательная смена двух поколений - спорофита и гаметофита с господством спорофита. Гаметофиты сильно редуцированы, причем мужские гаметофиты голо - и покрытосеменных растений не имеют антеридиев, чем резко отличаются от всех разноспоровых бессеменных растений.
Реклама

Голосеменные включают шесть классов, два из которых полностью исчезли, а остальные представлены ныне живущими растениями. Наиболее сохранившейся и самой многочисленной группой голосеменных является класс Хвойные, насчитывающий не менее 560 видов, образующих леса на обширных пространствах Северной Евразии и Северной Америки. Наибольшее число видов сосны, ели, лиственницы встречается у побережий Тихого океана.
Класс Хвойные. Все хвойные - вечнозеленые, реже листопадные (например, лиственница) деревья или кустарники с игольчатыми или чешуевидными {например, у кипариса) листьями. Игловидные листья (хвоя) плотные, кожистые и жесткие, покрыты толстым слоем кутикулы. Устьица погружены в углубления, заполненные воском. Все эти особенности строения листьев обеспечивают хорошее приспособление хвойных к произрастанию как в засушливых, так и в холодных местообитаниях.
У хвойных прямостоячие стволы, покрытые чешуйчатой корой. На поперечном разрезе стебля хорошо видны развитая древесина и менее развитые кора и сердцевина. Ксилема хвойных на 90-95% образована трахеидами. Шишки хвойных раздельнополые; растения - чаще однодомные, реже - двудомные.
Наиболее широко распространенными представителями хвойных в Беларуси и России являются сосна обыкновенная и ель обыкновенная, или европейская. Их строение, размножение, чередование поколений в цикле развития отражает характерные особенности всех хвойных.
Сосна обыкновенная - однодомное растение (рис.9.3). В мае у основания молодых побегов сосны образуются пучки зеленовато-желтых мужских шишек длиной 4-6 мм и диаметром 3-4 мм. На оси такой шишки расположены многослойные чешуйчатые листочки, или микроспорофиллы. На нижней поверхности микроспорофиллов находятся два микроспорангия - пыльцевых мешка, в которых образуется пыльца. Каждое пыльцевое зерно снабжено двумя воздушными мешками, что облегчает перенос пыльцы ветром. В пыльцевом зерне имеются две клетки, одна из которых впоследствии, при попадании на семязачаток, формирует пыльцевую трубку, другая после деления образует два спермия.
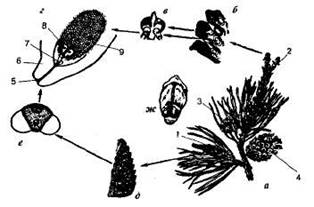
Цикл развития сосны обыкновенной: а - ветка с шишками; б - женская шишка в разрезе; в - семенная чешуя с семязачатками; г - семязачаток в разрезе; д - мужская шишка в разрезе; е - пыльца; ж - семенная чешуя с семенами; 1 - мужская шишка; 2 - молодая женская шишка; 3 - шишка с семенами; 4 - шишка после высыпания семян; 5 - пыльцевход; 6 - покров; 7 - пыльцевая трубка со спермиями; 8 - архегоний с яйцеклеткой; 9 - эндосперм.
Реклама

На других побегах того же растения образуются женские шишки красноватого цвета. На их главной оси располагаются мелкие прозрачные кроющие чешуйки, в пазухах которых сидят крупные толстые, впоследствии одревесневающие чешуи. На верхней стороне этих чешуй расположено по два семязачатка, в каждом из которых развивается женский гаметофит - эндосперм с двумя архегониями с крупной яйцеклеткой в каждом из них. На верхушке семязачатка, снаружи защищенного интегументом, имеется отверстие - пыльцевход, или микропиле.
Поздней весной или в начале лета созревшая пыльца разносится ветром и попадает на семязачаток. Через микропиле пыльца втягивается внутрь семязачатка, где и прорастает в пыльцевую трубку, которая проникает к архегониям. Образовавшиеся к этому времени два спермия по пыльцевой трубке попадают к архегониям. Затем один из спермиев сливается с яйцеклеткой, а другой отмирает. Из оплодотворенной яйцеклетки (зиготы) формируется зародыш семени, а семязачаток превращается в семя. Семена у сосны созревают на второй год, высыпаются из шишек и, подхваченные животными или ветром, переносятся на значительные расстояния.
По своему значению в биосфере и роли в хозяйственной деятельности человека хвойные занимают второе место после покрытосеменных, далеко превосходя все остальные группы высших растений.
Они помогают решать огромные водоохранные и ландшафтные задачи, служат важнейшим источником древесины, сырья для получения канифоли, скипидара, спирта, бальзамов, эфирных масел для парфюмерной промышленности, лекарственных и других ценных веществ. Некоторые хвойные культивируются как декоративные (пихты, туи, кипарисы, кедры и др.). Семена ряда сосен (сибирской, корейской, итальянской) употребляются в пищу, из них также получают масло.
Представители других классов голосеменных (саговниковые, гнетовые, гинкговые) встречаются значительно реже и менее известны, чем хвойные. Однако почти все виды саговниковых декоративны и пользуются широкой популярностью у садовников многих стран. Вечнозеленые безлистные невысокие кустарники эфедры (класс гнетовых) служат источником сырья для получения алкалоида эфедрина, который применяется как средство, возбуждающее центральную нервную систему, а также при лечении заболеваний аллергического характера.
Сейчас на Земле произрастают представители 4 классов голосеменных. Из них Гинкговые (единственный вид - Ginkgo biloba) и Саговниковые (120-130 видов 10 тропических родов) - это остатки процветавших в мезозое групп. Возраст и происхождение Гнетовых (71 вид 3 родов) неясны из-за отсутствия палеоботанических данных, и лишь класс Хвойных (Pinopsida) широко представлен ныне на нашей планете, по своему значению превосходя все остальные группы высших растений. В настоящее время насчитывается не менее 560 видов из 55 родов хвойных, распространенных в умеренных областях обоих полушарий. Подавляющее большинство представителей этого класса - вечнозеленые древесные растения, возраст и размеры которых потрясают воображение. Так, известны экземпляры секвойи вечнозеленой высотой более 100 м и диаметром ствола до 11 м. Некоторые экземпляры мамонтова дерева достигают диаметра 12 м, а таксодиума мексиканского - 16 м. Рекорд долгожительства принадлежит североамериканской сосне долговечной - до 4900 лет! Кроме деревьев, широко встречаются кустарниковые и даже кустарничковые хвойные (новозеландская сосна-пигмей).
Древняя группа (отдел) семенных растений, занимающая промежуточное положение между папоротниками и покрытосеменными (цветковыми) растениями. От папоротников отличаются наличием семезачатков (семяпочек), от покрытосеменных - отсутствием плодолистиков.
Все современные виды голосеменных имеют стробилы - укороченные и ограниченные в росте репродуктивные побеги. Большинство голосеменных - деревья или кустарники, нередко достигающие огромных размеров. Некоторые сильно разветвлены и несут много небольших или мелких, часто чешуевидных листьев (кордаитовые, гинкговые и хвойные), другие не ветвятся или ветвятся очень слабо и имеют относительно крупные, большей частью перистые, листья (лигиноптеридовые, беннеттитовые, цикадовые и вельвичия). Листья сильно различаются не только по числу и размерам, но и по анатомическому строению.
Голосеменные растения известны с верхнего девона; в каменноугольном и пермском периодах уже встречаются представители большинства порядков голосеменных, достигающих своего расцвета в мезозойскую эру. Ныне сохранилось только ок.600 видов.
Класс Гинкговые (Ginkgoopsida) ИЛИ Гинкгопсиды класс
Единственным современным представителем класса гинкговых является реликтовое растение - гинкго двулопастный (Ginkgo biloba). Оно было открыто для науки в 1690 г. в Японии врачом голландского посольства Е. Кемпфером и в 1712 г. описано им же под названием Ginkgo, что в переводе с японского означает "серебряный абрикос" или "серебряный плод". Так назывались продававшиеся в японских лавках съедобные семена этого дерева. С давних пор деревья гинкго, как весьма почитаемые и священные, растут во многих парках, окружающих старинные храмы в Японии, Китае и Корее. Около 1730 г. гинкго был ввезен в Западную Европу, а примерно лет через 50 - в Северную Америку. С этого времени гинкго неизменно пользуется пристальным вниманием и ботаников, и садоводов. Автором научного названия "гинкго" является Карл Линней. Один из английских садоводов послал необычное растение великому естествоиспытателю, и в 1771 г. Линней ввел его в ботаническую литературу под латинским названием Ginkgo biloba. Гинкго двулопастный - высокое дерево, достигающее более 30 м в высоту и более 3 м в диаметре. Молодые деревья имеют пирамидальную крону, с возрастом крона становится более раскидистой. Боковые ветви отходят от ствола почти под прямым углом, иногда они сближены, образуя подобие мутовок. Кора серая, шероховатая, у старых деревьев - с продольными трещинами. Основную массу ствола гинкго, как и у хвойных, составляет древесина, сердцевина развита слабо, кора узкая. Годичные кольца выражены довольно хорошо, хотя и не так четко, как у большинства хвойных. Трахеиды вторичной ксилемы имеют 1-2 ряда округлых окаймленных пор в супротивном, реже, в очередном расположении. Сердцевинные лучи однорядные, низкие. Существенным отличием от подавляющего большинства хвойных является отсутствие у гинкго способности к образованию смолы. Листья гинкго располагаются на побегах двух типов - на удлиненных конечных побегах, растущих быстро, и на укороченных побегах (брахибластах), отличающихся замедленным ростом. На длинных побегах листья одиночные, сидят по спирали. Брахибласты, как у некоторых хвойных (лиственница, кедр.), образуются на длинных побегах из пазушных почек листьев. Удлиненные и укороченные побеги отличаются и по анатомическому строению. В частности, у укороченных побегов сердцевина и кора занимают относительно большую площадь поперечного сечения. На верхушке укороченных побегов у гинкго образуется пучок из 5-7 листьев. Ни одно из современных голосеменных растений не имеет листья, похожие на листья гинкго. Они имеют вееровидную или широко клиновидную кожистую пластинку, пронизанную многократно дихотомирующими жилками, чем напоминают листья некоторых папоротников. Устьица на нижней стороне листа гаплохейльные. Черешок листа тонкий, упругий, длиной до 10 см. Пластинка большинства листьев более или менее глубоким срединным V-образным вырезом рассечена на две симметричные половинки. Эта их особенность отражена в видовом эпитете (от лат. biloba - двулопастный). Листья молодых экземпляров гинкго, особенно проростков, а также побегов прикорневой поросли, более сильно изрезаны, бывают 4-8-лопастными и нередко имеют клиновидное основание. Такие листья очень похожи на листья древних, вымерших представителей рода гинкго. Гинкго относится к числу немногих листопадных голосеменных. Ежегодно поздней осенью деревья сбрасывают листья, которые незадолго до этого приобретают привлекательный золотисто-желтый цвет. Гинкго - двудомное растение. Его мужские и женские репродуктивные органы образуются на разных деревьях. Впрочем, садоводы имеют возможность отчасти "исправить эту оплошность природы" путем прививки побегов с женского экземпляра на мужской и наоборот. В стадию зрелости деревья гинкго входят довольно поздно. При благоприятных условиях они начинают образовывать пыльцу и семена на 25-30-й год жизни. До этого времени неизвестно, какой перед вами экземпляр - мужской или женский. Микро - и мегастробилы гинкго образуются на укороченных побегах. Закладываются они в конце лета, но созревают в начале следующего вегетационного сезона. Ранней весной, когда еще не вполне распустились листья, на взрослых экземплярах гинкго появляются стробилы.
Микростробилы гинкго сережковидные и состоят из оси и спирально сидящих на ней микроспорофиллов. Микроспорофилл представляет собой тонкую ножку, на конце которой расположено два, реже 3-4 висячих микроспорангия. В микроспорангиях образуются многочисленные микроспоры, очень похожие на микроспоры саговниковых. Они однобороздные, эллипсоидальные или почти округлые. Развитие мужского гаметофита в микроспоре начинается еще внутри микроспорангия.
Ко времени раскрывания спорангия, а это происходит обычно в конце апреля - начале мая, пыльцевые зерна состоят, как правило, уже из четырех клеток: первой и второй проталлиальных клеток, генеративной клетки и клетки-гаустории. В таком состоянии пыльцевые зерна рассеиваются ветром и переносятся на семязачатки. К этому времени семязачаток выделяет каплю опылительной жидкости, к которой пыльца приклеивается. Зрелые мегастробилы гинкго состоят из длинной ножки и сидящих на ней двух семязачатков, из которых, как правило, развивается только один.
На одном укороченном побеге среди листьев может образоваться до 7 мегастробилов. В основании каждого семязачатка имеется валик, который называют воротничком. Изредка встречаются мегастробилы с 3-4 и более (до 15) семязачатками. Такие случаи обычно трактуются как свидетельство происхождения редуцированного мегастробила гинкго двулопастного от мегастробилов предковых форм, имевших большое количество семязачатков. Обратимся к биологическим процессам, которые происходят в семязачатке гинкго по мере его развития. Глубоко в ткани нуцеллуса, ближе к его основанию, закладывается материнская клетка мегаспоры. После редукционного деления из нее образуется линейная тетрада клеток. Дальнейшему развитию подвергается только нижняя из них. Она-то и становится функционирующей мегаспорой, из которой развивается женский гаметофит.
Остальные клетки тетрады дегенерируют. Исследования ультраструктуры материнской клетки мегаспоры показали, что перед делением в ней происходят сложные преобразования и перераспределение некоторых органелл. В частности, большая часть пластид и митохондрий концентрируется преимущественно в нижней (халазальной) части этой клетки, из которой образуется функционирующая мегаспора. На протяжении почти месяца в мегаспоре происходит свободное деление ядер. В результате в ней образуется более 8000 ядер. Примерно в середине июня вокруг ядер начинают формироваться клетки. Образование клеточных стенок осуществляется центростремительно, т.е. сначала формируется наружный слой ткани гаметофита и затем постепенно клетками заполняются внутренние участки.
Рыхлая ткань нуцеллуса постепенно абсорбируется разрастающимся гаметофитом. В конце июня на верхушке образовавшегося массивного гаметофита обычно формируются два архегония. В пространстве между ними в середине августа появляется столбикообразный выступ. Над этим участком разрушается оболочка мегаспоры, архегонии и столбикообразный выступ оказываются на дне пыльцевой камеры. Архегонии тем временем продолжают развиваться. Центральная клетка архегония делится неравновелико на верхнюю маленькую брюшную канальцевую клетку и располагающуюся под ней крупную клетку, которая становится яйцеклеткой. Брюшная канальцевая клетка недолговечна и вскоре разрушается. Задолго до процесса оплодотворения в гаметофите начинают накапливаться питательные вещества - крахмал, липиды, липопротеины. Аккумулируются они и в яйцеклетке. То же самое, как было сказано выше, характерно и для саговниковых.
Другой биологической особенностью гинкго, отличающей его от остальных голосеменных, кроме саговниковых, является ранняя дифференциация интегумента, которая происходит также задолго до оплодотворения. Вполне развитый интегумент семязачатка гинкго состоит из трех слоев. Примыкающий к нуцеллусу внутренний, самый тонкий слой (эндотеста) по внешнему виду напоминает тончайшую, пергаментную бумагу. Средний слой (склеротеста) твердый, сильно лигнифицированный, толщиной до 0,5 мм, образует "косточку" созревшего семязачатка, а впоследствии - семени.
Наружный слой интегумента (саркотеста) мясистый, сочный, достигает в толщину 5-6 мм и покрыт кутинизированной эпидермой с устьицами. Осенью саркотеста окрашивается в красивый янтарно-желтый цвет, но одновременно приобретает неприятный запах из-за присутствия масляной кислоты (которая, как известно, содержится и в прогорклом сливочном масле). По степени развития интегумента у гинкго, как и у саговниковых, невозможно отличить стерильный семязачаток от оплодотворенного, не сделав поперечного среза.
У остальных современных голосеменных полная дифференциация интегумента и отвердение склеротесты происходит только после образования зародыша. Вернемся к процессам, происходящим внутри семязачатка. Пыльцевые зерна, проникшие весной во время опыления в пыльцевую камеру, продолжают находиться в ней, плавая в опылительной жидкости. После прорастания пыльцевого зерна первая проталлиальная клетка разрушается, а генеративная клетка делится, образуя сперматогенную клетку и клетку-ножку. В дальнейшем развивается только сперматогенная клетка. Путем деления из нее образуются два подвижных сперматозоида. Один из концов сперматозоида заключает блефаропласт-своеобразное опорное образование, к которому прикрепляются многочисленные жгутики. Сперматозоиды очень эластичны, подвижны и "выплывают" через отверстие гаустории. Одновременно верхушка яйцеклетки образует небольшое выпячивание, проникающее между клетками шейки архегония.
Как только плавающий в пыльцевой камере сперматозоид коснется шейки, это выпячивание втягивается внутрь, как бы прокладывая дорогу сперматозоиду. Однако в яйцеклетку проникает только головка сперматозоида. В результате слияния ядер сперматозоида и яйцеклетки образуется зигота с диплоидным набором хромосом: 2п = 24. Гинкго - первое голосеменное растение, у которого были открыты подвижные сперматозоиды.
Честь этого открытия принадлежит японскому исследователю С. Xиразе. Работая художником в лаборатории ботаники Токийского университета, он увлекся ботаникой и начал изучать эмбриогенез гинкго. В 1896 г., сначала на микроскопических препаратах, а затем и в прижизненном состоянии он открыл подвижные сперматозоиды. И до сих пор в ботаническом саду Токийского университета растет огромное дерево гинкго, с которого Хиразе брал материал, приведший его к блестящему открытию. Около дерева - памятная доска. Оплодотворение с помощью подвижных сперматозоидов сближает гинкго с саговниковыми, а точнее, показывает, что и та и другая группа голосеменных в этом отношении стоят на одинаково низкой ступени эволюции.
Развитие зародыша, а нередко и оплодотворение происходит у гинкго уже в опавших с дерева семязачатках. Это тоже одна из архаичных особенностей этого растения, по-видимому сближающая его с вымершими семенными папоротниками и кордаитовыми. Семена гинкго не имеют периода покоя (тоже архаичный признак!) и могут прорасти, как только зародыш достигнет своего максимального развития. При наиболее благоприятном сочетании температурных условий и влажности это случается не ранее, чем через три месяца после оплодотворения. В природе семена гинкго сохраняют способность к прорастанию около года при условии их пребывания на достаточно влажном субстрате. При прорастании, как и у саговниковых, семядоли не выходят на дневную поверхность, а остаются внутри семени (подземное прорастание). Первые два листа обычно чешуевидные, третий и последующие листья - лопастные.
Из приведенного описания видно, что гинкго обладает целым рядом существенных, большей частью архаичных, особенностей, отличающих его от хвойных. Гинкго - одно из самых примитивных голосеменных растений современного растительного мира. По уровню эволюционного развития, как было показано, он ближе всего к саговниковым. Род гинкго с единственным сохранившимся представителем по праву выделяют в самостоятельное семейство гинкговых (Ginkgoaceae), а это семейство, в свою очередь, в самостоятельный порядок и класс. Но в мезозойскую эру этот род был представлен по крайней мере несколькими десятками видов и занимал огромный, почти космополитный ареал. По данным геологической летописи, история рода гинкго начинается в триасе. О существовании и распространении гинкго в прошлые геологические эпохи обычно судят по находкам ископаемых листьев. Репродуктивные органы, похожие на соответствующие структуры современного гинкго, также встречаются в ископаемом состоянии, но очень редко. Не менее редки достоверные ископаемые древесины гинкго. Палеоботанические данные свидетельствуют о том, что морфологическая эволюция листа гинкго шла в сторону уменьшения рассеченности листовой пластинки. Триасовые и большинство юрских гинкго имели листья, рассеченные на 4-12 и более четко оформленных лопастей. В середине юрского периода начинают появляться гинкго с почти цельной, лишь по краю неглубоко надрезанной листовой пластинкой. В раннем мелу цельнолистные гинкго встречаются уже довольно часто, одновременно сокращается количество видов с рассеченными листьями. В позднем мелу и третичном периоде остаются только гинкго с цельной или мало рассеченной пластинкой листа. К гинкговым с большей или меньшей степенью достоверности относятся еще несколько родов, также известных главным образом по листовым остаткам.
Это - байера (Baiera), эретмофиллум (Eretmophyllum), гинкгодиум (Ginkgodium), глоссофиллум (Glossophyllum), сфенобайера (Sphenobaiera), тореллия (Torel-lia). Самый древний из них - сфенобайера - появляется в пермских отложениях, т.е. в конце палеозойской эры. Расцвет гинкговых приходится на юрское и раннемеловое время, причем они были распространены преимущественно в северном полушарии на территории, входившей в зону умеренного и теплоумеренного климата с достаточно высокой степенью влажности.
В это время существовали и род гинкго, и все перечисленные выше роды предполагаемых гинкговых (кроме палеогеновой тореллии). Особенно много гинкговых было на территории нынешней Сибири. Пожалуй, нет ни одной юрской или раннемеловой сибирской флоры, где бы не встретились те или иные представители древних гинкговых. Наиболее широко были распространены роды гинкго, байера и сфенобайера. Нередко отпечатки листьев гинкговых переполняют плоскости наслоения геологических пород, наподобие того, как во время осенних листопадов листья устилают почву в лесу.
Такое обилие остатков позволяет думать, что в юрский период и раннемеловую эпоху растения этой группы были одними из основных лесообразующих листопадных пород умеренной и теплоумеренной зон северного полушария. Листья древних представителей гинкговых классифицируются по форме листовой пластинки, ее размерам и рассеченности, густоте жилкования, длине черешка, наличию или отсутствию секреторных каналов. Многие древние виды гинкговых, как и современный гинкго двулопастный, характеризуются значительной изменчивостью листьев. Но как бы она ни была велика, строение эпидермы листьев в пределах вида остается одинаковым. С другой стороны, сходные листья разных видов обнаруживают явные различия в строении эпидермы: в характере клеточных стенок, степени погруженности устьиц, особенностях трихомных образований. На границе ранне- и позднемеловой эпох, во время коренной перестройки растительности, вместе с другими группами типично мезозойских растений вымирает и большинство родов гинкговых. Начиная со второй половины мелового периода фактически остается только род гинкго. И хотя в позднемеловое и третичное время гинкго уже не играл сколько-нибудь значительной роли в растительном покрове Земли, все же ареал рода был еще весьма обширен. Третичные местонахождения гинкго известны даже на Аляске, в Гренландии, на Шпицбергене. Самый молодой из известных нам ископаемых гинкго - гинкго Флорина (Ginkgo florinii) - происходит из плиоценовых отложений Западной Европы и по строению эпидермы обнаруживает значительное сходство с современным видом. В четвертичный период в результате резкого похолодания, вызванного наступлением ледников, род гинкго был уже на грани полного исчезновения. В естественных условиях гинкго двулопастный сохранился ныне, очевидно, только на небольшой территории в Восточном Китае, в горах Дянь Му-Шань, вдоль границы между провинциями Чжэцзян и Аньхой, где он образует леса вместе с хвойными и широколиственными породами. В Китае, Японии и Корее гинкго известен с незапамятных времен. Он упоминается в китайских книгах VII и VIII вв., в китайских поэмах начала XI в., а в солидной медицинской монографии Ли Шичженя, изданной в Китае в XVI в., уже приведены описание и рисунок этого растения. Семена гинкго находят применение в китайской медицине. Кроме того, они издавна употребляются на Востоке в пищу (в отваренном или жареном виде), а мягкую древесину гинкго, легко поддающуюся обработке, используют в кустарных промыслах. В настоящее время гинкго двулопастный растет почти во всех ботанических садах и многих парках субтропической и теплоумеренной зон Европы и Северной Америки, как бы восстанавливая значительную часть ареала своих предков. В 1818 г. гинкго начали выращивать в Крыму, в Никитском ботаническом саду. В европейской части России гинкго хорошо растет и дает всхожие семена до широты Киева. Гинкго - весьма долговечное растение. В Китае, Японии и Корее известно много деревьев гинкго, возраст которых превышает 1000 лет, так что при благоприятных условиях деревья гинкго могут служить многим поколениям людей. Гинкго легко размножается как семенами, так и черенками и является чрезвычайно перспективным растением для озеленения южных городов. Он устойчив к промышленному задымлению воздуха, а также к грибковым и вирусным заболеваниям, редко поражается насекомыми. Деревья гинкго живописны и привлекательны с ранней весны до глубокой осени, одинаково хороши и для одиночных посадок и для создания тенистых аллей.
Гнетовые
Отдел
гнетовых
(Gnetophyta) содержит три своеобразных семейства голосеменных растений с неясными эволюционными связями. Растения рода
гнетум
- тропические лианы.
Эфедровые
(хвойники) - пустынные кустарники с чешуевидными листьями.
Вельвичия
- единственный представитель семейства вельвичиевых - имеет погруженный в песок стебель, от которого отходят два огромных лентовидных листа.
Гнетовые похожи на, однако отличаются от них более совершенной формой шишки, похожей на цветок.
Саговниковые
Саговниковые - причудливые растения, появившиеся не позднее 285 млн. лет назад, где-то в начале пермского периода. Они были столь многочисленны в мезозое, что это время образно назвали "временем саговников и динозавров". Ныне сохранилось всего 10 родов этого отдела, включающих около 100 видов. Большинство из них - крупные растения; некоторые достигают в высоту более 18м. Их ствол густо покрыт основаниями опавших листьев, а функционирующие листья собраны пучком на верхушке. Этим саговниковые напоминают пальмы (недаром некоторые из них так и называют - саговые пальмы). Однако в отличие от пальм у саговниковых наблюдается настоящий, хотя и медленный, вторичный рост в результате деятельности камбия. Репродуктивные органы саговниковых состоят из более или менее редуцированных листьев с прикрепленными к ним спорангиями, где созревают споры. Спорангии рыхло или кучно скучены в шишкоподобные образования у верхушки растения. Мужские и женские "шишки" находятся на разных особях
Типичный представитель - саговник поникающий (Cycas revolute). Это растение используется садоводами всех стран в качестве декоративного растения. Родом он из Южной Японии, иногда выращивается у нас на Черноморском побережье.
Из других саговниковых можно назвать замию карликовую (Zamia pumila), Encephalartos altensbinii.
Кора и сердцевина саговниковых богата крахмалом и раньше использовалась для получения крахмалистого пищевого продукта - саго. Семена большинства саговниковых также съедобны и до сих пор употребляются в некоторых странах в пищу
ХВОЙНЫЕ
- группа семенных сосудистых растений. Современные представители Хвойных это вечнозеленые, реже листопадные деревья и кустарники.
По одной из классификационных систем эта группа объединяется в класс Хвойные или Пинопсиды (Pinopsida) отдела Голосеменных растений (Gymnospermae). Этот класс разделяется на два подкласса: вымерший подклассКордаиты (Cordaitidae) и современный подкласс Хвойные (Pinidae или Coniferae).
По другой системе классификаций все современные Хвойные объединены в порядок Хвойные (Coniferales), входящий в класс Шишконосные (Coniferopsida) отдела голосеменных растений. Еще одна система относит современные Хвойные к самостоятельному отделу Coniferophyta.
Русское название "хвойные" происходит от слова "хвоя", которым обозначают игловидные листья некоторых их представителей (например, ели, пихты, сосны). Однако далеко не у всех Хвойных листья представляют собой "хвою". Столь же не точно название "шишконосные", представляющее собой перевод второго латинского названия Coniferae (от лат. conus - шишка и ferro - носить), поскольку и шишки есть далеко не у всех представителей этой группы.
Распространение.
Хвойные - растения преимущественно умеренно теплого климата, для существования которых, прежде всего, необходимо достаточное увлажнение. Поэтому в основном они произрастают в умеренном климатическом поясе. Северная граница распространения древесных Хвойных (как и вообще всех древесных видов) совпадает с изотермой июля +10° С. Дальнейшему продвижению леса (в том числе и хвойного леса) на север препятствует недостаток тепла и связанное с этим наличие многолетней мерзлоты с тонким почвенным слоем сезонного оттаивания, который не дает возможности корням деревьев получать из почвы достаточное количество влаги и питательных веществ. Однако по долинам рек и ручьев, где из-за отепляющего воздействия воды слой сезонного оттаивания увеличивается, хвойные леса продвигаются на север несколько дальше. В субтропическом и тропическом климате росту Хвойных мешает недостаточное атмосферное увлажнение, поэтому в этих климатических поясах они растут преимущественно в горах на таких высотах, на которых климат соответствует климату умеренного климатического пояса.
Ныне существующие Хвойные - самая многочисленная и наиболее распространенная группа среди современных голосеменных. Наибольшее число видов и родов Хвойных встречается в северном полушарии. Некоторые из них, например, сосны, ели, лиственницы, пихты формируют здесь обширные хвойные леса, состоящие иногда из одного или немногих видов. В южном полушарии Хвойные наиболее обильны в умеренных областях Новой Зеландии, Австралии и Южной Америки.
Большинство эндемичных родов Хвойных (родов, чье распространение ограничивается очень небольшой территорией) и все древние реликтовые роды (роды, которые на данной территории являются пережитком флоры прошлых эпох, их присутствие может находится в некотором несоответствии с современными условиями существования на этой территории) сосредоточены вдоль бассейна Тихого океана, особенно в Юго-Восточном и Центральном Китае, на Тайване, в Японии, Новой Каледонии, Тасмании, на тихоокеанском побережье Северной Америке, в Южном Чили, Новой Зеландии, Восточной Австралии и Новой Гвинее. Это объясняется тем, что климатически условия в области Тихого океана подверглись наименьшим изменениям после мезозоя, - эры, в которой эта группа достигли своего максимального развития. Не только число родов, но и число видов Хвойных возрастает по направлению к Тихому океану.
По своему значению как в природе, так и в жизни человека Хвойные занимают второе место после цветковых растений, далеко превосходя все остальные группы высших растений.
История развития.
Хвойные появились в каменноугольном периоде (около 370 млн. лет назад), причем в то время они росли, главным образом, в северном полушарии. Они пережили своих родичей кордаитовых и уже с триасового периода (ок.235-185 млн. лет назад) стали играть значительную роль в растительном покрове северного полушария. В юрском и особенно меловом периодах мезозойской эры (ок.185-66 млн. лет назад) Хвойные достигли наибольшего разнообразия и максимального распространения. В это время появилось большинство современных родов, причем в северном полушарии были сосредоточены семейства Сосновых (Pinaceae), Тиссовых (Taxaceae), Таксодиевых (Taxodiaceae) и Кипарисовых (Cupressaceae), а в южном - семейства Араукариевых (Araucariaceae) и Подокарповых (Podocarpaceae).
В третичный период кайнозойской эры (около 66-2,5 млн. лет назад) Хвойные были широко распространены по всей Земле, они не были столь заметно сконцентрированы вокруг Тихого Океана, как сейчас время. Они росли тогда даже в полярных областях, на территориях, лежащих сейчас за пределами границы распространения древесной растительности (Шпицберген, Западная Гренландия, Антарктида). Интересно, что в Антарктиде Хвойные произрастали еще в четвертичное время (2,5 млн. лет назад и позже).
Хвойные древнее всех ныне существующих групп семенных растений, в том числе даже Гинкговых, вполне достоверные остатки которых известны начиная с пермского периода (этот период начался 280 млн. лет назад и продолжался 45 млн. лет). Но в то время как от класса Гинкговых сохранился лишь один вид - гинкго (в сущности, подлинное "живое ископаемое"), Хвойные представляют собой процветающую группу. Среди современных Хвойных самыми древними семействами являются семейства Араукариевые, Подокарповые и Сосновые. Более или менее достоверные остатки представителей этих трех семейств известны уже из отложений конца пермского периода (около 250 млн. лет назад), причем Сосновые, по-видимому, появляются несколько раньше двух других. Древний возраст не только у семейств, но и некоторых родов. Так, растительные остатки, более или менее уверенно относимые к роду Сосна (Pinus), известны из отложений юрского периода (их возраст от 185 до 132 млн. лет), а пыльцевые зерна рода Кедр (Cedrus) - из отложений конца пермского периода (их возраст около 250 млн. лет). Это говорит о том, что некоторые современные роды Хвойных существовали еще до мелового периода, т.е. до появления цветковых растений.
Размеры.
Современные представители Хвойных - вечнозеленые, реже листопадные деревья и кустарники. Размеры Хвойных самые разные - встречаются все переходы от карликовых форм до настоящих гигантов.
По гигантским размерам первое место принадлежит секвойе вечнозеленой (Sequoia sempervirens), относимой к семейству Таксодиевых. Это самое высокое дерево в мире. Его средняя высота более 90 м, а рекордная - 117 м; диаметр ствола может достигать 11 м. По толщине ствола секвойя уступает только мексиканскому болотному кипарису (Taxodium mucronatum), толщина ствола которого может достигать 16 м, и мамонтову дереву (Sequoiadendron giganteum) самые могучие экземпляры которого имеют толщину до 12 м.
Среди карликовых Хвойных первенство принадлежит новозеландскому дакридиуму рыхлолистному (Dacrydium laxifolium) из семейства Подокарповых, высота которого менее 1 метра. Это крошечное хвойное с тонкими стелющимися стеблями образует заросли, главным образом, на горных и субальпийским торфяниках в районах с влажным климатом.
Время жизни.
Некоторые виды Хвойных - одни из самых долгоживущих растений в мире. Таким, например, долгожителем является мамонтово дерево, возраст некоторых его экземпляров - более 3000 лет, однако рекорд долгожительства побивает другой вид, а именно - североамериканская сосна долговечная (Pinus longaeva). В Восточной Неваде был найден экземпляр этого вида, возраст которого определяется приблизительно в 4900 лет, т. е почти пять тысячелетий. Значит, уже во время строительства пирамиды Хеопса это растение уже было довольно старым (ему было более 200 лет).
Строение.
Побеги.
У большинства Хвойных имеются два типа побегов: неограниченные в росте длинные побеги (ауксибласты) и ограниченные в росте укороченные побеги (брахибласты).
Ветвление побегов у Хвойных - моноподиальное. При таком типе ветвления развивающийся из семени главный стебель (моноподий) имеет неограниченный верхушечный рост, за счет чего растение растет в высоту. От моноподия отходят боковые побеги первого, второго и т.д. порядков. Побеги, отходящие от главного ствола, располагаются по спирали, однако они часто так сближаются, что превращаются в мутовки (кольца из побегов вокруг главного ствола), причем ежегодно образуется не больше одного такого кольца ветвей. Посчитав мутовки, можно определить возраст дерева, добавив к полученной цифре 2 года, поскольку первые 2 года жизни у хвойных деревьев мутовки не образуются. Однако этот метод применим для относительно молодых (до 50 лет) насаждений, в которых нижние мутовки на стволах еще не успели полностью затянуться за счет вторичного прироста ствола.
Ветви таких ложных мутовок постепенно укорачиваются по направлению кверху, что придает дереву характерную пирамидальную форму. Вместе с тем боковые ветви второго и следующих порядков располагаются двусторонне симметрично, делаясь иногда совершенно плоскими, что придает дереву ярусный характер. Если верхушечный побег повреждается, то одна из ветвей самой молодой мутовки боковых ветвей может начать расти вверх и принять роль главной. У старых деревьев обычно образуется широкая раскидистая крона, состоящая уже не из одной, а из нескольких главных ветвей, что хорошо заметно, например, у старых сосен.
По мере старения дерева, на открытом месте его нижние ветви могут сохраняться, доходя почти до земли (сбежистая крона), но в густом лесу они обычно достаточно быстро отмирают из-за недостатка света. В результате длинная нижняя часть ствола оголяется и остается практически без сучьев, что очень ценится при заготовке древесины.
У большинства Хвойных, растущих в холодных областях, верхушка побега защищена плотно сидящими тонкими чешуями, образующими в конце вегетационного сезона ясно выраженную почку. Почечные чешуи бывают покрыты защитным слоем смолы или плотно прикрыты толстым волосяным покровом. В других случаях, например, у араукарии и большинства кипарисовых, почечные чешуи не развиваются.
Стебель.
Анатомическое строение стеблей Хвойных относительно однообразно. У них достаточно тонкая кора и массивный древесинный цилиндр, внутри которого - сердцевина, едва различимая в более старых стволах. Как в коре, так и в самой древесине много смоляных ходов (каналов), состоящих из удлиненных межклетных пространств. Смоляные каналы наполнены смолой, которая выделяется выстилающими клетками.
У большинства Хвойных в стволе есть ясно выраженные кольца прироста древесины, образование которых связано с замедлением роста дерева в ежегодные периоды зимнего холода или летней засухи. Каждое кольцо соответствует приросту древесины за один вегетационный сезон. Эти кольца различимы на поперечных срезах ствола, ветвей и корней. Лучше всего они выражены у Хвойных умеренных и холодных широт. По числу годичных колец на распиле ствола, проведенном на высоте шейки корня, можно довольно точно определить возраст дерева. Более того, на основании ряда признаков строения кольца прироста, можно делать определенные выводы о климатических условиях прошлого. Изучение колец прироста Хвойных (так же как и древесных Цветковых) используют для датировки археологических остатков и природных явлений (такой метод называется дендрохронологией), а также для изучения древнего климата (дендроклиматология).
Корень.
Первичный корень у многих Хвойных сохраняется всю жизнь и развивается в виде мощного стержневого корня, от которого отходят боковые корни. Реже, например, у некоторых сосен, первичный корень недоразвит и заменяется боковыми. Кроме длинных корней (главных и боковых), у Хвойных есть и короткие, часто сильно ветвистые корни, которые являются главными абсорбирующими органами растения. Такие корни могут содержать микоризу- симбиоз мицелия гриба и корней растения. Грибы-микоризообразователи разлагают некоторые недоступные растению органические соединения почвы, способствуют усвоению фосфатов, соединений азота и вырабатывают вещества типа витаминов, а сами используют вещества, извлекаемые ими из корней растения.
На корнях Хвойных есть корневые волоски у узкой зоны верхушки корня, легко отпадающие при промывании корня.
Листья
у большинства Хвойных узкие и игольчатые, такие листья называются хвоей, однако у более древних родов (например, у некоторых видов Араукариевых и Подокарповых) листья ланцетные и даже широколанцетные. Так, у подокарпа наибольшего (Podocarpus maximus) наиболее крупные листья достигают 35 см длины и 9 см ширины.
Зеленые листья Хвойных чаще всего сидячие, но иногда - с коротким черешком. Почти всегда они цельные, и лишь у некоторых видов пихты листья на верхушке более или менее выемчатые. Их длина от 1-2 до 30-40 см. Самые длинные листья среди современных Хвойных - у североамериканской сосны болотной
(Pinus palustris), хвоя которой достигает 45 см длины. За исключением нескольких листопадных или веткопадных родов (араукария, агатис, таксодиум, метасеквойя и куннингамия), листья Хвойных вечнозеленые, плотные, болеe или менее жесткие и кожистые. Листорасположение, как правило, спиральное или очередное, реже - мутовчатое или супротивное. Узкие листья (хвоя) имеют одну жилку, широкие - много параллельных жилок. В сечение листья плоские, четырехгранные или округлые.
Кроме зеленых фотосинтезирующих листьев, у некоторых Хвойных имеются коричневые чешуевидные листья.
Размножение.
Органами размножения у Хвойных являются стробилы - видоизмененные укороченные побеги, несущие специальные листья - спорофиллы, на которых формируются спорообразующие органы - спорангии. Есть мужские стробилы (их называют микростробилами) и женские стробилы (мегастробилы). Мегастробилы, как правило, растут компактными собраниями, очень редко они растут по одиночке. Собрания мегастробилов и одиночные мегастробилы называются женскими шишками.
Микростробилы у большинства хвойных растут поодиночке, и, очень редко, в некоторых примитивных формах Хвойных, - компактными собраниями. Собрания микростробилов и одиночные микростробилы называются мужскими шишками.
Обычно Хвойные - однодомные растения (мужские и женские шишки развиваются на одном дереве), реже они двудомные (женские и мужские шишки развиваются на разных деревьях).
Мужские шишки чаще всего собраны группами и обычно располагаются в пазухах листьев, реже на верхушках боковых побегов. У основания шишек нередко есть чешуйки, выполняющие защитную роль. Микроспорофиллы очень редуцированы, чешуевидные или щитковидные. На каждом микроспорофилле образуется от 2 до 15 микроспорангиев. В микроспорангии образуется спорогенная ткань, окруженная со всех сторон тапетумом - слоем клеток, которые доставляют питательный материал для развивающихся микроспор. Число образующихся в микроспорангии микроспор обычно очень велико, они очень легкие, что помогает их распространению ветром.
Из каждой микроспоры развивается мужской гаметофит - половое поколение растений, жизненный цикл которых проходит с чередованием полового и бесполого поколений. Почти у всех высших растений (за исключением мхов) гаметофит слабо развит и недолговечен.
У большинства Хвойных развитие мужского гаметофита начинается еще до рассеивания микроспор, т.е. еще тогда, когда они находятся внутри микроспорангия. У остальных Хвойных (большинство Тиссовых и Кипарисовых) развитие мужского гаметофита начинается лишь после того, как микроспоры будут перенесены ветром на находящиеся в женских шишках семязачатки.
Каждая женская шишка, как правило, состоит из центральной оси, на которой сидят кроющие чешуи, в пазухе каждой из которых находится семенная чешуя, представляющая собой видоизмененный в процессе эволюции мегастробил. В различных линиях эволюции Хвойных происходит процесс постепенного срастания кроющей и семенной чешуи (т.е. чешуевидного мегастробила), что, в конце-концов, приводит к образованию простой, слитной чешуи, часто называемой фертильным комплексом.
На верхней стороне этих чешуевидных мегастробилов находятся семязачатки. Семязачаток представляет собой мегаспорангий (называемый здесь нуцеллусом), окруженный особым защитным веществом - интегументом. В каждом нуцеллусе развивается 3-4 потенциальных мегаспоры из которых только одна способна к развитию. Внутри мегаспорангия в результате многократного деления из функционирующей мегаспоры развивается женский гаметофит, называемый эндоспермом.
С помощью воздушных мешков пыльца из мужских шишек переносится на семязачатки. Происходит опыление и чешуйки женской шишки смыкаются. Мужской гаметофит продолжает свое развитие на мегаспорангии. Через определенный промежуток времени после опыления начинается процесс оплодотворения, который обычно происходит в течение того же сезона. Необычно велик этот промежуток времени у видов сосны, у которой между опылением и оплодотворением проходит от 12 до 14 месяцев. После оплодотворения образуется зигота (клетка, образующаяся в результате слияния мужской и женской половых клеток), из которой сразу же начинает развиваться сначала зачаточный зародыш (предзародыш), а затем и настоящий зародыш. Развитие зародыша происходит за счет запасных веществ эндосперма.
Сформировавшийся зародыш состоит из корешка, стебелька, нескольких семядолей (зародышевых листьев) и почечки. Зародыш окружен эндоспермом, который используется при прорастании. Интегумент образует твердую кожуру семени. Семязачаток плотно прирастает к семенной чешуе, из ткани которой образуется крыловидная пленка, способствующая распространению семян ветром. Таким образом, созревшее семя содержит зародыш спорофита, снабженный запасными веществами и защищенный кожурою. Спорофит - бесполое поколение растений, жизненный цикл которых проходит с чередованием полового и бесполого поколений. Все высшие растения (за исключением моховидных) представлены спорофитами.
Семена Хвойных очень сильно варьируют по величине, форме и окрасу. Кожура семени может быть деревянистой, кожистой или перепончатой. Семена, приспособленные для распространения ветром, снабжены одним большим крыловидным придатком или 2-3 небольшими крыльями. Семена, распространяемые животными, часто имеют сочный и ярко окрашенный покров.
Между опылением и созреванием семян проходит довольно много времени. Например, у сосны обыкновенной (Pinus sylvestris) созревание семян наступает осенью, на второй год после опыления. Шишки к этому времени достигают длины 4-6 см, их чешуи одревесневают и из зеленых становятся серыми. Следующей зимой шишки поникают, чешуйки расходятся и семена высыпаются. Отделившись от материнского растения, семя может длительное время находится в состоянии покоя и лишь с наступлением благоприятных условий трогается в рост.
Характеристика семейств.
В настоящее время насчитывают 7 семейств, около 55 родов и не менее 560 видов Хвойных.
Семейство Араукариевых (Araucariaceae) - очень древняя группа Хвойных, их геологическая история известна, начиная с конца пермского периода, хотя, возможно, они имеют еще более древнее происхождение. Это высокие деревья, растущие в тропической и субтропической зонах южного полушария. Листья обычно крупные, широколанцетной или яйцевидной, а иногда почти округлой формы; реже они мелкие, игольчатые. У некоторых видов зеленые листья растут не только на побегах, но и на стволе. К особенностям Араукариевых относится веткопад - свойство целиком сбрасывать облиственные боковые побеги или даже побеги с листьями.
Растения чаще двудомные. Микростробилы крупные, несут многочисленные спирально расположенные микроспорофиллы. Женская шишка состоит из большого числа чешуй, представляющих собой результат полного или почти полного срастания семенных и кроющих чешуй.
Семейство включает 2 рода: араукария (Araucaria) и агатис (Agatis).
Род араукария насчитывает 15 видов, растущих в Южной Америке (Чили, Патагония, Огненная Земля) и Австралии. Виды араукарии часто образуют обширные леса.
Род агатис насчитывает около 20 видов, распространенных в Новой Зеландии, Северной Австралии и Океании. Некоторые виды являются лесообразующими породами.
Араукариевые имеют важное практическое значение. Большинство видов дает ценную древесину, семена съедобны (являются пищевым продуктом населения Чили). Из видов агатиса добывают смолу. За пределами своего естественного ареала их часто разводят как декоративные породы.
Семейство Подокарповых или Ногоплодниковых (
Podocarpaceae)
История развития семейства начинается с конца пермского периода. В составе семейства 130 видов, распространенных во внетропических областях южного полушария, где они являются главными лесообразующими породами. В тропической зоне встречаются в горах Индии и Восточной Азии.
Это мощные прямостоячие деревья или стелющиеся кустарники. Листья чаще небольшие, широколанцетные, ланцетные, игловидные или чешуевидные, иногда большие и широкие, с несколькими жилками.
Растения обычно двудомные. Микростробилы мелкие, состоят из расположенных в спиральном порядке многочисленных маленьких микроспорофиллов, несущих по 2 микроспорангия. Типичной женской шишки обычно нет. Мегастробил очень редуцирован, состоит из одного семязачатка, окруженного сильно измененной семенной чешуей, которая или полностью обволакивает семязачаток, или сведена к маленькому влагалищному придатку у основания, иногда семенная чешуя и вовсе отсутствует.
Древесина Подокарповых очень плотная и широко используется для различных поделок.
Семейство Тиссовых (Taxaceae). За исключением двух видов Тиссовые распространенны в северном полушарии, где у семейства обширный, но разорванный ареал. Они встречаются и в Северной Америке, и в Европе, и в Азии. Геологическая история семейства начинается с юрского периода.
Тиссовые - вечнозеленые деревья или кустарники. В древесине более или менее отчетливо выражены годичные слои прироста. Листья ланцетовидные или линейные, иногда на коротких (1-2 мм) черешках.
Тиссовые двудомны, редко однодомны. Микростробилы у большинства тисовых одиночные, но встречаются также их сережковидные, колосовидные или шаровидные собрания, расположенные в пазухах листьев. Микроспорофиллы имеют различное строение, чаще всего они щитковидные, каждый из них несет от 2 до 9 микроспорангиев. Мегастробилы объединены в собрания, имеющие вид мелкой шишки с супротивно расположенными парами мегастробилов. Однако обычно эти собрания сильно редуцированы и сведены к одиночным мегастробилам. Они состоят из одного семязачатка, окруженного у основания воротничковидной кровелькой (ее называют ариллусом) красного, желтого или белого цвета. Морфологически ариллус соответствует семенной чешуе. Кровелька полностью покрывает зрелые семена, со временем она становится сочной и ярко окрашенной, что является приспособлением для распространения семян птицами.
Среди тиссовых встречаются настоящие долгожители. Так, продолжительность жизни тисса ягодного, или обыкновенного (Taxus baccata) - до 1500 лет, а иногда, по-видимому, и до 3-4 тысяч лет.
В семействе 5 родов. Это тисс (Taxus), торрея (Torrea), лжетисс, или псевдотаксус (Pseudotaxus), австротаксус (Austrotaxus) и аментотаксус (Amentotaxus).
Семейство Головчатотиссовых (Cephalotaxaceae). Семейство представлено одним родом - головчатотиссом (Cephalotaxus), состоящим всего из 6 видов. В давние геологические времена головчатотиссовые были широко распространены в северном полушарии, но сейчас встречаются только в Азии, где они растут, главным образом, в смешанных горных лесах на высоте 300-3300 м над уровнем моря.
Это вечнозеленые, двудомные, редко однодомные деревья небольшой высоты (10-15 м) или кустарники. Листья кожистые, узколинейные.
Особенностью семейства является наличие шаровидных собраний микростробилов. Эти шаровидные мужские шишки расположены на коротких ножках на побегах последнего года. Мегастробилы собраны в мелкие шишки, в каждой из которых развивается от одного до трех костянковидных семян.
В Японии из семян одного из видов семейства получают воскоподобное вещество, пригодное для технических целей (например, для свечного производства), а масло семян используют для производства красок и лаков. Иногда головчатотиссовые используются в декоративных целях.
Семейство Таксодиевых (Taxodiaceae). Современные таксодиевые являются настоящими "живыми ископаемыми", остатками некогда процветавшего семейства, возникшего более 140 млн. лет назад (самые древние остатки таксодиевых датируются концом юрского периода). Наибольшего расцвета они достигли в третичном периоде, когда многочисленные его представители были широко распространены по всему северному полушарию. Таксодиевые были важными компонентами лесов, простиравшихся на громадных территориях Северной Америки и Евразии и доходящих до Шпицбергена и Гренландии, теперь от них остались лишь небольшие островки в Северной Америке и Восточной Азии.
Сейчас это семейство представлено 10 родами и 14 видами. Благодаря декоративному виду и красивой прочной древесине большинство родов этого семейства культивируется во многих странах земного шара.
Современные таксодиевые - в основном крупные, часто исполинские деревья с одинаковыми или различающимися по длине побегами. Листья линейно-ланцетные, игольчатые или чешуевидные, на побеге расположены спирально. Некоторые виды листопадные, а у некоторых родов, имеющих побеги двух типов (удлиненные и укороченные), наблюдается явление веткопада - осенью укороченные побеги опадают вместе с листьями.
Микростробилы одиночные, лишь у одного вида они собраны в примитивное головчатое собрание. Женские шишки мелкие, одиночные, верхушечные, с плоскими или щитовидными чешуями.
К современным таксодиевым относится ряд интереснейших растений. Первыми из них стоит назвать - секвойядендрон или мамонтово дерево (Sequoiadendron giganteum) и секвойю вечнозеленую (Sequoia sempervirens) - одни из самых крупных и долгоживущих растений мира.
Семейство Кипарисовых (Cupressaceae). В семейство входит 19 родов и около 130 видов, широко распространенных как в южном, так и в северном полушарии. Из 19 родов только три включают в себя много видов (от 15 до 55) -это кипарис, каллитрис и можжевельник.
Кипарисовые - вечнозеленые деревья и кустарники. Деревья чаще всего средних размеров и низкорослые, хотя некоторые могут быть очень высокими, до 70 м высотой, со стволом, достигающим иногда 6 м в диаметре. Среди кустарниковых кипарисовых встречаются и стелющиеся формы. Листья мелкие, чешуевидные или игловидные. Микростробилы обычно одиночные. Женские шишки состоят из нескольких деревянистых, кожистых или сочных чешуй.
Семейство Сосновых (Pinaceae). Трудно назвать такое семейство даже среди цветковых растений, которое по освоенной им территории и по накапливаемой биомассе могло бы соперничать с сосновыми. За исключением одного вида все семейство распространено в северном полушарии, в основном, в умеренной и субтропических зонах (в последней преимущественно в горах). Некоторые виды сосны, ели, пихты и лиственницы забираются высоко в горы и заходят за полярный круг.
Это обширное семейство насчитывает 10 или 11 родов и около 250 видов. Выделяют 4 наиболее крупных рода - пихта (Abies), лиственница (Larix), ель (Picea) и сосна (Pinus), каждый из которых насчитывает по нескольку десятков, а то и сотню (сосна) видов. Другие роды (кедр, тсуга, псевдотсуга, кетелеерия, катайя, лжелиственница) содержат по одному или нескольку видов.
Сосновые - это вечнозеленые или, реже, листопадные деревья, иногда стелющиеся кустарники. Игловидные, чешуевидные, реже узколанцетные листья могут быть разных размеров от крошечных до сильно вытянутых, достигающих в длину 45 см (сосна болотная - P. palustris). Листья держаться на дереве от 2 до 7 лет и лишь у лиственницы и лжелиственницы ежегодно опадают на зиму.
У некоторых родов (например, у сосны, лиственницы, кедра) есть два типа побегов - длинные (неограниченные в росте) и укороченные. Длинные побеги покрыты многочисленными, быстро засыхающими чешуевидными листьями. У лиственницы, кедра и лжелиственницы они несут и зеленые листья. Короткие побеги возникают в пазухах чешуевидных листьев на удлиненных побегах, нередко они опадают вместе с растущими на них листьями.
У сосновых, растущих в суровых условиях Севера, почки защищены плотно прилегающими друг к другу тонкими чешуйками, покрытыми защитным слоем смолы.
Микростробилы одиночные, реже собраны в группы, состоят из многочисленных микроспорофиллов. Мегастробилы собраны в сложные образования - шишки, форма и величина которых для каждого рода разная. Размеры шишек - от 2,5-3 см в длину у лиственницы Лайелла (Larix lyallii) до 50 см в длину у американской сосны Ламберта (Pinus lambertiana).
У большинства сосновых - мощная корневая система, на которой часто встречается микориза (симбиоз мицелия гриба и корней растения). Грибы, вступающие в симбиоз с лесными деревьями, чаще всего относятся к группе шляпочных грибов. Грибы, которые собирают в лесу, представляют собой ни что иное, как плодовые тела грибов, связанных с корнями различных деревьев.
Древесные формы сосновых относятся к крупным деревьям, достигающим 40-50 м в высоту и 0,5-1,2 м в диаметре. Однако в неблагоприятных для себя условиях существования (далеко на Севере, высоко в горах, на болотах) они могут превращаться в низкорослых карликов, зато именно в таких местах можно встретить старейших долгожителей Земли. Так, самое долгоживущее растение на Земле (а именно, уже упоминавшийся экземпляр сосны долговечной, возраст которого был около 4900 лет) рос на горном пике и находился в весьма угнетенном состоянии.
Древесина сосновых разнообразна по фактуре и физическим свойствам. Ее издавна использовали в Евразии, Северной Америке и отчасти в Африке, где она всегда была основным материалом, из которого возводились жилища, хозяйственные постройки, культовые и общественные здания.
Сейчас значение этой древесины еще более возросло. Как и прежде ее используют в столярном и строительном деле, однако наибольшее значение она приобрела в целлюлозно-бумажной промышленности. Для использования этой древесины во многих странах мира искусственно разводятся различные виды этого семейства.
Литература
1. Хржановский В.Г. Основы ботаники с практикумом. Высшая школа. М., 1969.
2. Жизнь растений в шести томах. Том 4: Мхи, плауны, хвощи, папоротники, голосеменные растения. Просвещение. М., 1978
3. Общая ботаника с основами геоботаники. - Учебник для студентов вузов.
4. В. В. Петров, Л.И. Абрамова, С.А. Баландин, Н.А. Березина. М., Высш. шк., 1994.
5. Пчелин В.И. Определитель хвойных древесных растений. Учеб. пособие 2-е изд., перераб. и доп. Йошкар-ола, 1995.
6. Александрова М.С. Хвойные растения в вашем саду. Фитон+, 2000
7. Н.А. Лемеза, Л.В. Камлюк, Н.Д. Лисов "Пособие по биологии для поступающих в ВУЗы".
|
