ФЕДЕРАЛЬНОЕ АГЕНТСТВО ПО РЫБОЛОВСТВУ
Федеральное государственное образовательное учреждение высшего профессионального образования
«Мурманский государственный технический университет»
Вечерне- заочный факультет
КУРСОВАЯ РАБОТА
На тему:
«Особенности биологии и экологии Аphroditiformia Баренцева моря»
Выполнила: студентка
3курса БФ группы 351 Н.В. Дзюбан,
Научный руководитель:
старший преподаватель
кафедры «Биология», Д.К.Мамедов
Мурманск
2008
СОДЕРЖАНИЕ
Введение
1. Первые трактовки систематического положения Aphroditiformiа
2. Краткий морфо-фукциональный очерк Aphroditiformia
2.1 Форма тела и размеры
2.2 Сегментация и деление тела на отделы
2.2.1 Головной отдел
2.2.2 Глотка и ее вооружения
2.2.3 Параподиальный комплекс
2.2.4 Основные типы щетинок
2.2.5 Элитры и их защитная роль
3. Личиночные стадии
4. Экология Aphroditiformia
5. Главные направления в эволюции семейства полиноид
Выводы
Список литературы
УСЛОВНЫЕ ОБОЗНАЧЕНИЯ
а - ацикула к - кишечник
а.у - анальный усик кар - карункул
а.ч. – антеннальная чешуйка к.м - кольцевые мышцы
б.н. - брюшной нерв кт - ктенидии
б.с. - брюшной сосуд л.г - лобный гребень
б.у - буккальный усик л.р - лобные рога
в.у - вентральный усик нв - невроподия
гз - глаза нт - нотоподия
гл - глотка нт.щ - нотоподиальные щетинки
гн - ганглий н.п - нефридиальная папилла
д.б - дорсальный бугорок о - омматофоры
д.м - диагональные мышцы п - пальпа
д.у - дорсальный усик п.ж - паутинная железа
ж - жабра п.л - подиальная лопасть
з.с - затылочная (нухальная) складка II сегмента
п.м - продольные мышцы ч - челюсти
пп - папиллы цер - цератофор
пр - простомиум цир - циррофор
пф - пальпофор црс - цирростиль
р - ротовое отверстие щ - щупальце
с.о - слепой отросток кишечника щ.л - щупальцелатеральное
с.с - спинной сосуд щ.м - щупальце медиальное
сф - сфинктер э - элитра
т.у - тентакулярный усик эм - эмфибрион
ф - фасциальный бугорок эф - элитрофор
I,II,III…. – порядковый номер сегмента
ВВЕДЕНИЕ
До недавних пор про подкласс Чешуйчатые Многощетинковые черви (Aphroditiformia),или Афродита, она же «морская мышь» была известна лишь систематика, анатомические и географические особенности, связанные с местом обитания; в настоящее же время к «морской красавице» возник неподдельный интерес, связанный с удивительными особенностями этого червячка. Наиболее значимой она оказалась в области не только рыбного промысла, в качестве деликатесов пищи(пескожил Arenicola marina), но и в научной деятельности, а точнее в фотонной инженерии. Афродита обладает более сложной структурной организацией, по сравнению с предшествующими типами,
Реклама

Цель: Изучить биологию и экологию чешуйчатых многощетинковых червей рода полинойд.
Задачи:
1)Рассмотреть систематическое положение Чешуйчатых червей;
2) Рассмотреть морфо-функциональные особенности червей данного рода;
3) Описать эволюцию Aphroditiformia.
1. ПЕРВЫЕ ТРАКТОВКИ СИСТЕМАТИЧЕСКОГО ПОЛОЖЕНИЯ APHRODITIFORMIA
Чешуйчатые многощетинковые черви с давних пор привлекают внимание исследователей. Уже Линней в десятом издании своей Systema naturae (1758) выделил в качестве самостоятельного рода Aphrodita aculeata,
который считается типовым для всей группы чешуйчатых червей.
Афродита, или так называемая морская мышь, имеет продолговатую форму тела, нередко превышающую в длину 10 см, и со спины густо прикрыта войлоком из длинных капиллярных щетинок. Этот червь обычно почти полностью закапывается передним концом тела в мягкий ил, так что на его поверхности виден лишь небольшой пучок удлиненных спинных усиков нескольких задних сегментов. Он невольно поражает своей изумительной и неподдельной красотой. Особое впечатление производят его длинные волосовидные щетинки, образующие густую бахрому по бокам тела. Анатомию Aphrodita aculeata
впервые изучил академик П. Паллас, заведывавший зоологическими коллекциями Кунсткамеры Петербургской Академии наук. В своих Miscellanea Zoologica (1766) он обоснованно предложил объединить Aphrodita
с другими многощетинковыми червями — Nereis
и Serpula
— в особую самостоятельную группу, отделив ее от Mollusca, с которыми до этого они рассматривались совместно.
Начиная с самых первых классификаций аннелид многощетинковые черви, несущие на спинной стороне тела плоские чешуйки, или элитры, всегда выделялись как отдельное семейство или ряд семейств, а в некоторых случаях и как самостоятельный отряд. Наиболее обстоятельная первая характеристика чешуйчатых червей дана в работе французских натуралистов Одуэна и Мильн-Эдвардса, хотя их семейство Aphroditiens
насчитывало в тот период еще крайне небольшое количество известных видов и родов. В дальнейшем сведения по группе чешуйчатых начали быстро расширяться, и они некоторыми учеными принимались за самостоятельный отряд Aphroditea
или Aphroditacea
, Весьма высокий таксономический ранг чешуйчатым червям придал и Катрфаж, который семейства Aphroditiens
и Palyriens
выделил в особый подотряд Erraticae aberrantes,
противопоставив его всем остальным эррантным полихетам (подотряд Erraticae propriae).
Это было сделано им на том основании, что у чешуйчатых червей наблюдается неоднородность сегментов (anneaux dissimilaris), выражающаяся в чередовании усиковых и чешуйчатых сегментов, в то время как у всех остальных эррантных полихет все сегменты более или менее однотипные (anneaux similaires ou subsimilaires).
Реклама

Предложены и некоторые другие трактовки систематического положения чешуйчатых червей. Так, наиболее крупный немецкий полихетолог Элерс считал чешуйчатых червей лишь отдельным семейством в составе весьма обширного отряда Nereidea,
объединяющего всех современных эррантных полихет. При этом указанный автор для семейства использует то же название Aphroditea,
которое ранее Кинбергом было предложено для обозначения отряда. Французский исследователь Клапаред также рассматривал чешуйчатых червей лишь как семейство — Aphroditiens,
но в его составе выделял 6 триб — Aphrodites, Polynoides, Pholoides, Acoetides, Sigalionides
и Polylepides.
Шведский исследователь Мальмгрен различал 4 самостоятельных семейства чешуйчатых червей — Aphroditidae, Polynoidae, Acoetidae (=Polyodontidae)
и Sigalionidae,
которые в дальнейшем некоторыми авторами понимались как подсемейства. Как видно из всего сказанного, единое мнение о классификации чешуйчатых червей отсутствует. Подробная синонимия всех предложенных в прошлом столетии названий различных таксономических категорий дана в работе Дарбу. В более позднее время американскими учеными все чешуйчатые черви были выделены в надсемейство Aphroditoidea
, однако это же название иногда употребляется и в более широком понимании, а именно для объединения почти всех эррантных полихет, за исключением глицерид и эвяицид.
В данной работе для чешуйчатых червей я использую весьма удачный термин Aphroditiformia,
предложенный Левинсеном. По этому каталогу подотряд Aphroditiformia
состоит из трех надсемейств: Aphroditacea
(с 6 семействами), Chrysopetalacea
(с 2 семействами) и Pisionacea
(с 1 семейством). Общая схема классификации выглядит следующим образом (после названия семейств в скобках первая цифра обозначает количество родов вторая — видов):
Подотряд APHRODITIFORMIA Надсем. Aphroditacea
Сем. Aphroditidae
(7, 63)
Сем. Polynoidae
(93, 539)
Сем. Sigalionidae
(17, 156)
Сем. Pholoididae
(1, 4)
Сем. Eulepethidae
(4, 12)
Сем. Polyodontidae
(6, 39)
Надсем. Chrysopetalacea
Сем. Chrysopetalidae (4, 24)
Сем. Palmyridae (1,2)
Надсем. Pisionacea Сем. Pisionidae
(4, 14)
Надсемейство Aphroditacea
является центральной группой в филогенетическом отношении, обладает большим количеством родов и видов, отличающихся морфологическим разнообразием. Наибольшее в этом надсемействе значение имеет семейство Polynoidae.
Надсемейство Chrysopetalacea
представляет собой как бы связующее звено с высокоспециализированным семейством Amphinomida
. Надсемейство Pisionacea
занимает промежуточное положение между Aphroditiformia
и Glyceriformia
,причем по личиночному развитию пизиониды ближе всего к чешуйчатым червям.
2. КРАТКИЙ МОРФО-ФУНКЦИОНАЛЬНЫЙ ОЧЕРК APHRODITIFORMIA
2.1 ФОРМА ТЕЛА И РАЗМЕРЫ
Тело чешуйчатых червей, как правило, заметно сплющено дорсовентрально. Спинная сторона, прикрытая спинными чешуйками, или элитрами, несколько выпуклая (сводчатая), а брюшная — более плоская, с медиальным продольным желобком. К переднему и особенно к заднему концам тело немного суживается; на заднем конце тела все сегменты становятся более короткими и узкими. Многие чешуйчатые черви обладают относительно коротким телом, имеют овальную форму; у других же, наоборот, оно сильно вытянутое («червеобразное»). Форма тела связана с образом жизни. Удлиненное тело характерно для червей, глубоко зарывающихся в грунт (многие Sigalionidae),
строящих собственные длинные трубки (Polyodontidae),
а также для червей, ведущих комменсальный образ жизни (некоторые Polynoidae).
У чешуйчатых червей наблюдается вполне определенная тенденция к стабилизации количества сегментов и к укорочению тела, что для них, бесспорно, является эволюционно прогрессивным моментом.
Форма тела чешуйчатых червей, в противоположность многим другим поли-хетам, весьма постоянная, не подверженная сколько-нибудь заметному удлинению и сжиманию при передвижениях, причем она мало изменяется и при фиксации. Некоторые полиноиды с укороченным телом (в основном Lepidonotus)
при фиксации сворачиваются в колечко (уже по этой особенности они хорошо распознаются среди прочих полиноид).
Размеры чешуйчатых червей варьируют. Представители семейства Aphroditidae
s. str. характеризуются довольно крупными размерами. Так, Aphrodita aculeata,
весьма обычная у берегов Европы, нередко имеет в длину около 100 мм, при ширине 40—50 мм (отмечены экземпляры до 180 мм длины), a Laetmonice producta
из южного полушария достигает длины 135 мм и ширины до 40— 65 мм, включая параподии со щетинками. В семействе Polynoidae
также наблюдаются гиганты. В частности, к ним относится калифорнийская полиноида Hololepida magna
длиной 250 мм. Из антарктических районов в коллекциях ЗИН АН СССР имеются экземпляры Eulagisca gigantea,
достигающие в длину 210 мм при ширине 60 мм без щетинок и 100 мм со щетинками. В арктических морях одна из наиболее крупных полиноид — Eunoe nodosa
— до 100 мм длины и 35 мм ширины. Средняя же длина большинства видов полиноид около 30—40 мм. Наиболее тонкие и длинные полиноиды — Lepidasthenia
(до 200 мм длины), характеризующиеся также наибольшим количеством сегментов.
Великанами среди полихет справедливо считаются некоторые Polyodontidae,
живущие в очень крупных длинных илистых трубках, в связи, с чем их иногда называют «илистыми сосисками»; так, имеются указания, что они достигают в длину 2 метров при ширине 2 см. Среди сигалионид немало видов до 300 мм длины при ширине 8—9 мм (Sigalion squamatum
и др.). Но наряду с такими крупными формами среди чешуйчатых червей есть виды, которые характеризуются крайне небольшими размерами., не превышающими 20—25 мм. При этом длина тела не коррелирует с количеством сегментов. Например, Pholoe minuta
с 70 сегментами имеет в длину максимум 20 мм, а у упомянутого выше Laetmonice producta,
достигающего 135 мм, количество сегментов не свыше 50; у Eulagisca gigantea
и Harmothoe Imbricata
количество сегментов не превышает 37—38, в то же время длина особей первого вида иногда превосходит 200 мм, а второго — достигает только 50 мм, обычно около 30 мм.
Наиболее разнообразна окраска элитр. Последние могут иметь пестрый или узорчатый рисунок, непосредственно связанный со скульптурой самих элитр. Исключительно красив бросающийся в глаза рисунок в виде продольных полос на элитрах Hyperhalosydna striata.
Уже по этому рисунку удается безошибочно определить вид. Иногда элитры бесцветные и полупрозрачные, — например, у видов Aphrodita,
элитры которых прикрыты наружным войлоком из волосовидных щетинок, или у Bylgides sarsi,
у которого наличие тонких и нежных элитр связано с его факультативным планктонным образом жизни. У Psammolyce,
зарывающегося в песок, элитры, как правило, прочно обклеены мелкими песчинками. Это, по всей видимости, своего рода маскировка, (предохраняющая червей от врагов). Благодаря относительно темной окраске тела, в особенности элитр, соответствующей общему фону субстрата, на котором поселяются полиноиды, последних не всегда удается быстро обнаружить. К этому следует добавить, что элитры крупных ползающих по поверхности грунта полиноид часто покрыты разнообразными эпифитами и различными комменсалами, что еще больше скрывает их на морском дне.
Щетинки большей частью золотистого или светло-янтарного оттенка, могут быть и совершенно прозрачными. Наряду с этим крупные спинные игловидные щетинки Aphrodita
почти черного цвета.
2.2 СЕГМЕНТАЦИЯ И ДЕЛЕНИЕ ТЕЛА НА ОТДЕЛЫ
Тело многощетинковых червей поделено на сегменты (членики), которые внешне хорошо распознаются по наличию на каждом из них по одной паре параподий, снабженных обычно различными придатками. Наружная кольчатость тела тесно связана с внутренними септами (диссепиментами),:
обособляющими друг от друга отдельные сегменты (метамеры, или сомиты) (рис. 1). В каждом сегменте, как правило, закономерно повторяются наборы внутренних органов; комплекс нервных образований одного сегмента обозначается как невросомит, сплетение кровеносных сосудов — ангиосомит, система мышечных волокон — миосомит и т. д.; большей частью метамерно располагаются также выделительные органы и гонады. У чешуйчатых червей кишечник в каждом сегменте имеет боковые слепые выросты (карманы, или дивертикулы), так что и он у них обладает вполне определенной метамерией (рис. 2).

Рис. 1. Поперечный разрез через туловищный сомит (чешуйчатый сегмент).
Таким образом, наружная кольчатость, или наружная метамерия, червей обычно соответствует внутренней метамерии.1
Полная гомономность (однородность) всех туловищных сегментов у многощетинковых червей никогда не наблюдается. Наибольшая гомономность сегментов характерна для бродячих (эррантных) полихет, но и у них она в той или другой степени нарушена. Следует различать гетерономность первичную и вторичную. Первичная гетерономность возникает еще в процессе метаморфоза личинки и связана с образованием ларвальных и постларвальных сегментов, имеющих разное происхождение. Ларвальные сегменты закладываются обычно одновременно на начальной стадии метаморфоза личинок, а лостларвальные сегменты последовательно и постепенно возникают из предпигидиальной эктодермальной зоны роста, причем целом в ларвальных сегментах всегда формируется из мезодермальных полосок путем их разделения на мезодермальные блоки, которые затем становятся полыми.
1
При описании и на рисунках червей сегменты обозначаются римскими цифрами
Число ларвальных сегментов варьирует: у Polynoidae
их 8—10, а у Siga-lionidae
— 4—6. Большое и изменчивое количество ларвальных сегментов указывает на примитивный тип метаморфоза. Ларвальные сегменты в отличие от постларвальных не образуют половых желез, не имеют целомодуктов и хлора-гогенных клеток. Иногда ларвальные сегменты подвергаются слиянию, образуя перистомиум: например, крупный перистомиум нереид представляет собой продукт слияния трех передних ларвальных сегментов. Вторичная гетерономность относится к постларвальным сегментам, она целиком обусловлена особенностями обитания взрослых червей (адаптивные приспособления).
Количество сегментов у чешуйчатых червей, как и у большинства других полихет, сильно варьирует. Существуют виды, у которых сегментов относительно немного и их число строго постоянное (например, Lepidonotus
— всего 26 сегментов), но некоторые виды (многие Lepidasthenia, Arctonoe
и др.) состоят из 100 и более сегментов, причем количество последних увеличивается с возрастом и возникновение новых сегментов на заднем конце тела (в предпиги-диальной зоне роста) не прекращается с наступлением половой зрелости. Самые задние сегменты обычно меньше (тоньше и короче) остальных и имеют недоразвитые параподий. Вполне естественно, что черви с большим количеством сегментов одновременно и наиболее длинные. Однако, как уже отмечалось ранее, количество сегментов не всегда строго коррелирует с размерами самих червей. Неопределенно большое количество сегментов у полиноидных червей мы рассматриваем как примитивное состояние, а стабилизацию количества сегментов — как прогрессивный момент. Подсчет сегментов легче всего производить с брюшной стороны, так как спинная сторона червей нередко замаскирована различными придатками, в частности у чешуйчатых червей элитрами. Количество сегментов у представителей афродитид (Aphro-
f dita, Laetmonice, Hermonia, Pontogenia)
не превышает 50 (колеблется в пределах 32—47). Среди полипоид по числу сегментов выделяется три группы родов: 1) сегментов 25— 30 — Lepidonotus, Hermenia, Euphione, Iphione;
2) сегментов 30—50 — Harmothoe, Lagisca, Eunoe, Antinoe, Gattyana, Bylgides, Eucranta, Malm-grenia, Alentia
и др. и 3) сегментов свыше 50 (до 100 и более) — Perolepis, Arctonoe, Lepidasthenia
и др. Для большинства сигалионид характерно более 50 сегментов.
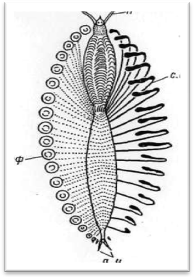
Рис.2. Схема строения пищеварительного канала
Помимо основных наружных перетяжек, отделяющих соседние сегменты друг от друга, в пределах одного сегмента преимущественно на его спинной стороне нередко наблюдается добавочная вторичная, или так называемая ложная кольчатость — псевдосегментация (annuli). У рассматриваемых нами чешуйчатых червей она имеет характер небольших метамерно повторяющихся бороздок или складок, к которым приурочены ресничные пояски и поперечные пигментные полосы, придающие некоторым полиноидам весьма пеструю окраску.
При беглом взгляде на чешуйчатых червей нередко создается впечатление, что они обладают гомономной сегментацией, т. е. на всем протяжении тела у них все сегменты однородны. Такое впечатление объясняется тем, что у многих полиноид вся их спинная сторона как бы равномерно прикрыта спинными чешуйками (элитрами). Однако это ложное представление, так как элитры имеются далеко не на всех сегментах. У чешуйчатых червей наблюдается вполне определенное чередование усиковых сегментов, лишенных элитр, и чешуйчатых сегментов, снабженных элитрами. Таким образом, говорить о полной гомономности сегментов у них не приходится.
Многократное повторение двух или трех смежных сегментов, имеющих разное морфологическое строение, называется метамерией 2-го порядка.
Метамерия 2-го порядка, создающая видимость гомономности, имеет место не только у чешуйчатых червей. Например, у некоторых силлид более длинные спинные усики нередко чередуются через сегмент с более короткими, а иногда (Parasphaerosyllis)
изменяется поочередно и форма самих усиков.
Метамерия 2-го порядка — это лишь один из примеров наиболее простой дифференциации сегментов. Она представляет собой начальную стадию образования гетерономности. Наиболее гомономными червями являются филло-доциды, отчасти нереиды. У большинства полихет, в особенности у седентарных, ведущих тубикольный образ жизни, процесс возникновения гетерономности идет значительно дальше и видоизмененные сегменты не чередуются между собой, а приурочены к различным участкам тела, выполняющим разные функции. В связи с этим у некоторых полихет резко обособляются передний участок тела — торакс (так называемые грудные сегменты) и задний участок — абдомен (брюшные сегменты), а иногда и дополнительный хвостовой участок. Наиболее сложная гетерономия наблюдается у ряда седентарных полихет, в частности у Chaetopteridae
и Sabellariidae,
что обусловлено тубикольным образом жизни. У чешуйчатых червей разделения тела на торакс и абдомен нет, однако XXIII сегмент у них все же играет какую-то роль, отделяя переднюю часть тела со строго закономерным расположением элитр через один сегмент от задней, где расположение элитр еще не стабилизировалось.
У всех чешуйчатых червей, как и у остальных полихет, различают три основных отдела тела. Во-первых, передний, или так называемый головной, отдел, слагающийся из простомиума, или головной лопасти, и двух передних сегментов — перистомиального и буккального. Во-вторых, туловищный отдел (метастомиум), состоящий из большого количества повторяющихся сегментов с развитыми параподиями; этот отдел является наиболее крупной частью червя. В-третьих, анальная лопасть, или пигидий, находящийся на самом заднем конце тела и представляющий собой неметамерный отдел, который образуется у личинок еще до метаморфоза. Впереди пигидия находится зона роста, где возникают новые сегменты. Анальная лопасть обычно снабжена двумя нитевидными анальными усиками, или уритами. У Aphrodita
уриты отсутствуют, но на параподиях последних сегментов имеются удлиненные спинные усики, образующие бахрому вокруг ануса.
2.2.1 ГОЛОВНОЙ ОТДЕЛ
Передний, или головной, отдел тела состоит из простомиума (prostomium), иначе головной лопасти, и расположенного за ним первого (перистомиального) и второго (буккального) сегментов (рис. 3, 4). Первый сегмент сильно редуцирован со спины, и от него сохраняются лишь направленные вперед подиаль-ные выросты с перистомиальными, иначе тентакулярными усиками (cirri peri-stomiales, cirri tentaculares). Второй сегмент отличается от последующих туловищных сегментов в основном наличием очень длинных брюшных усиков, которые обозначаются как ротовые (cirri buccales), так как они расположены по бокам ротового отверстия и непосредственно участвуют в обнаружении и захвате пиши. Второй сегмент у всех чешуйчатых червей снабжен первой парой элитр, или спинных чешуек. На переднем крае II сегмента нередко имеется полукруглая складка, или вырост, заходящий сзади на спинную сторону головной лопасти (так называемая затылочная, или нухальная складка). Преобразования I и II сегментов в единый перистомиум путем их слияния у чешуйчатых червей не наблюдается. В этом отношении процессы цефализации у поли-ноид стоят на более низком уровне, чем у нереид, имеющих хорошо выраженный перистомиум в виде широкого ошейника, который представляет собой результат слияния трех передних ларвальных сегментов.
Простомиум у всех чешуйчатых червей хорошо развит и имеет крупные размеры. В процессе метаморфоза личинки он образуется непосредственно из верхнего полушария трохофоры (эписферы), т. е. имеет независимое от всех остальных сегментов происхождение. В соответствии с этим головная лопасть всегда располагается впереди ротового отверстия, находящегося на брюшной стороне червя. У чешуйчатых червей простомиум обычно вытянутой или округлой формы, иногда ширина его может быть больше длины. У переднего края головной лопасти находится 1—3 головных щупальца, или антенны (antennae): одно медиальное и два латеральных.

Рис. 3. Продольный разрез через передний отдел тела полиноиды.
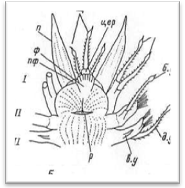

Рис. 4. Головной отдел полиноиды: вид сверху (А)
и вид снизу (Б).
Головные щупальца веретенообразной формы, гладкие или с многочисленными мелкими папиллами. Щупальца большей частью имеют обособленные подставки, которые обозначаются как цератофоры, а сами щупальца — цератостили. Наличие цератофор обеспечивает большую подвижность самих щупалец. При медиальном щупальце иногда наблюдаются своеобразные ложковидные органы с ресничным эпителием — так называемые ктенидии (Laetmonice, Sthenelais
и др.); назначение этих чувствительных органов точно не выяснено. Количество и характер расположения головных щупалец имеет большое систематическое значение: на этом основании в пределах семейства Polynoidae выделяются несколько подсемейств. У полиноид различают следующие типы головной лопасти (рис.5).
1.Лепидонотоидный тип
– три головных щупальца; боковые дольки головной лопасти сильно вытянуты вперед и непосредственно (без базальных подставок) переходят в латеральные щупальца (терминальное положение латеральных щупалец); медиальное щупальце отходит от крупной базальной подставки (цератофора), расположенной в промежутке между боковыми выростами головной лопасти, и находится в одной плоскости с латеральными щупальцами.
2.Арктоноидный тип
– три головных щупальца; латеральные щупальца имеют базальные подставки, отходящие от переднего края головной лопасти или несколько сдвинутые на брюшную сторону (субтерминальное положение);

Рис.5 Основные типы головного отдела полиноид. 1 – лепидонотоидный, 2 – арктоноидный, 3 – хармотоидный, 4 – ифионоиднный, 5 – мацеллицефалоидный, 6 - адметеолоидный.
Подставка медиального щупальца лежит поверх латеральных. Арктоноидный тип является промежуточным между лепидонотоидным и хармотоидным.
3.Хармотоидный тип
— три головных щупальца; латеральные щупальца с обособленными подставками - имеют вентральное положение, медиальное щупальце с крупной базальной подставкой сдвинуто на дорсальную сторону головной лопасти; дольки головной лопасти нередко с заостренными выростами, так называемыми лобными рогами.
4.Ифионоидный тип
— имеются лишь латеральные щупальца, сидящие на боковых выростах головной лопасти; вместо медиального щупальца в задней части головной лопасти под нухальной складкой находится небольшая папилла.
5.Мацеллицефалоидный тип
— имеется лишь одно хорошо выраженное медиальное щупальце, сильно сдвинутое на дорсальную сторону головной лопасти и сидящее на крупной базальной подставке (цератофоре); дольки головной лопасти округлые или с небольшими нитевидными придатками, которые представляют собой рудименты латеральных щупалец.
6.Адметеллоидный тип
— три головных щупальца; латеральные щупальца лепидонотоидного типа, медиальное щупальце сдвинуто на спинную сторону головной лопасти, при нем имеются особые придатки — так называемые антеннальные чешуйки.
Головная лопасть у всех чешуйчатых червей со спины прикрывает первый (перистомиальный) сегмент, который из-за этого не имеет четких границ. При тентакулярных усиках первого сегмента, иногда обозначаемых как щупальцевидные усики, нередко имеются ацикула и пучок щетинок. Головные щупальца и тентакулярные усики внешне могут мало различаться, однако головные щупальца иннервируются от переднего отдела головного мозга, а перистомиальные усики — от коннективов ротового нервного кольца.
Помимо головных щупалец и тентакулярных усиков, у всех чешуйчатых червей имеются так называемые щупики, или пальпы (palpi), по своей массивности превосходящие все остальные головные придатки. Пальпы отходят с брюшной стороны головного отдела червя, по бокам от ротового отверстия, и внутрь их может заходить целомическая полость. Пальпы имеют очень небольшие подставки — пальпофоры, придающие им дополнительную подвижность. Вдоль наружной поверхности пальп обычно наблюдаются многочисленные продольные ряды мягких чувствительных папилл.
Все указанные здесь головные придатки являются осязательными органами и играют определенную роль при обнаружении пищи, но основным орудием захвата пищи служит выпячивающаяся наружу мускулистая глотка (pharynx), вооруженная нередко мощными хитиноидными челюстями.
На дорсальной стороне головной лопасти расположены глаза; они в большинстве случаев простого типа, без глазных стебельков (омматофоров). Глазные пятна темного (почти черного) цвета, округлой или овальной формы. У полиноид их две пары, причем первая (передняя) несколько крупнее задней, которая к тому же может быть частично прикрыта спинной складкой второго (буккаль-ного) сегмента. Первая пара глаз располагается по бокам головной лопасти в наиболее расширенной ее части или у переднего края, они могут частично переходить и на вентральную сторону головной лопасти (например, у Иаг-mothoe imbricata).

Рис.6. Головной отдел у представителей различных семейств Aphroditiformia.
1
— Aphrodita (Aphroditidae), 2
— Harmothoe (Polynoidae), 3
— Stherwlepis (Sigalionidae), 4
— Eulepeihus (Eulepethidae),.
7 — Chrysopetalum (Chrysopetalidae), 6
— Pisione (Pisianidae),
7 — Polyodontes (PolyadontidaeJ.
Глазные пятна эпителиального характера и состоят из одного слоя светочувствительных клеток, имеющих сильно вытянутую призматическую форму с темным пигментом в средней своей части. От каждой призматической клетки отходит небольшое нервное волоконце, которое, переплетаясь с соседними, образует оптический нерв, связанный с головным мозгом. Сверху светочувствительные клетки (ретина) прикрыты прозрачной кутикулой (иногда утолщенной), причем у ряда форм под кутикулой наблюдается вдавление или «чаша» заполненная «стекловидным телом» (секретом клеток ретины). Стекловидное тело представляет собой светопреломляющую линзу, или хрусталик (диоптрический аппарат). Иногда под кутикулой дополнительно прослеживается слой эпидермиса и «чаша» преобразуется «в пузырек», который еще более глубоко опускается внутрь мышечных тканей головной лопасти. Наличие светопреломляющих линз хорошо распознается при наружном осмотре, они выглядят в виде светлого кружка в центре темного глаза. Такие линзы наблюдаются преимущественно у пелагических форм, которым требуется обозревать пространство в разных направлениях (например, Lagisca hubrechti).
У червей, обитающих на больших глубинах, наоборот, отмечается редукция глаз (Наг-mothoe derjugini, Weberia, Admetella,
различные представители мацеллицефалин). У представителей Polyodontidae,
живущих в длинных трубках, глаза, как правило, сильно гипертрофированы и находятся на передней овальной стороне вытянутых омматофоров — «глазных ножек». Последние похожи на «рачьи глаза» и служат для быстрой ориентировки, когда червь высовывается из трубки. Наиболее характерные примеры строения головного отдела для различных семейств Aphroditiformia
приведены на (рис. 6).
2.2.2 ГЛОТКА И ЕЕ ВООРУЖЕНИЕ
Глотка (pharynx) - мускулистая часть передней кишки, способная выпячиваться наружу через ротовое отверстие. У полихет различают два основных типа глотки: 1) мешковидная глотка (ventral proboscis) в виде относительно короткого слепого мешка (buccal bulb, bulbe pharyngien), расположенного под пищеводом, и 2) трубчатая глотка (axial proboscis, buccal tube), представляющая собой подвижную трубку, постепенно переходящую в среднюю кишку. Первый, весьма специализированный тип глотки среди эррантных полихет характерен лишь для эвницид и амфяномид; наиболее сложно устроена мешковидная глотка эвницид, вооруженная нижними и верхними челюстями (мандибулой и максиллами). У чешуйчатых червей глотка трубчатая. Механизм выпячивания сильно мускулистой глотки чешуйчатых червей такой же, как у других полихет с этим типом глотки, т. е. глотка выпячивается наружу наподобие пальца перчатки. Выпячивание происходит вследствие давления полостной жидкости и регулируется специализированными мышцами. Выпяченная глотка у некоторых видов достигает трети длины самого червя.
У наружного отверстия выпяченной глотки полиноид обычно имеются отдельные крупные округлые, мягкие краевые (оральные) папиллы или многочисленные удлиненные сосочки, образующие густую наружную бахрому.

Рис. 7. Глотка и челюстной аппарат.
1
—передний край выпяченной глотки Laetmonice:
вид сбоку (а), вид спереди (б) и отдельная краевая па-шглла (в); 2
— глотка Leyidonotus;
вид сбоку (а) и вид спереди (б); 3
— отпрепарированные челюстные крючки Lepidasthenm:
дорсальный правый (а)
и вентральный левый (б); 4
— передняя часть глотки Macellicephaloides; 5
— челюстной аппарат Рisione; 6
— челюстное вооружение Polyodontes,
вид глотки спереди (3, б)
.
Краевые папиллы богаты железами, выделяющими ферменты, которые участвуют в пищеварении. Наружная стенка выпяченной глотки обычно гладкая, без папилл. Примеры строения глотки и ее вооружения у чешуйчатых червей приведены на (рис.7).
Для большинства полиноид характерно наличие как на дорсальной, так и на вентральной стороне глотки по 9 крупных краевых папилл. Иногда их может быть несколько больше, а именно 11+11 (Lepidasthenia)
и даже до 14—22 пар (Hololepida).
У рода Hermilepidonotus
описан крайне необычный для полиноид характер краевых папилл: их до 30 пар, причем они не простой треугольной формы, а имеют с внутренней стороны до 20 добавочных пальцевидных отростков, образующих у входного отверстия как бы щетку, напоминающую таковую Aphroditidae
s. str. (Laetmonice)
(рис. 7, 1).
Несколько необычен характер папилл у мацеллицефалин: так, представители рода Macellicephaloides
(рис. 7, 4)
имеют на глотке две очень длинные боковые усиковидные папиллы, которые в морфологическом и функциональном отношениях весьма сходны с боковыми усиками на глотке пелагических альциопид.
Челюстной аппарат, представляющий собой локальные утолщения гиподермы, у чешуйчатых червей развит различно. Наиболее мощные челюсти в виде четырех крупных крючков (или когтей) наблюдаются у большинства полиноид, сигалионид и полиодонтид. При выпяченной глотке эти крючки частично выставлены наружу, располагаясь попарно на вентральной и дорсальной сторонах глотки. Каждый челюстной крючок имеет несколько расширенное основание и слегка оттопыренную утонченную боковую пластинку, или крылышко, так называемый элерон (aileron). Среди аберрантной группы преимущественно глубоководных полиноид — Macellicephalinae
— наблюдается постепенная редукция челюстного аппарата: Macellicephala,
так же как и все полиноиды, имеют 2 дорсальных и 2 вентральных крючка; Macellicephaloides
— 1 дорсальный и 2 вентральных крючка; у Macelloides
имеются лишь очень тонкие, слегка зазубренные утолщения по наружному краю глотки, a Macella
вообще лишена на глотке каких-либо следов хитиноидных утолщений. Незначительно развит челюстной аппарат у представителей Eulepethidae
. Для Aphroditidae
s. str. характерно отсутствие челюстных образований. Глотка Chrysopetalidae
снабжена лишь двумя небольшими стилетами.
Челюсти чешуйчатых червей, как и других полихет, состоят в основном из; кератина, или рогового вещества; они обычно темного, почти черного цвета. На некоторых челюстных пластинках хорошо видны так называемые годовые кольца, по которым можно судить о возрасте червей. Детали строения челюстей имеют значение для таксономии — диагностики родов и семейств, а также для филогенетических построений. Челюсти хорошо сохраняются в ископаемом состоянии. В онтогенезе челюсти у полиноид закладываются уже на второй стадии нектохеты.
Челюстной аппарат полихет выполняет различные функции. В довольно редких случаях он используется как орудие защиты и нападения (последнее было отмечено среди нереид). Глотка может играть существенную роль при ло-комоции и зарывании в грунт, в частности при постройке ходов — ranepefr в толще грунта; в этих случаях она действует как вспомогательное «якорное»-устройство (например, у глицерид и нефтиид).
Наличие или отсутствие челюстного аппарата у полихет еще ничего не говорит о том, являются ли данные черви плотоядными или детритоядными животными. Известно, что многие полихеты, лишенные челюстного аппарата, ведут хищнический образ жизни, например филлодоциды, и, наоборот, некоторые многощетинковые черви с развитыми челюстями используют последние в основном как грабли для сбора детрита, в частности некоторые нереиды. В желудках большинства полиноид, снабженных мощными челюстями, обычно обнаруживают не только мелких животных, но и различного рода растительные обрывки и разнообразный детрит. По всей видимости, полиноиды, как и большинство других полихет, всеядные животные. Челюстной аппарат полихет, как правило, не служит жевательным органом, а лишь служит для схватывания и удерживания добытой пищи, которая затем путем втягивания глотки поступает в кишечник. У основания челюстей глицерид и некоторых нереид имеются особые ядовитые железы, их яд умертвляет жертву. У полиноид такие железы не описаны, но наличие их вполне возможно.
2.2.3 ПАРАПОДИАЛЬНЫЙ КОМПЛЕКС
Все сегменты метастомиума, иначе туловищные сегменты, снабжены боковыми выростами — параподиями, вооруженными многочисленными щетинками. Пара-подии в основном служат локомоторными органами, но одновременно в не-которых случаях выполняют и дыхательные функции, а также являются орудиями защиты. Более подробно остановимся на морфологии параподий Aphroditiformia,
у которых, как и у большинства эррантных полихет, параподий моностикного характера, т. е. невроподии и нотоподии объединены общим подиальным стволом (в противоположность многим седентарным полихетам, у которых имеются дистикные параподий, характеризующиеся разобщенными ветвями).


Рис. 8. Усиковая (А)
и чешуйчатая (Б)
параподии полиноид.
Параподий чешуйчатых червей имеют большей частью сильно вытянутую форму и четко отделены от главного ствола тела самого червя (на брюшной стороне они ограничиваются нефридиальными папиллами). Длина параподий нередко равна ширине тела и даже может ее превосходить. Различают два основных типа параподий: 1) усиковые параподии, снабженные удлиненным верете-образной формы спинным усиком, выполняющим осязательную функцию, и 2) чешуйчатые параподии, у которых спинной усик заменен плоской чешуйкой, или элитрой, выполняющей преимущественно защитную функцию (рис. 8). Параподии снабжены двумя крупными внутренними осевыми щетинками, как правило, темного цвета. Это так называемые ацикулы (aciculum), которые соответствуют двум ветвям параподий: верхней, или спинной (нотоподия— notopodium), и нижней, или брюшной (невроподия — neuropidium). Такие параподии обозначаются как двуветвистые (biramus, biremis), однако верхняя ветвь нередко остается недоразвитой и выражена лишь небольшим бугорком на передне-верхней стороне подиального ствола. Параподии с недоразвитой нотоподией иногда называются «ложнодвуветвистые» или «полуразветвленные» (sub-biramus, sesquiramus). При беглом взгляде последние могут быть приняты за одноветвистые; они наблюдаются преимущественно лишь у многосегментных полиноид типа Arctonoe, Lepidasihenia
и Perolepis.
Короткие полиноиды — Harmotkoe
и большинство других родов имеют, напротив, мощную нотоподию, снабженную большим количеством крупных щетинок, образующих плотный пучок. В качестве «типичных» для чешуйчатых червей принято считать параподии Harmotkoe.
Однако за исходный тип параподий для всех полихет мы принимаем одноветвистые. В таксономической литературе при характеристике многих родов полиноид пишется, что параподии у них с редуцированной нотоподией. Нам представляется, что в большинстве случаев мы имеем дело с недоразвитой нотоподией. Последнее, в частности, подтверждается данными по развитию: в онтогенезе нотоподия закладывается позже невроподии.
Невроподия представляет собой непосредственное продолжение главного подиального ствола (truncus), служит как бы базой для нотоподии и несет основную локомоторную функцию. Нижняя (невроподиальная) ацикула доходит до дистального конца подиального выроста, так называемого ацикулярного бугорка (acicular lobe), придавая всей конструкции параподии упругий харак тер. Невроподии на дистальном конце обычно имеют две мягкие, разного размера и формы кожистые лопасти (lamella): переднюю, или предщетинковую (presetal), и заднюю, или послещетинковую (postsetal). Щетинки на невроподии расположены полукругом, причем когда невроподия сильно вытянута и прижата к субстрату, то и большинство ее щетинок также упирается в последний (это обеспечивается разной длиной самих щетинок и степенью их выдвижения). Выдвижение и втягивание щетинок происходит под воздействием специальной мускульной системы — протракторов и ретракторов. Мышечные волокна ретракторов плотно окружают основание ацикул, а таковые протракторов прикреплены к внутренней стенке параподии.
Нотоподия чешуйчатых червей развита различно: у комменсальных полиноид она обычно недоразвита, а у свободно ползающих по субстрату, так же как и у зарывающихся в грунт форм, напротив, нотоподия снабжена мощным пучком щетинок, образующим под элитрами как бы фильтр, предохраняющий респираторный канал на спинной стороне от засорения посторонними детритными и минеральными частицами. Дорсальная ветвь параподии не выполняет какую-либо локомоторную функцию, она обычно представлена в виде небольшой широкой подушечки с многочисленными рядами нотоподиальных щетинок, которые, как правило, сидят не так глубоко, как невроподиалъные щетинки, и к тому же они не втягиваются и не выдвигаются; поддерживающая их дорсальная ацикула значительно тоньше и короче вентральной.
Форма параподии в разных семействах весьма разнообразна (рис. 9), но во всех случаях она хорошо адаптирована к способам передвижения: ползанию-по твердому субстрату («маршированию»), зарыванию в мягкий грунт и обитанию-в трубках или иных убежищах. Параподии Aphrodita
(рис. 9,1)
вооружены мощными дорсальными щетинками в виде гладких или зазубренных игл, торчащих в разные стороны, и очень длинными волосовидными щетинками, образующими поверх спинных чешуек густой войлок; такое вооружение параподии, с одной стороны, способствует быстрому закапыванию червя передним своим концом в грунт путем поочередных полувинтовых движений тела, а с другой стороны — надежно предохраняет спинную сторону червя, выполняющую респираторную функцию, от засорения. Когда Aphrodita
зарыта в грунт, наружу торчат лишь спинные усики нескольких самых задних сегментов. Крупные черви Polyodontidae
(рис. 9, 7),
строящие собственные толстые илистые трубки, имеют параподии, слабо расчлененные на дорсальную и вентральную ветви, но снабженные специальными «паутинными железами», секрет которых в виде тонких золотистых волокон служит основой для стенок их трубок; эти золотистые волокна образуются хетобластами, а потому их следует рассматривать как одну из модификаций щетинок. Наиболее сложно устроенные параподии с большим набором различных щетинок наблюдаются у Sigalionidae
(рис. 9, 6)
и Eulepethidae
(рис. 9, 8),
что связано с закапыванием в грунт и обитанием в трубках. Представители Chrysopetalidae
имеют параподии (рис. 9, 9),
на спинной ветви которых в виде широкого веера располагаются золотистые крупные палены, прикрывающие наподобие элитр спинную сторону червя.
Один из наиболее обычных и простых способов передвижения чешуйчатых червей — «марширование» (шагательные движения), при котором невропо-диальные щетинки действуют как рычаги, все тело животного как бы передвигается на ходулях. Довольно сложная работа самих параподии, как показали недавние экспериментальные исследования, строго координируется туловищным мозгом.
Нам пришлось наблюдать движения Lepidonotus squamatus
в аквариуме: при спокойном состоянии животного тело его неподвижно, однако параподии все время поочередно выдвигаются наружу, а затем сокращаются — втягиваются обратно, что представляет собой чисто дыхательные движения. Когда животное потревожено, тело производит легкие волновые изгибы, начинающиеся от заднего конца, а параподии резко усиливают свою деятельность: во втянутом состоянии они выносятся вперед, а затем в расправленном виде с сильно выдвинутыми щетинками производят отталкивающий удар назад наподобие движения весла. При поступательном движении червя спинные его усики и головные щупальца обычно неподвижны. У всех чешуйчатых червей при поступательном движении их форма и размеры меняются незначительно.
Остановимся несколько подробнее на поисковых движениях Iphione muri-cata,
обитающего в прибрежной зоне Баренцева моря. Днем эти черви сидят неподвижно, плотно прикрепившись к камням, причем если их потревожить, они еще плотнее прижимаются к субстрату, от которого их можно оторвать лишь с большим трудом. Двигаться в поисках пищи они начинают через несколько часов после захода солнца, а перед восходом солнца сразу останавливаются. За ночь они успевают передвинуться всего на несколько сантиметров, причем некоторые из них к утру возвращаются на прежнее место. При ночных путешествиях элитры ритмично подымаются и опускаются, пропуская под собой ток воды. Параподии приступают к работе с заднего конца тела, одна волна движений параподии длится около 5 секунд, после чего наступает пауза в 2 секунды; параподии на каждом сегменте работают в противоположном направлении. Иногда вместо поступательного движения у Iphione
наблюдается верчение на одном месте.
2.2.4 ОСНОВНЫЕ ТИПЫ ЩЕТИНОК
Различают две категории «дефинитивных щетинок; простые — состоящие лишь из одной части, и сложные — образованные из двух частей. Последние имеют более длинный основной ствол (или рукоятку, иначе древко) и сочлененный с ним дистальный членик. Сочленение подвижное и при упоре щетинки в субстрат его членик своим зазубренным краем ложится параллельно субстрату, в результате чего усиливается сцепление. Сложные щетинки среди рассматриваемых нами семейств имеются лишь у Sigalionidae
и Chrysopetalidae.
Дистальный членик щетинок сигалионид иногда сильно вытянутый, со вторичными суставами и двузубым кончиком (рис. 10, 13).
Сложные щетинки с коротким члеником, напоминающим серп, обозначаются как серповидные, а с удлиненным, напоминающим косу, — косовидные. Сложные щетинки присущи только невроподии, выполняющей основную локомоторную функцию.
Некоторые типы простых щетинок характерны для вполне определенных родов и имеют большое систематическое значение. Так, крупные игловидные нотоподиальные щетинки темного цвета свойственны Aphrodita.
Такие же щетинки, но с загнутыми назад на их дистальном конце боковыми крючьями,
так называемые гарпунообразные (рис. 10, 1),
весьма типичны для Hermonia
и Laetmonice,
причем для последнего рода не менее характерны также невро-лодиальные «гребенчатые» щетинки (рис. 1.0, 2).

Рис. 10. Основные типы щетинок Aphroditiformia.
Polynoidae'.

Наиболее обычный тип нотоподиальных щетинок полиноид — слегка изогнутые разной толщины щетинки с многочисленными плотно расположенными поперечными рядами мелких зубчиков (рис. 10, 4, 5);
дистальный их кончик нередко волосовидный (Gattyana, Arcteobia
), в других случаях гладких закругленный или даже притуплений, иногда с расщепленной верхушкой.В невроподии наиболее обычны щетинки с загнутым однозубым или двузубым концом (рис. 10, 6, 7).
У этих щетинок различают гладкую рукоятку, или стержень, и несколько расширенную зазубренную дистальную часть. На дистальной части зубчики расположены с двух сторон, охватывая веером основную' часть щетинки; боковые зубчики могут иметь сложную дополнительную скульптуру, которая обнаруживается на фотографиях сканирующим микроскопом (рис. 11). Длина зазубренной части и количество поперечных рядов зубчиков на ней могут служить видовым признаком.

Рис. 1 Рис.11. Боковые зубчики на дистальной части щетинки Perolepis.
(Фото сканирующим микроскопом В. Г. Аверинцева).
Количество щетинок в одном пучке как в нотоподии, так и в невроподии достигает 50 и более. Верхние и нижние щетинки в одном пучке несколько различаются.
По мере износа в параподиях появляются новые тождественные щетинки. Каждая щетинка формируется одной базальной клеткой, или хетобластом, расположенным на дне так называемого щетинкового мешка эпидермального происхождения. На апикальной поверхности хетобласта имеется сложно устроенный аппарат микроворсинок (microvilli), который и определяет структуру секретируемого хетобластом вещества будущей щетинки. Рост щетинок — базальный, и по мере их роста аппарат микроворсинок может перестраиваться, в результате чего меняются форма и характер самих щетинок. Как показали еще Щепотьев (1903, 1904) и Любищев (1924а), а затем и ряд других исследователей, ствол щетинок пронизан многочисленными каналами, оставшимися в результате деятельности микроворсинок, т. е. имеет как бы трубчатую структуру. По своему тонкому строению полихетные щетинки весьма схожи с таковыми погонофор, эхиурид, брахиопод и хетогнат, и эта ультраструктура не может быть использована для филогенетических построений.
Щетинки многощетинковых червей содержат хитин в разных пропорциях. У Aphrodita aculeata
дорсальные крупные игловидные щетинки имеют хитина до 38% от сухого веса, а тонкие волосовидные латеральные и дорсальные щетинки только 19—20%. Хитин имеется также в щетинках эхиурид, погонофор и хетогнат. В других каких-либо морфологических структурах полихет, а также в их трубках хитин пока не отмечен; в частности, он не обнаружен в элитрах Aphrodita
и в челюстях полиноид.
2.2.5 ЭЛИТРЫ И ИХ ЗАЩИТНАЯ РОЛЬ
Элитры, или спинные чешуйки, представляют собой видоизмененные спинные усики, сплющенные дорсовентрально. Элитры, так же как и спинные усики, сидят на специальных выступах стенки тела — элитрофорах, которые, однако, располагаются несколько выше, чем подставки спинных усиков— циррофоры на усиковых сегментах. Такое расположение элитрофор и циррофор находит свое объяснение в том, что элитры предназначены для прикрытия спины (защитная функция), а спинные усики — осязательные органы и, возможно, хеморецепторы, в соответствии с чем они должны быть выставлены по возможности дальше в сторону.
Попытка гомологизировать элитрофоры с «дорсальными бугорками» (dorsal tubercle), расположенными на усиковых сегментах поверх спинных усиков, оказалась несостоятельной. В онтогенезе элитры и спинные усики закладываются однотипно. В морфо-функциональном отношении элитры следует рассматривать как новообразующиеся органы, а потому вначале они имели неупорядочную закладку и лишь затем, в процессе эволюции, путем олигомеризации произошло их упорядочение и строго закономерное расположение на определенных сегментах тела.
У большинства чешуйчатых червей элитры почти полностью прикрывают спинную сторону, вдоль которой непрерывно спереди назад движется ток свежей воды, используемый червями в респираторных целях. Задняя часть каждой чешуйки в той или иной степени налегает на передний край следующей за ней элитры, т. е. они образуют как бы черепичную кровлю. Лишь у представителей Polyodontidae
элитры располагаются в обратном направлении: задняя элитра находит на переднюю. Последнее у полиодонтид, возможно, связано с их постоянным обитанием в длинных трубках и необходимостью при нападении хищников быстро уходить задним концом тела в глубину своего домика, в то время как другие чешуйчатые черви преимущественно двигаются передним концом тела лишь вперед.
Элитры прикреплены к элитрофорам обычно центральной своей частью (место прикрепления округлое или слабоовальное). Однако бывает и боковое прикрепление. Последнее характерно для всех Aphroditidae,
а среди полиноид наблюдается у многосегментных Perolepis,
которых среди полиноид мы считаем наиболее примитивными формгми. Можно наметить постепенные переходы от латерально прикрепленных крохотных элитр Perolepis
и небольших, едва достигающих ширины сегментов округлых элитр Lepidasthenia
— к крупным жестким почковидным (бобовидным) элитрам Lepidonotus,
полностью прикрывающим всю спинную сторону червей (рис. 12). Этот ряд наглядно доказывает происхождение элитр из цирростилей спинных усиков.
Форма и размеры элитр несколько изменяются в зависимости от части тела, в которой они находятся: как правило, передние элитры несколько меньшего размера и округлой формы, в то время как элитры в средней части тела наиболее вытянутые и наиболее крупные. Но имеются и исключения: у некоторых Perolepis
и Lepidasthenia
наиболее крупными, в несколько раз превышающими все остальные, являются элитры первой пары. В большинстве случаев элитры покрывают друг друга вдоль всей медиальной линии спины, однако у некоторых видов средняя часть спины остается неприкрытой.
У всех чешуйчатых червей в передней части тела до XXIII сегмента расположение элитр одинаковое: II, IV, V, VII, IX, XI, XIII, XV, XVII, Х1Х,:
XXI и XXIII сегменты. Дальше чередование чешуйчатых и усиковых сегментов варьирует. Элитры нередко находятся через два усиковых сегмента, в других случаях — через один. Иногда элитры располагаются асимметрично, т. е. на одном и том же сегменте с одной стороны наблюдается элитра, а на другой стороне — спинной усик (Polyeunoa, Arctonoe).
У большинства червей элитры прикрывают тело почти до самого заднего его конца. Однако у рода Polynoe
значительная задняя часть тела свободна от элитр. Сигалиониды в задней части тела, начиная с XXIII (иногда с XXIX) сегмента, имеют элитры на каждом сегменте. Характер чередования чешуйчатых и усиковых сегментов в задней части тела используется как один из родовых признаков.
Морфология чешуек относительно простая. В элитрофор, представляющий собой простое выпячивание стенки тела, частично заходит целом. В своей ди-стальной части элитрофор системой мощных кольцевых мышц отделен от собственно элитры (своего рода сфинктер). Резкое сокращение этих мышц при раздражении червя обеспечивает аутотомию элитр. Сами элитры имеют разнообразную фактуру. Некоторые из них очень нежные, мягкие, другие же, напротив, жесткие, сильно склеротизированные.
В качестве крайнего примера жестких элитр можно привести элитры Iphione muricata.
Элитры этого червя имеют бросающуюся в глаза ячеистую (полигональную) структуру, напоминающую собой пчелиные соты («elytra reticulata»). Почти вся их наружная поверхность сложена из крупных, хорошо отгороженных друг от друга шестигранных ячеек, которые в свою очередь имеют многочисленные более мелкие ячейки. Последние не что иное, как отдельные клетки с толстой клеточной мембраной. Внутри этих клеток имеется по крупному ядру, причем плазма прижата к стенкам клеток. Под первым (верхним) слоем наиболее крупных клеток располагаются второй, третий, четвертый и даже пятый слои из более мелких клеток.

Рис.12 Изменение формы параподий и элитр в ряду Perolepis (А)
— Lepidasthenia (Б)
— Lepidonotus (В).
Все клетки плотно прижаты друг к другу, и вся эта конструкция напоминает растительную ткань. Крупные ячейки отделены друг от друг поперечными прослойками толстой кутикулы. Такого рода жесткие элитры, полностью прикрывающие спинную сторону червя, крепко прижавшегося к скалам, предохраняют его от подсыхания и позволяют ему в неподвижном состоянии находиться долгие часы под палящим тропическим солнцем. Ячеистая структура элитр, но менее жесткая, чем у Iphione muricata,
наблюдается также у Gattyana deludens
, G. iphionelloides
, Harmothoe areolata, H. reticulata, H. dictyophora
и некоторых других.
У мягких элитр, к которым относятся элитры H", lunulata
и некоторых других полиноид, лишь верхняя и нижняя их поверхности имеют небольшой слой кутикулы, богатой кератиноидами. Изнутри кутикула выстлана одним слоем призматического эпителия, а все остальное внутреннее пространство элитры заполнено соединительной тканью, пронизанной единичными мышечными волокнами. Через ствол элитрофора в элитры проникают нервы, имеющие связь с брюшной нервной цепочкой. У места прикрепления элитры к элитрофору обычно наблюдается густое нервное сплетение (plexus), от которого отходят в разные стороны древовидно ветвящиеся небольшие нервные волокна, хорошо заметные при рассмотрении элитр в проходящем свете. Тончайшие нервные фибриллы заканчиваются, с одной стороны, в фотогенных клетках (фотоцитах), находящихся у нижней поверхности элитры, а с другой — в мягких папиллах, нередко густо покрывающих почти всю верхнюю сторону элитр, в особенности у их наружного края (на нижней стороне элитр папилл нет, эта поверхность всегда гладкая, глянцевидная). Мягкие папиллы, представляющие собой небольшие чувствительные органы, могут быть как одноклеточными образованиями, так и многоклеточными.
На верхней стороне элитр, помимо мягких папилл, обычно наблюдаются и многочисленные склеротизированные папиллы весьма разнообразной формы. Последние не иннервированы и, видимо, не выполняют чувствительных функций. Склеротизированные образования могут быть в виде небольших острых иголочек, крупных заостренных шипов или наподобие грибовидных образований с закругленной или со звездчатой верхушкой, иногда в виде бугорчатых бородавок, древовидно разветвленных кустиков и т. д. Микроструктура этих образований и их расположение имеют большое видовое таксономическое значение. Детали скульптуры верхней поверхности элитр хорошо обнаруживаются с помощью растрового электронного микроскопа.
Элитры некоторых полиноид (Polynoe scolopendrina, Acholoe astericola,
различные Harmothoe, Lepidasthenia
и др.) при раздражении светятся. Излучение света внутриклеточное (гранулы в апикальных частях фотогенных клеток). Свечение происходит в виде коротких и повторных вспышек и контролируется ганглиями брюшной нервной цепочки (рефлекторные реакции). При сильном раздражении одновременно аутотомируются и сами элитры, которые продолжают люминесцировать в оторванном состоянии достаточно долгое время. Последнее в некоторых случаях, возможно, вводит в заблуждение напавшего на полиноиду хищника, и пока оторванные элитры светятся, сам червь «обманным путем» ускользает от врага. После аутотомии элитр червь их регенерируют полностью. Недавно было констатировано, что элитры могут служить своеобразной выводковой камерой для молоди, наподобие крышечек некоторых Spirorbis.
Так, в элитрах небольшой сигалиониды Pholoe swedmarki,
адаптированной к интерстициальному биотопу на Бермудских островах, а также у Ph. antipoda
Hartman, 1967 из антарктических вод (Аверинцев, in litt.) обнаружены вполне сформировавшиеся молодые черви, имеющие до 6 сегментов.
3. ЛИЧИНОЧНЫЕ СТАДИИ
Постэмбриональное развитие большинства Aphroditiformia
связано с пелагической трохофорой, претерпевающей ряд «возрастных» изменений, которым соответствуют следующие стадии: 1) трохофора (несегментированная личинка),  2) метатрохофора I и II и 3) нектохета I и II (сегментированные личинки). Трохофора у всех чешуйчатых червей монотрохного типа, т. е. с одним ресничным пояском. 2) метатрохофора I и II и 3) нектохета I и II (сегментированные личинки). Трохофора у всех чешуйчатых червей монотрохного типа, т. е. с одним ресничным пояском.
Личинки всех чешуйчатых червей построены по единому плану, отличному от личинок остальных многощетинковых червей. Этот тип личинок, характеризующийся длинным боковым пучком ресничек, расположенным слева от рта и похожим на бороду, предложено называть «barbularia».
Первые данные о личинке наиболее широко распространенного вида — Harmothoe imbricata
— были сообщены еще в середине прошлого столетия М. Сарсом. В настоящее время морфология и анатомия личиночных стадий этого вида подробно изучены многими исследователями; освещены также различные аспекты биологии размножения и постэмбрионального роста.
Размножение Н. imbricata
в большинстве районов ареала приходится в основном на холодные зимние и весенние месяцы. По нашим морям мы имеем следующие данные: в Кольском заливе половозрелые особи отмечены в январе—феврале, а трохофоры под элитрами — в марте, на Восточном Мурмане (Дальние Зеленцы) зрелые яйца в полости тела наблюдались в конце мая, а личинки в планктоне — до начала августа, в открытых районах Баренцева моря личинки в планктоне встречались в мае—июне, в Белом море примерно в те же сроки — с апреля по июнь, у южных Курильских островов экземпляры со зрелыми половыми продуктами в полости тела наблюдались с декабря по апрель, в Японском море (залив Посьета) трохофоры под элитрами отмечены с марта по май, а пелагические личинки — в мае, в Черном море в планктоне личинки встречались с декабря по март.
Характер размножения зависит от экологических условий географического района.
Зрелые яйца Н. imbricata,
имеющие беловато-розовую окраску, сильно варьируют в размерах в зависимости от количества желтка; в заливе Посьета диаметр яиц 120—135 мкм. Различные личиночные стадии Н. imbricata
изображены на рис. 13. Трохофора сферической формы, диаметром до 200—400 мкм, с парой коричнево-красных глаз, снабженных линзой. Ротовое отверстие прикрыто нависающей сверху широкой губой. Основной ресничный поясок — прототрох — трехрядный. От рта до анального отверстия по брюшной стороне узкой полоской тянется невротрох. На границе пищевода и средней кишки у трохофоры инвагинацией образуется пара глоточных дивертикулов, которые представляют собой зачаток будущей глотки. Стенка средней кишки заполнена желточными гранулами темно-карминного или сиреневого цвета. Метатрохофоры I и II имеют яйцевидную форму. Ресничный покров сохраняется в первоначальном виде, и только в центре эписферы возникает слабый апикальный пучок ресничек. К первой паре глаз прибавляются еще две пары новых глаз, также с преломляющими линзами. На стадии метатрохофоры образуется восемь пар щетинконосных мешков (восемь ларвальных сегментов). Будущие параподии закладываются в виде бугорков, расположенных в три ряда. Первый ряд с дорсальной стороны представляет собой зачатки спинных усиков и элитр, причем более крупные бугорки соответствуют будущим элитрам, а маленькие — спинным усикам. Во втором ряду бугорки одинакового размера, из них впоследствии развиваются невроподии. Третий ряд одинаковых по размеру бугорков дает начало брюшным усикам. На стадии нектохеты число сегментов сохраняется равным восьми. На простомиуме появляются зачатки трех щупалец и двух пальп. Формируется параподиальный комплекс: элитры становятся плоскими, а невроподиальные ветви со щетинками сильно выдвигаются в стороны. На этой стадии нотоподиальных выростов еще нет.

Рис 13. Личиночные стадии Harmothoe imbricata.
1
— трохофора, 2
, — метатрохофора, 4
— нектохета со спинной стороны. 5 — она же, с брюшной стороны, 6
— оседающая молодая особь, вид сверху, 7
— она же, с брюшной стороны.
Параподии постепенно начинают совершать движения и помогают работе ресничного пояса. Все развитие, от момента оплодотворения до оседания нектохеты на дно, в Белом море длится около полутора месяцев.
Трохофоры и личинки других видов полиноид весьма схожи с только что описанными, но число ларвальных сегментов варьирует: Lepi-donotus squamatus
и L. clava
— 8, Harmothoe lunulata
— 9, H. longisetis
— 10, Lagisca extenuata
— 9, Acholoe astericola
— 9, Halosydna brevisetosa
— 9, Alentia gelatinosa
— 9. Эписфера трохофоры последнего вида несколько отличная: она резко сплющена дорсовентрально, а потому трохофора имеет не яйцевидную форму, а напоминает скорее короткий усеченный конус.
Сходное развитие наблюдается и в сем. Sigalionidae
, трохофора Sthenelais boa
имеет чечевицеобразную форму, у нее эписфера несколько крупнее гипосферы. Ротовое отверстие с крупными губами. Одна пара коричневатых глаз с линзами. На 15-е сутки у почти сферической метатрохофоры I помимо пер.едней нары округлых бугорков (будущих тентакулярных усиков) намечается 6 двуветвистых параподий с ацику-лами и зачатками щетинок. Метатрохофора II уже снабжена тремя парами хорошо развитых элитр с удлиненными папиллами; добавляются две новых пары глаз, и на дорсальной стороне эписферы появляется крупная па- пилла (будущее медиальное щупальце). Нектохета I характеризуется наличием 4 пар элитр; последняя пара элитр принадлежит 7-му сегменту — первому постларвальному. Нектохета II имеет 8 сегментов, у нее появляются зачатки латеральных щупалец, пальпы достигают размеров медиального щупальца. На этой стадии личинка оседает на дно.
В отличие от Sthenelais,
нектохета I небольшой сигалиониды Pholoe
имеет шсего 4 пары параподий, не считая перистомиальных усиков. Развитие Pholoe minuta
из наших вод подробно изучено в Японском море.
Личинки хризопеталид (Chrysopetalum, Paleanotus)
характеризуются наличием у них двух крупных пучков весьма длинных зазубренных щетинок по бокам ротового отверстия под прототрохом.
Эти щетинки темного цвета н по своей длине значительно превосходят размеры тела личинок на стадиях трохофоры и метатрохофоры I. У метатрохофоры I Chrysopetalum debile,
по данным Казо, сразу закладываются 5 зачаточных параподий и через час появляется шестой ларвальный сегмент. Крупные подиаль-ные щетинки — палены, характерные для данного семейства, имеются уже у метатрохофоры II и они полностью прикрывают спинную сторону личинки на стадии нектохеты.
4. ЭКОЛОГИЯ
Многие полиноиды обитают в прибрежной зоне Баренцева моря, и крепко прикрепляются к камням; некоторые соответствуют планктонному образу жизни (Bylgides Sarsi).
Полиноиды (Psammolyce
) зарываются в песок и ползают по поверхности грунта, крупные черви (Polyodontida
e) строят свои собственные толстые илистые трубки.
5. ГЛАВНВЫЕ НАПРАВЛЕНИЯ ЭВОЛЮЦИИ СЕМЕЙСТВА ПОЛИНОИД
Положение в общей системе полихет чешуйчатых червей и близких к ним семейств, объединенных нами в подотряд Aphroditiformia,
уже было рассмотрено ранее. Здесь остановимся на филогенетических отношениях в наиболее богатом родами семействе Polynoidae.
По современным данным, в состав этого семейства входит около 100 родов. Некоторые из них еще нуждаются в глубокой ревизии, и таксономический их статус не вполне ясен. Однако несмотря на это в семействе полиноид более или менее четко намечаются несколько подсемейств, отличающихся между собой в основном особенностями строения головного отдела.
К ранее выделенным трем подсемействам — Lepidonotinaey
Harmothoinae
и Iphioninae
— в последнее время было добавлено два других: Macellicephalinae
и Admetellinae
. Из указанных подсемейств лишь два первых насчитывают большое количество родов (примерно по 40). При этом существенно, что в обоих основных подсемействах имеются как роды, представители которых характеризуются относительно коротким телом, состоящим из небольшого числа сегментов и снабженным строго определенным (ограниченным) количеством элитр, так и роды, для которых характерно весьма большое число сегментов и неопределенно большое количество элитр. Естественно возникает вопрос, какие формы полиноид — с коротким или длинным телом — следует считать за родоначальные — исходные в обоих подсемействах.
Высказываний по этому поводу в литературе немного. Французский исследователь Дарбу в основу системы ставит род Harmothoe,
от которого, он считает, произошли, с одной стороны, Lagisca—Polynoe,
а с другой — Lepidonotus—Lepidasthenia,
т. е. развитие шло от малосегментных к многосегментным. При этом, по его мнению, у Lepidasthenia
будто бы имела место редукция нотоподии, связанная с комменсализмом, и этот род как первоначально факультативный комменсал полихетных трубок дал начало Polyodontidae,
постоянно живущим в трубках, построенных самими червями.
Рис. 15. Полипоида Arctonoe vittata
(Grube)в мантийной полости моллюска Acmaea pallida
(Gould) из Японского моря.
Нам представляется более вероятным, что Polyodontidae
произошли от каких-то предковых форм Sigalionidae,
так как среди сигадионид имеется род Sthenelanella,
у которого наблюдаются такие же в высшей степени своеобразные «паутинные железы», как и у представителей полиодонтид. Что же касается параподий, то, первичными следует считать одноветвистые параподий, а образование у полиноид нотоподии связано с дополнительной функцией, возникшей в связи с появлением элитр. Немецкий исследователь Зайдлер касается вопроса об исходной форме полиноид весьма кратко. Для лепидонотин за таковую он считает Lepidonotus,
т. е., по его мнению, многосегментные также произошли от малосегментных.
Нам представляется, что процесс эволюции протекал в обратном направлении, современные малосегментные полиноиды с небольшим количеством элитр представляют собой конечную ветвь развития как в подсемействе Lepidonotinae,
так и в подсемействе Harmothoinae.
Уменьшение количества сегментов и их стабилизация у полиноид произошли в результате специализации, как это имело место и во многих других семействах полихет. В частности, среди фил-лодицид, для которых характерен большой комплекс примитивных черт, такая стабилизация количества сегментов наблюдается лишь у специализированных пелагических видов.
Первичные полиноиды, надо думать, произошли от каких-то многосегментных червей, имевших на первых этапах развития гомономное строение, т. е. все их сегменты вначале имели одинаковые подиальные придатки (лишь удлиненные циррусы). Преобразование части этих придатков в элитры было связано у свободноподвижных донных форм полиноид с необходимостью защиты спинной стороны их тела от засорения, аналогично тому, что мы наблюдаем и у крупных донных филлодоцид. Разница здесь лишь в том, что у филлодо-цид циррусы приобретали характер широких лепестков, прикрывающих спинную сторону червя, в результате бокового сплющивания, а элитры возникали из циррусов путем их дорсовентрального сплющивания. Преобразование у первичных полиноид части циррусов в элитры нарушило у этих червей начальную гомономность их сегментации. Как новообразования- элитры у родоначальных форм возникали, вероятно, вначале произвольно, т. е. беспорядочно на различных сегментах. Лишь в результате последующей олигомеризации, в понимании В. А. Догеля (1954), произошло их упорядочение. В соответствии с этим у большинства полиноид элитры располагаются на строго определенных сегментах, во всяком случае до XXIII сегмента. Однако у некоторых современных полиноид с длинным телoм чередование элитр и спинных усиков на задних сегментах все же иногда сохраняется весьма неправильным и сильно варьирует, причем у некоторых форм наблюдается даже асимметричное расположение элитр, т. е. с одной стороны сегмента имеется элитра, а с другой стороны того же сегмента спинной усик, например Lepidasthenia, Polyeunoa
и Arctonoe
. Указанный неустойчивый характер расположения элитр на задних сегментах у названных современных полиноид следует рассматривать как довольно примитивную особенность, сохранившуюся от родоначальных форм.
Общие процессы эволюции чешуйчатых червей нам представляются в следующем виде. Как всякие свободноползающие многосегментные животные, недостаточно защищенные от врагов, они были вынуждены отыскивать какие-то укрытия. Некоторые стали зарываться в грунт (Sigalionidae).
Многосегментные полиноиды начали залезать в чужие трубки, и некоторые из них стали факультативными комменсалами. Такой образ жизни сохранился и у ряда современных полиноид. Комменсализм для них оказался преимущественно консервативным фактором, задержавшим дальнейшую их эволюцию, которая шла по линии сокращения количества сегментов и более мощного развития чешуйчатого покрова. Можно наметить следующий общий ряд: роды с длинным телом, снабженным небольшими элитрами, а в соответствии с этим и со слабо развитыми нотоподиями, через промежуточные формы дали начало родам с коротким телом и небольшим количеством элитр, полностью прикрывающих спинную сторону червей. Элитры постепенно приобрели надежную защитную функцию, и черви смогли вести свободный хищнический образ жизни. Специализация у родов с коротким телом, как отмечалось в нашем общем морфологическом очерке, достигает наиболее высокого уровня.
Одним из наглядных доказательств правильности высказанных здесь соображений может служить род Perolepis,
сохранивший ряд наиболее примитивных черт для полиноид. Наряду с длинным телом, состоящим из большого числа сегментов, у представителей этого рода элитры зачаточного типа. На туловищных сегментах у этих червей элитры крайне небольших размеров и прикреплены к сильно вытянутым элитрофорам латерально, что необычно для других современных полиноид. Латеральный способ прикрепления элитр характерен для Aphroditidae
s. str. Элитрофоры и циррофоры у Perolepis
еще не различаются по своей форме и находятся на одном уровне (вдоль одной линии), подобно тому как расположены спинные усики у многих эррантных полихет (Phyllodocidae, Syllidae
и др.), для которых характерна полная гомономность туловищных сегментов. У остальных же полиноид элитрофоры по отношению к циррофорам обычно сильно сдвинуты на спинную сторону. Такое перемещение, способствующее более полному налеганию друг на друга чешуек вдоль спинной медиальной линии, следует рассматривать как дальнейший этап эволюции, непосредственно связанный с основным назначением самих элитр.
В роде Perolepis
хорошо представлены все постепенные переходы преобразования спинных усиков в элитры: вначале имело место укорочение цирро-стиля и образование на нем с брюшной стороны небольшого вдавления, в результате чего цирростиль получил «ложковидную» форму (Perolepis sibogae, P. pacified);
в дальнейшем произошло еще большее уплощение и расширение этого органа (P. ohshimai, P. pettiboneae).
От указанного зачаточного типа элитр Perolepis
(рис. 12) нетрудно перейти к элитрам Lepidasthenia,
которые хотя и очень небольших размеров и не прикрывают спинную сторону червя, но уже обычного для полиноид абриса и отходят от коротких элитрофоров, несколько сдвинутых на спинную сторону червя. У остальных полиноид элитры, как правило, широкой овальной формы и черепицеобразно находят друг на друга, надежно защищая спинную сторону червя от засорения (под элитрами располагается респираторный канал). Таким образом, по характеру строения элитр род Perolepis
можно условно принять за прототип, давший начало другим представителям полиноид, в частности Lepidonotinae,
у которых терминально расположенные латеральные щупальца еще не имеют цератофоров.
Роды, относящиеся к подсемейству Lepidonotinae,
хорошо укладываются в следующий общий ряд: Lepidasthenia
(элитр свыше 50 пар, сегментов более 100)~Parahalosydnopsis
(38—28, сегментов до 75)—Halosydnella
(24—20, сегментов до 47)—Alentiana
(20—15, сегментов до 39)—Halosydna
(18, сегментов 37—36)—Lepidonotus, Euphione, Hermione
(12, сегментов 27—26). В указанном ряду уменьшение количества элитр сопровождается усложнением их структуры. Некоторое исключение составляет Hermione,
у которого по не совсем ясным причинам элитры подверглись редукции в размерах. Центральное место по количеству видов в данной ветви развития занимает Lepidonotus
(около 75 видов), представляющий в морфологическом отношении, по всей видимости, наиболее приспособленную форму, а отнюдь не родоначальную.
По современной систиматике к Lepidonotinae
относят также роды с «аркто-ноидным» типом головной лопасти, у которых терминально и субтерминально расположенные латеральные щупальца имеют ясно выраженные цератофоры. Эти роды в филогенетическом отношении тесно связаны с многосегментной по-линоидой Arctonoe,
но не образуют четко выраженной самостоятельной линии развития, как роды с лепидонотоидньш типом головной лопасти. В этой группе не имеется центрального рода, наподобие Lepidonotus.
К ним, в частности, относятся как многосегментные Gastrolepidia, Pottsiscalisetosus,
так и формы с более коротким телом — Weberia, Alentia, Arctonoella, Parahalosydna, Paralepi-donotus, Drieschia
и некоторые др. Эти роды представляют собой переходную группу к Harmothoinae,
имеющим вентрально расположенные латеральные щупальца с цератофорами. Возможно, в дальнейшем указанные роды с аркто-ноидным типом головной лопасти потребуется выделить в самостоятельное подсемейство.
Аналогичная Lepidonotinae
картина постепенной стабилизации количества сегментов и элитр наблюдается и среди Harmothoinae:
здесь тоже имеются роды с неопределенно большим числом сегментов и элитр (Polyeunoa, Grubeopolynoe
и др.), причем они также преимущественно комменсалы. В подсемействе Harmothoinae
больше всего родов с 15 парами элитр, полностью прикрывающими спинную сторону червей. Центральное положение по количеству видов занимает Harmothoe
(около 100 видов). Следует отметить, что у Harmothoe
и у некоторых других родов с 15 парами элитр хорошо выражены лобные рога, в то время как у родов с длинным телом лобные рога еще отсутствуют. Последнее указывает на то, что Harmothoe
в некоторых отношениях более высоко организованный род, чем остальные. Несколько обособленное положение в этом ряду занимает Polynoe
s. lato, у которого количество сегментов может быть весьма большим, а количество элитр всего 15 пар. Родов с 12 парами элитр в этом ряду немного (Polynoella, Antinoana).
В нашей схеме (рис. 16), иллюстрирующей некоторые родовые филогенетические отношения в пределах семейства полиноид, хорошо видно, что в двух основных подсемействах — Lepidonotinae
s. str. и Harmothoinae
— роды образуют очень сходные параллельные ряды развития, что свидетельствует о наличии общих процессов эволюции в пределах всего семейства Polynoidae.
На этой схеме отмечена лишь часть родов, но она в какой-то степени охватывает и все остальные известные в настоящее время роды; длина линий для родов соответствует примерному количеству видов в этих родах.
В состав подсемейства Macellicephalinae
входит несколько родов, представители которых встречаются преимущественно в абиссали и обнаруживают многие общие морфологические особенности, связанные со специфическими условиями жизни на больших глубинах. Тело у представителей этой группы полиноид в большинстве случаев весьма мягкое, элитры очень нежные и легко отпадают (от 8 до 15 пар), щетинки тонкие и длинные, параподии сильно вытянутые, а спинные усики иногда достигают исключительной длины. Все это облегчает плавание в придонных слоях воды в поисках пищи. Как чисто адаптивное приспособление, связанное с добычей пищи на абиссальных глубинах, где пищевые ресурсы сильно разрежены, следует рассматривать образование на глотке Macellicephaloides
двух длинных боковых усиковидных придатков (как у некоторых альциопид). Наряду с этим у мацеллицефалин наблюдается процесс постепенного исчезновения глоточных зубов, характерных для остальных полиноид: у Macelloides
они заменены слабыми пластинками, а у Macella
вообще отсутствует какое-либо хитиноидное вооружение на глотке. Представители Macellicephalinae,
надо думать, проникли на большие глубины сравнительно недавно и ведут свое начало от каких-то батиальных (или батипелагических) предков, имевших уже относительно короткое тело. По не вполне понятным причинам у них произошла редукция латеральных щупалец. Заостренные окончания боковых долек простомиума, описанные для некоторых видов Macellicephala
как лобные рога, в действительности представляют собой не что иное, какрудименты латеральных щупалец (эти выросты легко отчленяются при слабом нажиме).

Рис.16. Основные направления развития полиноидных червей
В качестве одной из предковых форм мацеллицефалин можно рассматривать Bathymoorea,
у которого латеральные щупальца еще развиты, но медиальное щупальце сильно сдвинуто на дорсальную сторону простомиума.
Выделенное нами подсемейство Admetellinae
состоит из двух родов — Admettella
и Bathyadmetella,
у представителей которых на головной лопасти появляются своеобразные добавочные придатки в виде «antennal scales» и «tentacular sheath», наподобие ктенидий Sigalionidae.
Параподии адметеллин сильно вытянуты, с тонкими длинными щетинками, количество сегментов свыше 50 и элитр свыше 20 пар. Точное положение этого подсемейства в общей системе полиноид пока не вполне ясное. Но во всяком случае оно тесно связано с пред-ковыми многосегментными формами лепидонотин.
Высказанные здесь соображения о филогенетических отношениях в пределах полиноид имеют предварительное, чисто рабочее значение, но эти соображения смогут оказаться полезными при дальнейшей разработке классификации не только полиноид, но и некоторых других семейств полихет
ВЫВОДЫ
На основании рассмотренного материала и поставленных целей можно сделать несколько выводов:
1)На современном этапе используется систематическая трактовка, или термин, предложенный Левинсеном, под которым без точного пояснения его таксономического ранга этот автор объединил всех чешуйчатых червей и Palmyridae.
Aphroditiformia
трактуетсякак подотряд в системе отряда Phyllodocemorpha
. Указанный подотряд в нашем понимании в настоящее время принят и в сводном каталоге семейств и родов многощетинковых червей, опубликованном в США. По этому каталогу подотряд Aphroditiformia
состоит из трех надсемейств: Aphroditacea
(с 6 семействами), Chrysopetalacea
(с 2 семействами) и Pisionacea
(с 1 семейством).
2) одни из отличительных особенностей морфо-функционального очерка Aphroditiformia являются: спинная сторона прикрыта чешуйками (элитрами), благодаря которым при раздражении происходит свечение(внутриклеточное излучение); развитие личинки протекает со стадией трохофора; наличие двух пар глаз: задние и передние; наличие 6 типов головной лопасти, пальпов.
3)в направлениях эволюции есть несходства в представлениях о родоначальном классе. В наше время представляется более вероятным, что Polyodontidae
произошли от каких-то предковых форм Sigalionidae,
так как среди сигадионид имеется род Sthenelanella,
у которого наблюдаются такие же в высшей степени своеобразные «паутинные железы», как и у представителей полиодонтид;
Также нельзя не отметит, что процесс эволюции протекал в обратном направлении, современные малосегментные полиноиды с небольшим количеством элитр представляют собой конечную ветвь развития как в подсемействе Lepidonotinae,
так и в подсемействе Harmothoinae.
Уменьшение количества сегментов и их стабилизация у полиноид произошли в результате специализации, как это имело место и во многих других семействах полихет;
Можно наметить следующий общий ряд: роды с длинным телом, снабженным небольшими элитрами, а в соответствии с этим и со слабо развитыми нотоподиями, через промежуточные формы дали начало родам с коротким телом и небольшим количеством элитр, полностью прикрывающих спинную сторону червей. Элитры постепенно приобрели надежную защитную функцию, и черви смогли вести свободный хищнический образ жизни. Специализация у родов с коротким телом, как отмечалось в нашем общем морфологическом очерке, достигает наиболее высокого уровня.
ОСНОВНЫЕ ТЕРМИНЫ
Анальная лопасть, или пигидий (pygidium) — задний отдел тела, образующийся из нпжнего полушария трохофоры.
Анальные усики (cirri anales) — порошицевые усики, или уриты (urites), расположенные по краю анальной лопасти.
Антеннальная чеш]уйка (squamula antennalis) — лепестковидные придатки при церафоре медиального щупальца у Admetellinae.
Антенны (antennae) — см. головные щупальца.
Апикальный орган — пучок длинных чувствительных ресничек на макушке трохофоры.
А ц и к у л a (acicula) — внутренняя осевая щетинка, поддерживающая подиальнуго ветвь (нотоподию или невроподию); к ацикуле прикреплены мускульные волокна, обеспечивающие втягивание наружных щетинок.
Ацикуловпдная щетинка (seta acicularis) — простые толстые щетинки, иначе игловидные щетинки.
Ацикулярный бугорок, или вырост (lobus acicularis) — вырост на пара-подиях, в который упирается ацикула.
Бахромчатый орган (papilla dorsalis fimbriata) — у афродитид фестончатые выросты при дорсальных бугорках (см.), выполняющие функции жабр.
Буккальный сегмент (segmentum buccale) — у чешуйчатых червей второй сегмент, окружающий с боков на брюшной стороне ротовое отверстие; от последующих туловищных сегментов он отличается лишь наличием очень длинных брюшных усиков (cirri buccales).
Вентральный усик (cirrus ventralis) — усик на нижней (брюшной) стороне пара-подий, иначе брюшноД усик.
Гетерогомфная щетинка (seta heterogompha) — тип сложных (сочлененных) щетинок, у которых основной стержень (рукоятка) на дистальном конце имеет два неравных по величине сочленительных зубца.
Глазной черенок — см. омматофоры.
Глотка (pharynx) — мускулистая часть передней кишки, служащая для захвата пищи, у многих снабжена хитиноидными челюстями.
Головная лопасть, или простомиум (prostomium) — предротовая часть червя, образующаяся из верхнего полушария (эппсферы) трохофоры.
Головные щупальца (antennae, tentaculae) — тактильные выросты на головной лопасти; различают непарное среднее, или медиальное щупальце (antenna mediana). и боковые, или латеральные щупальца (antennae laterales),
Гомогомфная щетинка (seta homogompha) — тип сложных (сочлененных) щетинок, у которых основной стержень (рукоятка) на дпстальном конце имеет два одинаковых по величине сочленительных зубца.
Двуветвистая параподия (parapodium biremis, biranms) — параподпя, разделенная на две ветви: спинную, илп нотоподию (см.), и брюшную, или невроцодию (см.), каждая из которых снабжена собственным пучком щетинок. В тех случаях, когда спинная ветвь развита слабо (представлена лишь небольшим бугорком с ацику-лой), параподии называются «полуразветвленные» или «ложнодвуветвпстые» (sesqui-ramus, subbiramus).
Дорсальный усик (cirrus dorsalis) — усик на верхней (спинной) стороне параполш, иначе спинной усик. Жабры (branchiae) — различного рода отростки (усиковидные, древовидные и т. д.) преимущественно при нотоподиях; различают целомические жабры в виде простых выпячиваний стенки тела и жабры с богато разветвленной системой капиллярных сосудов.
Затылочная, или нухальная складка — передняя часть II сегмента. частично прикрывающая головную лопасть сзади. Капиллярная щетинка (seta capillaris) — простая, длинная щетинка, иначе волосовидная щетинка. Карункул (caruncula) — чувствительный орган в виде продольного или округлого валика, позади головной лопасти (преимущественно у амфиномид, имеется у хрпзо-петалид).
Краевые глоточные папиллы — см. оральная папилла.
Ктенидий (ctenidium) — различной формы небольшие выросты, покрытые ресничками как на головной лопасти, так и на параподиях (преимущественно у сигалионпд).
Ларвальные сегменты — передние сегменты, одновременно закладывающиеся при метаморфозе трохофорной личинки (остальные сегменты, образующиеся позднее из преданальной зоны роста, обозначаются как постларвальные).
Лобные рога (cornua frontalia) — заостренные выступы на дольках головной ло-' пасти у ряда полипоид.
Лобный гребень, или фасциальный бугорок (tuberculum faciale),— мягкий склал-чатый вырост на вентральной стороне простомнума, расположенный под медиальным головным щупальцем и направленный вперед.
Лож несложные щетинки — щетинки, у которых дистальный членик не отделен от основного стержня щетпнкп.
Мешковидная глотка (proboscis ventralis) — имеет характер слепого мешка (bulbus buccalis), расположенного под пищеводом.
Невроподиальная щетинка (neuroseta) — щетинка брюшной ветви пара-подий, неврохета.
Невроподия (neuropodium) — брюшная ветвь параподии (ramus inferus, ramus vou-tralis).
Нектохета — последняя стадия планктонных личинок полихет. Нефридиальная папилла (papilla nephridialis) — удлиненная папилла, обычно находящаяся на нижней или задней стороне невроподин; на дистальном ее конце расположено наружное отверстие нефридий — нефропора. Нотоподиальная щетпнка (notoseta) — щетинка спинной ветви параподии, нотохета.
Нотоподия (notopodium) — спинная ветвь параподии (ramus superus, ramus dorsalis).
Нухальная скл а'д к а — см. затылочная.
Одноветвистая параподия (parapodium uniremis, uniramus) — параподпя с одной ацикулой и с одним подиальным бугорком, вооруженным одним пучком щетинок.
Ом мат офоры (ommatophorae) — удлиненные выросты на головной лопасти, на коице которых находятся глаза (глазные черенки); наиболее хорошо развиты у Polyodon-
Оральная папилла (papilla oralis) — паппллы, окружающие входное отверстие глотки, пли хобота, иначе краевые глоточные паппллы.
Палены (palea) — крупные, слегка сплющенные золотистые щетинки, прикрывающие спинную сторону некоторых представителей Chrysopetalidae,
а также щетинки на пер1дних сегментах у амфаретпд, пектпнарий и сабелларпид, образующие опахало (веер) или своеобразную крышечку, замыкающую наружное отверстие трубки.
Пальп о фор (palpophorus) — базальная часть двучленистых пальп.
Пальцы, пли щупики (palpi) — парные придатки головной лопасти, расположенные у чешуйчатых червей и ряда других эррантных лолпхет с брюшной стороны близ ротового отверстия.
Паиv
ллa (papilla) — разнообразные мелкие выросты на поверхности тела; бывают различной формы — шаровидные, нитевидные, кеглевидные и т. д.
Параподия (parapodium) — боковые выросты сегментов, вооруженные щетинками п снаСжсьвые спинным и брюшным усиками.
Паутинная железа — подпальные железы у Polyodontidae,
выделяющие золотистые нглл, идущие на построение трубок,
Подвальная лопасть (lamella), иногда обозначается как подиальная губа — листовидный вырост на конце параподии впереди щетинок (предщетинковая лопасть — lamella presetalis) или позади их (послещетинковая лопасть — lamella post-setalis).
Подпальный ствол (truncus parapodialis) — вытянутое основание параподнй, до их разделения на нотоподию и нейроподию.
Подиальный язычок (ligula) — вытянутый, пальцевидной формы вырост на конце параподии.
Провентрикулус (proventriculus) — сильно мускулистая и богатая железами, бочковидной формы передняя часть пищеварительной трубки, расположенная сразу позади глотки.
Простомиум (prostomium) — см. головная лопасть.
Простая щетинка — щетинка, состоящая из одной, нерасчлененной части (в противоположность сложным, или сочлененным щетинкам, см.); форма простых щетинок самая разнообразная: игловидная, гребенчатая, гарпунообразная, крючковидная, клювовидная, шпателевидная, мотыговидная и т. д.
Прототрох (prototrochus) — поясок ресничек в передней части трохофоры (предрото-вое кольцо ресничек).
Рострум — дистальная расширенная часть стержня (рукоятки) сложных щетинок, обычно заканчивающаяся двумя сочленительными зубцами, поддерживающими конечный членик щетинок.
Ротовой, или буккальный усик (cirrus buccalis) — брюшной усик II сегмента.
Рукоятка щетинок — у сложных (сочлененных) щетинок основной стержень, или древко (hastile), к которому прикреплен подвижный дистальный членик.
Сложная, или сочлененная щетинка — щетинка, состоящая из двух частей: основного стержня, или рукоятки, и сочлененного с ним дистального членика.
Спинигерные щетинки (setae spinigerae) — сложные (сочлененные) щетинки, дистальный членик которых в виде клинка.
Спинная чешуйка, или э л и т р a (elytra, elytron) — сильно сплющенные дорсо-вентрально спинные усики; налегая черепицеобразно друг на друга, они прикрывают собой значительную часть спинной поверхности червя.
Спинной бугорок — см. дорсальный бугорок.
Спинной войлок — сильно переплетенные, нередко засоренные илом, тонкие длинные нотоподиальные щетинки, прикрывающие спинную сторону некоторых афродитпд.
Спинной усик — см. дорсальный усик.
Стилоды (stylodae) — различные, преимущественно пальцевидной формы выросты на конце параподии.
Телескопические глаза — крупные глаза с хрусталиками.
Телотрох — поясок ресничек в задней части трохофоры.
Тентакула (tentacula), или антенна (antenna) — см. головные щупальца; в старых работах под термином тентакулы обозначались как головные щупальца (tentacula imparia, tentacula media), так и пальпы (tentacula lateralia).
Тентакулярнщй усик (cirrus tentacularis) — см. перистомиальные усики (щу-пальцевидные усики).
Тубикольные черви —- полихеты, живущие в постоянных трубках (переносных или прикрепленных).
Туловищные сегменты — сегменты между головной лопастью и пигиднумом (см. метастомиум). "*
Уриты (urites) —см. анальные усики.
Усики, или циррусы (cirri)—см. вентральные, дорсальные, тентакулярные и другие усики.
Усиковые сегменты — сегменты, снабженные спинными усиками (в отличие от чешуйчатых сегментов, снабженных спинными чешуйками, или элитрамп).
Фальцигерные щетинки (setae falcigerae) — сложные (сочлененные) щетинки, дистальный членик которых в виде серпа или различной формы крючка
Фасциальный бугорок — см. лобный гребень.
Фронтальные отростки (Jilameata frontalia) — рудиментарные латеральные щупальца у мацеллицефалин, иногда они отмечаются как «лобные рога».
Хобот (proboscis) — вывороченное наружу хоботное влагалище, содержащее глотку.
Хоботное влагалище, иначе буккальный отдел передней кншкп, или предгло-точная полость — при втянутой глотке лежит впереди нее, а при выпячивании глотки образует наружную ее стенку.
Хоботные папиллы — различного рода мягкие выросты на наружной стенке хобота.
Челюсти (mandibulae) — хитиноидные образования в глотке в виде мощных заостренных крючков, различного рода пластинок и зубчиков.
Чешуйчатые сегменты — сегменты, снабженные спинными чешуйками, пли элитрами (в противоположность усиковым сегментам, снабженным спинными усиками).
Цератостиль (ceratostylus) – дистальная часть головных щупалец.
Цератофор (ceratophorus) – подставка головных щупалец.
СПИСОК ЛИТЕРАТУРЫ
Загс, И. Г. Новые добавления в фауне Polychaeta. – Л., 1925 – 35 с.
Ушаков, П. В. Фауна СССР: в 2т. Т.2: Многощетинковые черви. – Наука, 1982 – 11с.
Цетлин, А. Б. Практический определитель многощетинковых червей Белого моря. – Изд-во Моск. ун-та,1980 - 52 с.
|

